effect of hyperthermia and hypoxia on the expression of dysadherin, a cancer associated cell-membran
DESCRIPTION
ÂTRANSCRIPT

ReseaRch
2 | 2011-2012 | Volume 1
roadB treetSScientific
Pranav Haravu
tion between hypoxia and metastasis. These in vitro stud-ies have been re-confirmed by in vivo tests, which again show hypoxia-induced metastasis in cancerous tissue. (3) (4)These in vivo and in vitro tests have been even further validated by observing a strong correlation between tumor hypoxia and high levels of metastasis in clinical models. When used as a prognostic indicator, high levels of tumor hypoxia have generally led to poor prognoses and survival rates. These studies provide grounds for hypothesizing that hypoxia, by some mechanism, leads to metastasis and tumor invasion.
Effect of HyperthermiaMany currently practiced and developing methods of
cancer therapy involve treating a tumor with local hyper-thermia in conjunction with chemotherapy, for example temperature sensitive liposomes. (5) The current use of localized hyperthermia is primarily to increase vascula-ture permeability within a tumor and reduce the amount of drug necessary to attain the therapeutic threshold. In addition, cancer cells are more vulnerable to increases in temperature than normal cells, due to a lack of heat-shock proteins (HSPs). (6) (7)Normal cells, which un-dergo slower cell cycles, produce much larger quantities of HSPs than cancerous cells. The function of HSPs is to ubiquitously bind to hydrophobic proteins, preventing the protein from denaturing due to heat. Without HSPs, enzymatic and structural proteins within the cancer cells denature, generally resulting in apoptosis. (6) (7)Though hyperthermia is currently used as a clinical therapy,
IntroductionThough cancer treatments have significantly pro-
gressed, metastasized tumors still pose a significant chal-lenge to modern therapeutic methods, resulting in poor prognoses and lower survival rates for patients with me-tastasized cancers. (1) Despite improved therapeutic prac-tices, metastasized tumors remain extremely difficult to treat for two fundamental reasons. One, we are unable to know where additional tumors will grow, and two; we are unable to prevent future tumors from reoccurring. It ap-pears then, that the most promising method of treatment for malignant tumors would include preventing metastasis from occurring, for which we need to identify and under-stand the mechanism resulting in metastasis. In this report we seek to investigate the role of dysadherin in a possible mechanism for both hypoxia- and hyperthermia-induced metastasis.
Effect of HypoxiaA significant percentage of tumors express hypoxia, a
cellular deficiency in available oxygen, due to the sporadic growth of cancerous cells. Normal cells grow at a much slower rate than their cancerous counterparts, allowing for capillaries to form and supply all cells with a steady supply of oxygen. (2) However, in cancerous tumors, high rates of cell growth result in portions of the tumor being distant from tumor vasculature, decreasing the cells’ oxy-gen supplies due to limitations in the diffusion of oxygen. (2) Numerous studies have found that hypoxic cells show a much higher metastatic potential, indicating a correla-
Effect of Hyperthermia and Hypoxia on the Expression of Dysadherin, a Cancer Associated Cell-Membrane Glycoprotein Shown to Increase Metastasis
ABSTRACT: Though cancer treatments have significantly progressed, metastasized tumors still pose a significant chal-lenge to modern therapeutic methods, resulting in poor prognoses and lower survival rates for patients with metastasized cancers. In this study we investigated the role of dysadherin in a possible mechanism for both hypoxia- and hyperthermia-induced metastasis. Dysadherin is a cell-membrane protein found highly expressed and glycosylated in cancer cells, but expressed only in a few normal cells. It down-regulates E-cadherin mediated cell-cell adhesion, facilitating metastasis. We treated Panc-1 and PC3 cells with 43°C hyperthermia and .5% O2 hypoxia. Western Blots, with β-actin as a loading control, showed a significant increase in expression of glycosylated dysadherin in Panc-1 cells exposed to hypoxic and hyperthermic conditions. However, unglycosylated dysadherin expression was not affected, leading us to believe unglycosylated dysad-herin does not play a significant role in metastasis. IF imaging of PC3 cells also showed a significant increase in dysadherin expression in samples treated with hypoxia or hyperthermia, quantified by the ratio of fluorescence of the secondary, FITC, to that of DAPI. The results suggest hypoxia- and hyperthermia-induced metastasis function by increasing the expression of glycosylated dysadherin, which in turn down-regulates E-cadherin and promotes metastasis. Inhibiting key steps within this mechanism could serve as a potential drug target, greatly reducing the metastatic potential of malignant tumors. To our knowledge, this is the first report indicating that hypoxia- and hyperthermia-induced metastasis may occur by increasing glycosylated dysadherin expression.

ReseaRch
Volume 1 | 2011-2012 | 3
roadB treetSScientific
main, an extracellular domain, and a cytoplasmic region. (19) Dysadherin was first thought to be similar to the RIC protein in mice, and was later shown to be FXYD5 (FXYD domain-containing ion transport regulator 5), a member of the FXYD family of proteins. (20)
When first discovered, dysadherin was shown to be ex-pressed in the majority of cancers, such as breast, cervical, lung, stomach, colon, and bladder, while only expressed in a few normal cell types, including lymphocytes, endothe-lial cells, and basal cells. (21) (19) Later however, it was shown to also be expressed in other epithelial tissues, such as kidney, primarily in the cortex, intestine, primarily in the duodenum, and lung. (20)
In its original discovery, it was observed that cell lines transfected with increased amounts of dysadherin showed decreased cell-cell adhesion. Upon analysis of E-cadherin and catenin expression, it was found that compared to controls, E-cadherin and α-catenin expression was signifi-cantly decreased in cells over-expressing dysadherin. (19) In addition, it was found that dysadherin was over-ex-pressed in regions of loose cell-cell contact; further indicat-ing that dysadherin down regulates E-cadherin expression and promotes metastasis. (19) These initial results were replicated and observed numerous times, including in vivo and patient studies. Over expression of dysadherin leading to the down regulation of E-cadherin was observed in gas-tric, thyroid, head and neck, cervical, pancreatic, colorec-tal, and lung carcinomas, as well as a multitude of others. When high dysadherin levels were observed, metastasis rates increased and survival rates decreased. (23-31)
In this report we attempt to show that both hypoxia and hyperthermia may induce an increase in dysadherin expression, partly explaining both hypoxia- and hyper-thermia-induced metastasis. This provides the missing link (step A in the schematic below), between hypoxia, hyperthermia, and metastasis.
the full effect of hyperthermia on cancer cell growth is still debated; as it has been shown to increase the rate of metastasis of tumors both in vivo and in vitro. (8) (9) We hypothesize that hyperthermia induced metastasis may be similar to hypoxia induced metastasis, in that both may be due to up-regulation of dysadherin.
Role of Dysadherin In Cell-Cell AdhesionIn order for a benign tumor cell to become invasive and
malignant, it must overcome cell-cell adhesion and enter a tumor’s vasculature. Cells are normally connected to each other through many junctional structures, such as tight junctions, adherens-type junctions, and desmosomes. (10) One such mechanism for mediating cell-cell adhesion in epithelial cells is E-cadherin, illustrated below (10). E-cadherin molecules expressed on adjacent cells bind to each other in the extracellular domain in a zipperlike fash-ion. In the intracellular domain, E-cadherin links to β and γ-Catenin, both of which link to α – Catenin, ultimately attaching to the cell’s actin cytoskeleton. (11)
The presence of this E-cadherin cell-cell adhesion mechanism has shown to be crucial in preventing metas-tasis of tumors, indicating a causation rather than corre-lation. (12) (13) Mutations in the E-cadherin gene and lower expression levels in cancer cells in vivo and in vitro, as well as from clinical data, consistently show increased metastasis and tumor invasion. (14) (15) (16) (17) Hypox-ic tumors have been shown to down-regulate E-cadherin expression, decreasing cell-cell adhesion and facilitating metastasis. (18)
Discovered in 2002, dysadherin is a cancer associated glycoprotein that also has been shown to down-regulate E-cadherin mediated cell-cell adhesion. Dysadherin is comprised of a 178 amino acid sequence, located on 19q13.12, and consists of two hydrophobic regions, cor-responding to a signal peptide and the transmembrane do-
Figure 1. E-cadherin-mediated cell–cell adhesion. There is an extracellular component that links to other E-cadherin complexes in a zipperlike fashion, and an inner complex that links to the cell’s cytoskeleton.
Figure 2. Schematic showing possible major steps from hy-poxia and hyperthermia to metastasis. The red arrow, the missing link, has not yet been shown, and is the objective of this paper.

ReseaRch
4 | 2011-2012 | Volume 1
roadB treetSScientific
Immunofluorescence MicroscopyThe cover slips onto which cells were cultured were
first rinsed with PBS, fixed with 4% PFA, incubated for 2 hours with primary antibody, and then 2 hours with FITC conjugated goat secondary antibody. The slips were then mounted with a photo-bleacher reducer and DAPI nuclear stain. Images were taken for .5 second expo-sures at wavelengths corresponding to DAPI and FITC. Results
Western Blots
Initial Western Blots confirmed the expression of dys-adherin in both cell lines at a MW of around 50kDa. Fol-lowing the confirmation of dysadherin expression, we pro-ceeded to expose the cell lines to treatment. The Western Blot for Panc-1 cells exposed to the 4 different treatments is given in figure 3. When we blotted for dysadherin, shown in figure 3, we see two sets of bands in each lane; a strong region of bands around 50 kDa, and a weaker almost background level set of bands between 15 and 20 kDa. The bands with MW ≈ 50 kDa are dysadherin mol-ecules heavily O-glycosylated in the extracellular domain, while the much fainter bands between 15 and 20 kDa are most likely unglycosylated dysadherin. A β- actin loading control enabled us to compare lanes to each other.
Thus, we are able to see that the glycosylated dys-adherin bands from cells exposed to hypoxia and hy-perthermia (bands A” and D” respectively) are much darker than their controls, indicating that dysadherin expression increased when exposed to hypoxic and hy-perthermic conditions. In addition, we notice that the levels of unglycosylated dysadherin expression, bands A’, B’, C’, and D’, are relatively equal between all treat-ments, indicating that the treatment most likely had no impact on unglycosylated dysadherin expression. Immunofluorescence Microscopy
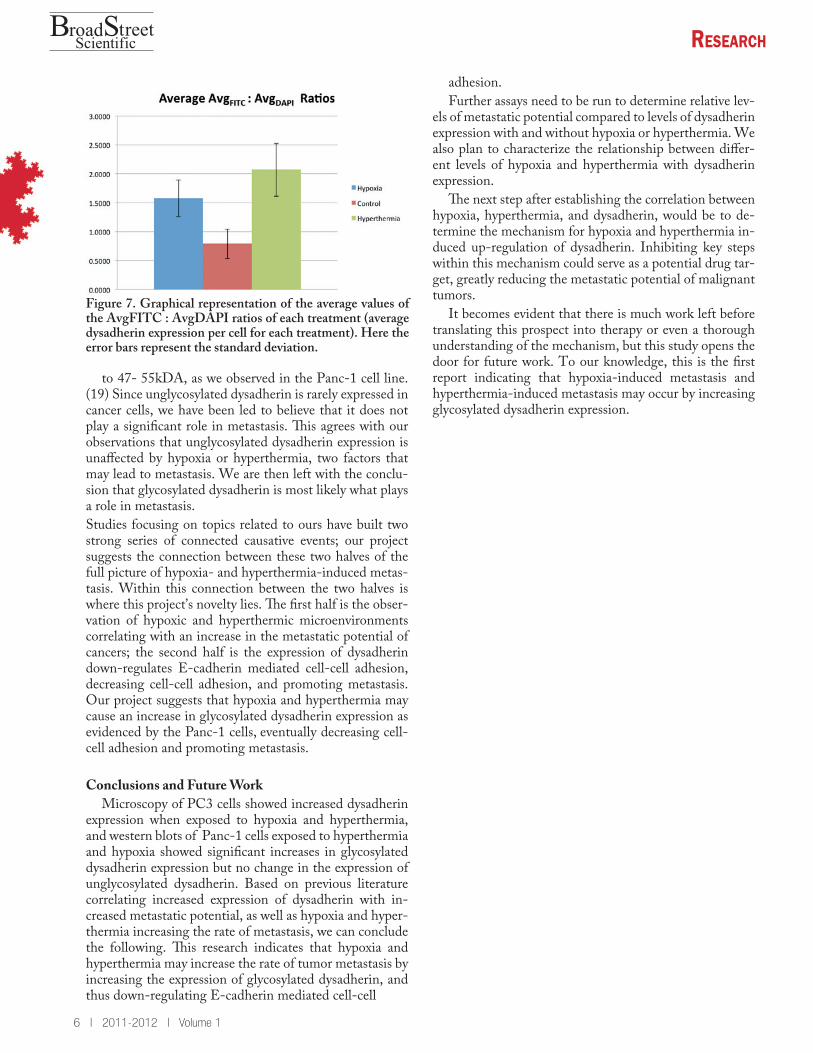
We used ImageJ to measure the average pixel count for the entire field. The average pixel count of the images taken at the FITC wavelengths, those in Figure 5 (Avg-FITC ) gave a ratio of conjugated FITC antibodies to total area, while that of the images taken at DAPI wave-lengths, those in Figure 6, (Avg DAPI ) gave the ratio of cell nuclei (cells) to total area. The ratio of AvgFITC : AvgDAPI gives us the average ratio of conjugated FITC antibodies per cell, an indicator of the amount of primary antibody binding and thus the levels of dysadherin expres-sion. These results are given in Figure 6, which shows the AvgFITC : AvgDAPI ratio of each sample, A, B, C, and D. The average AvgFITC : AvgDAPI ratios of all the samples from each set is shown in Figure 7. These results show statistically significant higher levels of FITC fluo-rescence per cell in the samples treated with hypoxia and hyperthermia than the control, indicating an increase in dysadherin expression.
Methods and Materials
Cell CulturingPC3 (prostate cancer) and Panc-1 (pancreatic can-
cer) cell lines were all established in our laboratory, cul-tured in F12K+ 10% Fetal Bovine Serum + 1% Anit-Anti and DMEM + 10% Fetal Bovine Serum + 1% Anit-Anti respectively. For IF imaging, cells were cul-tured onto BD Biocoat cover slips. All cell culturing and treatment was conducted in a sterile environment.
Cell TreatmentsEach cell line was split into 4 different 10mL treat-
ment plates (for cells to be harvested for Western blots) or 6-well plates (for cells cultured on cover slips for IF imaging). Hypoxia was induced for 24 hours at .5% O2. Hyperthermia was induced with 2 1 hour wa-ter baths, 24 hours apart, at 43°C. Two controls were also used, one set with no treatment (left in incubator), and another was exposed to the same dosage as the hy-perthermia, except the water bath was set at 37°C. The water bath control was not used in the IF imaging. Protein Harvesting
Existing media in cell plates was decanted, and each plate was rinsed with PBS. Cells were then scraped into 15mL centrifuge tubes and centrifuged at 4,000 rpm for 10 minutes. Remaining media and PBS was suctioned off, leaving only the cell pellet at the bottom. This pellet was then suspended in lysis buffer (10 mM PBS pH 7.4, 0.5% Triton X-10, 2 mM CaCl2, 10 ug/ml leupeptin, 2 ug/ml pepstatin A, 10 ug/mL aprotinin) (19), transferred to 1.5 mL tubes, and incubated on ice for 30 minutes. The samples were then centrifuged at 14,500 rpm for 20 minutes, and the supernatant, which contained the Triton X-100 soluble components of the cell (proteins), was col-lected and stored at -20°C, while the pellet was discarded. Western Blots
As the expected weight of the protein ranged from 40-55 kDa when glycosylated, and 19-20 kDa when unglycosylated, we separated 30μg protein samples in a 12% SDS/PAGE gel. We then transferred the proteins to poly-vinylidene difluoride (PVDF) membranes and blocked overnight at 4°C in a 5% milk blocking buf-fer. After washing in a 1% milk buffer, the membranes were incubated at room temperature for 2 hours with the primary antibody (1:200). The primary antibody used was Dysadherin (T-14) obtained from Santa Cruz BiotechnologyInc. (sc-30604). This antibody targets the extracellular domain of dysadherin, epitope 50-100. Membranes were then rinsed and incubated for 2 hours at room temperature with anti-goat secondary antibod-ies (1:1000), and then developed using an ECL kit.

ReseaRch
Volume 1 | 2011-2012 | 5
roadB treetSScientific
DiscussionIn the IF microscopy we saw that hypoxia and hyper-
thermia both significantly increased dysadherin expression levels in PC3 cells. We have observed that in Panc-1 cells, hypoxia and hyperthermia increase the expression of gly-cosylated dysadherin, as evidenced by the stronger bands at MW = 50 in A” and D” in Figure 4. In addition, we also see that the expression of unglycosylated dysadherin is seemingly not affected by hypoxia or hyperthermia. This has led us to believe that unglycosylated dysadherin would most likely not play a significant role in either hypoxia-induced or hyperthermia-induced metastasis.
Previous studies have found that when expressed in non-cancerous cells, dysadherin is highly unglycosylated and has an observed MW of around 20 kDa. (20) How-ever, when expressed in malignant cancerous cells, dysad-herin becomes highly glycosylated, raising its MW
Illustrations
Figure 3. Western Blot of Panc-1 cells blotted for dysad-herin. Each lane has a different treatment; A has hypoxia, B is the control with no treatment, C is the hyperthermia control (a water bath set at 37°C ), and D is treated with hy-perthermia. We see much stronger bands in regions A” and D” than in B” and C”, all of which correspond to glycosylated dysadherin (MW = 47-55 kDa). Regions A’, B’, C’, and D’ show similar band strengths and correlate to unglycosylated dysadherin (MW = 19 kDa). Thus, the expression of glyco-sylated dysadherin is shown to have increased under hypoxic and hyperthermic conditions.
Figure 4. Images taken at a wavelength corresponding to DAPI. The first row (1A-1D) consists of the samples treated with hypoxia, the second row (2A-2D) consists of the samples set as a control, and the third row (3A-3D) consists of the samples treated with hyperthermia.
Figure 5. Images taken at a wavelength corresponding to FITC. The ordering of the images is identical to that in Fig-ure 5.
Figure 6. Graphical representation of the AvgFITC : Avg-DAPI ratios (average dysadherin expression per cell in each sample). Each cluster is a different set of samples, and the error bars represent standard error.
ReseaRch
6 | 2011-2012 | Volume 1
roadB treetSScientific
to 47- 55kDA, as we observed in the Panc-1 cell line. (19) Since unglycosylated dysadherin is rarely expressed in cancer cells, we have been led to believe that it does not play a significant role in metastasis. This agrees with our observations that unglycosylated dysadherin expression is unaffected by hypoxia or hyperthermia, two factors that may lead to metastasis. We are then left with the conclu-sion that glycosylated dysadherin is most likely what plays a role in metastasis. Studies focusing on topics related to ours have built two strong series of connected causative events; our project suggests the connection between these two halves of the full picture of hypoxia- and hyperthermia-induced metas-tasis. Within this connection between the two halves is where this project’s novelty lies. The first half is the obser-vation of hypoxic and hyperthermic microenvironments correlating with an increase in the metastatic potential of cancers; the second half is the expression of dysadherin down-regulates E-cadherin mediated cell-cell adhesion, decreasing cell-cell adhesion, and promoting metastasis. Our project suggests that hypoxia and hyperthermia may cause an increase in glycosylated dysadherin expression as evidenced by the Panc-1 cells, eventually decreasing cell-cell adhesion and promoting metastasis.
Conclusions and Future WorkMicroscopy of PC3 cells showed increased dysadherin
expression when exposed to hypoxia and hyperthermia, and western blots of Panc-1 cells exposed to hyperthermia and hypoxia showed significant increases in glycosylated dysadherin expression but no change in the expression of unglycosylated dysadherin. Based on previous literature correlating increased expression of dysadherin with in-creased metastatic potential, as well as hypoxia and hyper-thermia increasing the rate of metastasis, we can conclude the following. This research indicates that hypoxia and hyperthermia may increase the rate of tumor metastasis by increasing the expression of glycosylated dysadherin, and thus down-regulating E-cadherin mediated cell-cell
Figure 7. Graphical representation of the average values of the AvgFITC : AvgDAPI ratios of each treatment (average dysadherin expression per cell for each treatment). Here the error bars represent the standard deviation.
adhesion. Further assays need to be run to determine relative lev-
els of metastatic potential compared to levels of dysadherin expression with and without hypoxia or hyperthermia. We also plan to characterize the relationship between differ-ent levels of hypoxia and hyperthermia with dysadherin expression.
The next step after establishing the correlation between hypoxia, hyperthermia, and dysadherin, would be to de-termine the mechanism for hypoxia and hyperthermia in-duced up-regulation of dysadherin. Inhibiting key steps within this mechanism could serve as a potential drug tar-get, greatly reducing the metastatic potential of malignant tumors.
It becomes evident that there is much work left before translating this prospect into therapy or even a thorough understanding of the mechanism, but this study opens the door for future work. To our knowledge, this is the first report indicating that hypoxia-induced metastasis and hyperthermia-induced metastasis may occur by increasing glycosylated dysadherin expression.

ReseaRch
Volume 1 | 2011-2012 | 7
roadB treetSScientific
17. E-cadherin complex and its abnormalities in human breast can-cer. Jiang, Wen G and Mansel, Robert E. 2000, Surgical Oncology, pp. 151-171.18. Regulation of E-cadherin: does hypoxia initiate the metastatic cascade? Beavon, I.R.G. s.l. : Journal of Clinical Pathology, April 1999, pp. 179-188.19. Dysadherin, a cancer-associated cell membrane glycoprotein, down-regulates E-cadherin and promotes metastasis. Ino, Yoshi-nori, et al. 2002, PNAS, pp. 265-270.20. Interaction with the Na, K-ATPase and Tissue Distribution of FXYD5 (Related to Ion Channel). Lubarski, Irina, et al. 2005, The Journal of Biological Chemistry, pp. 37717-37724.21. Dysadherin: a new player in cancer progression. Nam, Jeong-Seok, Hirohashi, Setsuo and Wakefield, Lalage M. 2007, Cancer Letters, pp. 161-169.22. Prognostic significance of dysadherin expression in patients with non-small cell lung cancer. Tamura, Masaya, et al. 2005, Gen-eral Thoracic Surgery, pp. 740-745.23. Dysadherin expression in gastrointestinal stromal tumors (GISTs). Liang, Jian, et al. 2009, Pathology - Research and Prac-tice, pp. 445-450.24. Dysadherin: Expression and Clinical Significance in Thyroid Carcinoma. Sato, Haruhiro, et al. 2003, The Journal of Clinical En-docrinology & Metabolism, pp. 4407-4412.25. Clinical Significance of Dysadherin Expression in Gastric Can-cer Patients. Shimada, Yutaka, et al. 2004, Clinical Cancer Re-search, pp. 2818-2823.26. Significance of dysadherin and E-cadherin expression in dif-ferentiated-type gastric carcinoma with submucosal invasion. Mae-hata, Yoshitomo, et al. 2011, Human Pathology, pp. 558-567.27. Prognostic Significance of Dysadherin Expression in Cervical Squamous Cell Carcinoma. Wu, Dan, et al. 2004, Pathology On-cology Research, pp. 212-218.28. Dysadherin Expression in Head and Neck Squamous Cell Carcinoma; Association With Lymphangiogenesis and Prognostic Significance. Kyzas, Panayiotis A, et al. 2006, American Journal of Surgical Pathology, pp. 185-193. 29. Prognostic significance of dysadherin expression in advanced colorectal carcinoma. Aoki, S, et al. 2003, British Journal of Can-cer, pp. 726-732.30. Dysadherin Overexpression in Pancreatic Ductal Adenocarci-noma Reflects Tumor Aggressiveness: Relationship to E-cadherin Expression. Shimamura, Takeshi, et al. 2003, Journal of Clinical Oncology, pp. 659-667.31. Dysadherin Expression Facilitates Cell Motilitiy and Metastatic Potential of Human Pancreatic Cancer Cells. Shimamura, Takeshi, et al. 2004, Cancer Research, pp. 6989-6995.
References 1. Metastasis Mechanism. Geiger, Thomas R and Peeper, Daniel S. 2009, Biochimical et Biophysica Acta, pp. 293-308.2. Hypoxia modulated gene expression: angiogenesis, metastasis and therapeutic exploitation. Dachs, G.U. and Tozer, G.M. 13, s.l. : European Journal of Cancer, 2000, Vol. 36, pp. 1649-1660.3. Progression and metastasis in a transgenic mouse breast cancer model: Effects of exposure to in vivo hypoxia. Kalliomaki, Tuula M, et al. 2009, Cancer Letters, pp. 98-108.4. Tumor Hypoxia Correlates with Metastatic Tumor Growth of Pancreatic Cancer in an Orthotopic Model. Buchler, Peter, et al. 2004, Journal of Surgical Research, pp. 295-303.5. The development and testing of a new temperature-sensitve drug delivery system for the treatment of solid tumors. Needham, David and Dewhirst, Mark W. s.l. : Advanced Drug Delivery Reviews, 2001, Advanced Drug Delivery Reviews, Vol. 53, pp. 285-305.6. The cellular and molecular basis of hyperthermia. Hildebrandt, Bert, et al. s.l. : Critical Reviews in Oncology/Hematology, 2002, Vol. 43, pp. 33-56.7. Effect of hyperthermia on the viability and the fibrinolytic poten-tial of human cancer cell lines. Fukao, Hideharu, et al. 2000, Clinica Chimica Acta, pp. 17-33.8. Hyperthermia increases the metastatic potential of murine mela-noma. Oliveira-Filho, R.S., Bevilacqua, R.G. and Chammas, R. 1997, Brazilian Journal of Medical and Biological Research, pp. 941-945.9. Effects of Total-Body Hyperthermia on Metastases from Ex-perimental Mouse Tumors. Oda, Masayuki, Koga, Shigemasa and Maeta, Michio. 1985, Cancer Research, pp. 1532-1535.10. The role of the cell-adhesion molecule E-cadherin as a tumour-suppressor gene. Christofori, Gerhard and Semb, Henrik. 1999, Trends in Biochemical Sciences, pp. 73-76.11. The E-cadherin-catenin complex in tumour metastasis: struc-ture, function and regulation. Beavon, I.R.G. 2000, European Jour-nal of Cancer, pp. 1607-1620.12. Restoring E-cadherin-mediated cell–cell adhesion increases PTEN protein level and stability in human breast carcinoma cells. Li, Zengxia, et al. 2007, Biochemical and Biophysical Research Communications, pp. 165-170.13. E-cadherin-mediated Cell-Cell Adhesion Prevents Invasive-ness of Human Carcinoma Cells. Frixen, Uwe H, et al. 1991, The Journal of Cell Biology, pp. 173-185.14. The E-cadherin cell–cell adhesion complex and lung cancer invasion, metastasis, and prognosis. Bremnes, Roy M, et al. 2002, Lung Cancer, pp. 115-124.15. Influence of E-cadherin dysfunction upon local in vasion and metastasis in non-small cell lung cancer. Shibanuma, Hiroyuki, et al. 1998, Lung Cancer, pp. 85-95.16. Immunohistological Analysis of E-cadherin, a-, B- and y-Catenin Expression in Colorectal Cancer: Implications for Cell Adhesion and Signaling. Ghadimi, B.M., et al.