effect of brain-derived neurotrophic factor, nerve growth factor, and neurotrophin-3 on functional...
TRANSCRIPT

JOURNAL OF NEUROTRAUMAVolume 17, Number 12, 2000Mary Ann Liebert, Inc.
Effect of Brain-Derived Neurotrophic Factor, Nerve GrowthFactor, and Neurotrophin-3 on Functional Recovery and
Regeneration After Spinal Cord Injury in Adult Rats
JUN NAMIKI, ATSUHIRO KOJIMA, and CHARLES H. TATOR
ABSTRACT
This study examined whether continuous intramedullary infusion of brain-derived neurotrophic fac-tor (BDNF), nerve growth factor (NGF), or neurotrophin-3 (NT-3) had either an early neuropro-tective effect or a delayed effect on regeneration after spinal cord injury (SCI) in adult rats. BDNF,NGF, NT-3 or vehicle was infused at a rate of 625 ng/h into the SCI site at T3 through an implantedcannula attached to an osmotic pump. This infusion was maintained for 14 days after a 35-g clipcompression injury. At 4 weeks after injury, the axonal tracer fluorogold (FG) was introduced intothe spinal cord caudal to the lesion and the animals sacrificed 3 days later following behavioral as-
sessment. The inclined plane score was significantly higher in BDNF-treated animals (45 ± 3°) com-
pared to control animals (36 ± Io) at 1 week after injury (p < 0.05), although the scores were not
significantly different at later times. BDNF-treated animals also showed more FG-labeled cells inthe red nucleus and sensorimotor cortex (1,638 ± 350 and 124 ± 83, respectively) compared to con-
trols (1,228 ± 217 and 36 ± 15, respectively) and a lower percent cavitation at the injury site (21.4 ±
10.4%) compared to control animals (32.3 ± 11.7%). Invasion & proliferation of Schwann cells andformation of peripheral myelin were more prominent at the injury site in the BDNF-treated ani-mals than in the other groups. These results indicate that continuous intramedullary infusion ofBDNF provides neuroprotection and enhances some regenerative activity after SCI.
Key words: brain-derived neurotrophic factor; fluorogold; myelin; nerve growth factor; neurotrophin-3;Schwann cell; spinal cord injury
INTRODUCTION 1999; Whittemore et al, 1999). Promotion of neuronalsurvival by the administration of exogenous neurotrophic
There are three main types of therapeutic strategies factors to counteract secondary injury is a promising ap-for spinal cord injury (SCI): rescue of injured neu- proach to treatment of SCI, and members of the neu-
rons from secondary injury with prevention of neuronal rotrophin family, including nerve growth factor (NGF),death; promotion of target-directed axonal regeneration; brain-derived neurotrophic factor (BDNF), and neuro-
and neural replacement by endogenous or transplanted tophin-3 (NT-3), have been shown to have significant ef-neural stem cells or other tissue transplantation (reviewed fects on the prevention of neural damage after SCI. Deathin Tator and Fehlings, 1991; Anderson and Hall, 1993; of rubrospinal neurons after spinal cord hemisection was
Tator, 1995, 1998; Namiki and Tator, 1999; Park et al, prevented by BDNF (transiently prevented by NGF or
University of Toronto and Toronto Western Research Institute, Toronto, Onatrio, Canada.
1219

NAMIKI ET AL.
NT-3), and retrograde cell death of spinocerebellar neu-
rons in newborn rats was prevented by NT-3 (Diener andBregman, 1994). Similarly, local administratioin ofBDNF prevented atrophy of rubrospinal neurons and pro-moted regeneration of rubrospinal axons after SCI in adultrats. (Tetzlaff et al., 1994; Kobayashi et al., 1997; Liu etal., 1999). BDNF also reduced the longitudinal extent ofthe lesion cavity after spinal cord hemisection in adult rats(Novikova et al., 1996). Kim et al. (1996) showed thatNGF or BDNF preserved neural tissue after spinal cordcontusion in adult rats, and this beneficial histological ef-fect at the injury site was accompanied by promotion offunctional recovery, although this was observed only 1week after injury. Houweling et al. (1998a), Jakeman etal. (1998), and Liu et al. (1999) also reported that the lo-cal application of BDNF after SCI resulted in a decreasedloss of function in the early postinjury phase. On the otherhand, Bradbury et al. (1998) delivered BDNF to the SCIsite via an osmotic minipump and reported that BDNFhad no neuroprotective effect. Nakahara et al. (1996) alsoreported that BDNF-secreting fibroblast grafts did not pro-mote axonal regeneration after adult rat SCI. Thus, the ef-fect of BDNF on the treatment of SCI remains contro-versial. Similarly, studies on the effect of NGF on
functional recovery after SCI have also produced incon-sistent results (Kim et al., 1996; Tuszynski et al., 1997).NGF has been shown to promote axonal regeneration inthe spinal cord (Fernandez et al., 1993; Diener and Breg-man, 1994; Tuszynski et al., 1996, 1997; Oudega et al.,1996; Hiraizumi et al., 1996; Kim et al., 1996; Grill et al.,1997a; Kobayashi et al., 1997). However, Lee et al. (1999)reported that NGF had no significant neuroprotective ef-fect on the injured cord. There have been two reports ofa neuroprotective effect of NT-3 in the injured adult spinalcord (Giehl and Tetzlaff, 1996; Bradbury et al., 1998),and several reports of axonal regeneration promotion withNT-3 after SCI in adult rats (Schnell et al., 1994; Breg-man et al., 1997; Grill et al., 1997b; Ye and Houle, 1997;McTigue et al., 1998). Based on these divergent reportsthere is an obvious need for further examination of theneuroprotective and regenerative effects of BDNF, NGFand NT-3 measured by functional and histological as-
sessment after SCI in adult rats.To date, neurotrophic factors have been administered
locally into the injured spinal cord through a variety ofmethods, including a single injection through a glass cap-illary (Schnell et al., 1994), placement in Gelfoam (Di-ener and Bregman, 1994; Bregman et al., 1997; Ye andHoule, 1997), and collagen (Houweling et al., 1998a,b),use of genetically modified neurotrophic factor-secretingfibroblasts or Schwann cells (Kim et al., 1996; Nakaharaet al., 1996; Tuszynski et al., 1996, 1997; Grill et al.,1997a,b; Menei et al., 1998; Liu et al., 1999; Weidner et
al., 1999), via adenoviral vectors encoding neurotrophicfactors (Zhang et al., 1998), or via osmotic pump (Fer-nandez et al., 1993; Xu et al., 1995; Novikova et al., 1996,1997; Oudega et al., 1996; Novikov et al., 1997;Kobayashi et al., 1997; Jakeman et al., 1998; Houwelinget al., 1998; Bradbury et al., 1998; Yick et al., 1999; Leeet al., 1999).
The present study focused on spinal cord function andhistological states, and was designed to examine whetherBDNF, NGF, or NT-3 prevented neural damage and pro-moted regenerative activity after clip compression SCI inadult rats. Continuous intramedullary infusion of the neu-
rotrophic factors into the spinal cord lesion site was testedwith an osmotic pump, which may provide more effec-tive delivery than epidural (Fernandez et al., 1993) or
subarachnoid delivery (Verge et al., 1990; Gold et al.,1993, 1997; Novikova et al., 1996, 1997; Novikov et al.,1997; Kishino et al., 1997; Jakeman, et al., 1998).
MATERIALS AND METHODS
AnimalsAdult female Wistar rats weighing 220-290 g were
used. All protocols were approved by the Animal CareCommittee of the Research Institute of the UniversityHealth Network in accordance with the policies establishedin the Guide to the Care and Use of Experimental Animalsprepared by the Canadian Council on Animal Care.
Normal Animals After Cannula Insertion andIntramedullary Infusion
To test for the effects of intramedullary delivery, twonormal animals without SCI had cannula implantation atT3 followed by continuous vehicle infusion into thespinal cord and were compared functionally and histo-logically with five normal animals which did not undergothese procedures.
Clip Compression InjurySeven animals were assigned to each treatment group
(NGF, BDNF, NT-3, and vehicle control). One animal ineach group died prior to the 4-week survival period, andthese were excluded from analysis. The animals were
anesthetized by inhalation of halothane, and a laminec-tomy performed at the T3 vertebral level under asepticconditions. With the aid of an operating microscope, a
modified aneurysm clip with blades 1.3 mm in width(Walsh Manufacturing, Oakville, ON) calibrated to de-liver a closing force of 35 g was positioned extradurallyaround the spinal cord at the T3 level, and the spinal cordwas then acutely compressed for 1 min, as described pre-
1220

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
viously (Rivlin and Tator, 1978; Theriault and Tator,1994). The animals were given buprenorphine (0.03mg/kg) subcutaneously for pain control before awaken-ing and in the morning after surgery. Penicillin G (10,000U/kg) was also injected subcutaneously after surgery. Theanimals were housed two per cage in a temperature-con-trolled room (27°C) with a 12-h light/dark cycle for 4weeks. After SCI, rats had manual bladder evacuationthree times daily and were monitored weekly for pH,leukocytes, and blood in the urine. Urinary tract infec-tions were treated with intramuscular gentamicin (8mg/kg, once daily).
Delivery of Neurotrophic FactorsThe neurotrophic factors were infused directly into the
lesion site through a stainless steel cannula (34 gauge;Plastic One, Roanoke, VA) attached to an osmotic pump(model 2002; Alza, Palo Alto, CA) via a polyethylenetube (PE-60 Intramedic; Clay Adams, Parsippany, NJ).NGF, BDNF or NT-3 (provided by Regeneran Pharma-ceuticals, Tarrytown, NY) was delivered beginning im-mediately after SCI and continuing for 14 days at a flowrate of 0.5 pL/h with an initial concentration of 1.25mg/mL, resulting in an infusion rate of 625 ng/h. Con-trol animals received equivalent amounts of vehicle(Ringer solution). The osmotic pumps were preincubatedin sterile saline at 37°C overnight before implantation toallow immediate pumping of the neurotrophic factors.Within 30 min after the compression injury, the cannulawas passed into the epicenter of the lesion site at T3 withthe aid of a stereotactic device. The tip was positioned atthe midlline of the cord, 1.0 mm in depth from the dor-sal surface of the cord, near the corticospinal tract. He-mostatic clips (Atrauclip; Pilling, Fort Washington, PA)were placed in the spinous processes of T2 and T4, andthe cannula and its connector were then fixed to thosespinous processes with methyl methacrylate (Bone Ce-ment, low viscosity; Zimmer, Warsaw, IN). The pumpwas then implanted subcutaneously over the thoracicspine slightly caudal to the scapulae. Fourteen days later,the animals were briefly anesthetized by inhalation ofhalothane, the pumps were removed without disturbanceof the cannula assembly, and the animals allowed to sur-
vive an additional 2 weeks without further infusion ofneurotrophic factor or vehicle.
Functional AssessmentAll outcome measures were assessed in a blinded fash-
ion. Neurological function was evaluated weekly with theinclined plane technique (Fehlings and Tator, 1992) fromthe first week following surgery. Briefly, rats were posi-tioned laterally with respect to the long axis of the in-
clined plane, and the maximum angle achieved withoutfalling for 5 sec was assessed.
Electrophysiological assessment of motor function was
performed by recording motor evoked potentials (MEPs)at 4 weeks after surgery, immediately before sacrifice forhistological assessment. MEPs were measured underanesthesia after the intraperitoneal injection of a mixtureof a-chloralose (75 mg/kg) and urethane (525 mg/kg).Muscle paralysis was produced with pancuronium bro-mide (1 mg/kg, every 30 min i.V.), and the rats were me-
chanically ventilated. A small craniectomy was made inthe left parietal region and anodal stimuli (10 mA, 50 ps,8.1 Hz) were applied extradurally over the hindlimb re-
gion of the sensorimotor cortex (SMC); (Paxinos andWatson, 1986; Hebel and Stromberg, 1986) with a ref-erence electrode under the hard palate. The T7 spinousprocess was removed, and the recording electrode was
placed extradurally on the dorsal surface of the spinalcord at the T7 level. The reference electrode was placedin the paraspinal muscles adjacent to the recording elec-trode. MEPs were amplified and filtered (band pass,30-3,000 Hz). A total of 512 responses were averagedand replicated. The peak latency and peak-to-peak am-
plitude were calculated.
Histology and ImmunohistochemistrySeventy-two hours prior to the end of the 4-week ob-
servation period, all animals received fluorogold (FG) forretrograde neuronal labeling. The animals were anes-
thetized by inhalation of halothane, and followinglaminectomy at T10, the spinal cord was transected atT10 and a Gelfoam pledget soaked in 4% FG (Fluo-rochrome, Englewood, CO) was inserted between thestumps of the transected spinal cord (Midha et al, 1987;Theriault and Tator, 1994; Agrawal and Fehlings, 1997).Seventy-two hours after FG introduction and immedi-ately after the MEP measurement described above, theanimals were deeply anesthetized by injection of an ad-ditional amount of a-chloralose and urethane and thentranscardially perfused with 4% paraformaldehyde (inphosphate buffer saline [PBS], pH 7.3-7.4). The brainwas removed and subsequently sectioned in the coronalplane at 40 pm in a cryostat. The number of FG-labeledcells was counted in the RN (magnocellular and parvi-cellular portions) and SMC in every alternate section withthe fluorescent microscope at X100. The criteria forcounting labeled cells were described previously (Theri-ault and Tator, 1994; Agrawal and Fehlings, 1997).
The spinal cord encompassing the injury site was post-fixed in 10% neutral buffered formalin, and then seg-ments of the spinal cord were embedded in paraffin andcut transversely at 8 pm. Every 20th section was stained
1221

NAMIKI ET AL.
with hematoxylin and eosin, and luxol fast blue(H&E/LFB), LFB in combination with periodic acidSchiff (PAS), Masson's trichrome, or used for immuno-histochemistry for demonstrating glial fibrillary acidicprotein (GFAP). After deparaffinization, sections forGFAP immunohistochemistry were blocked for endoge-nous peroxidase with 1% methanol peroxidase for 30min, washed in 0.1 M PBS, and then blocked in normalgoat serum containing 0.3% Triton-X 100 for 1 h at room
temperature. Mouse monoclonal anti-GFAP antibody(1:200 dilution; Boehringer Mannheim Canada, Laval,QC) was used with biotinylated anti-mouse secondary an-
tibody (Vector Laboratory, Burlingame, CA) and avidin-biotin-peroxidase complex (Vectastain Elite ABC KitStandard, Vector Laboratories). 3,3'-Diaminobenzidine(DAB, Vector Laboratories) was applied as the chro-mogen. Negative controls were obtained by omission ofthe primary antibody along with known positive controls.
One cross-section containing the lesion epicenter was
selected from each rat, and the percent cavitation was an-
alyzed with computer-assisted image analysis (MCIDM4; Imaging Research, ON). Sections were digitized atX10, and the spinal cord cross-section and residual tis-sue were outlined. The percent cavitation was calculatedas the area of cavitated tissue divided by the area of thetotal cross-section at the level of the injury.
Regenerative activity at the site of maximum injurywas assessed on the basis of the following: (1) presenceand degree of Schwann cell proliferation; (2) presence ofperipheral myelin formation; and (3) new central myelinformation. The identification of peripheral myelin andcentral myelin was based on morphology with H&E/LFBand on immunohistochemistry in sections stained withLFB in combination with PAS. Morphologically,Schwann cells produce thicker, more compact myelinthan those produced by oligodendrocytes (Gilmore andDuncan, 1968; Blakemore and Patterson, 1975; Griffithsand McCulloch, 1983; Wallace et al., 1987; West andCollins, 1989; Bunge et al., 1994; Salgado-Ceballos etal., 1998). Oligodendrocyte myelin typically shows a
"puffing" of the lamellae at intervals around its circum-ference and along its length, giving it a lighter, irregularappearance by comparison with Schwann cell myelin,which usually remained compact through fixation and de-hydration (Blight and Young, 1989). With LFB in com-bination with PAS, peripheral myelin stains blue in con-
trast to the blue-green of central nervous system myelin(Feigin and Cravioto, 1961; Gilmore and Duncan, 1968).Schwann cells had distinct morphological features in-cluding small size (smaller than astrocytes), elongatedshape, spindle-shaped nuclei, the tendency to pallisade,and densely blue cell bodies with H&E/LFB. Schwann
cells were more easily identified when they were associ-ated with adjacent peripheral myelin. Also, Schwanncells were not associated with collagen in the sectionsstained with Masson's stain or with GFAP immunoreac-tivity. A simple, subjective rating scale for Schwann cellproliferation and peripheral myelin formation was em-
ployed: A, marked activity; B, moderate activity; and C,minimal activity.Statistics
The inclined plane score at each time point, cell countdata for FG labeled cells, and percent cavitation at theinjury site were analyzed by one-way analysis of vari-ance (ANOVA). Post hoc analyses were performed us-
ing the Student-Newman-Keuls method. A variance sta-bilizing (square root) transformation of the cell count datawas performed prior to parametric analysis to correct forunequal variances (Fehlings and Tator, 1995; Agrawaland Fehlings, 1997). The results are expressed as mean ±SEM and differences were considered significant at p <0.05. Regenerative activity including invasion and pro-liferation of Schwann cells and new peripheral myelinformation was analyzed by Kruskal-Wallis one-wayANOVA on rank. Post hoc analyses were performed us-
ing the Student-Newman-Keuls method, and differenceswere considered significant at p < 0.05.
RESULTS
Effects of Cannula Implantation and ContinuousInfusion in the Normal Spinal Cord
To test the possible damaging effects on spinal cordmotor function of cannula implantation and the continu-ous infusion of solutions into the spinal cord, an additionalcontrol group of uninjured animals underwent cannula im-plantation at T3 followed by vehicle infusion into thespinal cord for 14 days. These animals (vehicle infusiongroup, n = 2) survived for 4 weeks, the same experi-mental period as the injured animals, and were comparedto nonimplanted normal control animals with respect tothe inclined plane score (n = 5), motor evoked potentials(n = 4), and retrograde neuronal labeling (n = 5). The ve-
hicle infusion group showed no significant differencesfrom the normal control group in inclined plane score
(normal control, 72 ± Io; vehicle infusion group, 73 ± 3°at 1,2, 3, or 4 weeks, respectively), or in latency and am-
plitude of MEPs (Table 1), indicating that the deliverysystem did not significantly impair these indices of mo-
tor function. However, the vehicle infusion group showeda significant decrease in the number of FG-labeled cellsin the SMC compared to the normal control group, al-
1222

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
Table 1. Functional and Histological Effects of Cannula Implantationand Continuous Infusion in the Normal Spinal Cord
Normal control(uninjured)
Vehicle infusion(uninjured)
n = 2
MEP latency (msec)
MEP amplitude (pV)
Number of FG-labeled cells
PIP2P3P4Pl-NlP2-N2P3-N3P4-N4RNSMC
1.4 :
2.3 :
3.0 :
3.9 :
6.810.214.26.5
2,81314,060
0.0 (n = 4)0.1 (n = 4)0.1 (n = 4)0.2 (n = 4)2.6 (n = 4)1.1 (n = 4)1.3 (n = 4)1.4 (n = 4)
: 286 (n = 5): 1,096 (n = 5)
1.5 ± 0.12.1 ± 0.13.0 ± 0.13.9 ± 0.23.2 ± 2.1
15.3 ± 0.314.1 ± 3.45.1 ± 1.8
1,962 ± 1,0162,693 ± 2,687
The normal control animals had no surgical procedures prior to FG introduction. The vehicle infusion group had no spinal cordinjury, but had vehicle infusion for 14 days into the spinal cord through a cannula implanted at T3 and then survived an additional14 days. In both groups, the spinal cord was transected at T10 for FG introduction 72 h prior to MEP measurement. MEPs were
recorded from the dorsal surface of the spinal cord at T7.
though there was no significant difference in the numberof FG-labeled cells in the RN between these groups (Table1). Histologically, there was evidence of mild injury tothe dorsal columns including the corticospinal tracts at thesite of cannula implantation. There was mild disruptionof tissue along the tract of the cannulae and an adjacentchronic inflammatory reaction including hemosiderinladen macrophages, although there was no cavitation.
Motor Function After InjuryAfter the 35-g clip injury at T3, the vehicle-treated an-
imals (vehicle control, n = 6) showed severe hindlimbparesis indicated by an inclined plane score of 36 ± 2°at 1 week after injury, which then recovered slightly toa final inclined plane score of 43 ± 2° at 4 weeks (Fig.1). The BDNF-treated animals (n = 6) showed a signif-icantly higher inclined plane score of 45 ± 3° at 1 weekcompared to the vehicle control (p < 0.05). Although theinclined plane scores of the BDNF-treated animals were
not significantly different at the later time intervals afterinjury, these animals showed a higher inclined planescore than the vehicle control animals at all time inter-vals after injury, and the final score at 4 weeks was 49 ±2°. The inclined plane scores of the NT-3-treated or
NGF-treated animals (n = 6, respectively) were not sig-nificantly different from the vehicle control animals at
any time interval after injury.The electrophysiological assessment using MEP at 4
weeks after injury failed to record any responses fromthe spinal cord caudal to the injury site in any animal(data not shown).
Retrograde Fluorogold TracingThe number of retrogradely labeled neurons in RN
and SMC with FG is a quantitative measure of the per-sistence of rubrospinal and corticospinal axons, re-
spectively, at the lesion site after SCI. Labeled cells inthe injured animals were seen in both the magnocellu-lar and parvicellular portions of the RN. The BDNF-treated animals suggested higher number of labeledcells in the RN (1,638 ± 350) and SMC (124 ± 83)compared with the vehicle control, however the differ-ences were not significant. Representative examples ofthe labeled cells in the RN are shown in Figure 3. Thenumber of labeled cells in the SMC in the BDNF-treated animals (Fig. 2) was small, with only 0.9% ofthe number seen in the uninjured controls (Table 1).The number of labeled cells in the NT-3-treated or
NGF-treated animals were also not significantly dif-ferent from the vehicle control, in either the RN or
SMC. The NT-3-treated animals had a lower numberof labeled cells (673 ± 150 in RN, 12 ± 9 in SMC)than the vehicle-treated animals, and the difference inthe numbers of retrogradely labeled rubrospinal neu-
rons between the NT-3-treated and BDNF-treated an-
imals was significant (Fig. 2). The NGF-treated ani-mals had similar numbers of labeled cells (1,288 ± 129in RN, 22 ± 3 in SMC) to the vehicle control. In thevehicle control, the numbers of labeled cells after in-jury were 44% (1,228 ±217) in the RN and 0.3%(36 ± 15) in the SMC compared with the numbers inthe uninjured normal control (Fig. 2, for vehicle con-
trol and Table 1 for the uninjured normal control).
1223

NAMIKI ET AL.
oo
o u
1 &o« o•a -oS"oc
VehicleNT-3
pre-injury
2w
Time after 35g clip injury
4w
FIG. 1. Inclined plane scores after the 35-g clip injury for the four experimental groups. The neurotrophic factors or vehiclewere infused into the spinal cord at the injury site for 14 days after injury.* BDNF-treated group was significantly different fromthe vehicle control at 1 week, p < 0.05 (Anova followed by the post hoc Student-Newman-Keuls method).
Histological Evaluation
In the vehicle control, the lesions produced by the 35-g clip injury were characterized by a major degree of necro-sis and cavity formation (Fig. 4). The cavities containedmacrophages with ingested myelin and hemosiderin. Thesefeatures were prominent at the epicenter of the lesion. Thelesion cavities extended rostrally and caudally for severalmm, especially in the central gray matter and dorsal hornsand in the central aspect of the dorsal funiculus. A rim ofresidual tissue was usually observed near the pial surfaceespecially ventrally and consisted mainly of white matter.Residual gray matter was infrequently present at the tip ofthe dorsal horns or in the most ventral aspects of the ven-
tral horns (Fig. 4). These morphological changes in thechronic phase are consistent with those previously reportedafter clip compression injury of the rat spinal cord (Midhaet al, 1987; Wallace et al, 1987; Tator, 1995, 1998;Agrawal and Fehlings, 1997; Koyanagi and Tator, 1997).A defect indicating the location of the implanted cannulatip was observed in the dorsal residual white matter in mostof the animals, and large cavities extended rostrally andcaudally from the defect presumably due to the cannulaand intramedullary infusion. Although we did not includean injured control group with an implanted cannula with-out infusion, it is our impression that these cavities were
larger than those usually seen after a 35-g clip compres-sion injury.
To examine whether intramedullary infusion of theneurotrophic factors prevented loss of neural structuresafter SCI, the proportion of the cavitated area to the to-
tal cross-sectional area at the epicenter of the injury sitewas measured 4 weeks after the 35-g clip injury. In thevehicle control, the percent cavitation at the lesion sitewas 32.3 ± 4.8% (n = 6). In the neurotrophic factor-treated animals, the percent cavitation at the lesion sitewere 30.7 ±5.1% after NT-3, 21.4 ± 4.3% after BDNF,and 24.7 ± 4.3% after NGF (n = 6 per group). Althoughthere was a suggestion of less cavitation in the neu-
rotrophic factor-treated animals, especially after BDNF,compared with the control animals, the differences were
not statistically significant.Many of the neurotrophic factor-treated animals, es-
pecially the BDNF group, showed more invasion and pro-liferation of Schwann cells and greater formation of pe-ripheral myelin which stained blue with LFB and PAScompared with the control animals (Table 2, and Figs. 4and 5). The difference in the invasion of Schwann cellsbetween the neurotrophic factor-treated animals and thecontrol animals was significant (p < 0.05), although theincrease in peripheral myelin formation was not signifi-cant. In the BDNF group, three of the six rats showed a
remarkable degree of Schwann cells and peripheralmyelin formation (Fig. 5). The Schwann cells invadedmainly from the dorsal roots and proliferated in the ar-
eas of necrosis and cavitation among the macrophages,and ultimately filled the cavity in some areas. None ofthe groups showed any new central myelin formation.Masson's trichrome stain showed collagen fibers aroundthe site of the cannulae, but fibroblasts were not presentaround the cavities in any group. Although there were
many GFAP-positive cells in the adjacent preserved tis-
1224

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
2000 ,
co Mo xr,_ Cl« Öü «
ta
-5 îooo J
Vehicle NT-3 BDNF NGF(n=6) (n=6) (n=6) (n=6)
B
300
200 ioo .SC D.
3 00S £ 100
0 €i IVehicle
(n-6)NT-3
(n=6)BDNF
(n=6)NGF
(n=6)
FIG. 2. Number of FG-labeled cells in red nucleus (RN); (A)and sensorimotor cortex (SMC); (B) 4 weeks after the 35-g clipinjury. Neurotrophic factors or vehicle were infused into the in-jury site for 14 days after injury. The numbers in parenthesesindicate the total number of animals in each group. There was
no significant difference between vehicle-treated and each neu-
rotrophic factor-treated group (Anova followed by the post hocStudent-Newman-Keuls method).
sue, there were only a small number of GFAP-positivecells in the central area of the injured cord. The findingswith GFAP and Masson's trichrome stain were similarin all four groups.
DISCUSSION
The present study demonstrates that the intramedullaryinfusion of BDNF significantly improved clinical neuro-
logical function as shown by the inclined plane score 1
week after injury compared with vehicle control (p <0.05), with the suggestion of a higher number of retro-gradely labeled cells in the RN and SMC at 4 weeks af-ter injury compared to the vehicle control (although theselatter differences did not reach statistical significance).Possible explanations for this improvement include thepossibility that BDNF prevented some of the damage toless severly injured axons and cell bodies or preventedthe death of some of the severly injured neurons. It is ofinterest that functional improvement based on the 21-point locomotor scale (BBB rating scale) was also shownby Kim et al. (1996), who reported that both BDNF andNGF secreted by genetically modified fibroblasts placedat the injury site accelerated recovery of motor function,and similar to the present study, the improvement was
significant only at 1 week after a contusion injury. Jake-man et al. (1998) reported that BDNF delivered in-trathecally with an osmotic pump at T8 near the lesionsite improved BBB scores at 15 days after mild injury,at 3 days after moderate injury, and at 8, 15, and 22 daysafter complete transection. They also suggested that therewas a general decrease in all the behavioral effects after2 weeks. Liu et al. (1999) also used transplants of ge-netically modified fibroblasts expressing BDNF andshowed that behavioral improvement was present by 1week after treatment and reached a plateau between 3 and4 weeks in rats with hemisection of the cervical cord. Asnoted above, Kim et al. (1996) also found that NGF ex-erted a similar neuroprotective effect on motor functionafter spinal cord contusion, which contrasts with the lackof a neuroprotective effect with NGF in the present study.This lack of effectiveness of NGF and the discrepancybetween BDNF and NGF in neuroprotective effects was
also found by Kobayashi et al. (1997) with respect to pre-vention of atrophy of axotomized rubrospinal neurons.
Thus, the neuroprotective effect of NGF, if any, on theinjured adult spinal cord appears to be less than BDNF.
In the present study, the intramedullary infusion ofBDNF into the SCI lesion site also induced a remarkableproliferation of Schwann cells and production of periph-eral myelin, and a concomitant decrease in the size of thecavities at the lesion site compared to NGF and NT-3, al-though these differences were not significant. It is wellrecognized that Schwann cells migrate from nerve roots,infiltrate into the lesion site, and produce abundant pe-ripheral type myelin after SCI (Lampert and Cressman,1964; Gilmore and Duncan, 1968; Hirano et al., 1969;Blakemore and Patterson, 1975; Harrison et al., 1975;Blakemore, 1977; Griffiths and McCulloch, 1983; Wal-lace et al., 1987; Blight and Young, 1989; West andCollins, 1989; Bunge et al., 1994; Beattie et al., 1996; Sal-gado-Ceballos et al., 1998; Tator 1998; Brook et al.,1998). The vehicle-treated animals in the present study
1225
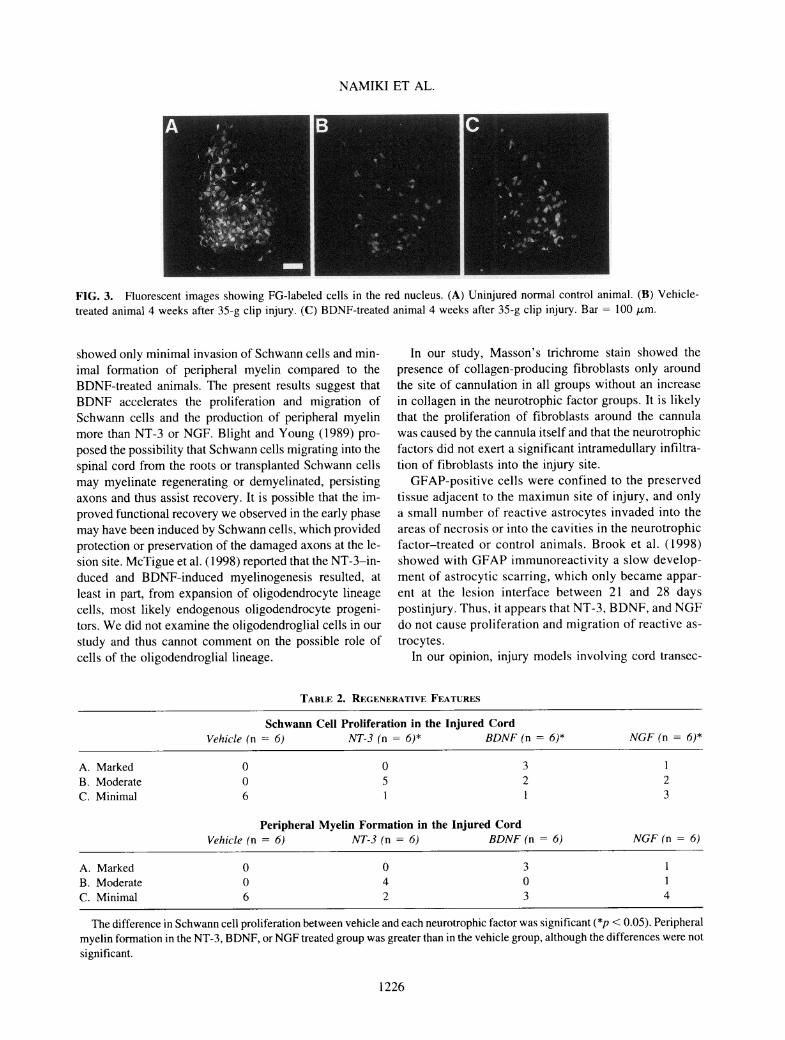
NAMIKI ET AL.
FIG. 3. Fluorescent images showing FG-labeled cells in the red nucleus. (A) Uninjured normal control animal. (B) Vehicle-treated animal 4 weeks after 35-g clip injury. (C) BDNF-treated animal 4 weeks after 35-g clip injury. Bar = 100 pm.
showed only minimal invasion of Schwann cells and min-imal formation of peripheral myelin compared to theBDNF-treated animals. The present results suggest thatBDNF accelerates the proliferation and migration ofSchwann cells and the production of peripheral myelinmore than NT-3 or NGF. Blight and Young (1989) pro-posed the possibility that Schwann cells migrating into thespinal cord from the roots or transplanted Schwann cellsmay myelinate regenerating or demyelinated, persistingaxons and thus assist recovery. It is possible that the im-proved functional recovery we observed in the early phasemay have been induced by Schwann cells, which providedprotection or preservation of the damaged axons at the le-sion site. McTigue et al. (1998) reported that the NT-3-in-duced and BDNF-induced myelinogenesis resulted, atleast in part, from expansion of oligodendrocyte lineagecells, most likely endogenous oligodendrocyte progeni-tors. We did not examine the oligodendroglial cells in our
study and thus cannot comment on the possible role ofcells of the oligodendroglial lineage.
In our study, Masson's trichrome stain showed thepresence of collagen-producing fibroblasts only aroundthe site of cannulation in all groups without an increasein collagen in the neurotrophic factor groups. It is likelythat the proliferation of fibroblasts around the cannulawas caused by the cannula itself and that the neurotrophicfactors did not exert a significant intramedullary infiltra-tion of fibroblasts into the injury site.
GFAP-positive cells were confined to the preservedtissue adjacent to the maximun site of injury, and onlya small number of reactive astrocytes invaded into theareas of necrosis or into the cavities in the neurotrophicfactor-treated or control animals. Brook et al. (1998)showed with GFAP immunoreactivity a slow develop-ment of astrocytic scarring, which only became appar-ent at the lesion interface between 21 and 28 dayspostinjury. Thus, it appears that NT-3, BDNF, and NGFdo not cause proliferation and migration of reactive as-
trocytes.In our opinion, injury models involving cord transec-
Table 2. Regenerative Features
Schwann Cell Proliferation in the Injured CordVehicle (n = 6) NT-3 (n = 6)* BDNF (n = 6)* NGF (n = 6)*
A. MarkedB. ModerateC. Minimal
0o6
Peripheral Myelin Formation in the Injured CordVehicle (n = 6) NT-3 (n = 6) BDNF (n = 6) NGF (n = 6)
A. MarkedB. ModerateC. Minimal
042
303
The difference in Schwann cell proliferation between vehicle and each neurotrophic factor was significant (*p < 0.05). Peripheralmyelin formation in the NT-3, BDNF, or NGF treated group was greater than in the vehicle group, although the differences were not
significant.
1226

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
FIG. 4. Photomicrographs of cross-sec-
tions of the SCI site of a vehicle control ratstained with H&E/LFB. In this animal, thenecrosis and cavity formation were seen 4weeks after SCI, and the proliferation ofSchwann cells and myelin formation were
not prominent compared with Fig. 5 (orig-inal magnification, X32).
FIG. 5. Photomicrographs of a cross-
section of the SCI site in a BDNF-treated animal at 4 weeks stained withH&E/LFB. There was extensive myelinformation, which stained blue with LFBcombined with PAS. The site of maxi-mum injury was almost completelyfilled with Schwann cells, and there was
no cavitation. The box in A indicates thelocation of B. Original magnifications,X32 (A), X400 (B).
ë <¡r\ *> w«^
jeWk^fíÉkíka\í^^aaMmr
1227

NAMIKI ET AL.
tion or hemisection are less suitable than the clip com-
pression model for studying the neuroprotective effects ofneurotrophic factors for producing functional improve-ment. Accordingly, we used a 35-g clip injury as a modelof moderate spinal cord contusion to detect functional im-provement in the neurotrophic factor treated animals. Thespinal cord clip compression model probably generatesmore ischemia than the transection or hemisection mod-els. The severity of clip compression we selected wasabove the threshold for causing unequivocal hindlimbparesis as detected by the inclined plane score (Fehlingsand Tator, 1995). In the vehicle control, the mean score4 weeks after injury (43 ± 2°) represented a 40% reduc-tion in the score observed in uninjured normal animals(Fig. 1). The present study failed to show any MEP re-
sponses in the neurotrophic factor-treated animals, indi-cating that there was insufficient conduction along motorpathways through the spinal cord lesion 4 weeks after in-jury to evoke any MEP responses. In previous studies, our
laboratory found that MEP responses were completelyabolished by a 56-g clip injury, while responses were de-tectable after a 20-g clip injury (Fehlings et al, 1987,1989). Thus, in the present experiment, the recording ofMEPs was too insensitive to detect a small amount ofresidual or recovered motor function.
Intramedullary delivery of the neurotrophic factors intothe lesion site was accomplished through a stainless steelcannula implanted into the injured spinal cord. The un-
injured animals with cannula implantation and vehicle in-fusion into the spinal cord showed incline plane and MEPresults similar to nonimplanted normal animals. How-ever, the number of retrogradely labeled corticospinalneurons was significantly decreased compared to the nor-
mal controls. Thus, it is highly likely that the deliverysystem caused significant damage to the corticospinaltracts. The 34-gauge cannula had an outer diameter ofapproximately 180 pm and was implanted 1 mm in depth,which is the approximate depth of the corticospinal tracts.It is likely that the implanted cannula itself caused some
damage, but that, in addition, the continuous infusion ofvehicle or the neurotrophic factors into the spinal cord ata flow rate of 0.5 pL/h for 14 days created additionaldamage and cavity formation which extended rostrallyand caudally from the cannula tip. The cavities also ap-peared to dissect the spinal cord tissue. In in vitro pilotstudies, we tested a smaller cannula of 36 gauge, withouter diameter of approximately 110 pm, but there waseither occlusion or high resistance to infusion of the neu-
rotrophic factors at a flow rate of 0.5 ph/h. Subarach-noid administration of neurotrophic factors would avoidthe tissue damage caused by the intramedullary infusion,and this route has been used experimentally (Verge et al,1990; Gold et al, 1993, 1997; Novikova et al, 1996,
1997; Novikov et al, 1997; Kishino et al, 1997; Jake-man et al, 1998) and in patients (Penn et al, 1977) todeliver neurotrophic factors to the spinal cord. However,it is not known whether similar beneficial results wouldhave been observed.
The neurotrophic factors were infused at a dose of 625ng/h, which totaled 210 pg for the 14-day infusion pe-riod. This dose was based on previous studies of the con-tinuous administration of neurotrophic factors with an os-motic pump (Fernandez et al, 1993; Xu et al, 1995;Novikova et al, 1996; Kobayashi et al, 1997). Fernan-dez et al. (1993) described no dose-response relation be-tween doses of epidural NGF infusion (total amount,100-500 pg) and effects on axonal regeneration afterspinal cord transection. Perhaps, a lower dose of neu-
rotrophic factors might have been sufficient for in-tramedullary infusion compared with epidural (Fernandezet al, 1993) or subarachnoid administration (Novikova etal, 1996). Thus, it is possible that the dose used in thepresent study could have been reduced, although, with theavailable evidence, it is difficult to determine the optimaldose for the administration of neurotrophic factors.
While there is evidence for the neuroprotective effectof BDNF on SCI, NT-3 has been found to promote ax-
onal regeneration into a peripheral nerve graft after SCIin adult animals (Ye and Houle, 1997), into a transplantof fetal spinal cord tissue (Bregman et al, 1997), or intoa graft of NT-3-secreting fibroblasts (Grill et al, 1997b;McTigue et al, 1998). NT-3 has also produced regener-ation of corticospinal tract axons in the spinal cord cau-dal to a lesion in the presence of an antibody againstmyelin-associated neurite growth inhibitory proteins(Schnell et al, 1994). Bradbury et al. (1998) showed thatNT-3 delivered continuously to the SCI lesion site via anosmotic pump promoted cell survival and completely re-versed cell atrophy in ascending systems. There are twostudies of the combined administration of BDNF and NT-3 in SCI (Xu et al, 1995; Bregman et al, 1997), althoughthe value of the combination has not been clarified. In-terestingly, the present study showed that the cell countsof retrogradely labeled rubrospinal and corticospinal neu-
rons in the NT-3-treated animals were lower than in thevehicle control, and the difference in the numbers of ret-rogradely labeled rubrospinal neurons between the NT-3-treated and BDNF-treated animals was significant.These findings suggest that NT-3 may have a toxic ef-fect on the repair of the rubrospinal tract. Furthermore,there is no report of a neuroprotective effect of NT-3 inthe injured adult spinal cord. In summary, NT-3 admin-istrated for the first 14 days after injury may not be use-ful for neuroprotection and the prevention of motor tract
damage after SCI in adults. Perhaps, a programmed, se-
quential delivery of neurotrophic factors, for example
1228

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
first BDNF for neuroprotection, then NT-3 for regener-ation, may be a useful approach to the treatment of SCI.
ACKNOWLEDGMENTS
This work was supported by the Canadian Spinal Re-search Organization, the Samuel Lunenfeld Foundation,and Regeneran Pharmaceuticals. We thank JamesLoukides, Linda Lee, Rita van Bendegem, and JoanneClark for excellent technical assistance, and Dr. PatrickShannon of the Division of Neuropathology, TorontoWestern Hospital, for valuable suggestions for the im-munohistochemistry.
REFERENCES
AGRAWAL, S.K., and FEHLINGS, M.G. (1997). The effectof the sodium channel blocker QX-314 on recovery afteracute spinal cord injury. J. Neurotrauma 14, 81-88.
ANDERSON, D.K., and HALL, E.D. (1993). Pathophysiologyof spinal cord trauma. Ann. Emerg. Med. 22, 987-992.
BEATTIE, M.S., BRESNAHAN, J.C, KOMON, J., et al.(1997). Endogenous repair after spinal cord contusion in-juries in the rat. J. Neurotrauma. 148, 453^463.
BLAKEMORE, W.F. (1977). Remyelination of CNS axons bySchwann cells transplanted from the sciatic nerve. Nature(Lond.) 266, 68-69.
BLAKEMORE, W.F., and PATTERSON, R.C (1975). Obser-vations on the interactions of Schwann cells and astrocytesfollowing X-irradiation of neonatal rat spinal cord. J. Neu-rocytol. 4, 573-585.
BLIGHT, A.R., and YOUNG, W. (1989). Central axons in in-jured cat spinal cord recover electrophysiological functionfollowing remyelination by Schwann cells. J. Neurol. Sei. 91,15-34.
BRADBURY, E.J., KING, V.R., SIMMONS, L.J., et al. (1998).NT-3, but not BDNF, prevents atrophy and death of axo-
tomized spinal cord projection neurons. Eur. J. Neurosci. 10,3058-3068.
BREGMAN, B.S., McATEE, M., DAI, H.N., et al. (1997). Neu-rotrophic factors increase axonal growth after spinal cord in-jury and transplantation in the adult rat. Exp. Neurol. 148,475^94.
BROOK, G.A., PLATE, D., FRANZEN, R., etal. (1998). Spon-taneous longitudinally orientated axonal regeneration is as-
sociated with the Schwann cell framework within the lesionsite following spinal cord compression injury of the rat. J.Neurosci. 53, 51-65.
BUNGE, M.B., HOLETS, V.R., BATES, M.L., et al. (1994).Characterization of photochemically induced spinal cord in-
jury in the rat by light and electron microscopy. Exp. Neu-rol. 127, 76-93.
DIENER, P.S., and BREGMAN, B.S. (1994). Neurotrophic fac-tors prevent the death of CNS neurons after spinal cord le-sions in newborn rats. Neuroreport 5, 1913-1917.
FEHLINGS, M.G., and TATOR, C.H. (1992). The effect of di-rect current field polarity on recovery after acute experi-mental spinal cord injury. Brain Res. 579, 32-42.
FEHLINGS, M.G., and TATOR, C.H. (1995). The relationshipsamong the severity of spinal cord injury, residual neurolog-ical function, axon counts, and counts of retrogradely labeledneurons after experimental spinal cord injury. Exp. Neurol.132, 220-228.
FEHLINGS, M.G., TATOR, C.H., LINDEN, R.D., et al.(1987). Motor evoked potentials recorded from normal andspinal cord-injured rats. Neurosurgery 20, 125-130.
FEHLINGS, M.G., TATOR, C.H., and LINDEN, R.D. (1989).The relationships among the severity of spinal cord injury,motor and somatosensory evoked potentials and spinal cordblood flow. Electroencephalogr. Clin. Neurophysiol. 74,241-259.
FEIGIN, I., and CRAVIOTO, H. (1961). A histochemical studyof myelin. A difference in the solubility of the glycolipidcomponents in the central and peripheral nervous systems. J.Neuropathol. Exp. Neurol. 20, 245-254.
FERNANDEZ, E., PALLINI, R., LAURETTI, L., et al. (1993).Spinal cord transection in adult rats: effects of local infusionof nerve growth factor on the corticospinal tract axons. Neu-rosurgery 33, 889-893.
GIEHL, K.M., and TETZLAFF, W. (1996). BDNF and NT-3,but not NGF, prevent axotomy-induced death of rat corti-cospinal neurons in vivo. Eur. J. Neurosci. 8, 1167-1175.
GILMORE, S.A., and DUNCAN, D. (1968). On the presenceof peripheral-like nervous and connective tissue within irra-diated spinal cord. Anat. Rec. 160, 675-690.
GOLD, B.G. (1997). Axonal regeneration of sensory nerves isdelayed by continuous intrathecal infusion of nerve growthfactor. Neuroscience 76, 1153-1158.
GOLD, B.G., STORM-DICKERSON, T., and AUSTIN, D.R.(1993). Regulation of the transcription factor c-JUN by nerve
growth factor in adult sensory neurons. Neurosci. Lett. 154,129-133.
GRIFFITHS, I.R., and McCULLOCH, M.C. (1983). Nerve fi-bres in spinal cord impact injuries. Part 1. Changes in themyelin sheath during the initial 5 weeks. J. Neurol. Sei. 58,335-349.
GRILL, R.J., BLESCH, A., and TUSZYNSKI, M.H. (1997a).Robust growth of chronically injured spinal cord axons in-duced by grafts of genetically modified NGF-secreting cells.Exp. Neurol. 148, 444-452.
GRILL, R., MURAI, K, BLESCH, A., et al. (1997b). Cellular
1229

NAMIKI ET AL.
delivery of neurotrophin-3 promotes corticospinal axonalgrowth and partial functional recovery after spinal cord in-jury. J. Neurosci. 17, 5560-5572.
HARRISON, B.M, GLEDHILL, R.F, and McDONALD, W.I.(1975). Remyelination after transient compression of thespinal cord. Proc. Aust. Assoc. Neural. 12, 117-122.
HEBEL, R, and STROMBERG, M.W. (1986). Anatomy andEmbryology of the Laboratory Rat. BioMed Verlag:Günzburg, pps. 68-69.
HIRAIZUMI, Y, FUJIMAKI, E, TRANSFELDT, E.E, et al.(1996). The effect of the platelet derived wound healing for-mula and the nerve growth factor on the experimentally in-jured spinal cord. Spinal Cord 34, 394-402.
HIRANO, A, ZIMMERMAN, H.M, and LEVINE, S. (1969).Electron microscopic observations of peripheral myelin in a
central nervous system lesion. Acta Neuropathol. (Berl.) 12,348-365.
HOWELING, D.A, VAN ASSELDONK, J.T.H, LANK-HORST, A.J, et al. (1998a). Local application of collagencontaining brain-derived neurotrophic factor decreases theloss of function after spinal cord injury in the adult rat. Neu-rosci. Lett. 251, 193-196.
HOWELING, D.A, LANKHORST, A.J, GISPEN, W.H, etal. (1998b). Collagen-containing neurotrophin-3 (NT-3) at-tract regrowing injured corticospinal axons in the adult rat
spinal cord and promotes partial functional recovery. Exp.Neural. 153, 49-59.
JAKEMAN, L.B, WEI, P, GUAN, Z, et al. (1998). Brain-de-rived neurotrophic factor stimulates hindlimb stepping andsprouting of cholinergic fibers after spinal cord injury. Exp.Neural. 154, 170-184.
KIM, D.H., GUTIN, P.H, NOBLE, L.J, et al. (1996). Treat-ment with genetically engineered fibroblasts producing NGFor BDNF can accelerate recovery from traumatic spinal cordinjury in the adult rat. Neuroreport 7, 2221-2225.
KISHINO, A, ISHIGE, Y, TATSUNO, T, et al. (1997). BDNFprevents and reverses adult rat motor neuron degenerationand induces axonal outgrowth. Exp. Neural. 144, 273-286.
KOBAYASHI, N.R, FAN, D.-P, GIEHL, K.M., et al. (1997).BDNF and NT-4/5 prevent atrophy of rat rubrospinal neu-
rons after cervical axotomy, stimulate GAP-43 and Tal-Tubulin mRNA expression, and promote axonal regenera-tion. J. Neurosci. 17, 9583-9595.
KOYANAGI, I, and TATOR, C.H. (1997). Effect of a singlehuge dose of methylprednisolone on blood flow, evoked po-tentials, and histology after acute spinal cord injury in therat. Neural. Res. 19, 289-299.
LAMPERT, P, and CRESSMAN, M. (1964). Axonal regener-ation in the dorsal columns of the spinal cord of adult rats.An electron microscopic study. Lab. Invest. 13, 825-839.
LEE, T.T., GREEN, B.A., DIETRICH, W.D, et al. (1999).Neuroprotective effects of basic fibroblast growth factor fol-
lowing spinal cord contusion injury in the rat. J. Neurotrauma16, 347-356.
LIU, Y, KIM, D, HIMES, B.T, et al. (1999). Transplants offibroblasts genetically modified to express BDNF promoteregeneration of adult rat rubrospinal axons and recovery offorelimb function. J. Neurosci. 19, 4370-4387.
McTIGUE, D.M., HORNER, P.J, STOKES, B.T, et al. (1998).Neurotrophin-3 and brain-derived neurotrophic factor induceoligodendrocyte proliferation and myelination of regenerat-ing axons in the contused adult rat spinal cord. J. Neurosci.18, 5354-5365.
MENEI, P, MONTERO-MENEI, C, WHITTEMORE, S.R, etal. (1998). Schwann cells genetically modified to secret hu-man BDNF promotes enhanced axonal regrowth across tran-sected adult rat spinal cord. Eur. J. Neurosci. 10, 607-621.
MIDHA, R, FEHLINGS, M.G, TATOR, C.H, et al. (1987).Assessment of spinal cord injury by counting corticospinaland rubrospinal neurons. Brain Res. 410, 299-308.
NAKAHARA, Y, GAGE, F.H, and TUSZYNSKI, M.H.(1996). Grafts of fibroblasts genetically modified to secrete
NGF, BDNF, NT-3, or basic FGF elicit differential responsesin the adult spinal cord. Cell Transplant. 5, 191-204.
NAMIKI, J, and TATOR, C.H. (1999). Cell proliferation andnestin expression in the ependyma of the adult rat spinal cordafter injury. J. Neuropathol. Exp. Neurol. 58, 489^198.
NOVIKOV, L, NOVIKOVA, L, and KELLERTH, J.-O.(1997). Brain-derived neurotrophic factor promotes axonalregeneration and long-term survival of adult rat spinal mo-
toneurons in vivo. Neuroscience 79, 765-774.
NOVIKOVA, L, NOVIKOV, L, and KELLERTH, J.-O.(1996). Brain-derived neurotrophic factor reduces necroticzone and supports neuronal survival after spinal cord hemi-section in adult rats. Neurosci. Lett. 220, 203-206.
NOVIKOVA, L, NOVIKOV, L, and KELLERTH, J.-O.(1997). Effects of neurotransplants and BDNF on the sur-
vival and regeneration of injured adult spinal motoneurons.Eur. J. Neurosci. 9, 2774-2777.
OUDEGA, M, and HAGG, T. (1996). Nerve growth factor pro-motes regeneration of sensory axons into adult rat spinal cord.Exp. Neurol. 140, 218-229.
PARK, K.I, LIU, S, FLAX, J.D, et al. (1999). Transplanta-tion of neural progenitor and stem cells: Developmental in-sights may suggest new therapies for spinal cord and otherCNS dysfunction. J. Neurotrauma 16, 675-687.
PAXINOS, G, and WATSON, C. (1986). The Rat Brain inStereotaxic Coordinates, 2nd ed. Academic Press: San Diego.
PENN, R.D, KROIN, J.S, YORK, M.M, et al. (1997). In-trathecal ciliary neurotrophic factor delivery for treatment ofamyotrophic lateral sclerosis (phase 1 trial). Neurosurgery40, 94-100.
RIVLIN, A.S, and TATOR, C.H. (1978). Effect of duration of
1230

NEUROTROPHIC FACTORS FOR SPINAL CORD INJURY
acute spinal cord compression in a new acute cord injurymodel in the rat. Surg. Neurol. 10, 39^13.
SALGADO-CEBALLOS, H, GUIZAR-SAHAGUN, G., FE-RIA-VELASCO, A., et al. (1998). Spontaneous long-term re-
myelination after traumatic spinal cord injury in rats. BrainRes. 782, 126-135.
SCHNELL, L., SCHNEIDER, R„ KOLBECK, R., et al. (1994).Neurotrophin-3 enhances sprouting of corticospinal tract dur-ing development and after adult spinal cord lesion. Nature367, 170-173.
TATOR, C.H. (1995). Update on the pathophysiology andpathology of acute spinal cord injury. Brain Pathol. 5,407^tl3.
TATOR, C.H. (1998). Biology of neurological recovery andfunctional restoration after spinal cord injury. Neurosurg. 42,696-708.
TATOR, C.H., and FEHLINGS, M.G. (1991). Review of thesecondary injury theory of acute spinal cord trauma with em-
phasis on vascular mechanisms. J. Neurosurg. 75, 15-26.
TETZLAFF, W., KOBAYASHI, N.R., GIEHL, K.M.G., et al.(1994). Response of rubrospinal and corticospinal neurons to
injury and neurotrophin. Prog. Brain Res. 103, 271-286.
THERIAULT, E., and TATOR, C.H. (1994). Persistence ofrubrospinal projections following spinal cord injury in therat. J. Comp. Neurol. 342, 249-258.
TUSZYNSKI, M.H., GABRIEL, K, GAGE, F.H., et al. (1996).Nerve growth factor delivery by gene transfer induces differ-ential outgrowth of sensory, motor, and noradrenergic neu-
rites after adult spinal cord injury. Exp. Neurol. 137,157-173.TUSZYNSKI, M.H., MURAI, K, BLESCH, A., et al. (1997).
Functional characterization of NGF-secreting cell grafts tothe acutely injured spinal cord. Cell Transplant. 6, 361-368.
VERGE, V.M.K., TETZLAFF, W., BISBY, M.A., et al. (1990).Influence of nerve growth factor on neurofilament gene ex-
pression in mature primary sensory neurons. J. Neurosci. 10,2018-2025.
WALLACE, M.C, TATOR, C.H., and LEWIS, A.J. (1987).Chronic regenerative changes in the spinal cord after cordcompresión injury in rats. Surg. Neurol. 27, 209-219.
WEIDNER, N., BLESCH, A., GRILL, R.J., et al. (1999). Nervegrowth factor-hypersecreting Schwann cell grafts augmentand guide spinal cord axonal growth and remyelinate centralnervous system axons in a phenotypically appropriate man-
ner that correlates with expression of LI. J. Comp. Neurol.413, 495-506.
WEST, N.R., and COLLINS, G.H. (1989). Cellular changesduring repair of a cryogenic spinal cord injury in the rat: an
electron microscopic study. J. Neuropathol. Exp. Neurol. 48,94-108.
WHITTEMORE, S.R. (1999). Neuronal replacement strategiesfor spinal cord injury. J. Neurotrauma 16, 667-673.
XU, X.M., GUÉNARD, V., KLEITMAN, N., et al. (1995). Acombination of BDNF and NT-3 promotes supraspinal ax-onal regeneration into Schwann cell grafts in adult rat tho-racic spinal cord. Exp. Neurol. 134, 261-272.
YE, J.H., and HOULE, J.D. (1997). Treatment of the chroni-cally injured spinal cord with neurotrophic factors can pro-mote axonal regeneration from supraspinal neurons. Exp.Neurol. 143, 70-81.
YICK, L.-W., WU, W., SO, K.-F., et al. (1999). Peripheralnerve graft and neurotrophic factors enhance neural survivaland expression of nitric oxide synthase in Clarke's nucleusafter hemisection of the spinal cord in adult rat. Exp. Neu-rol. 159, 131-138.
ZHANG, Y., DIJKHUIZEN, P.A., ANDERSON, P.N., et al.(1998). NT-3 delivered by an adenoviral vector induces in-jured dorsal root axons to regenerate into the spinal cord ofadult rats. J. Neurosci. Res. 54, 554-562.
Address reprint requests to:Charles H. Tator, M.D., Ph.D.
Toronto Western Hospital399 Bathurst St.
MP 2-435Toronto, Ontario M5T 2S8, Canada
E-mail: [email protected]
1231