
Review Article
Resident Tissue Leukocytes: Underappreciated Guards of the Body
Department of Evolutionary Immunobiology, Jagiellonian University, ul. Ingardena 6, 30-060 Krakow, Poland Elzbieta Kolaczkowska
Corresponding Author & Address:
Department of Evolutionary Immunobiology, Jagiellonian University, ul. Ingardena 6, 30-060 Krakow, Poland; Email:
Elzbieta Kolaczkowska
[email protected] Published: 25th May, 2010 Accepted: 25th May, 2010 Received: 19th March, 2010 Revised: 12th May, 2010 Open Journal of Hematology, 2010, 1-1 © Kolaczkowska E.; licensee Ross Science Publishers ROSS Open Access articles will be distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution, and reproduction in any medium, provided that the original work will always be cited properly. In memory of Jacek Kołaczkowski
ABSTRACT
Inflammatory reactions occurring in different milieu follow common phases with major highlights being local changes in vascular permeability and influx of inflammatory leukocytes that eliminate the source of inflammation while subsequent resolution is completed by tissue repair. The main inflammatory cells consist of blood-derived neutrophils and macrophages that arrive to the inflammatory site in response to a chemotactic signal. However, their arrival to the loci is initiated by a special set of leukocytes already present in various places of the body such as skin or body cavities that are called tissue or resident leukocytes. The cells consist of mast cells, macrophages and lymphocytes. Although the presence of resident leukocytes is commonly recognized their actual involvement in subsequent stages of inflammation is underappreciated while the cells are critical for all phases of the reaction. The current mini-review describes the role of resident leukocytes in the initiation, the course, and the termination of exemplary acute inflammatory reaction, namely zymosan-induced peritonitis. Additionally, facts on human resident peritoneal leukocytes are presented and discussed.
Inflammation is universally defined as a reaction of the body to injury or infection of vascularized tissues with the purpose to deliver defensive materials, such as blood cells and fluid, to the site of injury/infection [1]. The traces of understanding of this process can be found dating back as far as 2700 B.C., and the four cardinal signs of inflammation (redness, swelling, heat and pain) were put in written form already in the first
century A.D. by the physician Cornelius Celsus [1]. By now the mechanisms behind the signs of inflammation are generally known and many aspects of the inflammatory machinery are explained, yet not all. While the role of neutrophils and monocyte-derived macrophages newly infiltrating the sites of inflammation is well recognized and widely studied, the role of leukocytes residing in tissues prior to inflammation is often underestimated. This is
Open Journal of Hematology OPEN ACCESS

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 2 of 16 (Page number not for citation purposes)
especially inequitable as the cells are critical for the onset of the inflammatory reaction and, furthermore, play a significant role during the peak of the process and might have even impact on the resolution of the reaction.
RESIDENT LEUKOCYTES: LYMPHOCYTES, MACROPHAGES AND MAST CELLS
In certain vascularized tissues of the body that are exposed to the external environment such as skin, airways, gastrointestinal track, body cavities (e.g. pleural or peritoneal cavity) so called resident tissue leukocytes are located [1]. The cells migrate to the above tissues in physiological conditions and their function is to guard for any pathogens, allergens or other environmental agents (e.g. toxins) that might enter the body. Thus resident leukocytes provide the first line of cellular immune defense. Their composition and functions will be discussed on the example of peritoneal cavity that is located in the abdomen. Peritoneal cavity is a space between two layers of peritoneum that is the largest (with the area comparable to the skin) and most complexly arranged serous membrane of the body [2]. It contains a thin film of serous fluid, which is essentially an ultrafiltrate of plasma [3]. The fluid contains resident peritoneal leukocytes that in humans consist of approximately 70% macrophages, 10% B lymphocytes, 10% T lymphocytes, and 10% mast cells [4]. In mice the cellular composition is different with similar numbers of macrophages and lymphocytes (mostly B cells) and 2-4% of mast cells [5, 6]. The data on exact proportion of particular resident leukocyte populations in mice differs according to the applied identification methods e.g. detecting different surface markers of B cells (CD19, B220, CD5) [5-7]. However, there are clear strain differences in macrophage and B cell numbers e.g. peritoneum of Balb/c mice contains more B cells than macrophages in sharp contrast to NOD mice (the diabetes-prone strain) while other strains (e.g. C57Bl/6, CBA, CH/He, DBA1, NZW, MRL/Mp-Ipr) have similar numbers of both cell types [6].
B cells
Peritoneal cavity is populated by rather unique B lymphocyte populations. Commonly B cells are divided into (i) evolutionary conservative
B-1 cells rapidly producing T cell-independent low affinity antibodies, and (ii) more conventional (mostly follicular) B-2 cells producing T cell-dependent high affinity antibodies. In terms of surface phenotype they were initially showed to be CD5+ and CD5-, respectively [8]. However, the further studies of peritoneal cavity B cells revealed that at least some B-1 cells lack CD5 and for this B-1 cells were further subdivided into B-1a (CD5+B220+Mac-1+) and B-1b (CD5-B220+Mac-1+, “sister cells”) [9]. Therefore the phenotype criteria turned out not to be fundamental for distinguishing B-1 and B-2 cells. What further discriminates the two populations is their appearance in either fetal or adult life and for this it was recently proposed that the B cells that are readily generated from fetal/neonatal precursors (but poorly from adult precursors) are to be coined B-1 whereas the B cells generated throughout adult life - B-2 cells [10]. Peritoneal cavity is inhabited mostly by B-1 cells, however, B-2 cells are also present at this locus. Nevertheless, the peritoneal B-2 cells appear to be intermediate between splenic B-2 and peritoneal B-1b cells e.g. they require CD11b expression and ability to produce IgM antibodies as B-1 cells [11].
Fetal precursors of B-1 cells have unique characteristics such as lack of TdT (an enzyme responsible for inserting nucleotides to the VDJ exons during antibody gene recombination) and delayed expression of MHC II [10]. Moreover, the precise lineage restriction of early developing hemopoietic cells in fetal liver may differ from that in bone marrow and in fact a presence of novel B cell precursors was described. In particular, B/myeloid bipotential precursors were reported in fetal liver [12], but similar cells with a novel CD19+B220- phenotype were also detected in bone marrow [13]. In this regard this is not surprising that presence of biphenotypic CD5+B/macrophage cells was subsequently reported and until now in vivo they were observed only in peritoneum. The B/macrophage cells exhibit simultaneously characteristics of both B-1 lymphocytes (IgM, IgD, B220+, CD5+) and macrophages (phagocytosis, COX-1/COX-2, F4/80+, Mac-1+) [14, 15]. Their capacity to perform the most ancient defense mechanism – phagocytosis combined with the ability to produce antibodies suggests their direct link to pre-lymphocytes that emerged some 400-500 million years ago in
Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 3 of 16 (Page number not for citation purposes)
ancient vertebrates [16]. Although the exact role of B/macrophage cells was never fully explained it was shown that the cells release prostaglandins PGE2, PGF2α and PGD2 in a COX-2-dependent manner [15]. As PGD2 and its metabolites activate the nuclear receptor PPARγ that regulates lipid
metabolism, and PGE2 down-regulates IL-12 production, PGE2 synthesis by B/macrophage cells may shift the balance of an immune response towards Th2 and humoral immunity [15]. This, however, needs to be demonstrated empirically.
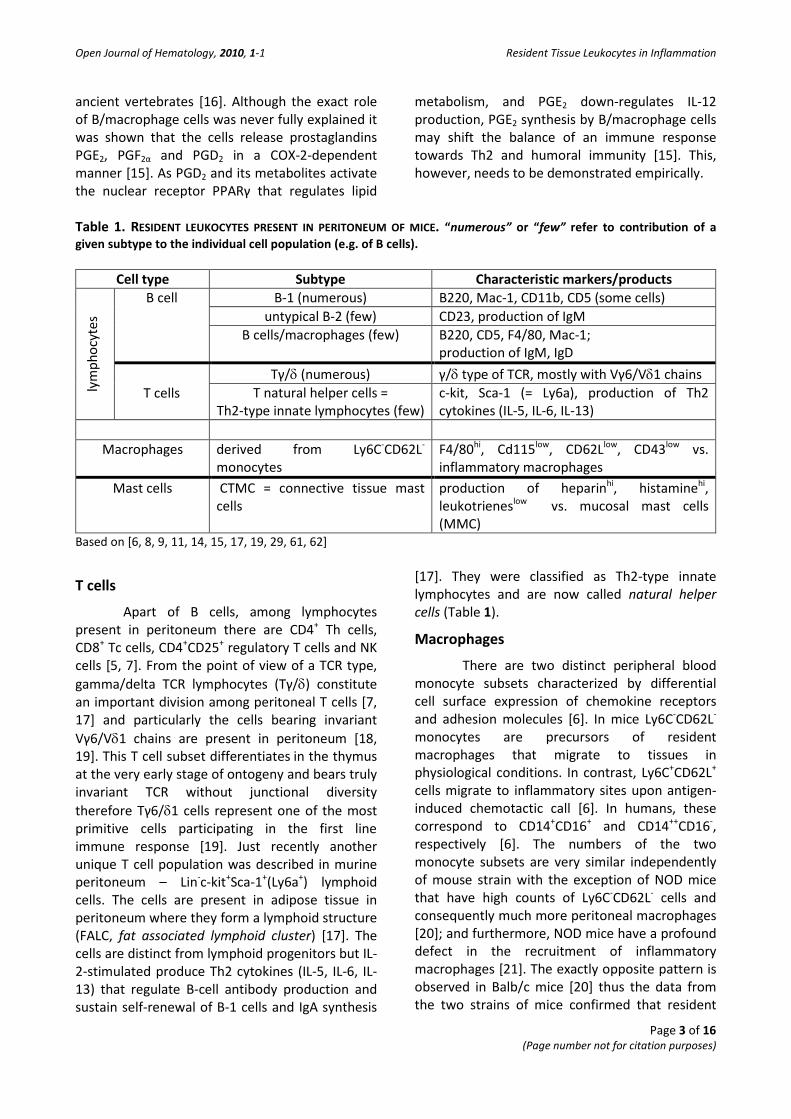
Table 1. RESIDENT LEUKOCYTES PRESENT IN PERITONEUM OF MICE. “numerous” or “few” refer to contribution of a given subtype to the individual cell population (e.g. of B cells).
Cell type Subtype Characteristic markers/products
lym
phoc
ytes
B cell B-1 (numerous) B220, Mac-1, CD11b, CD5 (some cells) untypical B-2 (few) CD23, production of IgM
B cells/macrophages (few) B220, CD5, F4/80, Mac-1; production of IgM, IgD
T cells
Tγ/δ (numerous) γ/δ type of TCR, mostly with Vγ6/Vδ1 chains T natural helper cells =
Th2-type innate lymphocytes (few) c-kit, Sca-1 (= Ly6a), production of Th2 cytokines (IL-5, IL-6, IL-13)
Macrophages derived from Ly6C-CD62L-
monocytes F4/80hi, Cd115low, CD62Llow, CD43low vs. inflammatory macrophages
Mast cells CTMC = connective tissue mast cells
production of heparinhi, histaminehi, leukotrieneslow vs. mucosal mast cells (MMC)
Based on [6, 8, 9, 11, 14, 15, 17, 19, 29, 61, 62]
T cells
Apart of B cells, among lymphocytes present in peritoneum there are CD4+ Th cells, CD8+ Tc cells, CD4+CD25+ regulatory T cells and NK cells [5, 7]. From the point of view of a TCR type, gamma/delta TCR lymphocytes (Tγ/δ) constitute an important division among peritoneal T cells [7, 17] and particularly the cells bearing invariant Vγ6/Vδ1 chains are present in peritoneum [18, 19]. This T cell subset differentiates in the thymus at the very early stage of ontogeny and bears truly
invariant TCR without junctional diversity therefore Tγ6/δ1 cells represent one of the most primitive cells participating in the first line immune response [19]. Just recently another unique T cell population was described in murine peritoneum – Lin-c-kit+Sca-1+(Ly6a+) lymphoid cells. The cells are present in adipose tissue in peritoneum where they form a lymphoid structure (FALC, fat associated lymphoid cluster) [17]. The cells are distinct from lymphoid progenitors but IL-2-stimulated produce Th2 cytokines (IL-5, IL-6, IL-13) that regulate B-cell antibody production and sustain self-renewal of B-1 cells and IgA synthesis
[17]. They were classified as Th2-type innate lymphocytes and are now called natural helper cells (Table 1).
Macrophages
There are two distinct peripheral blood monocyte subsets characterized by differential cell surface expression of chemokine receptors and adhesion molecules [6]. In mice Ly6C-CD62L- monocytes are precursors of resident macrophages that migrate to tissues in physiological conditions. In contrast, Ly6C+CD62L+ cells migrate to inflammatory sites upon antigen-induced chemotactic call [6]. In humans, these correspond to CD14+CD16+ and CD14++CD16-, respectively [6]. The numbers of the two monocyte subsets are very similar independently of mouse strain with the exception of NOD mice that have high counts of Ly6C-CD62L- cells and consequently much more peritoneal macrophages [20]; and furthermore, NOD mice have a profound defect in the recruitment of inflammatory macrophages [21]. The exactly opposite pattern is observed in Balb/c mice [20] thus the data from the two strains of mice confirmed that resident

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 4 of 16 (Page number not for citation purposes)
and inflammatory macrophages are derived from phenotypically and functionally different monocytes. This was further substantiated by an observation that LPS-stimulated resident peritoneal macrophages produce high levels of IL-6 and very low levels of IL-1β in sharp contrast to LPS-stimulated inflammatory macrophages induced by casein (one of the principal components of thioglycollate) [6]. The inflammatory macrophages are also slightly larger [6]. The two subsets of macrophages differ also in expression of some surface molecules as the resident cells express higher levels of 48/80 but lower of CD115, CD62L and CD43 than the inflammatory macrophages as shown in the latter case on thioglycollate-induced macrophages (Table 1) [6].
Moreover, there is diversity between resident peritoneal macrophages from various mouse strains with major differences detected in Balb/c mice. While expression of numerous macrophage surface molecules (CD11b, CD115, CD62L, CD43, Mac-3) is similar in different strains Balb/c mice are characterized by presence of numerous F4/80+ macrophages that are simultaneously expressing B220 (over 68% of the cells) which is considered to be a cell surface marker for B cells [6]. In contrast, in other investigated strains (C57Bl/6, CBA, CH/He, DBA1, NZW, MRL/Mp-Ipr) maximally 15% of peritoneal cells carry the F4/80+B220+ phenotype [6]. Although no further detailed studies were carried out on the double positive cells it cannot be excluded that these are related to biphenotypic B/macrophage cells described above. This would also correspond with the recognized dominance of the humoral immune response operating in Balb/c mice [22].
The secretory activity of resting resident peritoneal leukocytes is rather low and the major released protein is apoprotein E (ApoE) which is a ligand for low density lipoprotein receptors and thus participates in the transport of cholesterol and other lipids among various cells of the body [23]. The appearance of ApoE secretion correlates with the acquisition of mature macrophage phenotype, however, upon stimulation (e.g. with LPS or IFN-γ) resident macrophages limit the secretion of ApoE from app. 25% of total protein to < 2% [23].
Interestingly, some features of resident peritoneal macrophages change with age. For example, the cells from senescent mice express higher basal levels of transglutaminase (one of the markers of macrophage activation) and show increased phagocytic activity of zymosan particles indicating that aging resident macrophages are more activated than those from young mice [24, 25]. However, when the cells were challenged by inflammation in vivo they were not capable of reaching the same levels of activation as macrophages from young animals. Moreover, the inflammatory senescent macrophages were not capable of coping with a second challenge [26]. Most likely this reflects age-related changes in some basic mechanism e.g. signal transduction and/or protein synthesis/degradation but also altered responsiveness to environmental signals [24].
Mast cells
Mast cell-committed progenitors (MCP), i.e. Lyn-c-kit+Sca-1-Ly6c-FcεRIα-CD27-β7+T1/ST2+ cells are present in the bone marrow of adult mice [27]. In a similar manner as monocytes are precursors of macrophages MCPs are precursors of mast cells [27]. In what the two cell types differ is the capacity to proliferate as monocytes differentiate into macrophages without widespread expansion while MCPs proliferate extensively - one MCP can give rise to at least 103 mast cells [28]. Moreover, in contrast to mature macrophages, mature mast cells maintain high proliferative capacity. Another difference is that some macrophages die after completing their functioning while mast cells, which function by degranulation, restore the content of their granules in time and are then ready for new assignments [28]. MCPs circulate in blood and enter the tissues where they undergo differentiation into mast cells and maturate. The main survival and developmental factor for mast cells is stem-cell factor (SCF, c-kit L) but also many cytokines (especially IL-3), growth factors and chemokines influences phenotype of those cells [29].
One of the distinctive features of mast cells is their capacity to store preformed mediators (e.g. cytokines, chemokines, eicosanoids etc.) in their abundant granules that can be emptied upon activation in a process

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 5 of 16 (Page number not for citation purposes)
known as degranulation [30]. Degranulation can be induced by multiple mechanisms that include the crosslinking of IgE with its high affinity receptor (FεRI) on mast cell surface but in some circumstances IgE can also bind to the IgG receptors (FcγRII and FcγRIII) or galectin-3 [29]. Moreover, mast cells can be activated directly by pathogens, including bacteria, viruses and fungi or their derivatives. The microbial products can activate mast cells directly via Toll-like receptors (TLRs) or receptors for bacterial toxins, or indirectly, by activating the complement system which components bind to appropriate receptors on mast cells [29].
MCPs give rise to two types of mast cells present in mice – connective tissue mast cells (CTMC, e.g. residing in peritoneum and skin) and mucosal mast cells (MMC, e.g. present in gastro-intestinal tract) [27, 30]. The phenotypes of these two populations differ and they seem to be determined by factors present in the final site of differentiation [29]. CTMC are the major population of mast cells which contains heparine rather than chondroitin sulphate E or di-B side-chains proteoglycans as in the case of MMC. Heterogenic mast cells are also observed in humans, however, their diversity is rather limited and restricted to presence or absence of chymase (MCTC) while tryptase is present in both human mast cell types (MCT) [31]. Some studies suggest that lack of chymase might reflect the functional activation of human mast cells rather than phenotypic differentiation related to anatomic site [30].
The current view on mast cells focuses rather on their functioning than tissue distribution and subdivision. Mast cells were shown to function as effector cells in both innate and adaptive immunity via killing of pathogens, degradation of toxic endo- and exogenous peptides, regulation of numbers, viability, distribution and phenotype of various cells (for review see [29]). Those immunomodulatory functions are clearly not only pro-inflammatory as
also the anti-inflammatory role of mast cells was recently revealed. Such immunosuppressive effects of mast cells were shown to be induced by IL-10 and TGF-β [29].
The major findings presented here for mast cells and macrophages residing in the peritoneal cavity correspond to data from other body cavities such as pleural and pericardial. For example, it was shown that some major characteristics of pleural macrophages coincide with those of peritoneal macrophages [32]. On the other hand, heart mast cells have some unique features and are much more numerous (up to 25% of all pericardial leukocytes) [33].
INFLAMMATION IN BRIEF Inflammatory responses to injury or
infection in different tissues/organs consist of common stages, however, the processes might differ in quality and quantity of produced mediators, the duration of the reaction, and inflammation-related tissue damage [1]. The common stages of inflammation consist of (A) vascular changes in the tissue with ongoing inflammatory response and consequent changes in vessel permeability. The process depends on activity of vasoactive mediators that induce vasoconstriction and vasodilatation. The increase in vasopermeability leads to leakage of plasma proteins that include some chemoattractants such as complement components. Those factors together with chemokines facilitate (B) diapedesis and subsequent chemotaxis of blood-derived leukocytes. The first inflammatory cells to arrive to the inflamed tissue are neutrophils and subsequently macrophages infiltrate the site (Fig. 1). The aim of the leukocyte influx into the inflammatory site is (C) elimination of the pathogen and after the successful removal of the intruder (D) a resolution of inflammation takes place. If, however, the pathogen persists the blood- or lymph node-derived lymphocytes (Tα/β and B-2) can be engaged and a pathological chronic inflammatory reaction can be initiated [1].

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 6 of 16 (Page number not for citation purposes)
Fig. 1 Kinetics of inflammation. The course of inflammation is characterized by sequential changes of different parameters such as vascular permeability leading to fluid accumulation at the site of injury and influx of blood-derived inflammatory leukocytes – neutrophils and then macrophages. Each of the processes is initiated by selective factors, vasoactive and chemotactic mediators, respectively. The mediators are released by both resident tissue as well as inflammatory leukocytes (according to [1, 39-43, 50]).The involvement of resident immune cells in induction of the following stages of inflammation is indicated on the figure. The arbitrarily increasing (or decreasing) width of the arrows indicates stronger (or weaker) participation of resident leukocytes in a given process.
The data on resident leukocyte impact on inflammation will be discussed on the exemplary model of acute inflammation, namely zymosan-induced peritonitis. The model is well-described and many aspects of immunobiology of resident leukocytes were studied in this model. Zymosan is a component of the cell wall of yeast Saccharomyces cerevisiae consisting of two major polysaccharides: β-glucan and mannan [34]. Differential receptors were assigned to recognize the polysaccharides, including TLRs. In fact it was shown in vitro that TLR2 participates in macrophage recognition of zymosan and its was
reported that also in vivo TLR2 is partially responsible for zymosan binding during pleurisy and arthritis [35, 36]. However, zymosan recognition by mast cells during peritonitis was shown to be TLR2-independent whereas it depends on receptors for C3a and C5a components of the complement system [37] and dectin-1 [38].
There are three apparent phases of zymosan peritonitis (I) the onset of the reaction (0-2 h) accompanied by changes in permeability of peritoneal vessels (highest at 30 minutes) [39-41], (II) the peak (4-8 hours) with maximal neutrophil

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 7 of 16 (Page number not for citation purposes)
accumulation in peritoneum at 6 hours [39, 42] and (III) the resolution phase with high inflammatory macrophage numbers and low neutrophil counts (>24 hours) [39, 43].
ROLE OF RESIDENT LYMPHOCYTES IN INFLAMMATION
Involvement of lymphocytes in the course of zymosan peritonitis was investigated in mice deficient of Rag enzyme(s) thus deprived of mature lymphocytes as Rag-1 and -2 enzymes are critical for recombination of the VDJ genes encoding immunoglobulins and T cell receptors [44]. As zymosan peritoneal inflammation is generally resolved after 24-48 hours and after 3 days most of the parameters come back to the basal levels potential involvement of thymus-independent lymphocyte response could be expected [39]. Our group showed that there were no significant perturbations in the course of inflammation in Rag-1/2-/- mice as neutrophil numbers were not changed at 6 hours of peritonitis [45] when they reach highest counts [39]. This correlated with unaltered synthesis of chemokines (KC, MCP-1) and metalloproteinases (MMP-2 and MMP-9) that facilitate leukocyte diapedesis and chemotaxis [45]. There were, however, lowered levels of anti-inflammatory IL-10 while increased production of nitric oxide (NO) was detected. These data corresponded to studies on mice deficient of B-1 lymphocytes (Xid mice) in which decreased synthesis of IL-10 down-regulated NO production in response to zymosan [46] suggesting B-1 involvement in the anti-inflammatory phase of inflammation which, however, was not investigated in zymosan-inflamed Rag-1/2-/- mice [45]. Down-regulated IL-10 synthesis in Rag-1-/- mice was also observed by Rajakariar et al. who additionally described increased TNF-α levels at 2 h of zymosan peritonitis [7] (at 6 h we observed only a tendency to enhanced TNF-α release [45]). However, the studies by Rajakariar et al. revealed contradictory results concerning leukocyte infiltration which was increased at 2 and 6 h in Rag-1-/- mice and at least at 2 h concerned mostly neutrophils [7]. Moreover, the study reported that during zymosan peritonitis resident peritoneal lymphocytes at first disappear from peritoneum but repopulate it numerously during resolution of inflammation albeit do not participate in the latter
process; instead they seem to be important if the subsequent infection occurs [7].
ROLE OF RESIDENT MACROPHAGES AND MAST CELLS IN INFLAMMATION
Intraperitoneal injection of zymosan induces a reaction known as macrophage disappearance reaction (MDR) that is manifested by a transient decrease of resident macrophages [47]. The process starts as early as within first half an hour after zymosan injection and can last for several hours. Within this time only small numbers of resident peritoneal macrophages can be recovered from peritoneum [39]. MDR was also observed during different types of inflammation and in different compartments of the body, and moreover it might also concern inflammatory macrophages as well as neutrophils [47]. The detailed studies of MDR revealed that the “disappearance of macrophages” is due to their adherence to the peritoneal (mesothelial) lining and it concerns both aggregates of adhering macrophages and single cells [48]. MDR is linked to the process of coagulation as the macrophage aggregates are surrounded by fibrin/fibrinogen filaments and addition of anticoagulant heparin reverses the reaction. Moreover, during the reappearance of macrophages increased plasminogen activity is observed and there are gaps in the fibrin bulk [47]. The purpose of macrophage adherence to the peritoneal lining is not completely clear and there are two possible explanations of this reaction. Primarily, during MDR macrophages undergo a series of activation steps resulting in generation of fully activated cells. Alternatively, the reaction is aimed to sequestrate macrophages delaying the non-specific immune response; this theory is less favored [47]. The first hypothesis is also supported by observation of significantly increased activity of resident peritoneal macrophages as well as that of mast cells already minutes after induction of zymosan peritonitis (see following sections).
One of the commonly applied methods to study function or resident peritoneal cells in murine models is their selective depletion before induction of inflammation. In this way one can evaluate if the lack of the certain cell population changes the course of the inflammatory response in comparison to mice with physiological composition of peritoneal leukocytes; thus

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 8 of 16 (Page number not for citation purposes)
indirectly confirming or disproving their role. Peritoneal macrophages can be depleted by application of a compound that induces apoptosis of macrophages once phagocytosed. The drug (dichloromethylene diphosphonate) is delivered into peritoneum packed in multilamellar liposomes that are digested by a macrophage. Three doses applied on three constitutive days eliminate over 80% of peritoneal macrophages that are cleared from peritoneum by day 4 [49]. Whereas resident peritoneal mast cells can be functionally depleted by a single i.p. injection of compound 48/80 that induces mast cell degranulation, and substances/mediators released from mast cells are cleared up from the
peritoneum or expire by day 4 post-treatment, and after this time inflammation can be induced. Moreover, mast cells do not reappear in the peritoneum within next few days.
Alternatively, mice deficient in one population of leukocytes can be used. In studies on zymosan peritonitis one type of such mice was used - WBB6F1-W/Wv mice which lack c-kit receptor for SCF that is required to ensure mast cell survival in the tissues. Those mice, however, have several other phenotypical abnormalities, such as macrocytic anemia, a decrease in the number of bone marrow- and blood-derived neutrophils, sterility etc. [29].
Table 2. INVOLVEMENT OF RESIDENT PERITONEAL MAST CELLS AND MACROPHAGES IN DIFFERENT STAGES OF ZYMOSAN
PERITONITIS. – no effect, + effect on a given parameter (where arbitrary +, ++ and +++ show significance of the involvement), * in unstimulated cells, ° macrophages down-regulate (↓) neutrophil accumulation in peritoneum, in contrast to mast cells (↑), through release of anti-inflammatory IL-10.
Resident peritoneal: Phase of
zymosan peritonitis:
Process or parameter
mast cells
macrophages
onset
Histamine +++ - PGE2, PGF1α +-/ ++
LTC4-LTE4 - +++ Increase of vasopermeability +++ +++
peak
KC ++ - MCP-1 ++ -
NE* + + MMP-9 + ++/+++
Neutrophil infiltration ++ (↑) ++° (↓)
resolution IL-10 - ++ PGD2 + ++
According to [37-41, 47, 50, 53, 56-58].
Vascular permeability
The above methods were used to evaluate a role of mast cells and macrophages in changes in permeability of peritoneal vessels that occur within minutes after zymosan application and reach maximum at 30 minutes of peritonitis (Table 2) [39]. The studies showed that both mast cells and macrophages co-mediate the process that is significantly impaired if one of the populations is missing [40]. Moreover, some investigations were performed on WBB6F1-W/Wv mice and the mice were additionally depleted of macrophages. In those animals the increase of vasopermeability was limited by 80% [40] proving that the two
populations of resident peritoneal leukocytes play a major role in the process. It was also revealed that while the impact on permeability changes of mast cells is related mainly to the release of histamine, in the case of macrophages it depends mostly on prostaglandins (E2 and F1α), and cysteinyl leukotrienes (LTC4-E4) produced almost exclusively by those cells [39, 40]. Cyclooxygenases are enzymes required for prostaglandin production and it was further reported that the constitutive COX-1 isoform is expressed by both mast cells and macrophages as its content is down-regulated by their depletion [50]. In contrast, the inducible COX-2 responsible for synthesis of inflammatory prostaglandins

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 9 of 16 (Page number not for citation purposes)
operates mostly in macrophages whereas in mast cells only during the peak of the reaction (6 hours of peritonitis) [50]. This corresponds with the engagement of peritoneal macrophages in synthesis of PGE2 and PGD2 at the early and late stages of zymosan peritonitis, respectively [50].
Chemokine synthesis
Selective depletion of peritoneal mast cells had significant impact on infiltration of inflammatory neutrophils into zymosan inflamed peritoneum that was inhibited by over 60% (Table 2) [43]. Furthermore, it was shown that mast cells contribute significantly to synthesis of two important murine chemokines KC (CXCL1) and JE (MCP-1, CCL2) as their gene as well as protein expression were impaired in mice depleted of mast cells also by approximately 60% (Table 2) [43]. Rodents lack a direct homologue of IL-8 (CXCL8), but KC, along with MIP-2 (CXCL2) and LIX (CXCL5-6), are regarded as functional homologues of IL-8 that are responsible for recruitment of neutrophils [51]. On the other hand, MCP-1 attracts rather lymphocytes, natural killers and monocytes/macrophages, and moreover, monocytes/macrophages are found to be its major source [52], therefore data on significant contribution of mast cells into MCP-1 synthesis was unexpected [43]. The study analyzed only numbers of neutrophils and not those of subsequently infiltrating inflammatory macrophages thus the exact role of zymosan induced MCP-1 was not verified [43]. Decreased levels of MCP-1 in mice depleted of peritoneal mast cells were also confirmed by our group [53] but it cannot be excluded that the leukocytes are not direct producers of the chemokines but instead their mediators stimulate other cells to release KC and MCP-1. In contrast to mast cells, the depletion of macrophages did not modulate synthesis of the two chemokines [43]. However, removal of peritoneal macrophages up-regulated synthesis of MIP-1α [54]. This indicates that although peritoneal macrophages do not produce MIP-1α they control its synthesis.
Cytokine synthesis
Involvement of tissue resident macrophages and mast cells in synthesis of pro-inflammatory cytokines was negligibly investigated. It was shown that in WBB6F1-W/Wv mice with zymosan peritonitis levels of TNF-α, IL-
1β, IL-6 are significantly decreased suggesting mast cell involvement in their production [39]. However, this cannot be excluded that the down-regulation of the cytokine synthesis was a consequence of impaired neutrophil accumulation observed in zymosan-inflamed WBB6F1-W/Wv mice [39].
The study performed by Turchyn and co-workers demonstrated that ex vivo stimulation of resident peritoneal leukocytes with zymosan lead to time-dependent increase of TNF-α release [6]. Moreover, when macrophages were selectively removed from the pool of total resident peritoneal leukocytes TNF-α levels were reduced by over 90% during the 4-hour stimulation. Importantly, also depletion of mast cells reduced, slightly but significantly, TNF-α synthesis [6]. In fact during generalized immune complex-induced inflammation of the peritoneal cavity TNF-α production was reported to consists of two steps and mast cells were shown to be a major source of this cytokine during the first early peak [55]. Thus overall, the above data confirms that both peritoneal mast cells and macrophages are important producers of inflammatory cytokines.
Protease production and release
The influx of leukocytes into tissues is selectively initiated by chemokines. During their extravasation and further migration in response to the chemotactic signal the cells have to cross some barriers such as basement membranes and extracellular matrix (ECM) [1]. This can be achieved only by their degradation through the release of proteases. One of such proteases is carried in the azurophil granules of developing as well as mature neutrophils and for this is called neutrophil elastase (NE) [56]. However, it was shown that also unstimulated resident peritoneal mast cells and especially macrophages express NE (Table 2) [57]. The reason for NE expression by resting cells was not clarified but it was proposed that NE released in physiological conditions might be involved in the programmed cell death that occurs in the peritoneum as a part of a normal cell turnover cycle [57]. This was based on observation that NE decreases the mitochondrial membrane potential, releases cytochrome c, and cleaves caspases 9 and 3, leading to apoptosis of other cell types (e.g. epithelial cells) [58]. Although it was not investigated if NE is released also by zymosan-

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 10 of 16 (Page number not for citation purposes)
activated mast cells and macrophages this was done in the case of another proteinase - MMP-9. The enzyme belongs to matrix metalloproteinase family which members are well known for their capacity to degrade components of ECM and basement membranes [59]. MMP-9 levels increase during zymosan peritonitis and either pharmacological or genetic (MMP-9-/- mice) inhibition of the enzyme significantly decreases neutrophil infiltration of peritoneum [60]. Intracellular FACSCan analyses revealed that resting mast cells, in contrast to macrophages, contain MMP-9 in their granules. Upon intraperitoneal zymosan injection (after 30 minutes) mast cells slightly increase MMP-9 production but first of all it starts in peritoneal macrophages and then it increases dramatically [61]. In line with these data the removal of mast cells does not have significant impact on the MMP-9 pool in contrast to macrophages which are the major producers of the enzyme at these very early time points of inflammation (Table 2) [62]. Although mast cells do synthetise MMP-9 during zymosan peritonitis these are not significant quantities. Moreover, some fluctuations in levels of MMP-9 were detected in mast cell-depleted mice but these are mostly due to impaired neutrophil infiltration [62]. In contrast, removal of peritoneal macrophages has indeed a very significant down-regulating effect on MMP-9 levels released to the peritoneal cavity. This was shown that especially at 6 hours of peritonitis lack of resident peritoneal macrophages decreased MMP-9 release by half. The fact that it was accompanied by even enhanced neutrophil infiltration (see below) strengthens the data [62].
Resolution of inflammation
As mentioned above the lack of peritoneal mast cells had a considerable impact on numbers of neutrophils infiltrating zymosan inflamed peritoneum indicating that the cells are critical for the onset of the inflammatory reaction. Surprising results, however, were observed in the case of peritoneal macrophages as in mice depleted of those cells the accumulation of neutrophils was in fact increased. This turned out to be directly related to the synthesis of IL-10 [43, 63]. Interleukin 10 is produced predominantly by T cells and monocytes/macrophages and has immunosuppressive effects on release of pro-
inflammatory cytokines, chemokines and MMPs [55]. When IL-10-/- mice were depleted of peritoneal macrophages the zymosan-induced neutrophil accumulation was further increased suggesting involvement of also other macrophage products in regulation of neutrophil influx into peritoneum [43]. Those additional anti-inflammatory mediators were not identified in the study. However, recently our group showed that resident peritoneal macrophages are also important producers of another anti-inflammatory product - PGD2 [50]. Synthesis of this prostaglandin was reduced by some 80-90% in animals depleted of peritoneal macrophages. Therefore PGD2 might represent one of the additional mediators released by those cells that is responsible for negative control of neutrophil accumulation in zymosan inflamed peritoneum (Table 2).
HUMAN RESIDENT PERITONEAL LEUKOCYTES AND THEIR ROLE IN INFLAMMATION
Studies on human resident leukocytes are much less comprehensive. This is due to ethical issues and obstacles related to access to samples from some loci e.g. peritoneum. In fact most studies on peritoneal resident leukocytes were performed on samples collected during peritoneal dialysis (PD) used for treatment of patients with end-stage renal diseases. In those studies the PD patients without peritonitis serve most frequently as healthy controls, while repeated peritoneal inflammation is a common complication of PD. Therefore analysis of the data should be conscious as in the PD patients the low-grade chronic inflammation can be detected. Recently, it was demonstrated that this phenomenon was partially reduced after introduction of automated peritoneal dialysis (APD) and application of a new generation of PD solutions with physiological pH and low concentration of glucose degradation products (for review see [64]).
In one study on infection-free patients treated with continuous ambulatory peritoneal dialysis (CAPD) and on patients during bacterial peritonitis expression of adhesion molecules on leukocytes was investigated [65]. The study revealed that peritoneal macrophages from CAPD patients have higher expression of Mac-1 subunits

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 11 of 16 (Page number not for citation purposes)
CD11b and CD18 as well as of Mac-1 ligand – ICAM-1 (CD54) than blood monocytes [65]. The interaction of the two groups of adhesion molecules results in a firm adherence of leukocytes to the endothelium during their transmigration leading to diapedesis and further migration through the mesothelial cell layer into the peritoneal cavity [66]. Therefore the presence of Mac-1 and ICAM-1 on the peritoneal macrophages might suggest monocyte extravasation into the peritoneal cavity during dialysis. Moreover, only the expression of ICAM-1 was further increased on macrophages collected from patients with peritonitis showing that the cells are chronically activated during the renal failure [65]. In another study the elevated expression of CD14 on peritoneal macrophages was observed in CAPD patients as well as in bacterial peritonitis patients [67]. CD14 is a differentiation antigen and a co-receptor for LPS, and it plays a role in the transduction of activatory signals for the production of cytokines [67]. The report also revealed that in contrast to patients with bacterial peritonitis in which increased levels of neutrophils were observed, in CAPD patients there are more peritoneal macrophages [67]. On the other hand, however, it was documented that in vitro at least 525 macrophages per milliliter are required to suppress bacterial growth while effluents from PD patients contain only 104 cells/ml [68]. This might provide an explanation of frequent incidences of bacterial peritonitis in PD patients. Overall, the above results suggest that resident peritoneal macrophages are involved in the inflammatory process occurring in peritoneum.
Another feature of peritoneal leukocytes related to phagocytosis was evaluated in PD children [69] but similar results were observed in adults [70]. The process of phagocytosis might be enhanced by opsonins coating pathogens as opsonin receptors are present on the macrophage surface [1]. They include receptors for the constant region of IgG (FcγR) and for complement components (CR) [71]. It was also demonstrated that as a result of dilution and removal during PD, the concentration of opsonins in peritoneal effluent is decreased [72, 73]. In PD children peritoneal macrophages have less FcγR+ and CR+ cells but the number of the receptors is higher [69]. This indicates that although the FcγR+/CR+–
related function is impaired in those cells they are in a state of activation.
Furthermore, it was reported that human peritoneal macrophages treated ex vivo with LPS produce intensively pro-inflammatory TNF-α [74], IL-1β [75] and IL-6 [76], and increase superoxide generation [77]. Moreover, it was shown that human peritoneal macrophages produce 1a,25-dihydroxyvitamin D3 which controls the intensity of inflammation by inhibiting e.g. TNF-α synthesis [74].
Studies were also focused on peritoneal lymphocytes and it was shown that their numbers might be increased from 5-10% to 5-77% in the dialyzed patients [64], and moreover, the PD lymphocytes seem to be functionally impaired as they produce significantly less IL-2 [68]. Also mast cells were analyzed among human peritoneal leukocytes. There are, however, contradictory results on their numbers in PD patients. A study by Jimenez-Heffernan et al. showed that in comparison to “normal controls” (the samples were collected from autopsy cases and kidney donors) numbers of mast cells (evaluated by immunocytochemistry detecting tryptase) were higher by app. 50% in comparison to PD patients [78]. Although similar results were observed in PD-treated rats [79] the outcome surprised the authors. This was due to the known contribution of mast cells to peritoneal fibrosis accompanying long-term PD [80]. And furthermore, it was shown that histamine released from peritoneal resident mast cells induces calcium release in mesothelial cells which controls cytoskeleton organization and influences the performance of the peritoneal membrane during dialysis [81].
In contrast, Alscher and co-workers reported a 3-fold increase of mast cell numbers in PD patients [82]. In this study “normal controls” were collected from patients with diverticulosis of the sigma or colorectal carcinoma (without peritoneal metastasis) and mast cells were identified by staining with α-naphtol-AS-D-chloracetate-esterase and immunocytochemistry of tryptase [82]. Moreover, the study showed that mast cells were present not only in the peritoneal fluid but also in the peritoneal membrane and were partially degranulated in PD patients [82].
All the above described studies were concentrated on numbers and some basic

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 12 of 16 (Page number not for citation purposes)
activities of resident peritoneal leukocytes during PD and peritonitis. However, indirectly they provide important information on engagement of peritoneal mast cells and macrophages in immune processes in peritoneum. The fact that removal of peritoneal cells during dialysis leads to constant mobilization of blood monocytes to peritoneum further confirms their importance for normal functioning in humans.
CONCLUSIONS
As shown here on the basic model of inflammation that is provided by murine zymosan-induced peritonitis indeed resident tissue leukocytes do play a significant role in different stages of the process. One could predict their involvement in the initiation of inflammation that would also explain their presence in tissues other than blood in physiological conditions, nevertheless, their involvement in the late stages of inflammation is still not generally recognized. If however this is the case more attention should be directed to the resident tissue leukocytes as possible therapeutic targets, especially in chronic inflammatory disorders. In contrast to the acute inflammatory reaction that is a beneficial process – it leads to clearance of the pathogen and healing of the wounds – the chronic inflammation is considered to be a pathological process. As apart of inflammatory macrophages also resident macrophages were shown to have potent anti-inflammatory characteristics the selective or mutual up-regulation of the two cell types could improve remission or even resolution or the chronic process. This is reinforced by observations from studies on human resident leukocytes and their role in the immune-defense.
LIST OF ABBREVIATIONS
APD – automated peritoneal dialysis ApoE – apoprotein E C – complement factor CAPD – continuous ambulatory peritoneal dialysis CD – clusters of differentiation ECM – extracellular matrix
COX – cyclooxygenase CTMC – connective tissue mast cells (mice) FACS – fluorescence-activated cell sorting FALC – fat associated lymphoid cluster ICAM-1 – Inter-Cellular Adhesion Molecule 1 IFN – interferon Ig – immunoglobulin IL – interleukin KC – keratinocyte chemoattractant/keratinocyte-derived chemokine LIX – lipopolysaccharide-induced CXC chemokine LPS – lipopolysaccharide LT – leukotriene MCO – mast cell-committed progenitor MCP – monocyte chemotactic protein MCT – tryptase mast cells (human) MCTC – tryptase/chymase mast cells (human) MDR – macrophage disappearance reaction MHC – major histocompatibility complex MIP – macrophage inflammatory protein MMC – mucosal mast cells (mice) MMP – matrix metalloproteinase NE – neutrophil elastase NO – nitric oxide PD – peritoneal dialysis PG – prostaglandin PPAR-γ – Peroxisome proliferator-activated receptor gamma RI – receptor type I Rag – recombination activating gene SCF – stem cell factor VDJ – variable (V), diversity (D) and joining (J) genes encoding variable regions of Ig and TCR TCR – T-cell receptor TdT – terminal deoxynucleotidyl transferase TLR – Toll-like receptor TGF – transforming growth factor TNF – tumor necrosis factor.
CONFLICT OF INTEREST
There is no conflict of interest.
ACKNOWLEDGMENTS The work was supported by K/ZDS/000784.

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 13 of 16 (Page number not for citation purposes)
REFERENCES [1] Majno G, Joris I. Cells, Tissues, and Disease:
Principles of General Pathology. 2nd ed, Blackwell: Oxford, 2004.
[2] Sammour T, Kahokehr A, Soop M, Hill AG. Peritoneal damage: the inflammatory response and clinical implications of the neuro-immuno-humoral axis. World J Surg. 2010; 34: 704-720.
[3] Hills BA. Graphite-like lubrication of mesothelium by oligolamellar pleural surfactant. J Appl Physiol. 1992; 73: 1034-1039.
[4] Heel KA, Hall JC. Peritoneal defences and peritoneum-associated lymphoid tissue. Br J Surg. 1996; 83: 1031-1036.
[5] Kolaczkowska E, Koziol A, Plytycz B, Arnold B, Opdenakker G. Altered apoptosis of inflammatory neutrophils in MMP-9-deficient mice is due to lower expression and activity of caspase-3. Immunol Lett. 2009; 126: 73-82.
[6] Turchyn LR, Baginski TJ, Renkiewicz RR, Lesch CA, Mobley JL. Phenotypic and functional analysis of murine resident and induced peritoneal macrophages. Comp Med. 2007; 57: 574-580.
[7] Rajakariar R, Lawrence T, Bystrom J, Hilliard M, Colville-Nash P, Bellingan G, Fitzgerald D, Yaqoob MM, Gilroy DW. Novel biphasic role for lymphocytes revealed during resolving inflammation. Blood. 2008; 111: 4184-4192.
[8] Kantor AB. The development and repertoire of B-1 cells (CD5 B cells). Immunol Today. 1991; 12: 389-391.
[9] Stall AM, Adams S, Herzenberg LA, Kantor AB. Characteristics and development of the murine B-1b (Ly-1 B sister) cell population. Ann N Y Acad Sci. 1992; 651: 33-43.
[10] Hardy RR. B-1 B cell development. J Immunol. 2006; 177: 2749-2754.
[11] Hastings WD, Gurdak SM, Tumang JR, Rothstein TL. CD5+/Mac-1- peritoneal B cells: a novel B cell subset that exhibits characteristics of B-1 cells. Immunol Lett. 2006; 105: 90-96.
[12] Cumano A, Paige CJ, Iscove NN, Brady G. Bipotential precursors of B cells and macrophages in murine fetal liver. Nature. 1992; 356: 612-615.
[13] Montecino-Rodriguez E, Leathers H, Dorshkind K. Bipotential B-macrophage progenitors are present in adult bone marrow. Nat Immunol. 2001; 2: 83-88.
[14] Borrello MA, Palis J, Phipps RP. The relationship of CD5+ B lymphocytes to macrophages: insights from normal biphenotypic B/macrophage cells. Int Rev Immunol. 2001; 20: 137-155.
[15] Graf BA, Nazarenko DA, Borrello MA, Roberts LJ, Morrow JD, Palis J, Phipps RP. Biphenotypic B/macrophage cells express COX-1 and up-regulate COX-2 expression and prostaglandin E(2)
production in response to pro-inflammatory signals. Eur J Immunol. 1999; 29: 3793-3803.
[16] Plytycz B, Seljelid R. B-1/macrophages as 'living fossils'. Immunol Today. 1997; 18: 505.
[17] Moro, K, Yamada, T, Tanabe, M, Takeuchi, T, Ikawa, T, Kawamoto, H, Furusawa, J, Ohtani, M, Fujii, H, Koyasu S. Innate production of T(H)2 cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature. 2010; 463: 540-544.
[18] Shibata K, Yamada H, Hara H, Kishihara K, Yoshikai Y. Resident Vdelta1+ gammadelta T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J Immunol. 2007; 178: 4466-4472.
[19] Tagawa T, Nishimura H, Yajima T, Hara H, Kishihara K, Matsuzaki G, Yoshino I, Maehara Y, Yoshikai Y. Vdelta1+ gammadelta T cells producing CC chemokines may bridge a gap between neutrophils and macrophages in innate immunity during Escherichia coli infection in mice. J Immunol. 2004; 173: 5156-5164.
[20] Nikolic T, Bouma G, Drexhage HA, Leenen PJ. Diabetes-prone NOD mice show an expanded subpopulation of mature circulating monocytes, which preferentially develop into macrophage-like cells in vitro. J Leukoc Biol. 2005; 78: 70-79.
[21] Bouma G, Nikolic T, Coppens JM, van Helden-Meeuwsen CG, Leenen PJ, Drexhage HA, Sozzani S, Versnel MA. NOD mice have a severely impaired ability to recruit leukocytes into sites of inflammation. Eur J Immunol. 2005; 35: 225-235.
[22] Rose ME, Wakelin D, Hesketh P. Susceptibility to coccidiosis: contrasting course of primary infections with Eimeria vermiformis in BALB/c and C57/BL/6 mice is based on immune responses. Parasite Immunol. 1985; 7: 557-566.
[23] Takemura R, Werb Z. Modulation of apoprotein E secretion in response to receptor-mediated endocytosis in resident and inflammatory macrophages. J Exp Med. 1984; 159: 167-178.
[24] Lavie L, Weinreb O. Age- and strain-related changes in tissue transglutaminase activity in murine macrophages: the effects of inflammation and induction by retinol. Mech Ageing Dev. 1996: 90: 129-143.
[25] Lavie L, Gershon D. Oxygen free radical production by mouse peritoneal macrophages as a function of age. Mech Ageing Dev. 1988; 45: 177-189.
[26] Lavie L, Weinreb O, Gershon D. Age-related alterations in superoxide anion generation in mouse peritoneal macrophages studied by repeated stimulations and heat shock treatment. J Cell Physiol. 1992; 152: 382-388.
[27] Chen CC, Grimbaldeston MA, Tsai M, Weissman IL, Galli SJ. Identification of mast cell progenitors in

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 14 of 16 (Page number not for citation purposes)
adult mice. Proc Natl Acad Sci U S A. 2005; 102: 11408-11413.
[28] Kitamura Y, Ito A. Mast cell-committed progenitors. Proc Natl Acad Sci U S A. 2005; 102: 11129-11130.
[29] Galli SJ, Grimbaldeston M, Tsai M. Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nat Rev Immunol. 2008; 8: 478-486.
[30] Enerback L. The differentiation and maturation of inflammatory cells involved in the allergic response: mast cells and basophils. Allergy. 1997; 52: 4-10.
[31] Irani AA, Schechter NM, Craig SS, DeBlois G, Schwartz LB. Two types of human mast cells that have distinct neutral protease compositions. Proc Natl Acad Sci U S A. 1986; 83: 4464-4468.
[32] Shimotakahara A, Kuebler JF, Vieten G, Metzelder ML, Petersen C, Ure BM. Pleural macrophages are the dominant cell population in the thoracic cavity with an inflammatory cytokine profile similar to peritoneal macrophages. Pediatr Surg Int. 2007; 23: 447-451.
[33] Morgan LG, Levick SP, Voloshenyuk TG, Murray DB, Forman MF, Brower GL, Janicki JS. A novel technique for isolating functional mast cells from the heart. Inflamm Res. 2008; 57: 241-246.
[34] Di Carlo FJ, Fiore JV. On the composition of zymosan. Science. 1958; 127: 756-757.
[35] Takeshita K, Sakai K, Bacon KB, Gantner F. Critical role of histamine H4 receptor in leukotriene B4 production and mast cell-dependent neutrophil recruitment induced by zymosan in vivo. J Pharmacol Exp Ther. 2003; 307: 1072-1078.
[36] Frasnelli ME, Tarussio D, Chobaz-Peclat V, Busso N, So A. TLR2 modulates inflammation in zymosan-induced arthritis in mice. Arthritis Res Ther. 2005; 7: R370-379.
[37] Mullaly SC, Kubes P. Mast cell-expressed complement receptor, not TLR2, is the main detector of zymosan in peritonitis. Eur J Immunol. 2007; 37: 224-234.
[38] Yang Z, Marshall JS. Zymosan treatment of mouse mast cells enhances dectin-1 expression and induces dectin-1-dependent reactive oxygen species (ROS) generation. Immunobiology. 2009; 214: 321-330.
[39] Kolaczkowska E, Seljelid R, Plytycz B. Role of mast cells in zymosan-induced peritoneal inflammation in Balb/c and mast cell-deficient WBB6F1 mice. J Leukoc Biol. 2001; 69: 33-42.
[40] Kolaczkowska E, Shahzidi S, Seljelid R, van Rooijen N, Plytycz B. Early vascular permeability in murine experimental peritonitis is co-mediated by resident peritoneal macrophages and mast cells: crucial involvement of macrophage-derived
cysteinyl-leukotrienes. Inflammation. 2002; 26: 61-71.
[41] Doherty NS, Poubelle P, Borgeat P, Beaver TH, Westrich GL, Schrader NL. Intraperitoneal injection of zymosan in mice induces pain, inflammation and the synthesis of peptidoleukotrienes and prostaglandin E2. Prostaglandins. 1985; 30: 769-789.
[42] Ajuebor MN, Flower RJ, Hannon R, Christie M, Bowers K, Verity A, Perretti M. Endogenous monocyte chemoattractant protein-1 recruits monocytes in the zymosan peritonitis model. J Leukoc Biol. 1998; 63: 108-116.
[43] Ajuebor MN, Das AM, Virag L, Flower RJ, Szabo C, Perretti M. Role of resident peritoneal macrophages and mast cells in chemokine production and neutrophil migration in acute inflammation: evidence for an inhibitory loop involving endogenous IL-10. J Immunol. 1999; 162: 1685-1691.
[44] Jones JM, Simkus C. The roles of the RAG1 and RAG2 "non-core" regions in V(D)J recombination and lymphocyte development. Arch Immunol Ther Exp (Warsz). 2009; 57: 105-116.
[45] Kolaczkowska E, Barteczko M, Plytycz B, Arnold B. Role of lymphocytes in the course of murine zymosan-induced peritonitis. Inflamm Res. 2008; 57: 272-278.
[46] Popi AF, Lopes JD, Mariano M. Interleukin-10 secreted by B-1 cells modulates the phagocytic activity of murine macrophages in vitro. Immunology. 2004; 113: 348-354.
[47] Barth MW, Hendrzak JA, Melnicoff MJ, Morahan PS. Review of the macrophage disappearance reaction. J Leukoc Biol. 1995; 57: 361-367.
[48] Nelson DS, North RJ. The fate of peritoneal macrophages after the injection of antigen into guinea pigs with delayed-type hypersensitivity. Lab Invest. 1965; 14: 89-101.
[49] van Rooijen N, Sanders A. Liposome mediated depletion of macrophages: mechanism of action, preparation of liposomes and applications. J Immunol Methods. 1994; 174: 83-93.
[50] Kolaczkowska E, Goldys A, Kozakiewicz E, Lelito M, Plytycz B, van Rooijen N, Arnold B. Resident peritoneal macrophages and mast cells are important cellular sites of COX-1 and COX-2 activity during acute peritoneal inflammation. Arch Immunol Ther Exp (Warsz). 2009; 57: 459-466.
[51] Hol J, Wilhelmsen L, Haraldsen G. The murine IL-8 homologues KC, MIP-2, and LIX are found in endothelial cytoplasmic granules but not in Weibel-Palade bodies. J Leukoc Biol. 2010; 87: 501-508.
[52] Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 15 of 16 (Page number not for citation purposes)
overview. J Interferon Cytokine Res. 2009; 29: 313-326.
[53] Kolaczkowska E, Arnold B, Plytycz B. Mast cell involvement in zymosan-induced peritonitis in C57Bl/6 mice. Centr Eur J Immunol. 2008; 33: 91-97.
[54] Ajuebor MN, Das AM, Virag L, Szabo C, Perretti M. Regulation of macrophage inflammatory protein-1 alpha expression and function by endogenous interleukin-10 in a model of acute inflammation. Biochem Biophys Res Commun. 1999; 255: 279-282.
[55] Mosser DM, Zhang X. Interleukin-10: new perspectives on an old cytokine. Immunol Rev. 2008; 226: 205-218.
[56] Chua F, Laurent GJ. Neutrophil elastase: mediator of extracellular matrix destruction and accumulation. Proc Am Thorac Soc. 2006; 3: 424-427.
[57] Kolaczkowska E, Grzybek W, van Rooijen N, Piccard H, Plytycz B, Arnold B, Opdenakker G. Neutrophil elastase activity compensates for a genetic lack of matrix metalloproteinase-9 (MMP-9) in leukocyte infiltration in a model of experimental peritonitis. J Leukoc Biol. 2009; 85: 374-381.
[58] Preston GA, Zarella CS, Pendergraft WF 3rd, Rudolph EH, Yang JJ, Sekura SB, Jennette JC, Falk RJ. Novel effects of neutrophil-derived proteinase 3 and elastase on the vascular endothelium involve in vivo cleavage of NF-kappaB and proapoptotic changes in JNK, ERK, and p38 MAPK signaling pathways. J Am Soc Nephrol. 2002; 13: 2840-2849.
[59] Van den Steen PE, Dubois B, Nelissen I, Rudd PM, Dwek RA, Opdenakker G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9). Crit Rev Biochem Mol Biol. 2002; 37: 375-536.
[60] Kolaczkowska E, Chadzinska M, Scislowska-Czarnecka A, Plytycz B, Opdenakker G, Arnold B. Gelatinase B/matrix metalloproteinase-9 contributes to cellular infiltration in a murine model of zymosan peritonitis. Immunobiology. 2006; 211: 137-148.
[61] Kolaczkowska E, Arnold B, Opdenakker G. Gelatinase B/MMP-9 as an inflammatory marker enzyme in mouse zymosan peritonitis: comparison of phase-specific and cell-specific production by mast cells, macrophages and neutrophils. Immunobiology.2008; 213: 109-124.
[62] Kolaczkowska E, Lelito M, Kozakiewicz E, van Rooijen N, Plytycz B, Arnold B. Resident peritoneal leukocytes are important sources of MMP-9 during zymosan peritonitis: superior contribution of macrophages over mast cells. Immunol Lett. 2007; 113: 99-106.
[63] Chadzinska M, Kolaczkowska E, Scislowska-Czarnecka A, Van Rooijen N, Plytycz B. Effects of macrophage depletion on peritoneal inflammation in swiss mice, edible frogs and goldfish. Folia Biol (Krakow). 2004; 52: 225-231.
[64] Kazancioglu R. Peritoneal defense mechanisms--the effects of new peritoneal dialysis solutions. Perit Dial Int. 2009; 29 (Suppl 2): S198-201.
[65] Liberek T, Chmielewski M, Lichodziejewska-Niemierko M, Lewandowski K, Rutkowski B. Transmigration of blood leukocytes into the peritoneal cavity is related to the upregulation of ICAM-1 (CD54) and Mac-1 (CD11b/CD18) adhesion molecules. Perit Dial Int. 2004; 24: 139-146.
[66] Li FK, Davenport A, Robson RL, Loetscher P, Rothlein R, Williams JD, Topley N. Leukocyte migration across human peritoneal mesothelial cells is dependent on directed chemokine secretion and ICAM-1 expression. Kidney Int. 1998; 54: 2170-2183.
[67] Glorieux G, Lameire N, Van Biesen W, Dequidt C, Vanholder R. Specific characteristics of peritoneal leucocyte populations during sterile peritonitis associated with icodextrin CAPD fluids. Nephrol Dial Transplant. 2003; 18: 1648-1653.
[68] Lewis S, Holmes C. Host defense mechanisms in the peritoneal cavity of continuous ambulatory peritoneal dialysis patients. 1. Perit Dial Int. 1991; 11: 14-21.
[69] Bouts AH, Davin JC, Krediet RT, Schroder CH, Monnens LA, Nauta J, van de Winkel JG, Out TA. IgG and complement receptor expression in children treated by peritoneal dialysis. Pediatr Nephrol. 2005; 20: 1161-1167.
[70] Carcamo C, Fernandez-Castro M, Selgas R, Jimenez C, Molina S, Vara F. Long-term continuous ambulatory peritoneal dialysis reduces the expression of CD11b, CD14, CD16, and CD64 on peritoneal macrophages. Perit Dial Int. 1996; 16: 582-589.
[71] Vidarsson G, van de Winkel JG. Fc receptor and complement receptor-mediated phagocytosis in host defence. Curr Opin Infect Dis. 1998; 11: 271-278.
[72] Brulez HF, Verbrugh HA. First-line defense mechanisms in the peritoneal cavity during peritoneal dialysis. Perit Dial Int. 1995; 15: S24-33; discussion S33-24.
[73] McGregor SJ, Brock JH, Briggs JD, Junor BJ. Longitudinal study of peritoneal defence mechanisms in patients on continuous ambulatory peritoneal dialysis (CAPD). Perit Dial Int. 1989; 9: 115-119.
[74] Cohen ML, Douvdevani A, Chaimovitz C, Shany S. Regulation of TNF-alpha by 1alpha,25-

Open Journal of Hematology, 2010, 1-1 Resident Tissue Leukocytes in Inflammation
Page 16 of 16 (Page number not for citation purposes)
dihydroxyvitamin D3 in human macrophages from CAPD patients. Kidney Int. 2001; 59: 69-75.
[75] Douvdevani A, Rapoport J, Konforti A, Zlotnik M, Chaimovitz C. The effect of peritoneal dialysis fluid on the release of IL-1 beta and TNF alpha by macrophages/monocytes. Perit Dial Int. 1993; 13: 112-117.
[76] Steinhauer HB, Brugger U, Atmanspacher R, Lubrich-Birkner I, Schollmeyer P. Effect of CAPD dialysate on the release of eicosanoids and cytokines from human peritoneal macrophages. Adv Perit Dial., 1992; 8: 47-52.
[77] Deutsch A, Chaimovitzch C, Nagauker-Shriker O, Zlotnik M, Shany S, Levy R. Elevated superoxide generation in mononuclear phagocytes by treatment with 1 alpha hydroxyvitamin D3: changes in kinetics and in oxidase cytosolic factor p47. J Am Soc Nephrol. 1995; 6: 102-109.
[78] Jimenez-Heffernan JA, Bajo MA, Perna C, del Peso G, Larrubia JR, Gamallo C, Sanchez-Tomero JA, Lopez-Cabrera M, Selgas R. Mast cell quantification in normal peritoneum and during
peritoneal dialysis treatment. Arch Pathol Lab Med. 2006; 130: 1188-1192.
[79] Hekking LH, Zareie M, Driesprong BA, Faict D, Welten AG, de Greeuw I, Schadee-Eestermans IL, Havenith CE, van den Born J, Ter Wee PM, Beelen RH. Better preservation of peritoneal morphologic features and defense in rats after long-term exposure to a bicarbonate/lactate-buffered solution. J Am Soc Nephrol. 2001; 12: 2775-2786.
[80] Kondo S, Kagami S, Kido H, Strutz F, Muller GA, Kuroda Y. Role of mast cell tryptase in renal interstitial fibrosis. J Am Soc Nephrol. 2001; 12: 1668-1676.
[81] Bird SD, Walker RJ. Mast cell histamine-induced calcium transients in cultured human peritoneal mesothelial cells. Perit Dial Int. 1998; 18: 626-636.
[82] Alscher DM, Braun N, Biegger D, Fritz P. Peritoneal mast cells in peritoneal dialysis patients, particularly in encapsulating peritoneal sclerosis patients. Am J Kidney Dis. 2007; 49: 452-461.
Publish with ROSS Science Publishers and every scientist can easily read your work for free!
Your research papers will be: • available for free to the entire scientific community • peer reviewed and published immediately after
acceptance • cited in renowned open repositories upon
indexation of the journal • owned by yourself — author keep the copyright







