

Introduction
~. INTRODUCTION
1.1 CANDIDA AND CANDIDIASIS
It was as early as 1839 that J. L. Schoenlein isolated the
fungal strain Achorion schoenlein (today Trichophyton
schoenleinil) from a patient suffering of favus (honeycomb
ringworm), that the first case of human fungal infection came
into picture. Today, Candida is the most common cause of opportunistic mycoses
worldwide. It is also a frequent colonizer of human skin and mucous membranes
(Odds 1988; Thomas 1993). The genus Candida includes around 154 species.
Among these, six are most frequently isolated from human infections. While
Candida albicans is the most abundant and significant species, Candida
tropicalis, Candida glabrata, Candida parapsilosis, Candida krusei, and Candida
lusitaniae are also isolated as causative agents of Candida infections (Table 1.).
Importantly, there has been a recent emergence of infection of non-albicans
Candida species, such as Candida glabrata and Candida krusei (Hitchcock et al.
1993; Katiyar and Edlind 1998; Vanden Bossche et al. 1992). Nevertheless, the
diversity of Candida species that is encountered in infections is expanding and
the emergence of other species that were rarely in play in the past is now likely.
Table 1. Species Commonly Causing Invasive Candidiasis . . .,~
Candida albicans Candida tropicalis Candida glabrata Candida parapsilosis Candida krusei Candida lusitaniae
*~"'_UI'-' CY .......
50% 15-30% 15-30% 15-30%
-1% -1%
From: http://www.doctorfungus.orglindex.htm
Bloodstream infections with Candida species have increased and these
organisms account for 10% of all nosocomial bloodstream isolates (Odds 1988).
This incidence equals that of Escherichia coli and surpasses Klebsiella species.
In addition to hematogenously disseminated Candidal infections, mucocutaneous
1
Introduction
Candidal infections are becoming increasingly problematic (Table 2), especially in
patients with the acquired immunodeficiency syndrome (AIDS); 80% of these
patients have Candidal infections.
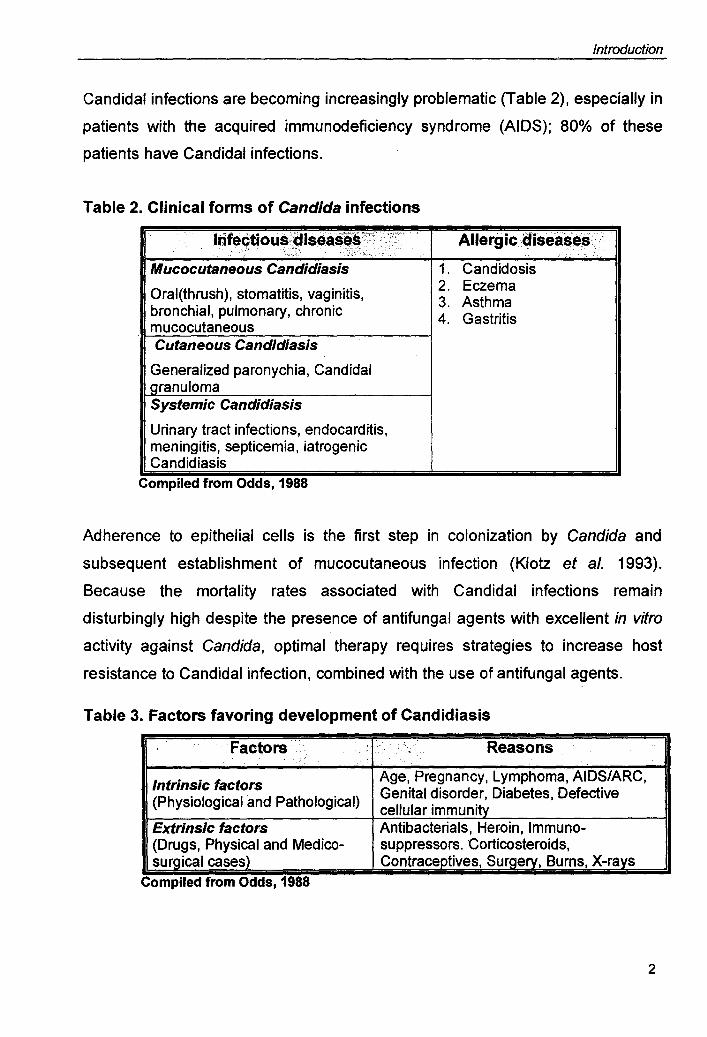
Table 2. Clinical forms of Candida infections
Irifectious;:diseases'X~< :\~,;, Allergic diseases; .. ~: <-r~. .... ,:;;;:: .. ,' ..
'\' ' '" I. "
Mucocutaneous Candidiasis 1. Candidosis Oral(thrush), stomatitis, vaginitis, 2. Eczema
3. Asthma bronchial, pulmonary, chronic 4. Gastritis mucocutaneous Cutaneous Candidiasis
Generalized paronychia, Candidal granuloma Systemic Candidiasis
Urinary tract infections, endocarditis, meningitis, septicemia, iatrogenic Candidiasis
Complied from Odds, 1988
Adherence to epithelial cells is the first step in colonization by Candida and
subsequent establishment of mucocutaneous infection (Klotz et al. 1993).
Because the mortality rates associated with Candidal infections remain
disturbingly high despite the presence of antifungal agents with excellent in vitro
activity against Candida, optimal therapy requires strategies to increase host
resistance to Candidal infection, combined with the use of antifungal agents.
Table 3. Factors favoring development of Candidiasis
Factors " ,,:' <' ~. ., Reasons "
Intrinsic factors Age, Pregnancy, Lymphoma, AIDS/ARC, (Physiological and Pathological) Genital disorder, Diabetes, Defective
cellular immunity Extrinsic factors Antibacterials, Heroin, Immuno-{Drugs, Physical and Medico- suppressors. Corticosteroids, surgical cases) Contraceptives, Surgery, Burns, X-rays . Complied from Odds, 1988
2

Introduction
Infections caused by Candida species are in general referred to as Candidiasis.
The clinical spectrum of Candidiasis is extremely diverse (Fig. 1). Almost any
organ or system in the body can be affected (Odds 1988). Candidiasis may be
superficial and local or deep-seated and disseminated (Thomas 1993).
Disseminated infections arise from hematogenous spread from the primarily
infected locus. C. albicans is the most pathogenic and most commonly
encountered species among all. Its ability to adhere to host tissues, produce
secretory aspartyl proteases and phospholipase enzymes, and transform from
yeast to hyphal phase are the major determinants of its pathogenicity. Several
host factors as listed in Table 4, predispose to Candidiasis.
Table 4. Table showing the host factors which promote Candida infection.
;P'REDISPOSING'FAC:rO~ ~?;":~~:n!?'~~0~;,i~~c::,.-~~~PL.;~~:'. ,
:~~~"':';'~ .": ,
".," "., " >, .' .. '<>:' 'I
Physiological Pregnancy, age (elderly and infancy) Trauma Maceration, infection, burn wound
Hematological Neutropenia, cellular immunodeficiency (leukemia, lymphoma, AIDS, aplastic anemia)
Endocrinological Diabetes Mellitus, hypoparathyroidism, Addison's disease
Iatrogenic Chemotherapeutics, corticosteroids, oral contraceptives, antibiotics, catheters, surgery
Other Intravenous drug addiction, malnutrition, malabsorption, thymoma
From: http://www.doctorfungus.org/thefungl/Candlda_spp.htm
Several factors contribute to the development of clinical resistance. First and
foremost is the underlying state of the infected individual's immune system which
has profound effects on the clinical outcome while treating fungal infections.
Azole resistant candidiasis occurs frequently in AIDS patients whose immune
systems are particularly debilitated. This correlation appears to be independent of
the individual's cumulative dose of fluconazole. Other medical complications
contribute to antifungal drug resistance.
3

Fig. 1. Candida albicans cells in various morphological forms. (a) Thick walled chlamydospore production on corn meal agar plate. 400X, 25C. (b) Yeast colony. Sabouraud glucose agar, 30°C. (c) Yeast cells and pseudohyphae in smear of heart tissue. Gram stain, 1000X (d) Yeast cells and pseudohyphae in material from the oral cavity, KOH preparation, phase-contrast microscopy

Introduction
1.2. TYPICAL CHARACTERISTICS OF CANDIDA
1.2.1. CANDIDA AND DIMORPHISM
On a petridish the fungus, C. albicans looks as harmless as common baker's
yeast. But when it encounters our cells, the fungus morphs
into a frightening foe, spreading out filamentous arms that
can impale immune cells and overpower unsuspecting hosts.
The result is life-threatening infections in AIDS and
chemotherapy patients.
Dimorphism is the ability to produce either separated yeast cells (blastopores) or
filamentous forms (hyphae and pseudohyphae) (Kerridge 1993). Switching
between different growth forms is considered an important virulence trait of many
pathogenic fungi. C. albicans is able to grow as budding yeast, but depending on
environmental conditions, growth can also occur in a filamentous form. Filaments
may be characterized as true hyphae, if they arise by continuous apical growth at
the filament tip; thereby unconstricted filaments are formed, which subsequently
are divided into mononucleate cells by septae that develop lateral buds or
branches, by budding hypha-like filaments are formed. Hyphal induction of
Candida proceeds in three phases (Fig. 2). In the first phase external signals are
sensed by specific receptors on the cell surface which in the second phase
activate intracellular signal transduction pathways. In the third phase the
structural and regulatory components necessary for the formation of the hyphal
form are produced, these include proteins catalyzing hyphal specific components
of the cell wall, as well as the proteins modulating the cell cycle.
4

Signals
(d)
Fig 2. Hypothetical model indicating that different microenvironments within the infected host exposing Candida albicans to a variety of signals which presumably lead to the differential activation of several distinct signaling pathways. The microenvironments shown, the brush border epithelium as well as the bloodstream, represent only two such microenvironments in which C. albicans can thrive within the host. Some signals will be specific to some microenvironments (A and C), whereas others will be common to several microenvironments (B). There is probably some overlap in the specificity of the signal transduction pathways with respect to their inputs as well as some redundancy in their outputs (multiple arrows). The outputs include (a) yeast to hyphal morphogenesis, (b) adherence to host tissue, (c) production of hydrolytic enzymes, (d) thigmotropism.[From Brown and Gow, 1999]

Introduction
Table 5. showing the transcript:on factors regulating yeast to hyphal
transition
FACTOR 1 ROLE +1-1 SIGNAL 1 INTERACTION
Efg1p + cAMP Disruption blocks serum-induced hyphae when coupled with deletion of CPH1 Disruption blocks activated Rim 101 p-induced pH-independent hyphae
Cph1p + Nutrients? Disruption blocks serum-induced hyphae when coupled with deletion of EFG1 Disruption blocks embedded-growth-induced hyphae when coupled with deletion of CZF1
Tup1p - ?
Czf1p + Embedded Disruption blocks embedded-growth-induced growth hyphae when coupled with deletion of CPH1
Tec1p + Nutrients?
Rim101p + pH Activated alleles need Efg 1 P for pH-independent hyphal induction
Rbf1p - ? .. +: null mutation mhlblts filamentation; - null mutation enhances filamentation, Taken from Whiteway, 2000
There are several signaling pathways leading to hyphal development (Fig. 3).
1.2.1.1. Efg1p pathway
EFG1 was discovered in a screen for C. albicans genes, whose expression in the
yeast S. cerevisiae increases filamentation (pseudohyphal growth) in this
organism (Stoldt et al. 1997). The designation EFG1 (enhanced filamentous
growth) was chosen because overexpression of Efg1 also led to a stimulation of
pseudohyphal growth in C. albicans. EFG 1 encodes a protein that belongs to a
conserved class of morphogenetic regulators in fungi (APSES proteins). A double
disruptant of Efg 1 P fails to form hyphae. With regard to virulence, the efg1 and
cph1 double mutant was almost completely avirulent in the mouse model of
systemic infection (Fig. 4) (Braun and Johnson 2000). The molecular
mechanisms by which Efg1p exerts its functio'n are not known. Autoregulation of
EFG1 expression may adjust cellular Efg1p levels, which may directly determine
the morphology of C. albicans.
5

- • • -+ + + + Histidine RimSp kinases Rim20p
+ + Rim101p Efg1p
~ YEAST HYPHAE
Cph1p Tec1p
t t t t
Fig. 3. The various signaling pathways leading to the transition from bud to hyphae in C. albicans. [From Whiteway, 2000]

Introduction
1.2.1.2. MAP Kinase Pathway
A MAP kinase module containing the kinases Ste20p, Ste11p, Ste7p and Kss1p,
which activates the
transcription factor
Ste12p, is essential for
pseudohyphal
morphogenesis of S.
cerevisiae (Madhani
and Fink 1997). In C.
albicans homologous
components of this
MAP kinase pathway
include Cst20p (Malathi
et al. 1994) (Ste12
homolog),
(Bardwell et
(Ste7p
Hst7p
al. 1998)
homolog),
Cek1p (MAP kinase
homolog of Kss 1 p)
(Csank et al. 1998) and
Cst20
e Hst7
RCAIIENTATION AND VIRULENCE
Fig 4. Signaling cascades regulating virulence of Candida albicans. Two parallel signaling cascades involving a MAP kinase and a cAMP-PKA signaling pathway regulate filamentation and virulence of this human fungal pathogen. AC: adenylate cydase. [From: Lengeler et al., 2000]
Cph1p (Liu et al. 1994) (Ste12p homolog). In contrast to the strong dependence
of S. cerevisiae on the MAP kinase pathway, this evolutionary conserved
cascade is not absolutely required for the hyphal morphogenesis and virulence of
C. albicans. Mutants lacking the MAP kinase components viz. Cst20p, Hst7p and
Cph1p show normal hyphal development in liquid medium and during serum
induction (Fig . 5) , but reduced hypha I formation on spider medium. Hst7 mutants
are fully virulent in the mouse infection model, while cst20 and cph1 mutants only
show attenuated virulence.
6

Introduction
1.2.1.3. cAMP Pathway
Exogenous cAMP stimulates hyphal growth of C. albicans suggesting a role for
cAMP and protein kinase A (PKA) in hypha I morphogenesis. Strong evidences
for the involvement of PKA in morphogenesis has recently been obtained by
~ Inducers: ~ ~~um,GI(;N~
• ?
? Ras1 • ?
? cAMP
?
~? Tpk2
~? Hst7 Tpk2 Cek1
Prr1
~ ?
~ TT ~ " ./ v V"
Fig. 5. A schematic showing the various cues which lead to fonnation of true hyphae in Candida albicans. The involvement of various signaling pathways is also shown. [From Ernst, 2000]
analysis of the CaTPK2 gene in C. albicans (Sonneborn et al. 2000). CaTPK2
encodes a protein highly homologous to the PKA isoform encoded by S.
cerevisiae. Overexpression of Ca TPK2 induces an elongated cellular form in C.
albicans, which resembles a germ tube. As expected , deletion of TPK2
significantly delays hyphal development. Heterologous expression of a C.
albicans gene, (CaCCT8), encoding an individual subunit of the cytosolic
chaperonin complex in S. cerevisiae, blocks pseudohyphal morphogenesis and
also blocks all phenotypes of an activated ras pathway, such as heat sensitivity,
lack of sporulation and lack of glycogen/trehalose accumulation. In C. albicans
overexpression of CaCCT8 also blocks morphogenesis in C. albicans suggesting
that PKA is involved in hyphal development of this fungus as well.
7

Introduction
1.2.1.4. Other regulators of morphogenesis
A gene encoding a homolog of Tup1 of S. cere visiae , which is a transcriptional
suppressor of more than 60 genes, was cloned in Candida (Braun and Johnson
1997). tup1 mutants show strong filamentous growth, mostly in the form of
pseudohyphae, which did not transform into true hyphae upon addition of serum.
In addition to the filamentous growth, tup1 mutant also showed additional
phenotypes compared to the wild type, such as misshapen cell walls and faster
growth on glycerol, indicating multiple regulatory effect of Tup1p. The Rbf protein
of C. albicans has functional similarities to S. cerevisiae Rap1 p in that it binds to
the regulatory RPG box and to telomeric repeat sequences (Ishii et al. 1997).
Deletion of RBF11ed to an altered cell form, presumably a pseudohyphal growth
form.
Table 6. Morphogenetic regulators of Candida albicans
EFG1
CPH1
CST20
Forms pseudohyphae, unable to form true hyphae Partial defect in hyphae formation Partial defect in hyphal formation
CaRSR1 Defective in hypha I formation
HSTl
CEK1
TPK2
CAP1
FLOB
TUP1
CLA4
Partial defect in hyphal formation Defective in hyphal formation Delays hyphal formation Oe!aY!5 hyphal formation Reduces filamentation Constitutive hyphal formation Unable to form hyphae under all tested conditions
Compiled from Emst and Schmidt, 2000
Virulen(:e Putati", Junction ..... /
Avirulent Transcriptional activator and repressor
Attenuated Transcription factor regulated virulence by MAPKinase Attenuated Protein kinase in MAP Kinase virulence pathway Attenuated RAS related (as S. cerevisiae virulence BUD1)
Virulent Protein kinase in MAP Kinase pathway
Avirulent MAP kinase
? Protein kinase A
Avirulent Cyclase-associated proteins
? Re_9ulates the FL011-.9..ene
Virulent Transcriptional repressor
Avirulent Protein kinal)e in MAP kinase pathway
8

Introduction
Several other genes required for hyphal morphogenesis and other cellular
functions have been characterized recently. Disruption of CaCLA4 in C. albicans
leads to a mutant unable to form filaments as well as misshapen cell forms
(Leberer et al. 1997). Deficiency of mannosyl transferase (CaPmt1 p) renders cell
incapable of forming hyphae on spider media, while still permitting
morphogenesis on serum (Timpel et al. 1998). Deletion of the CaKEX2 gene
encoding an endoproteinase of C. albicans lead to defective hyphal
morphogenesis (Newport and Agabian 1997), as well as an abnormal cell form:
thus CaKEXp may not be a specific component of morphogenetic pathways.
1.2.2. DIMORPHISM AND VIRULENCE
Both yeast cells and hyphae are found at the sites of infection. FungalInfections There are three observations, which led to the specific
hypothesis that filamentation, is required for virulence. First,
filament formation is stimulated at 37°C by either serum or
neutral pH «6.5); these circumstances mimic the host environment (Mitchell
1998). Second, newly formed filaments (called germ tubes) are more adherent to
mammalian cells than are yeast cells, and adherence certainly must set the stage
for tissue penetration. Third, yeast cells taken up by.· macrophages produce
filaments and lyse the macrophage, therefore filament formation provides a
mechanism for evasion of this host defense mechanism.
Filamentation and virulence are linked through a surface protein Int1 p (Gale et al.
1998), as well as through the MAPK cascade and Efg 1p. Int1 p has a positive role
in filamentation, because a C. albicans Int1 p-mutant has a medium-dependent
filamentation defect. The Int1 p-mutant also has reduced virulence in
intravenously inoculated mice. Expression of INT1 from a strong promoter in S.
cerevisiae (which lacks an Int1 homolog) causes formation of elongated
projections that resemble germ tubes. Therefore,lnt1 p may activate a conserved
filamentation regulatory pathway.
9

Introduction
1.2.3. HIGH FREQUENCY PHENOTYPIC SWITCHING IN C. ALB/CANS
C. a/bicans switches heritably and at a high frequency between at least seven
general phenotypes identified by colony morphology on agar (Soli et a/. 1993) as
seen in Fig. 6. Spontaneous conversion from the original smooth to variant
phenotypes (star, ring, irregular wrinkle, stipple and fuzzy) occurs at a combined
frequency of 1.4x1 0-4, but is increased 200 times by a low dose of UV. Switching
is therefore heritable, but also revertible at high frequency. The high frequency of
switching as well as complete reversibility suggests that reversible genetic
changes for instance, changes in the location of the mobile genetic elements may
be basic to the switching mechanism although heritable extrachromosomal
changes have not been ruled out. The differences in colony morphology appear
to be due to spatial, temporal, quantitative differences in bud and mycelium
formation (Berger et a/. 1990; Soli 1997). All of the seven switch phenotypes
retain the basic capacity of dimorphism and are therefore capable of forming
buds and mycelia, but the environmental constraints, such as pH regulated
dimorphism on the transitions between bud and hyphal forms vary markedly
between o-smooth
and switch
phenotypes.
Switching may
provide C. a/bicans
and related
infectious yeasts
with the diversity
that is expected of
such pervasive
and successful
pathogens.
Switching may
provide an
organism with the
~ White-phase specific,
bud-specific, trans-acting factors
Convergence of t regulatory circuitry
+ DAD PAD
Fig. 7. A model of the regulatory circuitry emanating from the switch event and dimorphism event. There seems to be a convergence to regulate the distal activation domain (DAD) and the proximal activation domain (PAD) of the phase specific gene WH11. From Soli et aI, 1993.
10

Fig. 6. Various colony phenotypes in the switching system of C. albicans. (A) O-smooth (8) Star (C) Ring (0) Irregular wrinkle (E) Stipple (F) Hat (G) Fuzzy (H) r-smooth. Adapted from Soli, 1990

Introduction
capacity to (1) invade diverse body locations (2) evade the immune system in a
fashion similar to Salmonella and trypanosomes or (3) change antibiotic
resistance.
A number of indirect observations strongly suggests that switching involves the
regulation of a number of diverse genes, that these genes are unlinked and that
there is no genomic reorganization at the regulated loci. In the case of white-
opaque transition, both a white-phase-specific gene, WH11, and an opaque-
phase-specific gene, OP4 (Morrow et al. 1993), are regulated through upstream
cis-acting transcription activation domains. In the case of WH11, white-phase-
specific factors have been demonstrated to bind to the two activation domains
(Srikantha et al. 1997). This suggests that the switch event in each direction
leads to deactivation of trans-acting factors specific to the 'switched-form' phase
and activation of trans-acting factors specific to the 'switched-to' phase.
Mass conversion experiments point to a single master switch, which gets
activated at either a low (SoC) or high temperature (390C). More importantly it was
demonstrated that co-incident with phenotypic commitment and the second cell
doubling, the transcription of white specific gene, WH11 is activated. Both OP4
and PEP 1 (opaque-phase specific genes) transcription is deactivated
immediately after the temperature is increased. Therefore, the point of phenotypic
commitment marks a major regulatory event both for the activation of WH11 and
the termination of the opaque gene inducibility, and this regulatory event, which is
believed to represent the master switch, coincides with the second cell doubling
(Fig. 7).
1.3. MECHANISM OF ACTION OF ANTIFUNGAL DRUGS
A breakneck increase in fungal infections has stimulated the search for
antifungals. The treatment of deeply invasive fungal infections has lagged behind
bacterial infections, as there are substantially fewer antifungals than antibacterial
drugs. It is important to note here that the fungal cells are more similar to human
cells than the bacteria and therefore it is necessary to look for drugs, which would
exciusively act on the fungal cells and not harm the human host.
11

Introduction
It is interesting to note that, unlike antibacterial drugs where the prinicipal target is
protein synthesis (the ribosome) and peptidoglycan synthesis (the cell wall) , the
MEMBRANE FUNCTION Polyenes: Amphotericin B Nystatin Primaricin
Cispentacin Difluoromethyl Ornithine
5-fluorocytosine Trimethoprim Sulfomethaxozole Pentamidine
CELL WALL SYNTHESIS Polyoxins Nikkomycins Echinocandins
ERGOSTEROL SYNTHESIS Azoles Allylaminesl Thiocarbamates Morphines
Griseofulvin Benomyll Benzimidazoles
Blasticidin, Sinefungin
Fig. 8. Mechanism of action of various classes of drugs on a typical fungal cell . [From Georgopapadakou and Walsh (1994)]
major targets for clinically important antimycotic drugs are the nucleus and
cellular membranes (Fig. 8). Physicians are desperate to find new ways to
combat fungal infections; however; developing broad-spectrum antibiotics against
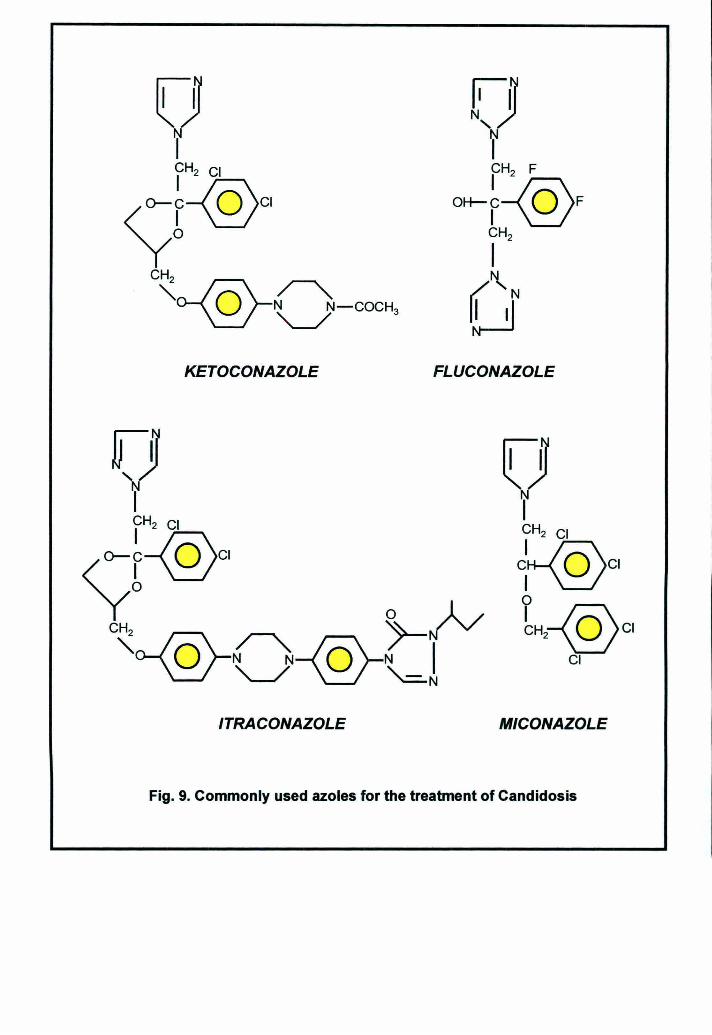
fungi is difficult and chiefly azoles are used to combat the fungal infections (Fig.
9). Even the few drugs that are available are of little use to combat emerging
drug-resistant strains. Adding new and improved drugs to the arsenal is therefore
important. In addition, developing drugs against new targets is crucial to remain a
step ahead of the emerging microbes. Table 7 summarizes the antifungal sin use
and the mechanism of resistance.
12
KETOCONAZOLE FLUCONAZOLE
ITRACONAZOLE MICONAZOLE
Fig. 9. Commonly used azoles for the treatment of Candidosis

Introduction
Table 7. Targets and mechanisms of resistance of some antifungals 1
Purirr,il'liine
5- Flucytosine
Polyenes Nystatin Amphotericin B
Azoles Fluconazole Ketooonazole Itraconazole Voriconazole Clotrimazole
Allylamines Naftifine Terbinafine Tolnaftate
Morpholines Amorolfine
Lipopeptides Echinocandins Pneumocandins Aculeacins
Membrane ergosterol
14a-demethylase(ERG 11, also designated as ERG16 earlier)
Squalene epoxidase (ERG 1)
d 14_ reductase(ERG24),d 8,7_
isomerase (ERG2)
j3-1,3-glucan synthetase (encoded by FKS1 and RHO 1
Compiled from (Marichal
~ Alteration in membrane lipids, mainly ~ Ergosterol (resistant clinical isolates
lack ergosterol and accumulate 3-(3-ergosta- 7,22-dienol and 3-13- ergosta-8-enol, due to defect in d 5,6_
desaturase gene (ERG3) ~ Enhanced catalase activity
~ Mutations in the target enzyme cytochrome P450 14a-demethylase which alters the affinity of this enzyme to the azoles
~ Overexpression of 14a-demethylase ~ Failure to accumulate azoles due to
rapid efflux mediated by ABC and MFS family of MDR transporters
~ Alteration of sterol d 5,6 -desaturase (ERG3)
~ Overexpression of CDR1, CDR2 and CaMDR1
~ Overexpression of /). 14-reductase (ERG24) .or sterol C-24 (28) reductase (ERG4) genes
~ Overexpression of CDR1 and CDR2 » Mutations in FKS1gene alters affinity
of the enzyme
1998).
13

Introduction
1.4. MUL TIDRUG RESISTANCE
Multidrug resistance is defined as resistance against a broad spectrum of drugs
that share neither a common target nor a common structure. The development of
drug resistance is a frequent impediment to the effective treatment of infectious
and malignant diseases (Peters 1996; Sternberg 1994; Van denbroucke-Grauls
1993). A primary goal in the study of chemotherapy is to understand how cells
can become drug resistant by lowering the intracellular concentration of drugs! or
altering the ability of the drugs to affect their targets. Mechanisms of multidrug
resistance are opportunistic in their manipulation of the normal pathways of
cellular homeostasis. Several alterations in the cell take place resulting in drug
resistance.
Many different resistance mechanisms have been described, but those that
involve proteins belonging to the ABC transporter superfamily have been of
particular interest because of the increasingly prominent role these proteins play
in such devastating and widespread diseases as malaria, leishmaniasis and
cancer (Foote et a/. 1989; Ouelette and Papadopoulou 1993; Skovsgaard et a/. 1994).
1.4.1. FUNGAL RESISTANCE
A drug-resistant, pathogenic fungus is an organism, according to Kerridge et a/.
that will grow and produce clinical symptoms of disease in the presence of the
drug at the maximal concentration, at the site of infection. From a mycological
viewpoint different classifications of resistance are made.
1.4.1.1. Intrinsic resistance
A species is regarded as intrinsically resistant when it is not included in the
normal spectrum of a given antifungal compOlind. For example, Candida krusei
can be regarded as intrinsically resistant to fluconazole and C. parapsi/osis to
Amphotericin B.
14

Introduction
1.4.1.2. Acquired resistance
Acquired resistance is found in isolates belonging to a species which is normally
susceptible to the compound.
1.4.1.3. Selective resistance
When a patient is colonized with multiple species or strains, during treatment the
most sensitive isolates are eradicated favoring the growth and selection of less
sensitive or resistant isolates. This type of resistance is called selective
resistance.
1.4.1.4. Phenotypic resistance
This occurs when a strain develops a progressive increase in resistance during
continued incubation in the presence of antifungal compound.
1.4.1.5. Genotypic resistance
It is a feature of the clone and is inherited by daughter cells.
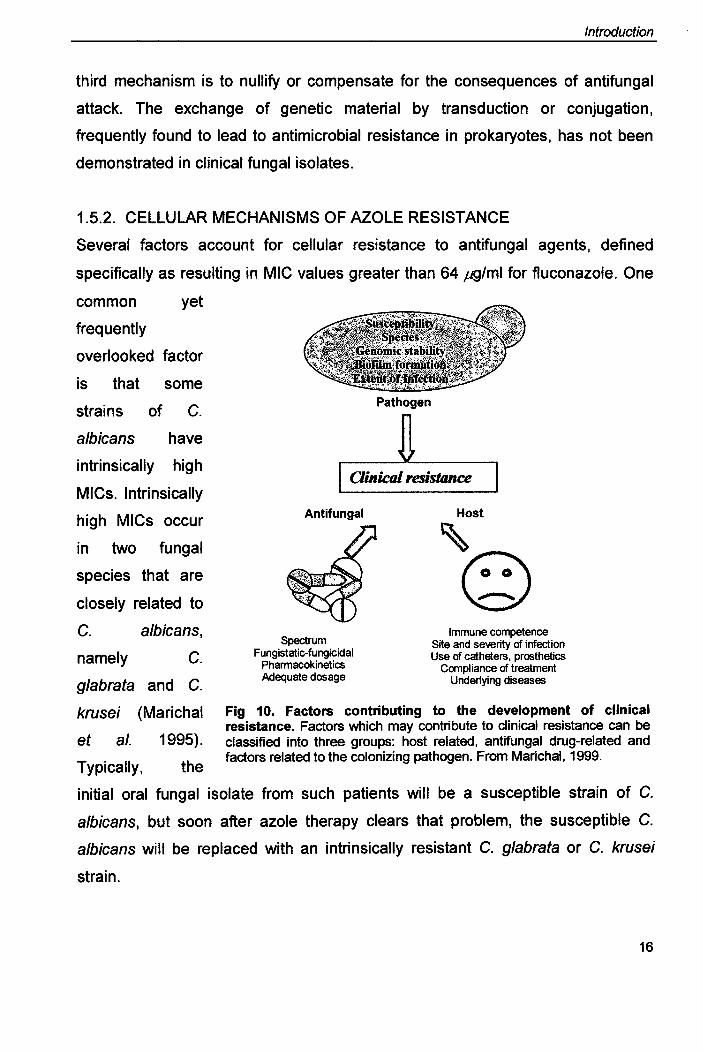
1.5. MECHANISMS CONTRIBUTING TO FUNGAL RESISTANCE
It is clear that the origin of antifungal resistance is multifactorial and results from a
combination of circumstances related to the host, the antifungal agent and the
pathogen (Fig. 10). Fungal resistance can be broadly attributed to biochemical,
cellular and molecular mechanisms (White 1997a).
1.5.1. BIOCHEMICAL MECHANISM OF RESISTANCE
From the biochemical point of view, factors which lead to a measurable decrease
in the susceptibility of a pathogen to an inhibitory agent in vitro is attributed to the
pathogen itself. The fungal pathogen can adopt three general mechanisms in
order to become less susceptible to antifungal attack. The first mechanism
prevents an adequate amount of active antifungal from reaching its target. The
second mechanism involves interference with the structure or quantity of the
antifungal target, thus altering the stoichiometry of the inhibitory effects and the
15
Introduction
third mechanism is to nUllify or compensate for the consequences of antifungal
attack. The exchange of genetic material by transduction or conjugation,
frequently found to lead to antimicrobial resistance in prokaryotes, has not been
demonstrated in clinical fungal isolates.
1.5.2. CELLULAR MECHANISMS OF AZOLE RESISTANCE
Several factors account for cellular resistance to antifungal agents, defined
specifically as resulting in MIC values greater than 64 ,uglml for fluconazole. One
common yet
frequently
overlooked factor
is that some
strains of C.
albicans have
intrinsically high
MICs. Intrinsically
high MICs occur
in two fungal
species that are
closely related to
C. albicans,
namely C.
glabrata and C.
krusei (Marichal
et al. 1995).
Typically, the
Antifungal
Spectrum Fungistatic-fungicidal
Pharmacokinetics Adequate dosage
Pathogen
Host , ~
Immune competence Site and severity of infection Use of catheters, prosthetics
Compliance of treatment Underlying diseases
Fig 10. Factors contributing to the development of clinical resistance. Factors which may contribute to clinical resistance can be classified into three groups: host related, antifungal drug-related and factors related to the colonizing pathogen. From Marichal, 1999.
initial oral fungal isolate from such patients will be a susceptible strain of C.
albicans, but soon after azole therapy clears that problem, the susceptible C.
albicans will be replaced with an intrinsically resistant C. glabrata or C. krusei
strain.
16

Introduction
c. albicans strains usually persist for long periods as commensals. Only rarely in
the abS~j--e of azole drug treatment are such strains replaced with other strains,
whereaF! ;train replacement occurs as often as 40% of the time when antifungal
drugFesistance enters the picture.
Table 8. Cellular mechanisms of antifungal resistance
Intrinsic resistance of endogenous strains Replacement with a more resistant Candida species Replacement with a more resistant C. albicans Genetic alteration that results in a more resistant strain Transient that renders a cell resistant
Alternatively, azole resistance sometimes is transient-apparent when fluconazole
is present but gone soon after the drug is removed. The transient or epigenetic
resistance is most likely the result of an increase or decrease in expression of a
gene important for drug resistance. Epigenetic expression is particularly difficult
to study in clinical setting since the trait disappears as soon as the clinical isolate
is removed from the patient.
1.5.3. MOLECULAR MECHANISMS OF ANTIFUNGAL RESISTANCE
How azoles enter susceptible fungal cells is not known, although their relative
hydrophobicity may facilitate entry by passive diffusion. Once inside the cells,
azoles interact with the 14a-DM enzyme in the ergosterol biosynthetic pathway,
allowing precursors to be incorporated into newly synthesized regions of plasma
membrane (Barrett-Bee and Dixon 1995). Not a!! of the azole entering the cell
remains there, because two low-level active efflux systems, the ABC transporters
and the major facilitators, pump free drug from the cell.
17

Table 9. Mechanisms (molecular) leading to antifungal resistance
.f ~~~~~~~~~~?·f~~,~,~!!u~'~~~·::R~~istance Change in sterol components of the plasma membrane Genetic changes in ERG16 including:
Point mutations Overexpression Gene amplification (leads to overexpression)
Alterations in other enzymes in ergosterol biosynthetic pathway Increased efflux of drug due to:
ABC transporters (CDR genes) Ma·or Facilitator MOR1 ene
Introduction
Presumably azole enters resistant cell in the same way as they ordinarily enter
susceptible cells. However, if the sterol composition of the cell membrane is
altered, perhaps the usual uptake process also becomes altered (Aoyama et al.
2000). Thus, for example, changes to other genes in the ergosterol biosynthetic
pathway may also affect a cell's susceptibility to azoles. Once the antifungal
molecules enter a cell, their interaction with the target enzyme,14DM, can be
modified in at least two ways. First, certain point mutations in the gene for 14DM
make the enzyme less sensitive to azole drugs (Favre et al. 1999). In addition to
point mutations, enzyme overexpression, which lead to more of the azole target
molecule per cell, necessitates higher doses of drug to achieve inhibitory effects
comparable to those seen in susceptible cells.
Although each of these factors contributes to azole resistance in one or the other
strain of C. albicans, independent clinical isolates, rarely, if ever, exhibit all of the
changes that have been identified so far, and still other mechanisms undoubtedly
have yet to be identified. It is also known that a combination of mechanisms may
contribute at a particular infection.
18

Introduction
1.6. PUMPS AS PROMINENT MECHANISM OF MDR
Efflux mechanisms, which reduce the cytoplasmic concentration of drugs and
other small molecules, are the major factor affecting a cell's
susceptibility to azoles. The ABC transporters (Keppler et al.
2000) and the Major Facilitators (Pao et al. 1998) are two
classes of efflux systems, which contribute to MOR by
transporting out drugs and can be regarded as a "first-line
defense barrier" in survival mechanisms.
The ATP-binding cassette (ABC) superfamily, also called the traffic ATPases,
comprises proteins, the majority of which mediate the selective transport of
substrates across biological membranes (Table 10 lists traffic ATPases found in
yeasts). ABC proteins are found in all organisms, including prokaryotes and
eukaryotes (Fig. 11). Table 10 lists the traffic ATPases found in yeasts. The
prototypical ABC protein is large ( .... 140 kOa) and contains four modules: (two
nucleotide binding domains (NBOs) that bind and hydrolyze ATP and two
membrane spanning domains (MSOs), each containing multiple (6)
transmembrane
segments (Fig. 12).
It is becoming
increasingly clear that
ABC proteins play a
significant role in
human health and
diseases. Several
genetic diseases can
be attributed to defects
in ABC proteins (see
Table 11) (Taglicht and
Michaelis 1998).
Because of the clinical
significance of ABC
Fig. 12. Schematic picture of different ABC type proteins depending on their predicted topology. Nucleotide binding domains are depicted in circles. Two hydrophobic domains are depicted as black lines. [From Decottignies and Goffeau (1997)]
19

EmrAB HlyBD
Bmr CDR1 PDRS P-glycoprotein
TPP+ Thiolactomycin ADP+Pi ATP
B. subtilis E.coll C. alblcans S. cerevlslae H. sapiens
Fig 11. Topology of bacterial, lower eukaryote and human ABC proteins. For comparison, the use of same color indicates the homologous sequences in different MDR proteins. From Lewis, 1992.

Table 10. Inventory of traffic ATPases in yeasts.
:X.~~t , ' -; 'Size .-:..: ... - .~ ~ 'f:' " -,"; . <,,f;' :"3" :.i . '
'.Geriename Chr. Topology Function :·F;: .
, , ' ;,j
Saccharomyces ADP1 11/ 1049 TM-NBF-TMs Small molecule transport cerevisiae
ATM1 XIII 690 TMs-NBF Mitochondrial DNA maintenance BPT1IYLL015 XII 1559 (TMs-NBF)2 Transport of un1conjugated bilirubin into vacuole CAF16IYFL028 VI 289 NBF-NBF Non transporter ABC protein GCN20 VI 752 NBF-NBF Interaction with tRNA & GCN2 MOL 1 XII 695 TMs-NBF Transport of peptides? MDL2ISSH1 XVI 812 ™s-NBF Transport of peptides? NEW1IYPL226 XVI 1196 NBF-NBF Non transporter ABC protein PDR10 XV 1564 (NBF-TMs)2 Drug efflux pump PDR11 IX 1411 (NBF-TMsh Drug efflux pump? PDR12 XVI 1511 (NBF-TMsh resistance to water-soluble, monocarboxylic acids with
chain lengths offrom C-1 to G-7 PDR15 IV 1529 (NBF-TMs)2 Drug efflux pump PDR5 XV 1511 (NBF-™sh Drug efflux pump PXA1/SSH2I XVI 870 ™s-NBF P.oxidation PAL1 PXA2IPAT1 XI 853 TMs-NBF Interaction with PXA1, small molecule transport RLl1IYDR091 IV 608 (TMs-NBFh Non transporter ABC protein SNQ2 IV 1501 (NBF-™s)2 Drug efflux pump STE6 XI 1290 (TMs-NBFh a-factor export YBT1IYLL048 XII 1661 (TMs-NBF)2 Small molecule transport YCF1 IV 1515 (TMs-NBFh Cd2+ & mdr glutathione S conjugate pump YDR061 IV 539 - unknown YEF3ITEF31 XII 1044 NBF-NBF Stimulation of aminoacyl tRNA binding to ribosome EFC1 YEF3BI XIV 1044 NBF-NBF Protein synthesis YNL014 YER036 V 610 NBF-NBF Unknown, non transporter ABC protein YHL035 VIII 1592 (TMs-NBF)2 Small molecule transport YKR103 XI 1524 (TMs-NBF)2 Possible pseudogene YKR104 XI - (TMs-NBFh Small molecule transport YNR070 XIV 1333 (NBF-TMs)2 Small molecule transport YOL075 XV 1294 (NBF-TMsh Small molecule transport YOR011 XV . 1394 .~ .(NBF-I!Y1'~)'~"M_". §r.!l~!!~_~~~.!Jle ~~~~~P~~~ __ ., ____ . .... ___ ,,",,"., .... _. ~~_ , .. ~,., ." .. , '''-,_ .... ". _ ..... '" . , ~, .... ... ,-"' :",....~ ~ .... ,,-"' ..... ,....-...... ,. ..... .. •• .,..: .. _ •• :..:t. •

· ....•... -_ .... ___ .,_.~. __ .,. ...... ,_, __ ... , .. __ ._ .. _._·._ .. · .... _.H· __ W.· ._ .. _._ ....... _ ... __ .~.. .. __ .... _ ..... _ .. _,.p .. _. ___ .. ~ ........................ _-........... __ ........ _ .................. ..... - ............. ; ............. ......................... , ...............
),OR1IYRS1 VII ' 1477 (™s-NBFh Oligomycin & mdr Schizosaccharomyces ABC1 II 1427 (™s-NBF)2 unknown pombe
BFR11HBA2 III 1530 (NBF-™sh Brefeldin A transport HMT1 III 830 TMs-NBF PhytochaloasinlCd++ transport MAM1 II 1336 (TMs-NBFh M-factor transport PMD1 III 1362 (TMs-NBFh Drug efflux pump
Candida albicans CDR1 III 1501 (NBF-™s)2 Drug efflux pump, phospholipid translocator CDR2 III 1499 (NBF-™s)2 Drug efflux pump,phospholipid translocator CDR3 IV 1501 (NBF-™sh Phospholipid translocator CDR4 I 1490 (NBF-™sh phospholipid translocator CDR5 VI ? Drug efflux pump? HST6 III 1323 (™s-NBFh Transport of a-factor, drugs? CaYOR1 ? NO ? Drug efflux pump? CaYCF1 ? 1606 ? Drug efflux pump?
Candida glabrata CgCDR1 ? 1499 (NBF-™sh Drug efflux pump CgCDR2 ? (NBF-™sh Drug efflux pump PDH1 ? 1542 (NBF-TMsh Drug efflux pump?
Candida dubliniensis CdCDR1 ? (NBF-™sh Drug efflux pump? CdCDR2 ? (NBF-™sh Drug efflux pump?
Candida krusei ABC1 ? ? Drug efflux pump ABC2 ? ? Drug efflux pump

Table 11. Traffic ATPases of yeasts and their homologues in human diseases
. YeastABC Homologues causing .... _, ..
1/: \ 'protein . disease . Organism Human health Jmpact ,
YCF1 CFTR Homo sapiens Cystic fibrosis PXA1 ALD Homo sapiens Adrenoleukodystrophy
PMP?O Homo sapiens Zellweger syndrome MDL1, MDL2 TAP1 Homo sapiens Behcet's disease, multiple sclerosis
TAP2 Homo sapiens Bare lymphocyte syndrome type 1 STE6 MDR1 Homo sapiens Cancer cells drug resistance YCF1, YLL015 MRP Homo sapiens Cancer cells drug resistance
SUR Homo sapiens Hyperinsulinemic hypoglycemia of infancy ATM1 ABC? Homo sapiens, Not known
Mus musculus ADP1 ABC8 Homo sapiens, Not known
Mus musculus PDR5, PDR10, CDR1 Candida albicans drug resistant candidosis PDR15 ATM1 PfMDR2 Plasmodium drug resistant malaria
falciparum STE6 EhPgp 1 Entamoeba drug resistant amoebiasis
histolyiica IdMDR Leishmania drug resistant kala-azar (visceral
donovani leish man iasis) SMDR2 Schistosoma drug resistant schistosomiasis
mansoni YER036 VgA Staphylococcus drug resistant wound infections, pneumoniae,
aureus impetigos MRP1 cMOAT Rattus norvegicus same phenotype as human Dubin-Johnson
syndrome (mild chronic conjugated hyperbilirubinemia

Introduction
proteins, it is critical to determine their role in no~mal cellular physiology and
disease. This presents a challenge, in that the normal physiological transport
substrate(s) for most ABC proteins are not known. For instance the normal role of
mammalian MOR1 could be for the clearance of xenobiotic compounds, as
suggested by knockout mouse; alternatively, it may transport an as-yet
undetermined native hormone (Ueda et al. 1992).
The study of ABC transporters in a tractable model organism such as yeast
provides a promising avenue of research for studying this issue.
The other family of efflux pumps are those belonging to the Major Facilitators
Superfamily (MFS). This family consists of over 50 transporters from bacteria to
higher eukaryotes involved in symport, antiport or uniport of various substrates
(Paulsen et al. 1996). These drug resistance proteins are PMF-dependent
antiporters which efflux out drugs exchanging one or more H+ ions with a
substrate molecule (Paulsen et al. 1998). The MFS type pumps involved in drug
resistance in yeast are S. cerevisiae FLR1 (Broco et al. 1999) and C. albicans
CaMOR1 (Gupta et al. 1998) and the recently cloned FLU1(Calabrese et 81.
2000).
1.6.1. PUMPS OF BUDDING YEAST INVOLVED IN MDR
S. cerevisiae is an ideal model organism for the functional
dissection of disease related genes such as those of the ABC
superfamily because of the ease of molecular manipulation of
genes in yeast. The completion of the entire genome
sequence of S. cerevisiae is a landmark achievement in modern biology
(Decottignies and Goffeau 1997). Through the functional studies of the yeast
ABC genes, together with genetic and biochemical analysis of the mammalian
ABC genes, heterologously expressed in yeast, a wealth of information can be
obtained that is directly applicable to understanding the role of the ABC proteins
in human health and disease.
20

Introduction
There are around 31
ABC proteins which are
known so far in S.
cerevisiae. (Table 10).
Yeast ABC proteins can
be classified on the
basis of sequence
similarity into six groups
designated the PDR,
ALDP, CFTR/MRP,
MDR, RLI and YEF3
subfamilies. These
designations reflect the
most prominent
mammalian (ALDP,
CFTR, MRP, MDR) or
Ste6p Yor1p Snq2p Pdr11p
yeast (PDR, YEF3) Fig.13. A generalized model of a yeast cell showing the members of each group. localization of various ABC proteins. From Bauer, 1999.
Two important features
that emerge in yeast ABC proteins is that the full length transporters predominate
over the half transporters and also not all ABC proteins contain membrane spans
e.g. YEF3 and GCN20 are known to be soluble proteins (Fig. 13).
PDR is the largest of the yeast subfamilies containing nine members (Balzi and
Goffeau 1995). The architecture of the PDR family members is distinct since they
are arranged in the reverse order (NBD1-MSD1-NBD2-MSD2). The
phenotypically best characterized PDR gene products are PDR5, SNQ2 and
recently YOR1. All the three genes are known to be involved in the phenomenon
of conferring drug resistance in yeast.
21

Introduction
1.6.1.1. PdrSp
The best-characterized yeast pleiotropic drug resistance ABC transporter is the .
product of the POR5 gene; PDR5 was isolated through its
property of conferring, upon amplification, resistance to
cycloheximide and sufomethuron methyl (SMM) (Balzi et al.
1994). Disruption of POR5 is not lethal but results in hypersensitivity to various
drugs such as cycloheximide, SMM and also to mitochondrial inhibitors
chloramphenicol, lincomycin, erythromycin and antimycin. The PDR gene
encodes a duplicated ABC protein consisting of the repeated alternance of two
hydrophilic domains and two hydrophobic domains, with six transmembrane
spans, each highly similar to the SNQ2 gene product. The POR5 gene transcript
is overexpressed in mutants of another pleiotropic drug resistance locus, PORt,
encoding a transcriptional regulator (Meyers et al. 1992). Subsequently the POR5
mRNA was found to be increased also in mutants of other POR loci, such as
POR3, also a transcriptional regulator homologous to POR1, as well as POR7
and POR9 (8alzi and Goffeau 1995; Dexter et al. 1994). The POR5 protein is
overexpressed in the plasma membranes of POR1 and POR3 mutants. This
shows that POR1 and POR3 factors control the expression of POR5 gene, the
product of which is responsible for the active pumping out of drugs, and possibly
other physiological substrates out of the cell. The PORt and POR3 mutations
found to confer drug resistance by hyperactivation of the expression of the mdr
pump POR5, represents the first reported identification of primary genetic sites of
lesions leading to overexpression of multidrug resistance pumps.
The PDR5 gene has been independently cloned and renamed STS1 and YDR1
respectively depending on their resistance to either sporidesmin, a mycotoxin
pathogenic for men and ruminants; or to cerulenin and cycloheximide (Bissinger
and Kuchler 1994). The PDR5 (STS1) transcript was reported to be reduced in a-
factor arrested cells indicating that the transcription of PDR5 might be under
hormonal control. The transcription of PDR5 (YDR1), as well as its homologue
SNQ2, was also shown to be induced by stress conditions, such as heat shock
and the presence of drugs. (Mahe et a/. 1995).
22

Introduction
1.6.1.2. Snq2p
Northern analyses performed after isolation of total RNA from a set of isogenic
strains harboring ,tjpdr1, ,tjpdr3 and ,tjpdr1,tjpdr3 deletion mutations (Mahe et al.
1996) demonstrated that SNQ2 mRNA level was severely hampered in the ,tjpdr1
strain while it was unaffected in the ,tjpdr3 strain. Nevertheless, PDR3 was
presumably involved in the regulation of SNQ2 because the SNQ2 mRNA was
even less in the double mutant strain compared to the single mutants. Identical
results were also obtained at the protein level. It was also demonstrated that a
bacterially expressed GST-Pdr3 fusion protein binds to the SNQ2 promoter at
three sites pointing towards a role of PDR3 in SNQ2 expression. (Mahe et al.
1995).
1.6.1.3. Yorlp
The oligomycin resistance gene, YOR1 was cloned on the basis of its ability to
strongly elevate tolerance to this compound when present in a high copy plasmid
(Katzmann et al. 1995). Recent experiments have also shown that this gene is
required for the resistance to the drug reveromycin A and that YOR1 transcription
is inducible by this drug. DNA sequence analysis of the YOR1 gene
demonstrated that this locus encodes a protein with striking sequence similarity
to the human cystic fibrosis transmembrane conductance regulator (CFTR) and
S. cerevisiae Ycf1 p (Cui et al. 1996). YOR1 is transcriptionally regulated by
Pdr1 p and Pdr3p and inspection of the YOR1 promoter region indicated the
presence of putative PDR11PDR3 response element located 215 bps upstream of
the transcription start site (Hallstrom and Moye-Rowley 1998). Although the
expression of both PDR5 and YOR1 are regulated by PDR1IPDR3, these two loci
do not exhibit common promoter structures. Both PDR5 and YOR1 contain
POREs, with PDR5 having three elements and YOR1 only one. Furthermore the
YOR1 PORE is identical to the second PORE site in the PDR5 promoter (Zhang
et al. 2001). PDR5 expression is strictly dependent on the presence of either
PDR1 or PDR3, whereas YOR1·expression is partially PDR1IPDR3-independent.
This finding suggests the presence of other positive regulatory elements for
23

Introduction
transcription in the YOR1 promoter. The second positive regulatory element for
YOR1 expression (UASYOR1) lies between positions -535 bp and -299 bp which
is under the negative control of URSYOR1 element at position -115 bp to -50 bp
region.
1.6.1.4. Ycf1p
The yeast cadmium factor gene (YCF1) from S. cerevisiae, which was isolated
according to its ability to confer cadmium resistance, encodes a 1,515 amino acid
ATP-binding cassette (ABC) protein with extensive sequence homology to the
human multidrug resistance-associated protein (MRP1) (Szczypka et al. 1994). It
encodes an MgATP-energized, uncoupler-insensitive vacuolar glutathione S-
conjugate transporter. YCF1 gene is required for YAP1 to exert its normal effects
on cadmium tolerance (Jungwirth et al. 2000). Mutant strains of yeast that lack
the YCF1 gene are hypersensitive to cadmium and this hypersensitivity is
epistatic to YAP1 overexpression. YCF1 mRNA levels and the expression of a
YCF1-lacZ reporter construct positively correlates with changes in YAP1 gene
dosage. A set of 5' truncation derivatives of the YCF1-lacZ fusion gene identified
the region from -201 bp to +47 bp as being sufficient for the YAP1-dependent
increase in expression. DNase I footprinting using a probe from this segment of
the YCF1 promoter showed that bacterially-produced YAP1 protein was capable
of binding a novel DNA element designated the YAP1 response element.
1.6.1.5. Flrlp
The adaptation of S. cerevisiae to growth in the presence of the antimitotic
fungicide benomyl involves the dramatic activation of FLR1 transcription, taking
place during benomyl-induced latency following sudden exposure to the
fungicide. FLR1 gene encodes a plasma membrane transporter of the major
facilitator superfamily (MFS) conferring resistance to multiple drugs, in particular
to benomyl (Alarco et al. 1997; Broco et al. 1999). FLR1 activation is completely
abolished in a mutant devoid of YAP1 gene. Effect of YAP-1 is presumably
exerted by Yap1 p either directly or via Pdr3p. The FLR1 promoter contains three
24

Introduction
potential Yap1p response elements (YREs) at positions -148 (YRE1), -167
(YRE2), and -364 (YRE3) (Nguyen et al. 2000). To address the function of these
YREs, the three sites have been individually mutated and tested in
transactivation assays. Results show that (i) each of the three YREs is functional
and important for the optimal transactivation of FLR1 by Yap1p and that (ii) the
three YREs are not functionally equivalent, mutation of YRE3 being the most
deleterious, followed by YRE2 and YRE 1. Simultaneous mutation of the three
YREs abolished transactivation of the promoter by Yap1 p, demonstrating that the
three sites are essential for the regulation of FLR1 by Yap1p.
1.6.2. EFFLUX PUMPS OF PATHOGENIC YEASTS
Aspergillus fumigatus is the most common species of Aspergillus causing
pulmonary disease. AmB and itraconazole are the two commonly used drugs to
which this fungus is sensitive (Denning et al. 1997) (Fig. 14). The role of
P45014DM and efflux pumps has been implicated in itraconazole resistance in A.
fumigatus. Recently two ABC transporter genes atrA and atrB have been cloned
from A. nidulans (Tobin et al. 1997). The proteins encoded by these genes share
the same topology as the ABC transporters PDR5 (from S. cerevisiae) and CDR1
(from C. albicans). Like the transcriptional activation of CDR1 by a variety of
stresses, atrA and atrB are also upregulated by several drugs, azoles and
fungicides (Del Sorbo et al. 1997). Another ABC transporter gene atrC has also
been cloned from A. nidulans. Northern analysis revealed that atrC mRNA levels
increased in response to cycloheximide. There are evidences suggesting the
presence of eight additional ABC pumps in A. nidulans. adr1, another ABC pump
has been characterized in A. fumigatus. The adr1 transcript is upregulated in
itraconazole resistant isolate, which also shows less accumulation of the drug.
ABC transporters, AfuMDR1 and AfuMDR2 from A. fumigatus and AflMDR1 from
A. flavus have also been identified.
CneMDR1, a gene encoding a protein related to MDR proteins has been cloned
and characterized from another pathogenic fungus, Cryptococcus neoformans
25

Fig. 14. The disease causing fungi 1. Stages in the development of fruiting bodies in Aspergillus as seen through differential interference contrast microscopy (630X) 2. Biseriate fruiting body in a lung cavity as seen in disease Aspergilloma GMS, 630X. 3. Cryptococcus neoformans: the round yeast cells are surrounded by a polysachharide capsule through differential interference contrast microscopy (630X) 4. Raised skin lesions resulting from dissemination of Cryptococcus in an immunocompromised patient. 5. Histoplasma: Macroconidia and microconidia as seen in phase contrast microscope on a slide culture at 25°C. 6. Histoplasmosis. Discoloration of the skin caused by the fungus.

Introduction
(Thornewell et al. 1997). Evidences also show the presence of a second MDR-
like gene (CneMDR2) in this fungus.
1.6.3. DRUG TRANSPORTERS FROM NON-ALBICANS SPECIES
A C. dubliniensis homolog of CaMDR1, termed CdMDR1, was cloned and while
its nucleotide sequence was found to be 92% identical to the corresponding
CaMDR1 sequence, the predicted CdMDR1 protein was found to be 96%
identical to the corresponding CaMDR1 protein. By PCR analysis, C. dubliniensis
was also shown to encode homologs of CDR1 and CDR2, termed CdCDR1 and
CdCDR2, respectively. Northern analysis of fluconazole-susceptible and -
resistant isolates of C. dubliniensis revealed that fluconazole resistance was
associated with increased expression of CdMDR1 mRNA.
In C. glabrata genes conferring resistance to azole antifungals were cloned in a
S. cerevisiae strain in which the ATP binding cassette (ABC) transporter gene
PDR5 was deleted . Three different genes were thus recovered , and among them,
only C. glabrata CDR1 (CgCDR1), a gene similar to the C. albicans ABC
transporter CDR genes, was upregulated by a factor of 5 to 8 in the azole-
resistant isolates (Sanglard et al. 1999). A correlation between upregulation of
this gene and azole resistance was thus established. The deletion of CgCDR1 in
an azole-resistant C. glabrata clinical isolate rendered the resulting mutant
(DSY1 041) susceptible to azole derivatives as the azole-susceptible clinical
parent.
Recently, two genes, ABC1 and ABC2, were identified in Candida krusei, which
code for the ABC type drug efflux pumps (Katiyar and Edlind 1998). ABC1 RNA
was constitutively expressed at low levels in log phase cells while ABC2 RNA
was undetectable. However, both genes were upregulated as cultures
approached stationary phase, and this upregulation was correlated with
decreased susceptibility to the lethal activity of the azole derivative miconazole.
Furthermore, ABC1 was upregulated following brief treatment of C. kruse; with
miconazole and clotrimazole (but not other azoles), and the unrelated
compounds albendazole and cycloheximide.
26

Introduction
1.6.4. EFFLUX PUMPS IN C. ALBICANS
Genes encoding several distinct ABC transporters and one major facilitator has
been cloned from C. albicans. Both CDR (COR1 and COR2) and MOR1 genes
are overexpressed in azole resistant isolates (Krishnamurthy et al. 1998; White
1997b). The pumps, which have been shown to be involved in antifungal
resistance, have 12 membrane-spanning helices in their protein structure. The
presence of the numerous post-translational modification sites suggests that the
activity of the pumps can be extensively modulated. In C. albicans more than ten
different ABC-type transporters have been predicted. CaMOR1 (also known as
Benr) and FLU1 are genes identified in C. albicans encoding for MFS-type pumps
(Calabrese et al. 2000; Gupta et al. 1998). Both types of pumps are widely
distributed in nature. While CDR proteins serve as efflux pumps for most of the
azole drugs, the MOR1 protein appears to prefer fluconazole, although it can
facilitate transport of unrelated nonazole drugs (Cannon et al. 1996). Because of
the clinical significance of ABC proteins, it is critical to determine their role in
normal cellular physiology. The physiological role of the CDRs is as mentioned
below.
Table 12. Known physiological functions of some ABC transporters of C.
albicans
ABC proteins Functions Exam~les Cdr1p • Drug transporter • Azoles, rhodamine123
• Steroid transporter • ,B-estradiol • Phospholipid • PtdEth & PtdCh
translocator(Floppase) In to out Cdr2p • Drug transporter • Azoles, rhodamine123
• Phospholipid • PtdEth & PtdCh translocater:(Flo~asel In to out
Cdr3p • Phospholipid • PtdEth & PtdCh Translocater(Flippase) Out to in
27

Introduction ------------------------------------------------------
1.6.4.1, CDR1
CDR1 gene of C. albicans was cloned by functional complementation of PDR5
gene of S. cerevisiae with Candida genomic library
::::::::;;;U1mt;tm~:: (Prasad et aJ. 1995a). After preliminary drug
resistance, a 6.2 kb clone was identified and
sequenced. This gene named CDR1 (Candida Drug Resistance) has an ORF of
4503 bp and 1210 bp of 5' promoter region. This flanking 5' promoter region
shows TATA region consisting of two overlapping TATA consensi (TATATA and
TATAAA). In the 3' 489 bp region sequenced, five sequences corresponding to
the Zaret and Sherman consensus for transcription and polyadenylation are
detected between 135 and 210 bp downstream of the stop codon. CDR1 maps to
chromosome 3 SfIl fragment P of C. albicans.
CDR1 ORF encodes a polypeptide of 1501 aminoacids with a molecular weight
of 169.9 kDa and an estimated isoelectric point of 6.53. The predicted secondary
structure and domain organization displays its belonging to ABC superfamily. It is
composed of two homologous halves, each comprising of one N-terminal
hydrophilic domain followed by a C-terminal hydrophobic domain. There are six
transmembrane spans for each of thG hydrophobic domain predicted. The
spacing between each of the six clustered TM spans are well conserved in the N-
terminal than the C-terminal half. Each hydrophilic domain contains ATP-binding
motifs typical ·of ATP binding cassette domains. Particularly Cdr1 pshares
sequence conservation with PDR5 and SNQ2 sequences. In the N-terminal ABC
domain, the Walker A motif (187 GPGAGCST) displays cysteine instead of the
frequently conserved lysine, and well conserved ABC signature (303
VSGGERKRVSIA), while in the Walker B motif (323 IQCD) the conserved
aspartate is present but not preceded by the typical four hydrophobic aminoacids
(Prasad et al. 1995a). However a strong conservation of sequence is observed in
the region imm~diately following the Walker B aspartate, where at/east the first
seven amino acids are well conserved in the NH2- and COOH-halves of CDR1p,
Pdr5p and Snq2p. The LD residues placed 5 aminoacids downstream from the
walker B aspartate are conserved in CDR1p as well as in other members of ABC
28

Introduction
:uperfamily. Similarly in the C-terminal domain, the walker A (895 GASGAGKT)
and Walker B (1022 LLFLD) motifs are well conserved. However, in place of the
ABC signature, a conserved sequence (1000 LNVEQRKRL TIGVEL) is observed
for COR1, POR5 and SNQ2. COR1 shows high sequence similarities to S.
cerevisiae POR5 (56% identity, 73% similarity) and SNQ2 (42% identity, 60%
similarity) over the entire length of the protein. Other proteins showing homology
to COR1, albeit at the level, is the white and brown pigment transporters of
Drosophila, AOP1 from S. cerevisiae, mammalian multidrug resistance protein
(MOR1) and human CFTR. The structural arrangement is identical to that of S.
cerevisiae ABC proteins, POR5 and SNQ2 as mentioned above. It mirrors the
architecture of the yeast a-mating pheromone transporter STE6, as well as the
mammalian drug resistance P-glycoprotein (MOR1) and cystic fibrosis factor
(CFTR). The significance of such domain inversion in some ABC proteins is not
clear. Many physiological functions like steroid transport and phospholipid
translocase are being discovered for COR1.
1.6.4.2. CDR2
The Cdr2p protein shares 84% identity and 92% similarity to CDR1p although the
N- and C-terminal portions of the proteins are more divergent. The ORF codes for
a protein 168 kDa in size with a conserved domain structure typical of ABC
proteins (Sanglard et al. 1997). CDR2 seems to have lower potency than CDR1
in the degree of resistance conferred in S. cerevisiae but gives specific resistance
to crystal violet. CDR2 is known to be overexpressedin resistant isolates. The
disruption of COR2 did not result in hypersusceptibility to the drugs tested. The
available information of the regulatory sequences in CDR2 promoter shows no
similarity with the CDR1 promoter indicating divergent regulation of the two
genes.
29

Introduction
1.7. REGULATOR'! NETWORKS INVOLVED IN MDR
As discussed above, the overexpression of drug efflux pumps is one of the major
contributors to the development of multidrug resistance across the evolutionary
scale (Higgins 1992). However, the molecular mechanisms underlying this
phenomenon have not been very well dissected. The elucidation of the regulatory
pathways controlling the expression of efflux pumps would not only give an
insight into the mechanism of regulation but may also lead to the discovery of
novel targets for antifungals.
1.7.1. REGULATION OF MDR IN BUDDING YEAST
Several regulatory networks deciphered in yeast include the PDR network which
forms a mesh of pathways regulating the expression of a number of genes,
related as well as unrelated to the phenomenon of multidrug resistance (Balzi
and Goffeau 1991). Other regulatory pathways in S. cerevisiae include the YAP
network and the recently studied YRR network. The elementary fact about all
these networks is the existence of cross talk within the regulatory networks.
1.7.1.1. The Master Regulators
1.7. 1. 1. 1. Pdr1 p
Various genetic interactions connecting PDR regulators to drug pumps have
been uncovered. As an example, the regulators POR1, POR3, POR7, and POR9
have been shown to control the transcription of the multidrug pump gene POR5,
encoding an ABC type protein (Meyers et al. 1992).
The pleiotropic drug resistance locus POR1 was first defined by a series of
nuclear mutations, initially isolated by selection in the presence of one or two
drugs and shown to display cross resistance to a total of nearby 30 different
inhibitors affecting unrelated, cytoplasmic or mitochondrial functions (Balzi et al.
1987). No fewer than twenty independent mutations conferring multidrug
resistance have been attributed to the POR1 locus. The very high frequency of
isolation of alleles of POR1 in the course of independent searches for mutations
suppressing toxicity suggests that POR1 plays a primary role in the multidrug
30

Introduction
resistance phenotypes of yeast (Hallstrom and Moye-Rowley 1998). The
phenotype associated to POR1 mutations is pleiotropic and not merely restricted
to multidrug resistance. The POR1-2 mutant allele was, for example, related to
physiological alterations such as respiratory deficiency and inability to grow under
adverse conditions, such as elevated pH, temperature, and osmolality (Carvajal
et al. 1997).
The POR1-8 multidrug resistance allele was shown to modulate the intracellular
availability in yeast cells of human hormone molecules, such as p-estradiol.
These facts suggest that POR1 affects a wide range of functions, encompassing
resistance to chemical and physical stresses, membrane transport, and organelle
functions. POR1 encodes a Zn2C6 binuclear cluster motif as DNA binding domain.
The first target gene to undergo transcriptional regulation by POR1 was the ABC
drug extrusion transporter POR5/STS1 or YOR1. DNasel protection assay
revealed the presence of three POR1 binding sites in the POR5 promoter
(Katzmann et al. 1996). This consensus (TCCGCGA) includes two rotationally
symmetric CCG triplets as typical traits for the binding of Zn2C6 cluster proteins.
By the use of a POR1::lexA fusion system, a transcription activation domain has
been identified in the carboxy terminus of the POR1 protein.
STE6, a homologue of POR5 is also transcriptionally influenced by POR1 (Mahe
et al. 1995). No typical POR1-binding consensus is present in STE6 implying
either that other POR1-binding sites exist or that the effect of the POR1 on the
STE6 transcript is indirect. Numerous other targets of POR1 are now known like
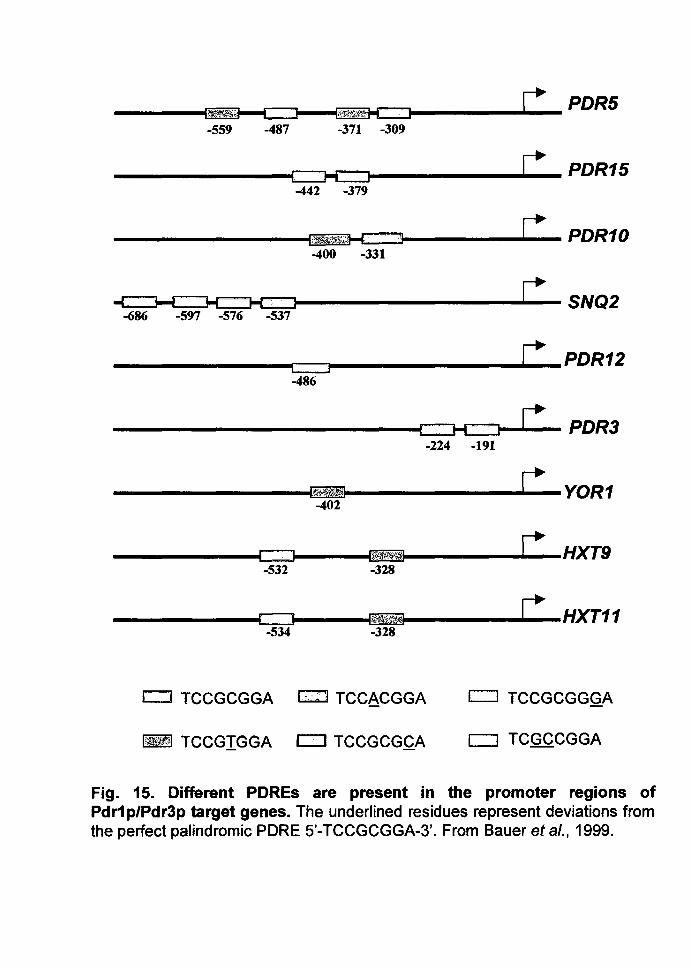
POR10, POR11, GAS1, G3PO, etc. (Fig. 15).
Recently Rowley and his group have shown that the Pdr1 p function is positively
regulated by Hsp70 protein encoded by POR13 (Hallstrom et al. 1998; Hallstrom
. and Moye-Rowley 2000a). Expression of deletion, insertion, and amino acid
. SUbstitution mutant variants of Pd r1 p suggest that the center region of the
transcription factor is the target for Pdr13p mediated response.
31
-559 -487 -371 -309
______ ~~~----......L..~- PDR15 -442 -379
--------------~I~~5~:~~~KE.:==~--------~~-PDR10 -400 -331
~~~~~~r=~-------------------SNQ2 -686 -597 -576 -537
___________ ~~------------.~--PDR12 c::::::J -486
----------------------------~~ PDR3 -224 -191
------------------~~~--------------~~---YOR1 -402
------------~r=JL_~. ~----~~~--------~~---H)(T9 -532 -328
c:J TCCGCGGA c::::J TCCACGGA c::J TCCGCGGGA
I~I TCCGIGGA c:::J TCCGCGCA [::J TCGCCGGA
Fig. 15. Different POREs are present in the promoter regions of Pdr1 p/Pdr3p target genes. The underlined residues represent deviations from the perfect palindromic PORE 5'-TCCGCGGA-3'. From Bauer et al., 1999.

Introduction
1.7.1.1.2. Pdr3p
A second site of mutations conferring multidrug resistance similar to POR1 is the
POR3 locus, localized in proximity of chromosome II centromere (Delaveau et al.
1994). POR3 encodes a transcription regulator of the Zn2C6 cluster protein family,
which is 36% homologous to POR1 and has a very highly conserved DNA
binding domain. Two transcription activation domains were identified respectively
near the N-terminal DNA binding domain and at the carboxy-terminus of POR3.
Wild type alleles of POR1 and POR3 cross complement POR3 or POR1
mutations respectively indicating functional overlapping between the two genes.
POR1 and POR3 genes regulate the expression of at least one common target,
the POR5 gene (Katzmann et al. 1994). Thus the findings that POR1 and POR3
potentially share common binding sites in the POR5 promoter validates the
observations of overlapping and complementary functions for these two
regulators. There exists an autoregulatory process for POR3 and a hierarchical
regulation by POR1 over the expression of POR3 (Delahodde et al. 1995). Two
POR113 binding sites are present in the POR3 promoter (Katzmann et a/. 1996).
Both POR1 and POR3 were shown to activate the promoter of POR3, fused to lac
Z. A reduced induction of this fusion gene is seen in a POR1 deleted background
suggesting a cooperation between POR1 and POR3 in the activation of the POR3
promoter. The autoactivation of the POR3 was shown to be involved in the
process of conferring resistance to cycloheximide. These provide the first
mechanistic model to explain how the homologous regulators POR1 and POR3,
which exhibit complementary but still somehow distinct functions, could interact
and lead to the fine regulation of expression of the multidrug resistance
determinant POR5.
The expression of FLR1, an MFS type of pump, is under the control of the
transcription factor Pdr3p (Broco et al. 1999). POR3 deletion severely reduces
benomyl-induced activation of the FLR1 gene expression (by 85%), while its
homologue Pdr1p is apparently not involved in this activation.
32

Introduction
1.7. 1. 1.3. The involvement of PDRs in regulation of non-ABC genes
Besides regulating the function of a number of ABC pumps, PDR1 and PDR3
also regulate the expression of genes involved in the normal functioning of the
cell (Fig. 16). HXT9 and HXT11 belong to the family of hexose transporters
whose basal expression is under the influence of PDR11PDR3 and the
overexpression of these genes leads to drug hypersensitivity (Nourani et al.
1997). This appears to be a contradiction to the other PDR11PDR3 targets whose
overexpression leads to drug resistance. But it is quite plausible, since hexose
transporters are supposed to mediate uptake rather than efflux of specific
molecules, increased expression of these permeases at the cell surface could
cause an increased
uptake (of toxic
substances).
Another recent finding
adding to the
multifaceted role of the
PDRs is that the loss of
the mitochondrial genome (rhoOcells)
causes up-regulation of
Pdr3p but not Pdr1p
function (Hallstrom and
Moye-Rowley 2000b).
Additionally, loss of the
mitochondrial inner
membrane protein
YCF1
Fig. 16. The PDR network. The genes in the center of the model represent ABC transporters and/or Pdr1 p/Pdr3p target genes, which are currently known to be regulated by the transcriptional regulators above (Pdr1 p/Pdr3p) and below (yap1p, Yrr1p, Yap2p, Msn2p/Msn4p). From Bauer, 1QQQ
Oxa1p generates a signal that results in increased Pdr3p activity. Both of these
mitochondrial defects, lead to increased expression of the PDR3 structural gene.
Importantly, the signaling pathway used to enhance Pdr3p function in rhoo cells is
not the same as in oxa1 cells. Loss of nuclear-mitochondrial signaling genes like
33

Introduction
RTG1 reduce the level o~ POR5 expression and drug resistance seen in rhoo cells
but has no effect on oxa 1-induced phenotypes. These data uncover a new
regulatory pathway connecting expression of multidrug resistance genes with
mitochondrial function.
Recent evidence also shows that both Pdr1 p and Pdr3p act to regulate
production of an enzyme involved in sphingolipid biosynthesis in S. cerevisiae
(Hallstrom et al. 2001). The last step in formation of the major sphingolipid in the
yeast plasma membrane, mannosyl-diinositol-phosphorylceramide (M(IP)2C) is
catalyzed by the product of the IPT1 gene, inositol phosphotransferase (lpt1 p).
Transcription of the IPT1 gene is responsive to changes in activity of Pdr1 p and
Pdr3p. A single Pdr1 pi Pdr3p response element (PORE) is present in the IPT1
promoter and is required for regulation by these factors. Loss of IPT1 has
complex effects on drug resistance of the resulting strain, consistent with an
important role for M(IP)2C in normal plasma membrane function. Direct assay for
lipid contents of cells demonstrated that changes in sphingolipid composition
correlate with changes in the activity of Pdr3p. These data suggest that Pdr1 p
and Pdr3p may act to modulate the lipid composition of membranes in S. cerevisiae through activation of sphingolipid biosynthesis along with other target
genes.
1.7.1.2. The Yrr Network
It has been previously shown that the YOR1 gene is regulated by the master
regulators Pdr1 p and Pdr3p by directly binding to the PORE located in the YOR1
promoter (Hallstrom and Moye-Rowley 1998). Recently Moye Rowley has shown
that a hyperactive form of Yrr1 p causes enhanced expression of a YOR1-lacZ
construct that is independent of the presence of POR1 and POR3 (Zhang et al.
2001). Mutant PDRE-YOR1 promoter construct was not compromised on the
increase in p-galactosidase activi.ty in the presence of the YRR1-1 allele. Deletion
mapping of the YOR1 promoter-lacZ constructs in the presence of co-transfected
YRR1-1 low copy number plasmid led to the identification of a region in between
34
Introduction
-222 and -190 bp which mediates the Yrr1p effect. Thus this is another example
of multiple levels of control of a particular gene by several transcription factors.
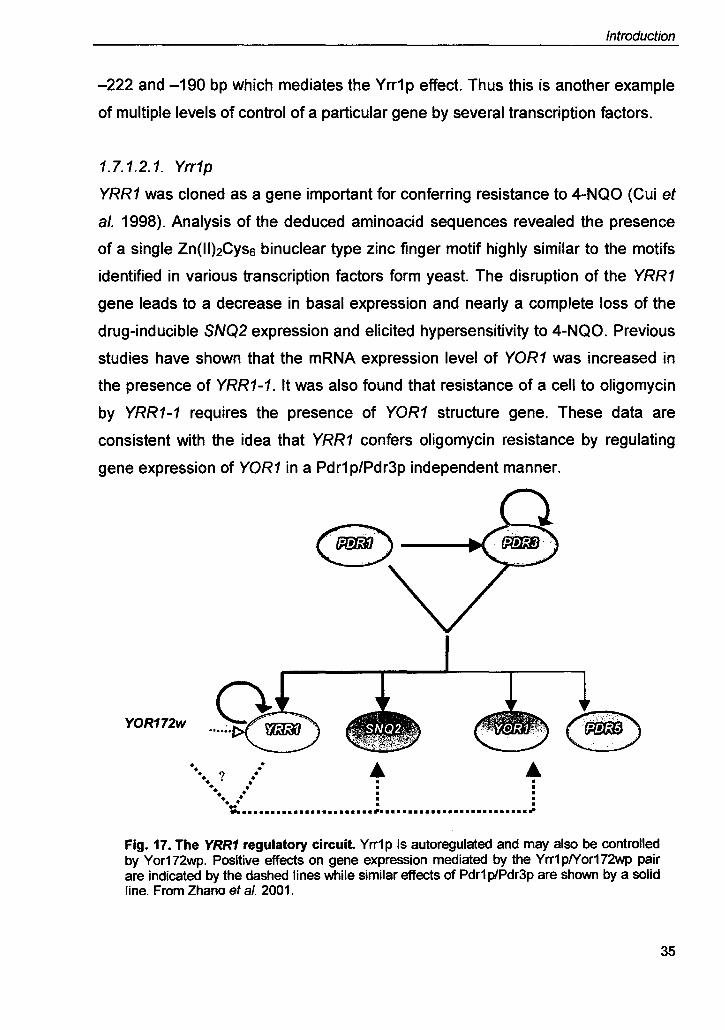
1.7.1.2.1. Yrr1p
YRR1 was cloned as a gene important for conferring resistance to 4-NQO (Cui et
al. 1998). Analysis of the deduced aminoacid sequences revealed the presence
of a single Zn(II)2Cyss binuclear type zinc finger motif highly similar to the motifs
identified in various transcription factors form yeast. The disruption of the YRR1
gene leads to a decrease in basal expression and nearly a complete loss of the
drug-inducible SNQ2 expression and elicited hypersensitivity to 4-NQO. Previous
studies have shown that the mRNA expression level of YOR1 was increased in
the presence of YRR1-1. It was also found that resistance of a cell to oligomycin
by YRR1-1 requires the presence of YOR1 structure gene. These data are
consistent with the idea that YRR1 confers oligomycin resistance by regulating
gene expression of YOR1 in a Pdr1 p/Pdr3p independent manner.
YOR172w
• •• ..? . ... .. +.. .• • • • . ~ .....•...••••....•••..• ; .......................... ;
Fig. 17. The YRR1 regulatory circuit Yrr1p is autoregulated and may also be controlled by Yor172wp. Positive effects on gene expression mediated by the Yrr1plYor172wp pair are indicated by the dashed lines while similar effects of Pdr1 p/Pdr3p are shown by a solid line. From Zhana et al. 2001.
35

Introduction
Along with identification of a Yrr1p response element in tile YOR1 promoter,
further analysis revealed the presence of a consensus PORE. ONasel protection
assay with the purifi6d Pdr1 p indicated that Pdr1 p regulates the YRR 1 gene. The
presence of the YRR1-1 allele increased expression of the YRR1-lacZ fusion by
nearly 600%. It appears that the PORE and Yrr1 p response elements are
physically linked in the YRR1 promoter (Zhang et al. 2001). Two base pairs
critical for the Yrr1 p activation, are located immediately adjacent to the YRR1
PORE. A similar tight linkage has been found for SNQ2 (Fig. 17). At least for
these three Yrr1 pI Pdr1 p co-regulated genes the possibility exists that these
regulatory proteins may directly communicate during gene regulation and thus
normal drug resistance.
Table 13. Known transcription factors in yeasts and their targets
Protein Organism Structure Target Gene Function Pdr1p S. cerevisiae TF (Zn(II)2Cys6) POR5 Regulation of PDR
POR10 POR15 SNQ2 YOR1 POR3
Pdr3p S. cerevisiae TF (Zn(II)~s6) POR5 Regulation of PDR POR10 POR15 SNQ2 YOR1 POR3
Pdr13p S. cerevisiae Hsp70 homologue Pdr1p Posttranslational Pdr1 p ~ulation
Ngg1p S. cerevisiae TF Pdr1p Inhibition of Pdr1 p activity
Pdr3p Yrr1p S. cerevisiae TF JZn(II)2~s61 SNQ2 Regulation of PDR
YOR1
Yap1p S. cerevisiae TF (bZip) POR5 Oxidative stress response
SNQ2 Cd2+ and diazoborine resistance
YCF1 Yap2p S. cerevisiae TF(bZip) YCF1 Cd2+ resistance
Regulation of arsenite Yap8p S. cerevisiae TF (bZip) YCF1? and arsenate
resistance Cmk1p S. cerevisiae CaM kinase Pdr12p Inhibition of Pdr12~
36

Yck1p S. cerevisiae Casein kinase I
Pap1p S.pombe TF (bZip)
Cap1p C. albicans TF (bZip)
CaTEC1p C. albicans TENATTS
RFG1p C. albicans HMG protein
Fcr1p C. albicans TF (Zn(II)2Cys6)
1.7.1.3. The Yap Network
1.7.1.3.1 Yap1p
Introduction
activity Modulation of keto-
Pdr5p and miconazole resistance
Snq2p Yor1p
Hba2p Oxidative stress response
Pmd1p
CaYCF1 Oxidative stress response
SAP4 Regulates virulence SAPS SAP6 Hyphal specific Represses genes filamentation
? Deletion confers azole resistance
The S. cerevisiae transcription factor YAP1 plays an important role in oxidative
stress response (OSR) and multidrug resistance by activating target genes
involved in cellular detoxification (WU et at. 1993). YAP1 belongs to the bZip
(Basic domain! leucine zipper) family of transcription factors which includes the
yeast Gcn4p and the mammalian AP-1 proteins, Fos and Jun (Moye-Rowley et
at. 1989). It activates transcription by binding to specific DNA sequences located
in the promoter of its targets. Yap1p targets involved in OSR include TRX2,
(thioredoxin), GSH1, (gamma glutamyl cysteine synthetase) GSH2, (glutathione
synthetase), TRR1 (thioredoxin reductase), GLR1, (glutathione reductase, GPX2
(glutathione peroxidase), TSA 1 (thioredoxin peroxidase) and A,4P1 (a!ky!
hydroperoxide reductase) as summarized in the table below. Yap1 p also
regulates the transcription of genes encoding membrane associated transporters
such as YCF1, coding for an ABC transporter which functions as a glutathione S
conjugate pump as well as ATR1 and FLR1, coding for MDR transporters of the
Major Facilitator (MF) superfamily (Nguyen et at. 2000). Large-scale studies
investigating Yap1 p-dependent transcription have identified several additional
37
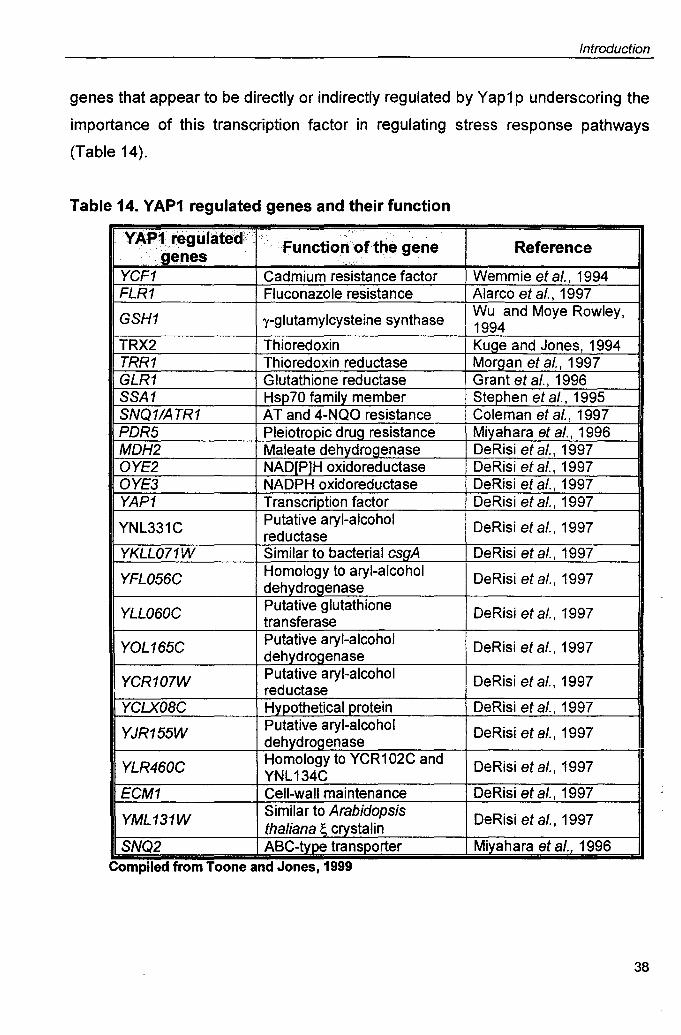
Introduction
genes that appear to be directly or indirectly regulated by Yap1 p underscoring the
importance of this transcription factor in regulating stress response pathways
(Table 14).
Table 14. YAP1 regulated genes and their function
YAP~lregulated>·.· , 'Functiorfof the gene Reference .g~nes .:.
YCF1 Cadmium resistance factor Wemmie et a/., 1994 FLR1 Fluconazole resistance Alarco et a/., 1997
GSH1 y-glutamylcysteine synthase Wu and Moye Rowley, 1994
TRX2 Thioredoxin Kuge and Jones, 1994 TRR1 Thioredoxin reductase Morgan et a/., 1997 GLR1 Glutathione reductase Grant et a/., 1996 SSA1 Hsp 70 family member Stephen et a/., 1995 SNQ11ATR1 AT and 4-NQO resistance Coleman et a/., 1997 PDR5 Pleiotropic drug resistance Mil'.-ahara et aI., 1996 MDH2 Maleate dehydrogenase DeRisi et a/., 1997 OYE2 NAD[P]H oxidoreductase DeRisi et a/., 1997 OYE3 NADPH oxidoreductase DeRisi et a/., 1997 YAP1 Transcription factor DeRisi et a/., 1997
YNL331C Putative aryl-alcohol DeRisi et a/., 1997 reductase YKLL071W Similar to bacterial csgA DeRisi et a/., 1997
YFL056C Homology to aryl-alcohol DeRisi et a/., 1997 dehydrogenase
YLL060C Putative glutathione DeRisi et a/., 1997 transferase
YOL165C Putative aryl-alcohol DeRisi et a/., 1997 dehyd rogenase
YCR107W Putative aryl-alcohol DeRisi et a/., 1997 reductase YCLX08C Hypothetical protein DeRisi et a/., 1997
YJR155W Putative aryl-alcohol DeRisi et a/., 1997 dehydrogenase
YLR460C Homology to VCR 1 02C and DeRisi et a/., 1997 YNL 134C ECM1 Cell-wall maintenance DeRisi et a/., 1997
YML131W Similar to Arabidopsis DeRisi et a/., 1997 thaliana ~ crystalin
SNQ2 ABC-type transporter Miyahara et a/., 1996 . Complied from Toone and Jones, 1999
38

Introduction
YAP1 was originally identified on the basis of its ability to bind to an AP-1
recognition element (ARE) found in the SV40 enhancer (5'-TGACTAA-3').
However, it has recently been demonstrated that Yap1 p preferentially interacts
with the sequence S'-TTAC/GTAA. This sequence, which is palindromic and
contains two identical TTA half-sites, has been shown to function in an orientation
independent manner. It thus appears that the consensus Yap1 p response
element (YRE) corresponds to the sequence 5'-TT/GAC/GTAA. Additionally
using the Yap1, Yap4 or YapS basic region, 8 members of the Yap family have
been unraveled so far (Fernandes et a/. 1997). Fernandes et a/. has shown that
the Yap proteins have common properties but are functionally distinct. The
magnitude of activation through the Yap site varies considerably, Yap1 activates
strongly, Yap2 activates moderately, Yap3 activates weakly, and YapS
dependent activation is barely detectable. PKA, the ultimate target of the ras
pathway in yeast inhibits Yap1-dependent transcription both by reducing Yap1
protein levels and by inhibiting Yap1 function.
The activity of Yap1 p is predominantly regulated at the level of nuclear export.
Under unstressed conditions, Yap1 p shuttles between the cytosol and the
nucleoplasm but is mainly cytosolic. It is actively exported from the nucleus by
the exportin Crm1 p, which interacts with the C-terminal cysteine rich domain
(CRD) of Yap1p (Kuge et a/. 1997). This domain contains a leucine rich nuclear
export sequence (NES) embedded within three cysteine residues invariably
conserved among homologues of the Yap1p family. Removal of the CRD,
mutation of the NES or of the cysteines as well as treatment of the cells with
oxidative agents such as hydrogen peroxide, diamide or diethylmaleate all disrupt
the Crm1 pINES interaction, resulting in the accumulation of Yap1 p in the nucleus
and Yap1 p dependent transcriptional activation. The CRD thought to behave as a
specialized export signal, which is sensitive to the redox state of cells, the
oxidation status of the cysteines affecting the accessibility of the NES to Crm1 p
thereby regulating Yap1 p activity (Fig. 18).
39

Non-stressed conditions Oxidative Stress
Nucleus Cytoplasm Nucleus Cytoplasm
' Cnnf >I @~ ttl YAP-t
@ -~~~ •
E~
Fig. 18. Yeast AP-1 proteins YAP-1, are regulated via oxidative stress-dependent changes in subcellular localization. Under normal growth conditions the YAP1 protein is cytoplasmic and maintained in the cytoplasm through the action of the Crm 1 nuclear export factor. Under oxidative stress conditions, th is export pathway is blocked and the YAP-1 protein accumulates in the nucleus resulting in increased YAP-1-dependent gene expression . [From Toone and Jones, 1999]

Introduction
1.7.2. CROSS-TALK BETWEEN TRANSCRIPTIONAL REGULATORS OF
MULTIDRUG RESISTANCE GENE
A link between the PDR and Yap transcription factors has also been established.
YAP1 overexpression gives rise to diazoborine resistance only in a wild type but
not in a L1pdr1 L1pdr3 background indicates that Yap1 p acts either in concert with
these two transcription factors on a common gene or participates in the
sequential Cadmium
activation of an
unknown target
gene (Wendler et
al. 1997). In
addition to
diazoborine
resistance, PDR3
is required for
Yap1 p-mediated
resistance to 4-
NQO but not for
the inhibitors
cycloheximide
Cycloheximde .... ~I--_
1, 10-O-Phen~
4-NQO
Fig. 19. A schematic showing the cross talk between the YAP and PDR networks in S. cerevisiae. [From Wendler et aI., 1997]
and 1, 10 o-phenanthroline. This shows that resistance to some drugs caused by
YAP1 overexpression is mediated via the PDR3 gene, while Yap1 p might directly
activate other detoxifying genes. Taken together, there appears to be a functional
connection of the Yap1 p activity with the master regulators of the pleiotropic drug
resistance network (Fig. 19).
40
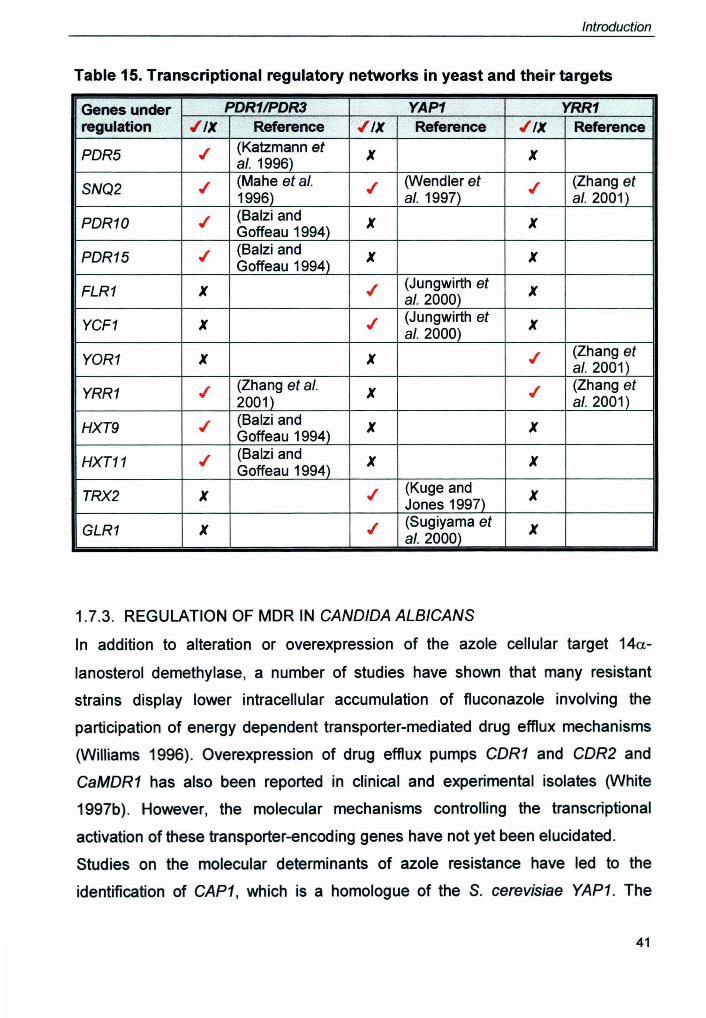
Introduction
Table 15. Transcriptional regulatory networks in yeast and their targets
Genes under PDR1IPDR3 YAP1 YRR1 regulation ./IX Reference ./IX Reference ./IX Reference
PDR5 ./ (Katzmann et X X al. 1996)
SNQ2 ./ (Mahe et al. ./ (Wendler et ./ (Zhang et 1996) al. 1997) a/. 2001)
PDR10 ./ (8alzi and X X Goffeau 1994)
PDR15 ./ (8alzi and X X Goffeau 1994)
FLR1 X ./ (Jungwirth et X al.2000)
YCF1 X ./ (Jungwirth et X a/.2000)
YOR1 X X ./ (Zhang et al.2001)
YRR1 ./ (Zhang et a/. X ./ (Zhang et 2001) al.2001)
HXT9 ./ (8alzi and X X Goffeau 1994)
HXT11 ./ (8alzi and X X Goffeau 1994)
TRX2 X .t (Kuge and X Jones 1997)
GLR1 X .t (Sugiyama et X al.2000)
1.7.3. REGULATION OF MDR IN CANDIDA ALB/CANS
In addition to alteration or overexpression of the azole cellular target 14a-
lanosterol demethylase, a number of studies have shown that many resistant
strains display lower intracellular accumulation of fluconazole involving the
participation of energy dependent transporter-mediated drug efflux mechanisms
(Williams 1996). Overexpression of drug efflux pumps CDR1 and CDR2 and
CaMDR1 has also been reported in clinical and experimental isolates (White
1997b). However, the molecular mechanisms controlling the transcriptional
activation of these transporter-encoding genes have not yet been elucidated.
Studies on the molecular determinants of azole resistance have led to the
identification of CAP1, which is a homologue of the S. cerevisiae YAP1. The
41

Introduction
isolation and characterization of a number of C. albicans genes involved in PDR
have revealed that their protein products possess structural and functional
homologues in S. cerevisiae suggesting some similarity between the S.
cerevisiae and C. albicans PDR ,systems. By analogy to the master regulators
Pdr1 and Pdr3, the FCRs were cloned by Raymond and group. The mode of
regulation of MDR pumps in non-albicans Candida species is still obscure. The
following section deals with the regulators of Candida MDR, the genes they
regulate and the new TFs recently discovered to playa role in virulence.
1.7.3.1. Fcrp
Three C. albicans genes, designated FCR (for fluconazole resistance), have
been isolated by their ability to complement the fluconazole hypersensitivity of a
S. cerevisiae mutant lacking the transcription factors Pdr1 p and Pdr3p (Talibi and
Raymond 1999). Complementation of the FCZ hypersusceptibility of the POR1
POR3 mutant strain by the three genes was only partial. Northern blot analysis
showed that each of these three genes is able to increase the expression of the
POR5 gene in the mutant strain and therefore is likely to encode a transcriptional
regulatory factor. Overexpression of these genes in the wild type POR1 POR3
parental strain did not result in the increased resistance to fluconazole, consistent
with the idea that they probably encode functional homologues of the Pdr1 p and
Pdr3p proteins. For FCR1, this hypothesis was supported by nucleotide
sequencing of the gene which was found to code for a regulatory factor,
homologous to S. cerevisiae Pdr1p, belonging to the yeast zinc cluster family.
Sequence comparison analyses indicate that the zinc cluster domains of Fcr1p
and Pdr1p are 53% identical and that this homology extends further downstream
into the linker and dimerization domain. Also, the C terminal domain of Fcr1 p
displays 29% identity with the C terminal domain of Pdr1 p, which has been
,shown to interact with the coactivator/repressor ADA complex and is believed to
function as a transcriptional activating domain. Moreover, two leucine residues,
which are present in gain-of-function mutants of Pdr1p and which are associated
with increased levels of POR5 mRNA in these mutants, are conserved in Fcr1p.
42

Introduction
A fcr11fcr1 double mutant was more resistant to a number of structurally
unrelated drugs, including FCZ, ketoconazole and brefeldin A as well as
fluphenazine and itraconazole than the wild type strain. Moreover it was found
that this double knockout strain was not more resistant to other drugs like 4-NQO
and 1, 10-phenanthroline, demonstrating that the drug resistant phenotype
resulting for the fcr11fcr1 deletion is not generalized, but is indeed specific for
certain drugs. Thus this shows that Fcr1 p behaves as a negative determinant of
PDR in C. albicans although it behaves as a positive regulator of PDR5 and drug
resistance in S. cerevisiae. This could be because of a heterologous background.
It is thus possible that Fcr1 p may negatively regulate the expression of CDR1
and CDR2 or other genes conferring similar phenotypes in C. albicans.
Alternatively it is possible that Fcr1 p positively regulates a gene whose
expression confers drug sensitivity.
1.7.3.2. Cap1p
Investigation of C. albicans molecular determinants of fluconazole resistance led
to the isolation of a gene coding for a new member of the yeast AP-1 family, that
is CAP1 (Alarco and Raymond 1999). CAP1 bears a very high degree of
homology to S. cerevisiae Yap2 and Schizosachharomyces pombe pap (Chang
and Kwon-Chung 1999). These proteins are not only structurally but also
functionally related, since their overexpression from a multicopy plasmid results
in a similar phenotype, namely an increased cellular resistance to different
toxicants. The sequence homology between CAP1 and YAP1 proteins translates
into functional homology because, CAP1 overexpression in a YAP1 mutant
partially restores the ability of the cells to grow on otherwise toxic concentrations
of cadmium and hydrogen peroxide and also because both CAP1 and YAP1 can
transactivate FLR1 to confer MDR (Alarco et al. 1997). Amino acid sequence
comparison of the CAP1, YAP1, yap2 and pap1 proteins indicates that the most
conserved regions among these proteins overlap the N-terminal blip and the C-
terminal acidic domains.
43

Introduction
To unravel the biological function of CAP1 gene it was deleted and its
consequence was studied. Deletion of CAP1 had no effect on the basal
transcription of the genes analyzed. However deletion led to increased
hypersensitivity to 4-NQO, 1, 10-phenanthroline, cadmium and hydrogen
peroxide which could be reverted by reintroduction of a plasmid borne copy of the
CAP1 gene in the CAP1 deletion strain. The essential role of CAP1 in cellular
tolerance to the reactive oxygen species (ROS) H202, is of particular interest
since it suggests that CAP1 may be involved in C. albicans response to the ROS
produced by the host immune system.
The production of ROS such as H202, hydroxyl radicals, and superoxide anions
by human phagocytic neutrophils is a major line of defense against fungal
infections.
1.7.3.3. CaTec1p
CaTEC1, a new member of the TEAlATTS family of transcription factors
regulates C. albicans virulence. The promoters of the type 4, 5 and 6 proteinase
isogenes (SAP4-6) contain repetitive TEAlATTS consensus sequence motifs.
CaTEC1 is predominantly expressed in the hyphal form of C. albicans. In vitro,
serum-induced hyphal formation as well as evasion from MPhi after phagocytosis
is suppressed in catec1/catec1 mutant cells. Furthermore, expression of the
proteinase isogenes SAP4-6 is no longer inducible in these mutant cells. The
deletion of the CaTEC1 gene attenuates virulence of C. albicans in a systemic
model of murine Candidiasis, although both mutant and revertant cells that were
prepared from infected tissues or the vaginal mucosa grew in a hyphal
morphology in vivo. Ca TEC1 complements the pseudohyphal and invasive
growth defect of haploid and diploid S.cerevisiae tec1/tec1 mutant cells and
strongly activates the promoter of FLO 11, a gene required for pseudohyphal
growth. Thus, this is the first evidence pointing to an essential role for a member
of the TEAlATTS transcription factor family that had so far only been ascribed to
function during development, as a virulence regulator in microbial pathogenesis.
44

Introduction
1.7.3.4. Rfg1 P
RFG1 is a repressor of hyphal growth in the pathogenic yeast C. albicans. The
gene was originally cloned in an attempt to characterize the homologue of the S.
cerevisiae Rox1, a repressor of hypoxic genes (Khalaf and Zitomer 2001). Rox1
is an HMG-domain, DNA binding protein with a repression domain that recruits
the Tup1/Ssn6 general repression complex to achieve repression. Rfg1 is
important for virulence of C. albicans in a mouse model and is shown to control a
number of genes that have been implicated in this process (Kadosh and Johnson
2001). Results demonstrate that a regulatory protein that controls the hypoxic
response in S. cerevisiae controls filamentous growth and virulence in C.
albicans.
45







