
ANKARA ÜNİVERSİTESİ
FEN BİLİMLERİ ENSTİTÜSÜ
DOKTORA TEZİ
Cr, TUZ VE KURAKLIK STRESİ ALTINDA GELİŞTİRİLMİŞ AYÇİÇEĞİ
(Helianthus annuus L.) BİTKİSİNDE HÜCRE DÖNGÜSÜ GENLERİNDEN
RETİNOBLASTOMA (RB), MYELOCYTOMATOSİS (MYC) VE E2F
GENLERİNE AİT mRNA İFADE SEVİYELERİNİN BELİRLENMESİ
Esin BAŞARAN
BİYOLOJİ ANABİLİM DALI
ANKARA
2016
Her hakkı saklıdır

i
ETİK
Ankara Üniversitesi Fen Bilimleri Enstitüsü tez yazım kurallarına uygun olarak
hazırladığım bu tez içindeki bütün bilgilerin doğru ve tam olduğunu, bilgilerin
üretilmesi aşamasında bilimsel etiğe uygun davrandığımı, yararlandığım bütün
kaynakları atıf yaparak belirttiğimi beyan ederim.
11.02.2016
Esin BAŞARAN

ii
ÖZET
Doktora Tezi
Cr, TUZ VE KURAKLIK STRESİ ALTINDA GELİŞTİRİLMİŞ AYÇİÇEĞİ
(Helianthus annuus L.) BİTKİSİNDE HÜCRE DÖNGÜSÜ GENLERİNDEN
RETİNOBLASTOMA (RB), MYELOCYTOMATOSİS (MYC) VE E2F GENLERİNE
AİT mRNA İFADE SEVİYELERİNİN BELİRLENMESİ
Esin BAŞARAN
Ankara Üniversitesi
Fen Bilimleri Enstitüsü
Biyoloji Anabilim Dalı
Danışman: Prof. Dr. E. Sümer ARAS
Ayçiçeği (Helianthus annuus L.) papatyagiller (Asteraceae) familyasına ait olup
çekirdekleri ve yağı için yetiştirilen önemli bir tarım bitkisidir. Bitkisel üretimde stres,
bitkilerin yaşadığı ortamda bir veya birden fazla biyotik ve abiyotik etkenin, büyüme ve
gelişmeyi olumsuz yönde etkileyerek, verim düşüklüğü ile sonuçlanan bir dizi
gerilemeye neden olmaktadır. Bitkilerin yaşamları sürecinde karşılaştıkları abiyotik
stres faktörlerinden en önemlileri tuz ve kuraklıktır. Ağır metal kirliliği de verimi
sınırlayan etmenler arasında yer almaktadır Bitkilerin büyüme süresince etkisi altında
kaldıkları abiyotik stres faktörleri hücre döngüsünde, hücrenin geleceğini belirleyen
kontrol mekanizmalarında birçok genin ekspresyonunda değişikliğe sebep olmaktadır.
Tuz, kuraklık ve Cr(VI) stresi altında yetiştirilmiş 4 farklı ayçiçeği çeşidinde (08-TR-
003, TR-3080, SARAY ve TARSAN 1018) hücre döngüsünün temel mekanizmalarında
görev alan, Retinoblastoma (Rb), Myelocytomatosis (Myc) ve E2F genlerinin mRNA
ifade seviyeleri Real Time PCR ile belirlenmiştir. Ayrıca MDA analizi ve Bradford
protein analizi yapılmıştır.
Tuz ve kuraklık stresi uygulanan tüm çeşitlerde, stres uygulama sürelerinde artışla
beraber genellikle total protein miktarında artış, MDA düzeyinde azalma görülmüştür.
Cr(VI) stresi uygulamasında ise artan Cr konsatrasyonlarında total protein miktarında
azalma, MDA düzeyinde artış gözlenmiştir. Rb, Myc ve E2F genlerinin mRNA ifade
seviyelerinde stres faktörleri ve bitki çeşitlerine göre farklılıklar gözlense de genel
olarak bu stres faktörlerinden etkilendiği söylenebilir. Sonuç olarak, MDA analizi,
Bradford analizi ve çalışılan genlerin ifade seviye farkları bir bütün olarak
değerlendirildiğinde, çalışılan ayçiçeği çeşitlerinin tuz ve kuraklık stresine karşı
toleranslı olduğu söylenebilir. Elde edilen sonuçlar, hücre döngüsü genleri ile yapılacak
ileriki çalışmalara yol gösterici olacaktır.
Şubat 2016, 239 sayfa
Anahtar Kelimeler: Ayçiçeği, Retinoblastoma (Rb), Myelocytomatosis (Myc), E2F,
Real Time PCR, Cr stresi, tuz stresi, kuraklık stresi

iii
ABSTRACT
Ph.D. Thesis
DETERMİNATİON OF MRNA EXPRESSİON LEVELS OF CELL CYCLE GENES
RETİNOBLASTOMA (RB), MYELOCYTOMATOSİS (MYC) AND E2F İN SUNFLOWER
(Helianthus annuus L.) PLANT GROWN UNDER Cr, SALİNİTY AND DROUGHT
STRESSES
Esin BAŞARAN
Ankara University
Graduate School of Natural and Aplied Sciences
Department of Biology
Supervisor: Prof. Dr. E. Sümer ARAS
Sunflower (Helianthus annuus L.) is an important agricultural plant which belongs to
Asteraceae family is being grown for the seeds and seed oil. Biotic and abiotic stress on
plant growth causes a series of regression resulting yield loss in sunflower plants by
affecting growth and development in a negative way. The most important abiotic stress
factors that plants encounter are salinity and drought. Heavy metal pollution is also
another yield limiting factor in sunflower plants. Abiotic stress factors may cause many
changes on the cell cycle, expression of developmental control gene mechanisms of the
cells during plant growth.
mRNA expression levels of Retinoblastoma (Rb), Myelocytomatosis (Myc) and E2F
genes which are responsible from basic mechanisms of cell cycle were studied by Real
Time PCR in four different sunflower plant samples (08-TR-003, TR-3080 SARAY and
TARSAN 1018) to be grown under yield limiting abiotic stress factors such as drought,
salinity and Cr(VI). In addition to that analysis MDA and Bradford protein analysis
were performed. Total protein levels of the stress treated plant samples under increasing
exposure time periods showed increase while MDA levels showed decrease at same
conditions. Increasing stress Cr(VI) application results showed decrease in total protein
amount and increase in MDA concentrations. Although some differences were observed
according to the types of plant and stress factors, mRNA expression levels of Rb, Myc
ve E2F genes were affected by abiotic stress factors.
According to the results of gene expression level differences of studied genes, MDA
and Bradford analysis, it can be concluded that studied samples of sunflower species are
tolerable to the salinity and drought stress conditions. The results of the current study
will shed valuable insight to the future studies on cell cycle genes.
February 2016, 239 pages
Key Words: Sunflower, Retinoblastoma (Rb), Myelocytomatosis (Myc), E2F, Real
Time PCR, Cr stress, salt stress, drought stress

iv
TEŞEKKÜR
Beni bu konuda çalışmaya yönlendiren, tez çalışmamın her aşamasında değerli bilgi,
öneri ve yardımlarını esirgemeyerek, sonsuz destek ve sabır gösteren, akademik
ortamda olduğu kadar beşeri ilişkilerde de engin fikirleriyle yetişme ve gelişmeme
katkıda bulunan danışman hocam, Ankara Üniversitesi Biyoloji Anabilim Dalı Öğretim
Üyesi Sayın Prof. Dr. E. Sümer ARAS’a,
Doktora tez izleme komitemde yer alan ve her konuda bana desteklerini esirgemeyen
Ankara Üniversitesi Öğretim Üyeleri, Sayın Doç Dr. Demet CANSARAN DUMAN’a
ve Doç. Dr. Nur KOÇBERBER’e,
Tez çalışmam süresince Gazi Üniversitesi Yaşam Bilimleri Uygulama ve Araştırma
Merkezi’nin olanaklarını kullanmama imkân sağlayan, Gazi Üniversitesi Tıbbi Biyoloji
ve Genetik Anabilim Dalı Öğretim Üyesi Sayın Doç. Dr. Turgay TEKİNAY’a,
Doktora eğitimim süresince bana her konuda desteklerini esirgemeyen Başkent
Üniversitesi SHMY çalışanlarına,
Her konudaki destekleriyle varlıklarını her an yanımda hissettirerek bana güç veren,
sevgili arkadaşlarım Dr. Bio. Çiğdem VARDAR, Dr. Bio. Esra GÜNDÜZER, Uzman
Bio. Çiğdem DÖNMEZ, Uzm. Bio. Nur ARSLAN, Uzm. Bio. Hatice KARABODUK
ve Dr. Bio. Şeyda FİKİRDEŞİCİ’ye
Her konuda gösterdikleri karşılıksız özveri ve maddi manevi her türlü destekle beni
bugünlere getiren canım aileme,
Doktora eğitimimde 2211 Yurtiçi Doktora Bursu ile bana maddi destek sağlayan
Türkiye Bilimsel ve Teknik Araştırma Kurumu Bilim İnsanı Destekleme Daire
Başkanlığı (TÜBİTAK BİDEB)’na,
Bu tez çalışmasına, Lisansüstü Tez Projesi kapsamında, 15L0430003 proje numarası ile
destekte bulunan Ankara Üniversitesi Bilimsel Araştırma Projeleri Koordinatörlüğü’ne
en derin duygularla teşekkür ederim.
Esin BAŞARAN
Ankara, Şubat 2016

v
İÇİNDEKİLER
TEZ ONAY SAYFASI
ETİK………………………………………….…………………………………….……i
ÖZET……………………………………………………………………………......…..ii
ABSTRACT…………………………………………………………………….….…..iii
TEŞEKKÜR…………………………………………………………………….….…..iv
SİMGELER DİZİNİ………………………………………….………………..…...…ix
ŞEKİLLER DİZİNİ……………………………………………………………..……...x
ÇİZELGELER DİZİNİ……………………………………………………………....xiv
1. GİRİŞ……………………………………………………………..…...………..…….1
1.1 Ayçiçeği (Helianthus annuus L.)……………………………..…………………….3
1.1.1 Tarihçe……………………………...……………………………………………..3
1.1.2 Sınıflandırılması………………………………...…………………………….…..3
1.1.3 Ayçiçeğinin toprak ve iklim istekleri……………………………………..……..4
1.1.4 Ekonomik önemi ve Türkiye’deki üretimi…………………………………...…5
1.1.5 Ayçiçeği çeşitleri……………………………...…………………………….…….7
1.1.5.1 08-TR-003…………………………………………………………………….…8
1.1.5.2 TR-3080………………………………………...………………………….……8
1.1.5.3 SARAY……………………………………...…………………………….…….8
1.1.5.4 TARSAN 1018……………………………………………………..……………9
1.2 Bitkilerde Stres…………………………………………..………………...……….9
1.2.1 Kuraklık stresi……………………………………………………………...…….9
1.2.2 Tuz stresi…………………………………………………..…………….………10

vi
1.2.3 Ağır metal stresi………………………………………..………………………..11
1.2.3.1 Krom (Cr) stresi…………………………………………………....………….12
1.3 Hücre Döngüsü……………………………………………………………...…….13
1.3.1 Bitkilerde temel hücre döngüsü mekanizması…………………………...…....14
1.3.2 Bitkilerde hücre döngüsü kontrolü…………………………………………….15
1.3.2.1 Hücre döngüsü düzenlenmesinde transkripsiyonel kontrol…………….….18
1.4 MYC Ailesi…………………………...……………………………………………21
2. KAYNAK ÖZETLERİ………………………………..…………………………....23
3. MATERYAL VE YÖNTEM……………………………………………………....33
3.1 Bitki Materyalinin Temin Edilmesi……………………………...……………....33
3.2 Hoagland Besi Ortamının Hazırlanması……………………...…………………34
3.3 Bitki Stres Koşulları…………………………………………………..………......36
3.3.1 100 mM ve 150 mM Tuz (NaCl) Stresi…………………………….…….…….36
3.3.2 100 mM ve 150 mM Kuraklık (PEG) Stresi………………………………..….36
3.3.3 Cr(VI) stresi…………………………………………………………..………....38
3.4 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)…………...………..….40
3.5 Total Protein Analizi…………………..…………………………………….........41
3.6 Total RNA İzolasyonu ………………………………………………………........42
3.6.1 TRIGent RNA İzolasyon Protokolü ……………………………………...…....42
3.7 DNaz Uygulaması…………………………………………………………………43
3.8 cDNA (Komplementer DNA) Sentezi…………………………………………...43
3.9 Primer Dizaynı………………………………………………………………..…...45
3.10 Real-time PCR Uygulaması……………………………………………….…….46
3.11 Normalizasyon ve İstatistiksel Hesaplamalar……………………...…………..48

vii
4. BULGULAR……………………………………………………...…………….…...50
4.1 Ayçiçeği Bitkisinde Stres Uygulamaları…………………………………….…...50
4.1.1 Tuz stresi………………………………………………………………….......…50
4.1.2 Kuraklık stresi………………………………………………………………......50
4.1.3 Cr(VI) stresi…………………………………………………………………......50
4.2 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)………………...…..….64
4.3 Total Protein İzolasyonu……………………………………………………….....68
4.4 Total RNA izolasyonu………………………………………………………….....72
4.5 DNaz Uygulaması……………………………………………………………..…..72
4.6 cDNA sentezi……………………………………………………….…………..….72
4.7 Real - Time PCR Reaksiyonu………………………………………………….....76
4.8 Normalizasyon ve İstatistiksel Analiz…………………………………………....81
4.8.1 100 mM tuz (NaCl) stresi sonuçlarının normalizasyon ve istatistiksel
analizi..….…………………………………………………………..…………...82
4.8.2 150 mM tuz (NaCl) stresi sonuçlarının normalizasyon ve istatistiksel
analizi………………………………..…………………………………………100
4.8.3 100 mM PEG stresi sonuçlarının normalizasyon ve istatistiksel analizi…...118
4.8.4 150 mM PEG stresi sonuçlarının normalizasyon ve istatistiksel analizi…...136
4.8.5 Cr (VI) stresi sonuçlarının normalizasyon ve istatistiksel analizi……..……154
5. TARTIŞMA VE SONUÇ…………………………………………………………190
5.1 Total Protein Miktarının Belirlenmesi (Bradford Yöntemi)……………….…190
5.2 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)………………………196
5.3 E2F Geni İfade Seviyesinin Belirlenmesi……………………………….………201
5.3.1 100 mM ve 150 mM NaCl stresi ile E2F seviyesinde meydana gelen
değişim.…………………………………………………………………………203

viii
5.3.2 100 mM ve 150 mM PEG stresi ile E2F seviyesinde meydana gelen
değişim………………………………………………………………………….205
5.3.2 Cr(VI) stresi ile E2F gen ifade seviyesinde meydana gelen değişim…...…...206
5.4 RBR Geni İfade Seviyesinin Belirlenmesi...........................................................208
5.4.1 100 mM ve 150 mM NaCl stresi ile RBR seviyesinde meydana gelen
değişim………………………………………………………………………….211
5.4.2 100 mM ve 150 mM PEG stresi ile RBR seviyesinde meydana gelen
değişim………………………………………………………………………….212
5.4.3 Cr (VI) stresi ile RBR seviyesinde meydana gelen değişim………………....214
5.5 MYC2 Geni İfade Seviyesinin Belirlenmesi........................................................216
5.5.1 100 mM ve 150 mM NaCl stresi ile MYC2 seviyesinde meydana gelen
değişim……………………………..…………………………………………...217
5.5.2 100 mM ve 150 mM PEG stresi ile MYC2 seviyesinde meydana gelen
değişim…………………...……………………………………...……………...220
5.5.3 Cr(VI) stresi ile MYC2 seviyesinde meydana gelen değişim……….……….222
6. SONUÇ…………………………………………………………………….………223
KAYNAKLAR……………………………………………………………...………..224
ÖZGEÇMİŞ……..………………………………………………………………..….238

ix
SİMGELER DİZİNİ
°C Santigrad
dH2O
µg
Distile su
Mikrogram
µl Mikrolitre
µM Mikromolar
Ca (NO3)2. 4H2O Kalsiyum Nitrit
Ca+2
Kalsiyum
Cl- Klor
CuSO4.5H2O Bakır Sülfat
H3BO3 Borik Asit
K Potasyum
K2SO4 Potasyum Sülfat
KCl Potasyum Klorür
KH2PO4 Potasyum Hidrojen Fosfat
MgSO4.7H2O Magnezyum Sülfat
mM Milimolar
MnSO4 Mangan Sülfat
Na Sodyum
NaCl Sodyum Klorür
NH4Mo Amonyum Molibdat
ZnSO4.7H2O Çinko Sülfat
Kısaltmalar
A Adenin
ABA Absisik asit
ACT Aktin
APC Anafaz ilerletici kompleks
C Sitozin
CDC Siklin bağımlı kinaz
cDNA Tamamlayıcı DNA
CYC Siklin
DEPC Dietil pyrokarnonat
DP Dimerizasyon partneri
E2F Adenovirus E2 promotor bağlanma faktörü
FeEDTA Demir Etilen Diamin Tetra Asetik Asit
G Guanin
MDA Malondialdehit
MYC Myelocytomatosis
PEG Polietilen Glikol
Rb Retinoblastoma
RBR Retinoblastoma ilişkili protein
T Timin

x
ŞEKİLLER DİZİNİ
Şekil 1.1 Türkiye bitkisel ham yağ üretimi(binton)…………………..…………….…...7
Şekil 1.2 Hücre döngüsünün evreleri………………………….…...….………….........14
Şekil 1.3 Bitkilerde temel hücre döngüsü mekanizması…………………………..........14
Şekil 1.4 Bitki hücre döngüsünün şematik görünümü ……..……………………..........16
Şekil 1.5 Durgun durumdan bölünme fazına girişte kromozom modellemesinin
olası rolü ………………………………………………...…...……………...17
Şekil 1.6 Bitkilerde hücre döngüsü ilerlemesi sırasında kilit kontrol
noktalarının modeli………………………………..……………………..….18
Şekil 1.7 Hücre döngüsünde transkripsiyonel kontrol………………………..…….......19
Şekil 1.8 Hücre döngüsü kontrolü………………………………………….………......20
Şekil 3.1 SANYO MLR-351H bitki büyütme kabininde bitkilerin geliştirilmesi...........33
Şekil 3.2 Her gün Hogland besin solüsyonu ile sulanarak on beş gün geliştirilen
ayçiçeği çeşitleri…………………………………………...……………...…35
Şekil 3.3 SANYO MLR-351H bitki büyütme kabininde yetiştirilen bitkilere
Hoagland besi ortamı içerisinde tuz (NaCl) ve kuraklık (PEG)
stresinin uygulanması…………………………………………………...…...37
Şekil 3.4 Farklı derişimlerde hazırlanan Cr(VI) çözeltileri……………….........……...38
Şekil 3.5. SANYO MLR-351H bitki büyütme kabininde yetiştirilen bitkilere
Hoagland besi ortamı içerisinde Cr(VI) stresinin uygulanması……….......39
Şekil 4.1 100 mM ve 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidi…...51
Şekil 4.2 100 mM ve 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidi……..52
Şekil 4.3 100 mM ve 150 mM NaCl stresi uygulanmış Saray ayçiçeği çeşidi………....53
Şekil 4.4 100 mM ve 150 mM NaCl stresi uygulanmış Tarsan 1018 ayçiçeği
çeşidi…………………………………………………………………….......54
Şekil 4.5 100 mM ve 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidi.,,,....55
Şekil 4.6 100 mM ve 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidi….......56

xi
Şekil 4.7 100 mM ve 150 mM PEG stresi uygulanmış Saray ayçiçeği çeşidi…………57
Şekil 4.8 100 mM ve 150 mM PEG stresi uygulanmış Tarsan 1018 ayçiçeği
çeşidi………………………...………………………………………………58
Şekil 4.9 Cr(VI) stresinin 24 saat uygulanmasının ardından bitkilerin genel
görüntüsü…………………..………………………………………………...59
Şekil 4.10 08-TR-003 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde
Cr(VI) stresi uygulanan örnekler…………………….……...……………..60
Şekil 4.11 TR-3080 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI)
stresi uygulanan örnekler……………………….………………………….61
Şekil 4.12 Saray ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI)
stresi uygulanan örnekler……………..……...………………………….....62
Şekil 4.13 Tarsan 1018 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde
Cr(VI) stresi uygulanan örnekler…………………………………………..63
Şekil 4.14 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri….....64
Şekil 4.15 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri…….65
Şekil 4.16 100 PEG stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri………....66
Şekil 4.17 150 PEG stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri,,,,,...........66
Şekil 4.18 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği
çeşitlerinde MDA düzeyleri……….…………………….………………...67
Şekil 4.19 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerine ait total
proteinlerin değişimi…………………...…………………………….…….68
Şekil 4.20 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerine ait total
proteinlerin değişimi…………………………………………………..…...69
Şekil 4.21 100mM PEG stresi uygulanmış ayçiçeği çeşitlerine ait total
proteinlerin değişimi……………….………………...………………..…...70
Şekil 4.22 150mM PEG stresi uygulanmış ayçiçeği çeşitlerine ait total
proteinlerin değişimi…………………………………………………….....70
Şekil 4.23 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği
çeşitlerine ait total proteinlerin değişimi……………..……………………71
Şekil 4.24 İzole edilen RNA örneklerinin DNase enzimi uygulaması öncesi
%1,2’lik agaroz jel görüntüsü……………………..…………………….....72

xii
Şekil 4.25 Çalışılan bazı örneklerde E2F geninin erime eğrisi analizi…………..…......77
Şekil 4.26 Çalışılan bazı örneklerde MYC2 geninin erime eğrisi analizi……………...77
Şekil 4.27 Çalışılan bazı örneklerde RBR geninin erime eğrisi analizi…………..…....78
Şekil 4.28 Çalışılan bazı örneklerde ACT geninin erime eğrisi analizi…………..…....78
Şekil 4.29 Çalışılan bazı örneklerde E2F geninin Real Time PCR pik profilleri……....79
Şekil 4.30 Çalışılan bazı örneklerde MYC2 geninin Real Time PCR pik
profilleri…………………………………………………………….…….....80
Şekil 4.31 Çalışılan bazı örneklerde ACT geninin Real Time PCR pik profilleri……..80
Şekil 4.32 Çalışılan bazı örneklerde RBR geninin Real Time PCR pik profilleri……..81
Şekil 4.33 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
E2F gen ifade düzeyleri…………………………………………….……...84
Şekil 4.34 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
RBR gen ifade düzeyleri…………………………………………………...90
Şekil 4.35 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
MYC2 gen ifade düzeyleri……………………...………………………….96
Şekil 4.36 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
E2F gen ifade düzeyleri……………………………………….……….....102
Şekil 4.37 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
RBR gen ifade düzeyleri………………………………………..………...108
Şekil 4.38 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
MYC2 gen ifade düzeyleri……………...…………………..…………….114
Şekil 4.39 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
E2F gen ifade düzeyleri……………………………………..…………....120
Şekil 4.40 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
RBR gen ifade düzeyleri……………………………………..…………...126
Şekil 4.41 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
MYC2 gen ifade düzeyleri……………………………………..………....132
Şekil 4.42 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
E2F gen ifade düzeyleri…………………………………………...……...138

xiii
Şekil 4.43 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
RBR gen ifade düzeyleri…………………………………..……………...144
Şekil 4.44 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı
MYC2 gen ifade düzeyleri…………………………………..…...……….150
Şekil 4.45 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği
çeşitlerinin zamana bağlı E2F gen ifade düzeyleri………..…….………..159
Şekil 4.46 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği
çeşitlerinin zamana bağlı RBR gen ifade düzeyleri ……………….…..…171
Şekil 4.47 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği
çeşitlerinin zamana bağlı MYC2 gen ifade düzeyleri …………….......….183

xiv
ÇİZELGELER DİZİNİ
Çizelge 1.1 Ayçiçeğinin Sınıflandırılması……………………………………………….4
Çizelge 1.2 Türkiye İstatistik Kurumu verilerine göre bölgelere göre 2014 yılında
yağlık ayçiçeği üretimi...................................................................................6
Çizelge1.3 Türkiye İstatistik Kurumu verilerine göre yıllara göre yağlık
ayçiçeği üretimi…………………………………...………………………...6
Çizelge 3.1 Hoagland besi ortamı makro besin çözeltisi içeriği……………………….34
Çizelge 3.2 Hoagland besi ortamı mikro besin çözeltisi içeriği………………………..34
Çizelge 3.3 Hoagland besi ortamında bulunan iyonların son konsantrasyonları……….34
Çizelge 3.4 Total RNA - primer karışımı……………………………………................43
Çizelge 3.5 Ters Transkripsiyon (RT-PCR) Reaksiyonu Komponentleri………...…....44
Çizelge 3.6 Ters transkripsiyon reaksiyonunun gerçekleştirildiği program……………44
Çizelge 3.7 Çalışmada kullanılan gen bölgeleri için tasarlanan primer dizileri………. 46
Çizelge 3.8 Real Time PCR reaksiyonu komponentleri………………………………..47
Çizelge 3.9 Real Time PCR reaksiyonunun gerçekleştiği program…………………....48
Çizelge 4.1 100 mM ve 150 mM NaCl (Tuz) ve 100mM ve 150 mM Polietilen
glikol (PEG) stresi uygulanmış ayçiçeği bitkilerinden elde edilen RNA
örneklerinden sentezlenmiş cDNA’ların saflık ve miktar tayinleri………..73
Çizelge 4.2 Farklı derişimlerde Cr(VI) stresi uygulanmış ayçiçeği bitkilerinden
elde edilen RNA örneklerinden sentezlenmiş cDNA’ların saflık ve
miktar tayinleri……………………………………………………………..75
Çizelge 4.3 100 mM Tuz (NaCl) stresi uygulanmış örneklerin E2F genine ait
normalize edilmiş gen ekspresyonu verileri…………….………………....82
Çizelge 4.4 100 mM Tuz (NaCl) stresi uygulanan örneklerin E2F genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart hata
ve standart sapma değerleri……………………………………………......83
Çizelge 4.5 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri.…….85

xv
Çizelge 4.6 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri………………………………..…………85
Çizelge 4.7 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri……..85
Çizelge 4.8 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……………………………….………….86
Çizelge 4.9 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri……..86
Çizelge 4.10 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……….……………..….………………86
Çizelge 4.11 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri........................................................................................................87
Çizelge 4.12 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi verileri……………………………………87
Çizelge 4.13 100 mM Tuz (NaCl) stresi uygulanmış örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verileri…………………………...…88
Çizelge 4.14 100 mM Tuz (NaCl) stresi uygulanan örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri……………………………….………...89
Çizelge 4.15 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………....91
Çizelge 4.16 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri…………………………..………91
Çizelge 4.17 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………...….91
Çizelge 4.18 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri……..…………………………....92
Çizelge 4.19 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………...……….92

xvi
Çizelge 4.20 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri…..………………………………92
Çizelge 4.21 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………...……….93
Çizelge 4.22 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri………………………..…………93
Çizelge 4.23 100 mM Tuz (NaCl) stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri……………...................……94
Çizelge 4.24 100 mM Tuz (NaCl) stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri …………………………….…………..95
Çizelge 4.25 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………...….97
Çizelge 4.26 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………………………...…97
Çizelge 4.27 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri……………………………………………………...…………….97
Çizelge 4.28 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………………………...…98
Çizelge 4.29 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri……………………………………………………….………..….98
Çizelge 4.30 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri…………………………..……98
Çizelge 4.31 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………...…99
Çizelge 4.32 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………….………….……99
Çizelge 4.33 150 mM Tuz (NaCl) stresi uygulanmış örneklerin E2F genine ait
normalize edilmiş gen ekspresyonu verileri………………...…………..100

xvii
Çizelge 4.34 150 mM Tuz (NaCl) stresi uygulanan örneklerin E2F genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri……………………………………….101
Çizelge 4.35 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
E2F genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………..103
Çizelge 4.36 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi Verileri………………………………….103
Çizelge 4.37 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..103
Çizelge 4.38 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi Verileri………………………………….104
Çizelge 4.39 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..104
Çizelge 4.40 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……………………………...…….….104
Çizelge 4.41 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………..105
Çizelge 4.42 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi Verileri………………………...………..105
Çizelge 4.43 150 mM Tuz (NaCl) stresi uygulanmış örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verileri…………………….……....106
Çizelge 4.44 150 mM Tuz (NaCl) stresi uygulanan örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri…………………………….………….107
Çizelge 4.45 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………...………………...109
Çizelge 4.46 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri……………………….………..109
Çizelge 4.47 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………...….…..109

xviii
Çizelge 4.48 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi Verileri………………………………...…….110
Çizelge 4.49 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..110
Çizelge 4.50 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri……………………………………….110
Çizelge 4.51 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………...……………...111
Çizelge 4.52 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri……………….………….……..111
Çizelge 4.53 150 mM Tuz (NaCl) stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri……………………...……..112
Çizelge 4.54 150 mM Tuz (NaCl) stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri…………………………………….….113
Çizelge 4.55 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………......115
Çizelge 4.56 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………………...………..115
Çizelge 4.57 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………...………...5
Çizelge 4.58 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…………………………………….....116
Çizelge 4.59 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri......116
Çizelge 4.60 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…………………………………...…..116
Çizelge 4.61 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………...………...117

xix
Çizelge 4.62 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………………………….117
Çizelge 4.63 100 mM PEG stresi uygulanmış örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verileri…………………………….……….....118
Çizelge 4.64 100 mM PEG stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri………………………………………………………….119
Çizelge 4.65 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..121
Çizelge 4.66 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri…………………...…………………..121
Çizelge 4.67 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..121
Çizelge 4.68 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……………………………………….122
Çizelge 4.69 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri......122
Çizelge 4.70 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri…………………..…………………...122
Çizelge 4.71 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………..123
Çizelge 4.72 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi verileri…………………………………..123
Çizelge 4.73 100 mM PEG stresi uygulanmış örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verileri………………………...……………...124
Çizelge 4.74 100 mM PEG stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri………………………………………………………….125
Çizelge 4.75 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..127
Çizelge 4.76 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri…...…………………………………..127

xx
Çizelge 4.77 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..127
Çizelge 4.78 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri……………………………………….128
Çizelge 4.79 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri......128
Çizelge 4.80 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri……………………………………….128
Çizelge 4.81 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………..129
Çizelge 4.82 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri……………………..…………..129
Çizelge 4.83 100 mM PEG stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri……………………..……...130
Çizelge 4.84 100 mM PEG stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri……….……………………………….131
Çizelge 4.85 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri……………………………………………………………...…...133
Çizelge 4.86 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri………………….………………..…..133
Çizelge 4.87 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri......133
Çizelge 4.88 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri………………...……………………..134
Çizelge 4.89 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri......134
Çizelge 4.90 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…………………………………….…134

xxi
Çizelge 4.91 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri…………………………………………………………………..135
Çizelge 4.92 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri…………………………...…..135
Çizelge 4.93 150 mM PEG stresi uygulanmış örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verileri………………………...……………...136
Çizelge 4.94 150 mM PEG stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri………………………………..……………….137
Çizelge 4.95 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..139
Çizelge 4.96 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri…………………...…………………..139
Çizelge 4.97 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..139
Çizelge 4.98 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri………………………………...……..140
Çizelge 4.99 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…..140
Çizelge 4.100 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri…………………...……………..…..140
Çizelge 4.101 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri……………………………………………………..……..……141
Çizelge 4.102 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
E2F genine ait Post Hoc Testi verileri………………………….……..141
Çizelge 4.103 150 mM PEG stresi uygulanmış örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verileri…………………………...142
Çizelge 4.104 150 mM PEG stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri……………………….…………...………….143

xxii
Çizelge 4.105 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………….……...145
Çizelge 4.106 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri………………………………..145
Çizelge 4.107 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....145
Çizelge 4.108 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri……………………………….……..146
Çizelge 4.109 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....146
Çizelge 4.110 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri…………………………….………..146
Çizelge 4.111 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………....147
Çizelge 4.112 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
RBR genine ait Post Hoc Testi verileri………………………………..147
Çizelge 4.113 150 mM PEG stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri…………………………...148
Çizelge 4.114 150 mM PEG stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart
hata ve standart sapma değerleri……………...……………………….149
Çizelge 4.115 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………....151
Çizelge 4.116 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri………………….…………..151
Çizelge 4.117 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…151
Çizelge 4.118 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…………………………….………..152
Çizelge 4.119 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....152

xxiii
Çizelge 4.120 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri………………………….…………..153
Çizelge 4.121 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri………………………………………………………………....153
Çizelge 4.122 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri……………………………..153
Çizelge 4.123 Cr (VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait normalize edilmiş gen ekspresyonu verileri………………………..154
Çizelge 4.124 Cr (VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait normalize edilmiş gen ekspresyonu verileri………………………..155
Çizelge 4.125 Cr (VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait normalize edilmiş gen ekspresyonu verileri………………………..156
Çizelge 4.126 Cr (VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait normalize edilmiş gen ekspresyonu verileri……………......157
Çizelge 4.127 Cr (VI) stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri…………………………………………….....158
Çizelge 4.128 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…...…....160
Çizelge 4.129 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…...……160
Çizelge 4.130 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri……………………………………………...161
Çizelge 4.131 Cr(VI) NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……………………………….……..162
Çizelge 4.132 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…...……163
Çizelge 4.133 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....163
Çizelge 4.134 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri……………………………………………...164

xxiv
Çizelge 4.135 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri……………………………….……..165
Çizelge 4.136 Cr (VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine
ait normalize edilmiş gen ekspresyonu verileri……………………..…166
Çizelge 4.137 Cr (VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait normalize edilmiş gen ekspresyonu verileri………………………..167
Çizelge 4.138 Cr (VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait normalize edilmiş gen ekspresyonu verileri………………………..168
Çizelge 4.139 Cr (VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait normalize edilmiş gen ekspresyonu verileri………….…….169
Çizelge 4.140 Cr (VI) stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri……………………………………………….170
Çizelge 4.141 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri…...……172
Çizelge 4.142 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri………...172
Çizelge 4.143 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri…………………………………………...…173
Çizelge 4.144 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri…………….………………………………..174
Çizelge 4.145 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) Verileri………...175
Çizelge 4.146 Cr (VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....175
Çizelge 4.147 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri……………………………………………...176
Çizelge 4.148 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri…………….………………………..177
Çizelge 4.149 Cr (VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait normalize edilmiş gen ekspresyonu verileri………………..178
Çizelge 4.150 Cr (VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait normalize edilmiş gen ekspresyonu verileri………………..179

xxv
Çizelge 4.151 Cr (VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait normalize edilmiş gen ekspresyonu verileri………………..180
Çizelge 4.152 Cr (VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait normalize edilmiş gen ekspresyonu verileri………..181
Çizelge 4.153 Cr (VI) stresi uygulanan örneklerin MYC2 genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri……………………………………………….182
Çizelge 4.154 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....184
Çizelge 4.155 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....184
Çizelge 4.156 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…….………………………………..185
Çizelge 4.157 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…….………………………………..186
Çizelge 4.158 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....187
Çizelge 4.159 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri....187
Çizelge 4.160 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri…………….………………………..188
Çizelge 4.161 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri………………………….…………..189

1
1. GİRİŞ
Bitkisel üretimde stres, bitkilerin yaşadığı ortamda bir veya birden fazla biyotik ve
abiyotik etkenin, büyüme ve gelişmeyi olumsuz yönde etkileyerek, verim düşüklüğü ile
sonuçlanan bir dizi gerilemeye neden olmaktadır. Kuraklık, yetersiz beslenme, tuzluluk,
düşük ve yüksek sıcaklık, toprak ve atmosfer kirliliği, radyasyon gibi etmenler bitkisel
üretimde verimi sınırlandıran abiyotik streslerdir (Lawlor 2002).
Abiyotik stres faktörleri özellikle ekonomik değeri olan bitkiler için en önemli
sınırlayıcı stres tipidir ve kuraklık bu stres tipinde en çok zarar veren faktördür (Bray
2007, Foolad 2007). Ülkemizdeki su kaynakları ve tarım sektörü son yirmi yılda
tekrarlanan kuraklık dönemlerinden büyük zarar görmüştür (Sönmez vd. 2005).
Özellikle bitkilerin büyüme evrelerinde oldukça etkili olan kuraklık, önemli miktarda
tarımsal ürün kayıplarına neden olmaktadır (Bray 2007). Kuraklık ve tuzluluk stresi,
dünya çapında ekilebilir alanların %10 azalmasına ve %50 ürün kaybına neden
olmaktadır (Bray vd. 2000).
Ağır metaller bitkilerde göze çarpan abiyotik stres kaynaklarından biridir. Ağır
metallerin çoğu biyolojik yapılarda birikme, protein ya da nükleik asitler gibi
biyomoleküllerle bağlanma eğilimlerinden dolayı oldukça zararlıdır (Yu 2005).
Stres etkenleri bitki gelişimini eş zamanlı olarak etkileyebilirken, bu stres etkenlerine
karşı tolerans ya da hassasiyet oluşumu anlaşılması güç olaylardır (Chinnusamy vd.
2004). Ayçiçeği bitkisinde çalışmada konu olan hücre döngüsü genleri Retinoblastoma
(Rb), Myelocytomatosis (Myc) ve E2F’dir.
E2F bir çeşit transkripsiyon faktörüdür. Abiyotik strese karşı tolerans oluşumunda
transkripriyon faktörlerinin rollerinin de olduğu tespit edilmiştir. E2F metabolik yolu
bitkilerde ve hayvanlarda korunmuş durumdadır (De Veylder vd. 2003, Dewitte ve
Murray 2003, Inze 2005). Heterodimerik E2F-DP transkripriyon faktörleri DNA

2
replikasyonu ve hücre döngüsü için gerekli genlerin transkripsiyonunu düzenleyerek
hücre döngüsünü kontrol ederler (Helin 1998, Lavia ve Jansen-Dürr 1999).
Bitki retinoblastoma proteinleri nükleerdir ve CDK fosforilasyon bölgeleri ile
karakterize edilirler. Hem hayvanlar hem de bitkilerde Rb proteinleri birçok E2F
transkripsiyon faktörü ailesi ile etkileşime girer. Rb proteini, hücre döngüsü kontrolü ve
hücre farklılaşması arasında bir bağlantı bulunmaktadır (Dewitte ve Murray 2003).
Myelocytomatosis ilişkili gen aile (MYC), bütün ökaryotlarda mevcut olan bir basic-
Helix-Loop-Helix (bHLH) domain içeren transkripsiyon faktörleri alt gen ailesini temsil
eder. Myc hücre döngüsü sırasında G1 fazında ortaya çıkmaya başlar ve G1/S kontrol
noktasındaki çeşitli anahtar hücre döngüsü genlerini hedef alır (Eilers 1999).
Çalışmada kullanılan bitki, Ayçiçeği (Helianthus annuus L.), dünyada ve ülkemizde
günümüzün en önemli yağ bitkilerinden biridir. Ülkemizin bitkisel yağ ihtiyacı, artan
nüfusla beraber artış göstermektedir. Ülkemizde genellikle ayçiçeği yağının tercih
edilmesi ve gerekli bitkisel yağın önemli bir miktarının dışarıdan ithal edilmesi,
ayçiçeğinin önemini daha da arttırmaktadır (Kaya 2004).
Ayçiçeği yağı yemeklik kalitesi yönünden en çok tercih edilen bitkisel yağdır.
Dolayısıyla Dünya’da birçok ülkede ekonomik düzeyde tarımı yapılmaktadır.
Yurdumuzda da yıllara göre değişmekle beraber yaklaşık 550-600.000 hektar arasında
ayçiçeği ekilmektedir. Ayçiçeği bitkisinde strese karşı hücre döngüsündeki
mekanizmalar çözüldüğünde tarımsal anlamda verimin artırılması kaçınılmaz
görülmektedir (Süzer 2014).
Bu çalışmada, ekonomik değeri olan bitkiler için en önemli sınırlayıcı stres tipleri olan
abiyotik stres faktörlerinden Cr(VI), tuz ve kuraklık stresi uygulanmış ayçiçeği
(Helianthus annuus L.) bitkisinde Retinoblastoma (Rb), Myelocytomatosis (Myc) ve
E2F genlerinin mRNA ifade seviyesinin araştırılması amaçlanmıştır. Bu arada total
protein tayini ve MDA analizi gibi fizyolojik parametreler de incelenmiştir.

3
Bu çalışma ile abiyotik stres faktörleri uygulanmış ayçiçeği bitkisinde, hücrenin en
temel yaşamsal olayları arasında yer alan, hücre döngüsündeki düzenleyici
mekanizmaların aydınlatılmasına, ayçiçeğinin ilgili stres koşullarına karşı olan savunma
mekanizmalarının ortaya çıkarılmasına katkı sağlayacak veriler elde edilmiştir.
1.1 Ayçiçeği (Helianthus annuus L.)
1.1.1 Tarihçe
Arkeolojik kanıtlar, Amerikan Kızılderilileri arasında ayçiçeği kullanımını ortaya
koymaktadır (Heiser 1955). Çalışmalar ayçiçeği ekiminin yaklaşık olarak M.Ö. 3000
yıllarında Arizona ve New Mexico’da başladığını gösterir (Semelczi- Kovacs 1975).
Coğrafik yayılım alanı genişlemiştir. Ürün Arkansas’tan Dakota’ya, Ontario’nun
doğusuna, Kanada ve Pensilvanya’ya yayılmıştır. Birçok kanıt evcilleştirilmiş
ayçiçeğinin Kuzey Amerika Kızılderili kültüründe tarih öncesi dönemde var olduğunu
göstermektedir. Whiting (1939) ayçiçeğinin mısırdan önce evcilleştirilmiş olabileceği
görüşündedir. Lees (1965), arkeologların, radyokarbon tarihleme yöntemi kullanarak,
2800 yıl önce Mississippi-Missoniuri Havzası’nda ayçiçeği kanıtlarını oluşturduklarını
belirtmektedir.
1.1.2 Sınıflandırılması
Ayçiçeği (Helianthus annuus), papatyagiller (Asteraceae) familyasından çekirdekleri ve
yağı için yetiştirilen sarı çiçekli bir tarım bitkisidir. Sistematik sınıflandırılması çizelge
1.1’de verilmiştir (Lınnaeus 1753).

4
Çizelge 1.1 Ayçiçeğinin sınıflandırılması
Domain: Eukarya (Ökaryotikler)
Alem: Plantae (Bitkiler)
Bölüm: Magnoliophyta (Kapalı tohumlular)
Sınıf: Magnoliopsida (İki çenekliler)
Takım: Asterales
Aile: Asteraceae (Papatyagiller)
Cins: Helianthus
Tür Helianthus annuus
İkili adı: Helianthus annuus L.
1.1.3 Ayçiçeğinin toprak ve iklim istekleri
Ayçiçeği yetişeceği toprak tipi yönünden çok seçici olmamasına rağmen organik
maddece zengin, derin ve su tutma kapasitesi iyi topraklarda yüksek verim
potansiyeline sahiptir. Kumlu topraklardan ağır yapıdaki killi topraklara kadar her türlü
iyi drenaj sağlanmış topraklarda tarımı yapılabilmektedir.
Ayçiçeğinin tuzluluğa karşı toleransı azdır. Tuzlu topraklarda yetiştirilen ayçiçeğinin
tohumlarının yağ yüzdesinde azalmalar görülmüştür. Ayrıca ayçiçeği yetişecek toprakta
%1-2 düzeyinde bulunacak tuz konsantrasyonunun çimlenmeyi önemli oranda
düşürdüğü belirlenmiştir (Süzer 2014).
Ayçiçeğinin en iyi yetiştiği pH aralığı, 6.0 ile 7.2 arasındadır. Ayçiçeği yüksek ve düşük
sıcaklıklara gelişme dönemine bağlı olarak oldukça toleranslıdır. Tohumlarının en iyi
çimlenebilmesi için 8-10oC’lik toprak sıcaklığı gerekir. Ayçiçeği bitkisi fideleri
kotiledon devresinde -4oC sıcaklığa dayanabilir. Ayçiçeği için en iyi yetişme sıcaklıkları
21 ile 24 oC arasıdır. Genellikle vejetatif dönemde serin, generatif dönemde ise açık ve
güneşli havalar ister.

5
Toprakta kök derinliğinde bitkilerin kullanabileceği faydalı rutubetin bulunup
bulunmaması, ayçiçeğinde tane ve yağ verimini etkileyen en önemli faktördür.
Ayçiçeği bitkisi kazık kök yapısı ile kurağa toleranslı bir bitki kabul edilse de yazlık bir
bitki olması ve bu mevsimde de yeterince yağış düşmemesi sonucu oluşan kuraklık
dekardan alınan verimi oldukça düşürmektedir.
Ayçiçeği yapraklarının ışığa yönelme özellikleri sayesinde fotosentez için gerekli ışığı
rahatlıkla alabilir. Bu ışığa yönelme özelliğinden dolayı ayçiçeğine Trakya ve Marmara
Bölgesinde “günebakan” veya “gündöndü” denilmektedir (Süzer 2014).
1.1.4 Ekonomik önemi ve Türkiye’deki üretimi
Ayçiçeği sağlıklı beslenme açısından önemli bir bitkidir. Ayçiçeği yağı, içerdiği
doymamış yağ asitleri oranının yüksek (% 69) olması nedeniyle de beslenme değeri en
yüksek olan bitkisel yağlardan birisidir. Küspesindeki protein içeriği nedeniyle, yem
olarak hayvan beslenmesinde de kullanılmaktadır. Ayçiçeği yağ, sabun ve boya
sanayinde, sapları da yakacak olarak kullanılmaktadır. Ayrıca ayçiçeği çerezlik olarak
da tüketilmekte olup, ayçiçeği üretiminin % 2,6’sı çerezlik olarak tüketilmektedir
(http://koop.gtb.gov.tr. 2015).
Ayçiçeği, ülkemizde ekimi yapılan yağlı tohumlu bitkiler arasında ekim alanı ve üretim
bakımından birinci sırada yer almaktadır. Ülkemizde yağlık ayçiçeği en fazla Trakya-
Marmara Bölgesinde (% 47,2) üretilmekte olup, Orta Anadolu Bölgesi, % 29,2’lik payla
ikinci sırada yer almaktadır. Geriye kalan ülkemiz ayçiçeği üretiminin % 12’si
Karadeniz, % 8,7 si Akdeniz ve % 2,8’i Doğu ve Güneydoğu Anadolu Bölgelerinde
gerçekleştirilmektedir. Hemen her bölgemizde yetişebilen ve tanelerinde yüksek oranda
ve kaliteli yağ içeren ayçiçeği, ülkemizde yağ bitkileri ekim alanında pamuktan sonra
ikinci sırayı almaktadır.
6
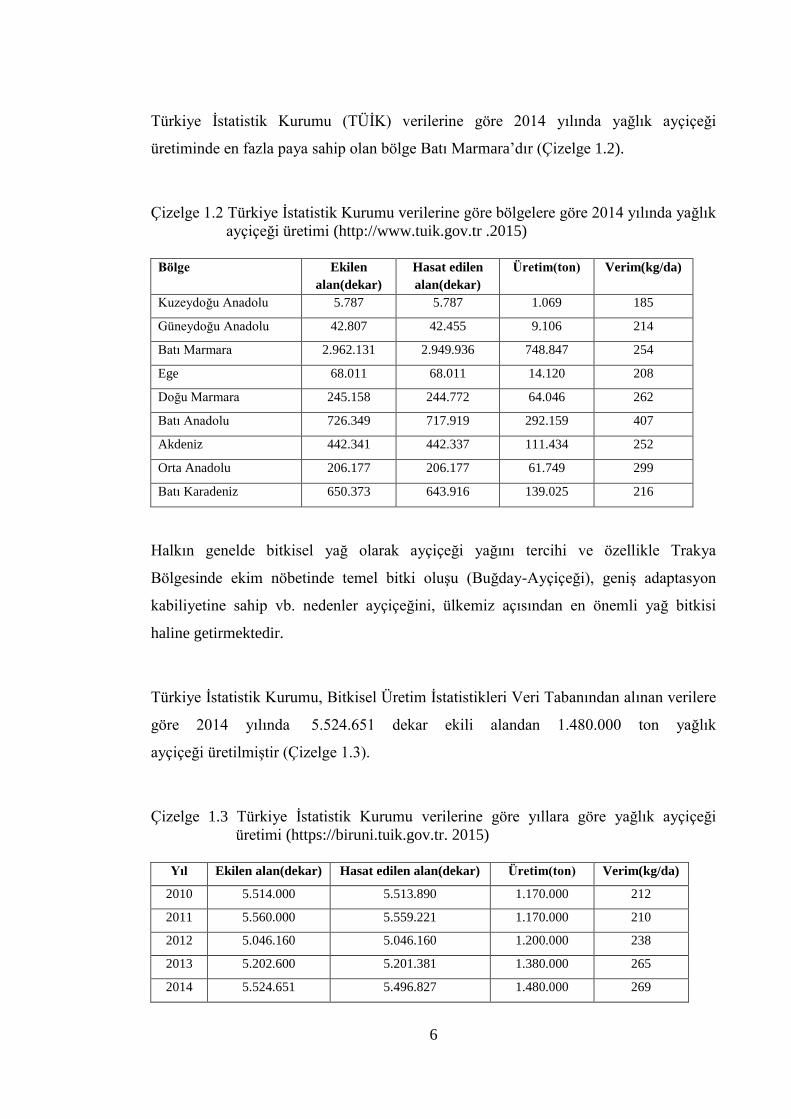
Türkiye İstatistik Kurumu (TÜİK) verilerine göre 2014 yılında yağlık ayçiçeği
üretiminde en fazla paya sahip olan bölge Batı Marmara’dır (Çizelge 1.2).
Çizelge 1.2 Türkiye İstatistik Kurumu verilerine göre bölgelere göre 2014 yılında yağlık
ayçiçeği üretimi (http://www.tuik.gov.tr .2015)
Bölge Ekilen
alan(dekar)
Hasat edilen
alan(dekar)
Üretim(ton) Verim(kg/da)
Kuzeydoğu Anadolu 5.787 5.787 1.069 185
Güneydoğu Anadolu 42.807 42.455 9.106 214
Batı Marmara 2.962.131 2.949.936 748.847 254
Ege 68.011 68.011 14.120 208
Doğu Marmara 245.158 244.772 64.046 262
Batı Anadolu 726.349 717.919 292.159 407
Akdeniz 442.341 442.337 111.434 252
Orta Anadolu 206.177 206.177 61.749 299
Batı Karadeniz 650.373 643.916 139.025 216
Halkın genelde bitkisel yağ olarak ayçiçeği yağını tercihi ve özellikle Trakya
Bölgesinde ekim nöbetinde temel bitki oluşu (Buğday-Ayçiçeği), geniş adaptasyon
kabiliyetine sahip vb. nedenler ayçiçeğini, ülkemiz açısından en önemli yağ bitkisi
haline getirmektedir.
Türkiye İstatistik Kurumu, Bitkisel Üretim İstatistikleri Veri Tabanından alınan verilere
göre 2014 yılında 5.524.651 dekar ekili alandan 1.480.000 ton yağlık
ayçiçeği üretilmiştir (Çizelge 1.3).
Çizelge 1.3 Türkiye İstatistik Kurumu verilerine göre yıllara göre yağlık ayçiçeği
üretimi (https://biruni.tuik.gov.tr. 2015)
Yıl Ekilen alan(dekar) Hasat edilen alan(dekar) Üretim(ton) Verim(kg/da)
2010 5.514.000 5.513.890 1.170.000 212
2011 5.560.000 5.559.221 1.170.000 210
2012 5.046.160 5.046.160 1.200.000 238
2013 5.202.600 5.201.381 1.380.000 265
2014 5.524.651 5.496.827 1.480.000 269
7
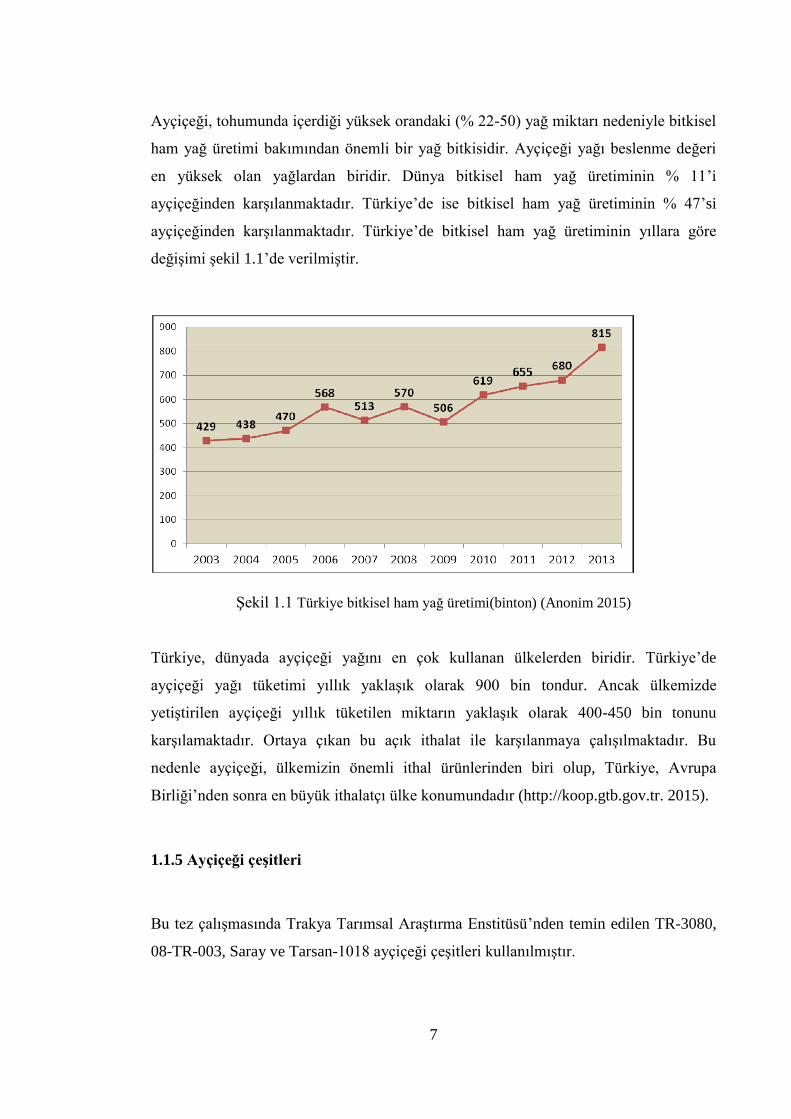
Ayçiçeği, tohumunda içerdiği yüksek orandaki (% 22-50) yağ miktarı nedeniyle bitkisel
ham yağ üretimi bakımından önemli bir yağ bitkisidir. Ayçiçeği yağı beslenme değeri
en yüksek olan yağlardan biridir. Dünya bitkisel ham yağ üretiminin % 11’i
ayçiçeğinden karşılanmaktadır. Türkiye’de ise bitkisel ham yağ üretiminin % 47’si
ayçiçeğinden karşılanmaktadır. Türkiye’de bitkisel ham yağ üretiminin yıllara göre
değişimi şekil 1.1’de verilmiştir.
Şekil 1.1 Türkiye bitkisel ham yağ üretimi(binton) (Anonim 2015)
Türkiye, dünyada ayçiçeği yağını en çok kullanan ülkelerden biridir. Türkiye’de
ayçiçeği yağı tüketimi yıllık yaklaşık olarak 900 bin tondur. Ancak ülkemizde
yetiştirilen ayçiçeği yıllık tüketilen miktarın yaklaşık olarak 400-450 bin tonunu
karşılamaktadır. Ortaya çıkan bu açık ithalat ile karşılanmaya çalışılmaktadır. Bu
nedenle ayçiçeği, ülkemizin önemli ithal ürünlerinden biri olup, Türkiye, Avrupa
Birliği’nden sonra en büyük ithalatçı ülke konumundadır (http://koop.gtb.gov.tr. 2015).
1.1.5 Ayçiçeği çeşitleri
Bu tez çalışmasında Trakya Tarımsal Araştırma Enstitüsü’nden temin edilen TR-3080,
08-TR-003, Saray ve Tarsan-1018 ayçiçeği çeşitleri kullanılmıştır.

8
1.1.5.1 08-TR-003
2008 yılında Türkiye’de ıslah edilmiştir. 2012 yılında tescil edilmiştir. Ortalama % 50.1
yağ oranı ile standart çeşitlerin yağ oranı ortalamasının % 5 üstünde yer almıştır. Orta
erkenci, yüksek verim potansiyeline sahip bir çeşittir.
İri ve dış bükey tablalıdır. Tabla eğimli gövde üzerinde ve kuş zararını azaltacak şekilde
hafifçe aşağı kıvrıktır. Bitkileri orta boya sahip olup sulamaya izin verecek şekildedir.
Orabanşın yeni ırklarına yüksek derecede toleranslı olduğundan, Trakya dahil tüm
ayçiçeği ekilen alanlara önerilmektedir. Pasa karşı, doğal koşullardaki gözlemlere göre
toleranslıdır. Mildiyöye karşı hassastır. Kurağa toleransı iyidir. Yağ oranı yüksek olup
iri tanelidir (http://www.betaziraat.com.tr. 2015).
1.1.5.2 TR-3080
2453-A X 01001-R melezinden geliştirilmiş, hibrit yağlık ayçiçeği çeşididir. Orta boylu,
tablası tam eğik, kuş zararlarına karşı dayanıklıdır. Stres koşullarına dayanaklıdır. Farklı
çevresel koşullara karşı adaptasyon kabiliyeti yüksektir. Çok yüksek yağ içeriğine
sahiptir (%48-50). Optimum bitki yoğunluğuna sahiptir (50,000-55,000 bitki/ha).
Verimi kuru şartlarda 180 - 320 kg/da, sulu şartlarda 360 kg/da’dır. Orobanşın eski
ırklarına dayanıklıdır. Tane verimi ve oranı yüksektir (http://arastirma.tarim.gov.tr.
2015).
1.1.5.3 SARAY
Trakya Tarımsal Araştırma Enstitüsü ve Macaristan Szeged Enstitüsü tarafından 2013
yılında ortaklaşa geliştirilen MO1A X 9758 R melezinden geliştirilen, orta oleik tip
hibrit ayçiçeği çeşididir.

9
İri ve dış bükey tablalıdır. Tabla gövde üzerinde aşağı doğru kıvrık olması nedeniyle,
güneş yanıklığı ve kuş zararına toleranslıdır. Erkenci, yüksek yağ ve tane verim
potansiyeline sahip olup, değişik çevre koşullarına adaptasyonu oldukça yüksektir.
Orobanşın yeni ırklarına yüksek derecede toleranslıdır. Mildiyöye dayanıksızdır. Ayrıca
herbisitlere karşı duyarlıdır. Kurağa toleransı iyidir. Taneler iri ve yağ oranı çok yüksek
olup, orta oleik asit içeriğine sahiptir (http://arastirma.tarim.gov.tr. 2015).
1.1.5.4 TARSAN 1018
2517-A X 0539-R melezinden geliştirilen, hibrit yağlık ayçiçeği çeşididir. Orta boylu,
tablası tam eğik olup erkenci bir çeşittir. Orabanşın eski ırkına dayanaklıdır. Kuraklık
stresine dayanıklıdır. Farklı çevresel koşullara karşı adaptasyon kabiliyeti yüksektir.
Yüksek ürün potansiyeline sahiptir. Çok yüksek yağ içeriğine sahiptir (%45-48).Kuru
şartlarda 160-330 kg/da, sulu şartlarda 360 kg/da verim elde edilmektedir. Marmara,
Karadeniz, Ege, Akdeniz’de kuru şartlarda, İç Anadolu bölgesinde sulu şartlarda
önerilmektedir(http://arastirma.tarim.gov.tr. 2015).
1.2 Bitkilerde Stres
Bitkisel üretimde stres, bitkilerin yaşadığı ortamda bir veya birden fazla biyotik ve
abiyotik etkenin, büyüme ve gelişmeyi olumsuz yönde etkileyerek, verim düşüklüğü ile
sonuçlanan bir dizi gerilemeye neden olmaktadır. Kuraklık, yetersiz beslenme, tuzluluk,
düşük ve yüksek sıcaklık, toprak ve atmosfer kirliliği, radyasyon gibi etmenler bitkisel
üretimde verimi sınırlandıran abiyotik stres faktörleridir (Lawlor 2002).
1.2.1 Kuraklık stresi
Kuraklık stresi, bitkilerde belirli bir süre içerisinde kaybedilen su miktarının çevreden
alınan su miktarından fazla olması durumunda oluşmaktadır. Dünya üzerindeki
kullanılabilir alanlar stres faktörlerine göre sınıflandırıldığında doğal bir stres faktörü

10
olan kuraklık stresi % 26’lık payıyla en büyük dilimi içermektedir (Blum 1986).
Kuraklık stresi büyümeyi ve verimi etkileyen en yaygın çevresel streslerden biri olup
bitkilerde birçok fizyolojik, biyokimyasal ve moleküler cevabı uyarmakta ve buna bağlı
olarak bitkiler, sınırlı çevresel koşullara adapte olmayı sağlayacak tolerans
mekanizmaları geliştirebilmektedir.
Stres durumunda turgor yitmesi, hücre büyümesini olumsuz şekilde etkileyerek
hücrelerin küçük kalmasına ve hücre duvarı sentezinde azalmaya yol açmaktadır
(Pugnaire vd. 1994). Kuraklık etkisi ile bitkilerde yaprakların nisbi nem içeriğinin ve
yaprak su potansiyelinin düşmesi fotosentez oranını azaltmaktadır (Lawlor ve Cornic
2002). Yaprakların absorbe ettiği ışık miktarı ve yararlanılan ışık arasındaki dengenin
bozulması nedeniyle de fotosentez aktivitesi engellenmektedir (Foyer ve Noctor 2000).
Kuraklık durumunda fotosentez oranının azalmasının temel nedenlerinden birinin de
genellikle stoma işlevlerinin sınırlanmasından kaynaklandığı kabul edilmektedir (Cornic
2000). Stomaların kapanması bitkilerin beslenme durumlarını da olumsuz
etkilemektedir (Oren vd. 1999).
1.2.2 Tuz stresi
Bitkilerin yaşamları sürecinde karşılaştıkları stres faktörlerinden biri de aşırı
tuzluluktur. Toprakta bulunan çözünebilir tuzların miktarı, bitkinin büyüme ve
gelişmesi için gerekli olan miktarın üzerine çıktığında sorunlar ortaya çıkmaya başlar.
Toprak çözeltisindeki tuz konsantrasyonu arttığında ve su potansiyeli azaldığında, bitki
hücrelerinin ozmotik potansiyeli düşer ve bitki hücrelerinin bölünmesi ya da uzaması
birden yavaşlar. Bu stres koşulları altında genellikle stomalar kapanır ve sonuç olarak
fotosentez azalır. Stres koşullarının devam etmesi halinde bitki büyümesi tamamen
durabilir (Ashraf 1994).

11
Tuzluluk bitki gelişmesini üç temel prensip çerçevesinde engellemektedir:
1) Su eksikliği (su stresi),
2) Na+ ve Cl
- iyonlarının fazla miktarda alınması nedeniyle oluşan iyon toksisitesi,
3) İyon taşınımında ortaya çıkan dengesizlik nedeniyle hücre içindeki sıvının mineral
yapısının ve Ca+2
dengesinin bozulması (Marschner 1995).
Tuz stresi kurak ve yarı kurak bölgeler ile sulama yapılan alanlarda önemli bir abiyotik
stres faktörüdür. Dünyanın yaklaşık %7’si, kültüre alınmış alanların %20’si ve sulama
yapılabilen alanların neredeyse yarısı tuzluluktan etkilenmektedir (Szabolcs 1994).
1.2.3 Ağır metal stresi
Ağır metaller bitkilerde göze çarpan abiyotik stres kaynalarından biridir. Ağır metallerin
çoğu biyolojik yapılarda birikme ve protein ya da nükleik asitler gibi biyomoleküllerle
bağlanma eğilimlerinden dolayı oldukça zararlıdır. Bazı ağır metallerin kanseri
başlattığı öne sürülmektedir (Yu 2005).
Pozitif yüklü metal iyonlarının, DNA’nın her iki zincirinde bulanan fosfat gruplarındaki
negatif yüklü oksijen atomları ve bazların azot ve oksijen gibi elektron donörü olan
atomları ile reaksiyona girebileceği bildirilmiştir. Buna göre metaller DNA’daki olası
bölgelere direk veya dolaylı olarak sıkıca bağlanabilmektedirler (Anastassopoulou
2003).
Metallerin DNA’ya bağlanmasının genellikle baz eşleşmesini sağlayan hidrojen
bağlarının kırılmasına ve çift zincirli yapının kararlılığının bozulmasına sebep olduğu
bilinmektedir. Nükleik asitlerin yapısında oluşan değişimler metal iyonunun çeşidine
bağlı olarak farklılık göstermektedir (Theophanides 1981).

12
1.2.3.1 Krom (Cr) stresi
Krom (Cr), dünyada en fazla bulunan yedinci elementtir (Katz ve Salem 1994). Yer
kabuğunda en çok bulunan, toksik ağır metallerden biridir (Kabata-Pendias ve Pendias
2001).
Kromun trivalent [kromik formu; Cr(III) veya Cr+3
] ve hekzavalent [kromat formu;
Cr(VI) veya Cr+6
] olarak adlandırılan formları bulunur. Kromun hem trivalent hem de
hekzavalent formları fitotoksiktir (Dixit vd. 2002). Cr(VI), en toksik form olup; kromat
(CrO4-2
) veya dikromat (Cr2O7-2
) şeklinde iki oksianyon formu halinde bulunmaktadır.
Buna karşın daha az mobil ve toksik olan Cr(III); oksitler, hidroksitler ve sülfatlar
şeklinde veya toprakta ve sucul çevrelerde organik bileşiklere bağlı halde bulunmaktadır
(Zayed ve Terry 2003).
Krom yer kabuğunda en çok bulunan, toksik ağır metallerden biridir. Toksik olmasına
rağmen kromun besleyici olmak gibi bir özelliği de vardır (Kabata-Pendias ve Pendias
2001). Bütün bunların yanında kurşun, civa, kadmiyum ve alüminyum gibi ağır metaller
çok uzun zamandır ve fazlaca çalışılmasına rağmen krom bilim insanları tarafından pek
dikkati çekmemiştir (Shanker vd. 2005).
Bitkilerde krom toksisitesi büyüme ve gelişmeyi etkileyip, çimlenme sürecinde
değişiklikler ortaya çıkmasına, bunun yanında kök, gövde ve yaprakların etkilenmesine
de neden olmaktadır. Ayrıca krom fitotoksisitesi pigment, beslenme dengesi,
antioksidan enzim bozulmasına neden olmakta ve oksidatif stresi uyarmaktadır (Panda
ve Khan 2003). Filizlenmenin erken gelişme evreleri krom gibi ağır metallerin toksik
etkilerinin belirlenmesinde çok önemlidir (Sharma vd. 1995, Pandey 2008).

13
1.3 Hücre Döngüsü
Hücre döngüsünün en temel işlevi, kromozomlarda büyük miktarda DNA’yı doğru
olarak eşlemek ve kopyaları genetik olarak eş iki yavru hücreye tam olarak dağıtmaktır.
Hücre döngüsü iki ana evreyi içerir. DNA eşlenmesinin gerçekleştiği S evresi ve
kromozom ayrılması ve hücre bölünmesinin gerçekleştiği M evresi.
Birçok hücre, DNA’sını kopyalamak ve bölünmek için ayırdığı zamandan çok daha
fazlasına, protein kütlesi ve organellerini iki katına çıkarmak ve büyümek için
gereksinim duyar. Büyümeye daha fazla zaman ayırmak için, çoğu hücre döngüsüne
ilave aralık evreleri yerleştirilmiştir. M evresi ve S evresi arasına bir G1 evresi, S evresi
ile mitoz arasına bir G2 evresi. Dolayısıyla ökaryot hücre döngüsü G1, S, G2 ve M
olmak üzere geleneksel olarak 4 ardışık evreye bölünmüştür. G1, S ve G2 bir arada
interfaz olarak da adlandırılır (Şekil 1.2).
G1 evresinin uzunluğu dış koşullar ile diğer hücrelerden gelen hücre dışı sinyallere
bağlı olarak büyük bir değişkenlik gösterir. Eğer hücre dışı koşullar elverişsizse,
hücreler G1’deki ilerlemeyi geciktirir ve hatta G0 (G sıfır) olarak da bilinen özel bir
durgunluk evresine girerler. Bu evrede yeniden çoğalmaya başlamadan önce günler,
haftalar, hatta yıllar boyunca kalabilirler. Eğer hücre dışı koşullar elverişli, büyüme ve
bölünme sinyalleri mevcutsa erken G1 veya G0’daki hücreler G1’in sonunda başlangıç
veya kısıtlama noktası olarak bilinen adanma noktasına doğru ilerlerler. Bu noktayı
geçtikten sonra, hücre büyümesi ve bölünmesini uyaran hücre dışı sinyallerin
yokluğunda dahi, hücreler DNA tıpkıyapımına adanır (Alberts 2008).

14
Şekil 1.2 Hücre döngüsünün evreleri (Alberts 2008)
1.3.1 Bitkilerde temel hücre döngüsü mekanizması
Bitki genomları birçok farklı siklin bağımlı kinazı (CDKs) ve siklinleri (CYCs) kodlar.
Arabidopsis thaliana, 12 farklı CDK ve 49’dan fazla farklı sikline sahiptir. Siklin
bağımlı kinazlar ve siklinler, sekanslarına dayanarak sınıflara ayrılmakta, işlevsel
farklılıkları ile uyumlu olarak sınıflandırılmaktadır.
A ve B tipi CDK’lar bitki hücre döngüsünün önemli yürütücüleridir. A tipi CDK’lar
(CDKA) memelilerdeki CDK1 ve CDK2 ile yakından ilişkilidir. Bunlar G1- S ve G2-M
geçiş noktalarını kontrol etmektedir. B tipi CDK’lar (CDKB) bitkiler için spesifiktir.
Hücre döngüsünün evresine bağlı olarak biraraya gelir. G2-M geçiş evresinde
maksimum değere ulaşır. Bu evredeki aktivitesi gereklidir (De Veylder vd. 2007).
Şekil 1.3 Bitkilerde temel hücre döngüsü mekanizması (De Veylder vd. 2007)

15
CDK’lar ile farklı tipteki siklinlerin sırasal birlikteliği hücre döngüsünün ilerlemesi için
gereklidir. D-tipi siklinler (CYCD), G1-S geçişini düzenler. Bunun yanında G2-M
geçişinde de rolleri olabilir. A-tipi siklinler (CYCA), çoğunlukla S fazından M fazına
geçişte bulunurlar. B-tipi siklinler (CYCB), G2-M geçişinde ve M fazı boyunca en üst
düzeyde bulunurlar (Şekil 1.3).
CDKA-CYCD komplekslerinin aktivitesi, stres uyarıcılar veya gelişimsel sinyallere
yanıt veren CDK inhibe edici proteinlerle (ICK/KRP ve SIM) ilişkileri sayesinde inhibe
edilebilirler. Anafaz ilerletici kompleks (APC) tarafından CYCA ve CYCB’nin seçici
yıkımı mitozdan çıkışı gösterir. Bitki dışındaki türler için tanımlandığı gibi, CDK aktive
edici kinazlar tarafından CDK alt biriminin fosforilasyonu, CDK aktivasyonu için
gereklidir (De Veylder vd. 2007).
1.3.2 Bitkilerde hücre döngüsü kontrolü
Farklı siklin tipleri, farklı substratlarda hedef potansiyeli ve düzenlemenin özgünlüğünü
sağlayan hücre döngüsünün farklı aşamalarının kontrolüne katılmaktadır. Spesifik
olarak, farklı siklinler karakteristik olarak ifade edilir ve G1 fazında, G1’den S fazı ve
G2’den M fazına geçişlerin kontrolünde önemlidir.
Bitkilerde, G1 siklinler, memelilerde mevcut olanlara göre genlerin çok daha büyük
grubu tarafından temsil edilir. Tüm yüksek bitkiler CYCD1-CYCD7 olarak
isimlendirilen siklin D genlerinin altı korunmuş alt grubuna sahiptir. CYCD2 ve
CYCD4 alt grupları dizi homolojisine göre ayırt edilemez (Menges vd. 2007).

16
Şekil 1.4 Bitki hücre döngüsünün şematik görünümü (Scofield vd. 2014)
G1 fazından ilerleme D-tipi siklinlerin (CYCD) bağlanması ile aktif hale gelen CDKA
tarafından RBR proteininin fosforilasyonu yoluyla başlatılır. KRP proteinleri, CYCD ve
CDKA alt birimlerine bağlanabilir ve kompleksin kinaz aktivitesini inhibe edebilir.
A-tipi siklinler S fazı boyunca CDKA ile ilişkilidir. Mitoz özgü CDKB (iki tip CDKB1
ve CDKB2) B-tipi siklinler (CYCB) ile birlikte mitoz için gereklidir. KRP proteinleri,
CDKB kinaz aktivitesi tarafından fosforlanabilir ve inaktive edilebilir, ayrıca mitozda
CDK aktivite seviyeleri artar (Şekil 1.4)(Scofield vd. 2014).
Bitki morfogenezi hücre çoğalması ve farklılaşmasının sıkı kontrolüne dayanır. Diğer
ökaryotlarda olduğu gibi, bitki hücre döngüsü boyunca ilerleme, siklinler (CYCs) olarak
adlandırılan kendi aktivatör proteinleri ile ilişkili sikline bağlı kinazlar (CDKler)
tarafından yönetilir ve CYC-CDK’nın etkinliği transkripsiyonal ve translasyon sonrası
düzeylerde modüle edilir. Hayvanlar ve mayalar ile karşılaştırıldığında, bitkiler, genel
olarak ana hücre döngüsü düzenleyicilerini kodlayan daha çok gen içerirler ve bunların
gelişmede veya çeşitli çevresel değişikliklere yanıtta nasıl fonksiyon gösterdikleri
karmaşıktır (Komaki ve Sugimoto 2012).

17
Hareketsiz veya G1’de durdurulmuş hücrelerde, retinoblastoma ilişkili (RBR) proteini,
G0 veya G1’den S fazına geçiş için gerekli olan düzenleyici genlerin promotorlarında
histon deasetilazları (HDACs) hedef almaktadır.
RBR, E2F-DP transkripsiyon faktörlerinin aktivitesini engeller. Kromatin, E2F-DP
hedef genlerinin transkripsiyonunu önlemek ve transkripsiyonal regülatör proteinlerin
erişilebilirliğini engellemek suretiyle, HDAC aktivitesinin bir sonucu olarak yoğunlaşır
(Şekil 1.5 a) (De Veylder vd. 2007).
Hücre döngüsü girişi, A-tipi siklin bağımlı kinaz (CDKA)- D-tipi siklin komplekslerinin
aktivasyonu ile karakterize edilir. Bu kompleks RBR proteinini fosforile eder ve
dolayısıyla E2F–DP transkripsiyon faktörlerinden HDAC ve RBR serbest bırakılır.
HDAC’ların yokluğunda, hala tanımlanamamış histon asetilazların aktivitesi baskın olur
ve histonların asetilasyonuna sebep olur. Sonuçta, daha az yoğun nükleozomal
konformasyon oluşur. Bu gevşek kromatin yapısı, kromatin kalıba, transkripsiyonal
regülatör proteinlerin bağlanmasına izin verir (Şekil 1.5b).
Şekil 1.5 Durgun durumdan bölünme fazına girişte kromozom modellemesinin olası rolü
(De Veylder vd. 2007)
Çevresel ve içsel sinyaller, Cdc2a ile ilişkili CycD artışına neden olur. Kompleksin
aktivitesi olasılıkla ICKs ve CDK aktive kinazlar (CAKs) tarafından düzenlenir. Geç
G1’de pRb’ı fosforilize eden CycD–CDK’nın artan seviyeleri, pRb tarafından

18
baskılanan E2F traskripsiyon faktörlerini serbest bırakır. E2F bağımlı genlerin
aktivasyonu, S fazına ilerlemeyle sonuçlanır (Şekil 1.6 a) (Meijer ve Murray 2001).
Başlangıçta, wee1 kinaz tarafından fosforilasyonu nedeniyle inaktif olan CycB–CDK
kompleksleri, Cdc25 fosfataz tarafından aktive edilir. Bu da hücrelerin mitoza girmesine
neden olur. Mitozdan çıkış APC ve muhtemelen ccs52 tarafından mitotik siklinlerin
yıkımını gerektirir (Şekil 1.6 b) (Meijer ve Murray 2001).
Şekil 1.6 Bitkilerde hücre döngüsü ilerlemesi sırasında kilit kontrol noktalarının modeli (Meijer
ve Murray 2001)
a. G1 ilerlemesi ve G1 → S kontrolünün aktivasyonu, b. G2→M geçişi.
1.3.2.1 Hücre döngüsü düzenlenmesinde transkripsiyonel kontrol
Bitki hücrelerinde iki önemli periyodik düzenleyici tanımlanmıştır. Bunlar; E2F–DP ve
MYB3R transkripsiyon faktörleridir. Bitki E2F’leri siklin bağımlı kinaz (CDK) – D tipi
siklin (CYCD) kompleksleri tarafından Retinoblastoma ilişkili proteinin (RBR)
fosforilasyonunu takiben aktive edilir (Şekil 1.7 a).
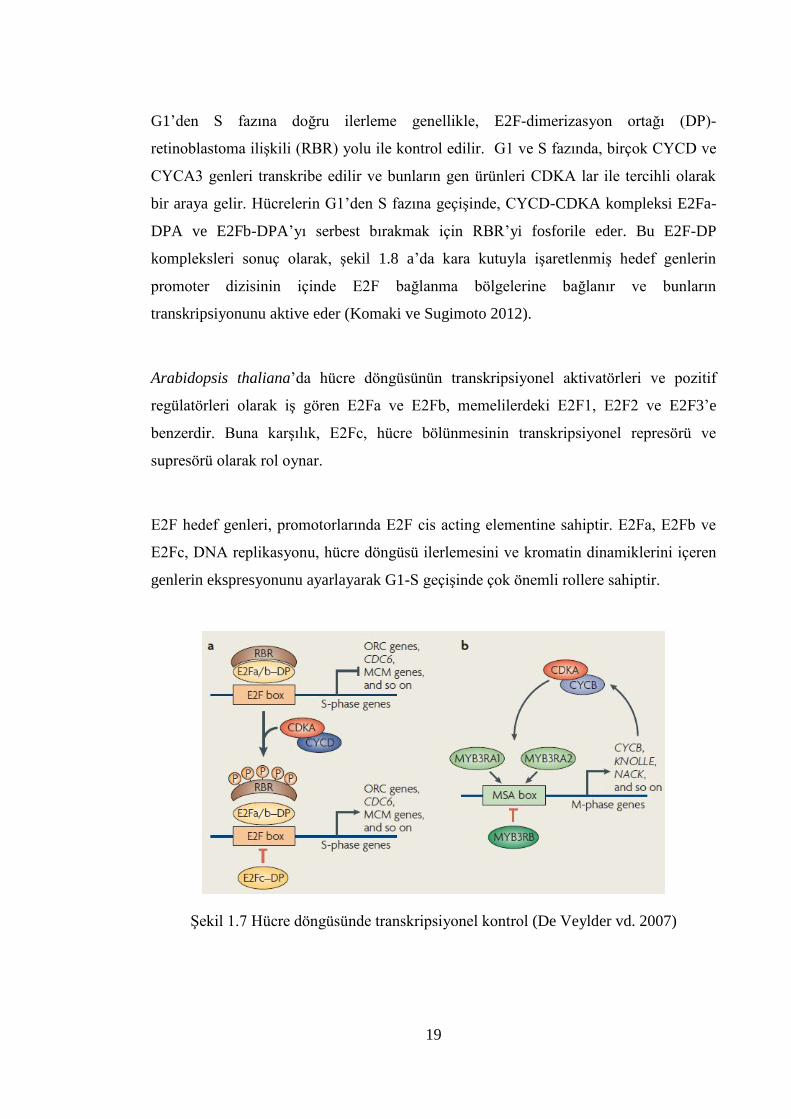
19
G1’den S fazına doğru ilerleme genellikle, E2F-dimerizasyon ortağı (DP)-
retinoblastoma ilişkili (RBR) yolu ile kontrol edilir. G1 ve S fazında, birçok CYCD ve
CYCA3 genleri transkribe edilir ve bunların gen ürünleri CDKA lar ile tercihli olarak
bir araya gelir. Hücrelerin G1’den S fazına geçişinde, CYCD-CDKA kompleksi E2Fa-
DPA ve E2Fb-DPA’yı serbest bırakmak için RBR’yi fosforile eder. Bu E2F-DP
kompleksleri sonuç olarak, şekil 1.8 a’da kara kutuyla işaretlenmiş hedef genlerin
promoter dizisinin içinde E2F bağlanma bölgelerine bağlanır ve bunların
transkripsiyonunu aktive eder (Komaki ve Sugimoto 2012).
Arabidopsis thaliana’da hücre döngüsünün transkripsiyonel aktivatörleri ve pozitif
regülatörleri olarak iş gören E2Fa ve E2Fb, memelilerdeki E2F1, E2F2 ve E2F3’e
benzerdir. Buna karşılık, E2Fc, hücre bölünmesinin transkripsiyonel represörü ve
supresörü olarak rol oynar.
E2F hedef genleri, promotorlarında E2F cis acting elementine sahiptir. E2Fa, E2Fb ve
E2Fc, DNA replikasyonu, hücre döngüsü ilerlemesini ve kromatin dinamiklerini içeren
genlerin ekspresyonunu ayarlayarak G1-S geçişinde çok önemli rollere sahiptir.
Şekil 1.7 Hücre döngüsünde transkripsiyonel kontrol (De Veylder vd. 2007)

20
MYB3R proteinleri, transkripsiyonal aktivatörlerdir ve onların spesifik hedef genlerinin
promotor bölgesindeki M-spesifik aktivatör (MSA) kutularına bağlanırlar. (Şekil 1.7b)
(De Veylder vd. 2007).
G2 ve M fazı boyunca, promotorlarında, M fazı spesifik aktivator (MSA) elementleri
içeren genler ifade edilir (Şekil 1.8b). G2 ve M fazında, CYCA2, CYCB1 ve CYCB2
genleri fazlaca ifade edilir ve bunların gen ürünleri ve CDKB1’ler ve CDKB2’ler ile bir
araya gelir. CYCD3; 1 geni M fazında da transkribe edilir ve gen ürünleri CDKA ile bir
araya gelir. MYB3R1 ve MYB3R4 transkripsiyon faktörleri, şekil 1.8 b’de kara kutuyla
işaretlenmiş hedef genlerin promotor dizisinin içinde MSA bölgesini tanır ve onların
transkripsiyonunu aktive eder. G1, S, G2 ve M fazında biraraya gelen genler, pembe,
sarı, mavi ve yeşil ile işaretlenmiştir. Hücre döngüsü boyunca ifade edilen genler, mor
ile işaretlenmiştir (Komaki ve Sugimoto 2012).
Şekil 1.8 Hücre döngüsü kontrolü (Komaki ve Sugimoto 2012)
a. G1’den S fazına geçiş, b. G2’den M fazına geçiş
G2-M spesifik gen transkripsiyonunun sürmesi için MSA kutuları gereklidir. Tütün
bitkisinde tanımlanan üç MYB3R proteini (MYB3RA1, MYB3RA2 ve MYB3RB),
hayvan c-Myb proteinleri ile yapısal olarak ilişkilidir. MYB3RA1 ve MYB3RA2 hücre
döngü fazı bağımlı transkripsiyon profilini gösterirken, MYB3RB düzeyleri hücre
döngüsü boyunca sabit kalır ve MYB3RB MSA içeren promotörleri aktive edemez.

21
MYB3RB ve diğer MYB3R proteinleri G2-M spesifik genlerinin ekspresyonunun aksi
yönünde kontrol ederler. MYB3RA2 tarafından, MSA içeren promotorların
transkripsiyonel aktivasyon derecesi, CDK için hedef olan C-terminal domaine ve hücre
döngüsü fazına bağlıdır. Bu, MYB3R proteinleri tarafından indüklenen siklinlerin,
CDK’lar ile kompleks oluşturması MYB3R aktivitesini üst düzeyde etkinleştiren geri
beslemeli mekanizmalar (feedback-loop mechanisms) anlamına gelir. Benzer şekilde,
siklinlerin proteoliz kontrolü, MYB3R proteinlerinin, hızlı inaktivasyonu ile
sonuçlanabilir. Her iki mekanizmada, hücre döngüsü boyunca, mitoza özgü siklinlerin
doruğa ulaştığının gözlenmesiyle sonuçlanabilir. Bu geri bildirim mekanizması MYB3R
transkripsiyon faktörlerinin promotorlarındaki MSA elementlerinin varlığı ile
güçlendirilir. MYB3RA1 ve MYB3RA2 kalıntılarının başlangıçtaki transkripsiyonel
aktivasyon nedeni bilinmemektedir (De Veylder vd. 2007).
1.4 MYC Ailesi
Myelocytomatosis ilişkili gen ailesi (MYC), bütün ökaryotlarda mevcut olan bir basic-
Helix-Loop-Helix (bHLH) domain içeren transkripsiyon faktörleri alt gen ailesini temsil
eder. N-terminal bölgesinde temel (basic) DNA bağlanma bölgesine sahip olmasıyla
karakterize edilir. C-terminal bölgesindeki hidrofobik kalıntılar proteinin dimerizasyon
kapasitesini belirleyen, bir döngü ile ayrılmış iki alfa heliks oluşturur. bHLH domain,
MYC’nin ait olduğu bHLH transkripsiyon faktörlerinin büyük ailesine karakteristiktir
(Toledo-Ortiz vd. 2003).
bHLH proteinleri, ağırlıklı olarak antosiyanin biyosentezi, fitokrom sinyalizasyonu,
tohum globulin ifadesi, meyve açılması, ve karpel ve epidermal gelişme işleviyle
karakterize edilmektedir (Buck ve Atchley 2003).
Myc hücre döngüsü sırasında G1 fazında ortaya çıkmaya başlar ve G1/S kontrol
noktasındaki çeşitli anahtar hücre döngüsü genlerini hedef alır. c-Myc, G1/S kontrol
noktası için uygun proteindir ve mitotik uyarı oluşmadığı durumlarda S fazındaki
hücreleri olumlu yönde uyarır (Eilers 1999).

22
Myc, sırasıyla Rb proteinini (pRb) fosforile eden, E2F aktivasyonuyla sonuçlanan,
Siklin E/Cdk2 kompleksin aktivasyonunu yönlendiren fonksiyonlara sahiptir. Bunun
yanında, Myc, Cdk aktivitesinden bağımsız olan E2F1, E2F2 ve E2F3 genlerinin
ekspresyonunu da aktive edebilmektedir. pRb ve E2F’nin indüksiyonu için Myc
bağlanma bölgeleri gerektiğinden Myc nin ortaya çıkardığı E2F2 ve E2F3 indüksiyonu
direk gerçekleşmektedir (Sears vd. 1997, Leone vd. 1997, Adams vd. 2000). İlginç bir
şekilde, E2F2 ve E2F3, Myc’nin S fazı girişini indüklemesi için gereklidir (Leone vd.
2001).

23
2. KAYNAK ÖZETLERİ
Bitkilerde büyüme, gelişme ve metabolizmayı etkileyen ya da engelleyen durumlara
stres adı verilmektedir. Stres faktörleri, orijinlerine göre abiyotik ve biyotik stres
faktörleri olmak üzere iki grupta incelenmektedir. Abiyotik stres faktörleri soğuk, sıcak,
kuraklık, tuzluluk, su fazlalığı, radyasyon, çesitli kimyasallar, oksidatif stres, rüzgar ve
toprakta besin yetersizliği gibi çevresel faktörlerdir. Biyotik stres faktörleri ise virüs,
bakteri ve fungusları içeren patojenler, böcekler ve herbivorlardır (Mahajan ve Tuteja
2005).
Abiyotik stres faktörleri özellikle ekonomik değeri olan bitkiler için en önemli
sınırlayıcı stres tipidir ve kuraklık bu stres tipinde en çok zarar veren faktördür (Bray
2007, Foolad 2007).
Ülkemizdeki su kaynakları ve tarım sektörü son yirmi yılda tekrarlanan kuraklık
dönemlerinden büyük zarar görmüştür (Sönmez vd. 2005). Özellikle bitkilerin büyüme
evrelerinde oldukça etkili olan kuraklık, önemli miktarda tarımsal ürün kayıplarına
neden olmaktadır (Bray 2007). Kuraklık ve tuzluluk stresi, dünya çapında ekilebilir
alanların %10 azalmasına ve % 50 ürün kaybına neden olmaktadır (Bray vd. 2000).
Stres etkenleri bitki gelişimini eş zamanlı olarak etkileyebilirken, bu stres etkenlerine
karşı tolerans ya da hassasiyet oluşumu anlaşılması güç olaylardır (Chinnusamy vd.
2004). Ayçiçeği bitkisinde çalışmada konu olan hücre döngüsü genleri Retinoblastoma
(Rb), Myelocytomatosis (Myc) ve E2F ‘dir.
E2F bir çeşit transkripsiyon faktörüdür. Abiyotik strese karşı tolerans oluşumunda
transkripriyon faktörlerinin rollerinin de olduğu tespit edilmiştir. E2F metabolik yolu
bitkilerde ve hayvanlarda korunmuş durumdadır (De Veylder vd. 2003, Dewitte ve
Murray 2003, Inze 2005).

24
Heterodimerik E2F-DP transkripriyon faktörleri DNA replikasyonu ve hücre döngüsü
için gerekli genlerin transkripsiyonunu düzenleyerek hücre döngüsünü kontrol ederler
(Helin 1998, Lavia ve Jansen-Dürr 1999).
pRb/E2F metabolik yolu bitkilerde de korunmuştur. S fazındaki spesifik genlerin
ekspresyonunu kontrol eden E2F transkripsiyon faktörleri G1 evresindeki genleri
susturmak için Rb proteine bağlı korepressör RBR ile birleşir. Geç G1 evresinde,
CYCD/CDKA, RBR’yi fosforile eder, E2F’yi serbest bırakır ve S-fazı genlerini aktive
eder (De Veylder vd. 2007).
G1’den S fazına doğru ilerleme genellikle, E2F-dimerizasyon ortağı (DP)-
retinoblastoma ilişkili (RBR) yolu ile kontrol edilir. Arabidopsis genomu üç tipik
E2F(E2Fa, E2Fb ve E2Fc) transkripsiyon faktörünü kodlar. Bu formlar, kendi
transkripsiyonel hedef genlerin promotorlarında spesifik E2F bölgelerine bağlanmak
için DPa veya DPb proteinleri ile dimerler oluştururlar. E2Fa ve E2Fb; DNA
replikasyonu, DNA tamiri ve kromatin korunması için gerekli genleri de içeren,
bunların muhtemel hedef genlerinde G1’den S fazına geçişi teşvik eden transkripsiyonel
aktivatörler olarak görev alırlar (Ramirez-Parra vd. 2003, Vlieghe vd. 2003, Vandepoele
vd. 2005, Naouar vd. 2009). G1 fazı boyunca, CYCD-CDKA kompleksleri RBR
fosforile eder ve böylece, G1’den S fazına geçişte gerekli genlerinin transkripsiyonunu
sağlamak için E2Fa–DPa ve E2Fb–DPa serbest bırakılır (Boniotti ve Gutierrez 2001).
Öte yandan, E2Fc-DPb dimerleri transkripsiyonel baskılayıcılar olarak fonksiyon
gösterirler (del Pozo vd. 2002, 2006, de Jager vd. 2009).
MYC transkripsiyon faktörleri, jasmonat (JA) sorumlu genlerin ifadesini düzenleyen,
yara direnç genlerini pozitif yönde düzenleyen ve patojen savunma genlerinin ifadesi
sırasında negatif düzenleyici olarak davranan, önemli transkripsiyonel düzenleyicileridir
(Van Verk 2009, Lorenzo 2004).
RNA pol-I transkripsiyonundaki rolünün yanında, c-Myc ribozomal DNA düzenleyici
bölgesindeki E-kutuları ile birleşir (Arabi vd. 2005). Ayrıca, Myc Pol III tarafından

25
transkripsiyonu yapılan genlere bağlanır ve aktivitesini indükler (Gomez-Roman vd.
2003).
Abe vd. (1997, 2003) Arabidopsis thaliana ile yaptıkları çalışmalarda dehidrasyon
cevap geni olan Rd22 geninin ekspesyonunda Myc ve Myb proteinlerinin
transkripsiyonel aktivatörler olarak görev aldıklarını rapor etmişlerdir.
Zeid (2001) Phaseolus vulgaris ‘te krom (Cr) uygulamasının α- ve β-amilaz aktivitesini
azaltarak embriyo ekseninin gelişimi için gerekli şekerlerin taşınmasını engellediğini ve
tohum çimlenmesini inhibe ettiğini, kromun düşük konsantrasyonlarının ise α-amilaz
aktivitesini arttırdığını rapor etmişlerdir.
Mariconti vd. (2002) Arabidopsis thaliana’daki E2F transkripsiyon faktörleri ailesini
konu alan çalışmalar yapmışlardır. Arabidopsis’te 6 tane fonksiyonel E2F geni
göstermişler, bunların hücre döngüsü işlemi sırasında farklı ekspresyon paternleri
gösterdiklerini öne sürmüşlerdir. 6 tane E2F grubunun, yapısal ve fonksiyonel
özelliklerine göre kendi içinde ikiye ayrılmış olduğunu, bunlardan ilk üçünün (AtE2Fa,
AtE2Fb ve AtE2Fc) diğer bitki ve hayvanlarda da korunmuş domainlere sahip
olduklarını rapor etmişlerdir.
Dewitte ve Murray (2003) tarafından, mısır bitkisinde yapılan çalışmalarda Rb proteini
sentezinden sorumlu iki gen bulunduğu bu genlerin bütün dokularda eksprese edildiği
fakat en güçlü ekspresyonun filiz uçlarında olduğu ortaya konmuştur. Gelişen mısır
yapraklarında Rb proteini fazlalığı tespit edildiğinde yaprak laminasında dereceli
uzamanın, Rb en yüksek düzeylere çıktığında arka yaprak sonunda hücre döngüsünün
uyarılarak hücre farklılaşmasının uyarıldığı, en düşük düzeylere indiğinde yaprak
tabanının proliferatif zonunun etkilendiği görülmüştür.
Ebel vd. (2004)’e göre Rb proteini yüksek bitkilerde bulunur ve korunmuştur.
Arabidopsis, sadece tek bir retinoblastoma-ilişkili gen (RBR) içerirken, hayvanlar, iki

26
akraba gene (p107 ve p130) sahiptir. Protein olarak, hem dişi ve hem de erkek gametofit
gelişimi için gereklidir.
Ramirez-Parra vd. (2004)’ye göre E2Ff/DEL3 proteini, hücre uzamasının kontrolü için
kendi promotoruna doğrudan bağlanarak, üç ekspansin geni (bitki hücre duvarında
bulunan ve büyüme ile ilgili birçok fonksiyona sahip genler) de içine alan birkaç hücre
duvarı biyosentez geninin ve UDP-glikoz-glikozil transferaz geninin ekspresyonunu
doğrudan baskılar.
West vd. (2004) yaptıkları çalışmada, tuz stresinin Arabidopsis’te birincil kök
büyümesinden sorumlu tepkiye aracılık eden hücre döngüsü düzenlemesinin önemi
üzerinde durmuşlardır. Tuz stresi, bitki büyüme ve gelişimini engellemiş, fideler artan
konsantrasyonlardaki NaCI ortamına transfer edildiğinde, kök büyüme oranı aşamalı
düşmüştür. Fidelerin 0.5% NaCl içeren büyüme ortamına aktarılmasının 3. gününde,
birincil kökleri transferden önceki sabit hızda büyürken, kontrol ortamına transfer
edilenlerin büyümeleri hızlanmıştır. Kinematik analizler, stresli köklerde büyüme
azalmasının, hücre üretiminde azalma ve daha küçük olgun hücre uzunluğuna bağlı
olduğunu ortaya koymuştur. Ortalama hücre döngüsü süresi etkilenmemiştir.
Dolayısıyla, bölünen hücre sayısının azalması nedeniyle hücre üretimi azalmıştır. 3 gün
önceki meristem boyutu adaptasyon mekanizmasını analiz etmek için, tuzlu ortama
transferi takip eden kısa vadeli hücre döngüsü olayları araştırılmıştır. Doğrudan siklin
bağımlı kinaz (CDK) aktivitesi ve CYCB1;2 promotor aktivitesi geçici olarak
azalmıştır. Çünkü CDKA;1 and CDKB1;1 her ikisinin de protein seviyeleri
etkilenmemiştir. Araştırmacılara göre, mitotik aktivitenin geçici inhibisyonu, CDK
aktivitesinin post translasyonel kontrolü aracılığıyla stres koşullarına uyumu olanak
sağlar. Bu nedenle tuz stresine adaptasyon iki aşama içerir. Birincisi meristemde daha
az hücrenin kalmasıyla sonuçlanan hücre döngüsünün geçici inhibisyonudur.
Meristemler verilen koşullar için uygun boyuta ulaştığında, hücre döngüsü süresi
varsayılana geri döner.
Santos (2004) Helianthus annuus L. var. SH222 fidelerinde, farklı tuz
konsantrasyonlarının (0–100 mM NaCl) klorofil içeriğindeki değişimini araştırdığı

27
çalışmada 21 gün süreyle 25 mM NaCl uygulamasının yalnızca klorofil b içeriğinde
önemli düzeyde bir artışa neden olduğu, 50 ve 100 mM NaCl uygulamalarının klorofil a
ve b içeriğinde önemli düzeyde azalmalara neden olduğunu rapor etmiştir. Araştırmacı,
50 ve 100 mM NaCl uygulamalarında klorofil sentezindeki azalmanın klorofilaz ilişkili
degradasyondan ziyade klorofillerin öncüsü olan ALA miktarındaki azalmadan
kaynaklandığını ileri sürmüştür.
Katerji vd. (2004), mısırda tuz ve kuraklık stresinin verim üzerinde oluşturduğu
etkilerini inceledikleri çalışmalarında, her iki stresin de verim ve yaprak su
potansiyelinde azalma meydana getirdiğini, verim üzerinde kuraklık ve tuzluluk
streslerinin benzer etki yaptıklarını bildirmişlerdir.
Park vd. (2005) Nicotiana benthamiana’da, Rb’nın hücre döngüsündeki düzenleme
rolünü virüs indüklü gen susturma ile inhibe ederek açıklamışlardır. Gen susturma
sonucu yaprak ve gövde hücrelerinde farklılaşmanın geciktiği, hücre proliferasyonunun
uzadığı görülmüştür. Çiçek oluşumunda da gecikmeler gözlenmiştir.
Vandepoele vd. (2005)’e göre hücre döngüsünün S fazına girişi, hücre döngüsü
ilerlemesi ve DNA replikasyonu için gerekli genlerin transkripsiyonunu teşvik eden,
E2F transkripsiyon faktörleri ile kontrol edilir. E2F yolu yüksek ökaryotlarda oldukça
korunmuştur. Yapılan çalışmada bitki E2F genlerini tanımlamak için mikroarray analizi
yapılmıştır. E2Fa ve DPa transkripsiyon faktörlerinin Arabidopsis thaliana fidelerinde
ektopik ifade genlerinin transkripsiyonel düzeyde promotor bölgelerini uyardığı tespit
edilmiştir. E2F hedef genlerinin G1 ve S fazları boyunca özel olarak ifade edildiğini ve
hareketsiz hücrelerin yeniden aktive edilerek hücre döngüsüne girdiğini göstermişlerdir.
Arabidopsis E2F hedef genleri için verilerin sağlamlığını test etmek için, ortolog pirinç
(Oryza sativa) genlerinin promotorlarında E2F cis etkili elementlerin varlığı da
araştırılmıştır. Bu yaklaşım kullanılarak, 70 potansiyel korunmuş bitki E2F hedef geni
belirlenmiştir. Bu genler, hücre döngüsü regülasyonu, DNA replikasyonu, kromatin
dinamikleri ile ilgili proteinleri kodlamaktadır. Ek olarak, potansiyel yeni S fazı
düzenleyici proteinler için birkaç gen tanımlanmıştır.

28
Magyar vd. (2005) oksine bağlı hücre bölünmesinin düzenlenmesinde E2FB
transkripsiyon faktörlerinin rolünü Arabidopsis bitkisinde araştırmışlardır.
Araştırmacılara göre E2FB’nin biriktirilmesi ve stabilitesi oksin tarafından pozitif
olarak düzenlenmektedir. E2FB, bitkiye özgü mitotik düzenleyicinin (CDKB1;1)
aktivasyonu ile M ve S fazlarına girişi düzenler. Artan E2F seviyeleri, kısa hücre döngü
süresine, hücre sayılarında artışa ve oldukça küçük hücre boyutlarına yol açmıştır.
Oksinin yokluğunda, hücrelerde poliploidi seviyesinde artışa eşlik eden uzama
görülmüştür. Ancak her ikisi de E2FB tarafından güçlü bir şekilde inhibe edilmiştir.
E2FB’nin hücrelerin çoğalmakta olduklarının mı yoksa hücre döngüsü, büyüme ve
DNA larının endoreduplikasyondan çıktıklarının mı belirlenmesinde oksin için bir
anahtar hedef olduğu sonucuna varmışlardır.
Kaya vd. (2006) ayçiçeği bitkisinin çimlenmesinde tuz ve kuraklığın etkisini
araştırdıkları çalışmada, tuzluluk ve kuraklık koşullarında çimlenmeye alınan ayçiçeği
tohumlarında çimlenmenin, her iki ortamda da geciktiğini ortaya koymuşlardır. Düşük
NaCl ortamında çimlenme ile kök ve gövde uzunluğunun daha fazla olmasına karşın,
çimlenme zamanının olumsuz etkilediğini belirtmişlerdir. Tohumların tüm NaCl
ortamlarında çimlendiğini, ancak -1.2 MPa PEG uygulamasında çimlenme olmadığını
gözlemlemişlerdir. Araştırıcılar, çimlenme süresinde ozmotik etkinin tuz toksisitesinden
daha etkili olabileceği sonucuna varmışlardır.
Lammens vd. (2008)’e göre, Arabidopsis’te üç atipik E2F, E2Fe/DEL1, E2Fd/DEL2 ve
E2Ff/DEL3, transkripsiyonel baskılayıcılar olarak işlev görürler. Ancak bunların
hiçbirinin bugüne kadar hücre döngüsü genlerinin sentezlenmesini doğrudan kontrol
ettiği gösterilmemiştir. Tüm bu atipik E2F’ler iki DNA bağlayıcı bölgeye sahiptir ve
monomerler olarak hedef promotorlara bağlanabilirler. E2Fe/DEL1 proteini, anafaz
teşvik edici kompleks/cyclosome (APC/C)’un aktivatörü olan CCS52A2 geninin
promotoruna bağlanır ve döngüye girişi baskılar.

29
Karuppanapandian ve Manoharan (2008) Cr(VI)’un fitotoksik etkilerinin kökler kadar
gövde dokusunda da oldukça belirgin olduğunu; Cr(III)’un ise sadece kök dokusunda
toksik olduğunu bildirmişlerdir.
Moussa ve Abdel-Aziz (2008), mısırda PEG solüsyonu ile oluşturdukları kuraklık
stresinde genotiplerin MDA düzeylerinin artış gösterdiğini, tolerant olan Giza 2
genotipinde ise MDA birikiminin daha düşük düzeyde görüldüğünü ifade etmişlerdir.
Day vd. (2008)’nin bazı çerezlik ayçiçeği çeşit ve genotiplerinin çimlenmesi üzerine
NaCl konsantrasyonlarının etkilerini araştırdıkları çalışmada, 10 farklı ayçiçeği
genotipine, 0, 5, 10 ve 20 dS/m konsantrasyonlarında NaCl stresi uygulamışlardır.
Ekimden itibaren 10. günde çimlenme yüzdesi (%), ortalama çimlenme zamanı (gün),
kök uzunluğu (cm), fide uzunluğu (cm), fide yaş ağırlığı (mg) ve fide kuru ağırlığı (mg)
ölçümleri yapılmıştır. Araştırma sonucunda, genotiplerin NaCl konsantrasyonlarına
farklı tepkiler gösterdiği belirlenmiştir. Artan NaCl seviyeleri çimlenme yüzdesinin
azalmasına, ortalama çimlenme zamanının uzamasına ve fide gelişiminin
engellenmesine neden olmuştur. Ayrıca tohumların kabuk oranının, tuz
konsantrasyonlarına bakılmaksızın, su alımı ve çimlenmede önemli bir etken olduğu
sonucuna varılmıştır.
Parvaiz ve Satyawaki (2008) yaptıkları çalışmada, arpa, ayçiçeği, çeltik gibi bitkilerin
tuza toleranslı genotiplerinin daha yüksek çözülebilir protein içeriğine sahip olduklarını
gözlemlemişlerdir.
Sozzani vd. (2010)’nin fonksiyon kazanımı ve fonksiyon kaybı çalışmalarına göre,
E2Fd/DEL2 proteini, hücre çoğalmasının kontrolünden sorumludur, ancak çalışmada
doğrudan hedef genler tespit edilememiştir.
Borghi vd. (2010) post embriyonik organ gelişiminin başlatılmasının ve devam
etmesinin düzenlenmesinde görev alan birçok genin varlığından söz etmişlerdir. Tümör
repressör retinoblastoma’nın bitki homoloğu olan retinoblastoma’nın hücre döngüsünün

30
anahtar düzenleyicisi olduğu ortaya koymuşlardır. Arabidopsis thaliana’da hücre
döngüsünde RNAi ile farklı gelişme evrelerinde retinoblastomanın ekspresyon
seviyelerinin azaldığını ve meristem aktivitesinin kökte, gövdede değiştiğini tespit
etmişlerdir. Bu çalışmada retinoblastomanın hücre döngüsü, proliferasyonu ve
gelişimindeki önemli rolleri gösterilmiştir.
Hichri vd. (2010) asma bitkisinde Myc geninin flavanoid biyosentez yolunun
düzenlenmesini konu alan çalışmalarında, Myc1 geninin flavanoid metabolik yolu
genlerinin promotörüne bağlanarak kontrolü sağladığını ortaya koymuşlardır.
Costas vd. (2011) bitkilerde hücre proliferasyonunda epigenetik modifikasyonları
tartışmış, hücre döngüsü süreçlerini incelemiş, hücre sinyalleri dışında transkripsiyonal
faktörlerin hücre döngüsünün yönetiminde önemli rol aldıklarını belirtmişlerdir.
Radziejwaski vd. (2011) E2F transkripsiyon faktörleri ile ilgili yaptıkları çalışmalarda,
bitkilerin zor çevre şartlarına uyabilecek biçimde evrimleştikleri ortaya koymuşlardır.
Strese uyum sağlamada dikkati çeken en önemli olaylardan birinin optimum gelişmeye
ulaşmak için hücre döngüsündeki yeniden programlanmalar olduğunu ve E2F
transkripsiyon faktörlerinin en önemli düzenleyiciler olduğunu savunmuşlardır.
Dudits vd. (2011) bitkilerdeki hayat döngüsünün hücre bölünmesi ve hücre genişlemesi
gibi gelişimsel olaylara dayalı olduğunu belirtmişlerdir. Hücresel döngüler, çevresel ve
hormonal faktörlerin etkisi altındadır. Aynı zamanda hormonal ve gelişimsel kontrol ile
hücre döngüsü işlemi arasında bağlantı sağlayan moleküler olaylar sınırlıdır.
Çalışmalar, organ gelişiminde hücre bölünmesi parametrelerinden, hormonal sinyal
ağları ile organ gelişimindeki hücre döngüsü genleri proteinlerinden etkilenen birçok
alternatif yol olduğunu göstermiştir. Araştırmacılar, hücre döngüsü işleyişinde
RBR’lerin ana rolünün fosforilasyona bağlı olduğunu ve RBR ve fosfo-RBR
proteinlerinin interfaz ve mitotik evrelerde var olduğunu ortaya koymuşlardır.

31
Kurzbauer ve Schlögelhofer (2011) Arabidopsis bitkisinde Rb’nın mayozdaki rolünü
çalışmışlardır. Fosforile olmuş Rb’nın kromozom yoğunlaşmasında, sentromerik
fonksiyonlarda ve kromozom stabilitesinde önemli rolleri olduğunu, ayrıca bu genin
yüksek bitkilerde korunduğunu ifade etmişlerdir.
Polit vd. (2012) Vicia faba’ nın kök meristem hücrelerinde Rb tipi proteinlerin hücre
döngüsüne bağlı değişimlerini incelenmişlerdir. Bu proteinin fosforilasyonundaki artışın
G1-S faz geçişiyle korelasyon halinde olduğu bulmuşlardır.
Hem bHLH hem de ZIP motiflerini içeren Myc genleri, Absisik asit (ABA)-duyarlı
genlerin düzenlenmesinde önemli rol oynayan transkripsiyon faktörleri grubudur. Ji vd.
(2012)’de yaptıkları çalışmada, NaCl, osmotik ve ABA stres koşulları altında MYC
genlerinin rolünü araştırmak için Tamarix hispida’dan dokuz MYC geni
klonlamışlardır. (RT)-PCR analizi kök, gövde ve yaprak dokularında dokuz MYC
geninin de ifade edildiğini, ancak bu genlerin ifade düzeylerinin çeşitli dokularda farklı
olduğunu göstermiştir. ABA, NaCl ve ozmotik streslerinin 3 saat uygulanmasından
sonra, Myc genleri köklerde yüksek derecede uyarılmıştır. Bununla birlikte bu streslerin
6 saat süreyle uygulanmasıyla gövde ve yaprak dokularında MYC genleri yüksek
derecede uyarılmıştır. Buna ek olarak, bu MYC genlerin çoğu en çok yapraklarda ve
gövdeleri ile karşılaştırıldığında köklerinde yüksek derecede ifade edilmiştir. Ayrıca,
MYC genleri köklerde, yaprak ve gövde dokularından daha çok uyarılmıştır. Bu
durum, bu genlerin stres cevabında gövde ve yapraklardan ziyade köklerde rol
oynayabileceğini göstermektedir. Bu çalışmanın sonuçları MYC lerin tuz ve ozmotik
stres toleransında rol oynadığını ve ABA sinyal transdüksiyon yolu tarafından kontrol
edildiğini göstermektedir.
Jasmonatlar (JAS) bitki büyümesi ve biyotik ve abiyotik streslere tepkileri arasındaki
dengeyi düzenleyen bitki hormonlarıdır. Son çalışmalar, Arabidopsis thaliana’da JA
kaynaklı yanıtlar için mekanizmaları ortaya çıkarmasına rağmen, hangi mekanizmaların
JA kaynaklı yanıtları azalttığı belirsizdir. Nakata vd. (2013) yaptıkları çalışmada, basic
helix-loophelix-tipi transkripsiyon faktörü, ABA-İNDUCİBLE bHLH-tipi transkripsyon

32
faktörü / JA-ASSOCIATED MYC2-LIKE1 (JAM1), transkripsiyonel baskılayıcılar
gibi davrandıkları ve JA sinyalizasyonunu olumsuz düzenlediklerini rapor etmişlerdir.
Fonksiyon kazanmış transgenik bitkiler, JA-kaynaklı kök büyüme inhibisyonu,
antosiyanin birikimi ve erkek fertilitesini içeren JA yanıtlarında önemli bir azalma
sergileyen JAM1 için kimerik represörü sentezler. Bu bitkiler aynı zamanda, böcek
otçul (herbivor) Spodoptera exigua tarafından saldırıya karşı direnmede tehlikededir.
Bunun aksine, jam1 fonksiyon kayıp mutantları böcek saldırısına karşı arttırılmış
dayanıklılığı da içeren, geliştirilmiş JA yanıt göstermiştir. JAM1 ve MYC2, MYC2’nin
hedef sekansına rekabetçi bağlanması, JAM1 tarafından MYC2 fonksiyonlarının
baskılanması ve JA sinyalizasyonunun negatif düzenlenmesi için mekanizma sağlıyor
olabilir. Bu sonuçlar JAM1’in JA sinyalini olumsuz düzenlediğini, böylece JA-aracılı
stres yanıtlarının ince ayarında ve bitki büyümesinde önemli bir rol oynadığını
göstermektedir.
Kaya ve Yiğit (2014) flurokloridona maruz kalmış ayçiçeği üzerinde salisilik asitin
fizyolojik ve biyokimyasal etkilerini inceledikleri çalışmalarında MDA içeriğindeki
değişimini araştırmışlardır. MDA içeriğinde Salisilik asit (SA) uygulaması ne olursa
olsun kontrol grubuna göre artış gözlenmiştir. En yüksek MDA düzeyi 72 mM salisilik
asit uygulanan grupta görülmüştür.

33
3. MATERYAL VE YÖNTEM
3.1 Bitki Materyalinin Temin Edilmesi
Trakya Tarımsal Araştırma Enstitüsü’nden 08-TR-003, TR-3080, SARAY, TARSAN
1018 olmak üzere 4 farklı ayçiçeği (Helianthus annuus L.) çeşidi temin edilmiştir.
08-TR-003, TR-3080, SARAY, TARSAN 1018 çeşitlerine ait tohumlar yüzey
sterilizasyonunun ardından, plastik kaplardaki perlit içine ekilmiştir. Her gün Hoagland
besin solüsyonu (Hoagland ve Arnon 1938) ile sulanarak, SANYO MLR-351H bitki
büyütme kabininde, 16 saat gündüz, 8 saat gece fotoperiyodunda, 125 µE/ m2.S ışıkta,
22 OC’de on beş gün süreyle geliştirilmiştir (Şekil 3.1).
Herbir ayçiçeği çeşidi, üç ayrı kaba ekilerek üç biyolojik tekrar yapılmıştır (Şekil 3.2).
Şekil 3.1 SANYO MLR-351H bitki büyütme kabininde bitkilerin geliştirilmesi
a. Kabin, b, c. Bitkilerin kabinde gelişim aşamaları

34
3.2 Hoagland Besi Ortamının Hazırlanması
Hoagland besi ortamını hazırlamak için, 10 L 10X makro besin çözeltisi içine 50 ml
mikro besin çözeltisi eklendikten sonra hazırlanan bu çözelti 1/10 seyreltilmiştir. Elde
edilen 1L 1X konsantrasyonundaki çözeltiye 1 ml 1000X FeEDTA çözeltisi
kullanımdan hemen önce eklenerek kontrol çalışma ortamı olarak kullanılmıştır. 1000X
FeEDTA; 365 g FeEDTA 1 L dH2O içerisinde çözdürülerek hazırlanmıştır.
Hoagland besi ortamı içeriği aşağıdaki gibidir:
Çizelge 3.1 Hoagland besi ortamı makro besin çözeltisi içeriği
Makro besin çözeltisi
K2SO4 15.7 g
2 L saf su
içerisinde
çözülür
KH2PO4 2.7 g
MgSO4.7H2O 24 g
Ca (NO3)2. 4H2O 47.23 g
KCl 0.0746 g
Çizelge 3.2 Hoagland besi ortamı mikro besin çözeltisi içeriği
Mikro besin çözeltisi
H3BO3. 0.124 g
2 L saf su
içerisinde
çözülür
MnSO4 0.066 g
CuSO4.5H2O 0.100 g
NH4Mo 0.048 g
ZnSO4.7H2O 0.1553 g
Çizelge 3.3 Hoagland besi ortamında bulunan iyonların son konsantrasyonları
Çözelti içerisinde iyonların son konsantrasyonları:
Ca 2 mM Mn 10-6
M
NO3 4 mM Cu2+
10-7
M
Mg 1 mM NH4 10-8
M
K 2 mM Zn 10-6
M
P 0.2 mM Fe 10-4
M
B 10-6
M

35
Şekil 3.2 Hergün Hogland besin solüsyonu ile sulanarak on beş gün geliştirilen
ayçiçeği çeşitleri

36
3.3 Bitki Stres Koşulları
3.3.1 100 mM ve 150 mM tuz (NaCl) stresi
1/10 Hoagland besi ortamına NaCl son konsantrasyonu 100 mM ve 150 mM olacak
şekilde NaCl eklenmiştir. 15 günlük bitkiler, her kap içerisinde 5 fide olacak şekilde
100 mM ve 150 mM NaCl içeren kapların içerisine alınmış, SANYO MLR-351H bitki
büyütme kabininde tutularak 0., 3. ve 27. saatlerde örneklem alımı gerçekleştirilmiştir
(Şekil 3.3). Her iki farklı konsantrasyondaki NaCl stresi bitkilere her çeşit için 3 farklı
kapta toplam 15 örnek olacak şekilde uygulanmıştır. Stres uygulama süreleri sonunda
örnekler alınarak RNA izolasyonuna kadar -80 derecede dondurucuda muhafaza
edilmiştir.
3.3.2 100 mM ve 150 mM kuraklık (PEG) stresi
1/10 Hoagland besi ortamına PEG 6000 son konsantrasyonu 100 mM ve 150 mM
olacak şekilde PEG 6000 eklenmiştir. 15 günlük bitkiler, her kap içerisinde 5 fide
olacak şekilde 100 mM ve 150 mM PEG içeren kapların içerisine alınmış, SANYO
MLR-351H bitki büyütme kabininde tutularak 0., 3. ve 27. saatlerde örneklem alımı
gerçekleştirilmiştir (Şekil 3.3). Her iki farklı konsantrasyondaki PEG stresi bitkilere her
çeşit için 3 farklı kapta toplam 15 örnek olacak şekilde uygulanmıştır. Stres uygulama
süreleri sonunda örnekler alınarak RNA izolasyonuna kadar -80 derece dondurucuda
muhafaza edilmiştir.

37
Şekil 3.3 SANYO MLR-351H bitki büyütme kabininde yetiştirilen bitkilere Hoagland
besi ortamı içerisinde tuz (NaCl) ve kuraklık (PEG) stresinin uygulanması

38
3.3.3 Cr(VI) stresi
Hoagland besi ortamı içerisinde yetiştirilmiş 15 günlük ayçiçeği fideleri stres
uygulaması amacıyla sırasıyla Hoagland, 80, 160, 320, 640 ve 1280 µM Cr(VI)çözeltisi
içeren, her bir derişimden üçer tane olmak üzere kaplara alınmış ve 24 saat SANYO
MLR-351H bitki büyütme kabininde tutulmaya devam edilmiştir (Şekil 4, 5). Farklı
derişimlerdeki Cr(VI) stresinden 24 saatin sonunda örneklem alımı gerçekleştirilmiştir
Alınan örnekler RNA izolasyonu aşamasına kadar saklanmak üzere -80oC’de muhafaza
edilmiştir.
Şekil 3.4 Farklı derişimlerde hazırlanan Cr(VI) çözeltileri

39
Şekil 3.5 SANYO MLR-351H bitki büyütme kabininde yetiştirilen bitkilere Hoagland
besi ortamı içerisinde Cr(VI) stresinin uygulanması

40
3.4 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)
Lipid peroksidasyon olayı, hücre zarı fosfolipitlerinin doymamış yağ asitlerinde
meydana gelen tepkimelerdir. Bu olayın temel mekanizması, oluşan toksik aldehitlerin
proteinler ve protein yapısında olmayan yapılarla etkileşmesi ve hücre zarlarına yönelik
istenmeyen etkiler meydana getirmesidir (Yarsan 1998).
Oksidatif stres sonucu oluşan serbest radikallere bağlı olarak hücre zarındaki lipidler
peroksidasyona uğramakta ve bunun son ürünü olarak malondialdehid (MDA) ortaya
çıkmaktadır. Oluşan MDA, hücre membranlarında gerçekleşen iyon alışverişi sırasında
membrandaki bileşiklerin çapraz bağlanmasına, hücre içi ve dışındaki protein, nükleik
asit gibi birçok biyomoleküle etki ederek geri dönüşümü mümkün olmayan hasarlara
neden olmaktadır (Rio vd. 2005). Plazma MDA düzeyinin belirlenebilmesi dokulardaki
lipid peroksidasyonunun ve dolayısıyla oksidatif stresin hassas göstergelerinden biridir
(Knight vd. 1988).
Bitkide stres oluşumunu göstermek amacıyla malondialdehit (MDA) analizi ile lipit
peroksidasyonu belirlenmiştir (Hodges vd. 1999).
1. 100 mg yaprak örneği %80’lik 1 ml alkol ile homojenize edilir. 3000 g de +4 oC’de
10 dk santrifüj edilir.
2. Santrifüj sonrası elde edilen süpernatant ikiye bölünür;
a. 1 hacim alınır + 1 hacim %20’lik TCA (trichloroaceticacid) + 1 hacim
%0,01’lik BHT (butylated hydroxytoluene) eklenir. (-TBA)
b. 1 hacim alınır + 1 hacim %0,65 TBA (2-thiobarbituric acid) içeren %20’lik
TCA + 1 hacim %0,01’lik BHT eklenir. (+TBA)
3. a ve b şeklinde ayrılmış ve yukarıdaki şekilde hazırlanmış olan tüpler 95 oC’de 25
dk inkübasyon sonrası ani bir şekilde soğutma amacıyla buza alınır.
4. Soğuyan örnek 3000 g’de 10 dk santrifüj edilir ve süpernatant alınır.
5. Birinci aşamada 532-600 nm’de, ikinci aşamada ise 532-600-440 nm’de ölçülür.

41
Spektrofotometrede okunan değerler ile yüzde MDA seviyelerinin hesaplanmasında
aşağıdaki formüller kullanılacaktır.
ABS=Absorbans
MDA=Malondialdehit
1- [(ABS 532+TBA)-(ABS 600+TBA)-(ABS 532-TBA)-(ABS 600-TBA)]= A
2- [(ABS 440+TBA)-(ABS 600+TBA)x0,0971]=B
3- nmolMDA/ml= (A-B)/157000x106
3.5 Total Protein Analizi
Cr, tuz ve kuraklık stresi uygulanmış ayçiçeği bitkilerinde Bradford metoduna
(Bradford 1976) göre total protein analizi yapılmıştır.
1) 0.08 g bitki örneği, kum yardımıyla havanda ezilir.
2) 700 µl fosfat tamponu (0.2 M, pH 7.0) ile homojenize edilir.
3) 14.000 rpm’de 20 dk. santrifüj edilerek süpernatantlar yeni tüplere alınır.
4) 2000 μl 1X Bradford çözeltisi ve 200 μl örnek çözeltisi ~10 saniye vortekslenerek
karıştırıldıktan sonra 10 dakika oda sıcaklığında bekletilir. Kör olarak 200 µl PB ve
2000 μl 1X Bradford karışımı kullanılır.
5) Absorbans değerleri 595 nm dalga boyunda SpectraMax M5 spektrofotometrede
ölçümler her örnek için 4 tekrarlı olarak gerçekleştirilmiştir.
Örneklerin Toplam Çözünür Protein Konsantrasyonunun Hesaplanması:

42
3.6 Total RNA İzolasyonu
Cr, tuz ve kuraklık stresi uygulanmış ayçiçeği bitkilerinden total RNA izolasyonu
TRIGent protokolüne göre yapılmış ve RNA miktarı spektrofotometrik ölçümlerle
belirlenmiş ve %1,2’lik agaroz jelde yürütülerek teyit edilmiştir.
3.6.1 TRIGent RNA İzolasyon Protokolü
1. Her 50 ~ 100 mg doku başına 1 ml TRIGent eklenir.
2. Örnekler 2 ila 8 °C sıcaklıkta 10 dakika boyunca 12,000´g santrifüj edilir.
3. Yağlı dokularda üstte toplanan tabaka uzaklaştırılmalıdır. Temiz süpernatan yeni
bir tüpe alınır.
4. Örnekler 15 ~ 30 °C ‘de 5 dakika inkübe edilir.
5. Her 1 ml TRIGent için 0.2 mL kloroform eklenir. Tüpler 15 saniye boyunca elle
kuvvetli bir şekilde çalkalanır ve 2 ila 3 dakika boyunca 15 ~ 30 °C’de inkübe
edilir.
6. Örnekler 2 ~ 8 °C sıcaklıkta 15 dakika boyunca 12,000´g santrifüj edilir.
7. Süpernatan yeni bir tübe alınır.
8. Her 1 ml TRIGent için 0.5ml izopropil alkol eklenir.
9. Örnekler 15 ~ 30 °C’ de 10 dakika inkübe edilir.
10. Örnekler 2 ~ 8 °C sıcaklıkta 15 dakika 12,000´g santrifüj edilir.
11. Her 1 ml TRIGent için 1 ml %75 lik etanol eklenir.
12. Örnekler vortekslenir ve 2 ~ 8 °C sıcaklıkta 7500 g’de 5 dakika santrifüj edilir.
13. Üst sıvı dökülür ve 5-10 dakika oda sıcaklığında kurumaya bırakılır.
14. RNA’nın çözülmesi için 50 µl DEPC ile muamele edilmiş su eklenir.
15. 10 dakika 55-60 °C’de inkübe edilir.
16. -80 °C’ de saklanır.

43
3.7 DNaz Uygulaması
Thermo Scientific EN0521 kodlu DNase enzimi kullanılarak izole edilen tüm RNA
örneklerinde olması muhtemel DNA kontaminasyonu minumum düzeye getirilmiştir.
1. Aşağıda yer alan bileşenler RNase free tüpe eklenir.
Bileşenler Kullanılan miktar
RNA 1 µg
10 X Reaksiyon tamponu (MgCl2 ‘lü) 1 µL
DNase I RNase free 1 µL (1 U)
DEPC ile muamele edilmiş ultra saf su ile toplam hacim 10 µl’ye ayarlanır.
2. 37 oC’de 30 dakika inkübe edilir.
3. 1 µl 50 mM EDTA eklenir.
4. 65 oC’de 10 dakika inkübe edilerek DNase inhibe edilir.
3.8 cDNA (Komplementer DNA) Sentezi
cDNA sentezi, RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific)
kullanılarak gerçekleştirilmiştir. Retinoblastoma(Rb), E2F, Myelocytomatosis (MYC)
ve Actin (ACT) gen bölgeleri anchored-oligo(dT)18 ile çalışılmıştır. Çizelge 3.4’de
cDNA sentezinin ilk aşamasında hazırlanan total RNA – primer karışımı içerisinde
bulunan bileşenler ve final konsantrasyonları verilmektedir.
1. Çizelge 3.4’te yer alan bileşenler buz üzerinde tutulan nükleaz free tüpe sırasıyla
eklenir.
Çizelge 3.4 Total RNA - primer karışımı
Komponent Final Konsantrasyon
Total RNA 0.1 ng – 5 µg total RNA
Anchored-oligo(dT)18 Primer,
50 pmol/µl
1 µL
Saf su Toplam hacim 12 µl’ye ayarlanır

44
2. Total RNA – primer karışımı ısı döngü cihazında, 65 oC’de 5 dakika bekletilerek
RNA’nın ikincil yapısının denatürasyonu sağlanır.
3. Ters transkrpsiyon reaksiyonunun (RT-PCR) gerçekleştirilmesi için çizelge 3.5 ‘de
yer alan komponentler sırasıyla eklenir.
Çizelge 3.5 Ters Transkrpsiyon (RT-PCR) Reaksiyonu Komponentleri
Komponent Miktar
5x Reaksiyon tamponu 4 µL
RiboLock RNase İnhibitör (20 U/µL) 1 µL
10 mM dNTP karışımı 2 µL
RevertAid H Minus M- MuL V Ters Transkriptaz (200 U/ µL) 1 µL
Toplam hacim 20 µL
4. Pipetle hafifçe karıştırılıp, santrifüj edilir.
5. Ters transkripsiyon reaksiyonu çizelge 3.6’da belirtilen programda, ısı döngü
cihazında gerçekleştirilir.
Çizelge 3.6 Ters transkripsiyon reaksiyonunun gerçekleştirildiği program
Primer Hedef
mRNA
RT Reaksiyonu İnkübasyon
Süresi
Oligo(dT)18 primer 4 kb a kadar 42 oC’de 60 dk
70 oC’de 5 dk
6. Elde edilen cDNA’lar Real time PCR reaksiyonu gerçekleştirilene kadar -80 oC’de
muhafaza edilmiştir.

45
3.9 Primer Dizaynı
Çalışmada housekeeping gen olarak kullanılan Actin(ACT) ve MYC2 gen bölgerine ait
primerlerin sentezi, gen bankasında (NCBI) yer alan ayçiçeği (Helianthus annuus L.)
bitkisine ait diziler kullanılarak gerçekleştirilmiştir. Ayçiçeği genomunda E2F ve
Retinoblastoma (Rb) genlerine ait diziler gen bankasında (NCBI) yer almadığından, bu
bölgeler için primerler model organizma olan Arabidopsis thaliana üzerinden
sentezlenmiştir. Çok sayıda primer sentezlenmiş olup en uygun primerin tercih
edilebilmesi için erime eğrisi (Melting curve) analizi yapılmış ve primer dimer oluşumu
ve özgül olmayan amplifikasyon ürünleri gibi hatalar gözlemlenmiştir. Bu analiz
sonuçları değerlendirilerek çalışma için en uygun primerler seçilmiş ve çizelge 3.7’de
dizisi verilen primerlerden * işaretli olanlar çalışmada kullanılmıştır.
Çalışmada kullanılan primerler için, Retinoblastoma ilişkili protein (RBR) genine ait
3561 baz uzunluğundaki, NM_112064.4 GenBank numaralı dizi, E2F genine ait 1643
uzunluğundaki AJ271597.1 GenBank numaralı dizi, MYC2 genine ait 465 baz
uzunluğundaki HQ428989.1 GenBank numaralı dizi ve Actin (ACT) genine ait 1134
baz uzunluğundaki FJ487620.1 GenBank numaralı dizi belirlenmiş ve bu dizilere uyan
primer dizileri tasarlanmıştır.

46
Çizelge 3.7 Çalışmada kullanılan gen bölgeleri için tasarlanan primer dizileri
Primer Sekans (5’→3’) Primer
uzunluğu (bç)
Bağlanma
Sıcaklığı (Tm)
*E2F-F ACCTCCGTTTCATCCTTCCG 20 60.5 oC
*E2F-R ACAAGTAGCAATCGAAATCCCC 22 60.3 oC
E2FB-F CAGGTTGTCTCTGACGCGAT 20 57.1 oC
E2FB-R GTTCCATGCGGTGCCTTTAC 20 56.9 oC
E2F-F CACCATCCACCGTCATCTCC 20 57.7 oC
E2F-R TACGTCGTTTCTGCACCTCC 20 57.1 oC
E2FA-F ACAGCTGCAGGCAGAAATTG 20 56.3 oC
E2FA-R TGGAGTACTCACGTCGGCTA 20 57.3 oC
*RBR-F AGAAGCGGATGGTCTGTC 18 56.1 oC
*RBR-R CCTCCTTTGCCTGAAGTCTG 20 60.5 oC
RBR1-F GGATGCGTTACAATGTCGCC 20 56.8 oC
RBR1-R GTTGCTGCTGCTGTTGTTCA 20 56.7 oC
*Myc2 F GACAAGGCGTCACTCTTAGG 20 60.5 oC
*Myc2 R TCCCAGCCAATGACTTTCAC 20 58.4 oC
MYC2-F GGAAGCGGAGAGACAAAGAA 20 54.6 oC
MYC2-R CGCATGGTGCACTTCTAAATC 21 54.4 oC
*AKTİN-F CAATGTTCCCGCCATGTAT 19 53 oC
*AKTİN-R CGACCACTGGCATAAAGAGAA 21 54.8 oC
*Çalışmada kullanılan primerler
(F: Forward/İleri, R: Reverse/Geri)
3.10 Real-time PCR Uygulaması
Real-Time PCR uygulaması ABI 7500 Fast Real Time cihazı aracılığıyla SYBR Green I
Master boyası kullanılarak gerçekleştirilmiştir. Çalışılması hedeflenen genlere ait diziler
NCBI veritabanında araştırılıp çıkarılmış ve bölgeye özgü primerler tasarlanarak
sentezlettirilmiştir. Ayrıca ekspresyon çalışmaları için gerekli şartlardan birisi olan
normalizasyon işleminde bir housekeeping gen olan Actin (ACT) geni kullanılmıştır.

47
Real-Time PCR yöntemi ile kantitasyon (miktar belirleme = gen ekpresyonu miktarı
belirleme) çalışması SYBR Green I boyası kullanılarak gerçekleştirilmiş ve
kantitasyonu takiben PCR’ın etkinliğini saptamak ve herhangi bir dimer oluşumu olup
olmadığını gözlemlemek amacıyla Erime Eğrisi (Melting curve) Analizi yapılmıştır.
Kontrol grubu ile E2F, Myc ve Retinoblastoma (Rb) gen bölgesi için tasarlanan bütün
primerlerin, Real time PCR reaksiyonları denenerek optimizasyonu sağlanmıştır. En iyi
sonucu veren E2F, Myc2 ve RBR primeleri ile çoğaltma işlemi gerçekleştirilmiştir.
Kontrol örnekleri ile gerçekleştirilen primer optimizasyonu çalışmaları sonrasında bütün
örneklerin Real time PCR reaksiyonu yapılmıştır. Stres uygulamaları sonrasında seçilen
örneklerin real time PCR reaksiyonları üç tekrarlı (teknik tekrar) olacak şekilde
gerçekleştirilmiştir.
Real Time PCR reaksiyonu çizelge 3.8’de verilen komponentler ile hazırlanmış olup saf
su ile toplam hacim 20 µl’ye ayarlanarak ABI 7500 Fast Real-Time cihazında çizelge
3.9’da verilen programda çoğaltılmıştır.
Çizelge 3.8 Real Time PCR reaksiyonu komponentleri
Komponent Son Konsantrasyon
cDNA 2 µg
İleri (Forward) Primer 10 µM
Geri (Reverse) Primer 10 µM
SYBR® Green Master Mix 1X
Toplam hacim 20 µl’dir.

48
Çizelge 3.9 Real Time PCR reaksiyonunun gerçekleştiği program
Primer Sıcaklık Süre Döngü sayısı
Ön denatürasyon 95 oC 7 dk 1
Denatürasyon 95 oC 10 s
40
Bağlanma 58-60 oC 30 s
Erime
Eğrisi Analizi
58-95 oC
Sürekli okuma 1
ACT, E2F, RBR ve MYC2 genlerinin tuz ve kuraklık stresi (100-150 mM) ve Cr(VI)
uygulamalarında farklı zamanlarda alınmış örneklerin Real time PCR reaksiyonu ABI
7500 Fast Real Time cihazında, Relative Standard Curve’de gerçekleştirilmiş ve gerçek
zamanlı olarak izlenerek pik profilleri kaydedilmiştir. Cycle Treshold (Ct) değeri
polimeraz zincir reaksiyonuna ait pik profilinde logaritmik artışa geçilen ilk noktayı
belirtmektedir. Real time PCR reaksiyonları sonrasında elde edilen pik profillerinden
her bir örneğe ait Ct değerleri belirlenmiştir.
3.11 Normalizasyon ve İstatistiksel Hesaplamalar
Herbir çeşide ait bitkiler on beş gün süreyle 3 bağımsız kapta yetiştirilmiştir. Stres
uygulamalarında örnekler her kapta 5 örnek olacak şekilde 3 ayrı kaba alınmıştır.
Herbir stres için 15 bitki stres uygulama süreleri sonunda alınarak bir havuz
oluşturulmuş, daha sonra her bir parametre için seçilen 1’er örnekten RNA izolasyonu
yapılmıştır. Seçilen bu örneklerden de 3’er tekrarlı (teknik tekrar) olacak şekilde real
time PCR reaksiyonları gerçekleştirilmiştir.
Farklı süre ve miktarda tuz, kuraklık ve Cr(VI) stresi uygulanmış bitkilere ait
transkript profilleri kontrol profilleri ve housekeeping gen olarak seçilen aktin ile
karşılaştırılmıştır. Elde edilen verilerin istatistiksel olarak değerlendirilmesi Livak ve
Schmittgen’in 2-ΔΔCT
metoduna göre ve One Way ANOVA (Dunnett) ile yapılmıştır
(Livak ve Schmittgen 2001). Araştırmacıların bir kontrol grubunu birden fazla deney

49
grubu ile karşılaştırması gerektiğinde Dunnet testini kullanmaları önerilmektedir
(Dunnet 1955). Post-Hoc testi ise (Multiple Comparisons Dependent Variable:
Expression Dunnet T3) eğer varyans analizi sonucunda gruplar arasında bir fark
bulunmuşsa, farklılığın hangi gruplardan kaynaklandığını tespit etmek için
kullanılmaktadır (Roscoe 1975).
Konsantrasyon ve Ct değeri olarak belirlenen gen ekpresyonu sonuçları housekeeping
gen olarak çalışmada kullanılan ACT (aktin) ve kontrol şartları dikkate alınarak
normalize edilmiştir (Livak ve Schmittgen 2001). Bu verilerin ortalama, standart
sapma, standart hata ve istatistiksel olarak anlamlılık dereceleri istatistik programı
(SPSS 17) ile hesaplanmıştır. Bu şekilde tuz (NaCl), kuraklık (PEG) ve Cr(VI) stresi
uygulanmış örneklerde RBR, E2F ve MYC2 geni mRNA ifade seviyeleri tespit
edilmeye çalışılmıştır.

50
4. BULGULAR
4.1 Ayçiçeği Bitkisinde Stres Uygulamaları
4.1.1 Tuz stresi
On beş gün süreyle geliştirilen bitkilere, NaCl (100, 150 mM) aracılığı ile tuz stresi
uygulanmıştır. NaCl stresi 27 saat süre ile uygulanmış ve her bir farklı molaritedeki
NaCl stresinden, her bir çeşit ayçiçeği için 0., 3., 27. saatlerde örneklem alımı
gerçekleştirilmiştir (Şekil 4.1-4.4).
4.1.2 Kuraklık stresi
On beş gün süreyle geliştirilen bitkilere, Polietilen Glikol (PEG) (100, 150 mM)
aracılığı ile kuraklık stresi uygulanmıştır. PEG stresi 27 saat süre ile uygulanmış ve
her bir farklı molaritedeki PEG stresinden, her bir çeşit ayçiçeği için 0., 3., 27.
saatlerde örneklem alımı gerçekleştirilmiştir (Şekil 4.5-4.8).
4.1.3 Cr(VI) stresi
15 günlük ayçiçeği fideleri stres uygulaması amacıyla sırasıyla Hoagland, 80, 160,
320, 640 ve 1280 µM Cr(VI) çözeltisi içeren kaplara alınmış ve 24 saat SANYO
MLR-351H bitki büyütme kabininde tutulmaya devam edilmiştir. Farklı
derişimlerdeki Cr(VI) stresinden 24 saatin sonunda örneklem alımı gerçekleştirilmiştir
(Şekil 4.9-4.13).

51
Şekil 4.1 100 mM ve 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidi

52
Şekil 4.2 100 mM ve 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidi

53
Şekil 4.3 100 mM ve 150 mM NaCl stresi uygulanmış Saray ayçiçeği çeşidi

54
Şekil 4.4 100 mM ve 150 mM NaCl stresi uygulanmış Tarsan 1018 ayçiçeği çeşidi

55
Şekil 4.5 100 mM ve 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidi

56
Şekil 4.6 100 mM ve 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidi

57
Şekil 4.7 100 mM ve 150 mM PEG stresi uygulanmış Saray ayçiçeği çeşidi

58
Şekil 4.8 100 mM ve 150 mM PEG stresi uygulanmış Tarsan 1018 ayçiçeği çeşidi

59
Şekil 4.9 Cr(VI) stresinin 24 saat uygulanmasının ardından bitkilerin genel görüntüsü

60
Şekil 4.10 08-TR-003 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI) stresi
uygulanan örnekler

61
Şekil 4.11 TR-3080 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI) stresi
uygulanan örnekler

62
Şekil 4.12 Saray ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI) stresi
uygulanan örnekler

63
Şekil 4.13 Tarsan 1018 ayçiçeği çeşidine 24 saat süreyle farklı derişimlerde Cr(VI)
stresi uygulanan örnekler
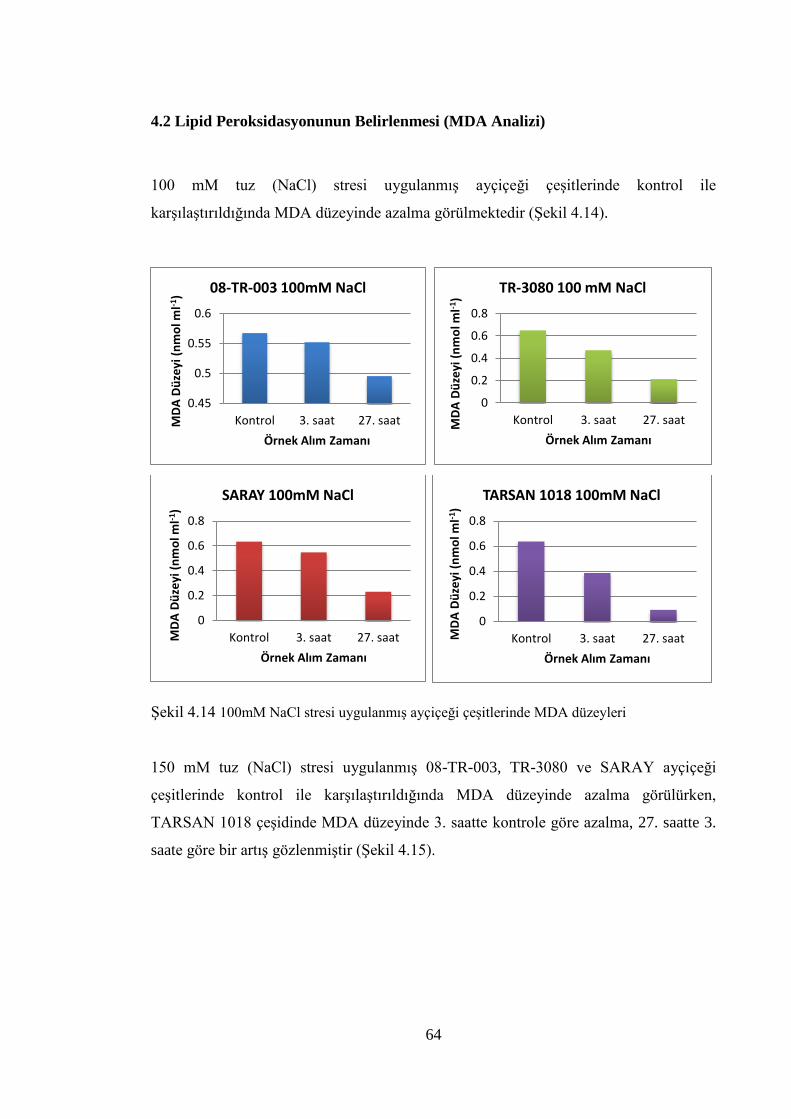
64
4.2 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)
100 mM tuz (NaCl) stresi uygulanmış ayçiçeği çeşitlerinde kontrol ile
karşılaştırıldığında MDA düzeyinde azalma görülmektedir (Şekil 4.14).
Şekil 4.14 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri
150 mM tuz (NaCl) stresi uygulanmış 08-TR-003, TR-3080 ve SARAY ayçiçeği
çeşitlerinde kontrol ile karşılaştırıldığında MDA düzeyinde azalma görülürken,
TARSAN 1018 çeşidinde MDA düzeyinde 3. saatte kontrole göre azalma, 27. saatte 3.
saate göre bir artış gözlenmiştir (Şekil 4.15).
0.45
0.5
0.55
0.6
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
08-TR-003 100mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
TR-3080 100 mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
SARAY 100mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
TARSAN 1018 100mM NaCl
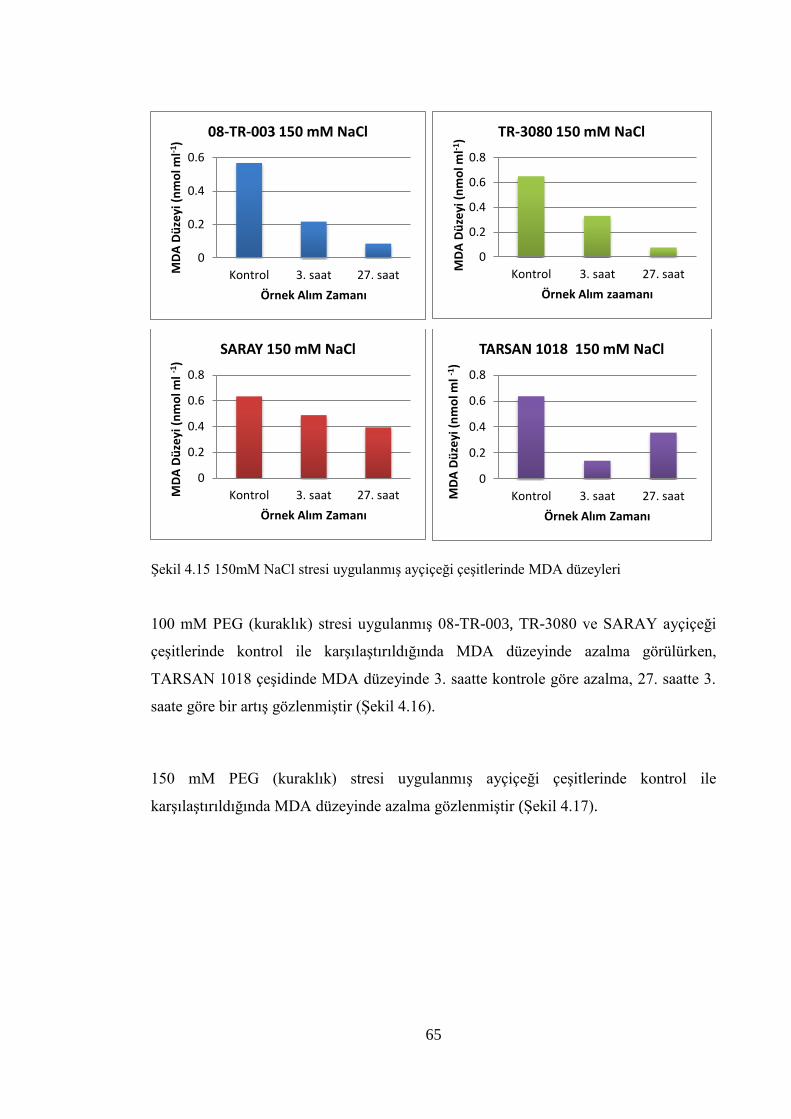
65
Şekil 4.15 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri
100 mM PEG (kuraklık) stresi uygulanmış 08-TR-003, TR-3080 ve SARAY ayçiçeği
çeşitlerinde kontrol ile karşılaştırıldığında MDA düzeyinde azalma görülürken,
TARSAN 1018 çeşidinde MDA düzeyinde 3. saatte kontrole göre azalma, 27. saatte 3.
saate göre bir artış gözlenmiştir (Şekil 4.16).
150 mM PEG (kuraklık) stresi uygulanmış ayçiçeği çeşitlerinde kontrol ile
karşılaştırıldığında MDA düzeyinde azalma gözlenmiştir (Şekil 4.17).
0
0.2
0.4
0.6
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
08-TR-003 150 mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım zaamanı
TR-3080 150 mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l -1)
Örnek Alım Zamanı
SARAY 150 mM NaCl
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l -1)
Örnek Alım Zamanı
TARSAN 1018 150 mM NaCl
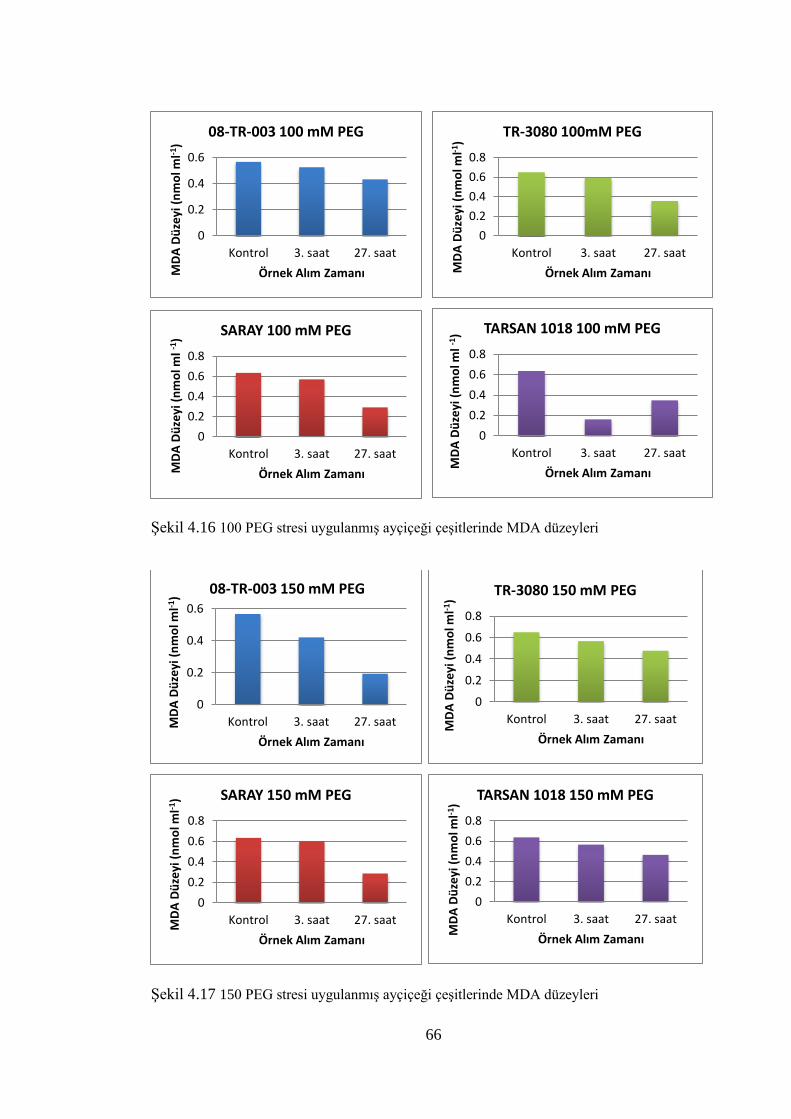
66
Şekil 4.16 100 PEG stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri
Şekil 4.17 150 PEG stresi uygulanmış ayçiçeği çeşitlerinde MDA düzeyleri
0
0.2
0.4
0.6
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
08-TR-003 100 mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
TR-3080 100mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l -1)
Örnek Alım Zamanı
SARAY 100 mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatM
DA
Dü
zeyi
(n
mo
l ml -1
)
Örnek Alım Zamanı
TARSAN 1018 100 mM PEG
0
0.2
0.4
0.6
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
08-TR-003 150 mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
TR-3080 150 mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saatMD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
SARAY 150 mM PEG
0
0.2
0.4
0.6
0.8
Kontrol 3. saat 27. saat
MD
A D
üze
yi (
nm
ol m
l-1)
Örnek Alım Zamanı
TARSAN 1018 150 mM PEG
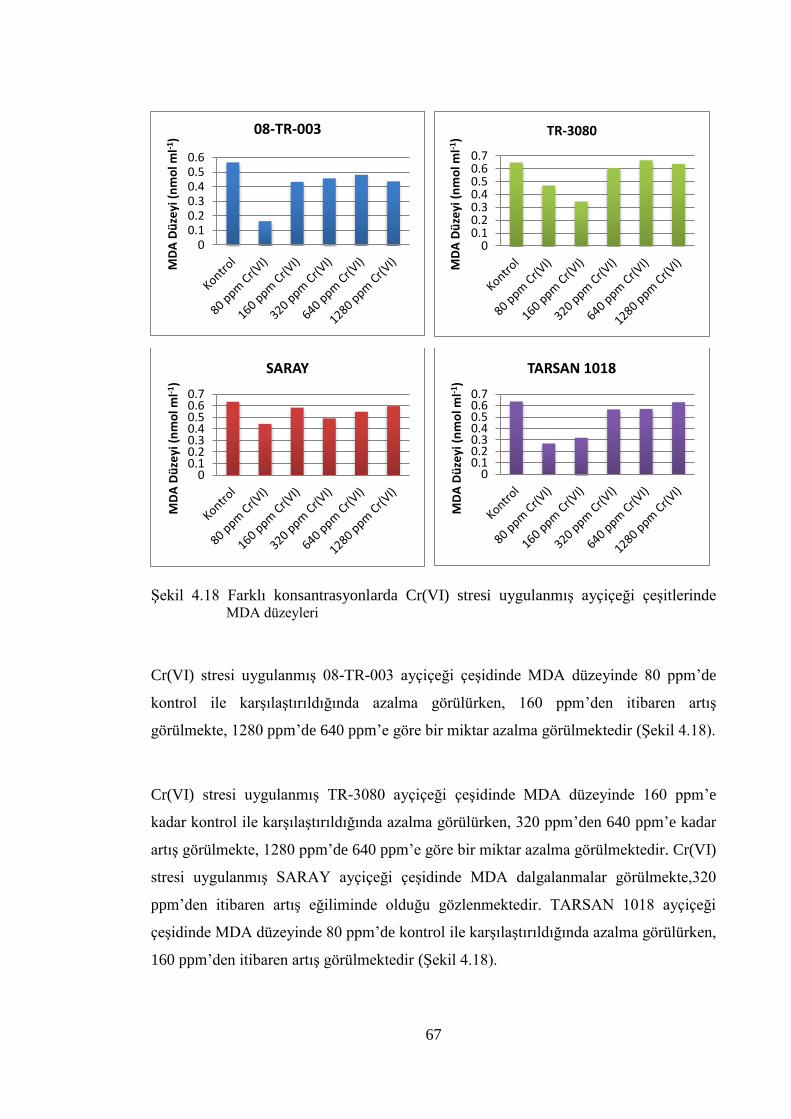
67
Şekil 4.18 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği çeşitlerinde
MDA düzeyleri
Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinde MDA düzeyinde 80 ppm’de
kontrol ile karşılaştırıldığında azalma görülürken, 160 ppm’den itibaren artış
görülmekte, 1280 ppm’de 640 ppm’e göre bir miktar azalma görülmektedir (Şekil 4.18).
Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinde MDA düzeyinde 160 ppm’e
kadar kontrol ile karşılaştırıldığında azalma görülürken, 320 ppm’den 640 ppm’e kadar
artış görülmekte, 1280 ppm’de 640 ppm’e göre bir miktar azalma görülmektedir. Cr(VI)
stresi uygulanmış SARAY ayçiçeği çeşidinde MDA dalgalanmalar görülmekte,320
ppm’den itibaren artış eğiliminde olduğu gözlenmektedir. TARSAN 1018 ayçiçeği
çeşidinde MDA düzeyinde 80 ppm’de kontrol ile karşılaştırıldığında azalma görülürken,
160 ppm’den itibaren artış görülmektedir (Şekil 4.18).
00.10.20.30.40.50.6
MD
A D
üze
yi (
nm
ol m
l-1)
08-TR-003
00.10.20.30.40.50.60.7
MD
A D
üze
yi (
nm
ol m
l-1)
TR-3080
00.10.20.30.40.50.60.7
MD
A D
üze
yi (
nm
ol m
l-1)
SARAY
00.10.20.30.40.50.60.7
MD
A D
üze
yi (
nm
ol m
l-1)
TARSAN 1018
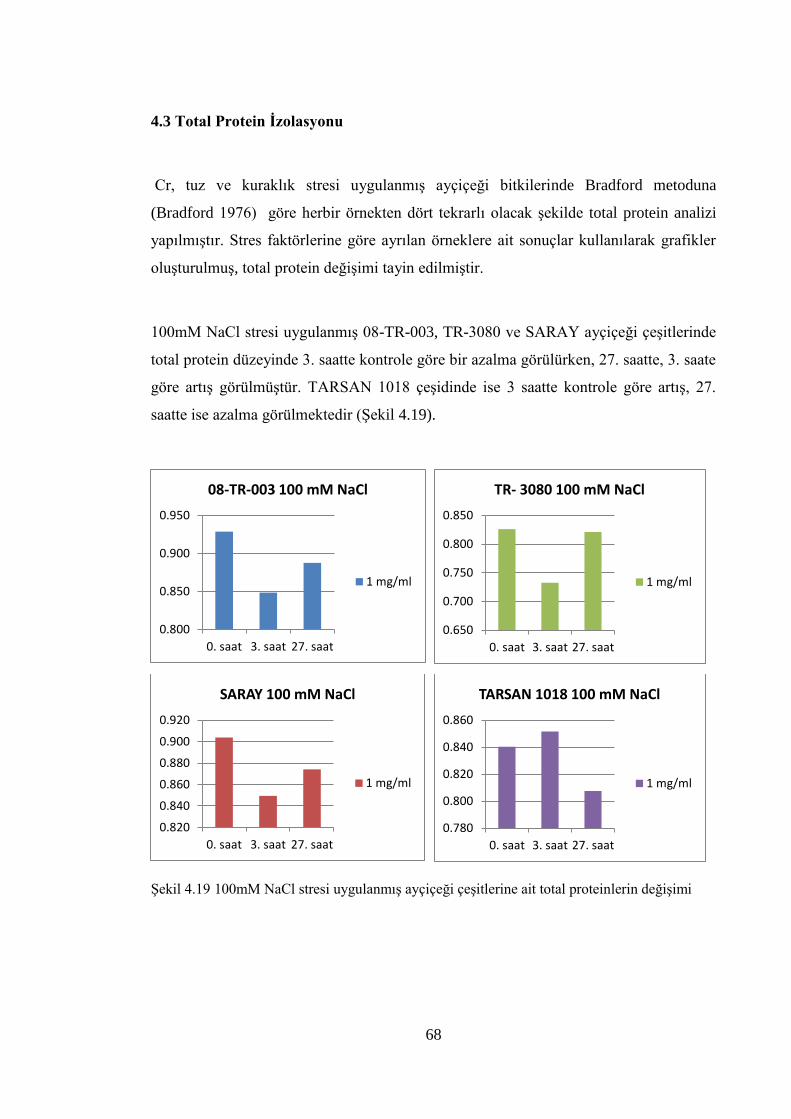
68
4.3 Total Protein İzolasyonu
Cr, tuz ve kuraklık stresi uygulanmış ayçiçeği bitkilerinde Bradford metoduna
(Bradford 1976) göre herbir örnekten dört tekrarlı olacak şekilde total protein analizi
yapılmıştır. Stres faktörlerine göre ayrılan örneklere ait sonuçlar kullanılarak grafikler
oluşturulmuş, total protein değişimi tayin edilmiştir.
100mM NaCl stresi uygulanmış 08-TR-003, TR-3080 ve SARAY ayçiçeği çeşitlerinde
total protein düzeyinde 3. saatte kontrole göre bir azalma görülürken, 27. saatte, 3. saate
göre artış görülmüştür. TARSAN 1018 çeşidinde ise 3 saatte kontrole göre artış, 27.
saatte ise azalma görülmektedir (Şekil 4.19).
Şekil 4.19 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerine ait total proteinlerin değişimi
0.800
0.850
0.900
0.950
0. saat 3. saat 27. saat
08-TR-003 100 mM NaCl
1 mg/ml
0.650
0.700
0.750
0.800
0.850
0. saat 3. saat 27. saat
TR- 3080 100 mM NaCl
1 mg/ml
0.820
0.840
0.860
0.880
0.900
0.920
0. saat 3. saat 27. saat
SARAY 100 mM NaCl
1 mg/ml
0.780
0.800
0.820
0.840
0.860
0. saat 3. saat 27. saat
TARSAN 1018 100 mM NaCl
1 mg/ml
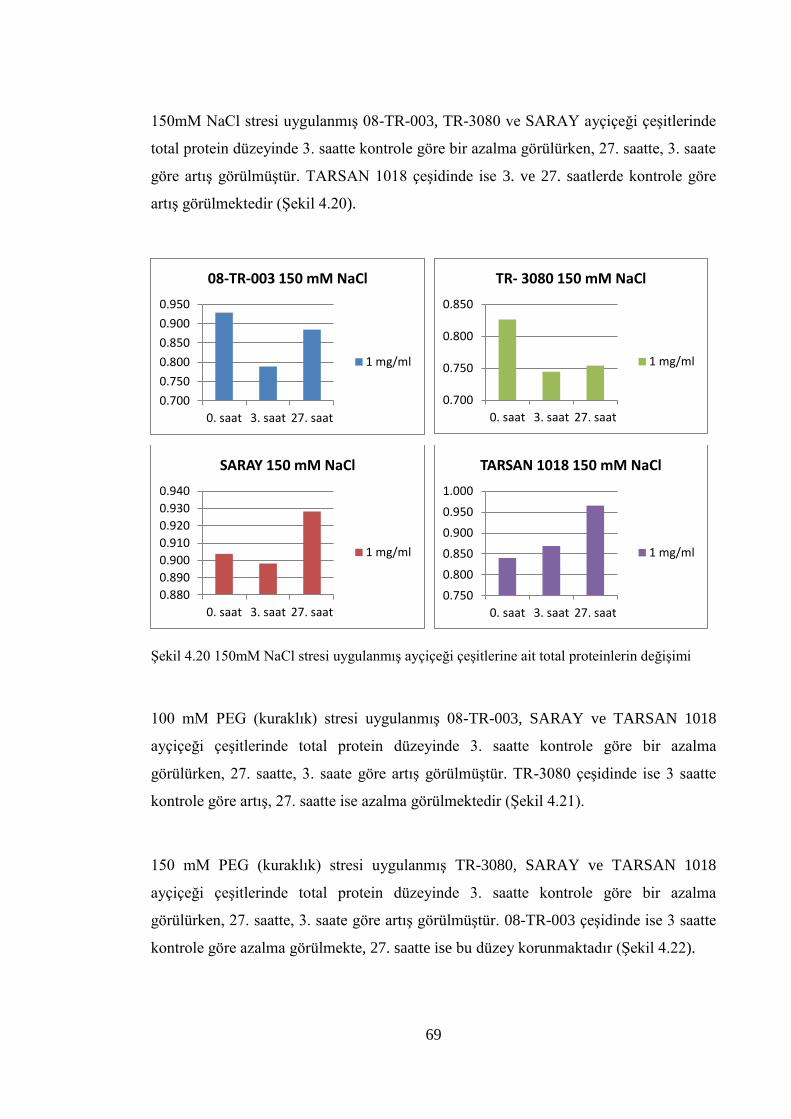
69
150mM NaCl stresi uygulanmış 08-TR-003, TR-3080 ve SARAY ayçiçeği çeşitlerinde
total protein düzeyinde 3. saatte kontrole göre bir azalma görülürken, 27. saatte, 3. saate
göre artış görülmüştür. TARSAN 1018 çeşidinde ise 3. ve 27. saatlerde kontrole göre
artış görülmektedir (Şekil 4.20).
Şekil 4.20 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerine ait total proteinlerin değişimi
100 mM PEG (kuraklık) stresi uygulanmış 08-TR-003, SARAY ve TARSAN 1018
ayçiçeği çeşitlerinde total protein düzeyinde 3. saatte kontrole göre bir azalma
görülürken, 27. saatte, 3. saate göre artış görülmüştür. TR-3080 çeşidinde ise 3 saatte
kontrole göre artış, 27. saatte ise azalma görülmektedir (Şekil 4.21).
150 mM PEG (kuraklık) stresi uygulanmış TR-3080, SARAY ve TARSAN 1018
ayçiçeği çeşitlerinde total protein düzeyinde 3. saatte kontrole göre bir azalma
görülürken, 27. saatte, 3. saate göre artış görülmüştür. 08-TR-003 çeşidinde ise 3 saatte
kontrole göre azalma görülmekte, 27. saatte ise bu düzey korunmaktadır (Şekil 4.22).
0.700
0.750
0.800
0.850
0.900
0.950
0. saat 3. saat 27. saat
08-TR-003 150 mM NaCl
1 mg/ml
0.700
0.750
0.800
0.850
0. saat 3. saat 27. saat
TR- 3080 150 mM NaCl
1 mg/ml
0.880
0.890
0.900
0.910
0.920
0.930
0.940
0. saat 3. saat 27. saat
SARAY 150 mM NaCl
1 mg/ml
0.750
0.800
0.850
0.900
0.950
1.000
0. saat 3. saat 27. saat
TARSAN 1018 150 mM NaCl
1 mg/ml
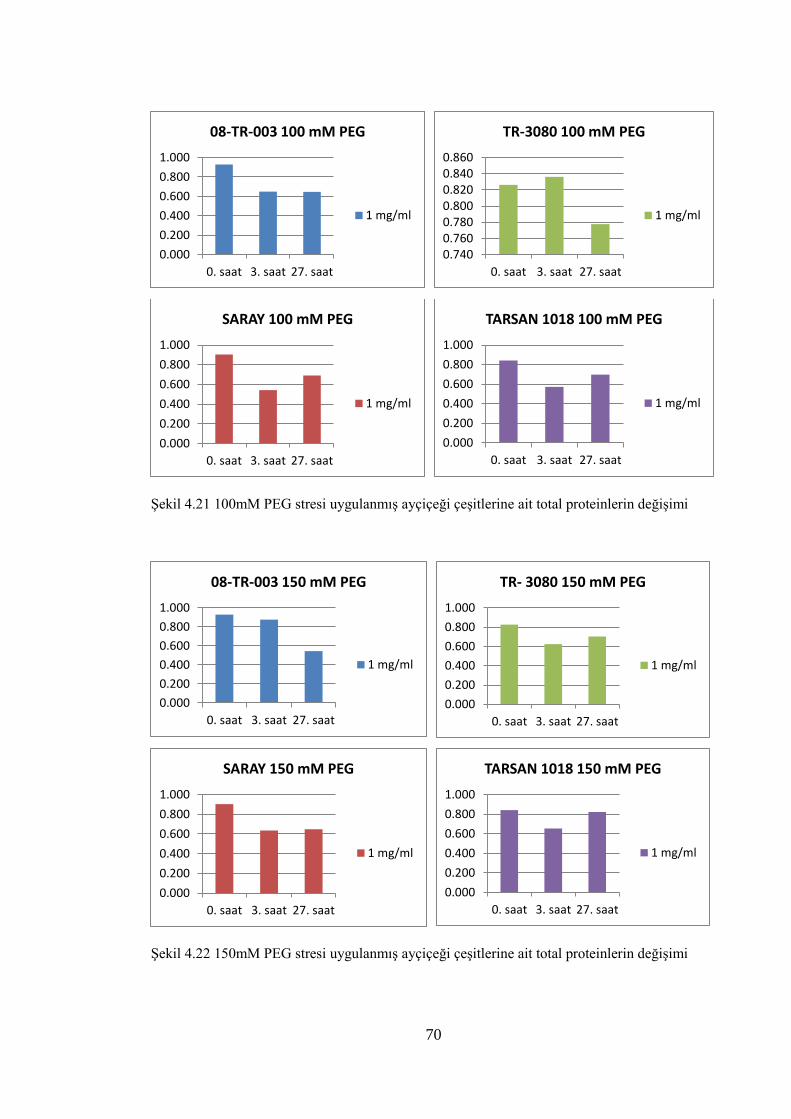
70
Şekil 4.21 100mM PEG stresi uygulanmış ayçiçeği çeşitlerine ait total proteinlerin değişimi
Şekil 4.22 150mM PEG stresi uygulanmış ayçiçeği çeşitlerine ait total proteinlerin değişimi
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
08-TR-003 100 mM PEG
1 mg/ml
0.7400.7600.7800.8000.8200.8400.860
0. saat 3. saat 27. saat
TR-3080 100 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
SARAY 100 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
TARSAN 1018 100 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
08-TR-003 150 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
TR- 3080 150 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
SARAY 150 mM PEG
1 mg/ml
0.000
0.200
0.400
0.600
0.800
1.000
0. saat 3. saat 27. saat
TARSAN 1018 150 mM PEG
1 mg/ml
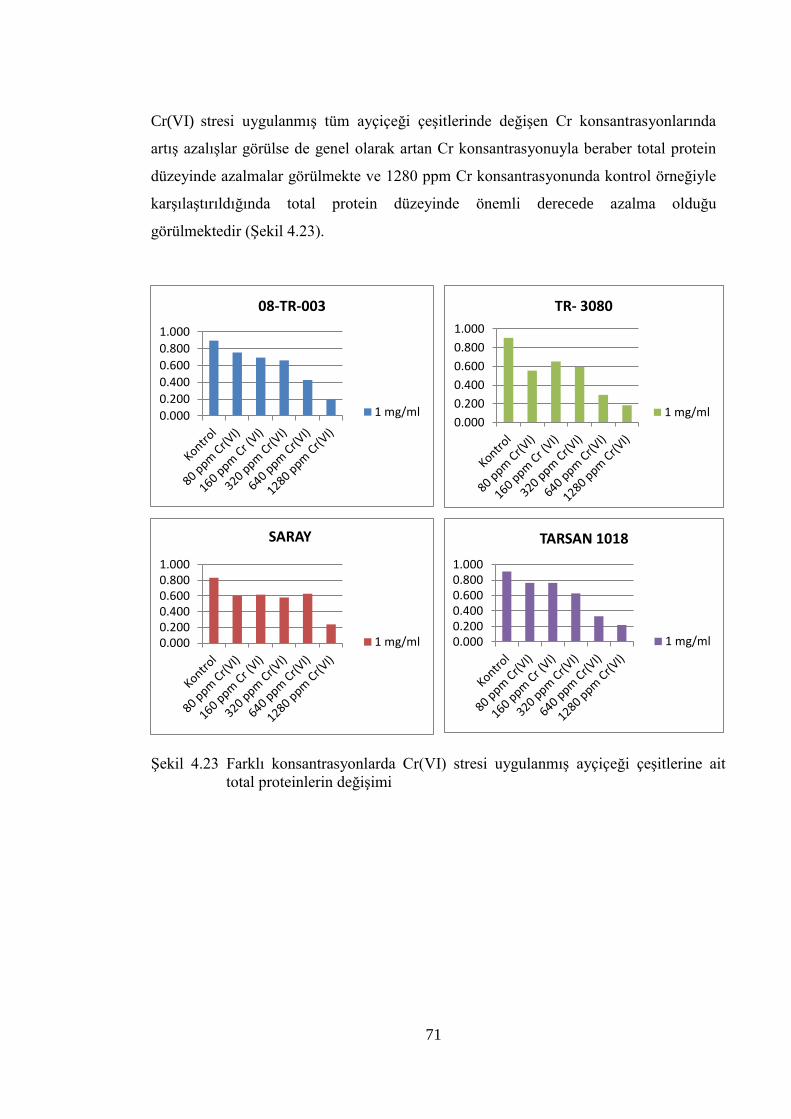
71
Cr(VI) stresi uygulanmış tüm ayçiçeği çeşitlerinde değişen Cr konsantrasyonlarında
artış azalışlar görülse de genel olarak artan Cr konsantrasyonuyla beraber total protein
düzeyinde azalmalar görülmekte ve 1280 ppm Cr konsantrasyonunda kontrol örneğiyle
karşılaştırıldığında total protein düzeyinde önemli derecede azalma olduğu
görülmektedir (Şekil 4.23).
Şekil 4.23 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği çeşitlerine ait
total proteinlerin değişimi
0.000
0.200
0.400
0.600
0.800
1.000
08-TR-003
1 mg/ml0.000
0.200
0.400
0.600
0.800
1.000
TR- 3080
1 mg/ml
0.0000.2000.4000.6000.8001.000
SARAY
1 mg/ml 0.0000.2000.4000.6000.8001.000
TARSAN 1018
1 mg/ml

72
4.4 Total RNA izolasyonu
Cr, tuz ve kuraklık stresi uygulanmış ayçiçeği bitkilerinden total RNA izolasyonu
TRIGent protokolüne göre yapılmış ve RNA miktarı spektrofotometrik ölçümlerle
Thermo Scientific Nanodrop 1000 Spectrophotometer cihazında belirlenmiştir.
İzole edilen RNA’lar % 1,2’lik agaroz jelde yürütülerek kontrol edilmiştir (Şekil 4.24).
Şekil 4.24 İzole edilen RNA örneklerinin DNase enzimi uygulaması öncesi %1,2’lik
agaroz jel görüntüsü
4.5 DNaz Uygulaması
Thermo Scientific EN0521 kodlu DNase enzimi kullanılarak izole edilen tüm RNA
örneklerine DNaz uygulaması gerçekleştirilmiş, olması muhtemel DNA
kontaminasyonu minumum düzeye getirilmiştir.
4.6 cDNA Sentezi
RNA örneklerinden cDNA sentezi, RevertAid First Strand cDNA Synthesis Kit
(Thermo Scientific) kullanılarak gerçekleştirilmiştir. Çizelge 4.1-4.2’de RNA
örneklerinden sentezlenmiş cDNA’ların saflık ve miktar ölçümleri yer almaktadır.
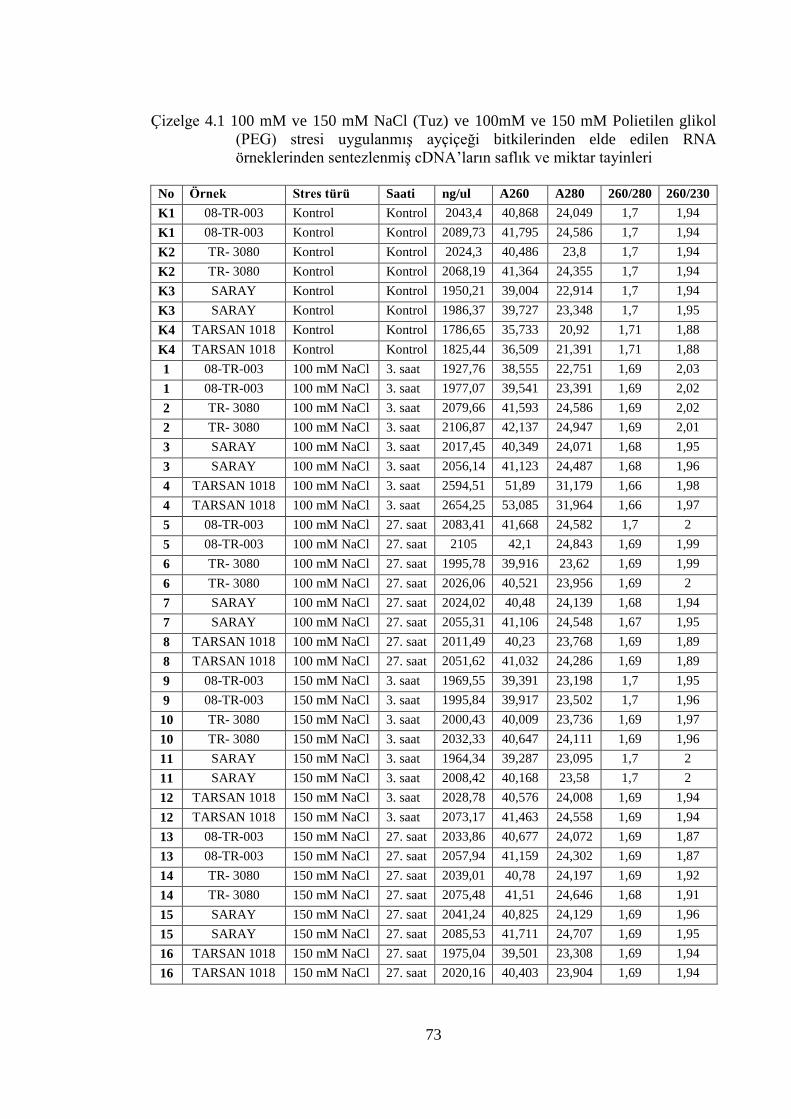
73
Çizelge 4.1 100 mM ve 150 mM NaCl (Tuz) ve 100mM ve 150 mM Polietilen glikol
(PEG) stresi uygulanmış ayçiçeği bitkilerinden elde edilen RNA
örneklerinden sentezlenmiş cDNA’ların saflık ve miktar tayinleri
No Örnek Stres türü Saati ng/ul A260 A280 260/280 260/230
K1 08-TR-003 Kontrol Kontrol 2043,4 40,868 24,049 1,7 1,94
K1 08-TR-003 Kontrol Kontrol 2089,73 41,795 24,586 1,7 1,94
K2 TR- 3080 Kontrol Kontrol 2024,3 40,486 23,8 1,7 1,94
K2 TR- 3080 Kontrol Kontrol 2068,19 41,364 24,355 1,7 1,94
K3 SARAY Kontrol Kontrol 1950,21 39,004 22,914 1,7 1,94
K3 SARAY Kontrol Kontrol 1986,37 39,727 23,348 1,7 1,95
K4 TARSAN 1018 Kontrol Kontrol 1786,65 35,733 20,92 1,71 1,88
K4 TARSAN 1018 Kontrol Kontrol 1825,44 36,509 21,391 1,71 1,88
1 08-TR-003 100 mM NaCl 3. saat 1927,76 38,555 22,751 1,69 2,03
1 08-TR-003 100 mM NaCl 3. saat 1977,07 39,541 23,391 1,69 2,02
2 TR- 3080 100 mM NaCl 3. saat 2079,66 41,593 24,586 1,69 2,02
2 TR- 3080 100 mM NaCl 3. saat 2106,87 42,137 24,947 1,69 2,01
3 SARAY 100 mM NaCl 3. saat 2017,45 40,349 24,071 1,68 1,95
3 SARAY 100 mM NaCl 3. saat 2056,14 41,123 24,487 1,68 1,96
4 TARSAN 1018 100 mM NaCl 3. saat 2594,51 51,89 31,179 1,66 1,98
4 TARSAN 1018 100 mM NaCl 3. saat 2654,25 53,085 31,964 1,66 1,97
5 08-TR-003 100 mM NaCl 27. saat 2083,41 41,668 24,582 1,7 2
5 08-TR-003 100 mM NaCl 27. saat 2105 42,1 24,843 1,69 1,99
6 TR- 3080 100 mM NaCl 27. saat 1995,78 39,916 23,62 1,69 1,99
6 TR- 3080 100 mM NaCl 27. saat 2026,06 40,521 23,956 1,69 2
7 SARAY 100 mM NaCl 27. saat 2024,02 40,48 24,139 1,68 1,94
7 SARAY 100 mM NaCl 27. saat 2055,31 41,106 24,548 1,67 1,95
8 TARSAN 1018 100 mM NaCl 27. saat 2011,49 40,23 23,768 1,69 1,89
8 TARSAN 1018 100 mM NaCl 27. saat 2051,62 41,032 24,286 1,69 1,89
9 08-TR-003 150 mM NaCl 3. saat 1969,55 39,391 23,198 1,7 1,95
9 08-TR-003 150 mM NaCl 3. saat 1995,84 39,917 23,502 1,7 1,96
10 TR- 3080 150 mM NaCl 3. saat 2000,43 40,009 23,736 1,69 1,97
10 TR- 3080 150 mM NaCl 3. saat 2032,33 40,647 24,111 1,69 1,96
11 SARAY 150 mM NaCl 3. saat 1964,34 39,287 23,095 1,7 2
11 SARAY 150 mM NaCl 3. saat 2008,42 40,168 23,58 1,7 2
12 TARSAN 1018 150 mM NaCl 3. saat 2028,78 40,576 24,008 1,69 1,94
12 TARSAN 1018 150 mM NaCl 3. saat 2073,17 41,463 24,558 1,69 1,94
13 08-TR-003 150 mM NaCl 27. saat 2033,86 40,677 24,072 1,69 1,87
13 08-TR-003 150 mM NaCl 27. saat 2057,94 41,159 24,302 1,69 1,87
14 TR- 3080 150 mM NaCl 27. saat 2039,01 40,78 24,197 1,69 1,92
14 TR- 3080 150 mM NaCl 27. saat 2075,48 41,51 24,646 1,68 1,91
15 SARAY 150 mM NaCl 27. saat 2041,24 40,825 24,129 1,69 1,96
15 SARAY 150 mM NaCl 27. saat 2085,53 41,711 24,707 1,69 1,95
16 TARSAN 1018 150 mM NaCl 27. saat 1975,04 39,501 23,308 1,69 1,94
16 TARSAN 1018 150 mM NaCl 27. saat 2020,16 40,403 23,904 1,69 1,94

74
Çizelge 4.1 100 mM ve 150 mM NaCl (Tuz) ve 100mM ve 150 mM Polietilen glikol
(PEG) stresi uygulanmış ayçiçeği bitkilerinden elde edilen RNA
örneklerinden sentezlenmiş cDNA’ların saflık ve miktar tayinleri (devam)
No Örnek Stres türü Saati ng/ul A260 A280 260/280 260/230
17 08-TR-003 100 mM PEG 3. saat 1905,22 38,104 22,666 1,68 1,95
17 08-TR-003 100 mM PEG 3. saat 1937,28 38,746 23,079 1,68 1,95
18 TR- 3080 100 mM PEG 3. saat 2042,95 40,859 24,189 1,69 1,98
18 TR- 3080 100 mM PEG 3. saat 2078,1 41,562 24,712 1,68 1,99
19 SARAY 100 mM PEG 3. saat 2014,5 40,29 23,89 1,69 1,99
19 SARAY 100 mM PEG 3. saat 2047,34 40,947 24,368 1,68 1,99
20 TARSAN 1018 100 mM PEG 3. saat 1994,8 39,896 23,655 1,69 1,98
20 TARSAN 1018 100 mM PEG 3. saat 2024,37 40,487 23,976 1,69 1,97
21 08-TR-003 100 mM PEG 27. saat 2057,75 41,155 24,303 1,69 2,01
21 08-TR-003 100 mM PEG 27. saat 2105,04 42,101 24,947 1,69 2,01
22 TR- 3080 100 mM PEG 27. saat 2005,49 40,11 23,736 1,69 1,99
22 TR- 3080 100 mM PEG 27. saat 2037,19 40,744 24,119 1,69 2
23 SARAY 100 mM PEG 27. saat 1898,97 37,979 22,634 1,68 1,96
23 SARAY 100 mM PEG 27. saat 1933,29 38,666 23,011 1,68 1,96
24 TARSAN 1018 100 mM PEG 27. saat 2115,73 42,315 25,237 1,68 1,96
24 TARSAN 1018 100 mM PEG 27. saat 2147,44 42,949 25,668 1,67 1,98
25 08-TR-003 150 mM PEG 3. saat 2122,88 42,458 25,138 1,69 1,98
25 08-TR-003 150 mM PEG 3. saat 2147,17 42,943 25,506 1,68 1,98
26 TR- 3080 150 mM PEG 3. saat 1979,11 39,582 23,286 1,7 1,99
26 TR- 3080 150 mM PEG 3. saat 2002 40,04 23,61 1,7 1,99
27 SARAY 150 mM PEG 3. saat 2049,27 40,985 24,293 1,69 1,98
27 SARAY 150 mM PEG 3. saat 2090,36 41,807 24,752 1,69 1,98
28 TARSAN 1018 150 mM PEG 3. saat 2031,82 40,636 24,146 1,68 1,96
28 TARSAN 1018 150 mM PEG 3. saat 2059,19 41,184 24,492 1,68 1,96
29 08-TR-003 150 mM PEG 27. saat 2004,84 40,097 23,641 1,7 2
29 08-TR-003 150 mM PEG 27. saat 2043,92 40,878 24,131 1,69 2
30 TR- 3080 150 mM PEG 27. saat 1862,05 37,241 22,169 1,68 1,94
30 TR- 3080 150 mM PEG 27. saat 1898,66 37,973 22,649 1,68 1,94
31 SARAY 150 mM PEG 27. saat 2014,22 40,284 23,787 1,69 1,97
31 SARAY 150 mM PEG 27. saat 2051,8 41,036 24,222 1,69 1,97
32 TARSAN 1018 150 mM PEG 27. saat 1897,81 37,956 22,529 1,68 1,93
32 TARSAN 1018 150 mM PEG 27. saat 1935,67 38,713 23,074 1,68 1,94
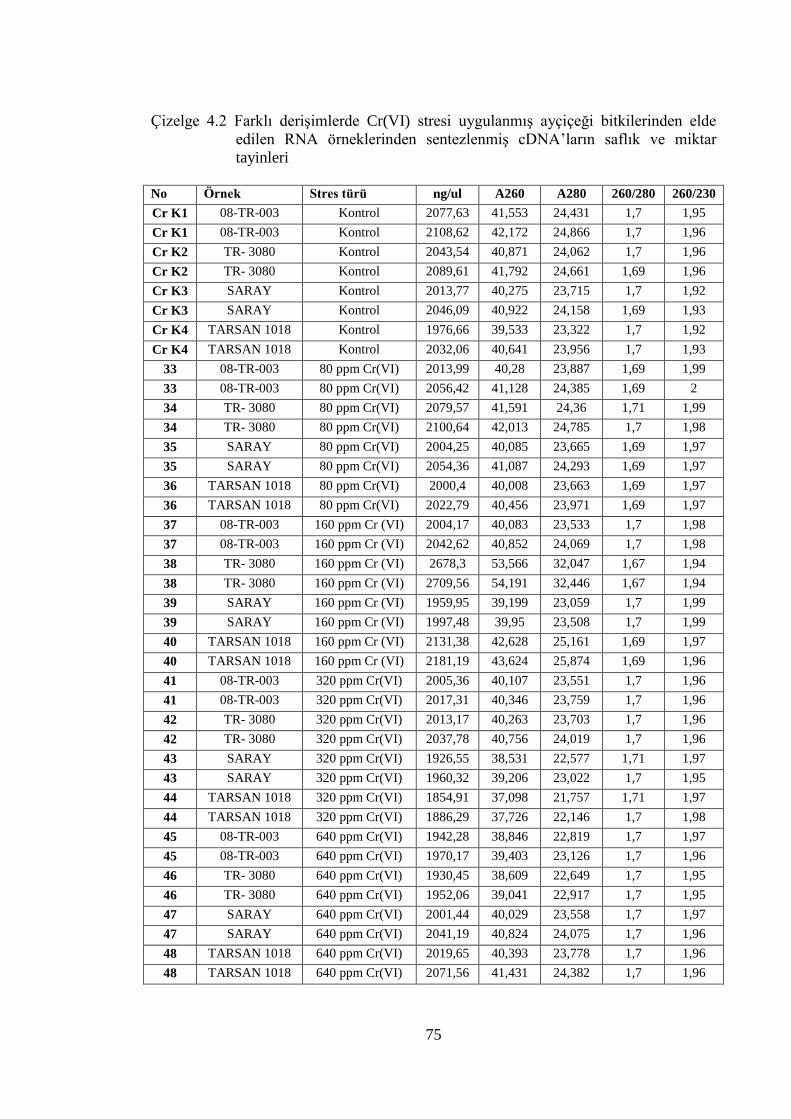
75
Çizelge 4.2 Farklı derişimlerde Cr(VI) stresi uygulanmış ayçiçeği bitkilerinden elde
edilen RNA örneklerinden sentezlenmiş cDNA’ların saflık ve miktar
tayinleri
No Örnek Stres türü ng/ul A260 A280 260/280 260/230
Cr K1 08-TR-003 Kontrol 2077,63 41,553 24,431 1,7 1,95
Cr K1 08-TR-003 Kontrol 2108,62 42,172 24,866 1,7 1,96
Cr K2 TR- 3080 Kontrol 2043,54 40,871 24,062 1,7 1,96
Cr K2 TR- 3080 Kontrol 2089,61 41,792 24,661 1,69 1,96
Cr K3 SARAY Kontrol 2013,77 40,275 23,715 1,7 1,92
Cr K3 SARAY Kontrol 2046,09 40,922 24,158 1,69 1,93
Cr K4 TARSAN 1018 Kontrol 1976,66 39,533 23,322 1,7 1,92
Cr K4 TARSAN 1018 Kontrol 2032,06 40,641 23,956 1,7 1,93
33 08-TR-003 80 ppm Cr(VI) 2013,99 40,28 23,887 1,69 1,99
33 08-TR-003 80 ppm Cr(VI) 2056,42 41,128 24,385 1,69 2
34 TR- 3080 80 ppm Cr(VI) 2079,57 41,591 24,36 1,71 1,99
34 TR- 3080 80 ppm Cr(VI) 2100,64 42,013 24,785 1,7 1,98
35 SARAY 80 ppm Cr(VI) 2004,25 40,085 23,665 1,69 1,97
35 SARAY 80 ppm Cr(VI) 2054,36 41,087 24,293 1,69 1,97
36 TARSAN 1018 80 ppm Cr(VI) 2000,4 40,008 23,663 1,69 1,97
36 TARSAN 1018 80 ppm Cr(VI) 2022,79 40,456 23,971 1,69 1,97
37 08-TR-003 160 ppm Cr (VI) 2004,17 40,083 23,533 1,7 1,98
37 08-TR-003 160 ppm Cr (VI) 2042,62 40,852 24,069 1,7 1,98
38 TR- 3080 160 ppm Cr (VI) 2678,3 53,566 32,047 1,67 1,94
38 TR- 3080 160 ppm Cr (VI) 2709,56 54,191 32,446 1,67 1,94
39 SARAY 160 ppm Cr (VI) 1959,95 39,199 23,059 1,7 1,99
39 SARAY 160 ppm Cr (VI) 1997,48 39,95 23,508 1,7 1,99
40 TARSAN 1018 160 ppm Cr (VI) 2131,38 42,628 25,161 1,69 1,97
40 TARSAN 1018 160 ppm Cr (VI) 2181,19 43,624 25,874 1,69 1,96
41 08-TR-003 320 ppm Cr(VI) 2005,36 40,107 23,551 1,7 1,96
41 08-TR-003 320 ppm Cr(VI) 2017,31 40,346 23,759 1,7 1,96
42 TR- 3080 320 ppm Cr(VI) 2013,17 40,263 23,703 1,7 1,96
42 TR- 3080 320 ppm Cr(VI) 2037,78 40,756 24,019 1,7 1,96
43 SARAY 320 ppm Cr(VI) 1926,55 38,531 22,577 1,71 1,97
43 SARAY 320 ppm Cr(VI) 1960,32 39,206 23,022 1,7 1,95
44 TARSAN 1018 320 ppm Cr(VI) 1854,91 37,098 21,757 1,71 1,97
44 TARSAN 1018 320 ppm Cr(VI) 1886,29 37,726 22,146 1,7 1,98
45 08-TR-003 640 ppm Cr(VI) 1942,28 38,846 22,819 1,7 1,97
45 08-TR-003 640 ppm Cr(VI) 1970,17 39,403 23,126 1,7 1,96
46 TR- 3080 640 ppm Cr(VI) 1930,45 38,609 22,649 1,7 1,95
46 TR- 3080 640 ppm Cr(VI) 1952,06 39,041 22,917 1,7 1,95
47 SARAY 640 ppm Cr(VI) 2001,44 40,029 23,558 1,7 1,97
47 SARAY 640 ppm Cr(VI) 2041,19 40,824 24,075 1,7 1,96
48 TARSAN 1018 640 ppm Cr(VI) 2019,65 40,393 23,778 1,7 1,96
48 TARSAN 1018 640 ppm Cr(VI) 2071,56 41,431 24,382 1,7 1,96
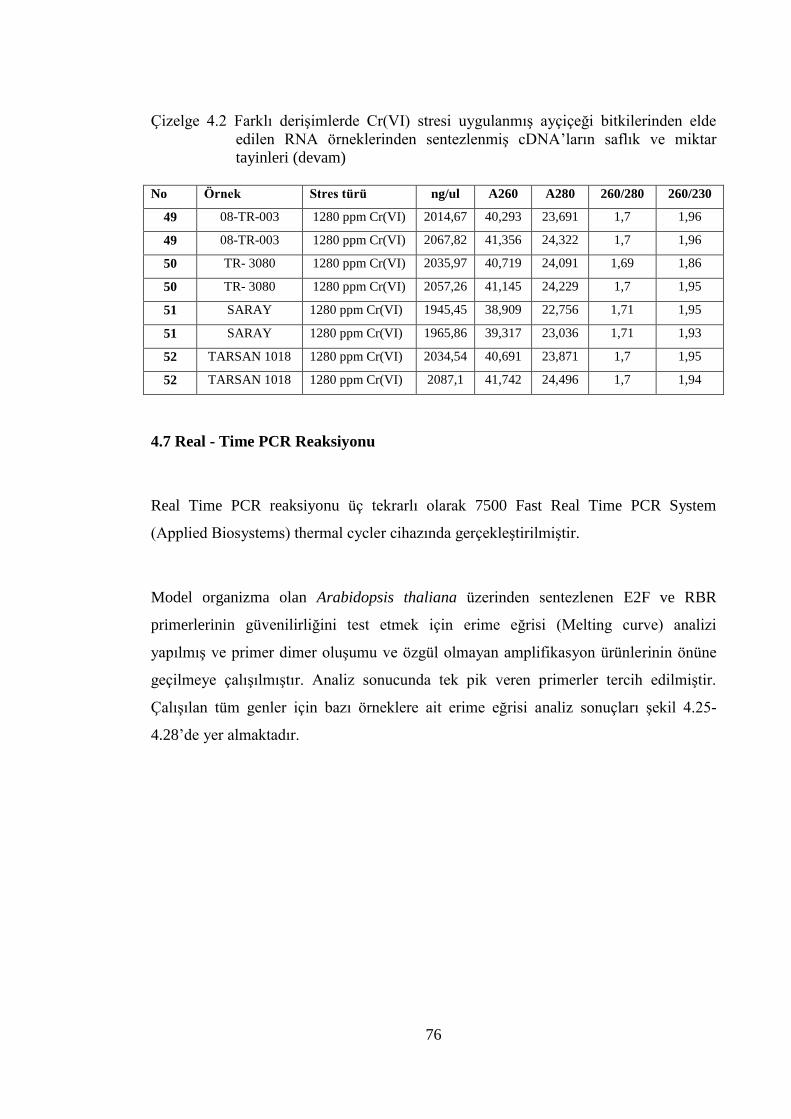
76
Çizelge 4.2 Farklı derişimlerde Cr(VI) stresi uygulanmış ayçiçeği bitkilerinden elde
edilen RNA örneklerinden sentezlenmiş cDNA’ların saflık ve miktar
tayinleri (devam)
No Örnek Stres türü ng/ul A260 A280 260/280 260/230
49 08-TR-003 1280 ppm Cr(VI) 2014,67 40,293 23,691 1,7 1,96
49 08-TR-003 1280 ppm Cr(VI) 2067,82 41,356 24,322 1,7 1,96
50 TR- 3080 1280 ppm Cr(VI) 2035,97 40,719 24,091 1,69 1,86
50 TR- 3080 1280 ppm Cr(VI) 2057,26 41,145 24,229 1,7 1,95
51 SARAY 1280 ppm Cr(VI) 1945,45 38,909 22,756 1,71 1,95
51 SARAY 1280 ppm Cr(VI) 1965,86 39,317 23,036 1,71 1,93
52 TARSAN 1018 1280 ppm Cr(VI) 2034,54 40,691 23,871 1,7 1,95
52 TARSAN 1018 1280 ppm Cr(VI) 2087,1 41,742 24,496 1,7 1,94
4.7 Real - Time PCR Reaksiyonu
Real Time PCR reaksiyonu üç tekrarlı olarak 7500 Fast Real Time PCR System
(Applied Biosystems) thermal cycler cihazında gerçekleştirilmiştir.
Model organizma olan Arabidopsis thaliana üzerinden sentezlenen E2F ve RBR
primerlerinin güvenilirliğini test etmek için erime eğrisi (Melting curve) analizi
yapılmış ve primer dimer oluşumu ve özgül olmayan amplifikasyon ürünlerinin önüne
geçilmeye çalışılmıştır. Analiz sonucunda tek pik veren primerler tercih edilmiştir.
Çalışılan tüm genler için bazı örneklere ait erime eğrisi analiz sonuçları şekil 4.25-
4.28’de yer almaktadır.
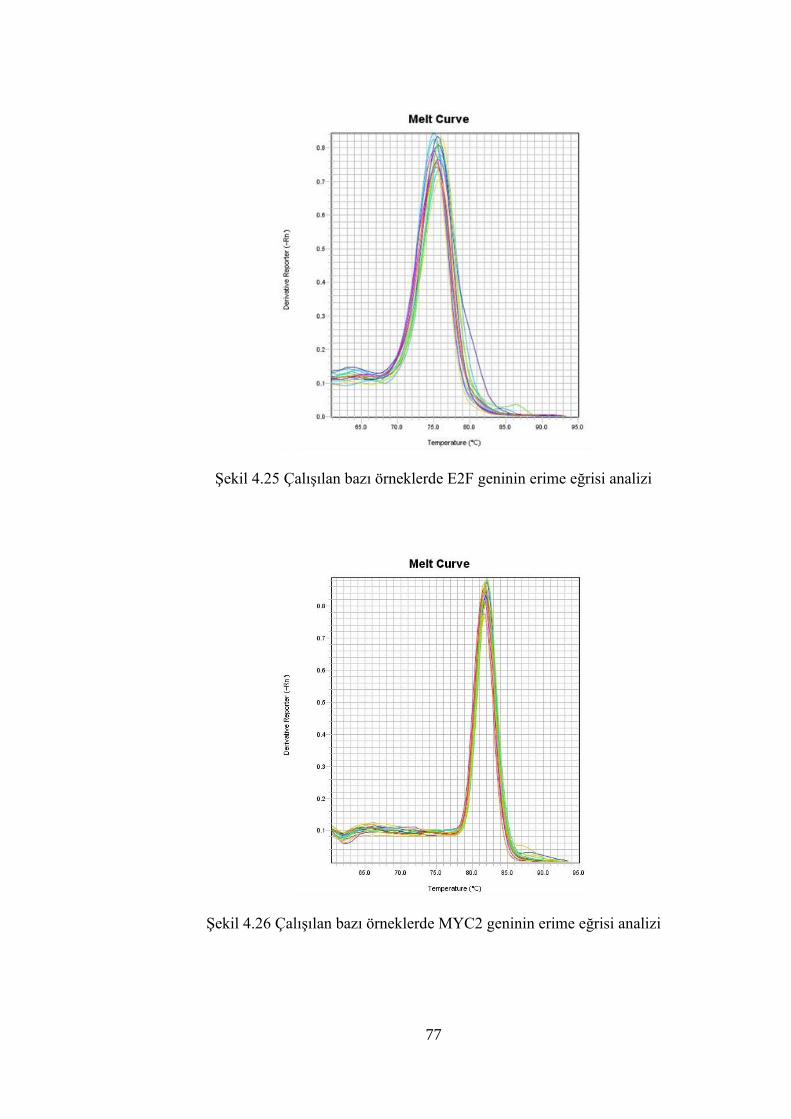
77
Şekil 4.25 Çalışılan bazı örneklerde E2F geninin erime eğrisi analizi
Şekil 4.26 Çalışılan bazı örneklerde MYC2 geninin erime eğrisi analizi
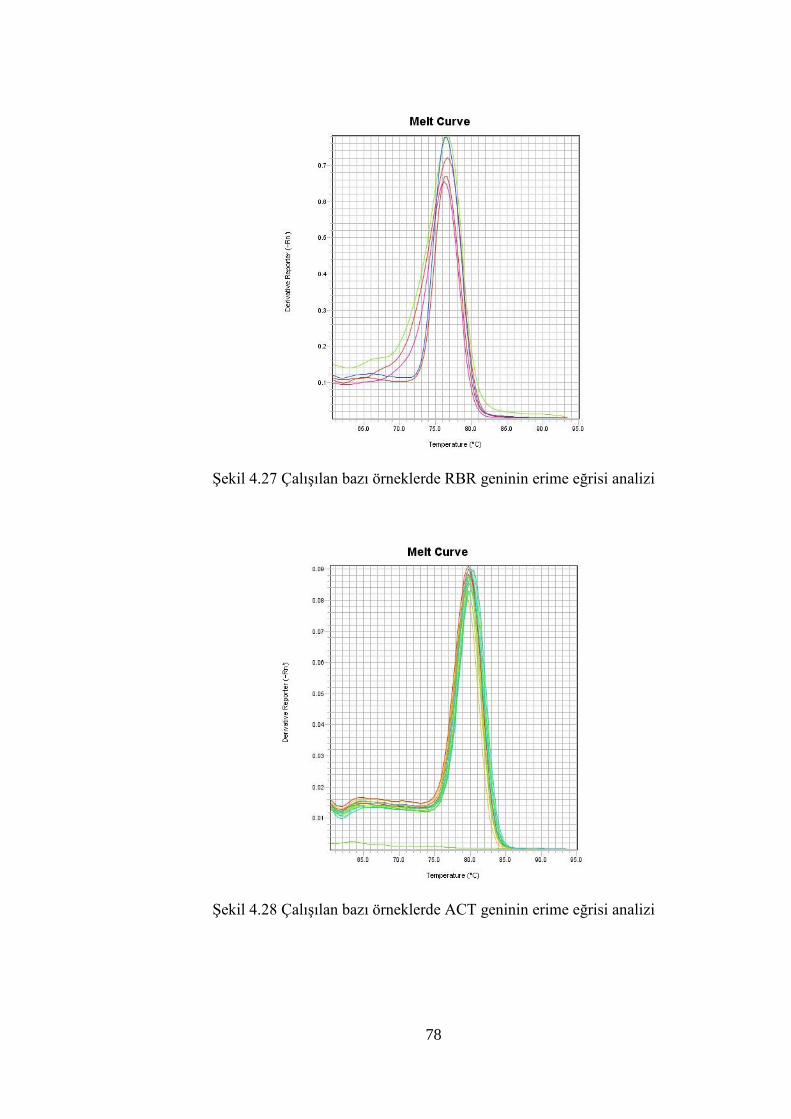
78
Şekil 4.27 Çalışılan bazı örneklerde RBR geninin erime eğrisi analizi
Şekil 4.28 Çalışılan bazı örneklerde ACT geninin erime eğrisi analizi
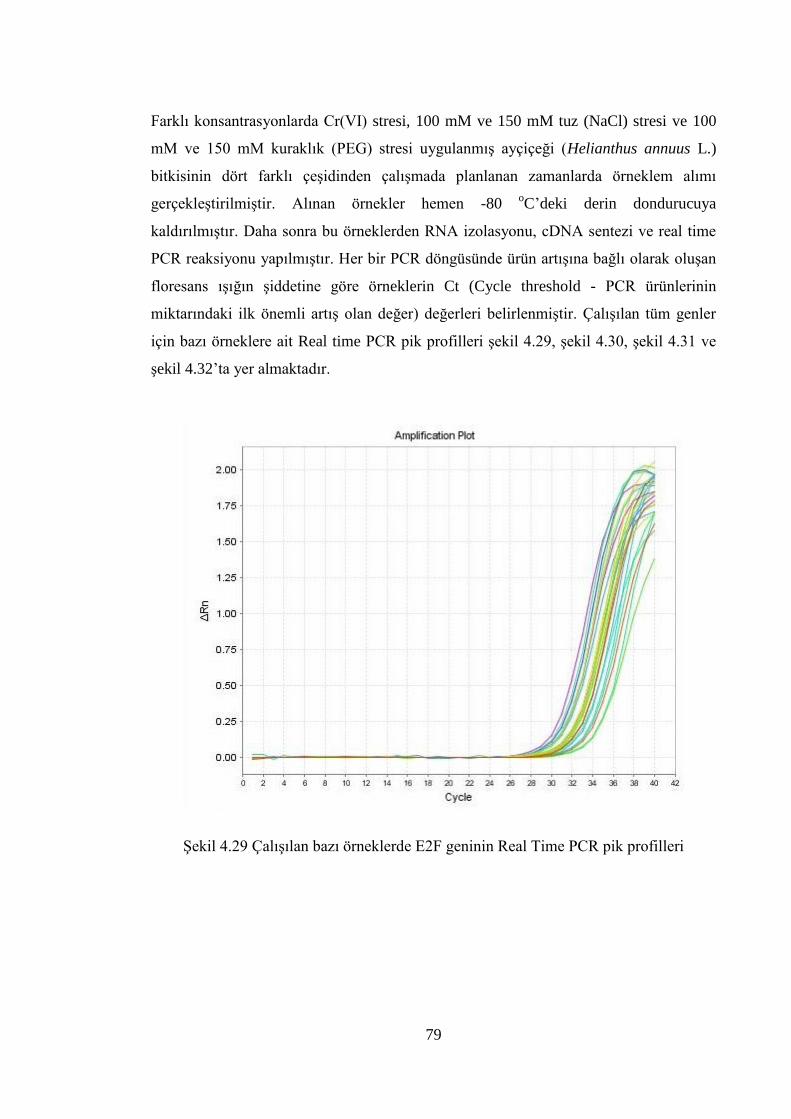
79
Farklı konsantrasyonlarda Cr(VI) stresi, 100 mM ve 150 mM tuz (NaCl) stresi ve 100
mM ve 150 mM kuraklık (PEG) stresi uygulanmış ayçiçeği (Helianthus annuus L.)
bitkisinin dört farklı çeşidinden çalışmada planlanan zamanlarda örneklem alımı
gerçekleştirilmiştir. Alınan örnekler hemen -80 oC’deki derin dondurucuya
kaldırılmıştır. Daha sonra bu örneklerden RNA izolasyonu, cDNA sentezi ve real time
PCR reaksiyonu yapılmıştır. Her bir PCR döngüsünde ürün artışına bağlı olarak oluşan
floresans ışığın şiddetine göre örneklerin Ct (Cycle threshold - PCR ürünlerinin
miktarındaki ilk önemli artış olan değer) değerleri belirlenmiştir. Çalışılan tüm genler
için bazı örneklere ait Real time PCR pik profilleri şekil 4.29, şekil 4.30, şekil 4.31 ve
şekil 4.32’ta yer almaktadır.
Şekil 4.29 Çalışılan bazı örneklerde E2F geninin Real Time PCR pik profilleri
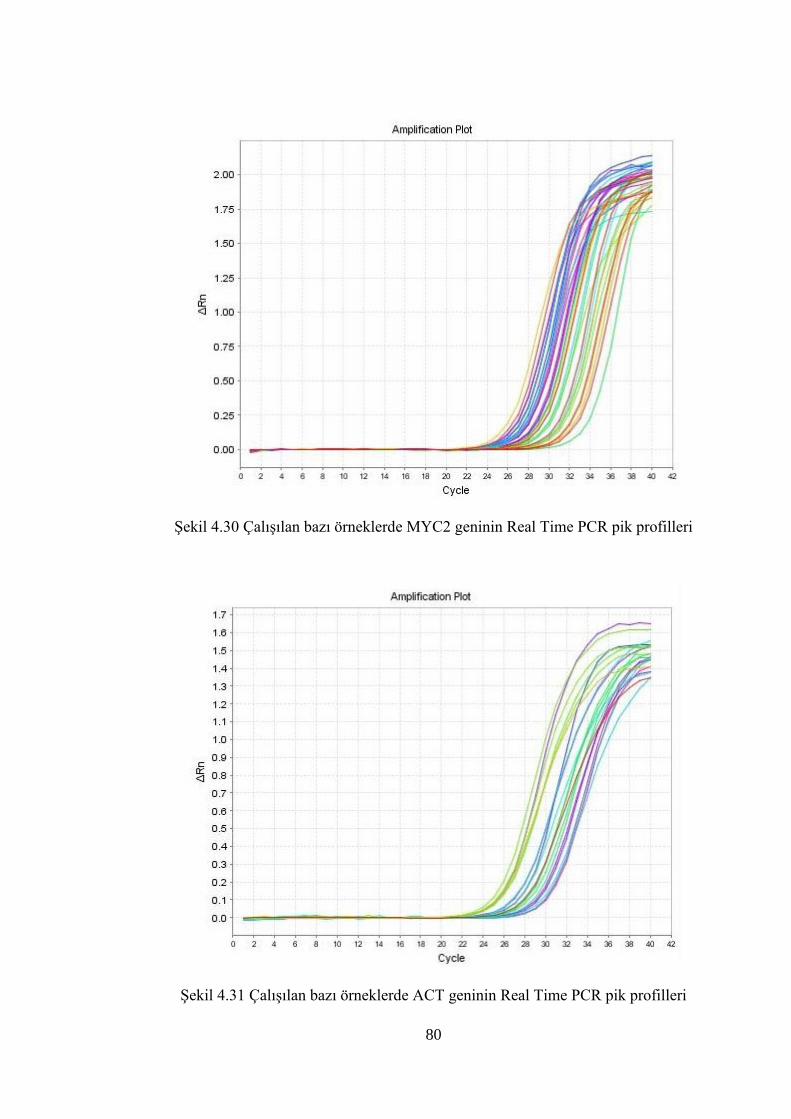
80
Şekil 4.30 Çalışılan bazı örneklerde MYC2 geninin Real Time PCR pik profilleri
Şekil 4.31 Çalışılan bazı örneklerde ACT geninin Real Time PCR pik profilleri

81
Şekil 4.32 Çalışılan bazı örneklerde RBR geninin Real Time PCR pik profilleri
4.8 Normalizasyon ve İstatistiksel Analiz
Farklı sürelerde tuz (NaCl), kuraklık (PEG), Cr(VI) stresi uygulanmış 08-TR-003, TR-
3080, SARAY ve TARSAN 1018 ayçiçeği bitkilerinde E2F, RBR ve MYC2 hedef
genlerine ait ifade profilleri, çalışmada housekeeping gen olarak kullanılan aktin (ACT)
ve kontrol şartları dikkate alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Livak ve
Schmittgen 2001). Normalizasyon ile elde edilen gen ekspresyonu verilerinin ortalama,
standart sapma, standart hata ve istatistiksel olarak anlamlılık dereceleri SPSS 17
istatistik programında yer alan One Way ANOVA (Dunnett) testi ile hesaplanmıştır.
Çalışmada kullanılan tüm örneklerin E2F, RBR ve MYC2 genlerine ait normalize
edilmiş gen ekspresyonu verilerinin ortalaması ve standart hataları alınarak elde edilen
veriler ile her bir gene ait ifade düzeyini gösteren grafikler çizilmiş ve mRNA ifade
seviyelerindeki değişimler tespit edilmeye çalışılmıştır.
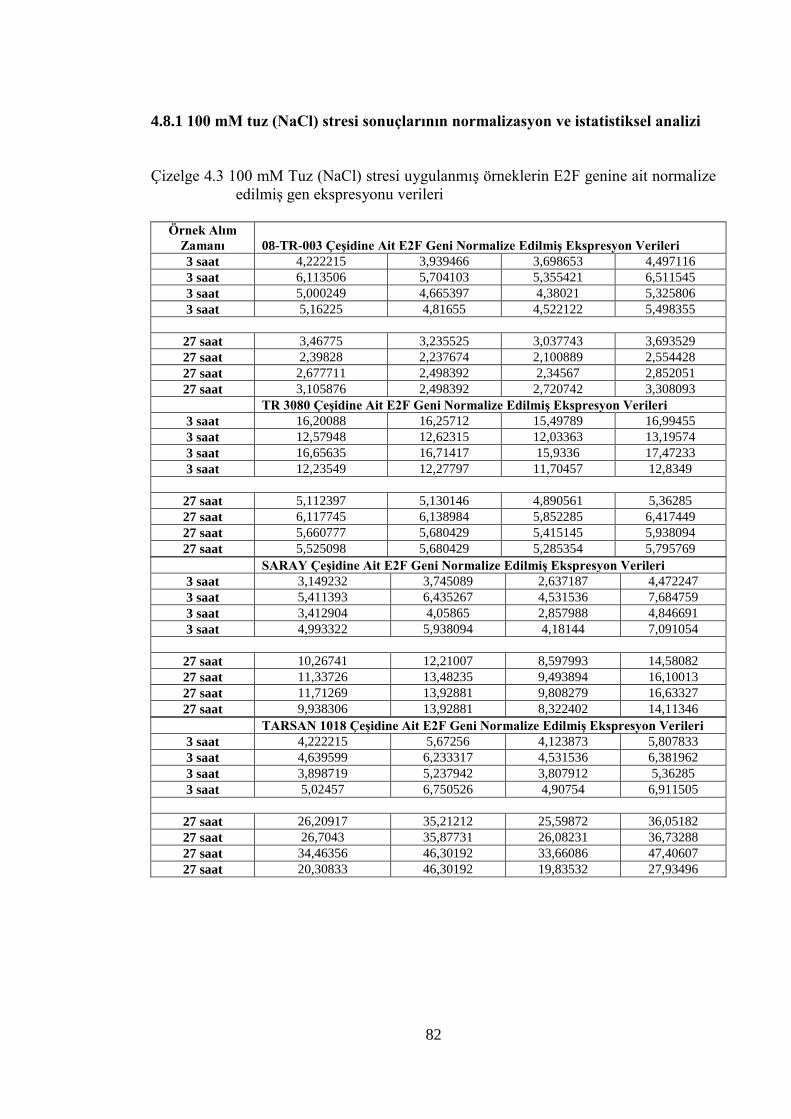
82
4.8.1 100 mM tuz (NaCl) stresi sonuçlarının normalizasyon ve istatistiksel analizi
Çizelge 4.3 100 mM Tuz (NaCl) stresi uygulanmış örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 4,222215 3,939466 3,698653 4,497116
3 saat 6,113506 5,704103 5,355421 6,511545
3 saat 5,000249 4,665397 4,38021 5,325806
3 saat 5,16225 4,81655 4,522122 5,498355
27 saat 3,46775 3,235525 3,037743 3,693529
27 saat 2,39828 2,237674 2,100889 2,554428
27 saat 2,677711 2,498392 2,34567 2,852051
27 saat 3,105876 2,498392 2,720742 3,308093
TR 3080 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 16,20088 16,25712 15,49789 16,99455
3 saat 12,57948 12,62315 12,03363 13,19574
3 saat 16,65635 16,71417 15,9336 17,47233
3 saat 12,23549 12,27797 11,70457 12,8349
27 saat 5,112397 5,130146 4,890561 5,36285
27 saat 6,117745 6,138984 5,852285 6,417449
27 saat 5,660777 5,680429 5,415145 5,938094
27 saat 5,525098 5,680429 5,285354 5,795769
SARAY Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 3,149232 3,745089 2,637187 4,472247
3 saat 5,411393 6,435267 4,531536 7,684759
3 saat 3,412904 4,05865 2,857988 4,846691
3 saat 4,993322 5,938094 4,18144 7,091054
27 saat 10,26741 12,21007 8,597993 14,58082
27 saat 11,33726 13,48235 9,493894 16,10013
27 saat 11,71269 13,92881 9,808279 16,63327
27 saat 9,938306 13,92881 8,322402 14,11346
TARSAN 1018 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 4,222215 5,67256 4,123873 5,807833
3 saat 4,639599 6,233317 4,531536 6,381962
3 saat 3,898719 5,237942 3,807912 5,36285
3 saat 5,02457 6,750526 4,90754 6,911505
27 saat 26,20917 35,21212 25,59872 36,05182
27 saat 26,7043 35,87731 26,08231 36,73288
27 saat 34,46356 46,30192 33,66086 47,40607
27 saat 20,30833 46,30192 19,83532 27,93496
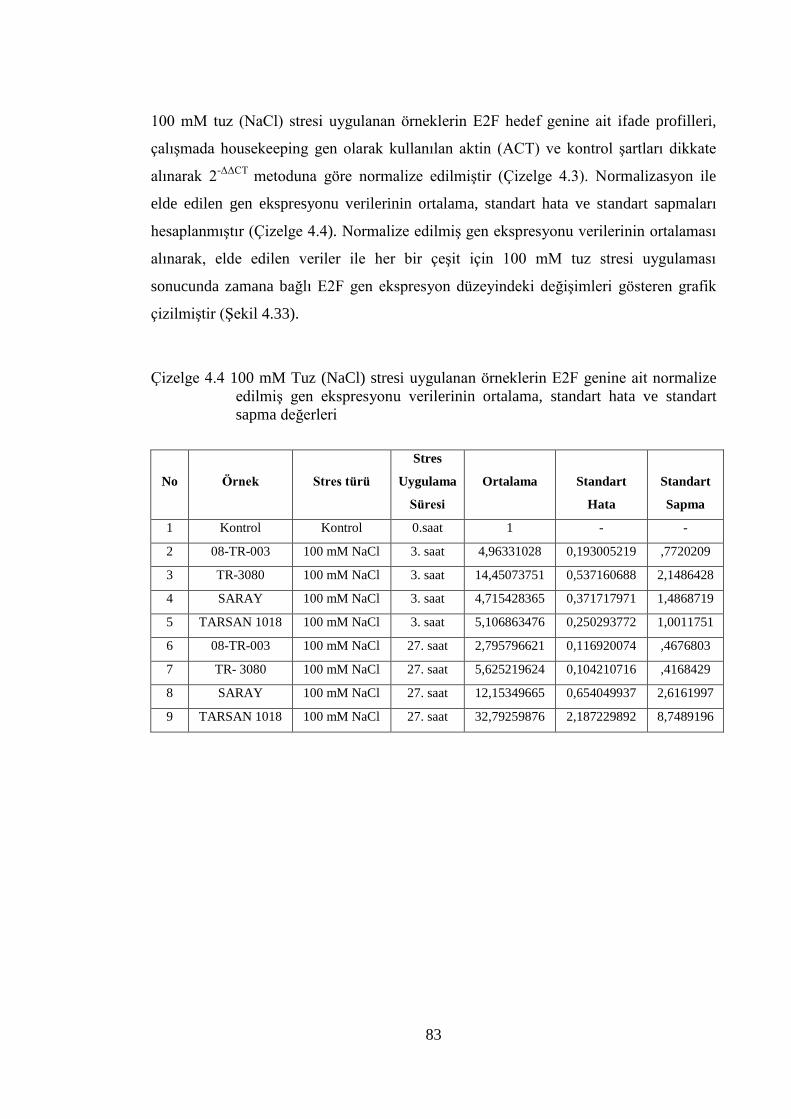
83
100 mM tuz (NaCl) stresi uygulanan örneklerin E2F hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.3). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.4). Normalize edilmiş gen ekspresyonu verilerinin ortalaması
alınarak, elde edilen veriler ile her bir çeşit için 100 mM tuz stresi uygulaması
sonucunda zamana bağlı E2F gen ekspresyon düzeyindeki değişimleri gösteren grafik
çizilmiştir (Şekil 4.33).
Çizelge 4.4 100 mM Tuz (NaCl) stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM NaCl 3. saat 4,96331028 0,193005219 ,7720209
3 TR-3080 100 mM NaCl 3. saat 14,45073751 0,537160688 2,1486428
4 SARAY 100 mM NaCl 3. saat 4,715428365 0,371717971 1,4868719
5 TARSAN 1018 100 mM NaCl 3. saat 5,106863476 0,250293772 1,0011751
6 08-TR-003 100 mM NaCl 27. saat 2,795796621 0,116920074 ,4676803
7 TR- 3080 100 mM NaCl 27. saat 5,625219624 0,104210716 ,4168429
8 SARAY 100 mM NaCl 27. saat 12,15349665 0,654049937 2,6161997
9 TARSAN 1018 100 mM NaCl 27. saat 32,79259876 2,187229892 8,7489196
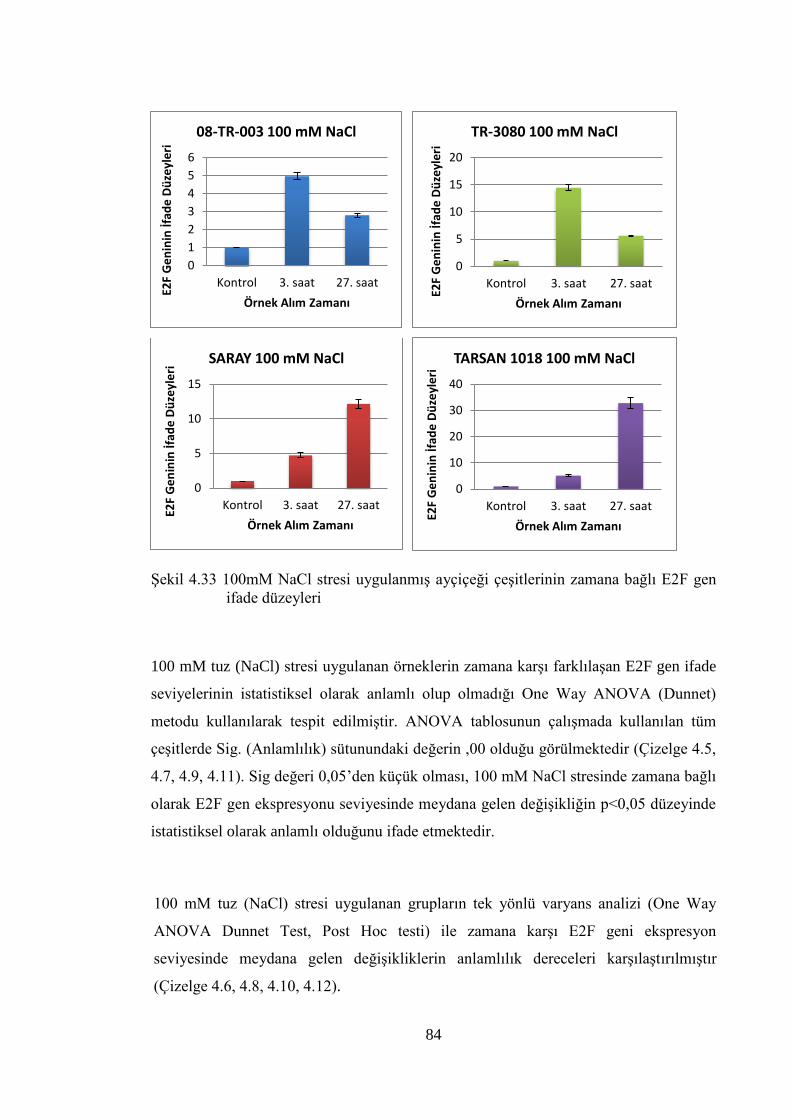
84
Şekil 4.33 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı E2F gen
ifade düzeyleri
100 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge 4.5,
4.7, 4.9, 4.11). Sig değeri 0,05’den küçük olması, 100 mM NaCl stresinde zamana bağlı
olarak E2F gen ekspresyonu seviyesinde meydana gelen değişikliğin p<0,05 düzeyinde
istatistiksel olarak anlamlı olduğunu ifade etmektedir.
100 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı E2F geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.6, 4.8, 4.10, 4.12).
0
1
2
3
4
5
6
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 100 mM NaCl
0
5
10
15
20
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 100 mM NaCl
0
5
10
15
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
SARAY 100 mM NaCl
0
10
20
30
40
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 100 mM NaCl
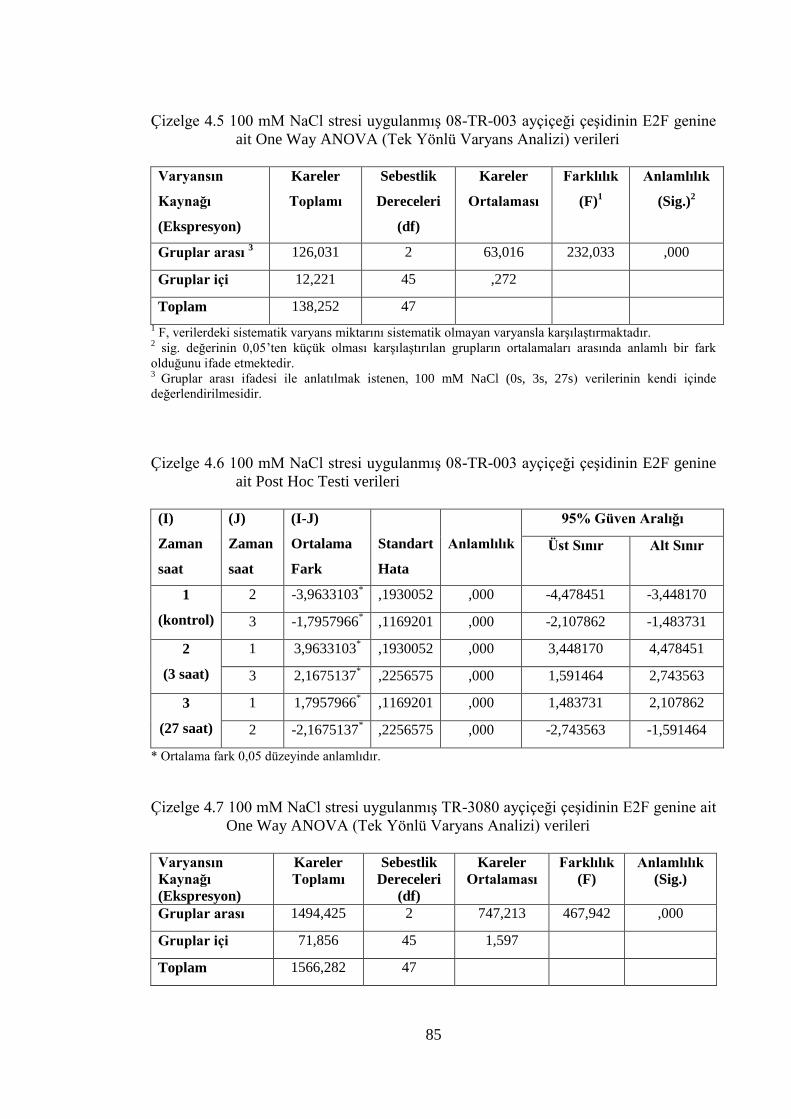
85
Çizelge 4.5 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 126,031 2 63,016 232,033 ,000
Gruplar içi 12,221 45 ,272
Toplam 138,252 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3
Gruplar arası ifadesi ile anlatılmak istenen, 100 mM NaCl (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.6 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -3,9633103* ,1930052 ,000 -4,478451 -3,448170
3 -1,7957966* ,1169201 ,000 -2,107862 -1,483731
2
(3 saat)
1 3,9633103* ,1930052 ,000 3,448170 4,478451
3 2,1675137* ,2256575 ,000 1,591464 2,743563
3
(27 saat)
1 1,7957966* ,1169201 ,000 1,483731 2,107862
2 -2,1675137* ,2256575 ,000 -2,743563 -1,591464
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.7 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 1494,425 2 747,213 467,942 ,000
Gruplar içi 71,856 45 1,597
Toplam 1566,282 47
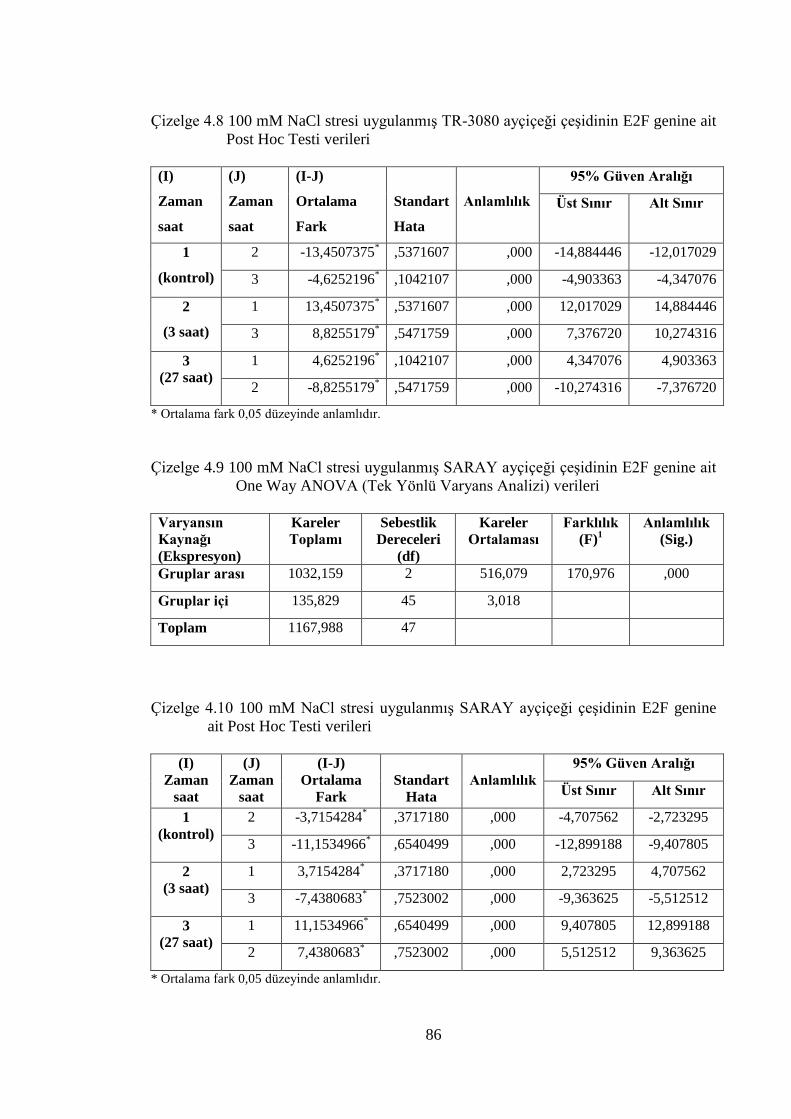
86
Çizelge 4.8 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine ait
Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -13,4507375* ,5371607 ,000 -14,884446 -12,017029
3 -4,6252196* ,1042107 ,000 -4,903363 -4,347076
2
(3 saat)
1 13,4507375* ,5371607 ,000 12,017029 14,884446
3 8,8255179* ,5471759 ,000 7,376720 10,274316
3
(27 saat)
1 4,6252196* ,1042107 ,000 4,347076 4,903363
2 -8,8255179* ,5471759 ,000 -10,274316 -7,376720
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.9 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)
Gruplar arası 1032,159 2 516,079 170,976 ,000
Gruplar içi 135,829 45 3,018
Toplam 1167,988 47
Çizelge 4.10 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -3,7154284* ,3717180 ,000 -4,707562 -2,723295
3 -11,1534966* ,6540499 ,000 -12,899188 -9,407805
2
(3 saat)
1 3,7154284* ,3717180 ,000 2,723295 4,707562
3 -7,4380683* ,7523002 ,000 -9,363625 -5,512512
3
(27 saat)
1 11,1534966* ,6540499 ,000 9,407805 12,899188
2 7,4380683* ,7523002 ,000 5,512512 9,363625
* Ortalama fark 0,05 düzeyinde anlamlıdır.
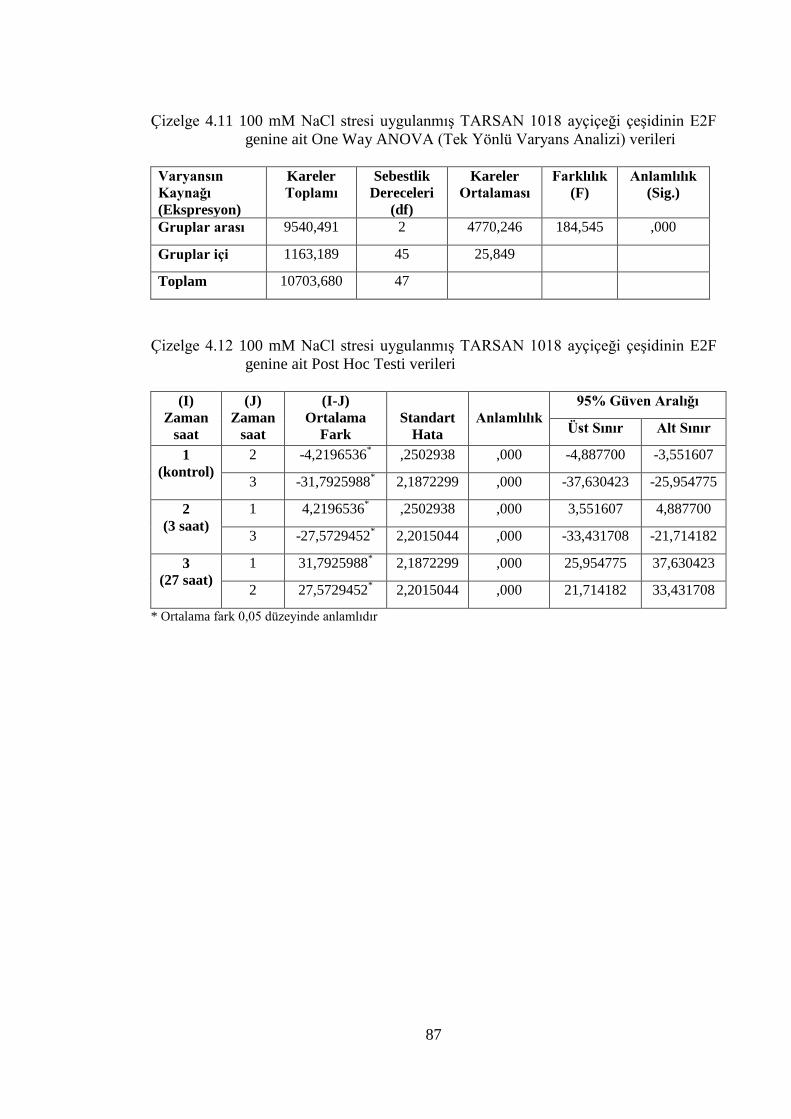
87
Çizelge 4.11 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 9540,491 2 4770,246 184,545 ,000
Gruplar içi 1163,189 45 25,849
Toplam 10703,680 47
Çizelge 4.12 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -4,2196536* ,2502938 ,000 -4,887700 -3,551607
3 -31,7925988* 2,1872299 ,000 -37,630423 -25,954775
2
(3 saat)
1 4,2196536* ,2502938 ,000 3,551607 4,887700
3 -27,5729452* 2,2015044 ,000 -33,431708 -21,714182
3
(27 saat)
1 31,7925988* 2,1872299 ,000 25,954775 37,630423
2 27,5729452* 2,2015044 ,000 21,714182 33,431708
* Ortalama fark 0,05 düzeyinde anlamlıdır
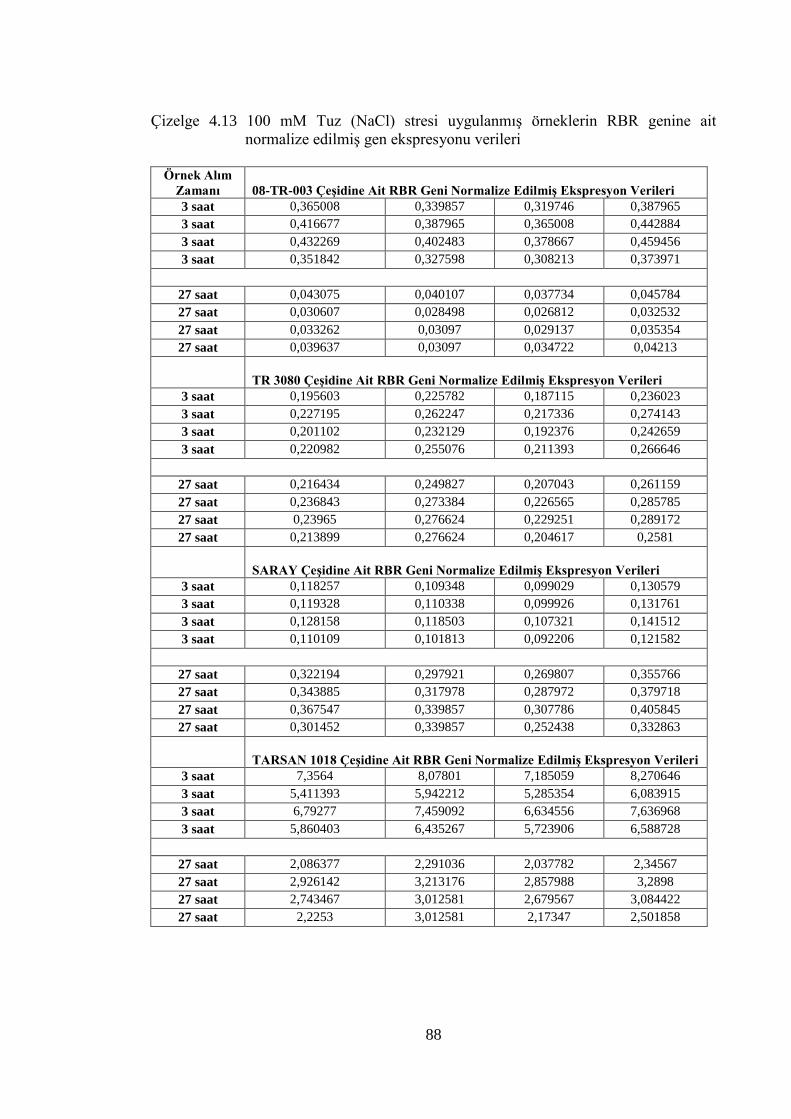
88
Çizelge 4.13 100 mM Tuz (NaCl) stresi uygulanmış örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,365008 0,339857 0,319746 0,387965
3 saat 0,416677 0,387965 0,365008 0,442884
3 saat 0,432269 0,402483 0,378667 0,459456
3 saat 0,351842 0,327598 0,308213 0,373971
27 saat 0,043075 0,040107 0,037734 0,045784
27 saat 0,030607 0,028498 0,026812 0,032532
27 saat 0,033262 0,03097 0,029137 0,035354
27 saat 0,039637 0,03097 0,034722 0,04213
TR 3080 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,195603 0,225782 0,187115 0,236023
3 saat 0,227195 0,262247 0,217336 0,274143
3 saat 0,201102 0,232129 0,192376 0,242659
3 saat 0,220982 0,255076 0,211393 0,266646
27 saat 0,216434 0,249827 0,207043 0,261159
27 saat 0,236843 0,273384 0,226565 0,285785
27 saat 0,23965 0,276624 0,229251 0,289172
27 saat 0,213899 0,276624 0,204617 0,2581
SARAY Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,118257 0,109348 0,099029 0,130579
3 saat 0,119328 0,110338 0,099926 0,131761
3 saat 0,128158 0,118503 0,107321 0,141512
3 saat 0,110109 0,101813 0,092206 0,121582
27 saat 0,322194 0,297921 0,269807 0,355766
27 saat 0,343885 0,317978 0,287972 0,379718
27 saat 0,367547 0,339857 0,307786 0,405845
27 saat 0,301452 0,339857 0,252438 0,332863
TARSAN 1018 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 7,3564 8,07801 7,185059 8,270646
3 saat 5,411393 5,942212 5,285354 6,083915
3 saat 6,79277 7,459092 6,634556 7,636968
3 saat 5,860403 6,435267 5,723906 6,588728
27 saat 2,086377 2,291036 2,037782 2,34567
27 saat 2,926142 3,213176 2,857988 3,2898
27 saat 2,743467 3,012581 2,679567 3,084422
27 saat 2,2253 3,012581 2,17347 2,501858
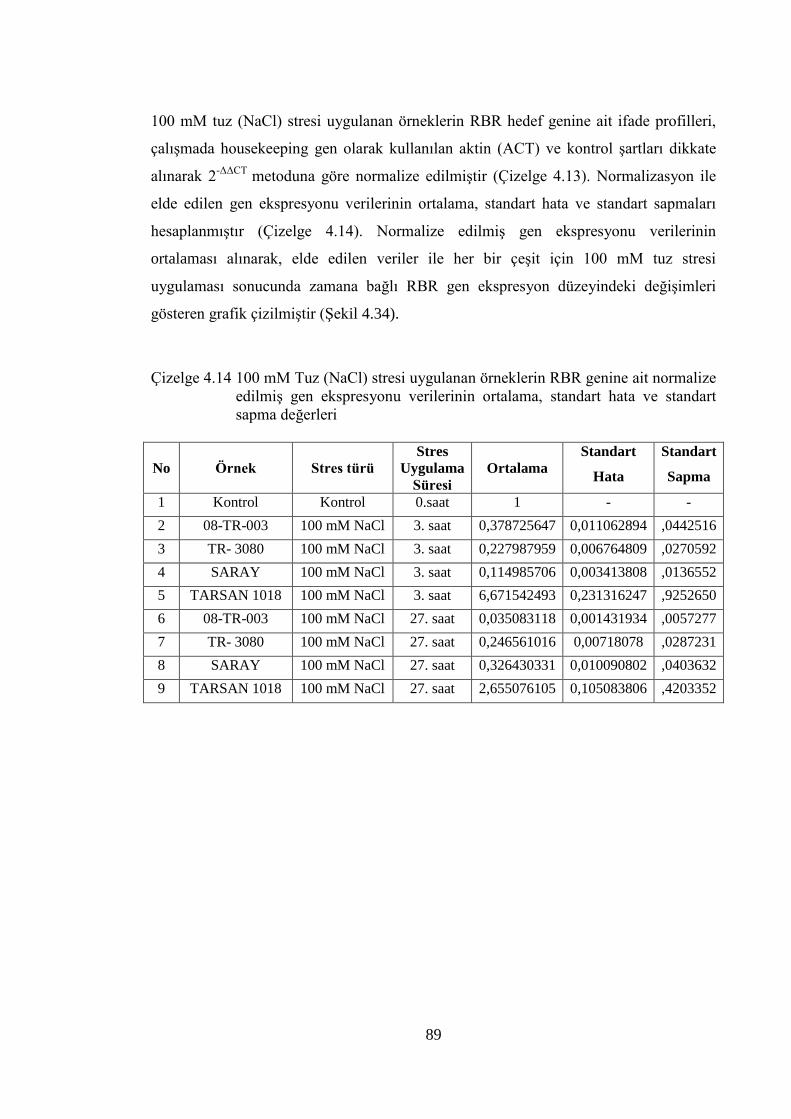
89
100 mM tuz (NaCl) stresi uygulanan örneklerin RBR hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.13). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.14). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 100 mM tuz stresi
uygulaması sonucunda zamana bağlı RBR gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.34).
Çizelge 4.14 100 mM Tuz (NaCl) stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM NaCl 3. saat 0,378725647 0,011062894 ,0442516
3 TR- 3080 100 mM NaCl 3. saat 0,227987959 0,006764809 ,0270592
4 SARAY 100 mM NaCl 3. saat 0,114985706 0,003413808 ,0136552
5 TARSAN 1018 100 mM NaCl 3. saat 6,671542493 0,231316247 ,9252650
6 08-TR-003 100 mM NaCl 27. saat 0,035083118 0,001431934 ,0057277
7 TR- 3080 100 mM NaCl 27. saat 0,246561016 0,00718078 ,0287231
8 SARAY 100 mM NaCl 27. saat 0,326430331 0,010090802 ,0403632
9 TARSAN 1018 100 mM NaCl 27. saat 2,655076105 0,105083806 ,4203352
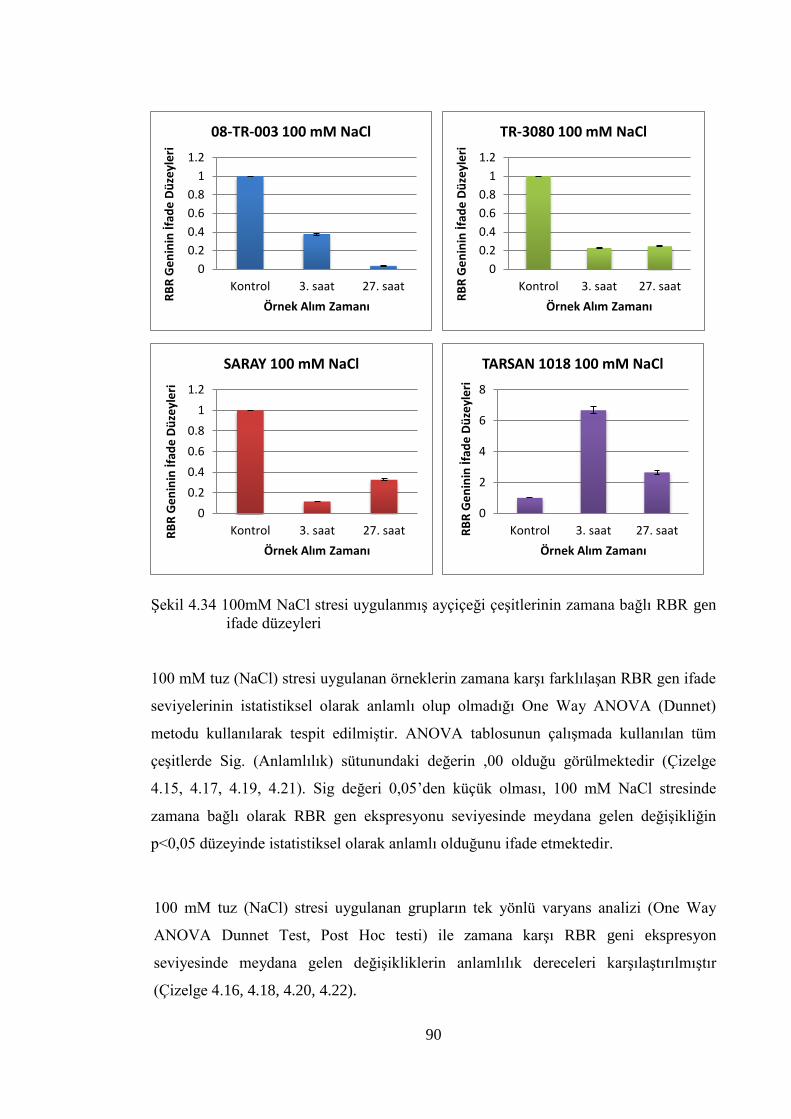
90
Şekil 4.34 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı RBR gen
ifade düzeyleri
100 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan RBR gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.15, 4.17, 4.19, 4.21). Sig değeri 0,05’den küçük olması, 100 mM NaCl stresinde
zamana bağlı olarak RBR gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
100 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı RBR geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.16, 4.18, 4.20, 4.22).
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
08-TR-003 100 mM NaCl
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 100 mM NaCl
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
SARAY 100 mM NaCl
0
2
4
6
8
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018 100 mM NaCl
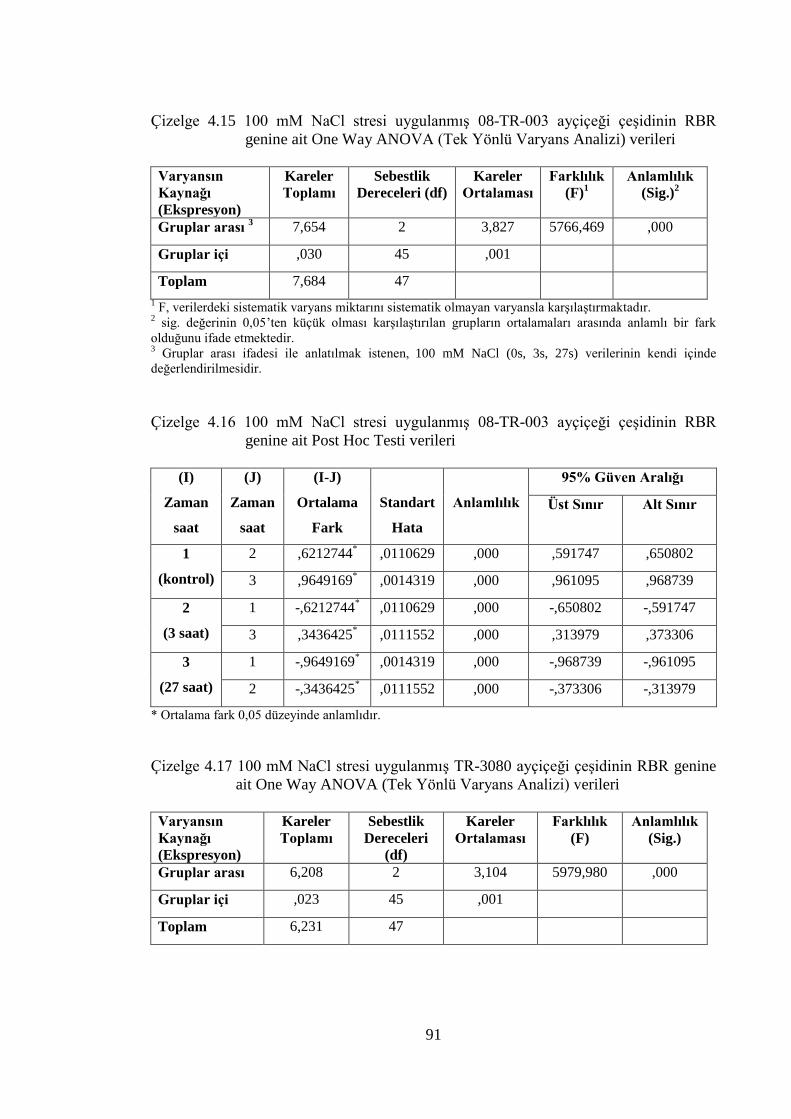
91
Çizelge 4.15 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri (df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 7,654 2 3,827 5766,469 ,000
Gruplar içi ,030 45 ,001
Toplam 7,684 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM NaCl (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.16 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,6212744* ,0110629 ,000 ,591747 ,650802
3 ,9649169* ,0014319 ,000 ,961095 ,968739
2
(3 saat)
1 -,6212744* ,0110629 ,000 -,650802 -,591747
3 ,3436425* ,0111552 ,000 ,313979 ,373306
3
(27 saat)
1 -,9649169* ,0014319 ,000 -,968739 -,961095
2 -,3436425* ,0111552 ,000 -,373306 -,313979
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.17 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 6,208 2 3,104 5979,980 ,000
Gruplar içi ,023 45 ,001
Toplam 6,231 47
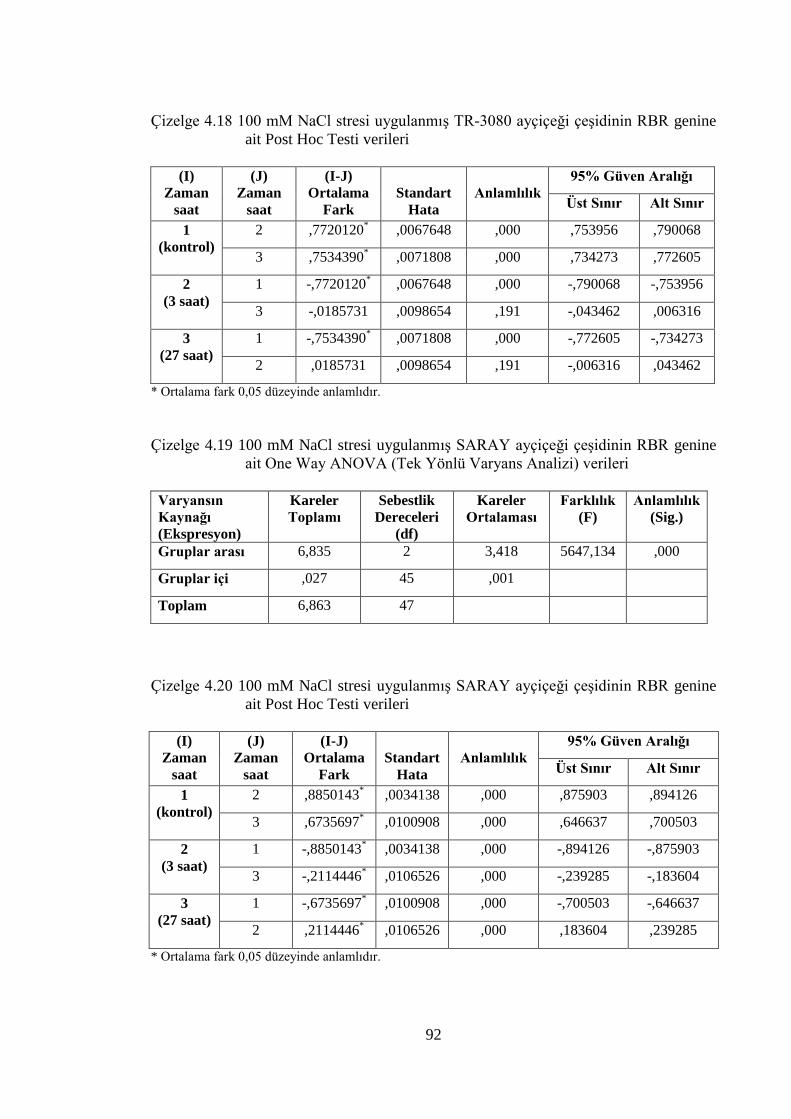
92
Çizelge 4.18 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,7720120* ,0067648 ,000 ,753956 ,790068
3 ,7534390* ,0071808 ,000 ,734273 ,772605
2
(3 saat)
1 -,7720120* ,0067648 ,000 -,790068 -,753956
3 -,0185731 ,0098654 ,191 -,043462 ,006316
3
(27 saat)
1 -,7534390* ,0071808 ,000 -,772605 -,734273
2 ,0185731 ,0098654 ,191 -,006316 ,043462
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.19 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 6,835 2 3,418 5647,134 ,000
Gruplar içi ,027 45 ,001
Toplam 6,863 47
Çizelge 4.20 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8850143* ,0034138 ,000 ,875903 ,894126
3 ,6735697* ,0100908 ,000 ,646637 ,700503
2
(3 saat)
1 -,8850143* ,0034138 ,000 -,894126 -,875903
3 -,2114446* ,0106526 ,000 -,239285 -,183604
3
(27 saat)
1 -,6735697* ,0100908 ,000 -,700503 -,646637
2 ,2114446* ,0106526 ,000 ,183604 ,239285
* Ortalama fark 0,05 düzeyinde anlamlıdır.
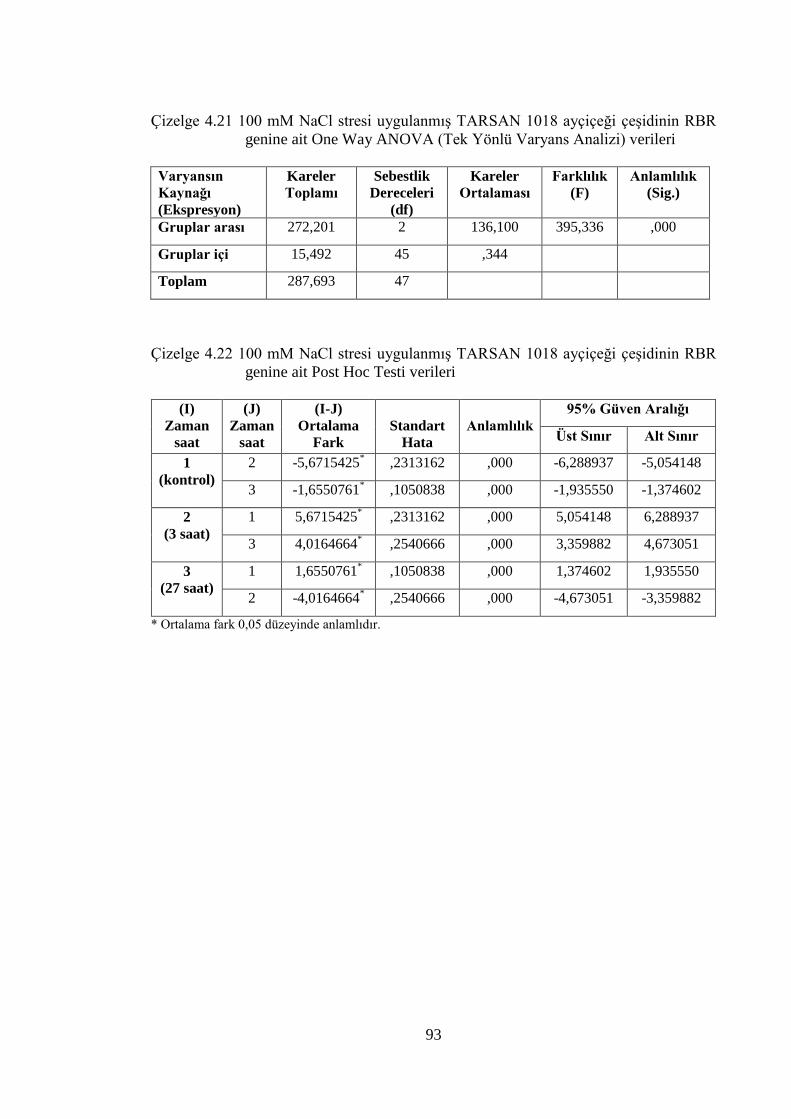
93
Çizelge 4.21 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 272,201 2 136,100 395,336 ,000
Gruplar içi 15,492 45 ,344
Toplam 287,693 47
Çizelge 4.22 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -5,6715425* ,2313162 ,000 -6,288937 -5,054148
3 -1,6550761* ,1050838 ,000 -1,935550 -1,374602
2
(3 saat)
1 5,6715425* ,2313162 ,000 5,054148 6,288937
3 4,0164664* ,2540666 ,000 3,359882 4,673051
3
(27 saat)
1 1,6550761* ,1050838 ,000 1,374602 1,935550
2 -4,0164664* ,2540666 ,000 -4,673051 -3,359882
* Ortalama fark 0,05 düzeyinde anlamlıdır.
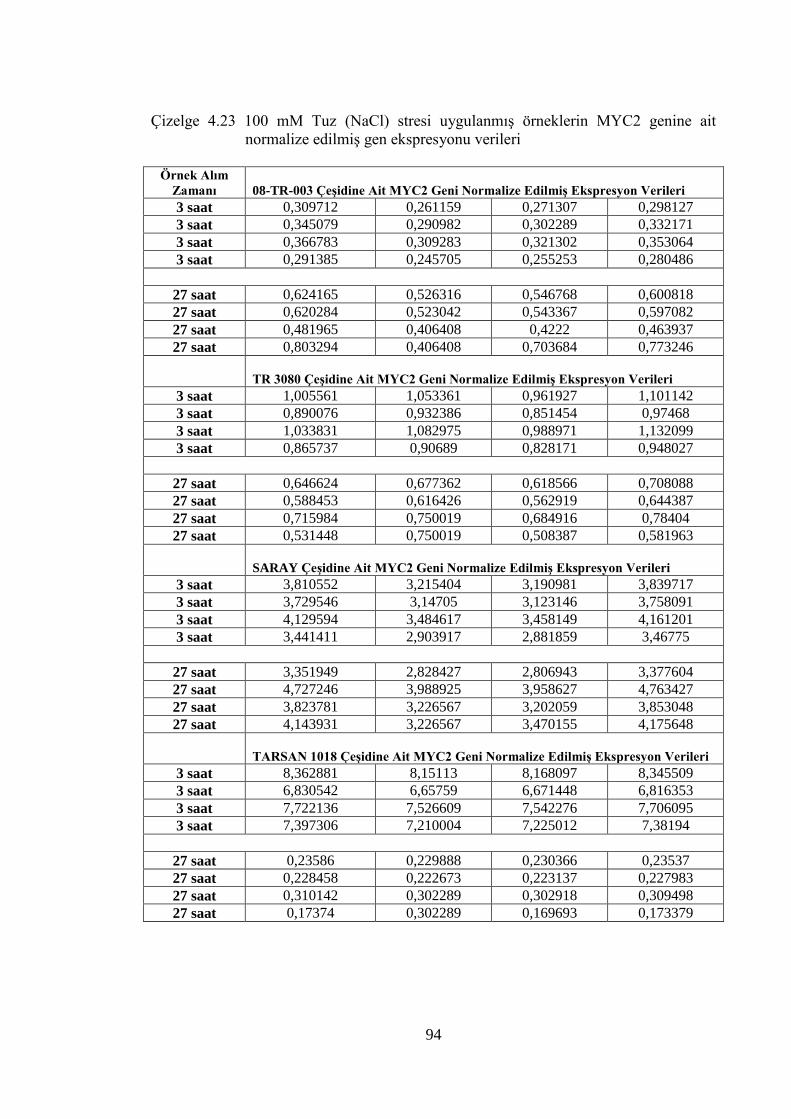
94
Çizelge 4.23 100 mM Tuz (NaCl) stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,309712 0,261159 0,271307 0,298127
3 saat 0,345079 0,290982 0,302289 0,332171
3 saat 0,366783 0,309283 0,321302 0,353064
3 saat 0,291385 0,245705 0,255253 0,280486
27 saat 0,624165 0,526316 0,546768 0,600818
27 saat 0,620284 0,523042 0,543367 0,597082
27 saat 0,481965 0,406408 0,4222 0,463937
27 saat 0,803294 0,406408 0,703684 0,773246
TR 3080 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,005561 1,053361 0,961927 1,101142
3 saat 0,890076 0,932386 0,851454 0,97468
3 saat 1,033831 1,082975 0,988971 1,132099
3 saat 0,865737 0,90689 0,828171 0,948027
27 saat 0,646624 0,677362 0,618566 0,708088
27 saat 0,588453 0,616426 0,562919 0,644387
27 saat 0,715984 0,750019 0,684916 0,78404
27 saat 0,531448 0,750019 0,508387 0,581963
SARAY Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 3,810552 3,215404 3,190981 3,839717
3 saat 3,729546 3,14705 3,123146 3,758091
3 saat 4,129594 3,484617 3,458149 4,161201
3 saat 3,441411 2,903917 2,881859 3,46775
27 saat 3,351949 2,828427 2,806943 3,377604
27 saat 4,727246 3,988925 3,958627 4,763427
27 saat 3,823781 3,226567 3,202059 3,853048
27 saat 4,143931 3,226567 3,470155 4,175648
TARSAN 1018 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 8,362881 8,15113 8,168097 8,345509
3 saat 6,830542 6,65759 6,671448 6,816353
3 saat 7,722136 7,526609 7,542276 7,706095
3 saat 7,397306 7,210004 7,225012 7,38194
27 saat 0,23586 0,229888 0,230366 0,23537
27 saat 0,228458 0,222673 0,223137 0,227983
27 saat 0,310142 0,302289 0,302918 0,309498
27 saat 0,17374 0,302289 0,169693 0,173379
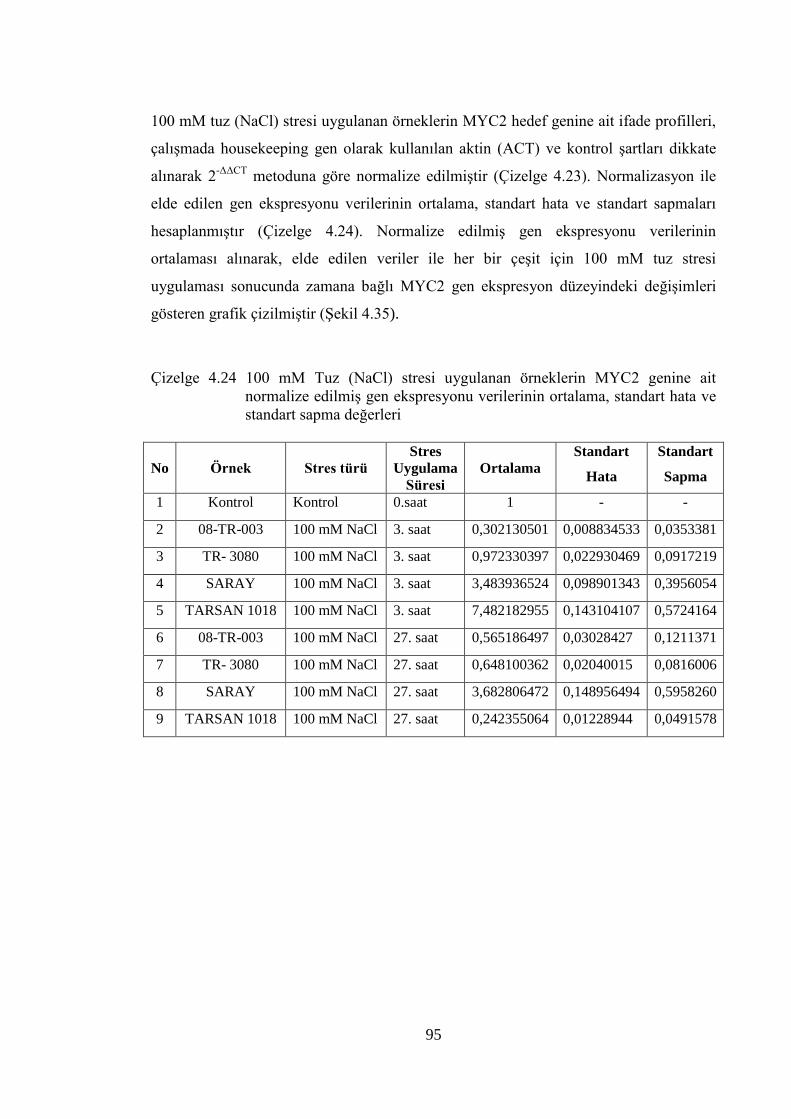
95
100 mM tuz (NaCl) stresi uygulanan örneklerin MYC2 hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.23). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.24). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 100 mM tuz stresi
uygulaması sonucunda zamana bağlı MYC2 gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.35).
Çizelge 4.24 100 mM Tuz (NaCl) stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM NaCl 3. saat 0,302130501 0,008834533 0,0353381
3 TR- 3080 100 mM NaCl 3. saat 0,972330397 0,022930469 0,0917219
4 SARAY 100 mM NaCl 3. saat 3,483936524 0,098901343 0,3956054
5 TARSAN 1018 100 mM NaCl 3. saat 7,482182955 0,143104107 0,5724164
6 08-TR-003 100 mM NaCl 27. saat 0,565186497 0,03028427 0,1211371
7 TR- 3080 100 mM NaCl 27. saat 0,648100362 0,02040015 0,0816006
8 SARAY 100 mM NaCl 27. saat 3,682806472 0,148956494 0,5958260
9 TARSAN 1018 100 mM NaCl 27. saat 0,242355064 0,01228944 0,0491578
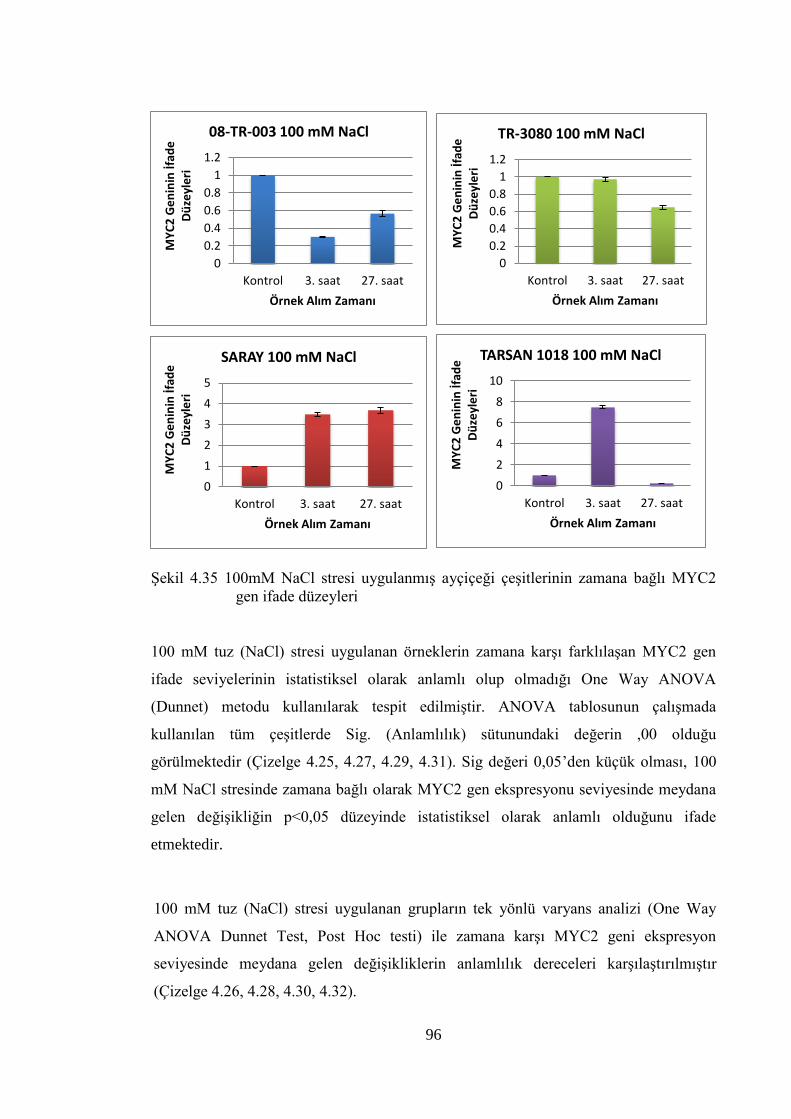
96
Şekil 4.35 100mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı MYC2
gen ifade düzeyleri
100 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan MYC2 gen
ifade seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA
(Dunnet) metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada
kullanılan tüm çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu
görülmektedir (Çizelge 4.25, 4.27, 4.29, 4.31). Sig değeri 0,05’den küçük olması, 100
mM NaCl stresinde zamana bağlı olarak MYC2 gen ekspresyonu seviyesinde meydana
gelen değişikliğin p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade
etmektedir.
100 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.26, 4.28, 4.30, 4.32).
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
08-TR-003 100 mM NaCl
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 100 mM NaCl
0
1
2
3
4
5
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
SARAY 100 mM NaCl
0
2
4
6
8
10
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018 100 mM NaCl
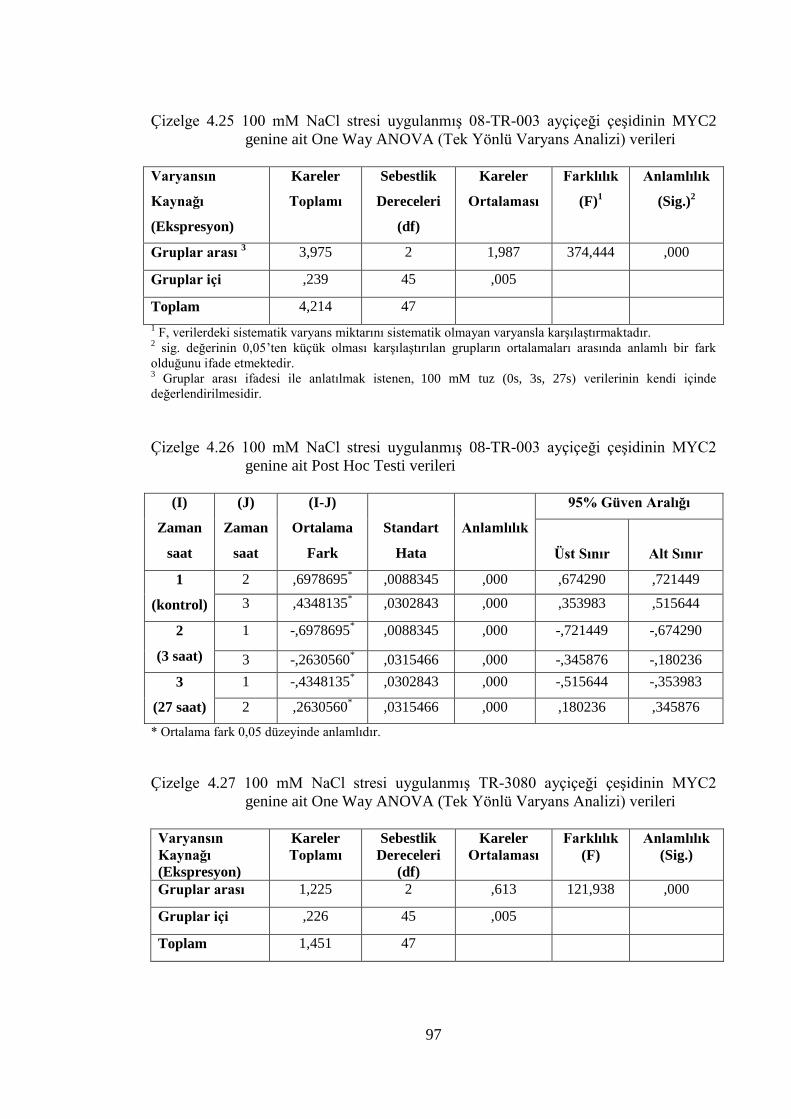
97
Çizelge 4.25 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 3,975 2 1,987 374,444 ,000
Gruplar içi ,239 45 ,005
Toplam 4,214 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM tuz (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.26 100 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır
Alt Sınır
1
(kontrol)
2 ,6978695* ,0088345 ,000 ,674290 ,721449
3 ,4348135* ,0302843 ,000 ,353983 ,515644
2
(3 saat)
1 -,6978695* ,0088345 ,000 -,721449 -,674290
3 -,2630560* ,0315466 ,000 -,345876 -,180236
3
(27 saat)
1 -,4348135* ,0302843 ,000 -,515644 -,353983
2 ,2630560* ,0315466 ,000 ,180236 ,345876
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.27 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 1,225 2 ,613 121,938 ,000
Gruplar içi ,226 45 ,005
Toplam 1,451 47
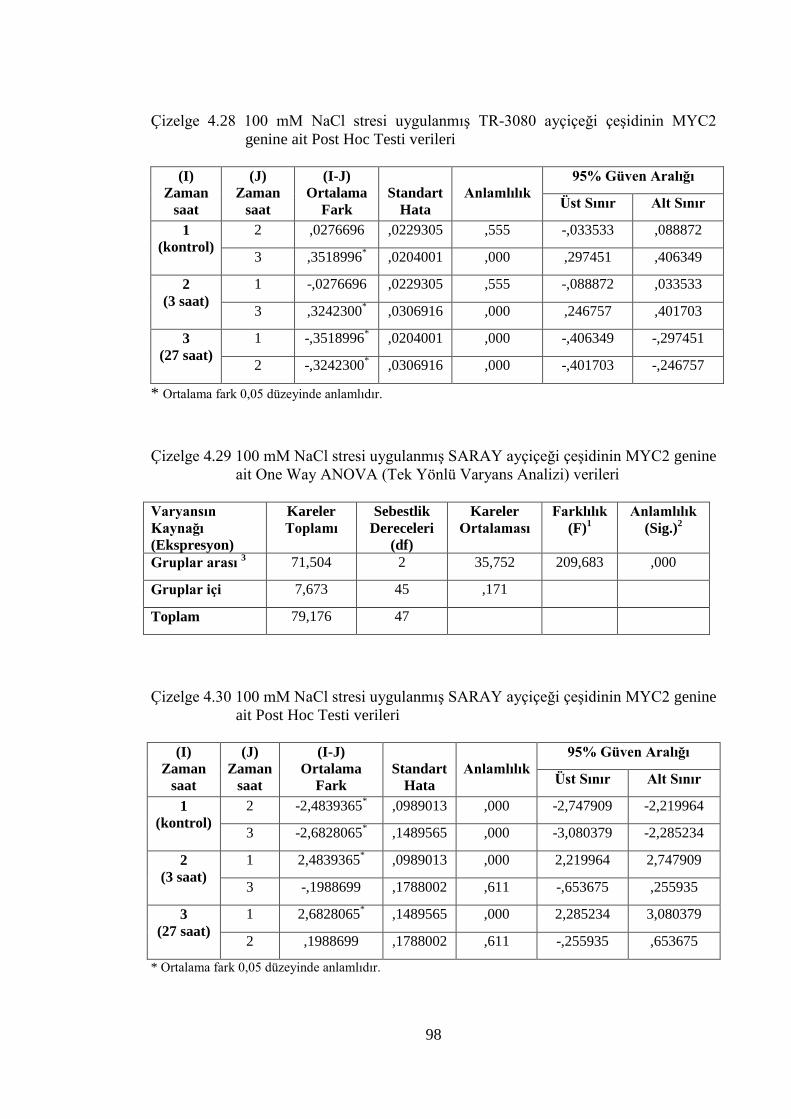
98
Çizelge 4.28 100 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,0276696 ,0229305 ,555 -,033533 ,088872
3 ,3518996* ,0204001 ,000 ,297451 ,406349
2
(3 saat)
1 -,0276696 ,0229305 ,555 -,088872 ,033533
3 ,3242300* ,0306916 ,000 ,246757 ,401703
3
(27 saat)
1 -,3518996* ,0204001 ,000 -,406349 -,297451
2 -,3242300* ,0306916 ,000 -,401703 -,246757
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.29 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 71,504 2 35,752 209,683 ,000
Gruplar içi 7,673 45 ,171
Toplam 79,176 47
Çizelge 4.30 100 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -2,4839365* ,0989013 ,000 -2,747909 -2,219964
3 -2,6828065* ,1489565 ,000 -3,080379 -2,285234
2
(3 saat)
1 2,4839365* ,0989013 ,000 2,219964 2,747909
3 -,1988699 ,1788002 ,611 -,653675 ,255935
3
(27 saat)
1 2,6828065* ,1489565 ,000 2,285234 3,080379
2 ,1988699 ,1788002 ,611 -,255935 ,653675
* Ortalama fark 0,05 düzeyinde anlamlıdır.
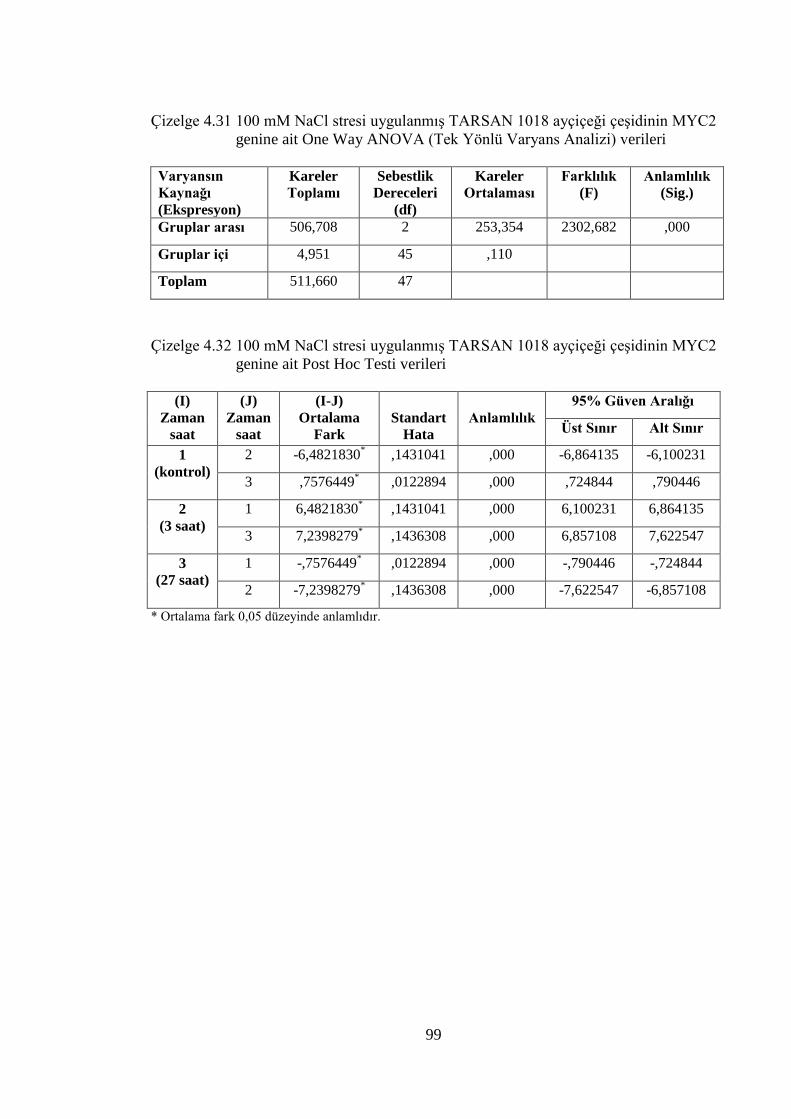
99
Çizelge 4.31 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 506,708 2 253,354 2302,682 ,000
Gruplar içi 4,951 45 ,110
Toplam 511,660 47
Çizelge 4.32 100 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -6,4821830* ,1431041 ,000 -6,864135 -6,100231
3 ,7576449* ,0122894 ,000 ,724844 ,790446
2
(3 saat)
1 6,4821830* ,1431041 ,000 6,100231 6,864135
3 7,2398279* ,1436308 ,000 6,857108 7,622547
3
(27 saat)
1 -,7576449* ,0122894 ,000 -,790446 -,724844
2 -7,2398279* ,1436308 ,000 -7,622547 -6,857108
* Ortalama fark 0,05 düzeyinde anlamlıdır.
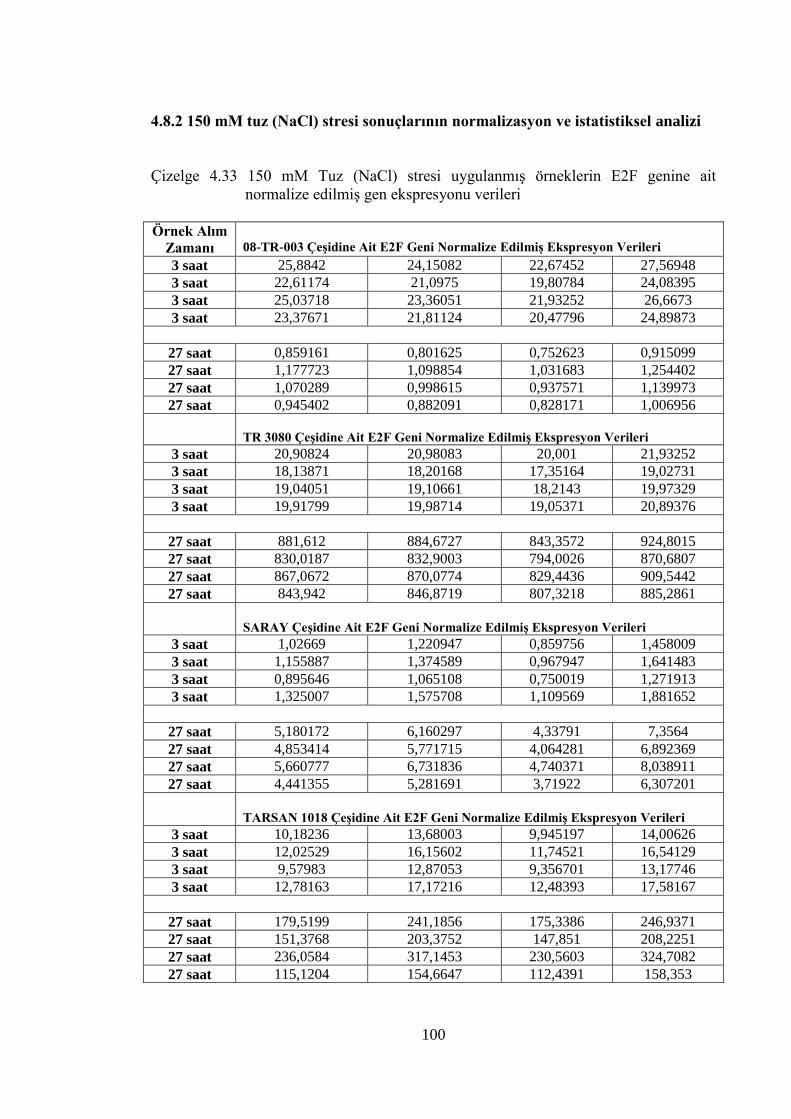
100
4.8.2 150 mM tuz (NaCl) stresi sonuçlarının normalizasyon ve istatistiksel analizi
Çizelge 4.33 150 mM Tuz (NaCl) stresi uygulanmış örneklerin E2F genine ait
normalize edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 25,8842 24,15082 22,67452 27,56948
3 saat 22,61174 21,0975 19,80784 24,08395
3 saat 25,03718 23,36051 21,93252 26,6673
3 saat 23,37671 21,81124 20,47796 24,89873
27 saat 0,859161 0,801625 0,752623 0,915099
27 saat 1,177723 1,098854 1,031683 1,254402
27 saat 1,070289 0,998615 0,937571 1,139973
27 saat 0,945402 0,882091 0,828171 1,006956
TR 3080 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 20,90824 20,98083 20,001 21,93252
3 saat 18,13871 18,20168 17,35164 19,02731
3 saat 19,04051 19,10661 18,2143 19,97329
3 saat 19,91799 19,98714 19,05371 20,89376
27 saat 881,612 884,6727 843,3572 924,8015
27 saat 830,0187 832,9003 794,0026 870,6807
27 saat 867,0672 870,0774 829,4436 909,5442
27 saat 843,942 846,8719 807,3218 885,2861
SARAY Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,02669 1,220947 0,859756 1,458009
3 saat 1,155887 1,374589 0,967947 1,641483
3 saat 0,895646 1,065108 0,750019 1,271913
3 saat 1,325007 1,575708 1,109569 1,881652
27 saat 5,180172 6,160297 4,33791 7,3564
27 saat 4,853414 5,771715 4,064281 6,892369
27 saat 5,660777 6,731836 4,740371 8,038911
27 saat 4,441355 5,281691 3,71922 6,307201
TARSAN 1018 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 10,18236 13,68003 9,945197 14,00626
3 saat 12,02529 16,15602 11,74521 16,54129
3 saat 9,57983 12,87053 9,356701 13,17746
3 saat 12,78163 17,17216 12,48393 17,58167
27 saat 179,5199 241,1856 175,3386 246,9371
27 saat 151,3768 203,3752 147,851 208,2251
27 saat 236,0584 317,1453 230,5603 324,7082
27 saat 115,1204 154,6647 112,4391 158,353
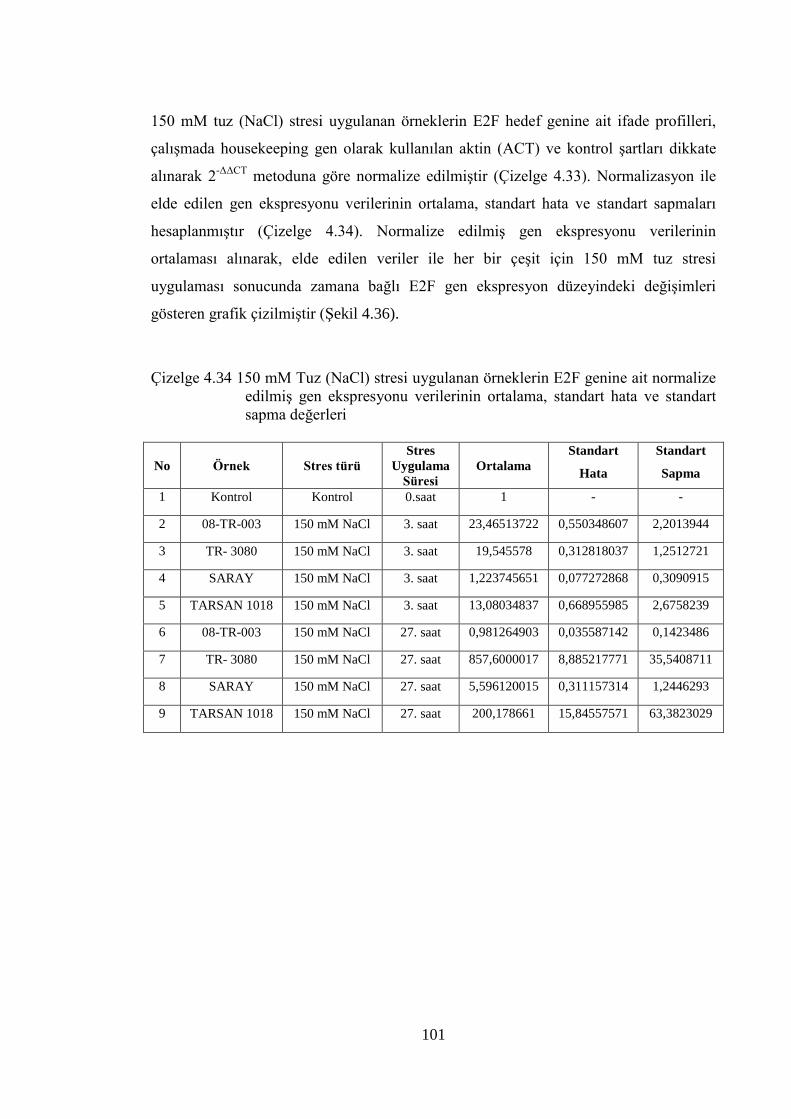
101
150 mM tuz (NaCl) stresi uygulanan örneklerin E2F hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.33). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.34). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM tuz stresi
uygulaması sonucunda zamana bağlı E2F gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.36).
Çizelge 4.34 150 mM Tuz (NaCl) stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM NaCl 3. saat 23,46513722 0,550348607 2,2013944
3 TR- 3080 150 mM NaCl 3. saat 19,545578 0,312818037 1,2512721
4 SARAY 150 mM NaCl 3. saat 1,223745651 0,077272868 0,3090915
5 TARSAN 1018 150 mM NaCl 3. saat 13,08034837 0,668955985 2,6758239
6 08-TR-003 150 mM NaCl 27. saat 0,981264903 0,035587142 0,1423486
7 TR- 3080 150 mM NaCl 27. saat 857,6000017 8,885217771 35,5408711
8 SARAY 150 mM NaCl 27. saat 5,596120015 0,311157314 1,2446293
9 TARSAN 1018 150 mM NaCl 27. saat 200,178661 15,84557571 63,3823029
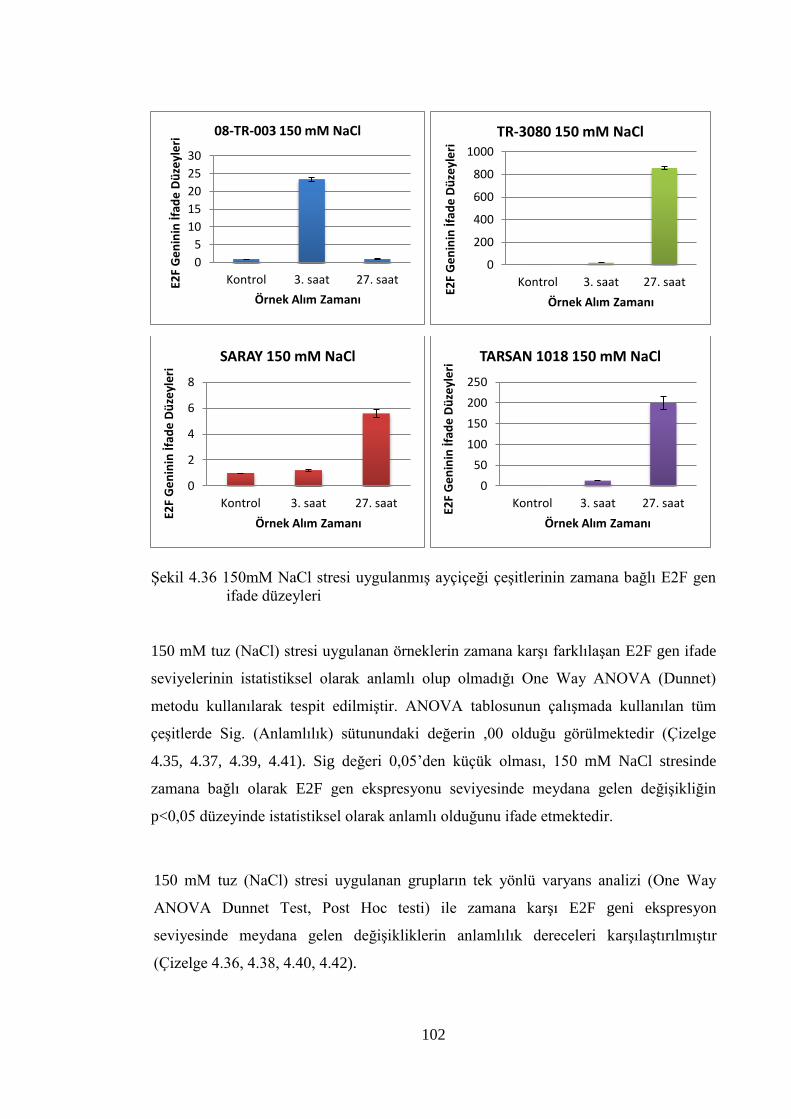
102
Şekil 4.36 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı E2F gen
ifade düzeyleri
150 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.35, 4.37, 4.39, 4.41). Sig değeri 0,05’den küçük olması, 150 mM NaCl stresinde
zamana bağlı olarak E2F gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
150 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı E2F geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.36, 4.38, 4.40, 4.42).
0
5
10
15
20
25
30
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 150 mM NaCl
0
200
400
600
800
1000
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 150 mM NaCl
0
2
4
6
8
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
SARAY 150 mM NaCl
0
50
100
150
200
250
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 150 mM NaCl

103
Çizelge 4.35 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 5387,772 2 2693,886 1660,705 ,000
Gruplar içi 72,996 45 1,622
Toplam 5460,768 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM Tuz (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.36 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi Verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -22,4651372* ,5503486 ,000 -23,934045 -20,996230
3 ,0187351 ,0355871 ,934 -,076249 ,113719
2
(3 saat)
1 22,4651372* ,5503486 ,000 20,996230 23,934045
3 22,4838723* ,5514980 ,000 21,013296 23,954448
3
(27 saat)
1 -,0187351 ,0355871 ,934 -,113719 ,076249
2 -22,4838723* ,5514980 ,000 -23,954448 -21,013296
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.37 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar
arası
7661027,833 2 3830513,916 9086,240 ,000
Gruplar içi 18970,788 45 421,573
Toplam 7679998,621 47
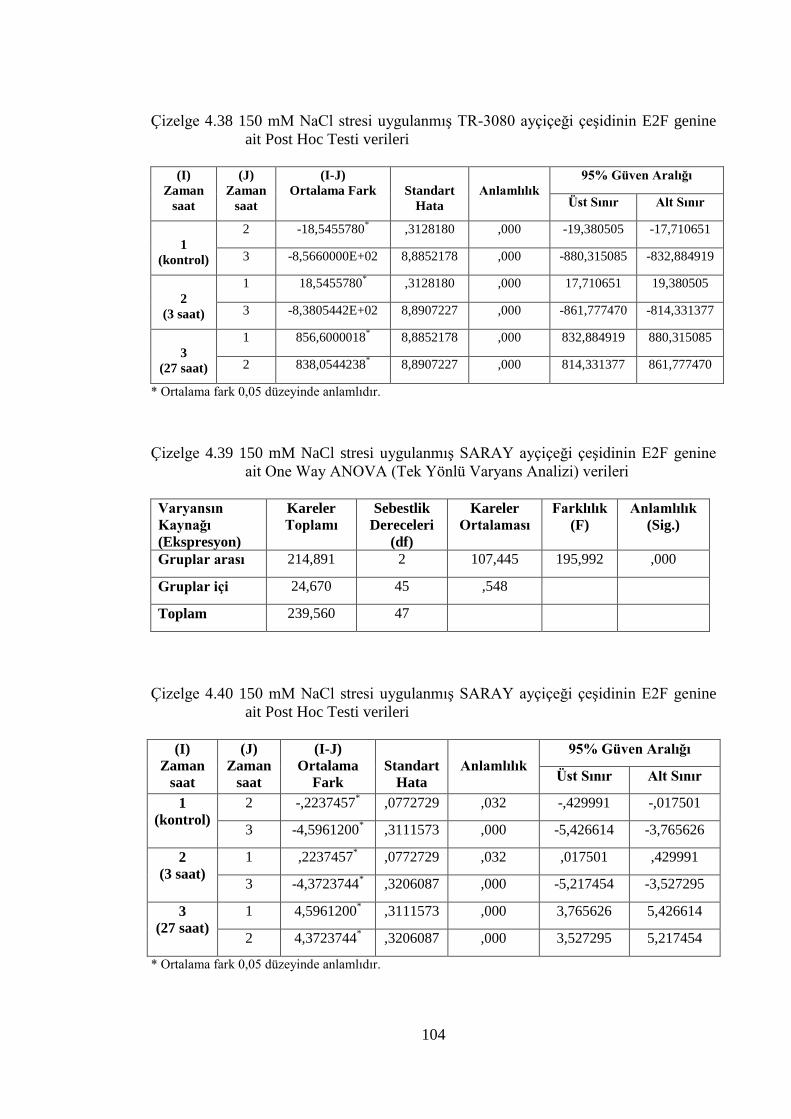
104
Çizelge 4.38 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -18,5455780* ,3128180 ,000 -19,380505 -17,710651
3 -8,5660000E+02 8,8852178 ,000 -880,315085 -832,884919
2
(3 saat)
1 18,5455780* ,3128180 ,000 17,710651 19,380505
3 -8,3805442E+02 8,8907227 ,000 -861,777470 -814,331377
3
(27 saat)
1 856,6000018* 8,8852178 ,000 832,884919 880,315085
2 838,0544238* 8,8907227 ,000 814,331377 861,777470
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.39 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 214,891 2 107,445 195,992 ,000
Gruplar içi 24,670 45 ,548
Toplam 239,560 47
Çizelge 4.40 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,2237457* ,0772729 ,032 -,429991 -,017501
3 -4,5961200* ,3111573 ,000 -5,426614 -3,765626
2
(3 saat)
1 ,2237457* ,0772729 ,032 ,017501 ,429991
3 -4,3723744* ,3206087 ,000 -5,217454 -3,527295
3
(27 saat)
1 4,5961200* ,3111573 ,000 3,765626 5,426614
2 4,3723744* ,3206087 ,000 3,527295 5,217454
* Ortalama fark 0,05 düzeyinde anlamlıdır.
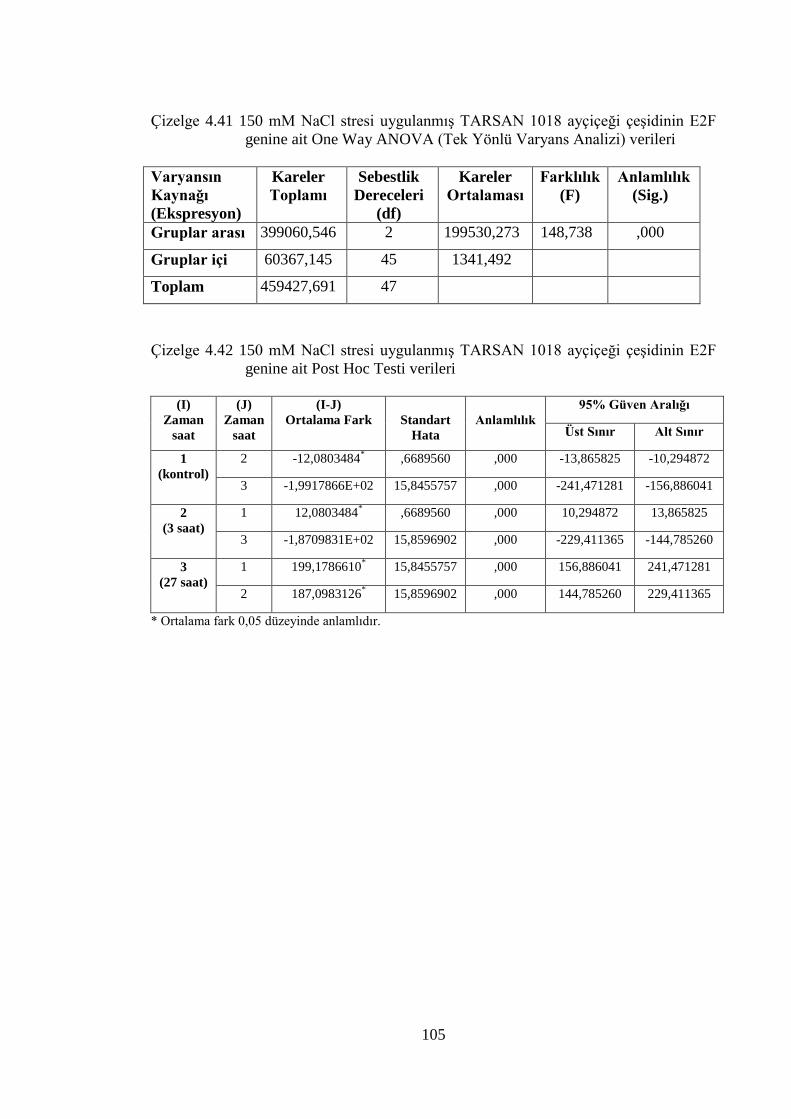
105
Çizelge 4.41 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 399060,546 2 199530,273 148,738 ,000
Gruplar içi 60367,145 45 1341,492
Toplam 459427,691 47
Çizelge 4.42 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -12,0803484* ,6689560 ,000 -13,865825 -10,294872
3 -1,9917866E+02 15,8455757 ,000 -241,471281 -156,886041
2
(3 saat)
1 12,0803484* ,6689560 ,000 10,294872 13,865825
3 -1,8709831E+02 15,8596902 ,000 -229,411365 -144,785260
3
(27 saat)
1 199,1786610* 15,8455757 ,000 156,886041 241,471281
2 187,0983126* 15,8596902 ,000 144,785260 229,411365
* Ortalama fark 0,05 düzeyinde anlamlıdır.
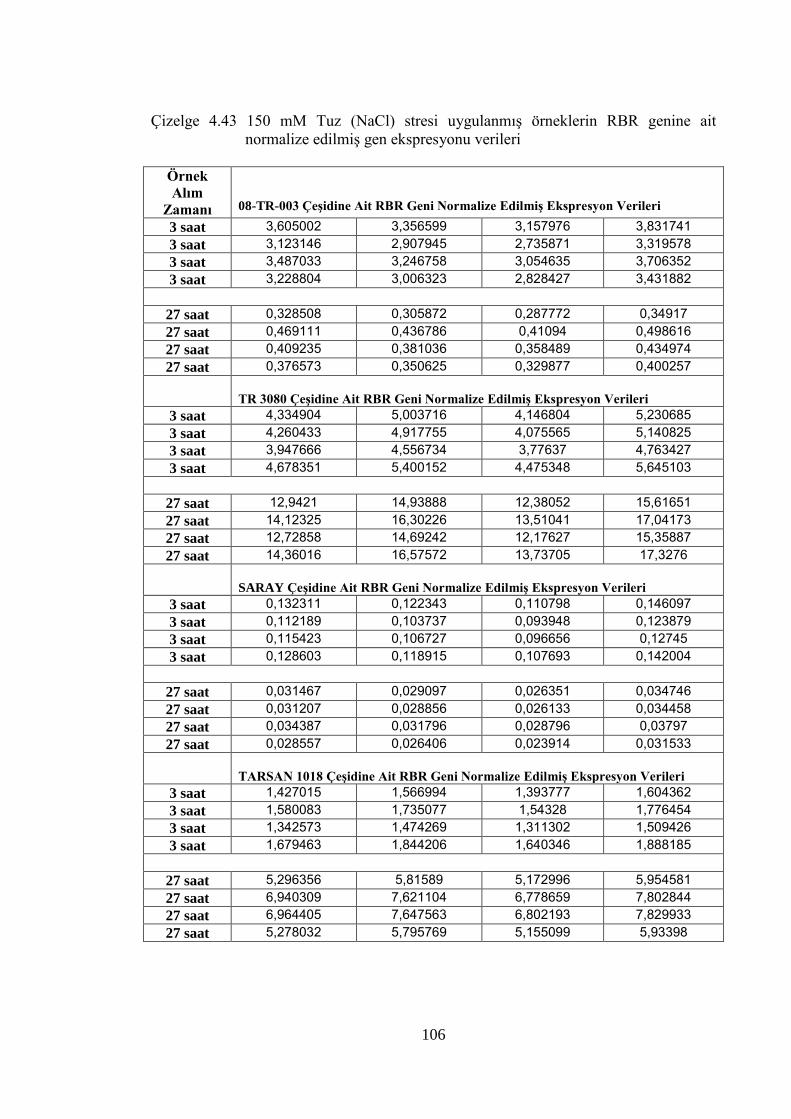
106
Çizelge 4.43 150 mM Tuz (NaCl) stresi uygulanmış örneklerin RBR genine ait
normalize edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 3,605002 3,356599 3,157976 3,831741
3 saat 3,123146 2,907945 2,735871 3,319578
3 saat 3,487033 3,246758 3,054635 3,706352
3 saat 3,228804 3,006323 2,828427 3,431882
27 saat 0,328508 0,305872 0,287772 0,34917
27 saat 0,469111 0,436786 0,41094 0,498616
27 saat 0,409235 0,381036 0,358489 0,434974
27 saat 0,376573 0,350625 0,329877 0,400257
TR 3080 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 4,334904 5,003716 4,146804 5,230685
3 saat 4,260433 4,917755 4,075565 5,140825
3 saat 3,947666 4,556734 3,77637 4,763427
3 saat 4,678351 5,400152 4,475348 5,645103
27 saat 12,9421 14,93888 12,38052 15,61651
27 saat 14,12325 16,30226 13,51041 17,04173
27 saat 12,72858 14,69242 12,17627 15,35887
27 saat 14,36016 16,57572 13,73705 17,3276
SARAY Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,132311 0,122343 0,110798 0,146097
3 saat 0,112189 0,103737 0,093948 0,123879
3 saat 0,115423 0,106727 0,096656 0,12745
3 saat 0,128603 0,118915 0,107693 0,142004
27 saat 0,031467 0,029097 0,026351 0,034746
27 saat 0,031207 0,028856 0,026133 0,034458
27 saat 0,034387 0,031796 0,028796 0,03797
27 saat 0,028557 0,026406 0,023914 0,031533
TARSAN 1018 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,427015 1,566994 1,393777 1,604362
3 saat 1,580083 1,735077 1,54328 1,776454
3 saat 1,342573 1,474269 1,311302 1,509426
3 saat 1,679463 1,844206 1,640346 1,888185
27 saat 5,296356 5,81589 5,172996 5,954581
27 saat 6,940309 7,621104 6,778659 7,802844
27 saat 6,964405 7,647563 6,802193 7,829933
27 saat 5,278032 5,795769 5,155099 5,93398
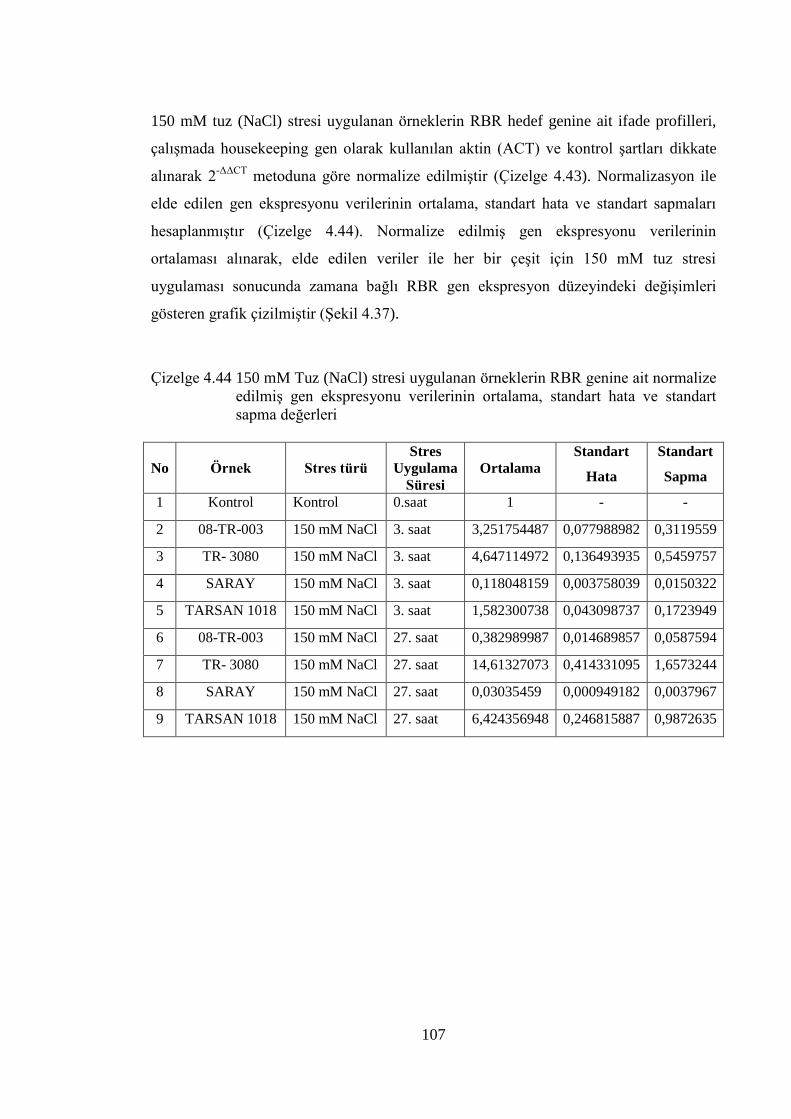
107
150 mM tuz (NaCl) stresi uygulanan örneklerin RBR hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.43). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.44). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM tuz stresi
uygulaması sonucunda zamana bağlı RBR gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.37).
Çizelge 4.44 150 mM Tuz (NaCl) stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM NaCl 3. saat 3,251754487 0,077988982 0,3119559
3 TR- 3080 150 mM NaCl 3. saat 4,647114972 0,136493935 0,5459757
4 SARAY 150 mM NaCl 3. saat 0,118048159 0,003758039 0,0150322
5 TARSAN 1018 150 mM NaCl 3. saat 1,582300738 0,043098737 0,1723949
6 08-TR-003 150 mM NaCl 27. saat 0,382989987 0,014689857 0,0587594
7 TR- 3080 150 mM NaCl 27. saat 14,61327073 0,414331095 1,6573244
8 SARAY 150 mM NaCl 27. saat 0,03035459 0,000949182 0,0037967
9 TARSAN 1018 150 mM NaCl 27. saat 6,424356948 0,246815887 0,9872635
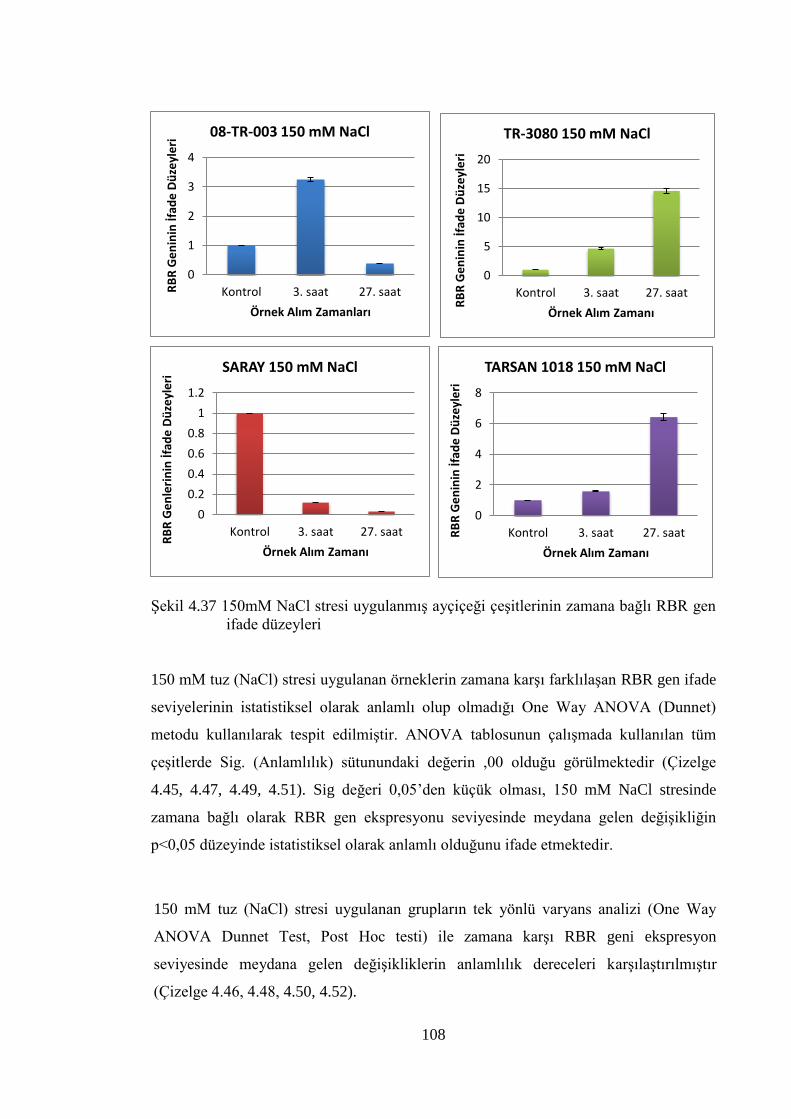
108
Şekil 4.37 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı RBR gen
ifade düzeyleri
150 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan RBR gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.45, 4.47, 4.49, 4.51). Sig değeri 0,05’den küçük olması, 150 mM NaCl stresinde
zamana bağlı olarak RBR gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
150 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı RBR geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.46, 4.48, 4.50, 4.52).
0
1
2
3
4
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanları
08-TR-003 150 mM NaCl
0
5
10
15
20
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 150 mM NaCl
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
RB
R G
en
leri
nin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
SARAY 150 mM NaCl
0
2
4
6
8
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018 150 mM NaCl
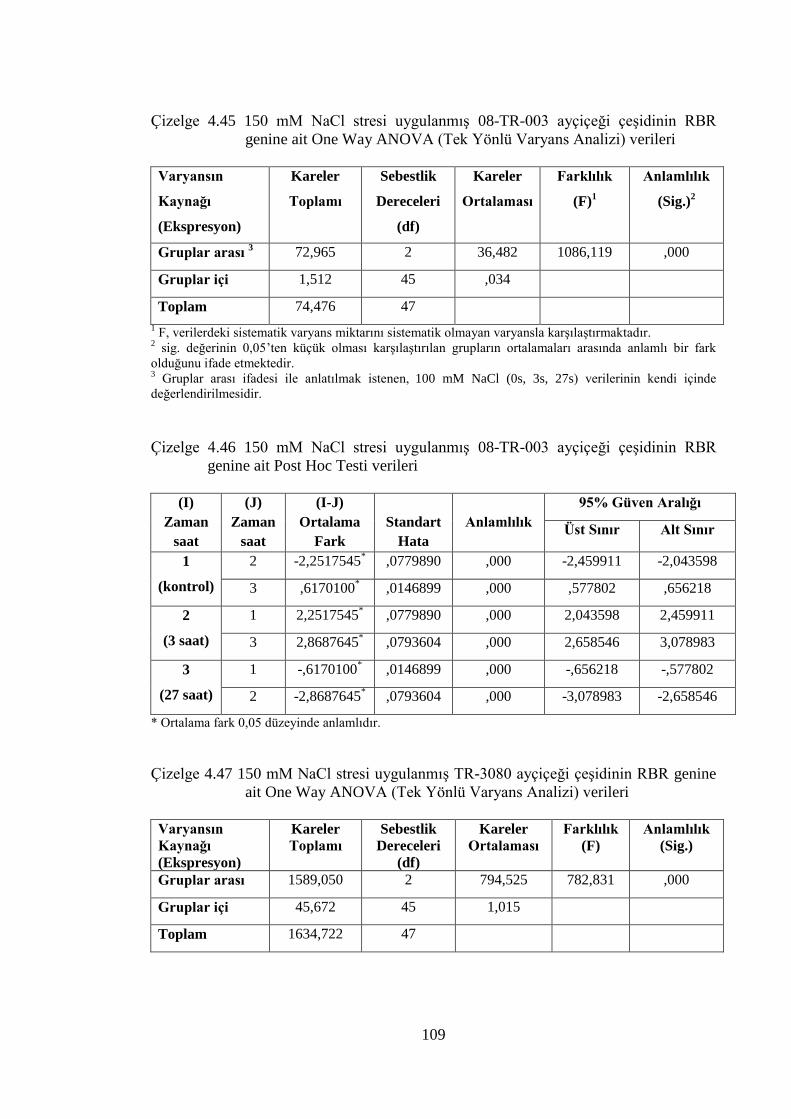
109
Çizelge 4.45 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 72,965 2 36,482 1086,119 ,000
Gruplar içi 1,512 45 ,034
Toplam 74,476 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM NaCl (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.46 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -2,2517545* ,0779890 ,000 -2,459911 -2,043598
3 ,6170100* ,0146899 ,000 ,577802 ,656218
2
(3 saat)
1 2,2517545* ,0779890 ,000 2,043598 2,459911
3 2,8687645* ,0793604 ,000 2,658546 3,078983
3
(27 saat)
1 -,6170100* ,0146899 ,000 -,656218 -,577802
2 -2,8687645* ,0793604 ,000 -3,078983 -2,658546
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.47 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 1589,050 2 794,525 782,831 ,000
Gruplar içi 45,672 45 1,015
Toplam 1634,722 47
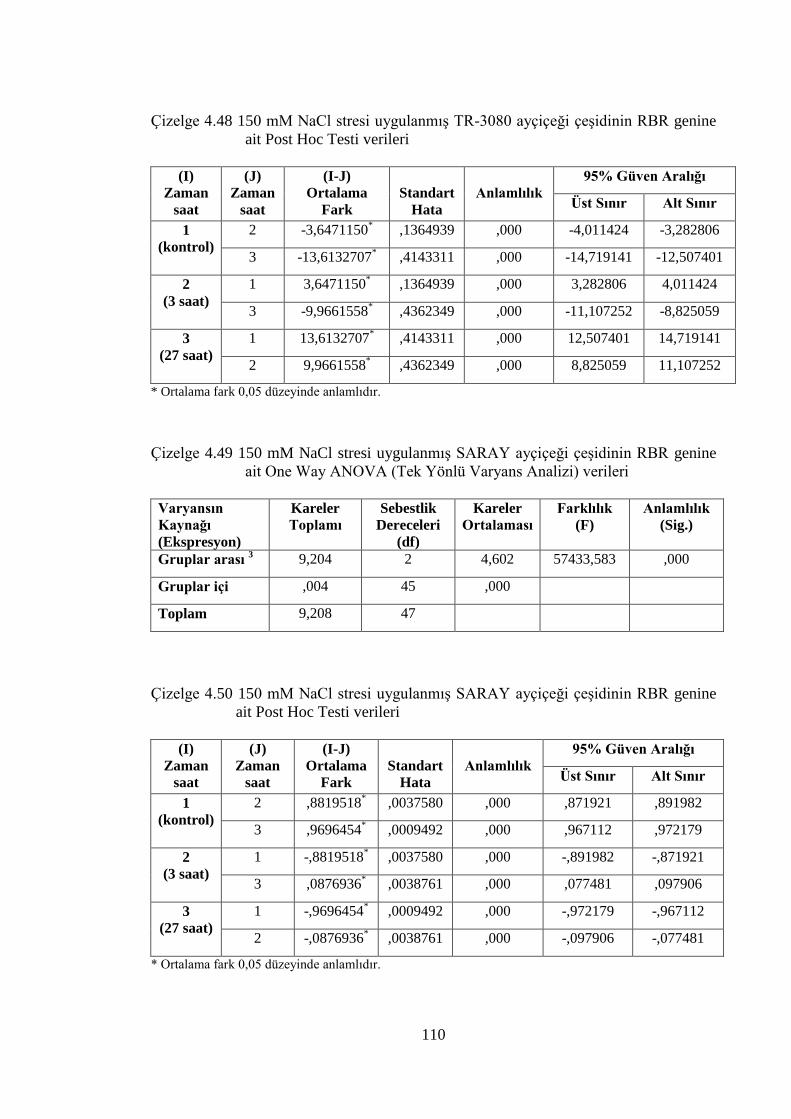
110
Çizelge 4.48 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -3,6471150* ,1364939 ,000 -4,011424 -3,282806
3 -13,6132707* ,4143311 ,000 -14,719141 -12,507401
2
(3 saat)
1 3,6471150* ,1364939 ,000 3,282806 4,011424
3 -9,9661558* ,4362349 ,000 -11,107252 -8,825059
3
(27 saat)
1 13,6132707* ,4143311 ,000 12,507401 14,719141
2 9,9661558* ,4362349 ,000 8,825059 11,107252
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.49 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 3 9,204 2 4,602 57433,583 ,000
Gruplar içi ,004 45 ,000
Toplam 9,208 47
Çizelge 4.50 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8819518* ,0037580 ,000 ,871921 ,891982
3 ,9696454* ,0009492 ,000 ,967112 ,972179
2
(3 saat)
1 -,8819518* ,0037580 ,000 -,891982 -,871921
3 ,0876936* ,0038761 ,000 ,077481 ,097906
3
(27 saat)
1 -,9696454* ,0009492 ,000 -,972179 -,967112
2 -,0876936* ,0038761 ,000 -,097906 -,077481
* Ortalama fark 0,05 düzeyinde anlamlıdır.
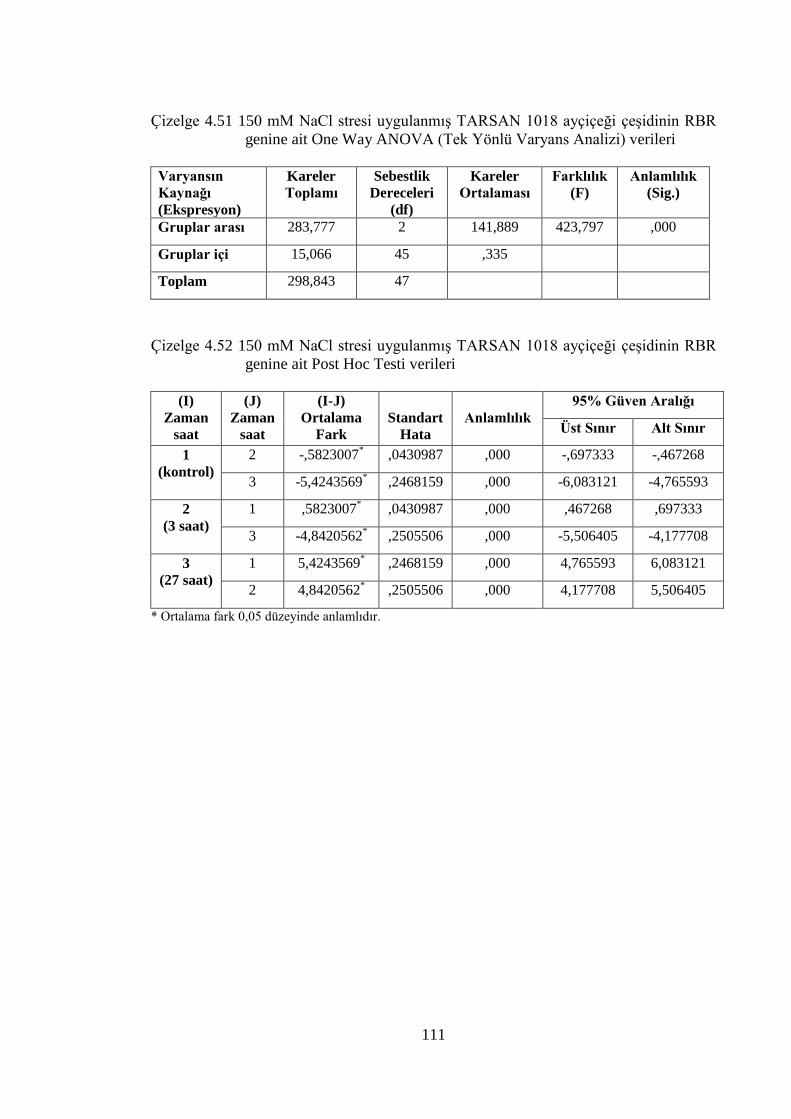
111
Çizelge 4.51 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 283,777 2 141,889 423,797 ,000
Gruplar içi 15,066 45 ,335
Toplam 298,843 47
Çizelge 4.52 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,5823007* ,0430987 ,000 -,697333 -,467268
3 -5,4243569* ,2468159 ,000 -6,083121 -4,765593
2
(3 saat)
1 ,5823007* ,0430987 ,000 ,467268 ,697333
3 -4,8420562* ,2505506 ,000 -5,506405 -4,177708
3
(27 saat)
1 5,4243569* ,2468159 ,000 4,765593 6,083121
2 4,8420562* ,2505506 ,000 4,177708 5,506405
* Ortalama fark 0,05 düzeyinde anlamlıdır.
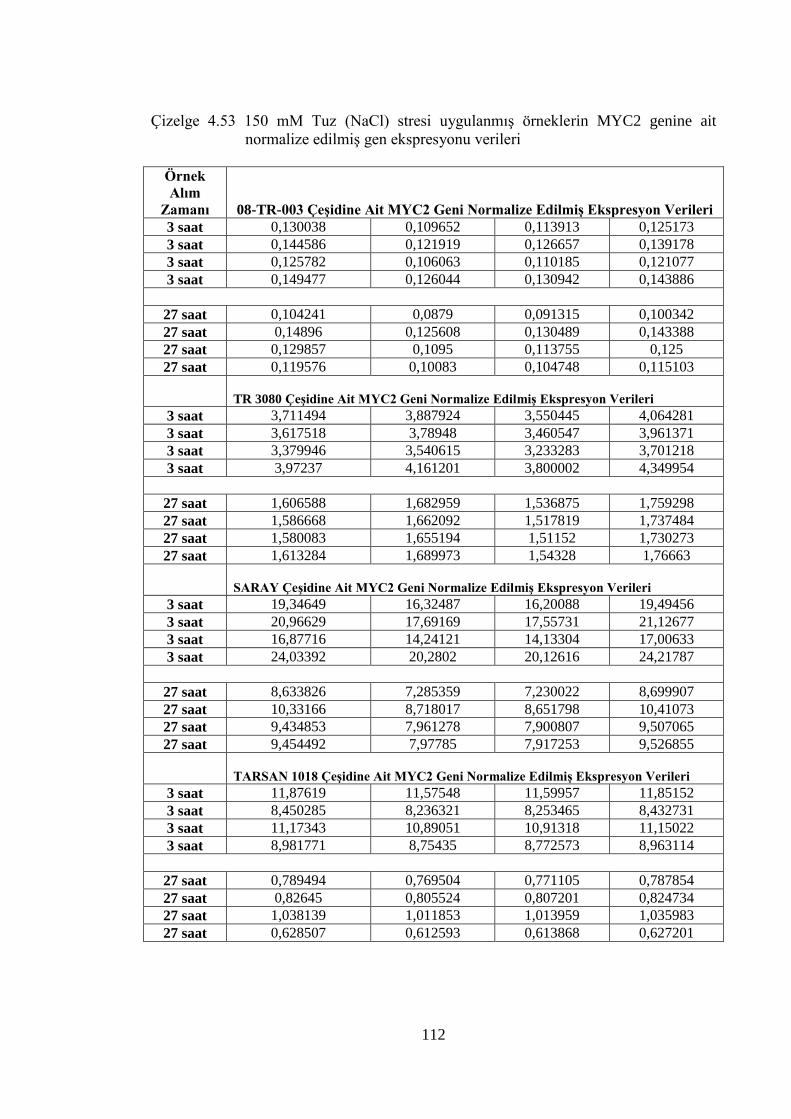
112
Çizelge 4.53 150 mM Tuz (NaCl) stresi uygulanmış örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,130038 0,109652 0,113913 0,125173
3 saat 0,144586 0,121919 0,126657 0,139178
3 saat 0,125782 0,106063 0,110185 0,121077
3 saat 0,149477 0,126044 0,130942 0,143886
27 saat 0,104241 0,0879 0,091315 0,100342
27 saat 0,14896 0,125608 0,130489 0,143388
27 saat 0,129857 0,1095 0,113755 0,125
27 saat 0,119576 0,10083 0,104748 0,115103
TR 3080 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 3,711494 3,887924 3,550445 4,064281
3 saat 3,617518 3,78948 3,460547 3,961371
3 saat 3,379946 3,540615 3,233283 3,701218
3 saat 3,97237 4,161201 3,800002 4,349954
27 saat 1,606588 1,682959 1,536875 1,759298
27 saat 1,586668 1,662092 1,517819 1,737484
27 saat 1,580083 1,655194 1,51152 1,730273
27 saat 1,613284 1,689973 1,54328 1,76663
SARAY Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 19,34649 16,32487 16,20088 19,49456
3 saat 20,96629 17,69169 17,55731 21,12677
3 saat 16,87716 14,24121 14,13304 17,00633
3 saat 24,03392 20,2802 20,12616 24,21787
27 saat 8,633826 7,285359 7,230022 8,699907
27 saat 10,33166 8,718017 8,651798 10,41073
27 saat 9,434853 7,961278 7,900807 9,507065
27 saat 9,454492 7,97785 7,917253 9,526855
TARSAN 1018 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 11,87619 11,57548 11,59957 11,85152
3 saat 8,450285 8,236321 8,253465 8,432731
3 saat 11,17343 10,89051 10,91318 11,15022
3 saat 8,981771 8,75435 8,772573 8,963114
27 saat 0,789494 0,769504 0,771105 0,787854
27 saat 0,82645 0,805524 0,807201 0,824734
27 saat 1,038139 1,011853 1,013959 1,035983
27 saat 0,628507 0,612593 0,613868 0,627201
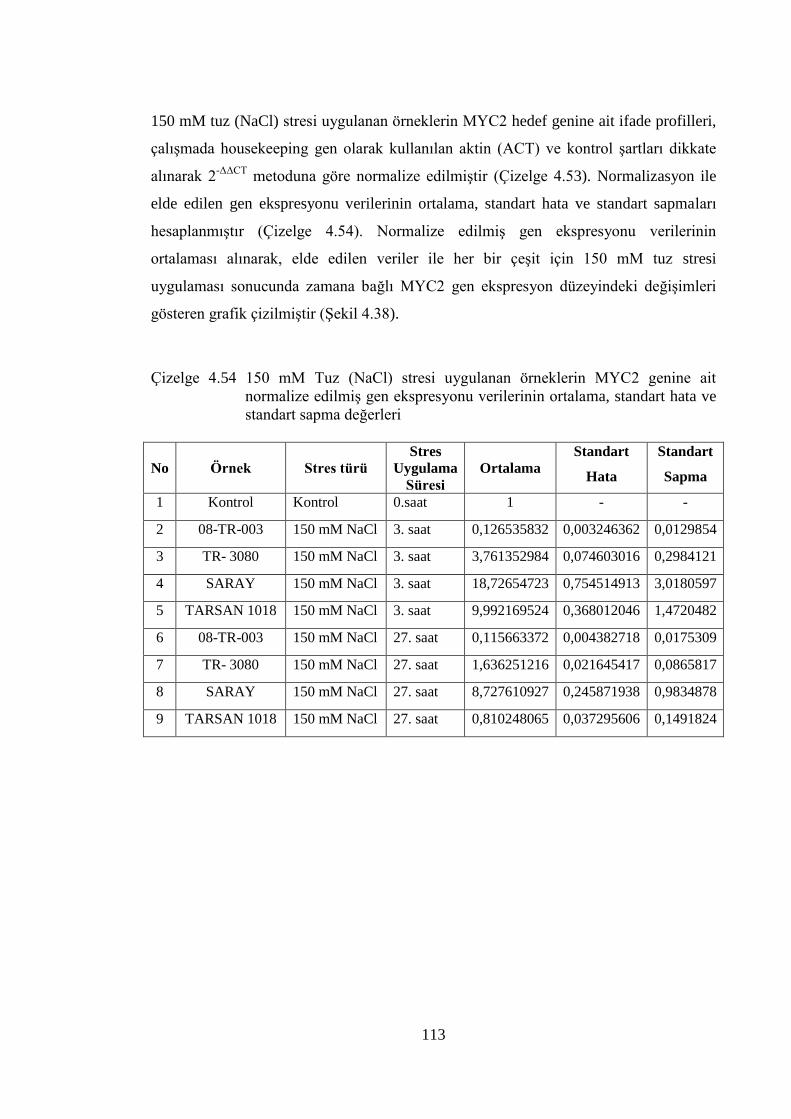
113
150 mM tuz (NaCl) stresi uygulanan örneklerin MYC2 hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.53). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.54). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM tuz stresi
uygulaması sonucunda zamana bağlı MYC2 gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.38).
Çizelge 4.54 150 mM Tuz (NaCl) stresi uygulanan örneklerin MYC2 genine ait
normalize edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve
standart sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM NaCl 3. saat 0,126535832 0,003246362 0,0129854
3 TR- 3080 150 mM NaCl 3. saat 3,761352984 0,074603016 0,2984121
4 SARAY 150 mM NaCl 3. saat 18,72654723 0,754514913 3,0180597
5 TARSAN 1018 150 mM NaCl 3. saat 9,992169524 0,368012046 1,4720482
6 08-TR-003 150 mM NaCl 27. saat 0,115663372 0,004382718 0,0175309
7 TR- 3080 150 mM NaCl 27. saat 1,636251216 0,021645417 0,0865817
8 SARAY 150 mM NaCl 27. saat 8,727610927 0,245871938 0,9834878
9 TARSAN 1018 150 mM NaCl 27. saat 0,810248065 0,037295606 0,1491824
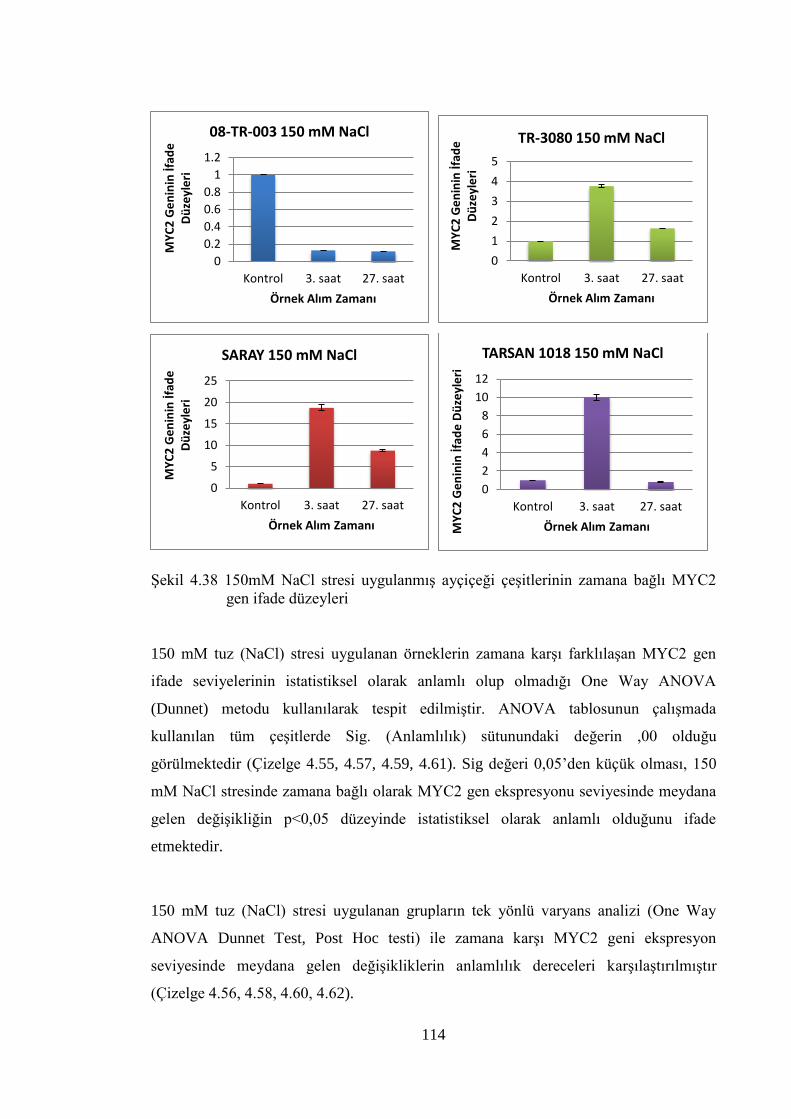
114
Şekil 4.38 150mM NaCl stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı MYC2
gen ifade düzeyleri
150 mM tuz (NaCl) stresi uygulanan örneklerin zamana karşı farklılaşan MYC2 gen
ifade seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA
(Dunnet) metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada
kullanılan tüm çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu
görülmektedir (Çizelge 4.55, 4.57, 4.59, 4.61). Sig değeri 0,05’den küçük olması, 150
mM NaCl stresinde zamana bağlı olarak MYC2 gen ekspresyonu seviyesinde meydana
gelen değişikliğin p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade
etmektedir.
150 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.56, 4.58, 4.60, 4.62).
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
08-TR-003 150 mM NaCl
0
1
2
3
4
5
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 150 mM NaCl
0
5
10
15
20
25
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
D
üze
yle
ri
Örnek Alım Zamanı
SARAY 150 mM NaCl
0
2
4
6
8
10
12
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 150 mM NaCl
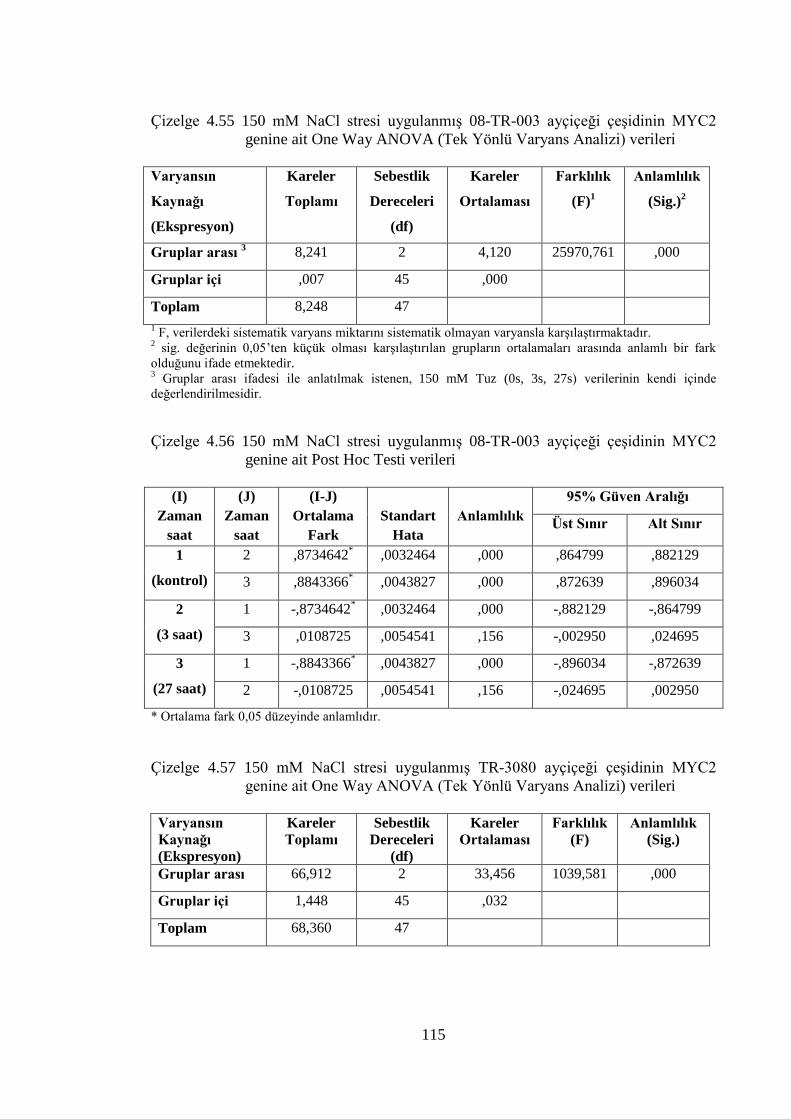
115
Çizelge 4.55 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 8,241 2 4,120 25970,761 ,000
Gruplar içi ,007 45 ,000
Toplam 8,248 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
150 mM Tuz (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.56 150 mM NaCl stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8734642* ,0032464 ,000 ,864799 ,882129
3 ,8843366* ,0043827 ,000 ,872639 ,896034
2
(3 saat)
1 -,8734642* ,0032464 ,000 -,882129 -,864799
3 ,0108725 ,0054541 ,156 -,002950 ,024695
3
(27 saat)
1 -,8843366* ,0043827 ,000 -,896034 -,872639
2 -,0108725 ,0054541 ,156 -,024695 ,002950
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.57 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 66,912 2 33,456 1039,581 ,000
Gruplar içi 1,448 45 ,032
Toplam 68,360 47
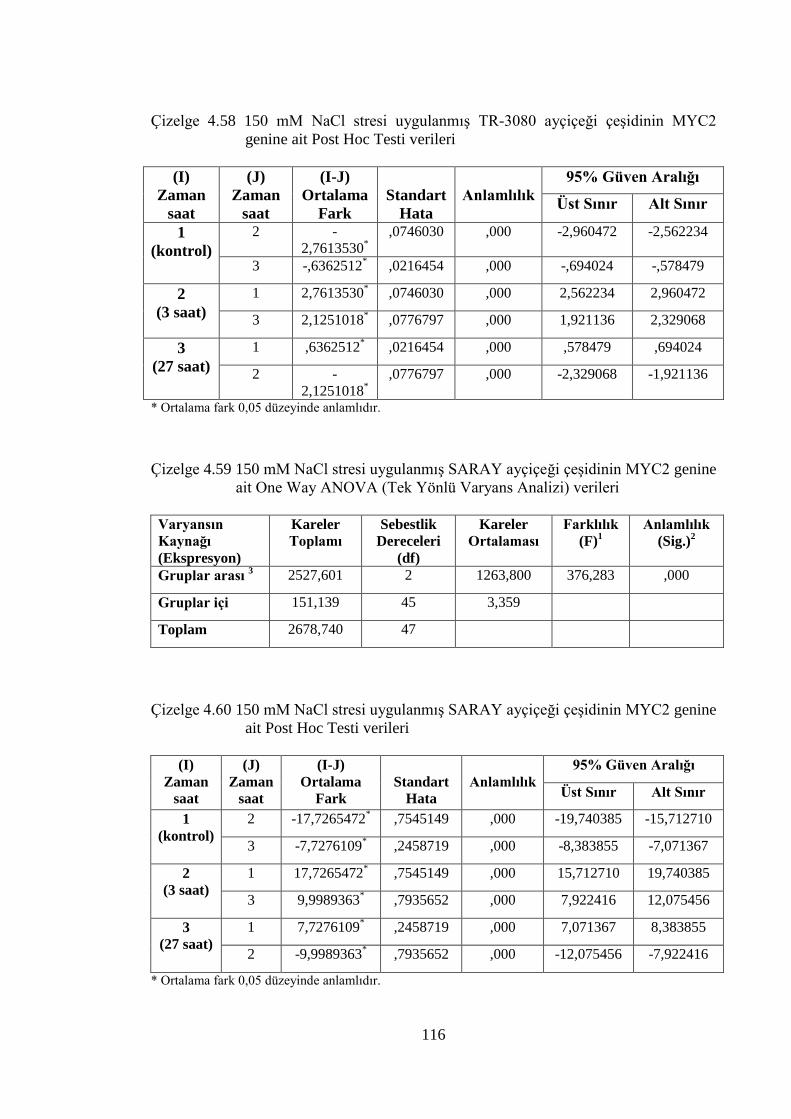
116
Çizelge 4.58 150 mM NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -
2,7613530*
,0746030 ,000 -2,960472 -2,562234
3 -,6362512* ,0216454 ,000 -,694024 -,578479
2
(3 saat)
1 2,7613530* ,0746030 ,000 2,562234 2,960472
3 2,1251018* ,0776797 ,000 1,921136 2,329068
3
(27 saat)
1 ,6362512* ,0216454 ,000 ,578479 ,694024
2 -
2,1251018*
,0776797 ,000 -2,329068 -1,921136
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.59 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 2527,601 2 1263,800 376,283 ,000
Gruplar içi 151,139 45 3,359
Toplam 2678,740 47
Çizelge 4.60 150 mM NaCl stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -17,7265472* ,7545149 ,000 -19,740385 -15,712710
3 -7,7276109* ,2458719 ,000 -8,383855 -7,071367
2
(3 saat)
1 17,7265472* ,7545149 ,000 15,712710 19,740385
3 9,9989363* ,7935652 ,000 7,922416 12,075456
3
(27 saat)
1 7,7276109* ,2458719 ,000 7,071367 8,383855
2 -9,9989363* ,7935652 ,000 -12,075456 -7,922416
* Ortalama fark 0,05 düzeyinde anlamlıdır.
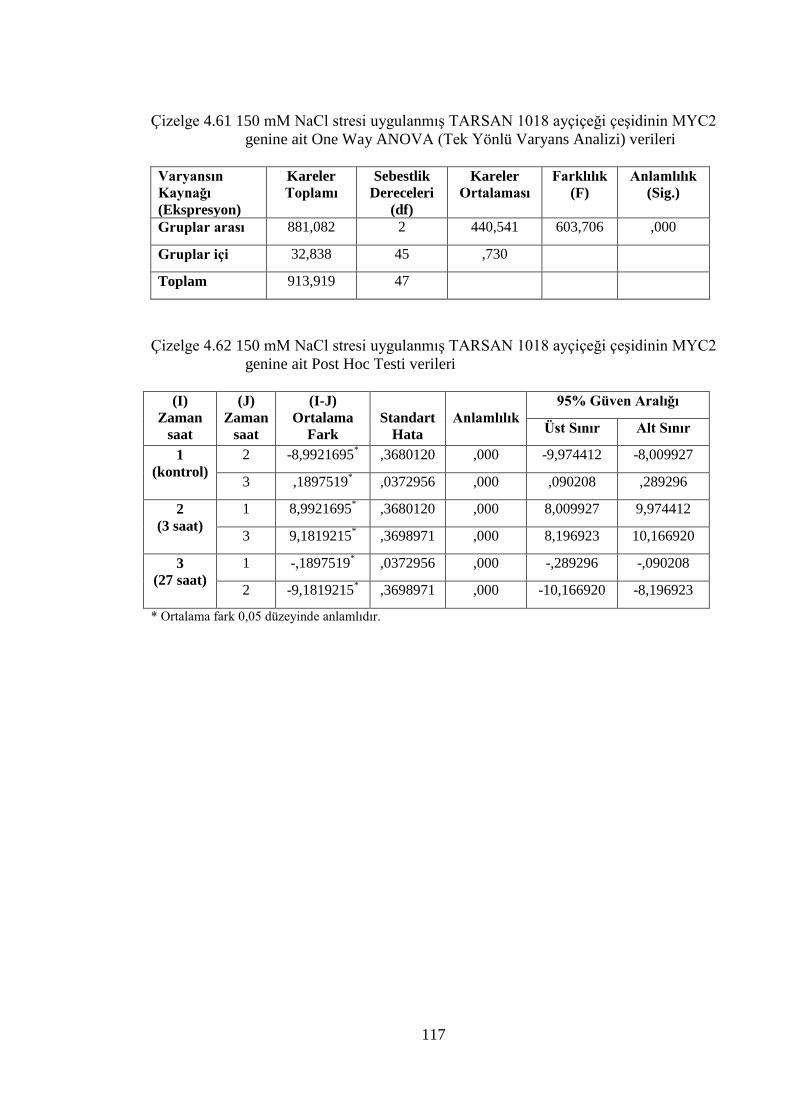
117
Çizelge 4.61 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 881,082 2 440,541 603,706 ,000
Gruplar içi 32,838 45 ,730
Toplam 913,919 47
Çizelge 4.62 150 mM NaCl stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -8,9921695* ,3680120 ,000 -9,974412 -8,009927
3 ,1897519* ,0372956 ,000 ,090208 ,289296
2
(3 saat)
1 8,9921695* ,3680120 ,000 8,009927 9,974412
3 9,1819215* ,3698971 ,000 8,196923 10,166920
3
(27 saat)
1 -,1897519* ,0372956 ,000 -,289296 -,090208
2 -9,1819215* ,3698971 ,000 -10,166920 -8,196923
* Ortalama fark 0,05 düzeyinde anlamlıdır.
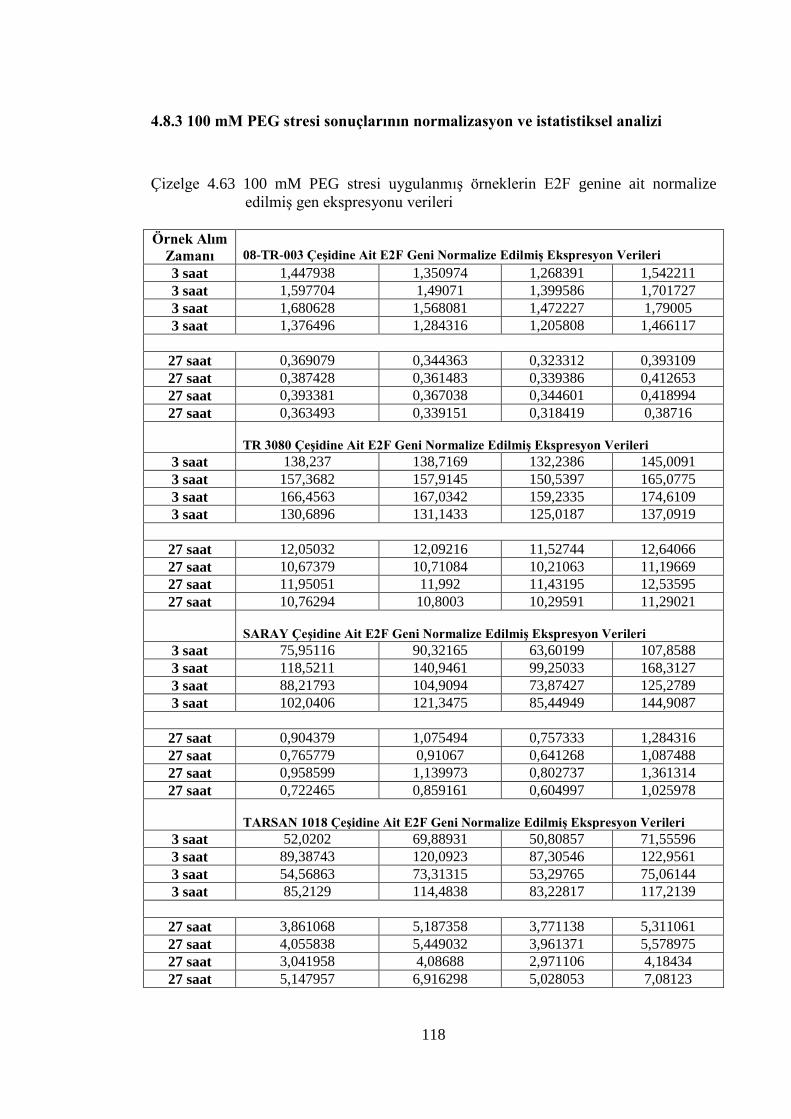
118
4.8.3 100 mM PEG stresi sonuçlarının normalizasyon ve istatistiksel analizi
Çizelge 4.63 100 mM PEG stresi uygulanmış örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,447938 1,350974 1,268391 1,542211
3 saat 1,597704 1,49071 1,399586 1,701727
3 saat 1,680628 1,568081 1,472227 1,79005
3 saat 1,376496 1,284316 1,205808 1,466117
27 saat 0,369079 0,344363 0,323312 0,393109
27 saat 0,387428 0,361483 0,339386 0,412653
27 saat 0,393381 0,367038 0,344601 0,418994
27 saat 0,363493 0,339151 0,318419 0,38716
TR 3080 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 138,237 138,7169 132,2386 145,0091
3 saat 157,3682 157,9145 150,5397 165,0775
3 saat 166,4563 167,0342 159,2335 174,6109
3 saat 130,6896 131,1433 125,0187 137,0919
27 saat 12,05032 12,09216 11,52744 12,64066
27 saat 10,67379 10,71084 10,21063 11,19669
27 saat 11,95051 11,992 11,43195 12,53595
27 saat 10,76294 10,8003 10,29591 11,29021
SARAY Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 75,95116 90,32165 63,60199 107,8588
3 saat 118,5211 140,9461 99,25033 168,3127
3 saat 88,21793 104,9094 73,87427 125,2789
3 saat 102,0406 121,3475 85,44949 144,9087
27 saat 0,904379 1,075494 0,757333 1,284316
27 saat 0,765779 0,91067 0,641268 1,087488
27 saat 0,958599 1,139973 0,802737 1,361314
27 saat 0,722465 0,859161 0,604997 1,025978
TARSAN 1018 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 52,0202 69,88931 50,80857 71,55596
3 saat 89,38743 120,0923 87,30546 122,9561
3 saat 54,56863 73,31315 53,29765 75,06144
3 saat 85,2129 114,4838 83,22817 117,2139
27 saat 3,861068 5,187358 3,771138 5,311061
27 saat 4,055838 5,449032 3,961371 5,578975
27 saat 3,041958 4,08688 2,971106 4,18434
27 saat 5,147957 6,916298 5,028053 7,08123
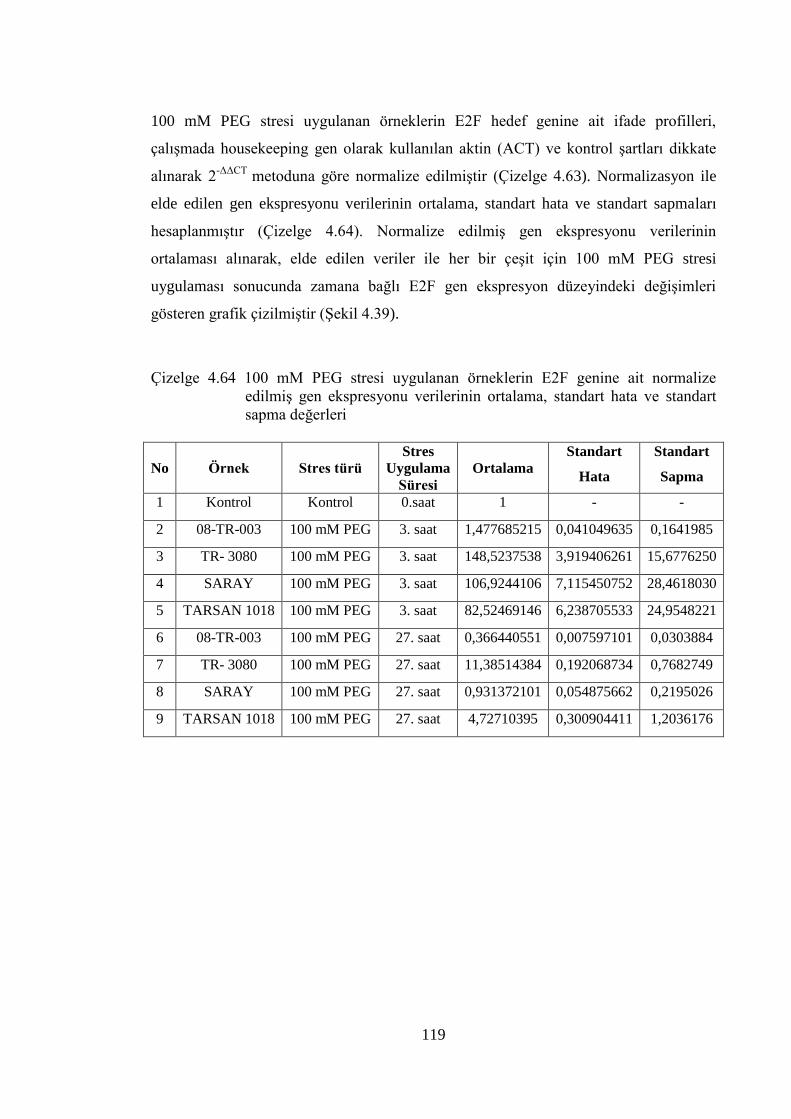
119
100 mM PEG stresi uygulanan örneklerin E2F hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.63). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.64). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 100 mM PEG stresi
uygulaması sonucunda zamana bağlı E2F gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.39).
Çizelge 4.64 100 mM PEG stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM PEG 3. saat 1,477685215 0,041049635 0,1641985
3 TR- 3080 100 mM PEG 3. saat 148,5237538 3,919406261 15,6776250
4 SARAY 100 mM PEG 3. saat 106,9244106 7,115450752 28,4618030
5 TARSAN 1018 100 mM PEG 3. saat 82,52469146 6,238705533 24,9548221
6 08-TR-003 100 mM PEG 27. saat 0,366440551 0,007597101 0,0303884
7 TR- 3080 100 mM PEG 27. saat 11,38514384 0,192068734 0,7682749
8 SARAY 100 mM PEG 27. saat 0,931372101 0,054875662 0,2195026
9 TARSAN 1018 100 mM PEG 27. saat 4,72710395 0,300904411 1,2036176
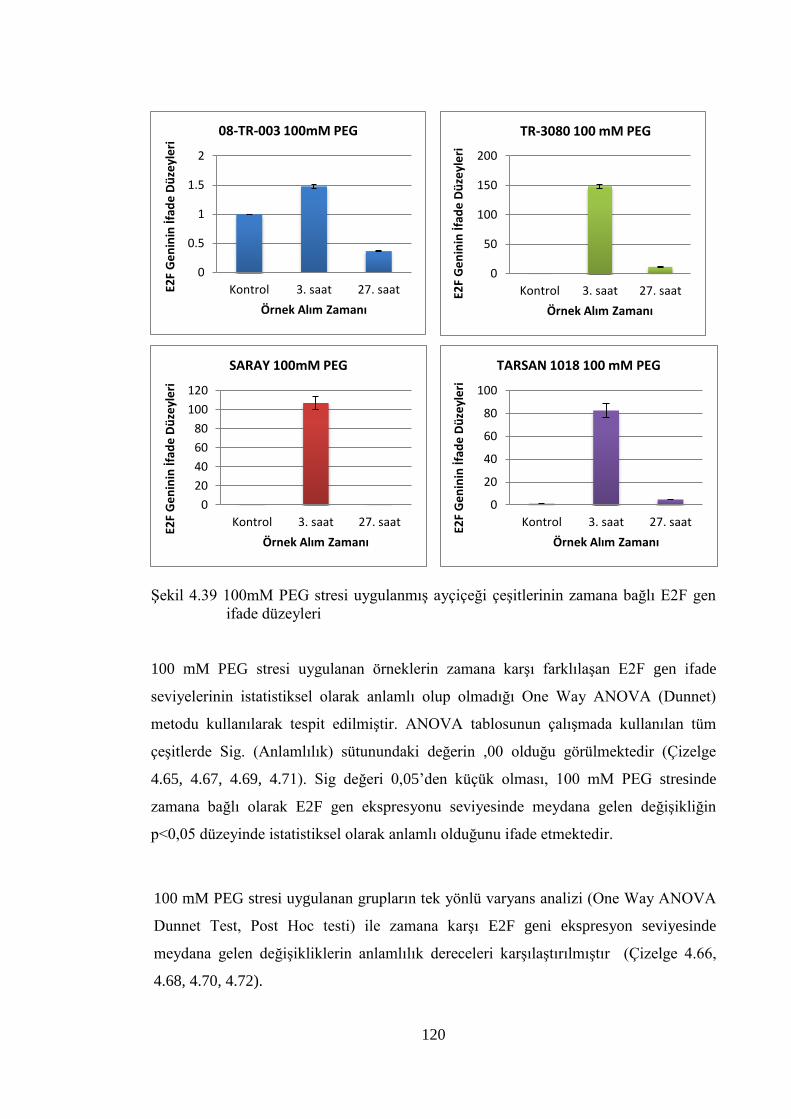
120
Şekil 4.39 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı E2F gen
ifade düzeyleri
100 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.65, 4.67, 4.69, 4.71). Sig değeri 0,05’den küçük olması, 100 mM PEG stresinde
zamana bağlı olarak E2F gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
100 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı E2F geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.66,
4.68, 4.70, 4.72).
0
0.5
1
1.5
2
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 100mM PEG
0
50
100
150
200
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 100 mM PEG
0
20
40
60
80
100
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 100 mM PEG
0
20
40
60
80
100
120
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
SARAY 100mM PEG
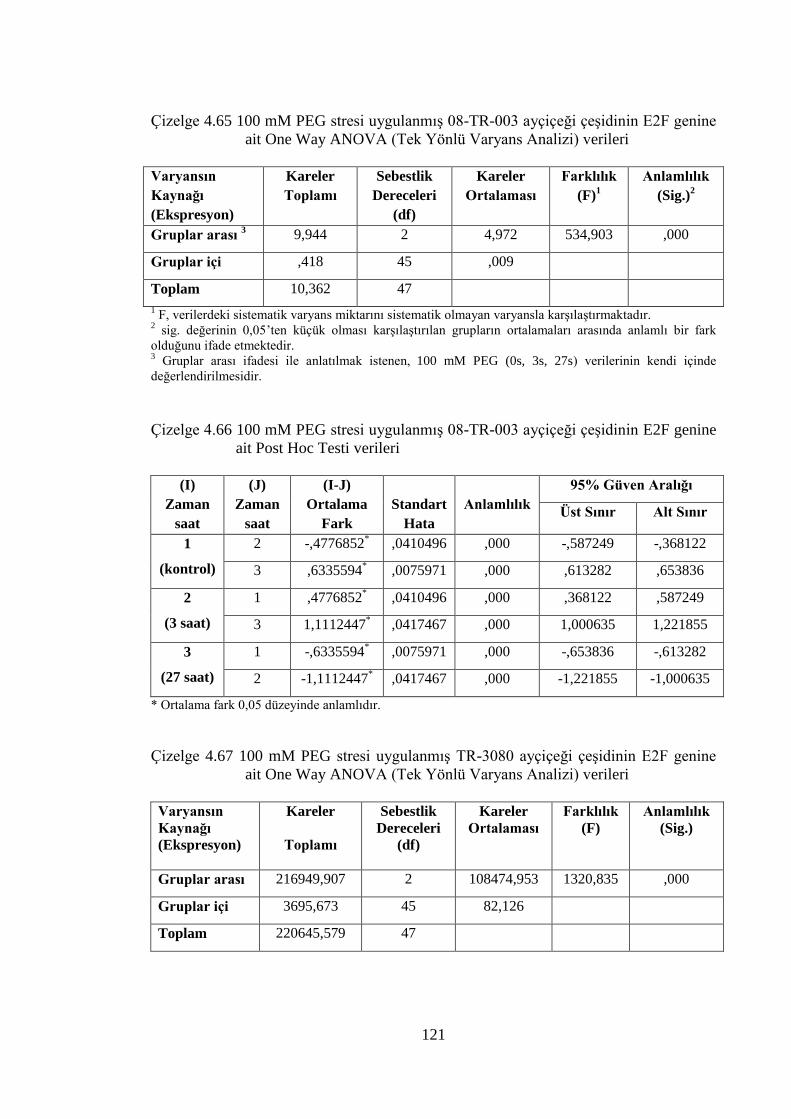
121
Çizelge 4.65 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 9,944 2 4,972 534,903 ,000
Gruplar içi ,418 45 ,009
Toplam 10,362 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.66 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,4776852* ,0410496 ,000 -,587249 -,368122
3 ,6335594* ,0075971 ,000 ,613282 ,653836
2
(3 saat)
1 ,4776852* ,0410496 ,000 ,368122 ,587249
3 1,1112447* ,0417467 ,000 1,000635 1,221855
3
(27 saat)
1 -,6335594* ,0075971 ,000 -,653836 -,613282
2 -1,1112447* ,0417467 ,000 -1,221855 -1,000635
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.67 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 216949,907 2 108474,953 1320,835 ,000
Gruplar içi 3695,673 45 82,126
Toplam 220645,579 47
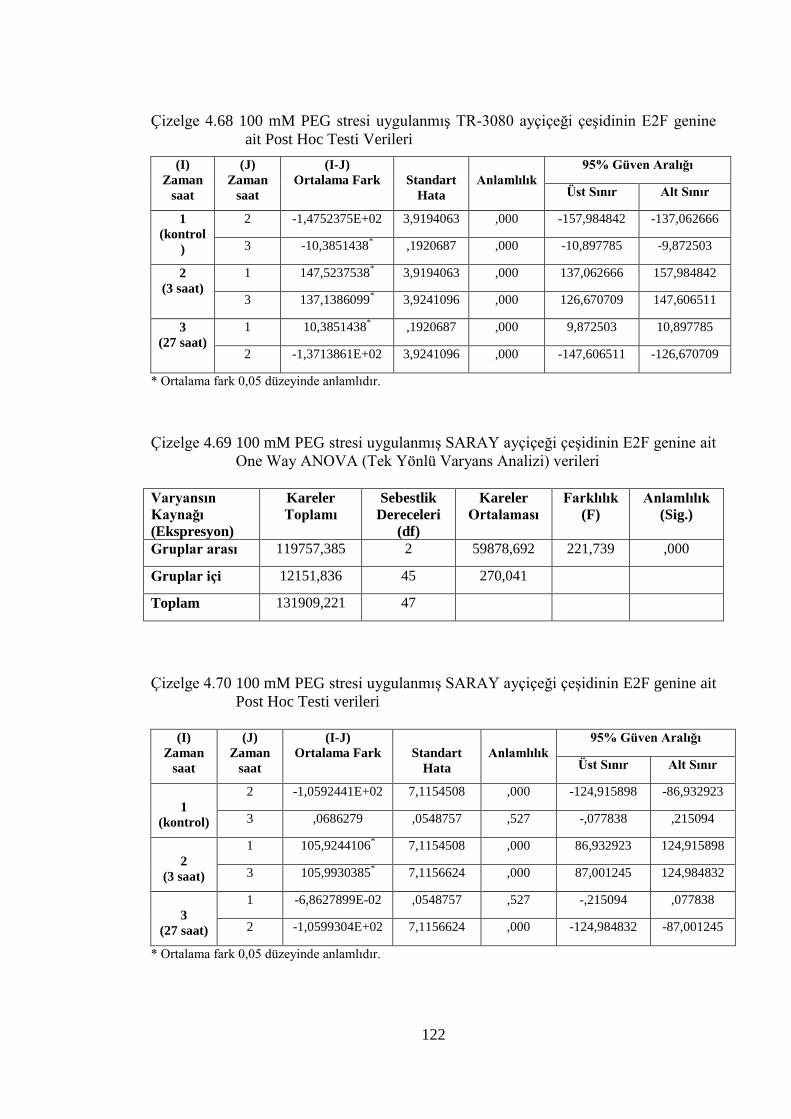
122
Çizelge 4.68 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi Verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol
)
2 -1,4752375E+02 3,9194063 ,000 -157,984842 -137,062666
3 -10,3851438* ,1920687 ,000 -10,897785 -9,872503
2
(3 saat)
1 147,5237538* 3,9194063 ,000 137,062666 157,984842
3 137,1386099* 3,9241096 ,000 126,670709 147,606511
3
(27 saat)
1 10,3851438* ,1920687 ,000 9,872503 10,897785
2 -1,3713861E+02 3,9241096 ,000 -147,606511 -126,670709
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.69 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 119757,385 2 59878,692 221,739 ,000
Gruplar içi 12151,836 45 270,041
Toplam 131909,221 47
Çizelge 4.70 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait
Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -1,0592441E+02 7,1154508 ,000 -124,915898 -86,932923
3 ,0686279 ,0548757 ,527 -,077838 ,215094
2
(3 saat)
1 105,9244106* 7,1154508 ,000 86,932923 124,915898
3 105,9930385* 7,1156624 ,000 87,001245 124,984832
3
(27 saat)
1 -6,8627899E-02 ,0548757 ,527 -,215094 ,077838
2 -1,0599304E+02 7,1156624 ,000 -124,984832 -87,001245
* Ortalama fark 0,05 düzeyinde anlamlıdır.
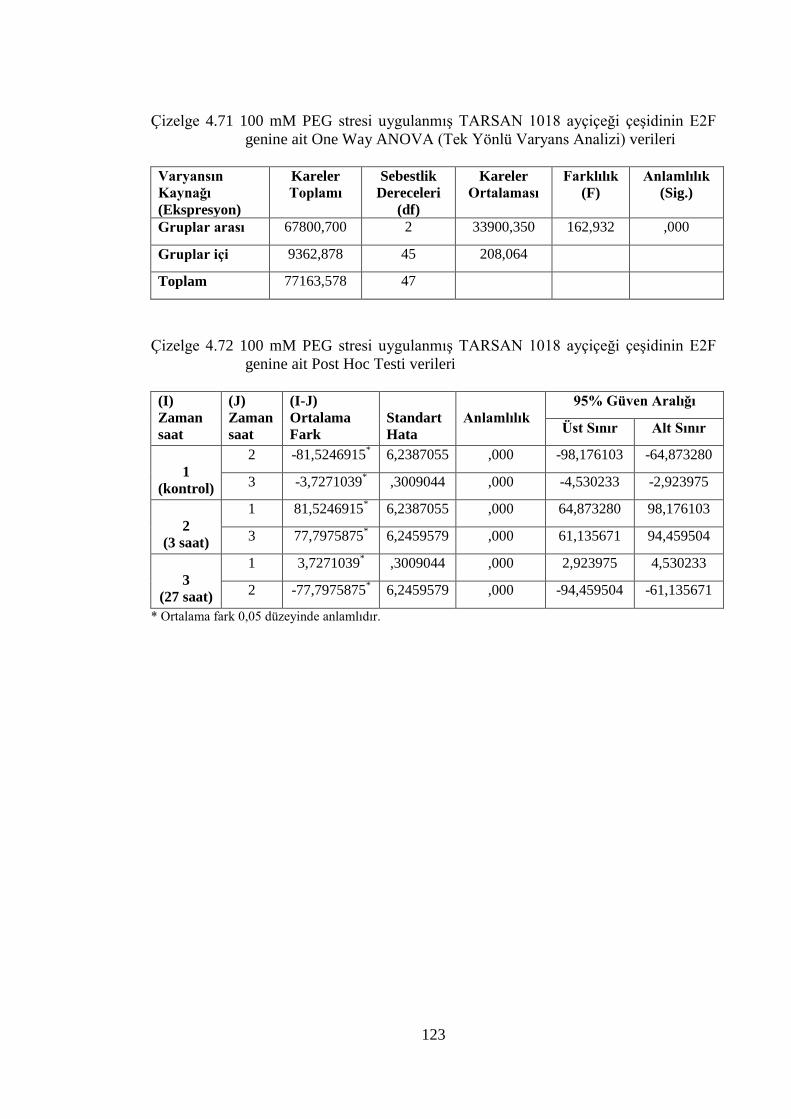
123
Çizelge 4.71 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 67800,700 2 33900,350 162,932 ,000
Gruplar içi 9362,878 45 208,064
Toplam 77163,578 47
Çizelge 4.72 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -81,5246915* 6,2387055 ,000 -98,176103 -64,873280
3 -3,7271039* ,3009044 ,000 -4,530233 -2,923975
2
(3 saat)
1 81,5246915* 6,2387055 ,000 64,873280 98,176103
3 77,7975875* 6,2459579 ,000 61,135671 94,459504
3
(27 saat)
1 3,7271039* ,3009044 ,000 2,923975 4,530233
2 -77,7975875* 6,2459579 ,000 -94,459504 -61,135671
* Ortalama fark 0,05 düzeyinde anlamlıdır.
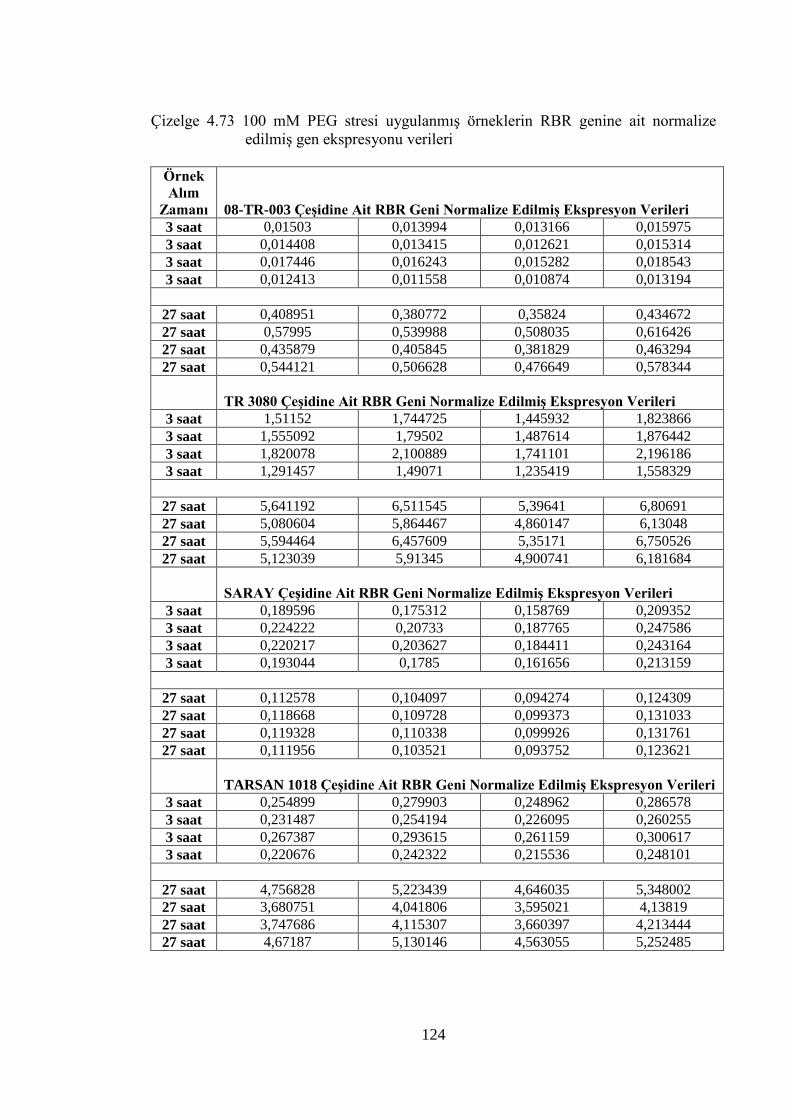
124
Çizelge 4.73 100 mM PEG stresi uygulanmış örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,01503 0,013994 0,013166 0,015975
3 saat 0,014408 0,013415 0,012621 0,015314
3 saat 0,017446 0,016243 0,015282 0,018543
3 saat 0,012413 0,011558 0,010874 0,013194
27 saat 0,408951 0,380772 0,35824 0,434672
27 saat 0,57995 0,539988 0,508035 0,616426
27 saat 0,435879 0,405845 0,381829 0,463294
27 saat 0,544121 0,506628 0,476649 0,578344
TR 3080 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,51152 1,744725 1,445932 1,823866
3 saat 1,555092 1,79502 1,487614 1,876442
3 saat 1,820078 2,100889 1,741101 2,196186
3 saat 1,291457 1,49071 1,235419 1,558329
27 saat 5,641192 6,511545 5,39641 6,80691
27 saat 5,080604 5,864467 4,860147 6,13048
27 saat 5,594464 6,457609 5,35171 6,750526
27 saat 5,123039 5,91345 4,900741 6,181684
SARAY Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,189596 0,175312 0,158769 0,209352
3 saat 0,224222 0,20733 0,187765 0,247586
3 saat 0,220217 0,203627 0,184411 0,243164
3 saat 0,193044 0,1785 0,161656 0,213159
27 saat 0,112578 0,104097 0,094274 0,124309
27 saat 0,118668 0,109728 0,099373 0,131033
27 saat 0,119328 0,110338 0,099926 0,131761
27 saat 0,111956 0,103521 0,093752 0,123621
TARSAN 1018 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,254899 0,279903 0,248962 0,286578
3 saat 0,231487 0,254194 0,226095 0,260255
3 saat 0,267387 0,293615 0,261159 0,300617
3 saat 0,220676 0,242322 0,215536 0,248101
27 saat 4,756828 5,223439 4,646035 5,348002
27 saat 3,680751 4,041806 3,595021 4,13819
27 saat 3,747686 4,115307 3,660397 4,213444
27 saat 4,67187 5,130146 4,563055 5,252485
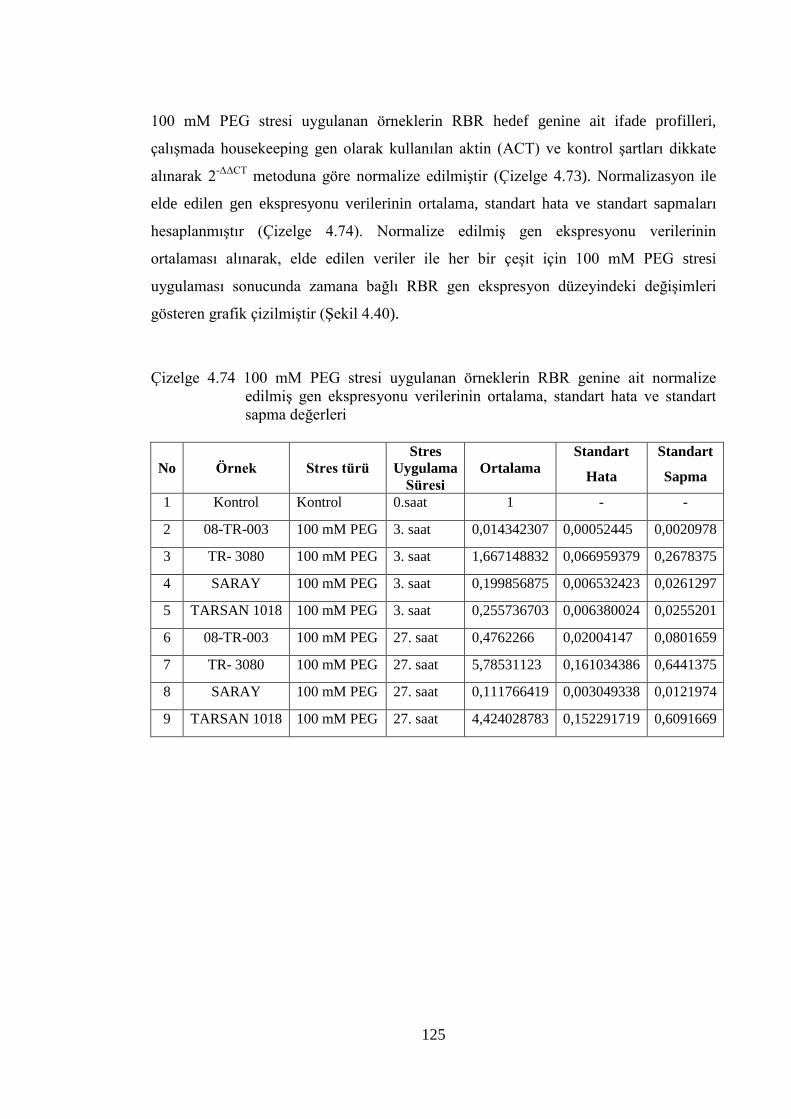
125
100 mM PEG stresi uygulanan örneklerin RBR hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.73). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.74). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 100 mM PEG stresi
uygulaması sonucunda zamana bağlı RBR gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.40).
Çizelge 4.74 100 mM PEG stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM PEG 3. saat 0,014342307 0,00052445 0,0020978
3 TR- 3080 100 mM PEG 3. saat 1,667148832 0,066959379 0,2678375
4 SARAY 100 mM PEG 3. saat 0,199856875 0,006532423 0,0261297
5 TARSAN 1018 100 mM PEG 3. saat 0,255736703 0,006380024 0,0255201
6 08-TR-003 100 mM PEG 27. saat 0,4762266 0,02004147 0,0801659
7 TR- 3080 100 mM PEG 27. saat 5,78531123 0,161034386 0,6441375
8 SARAY 100 mM PEG 27. saat 0,111766419 0,003049338 0,0121974
9 TARSAN 1018 100 mM PEG 27. saat 4,424028783 0,152291719 0,6091669
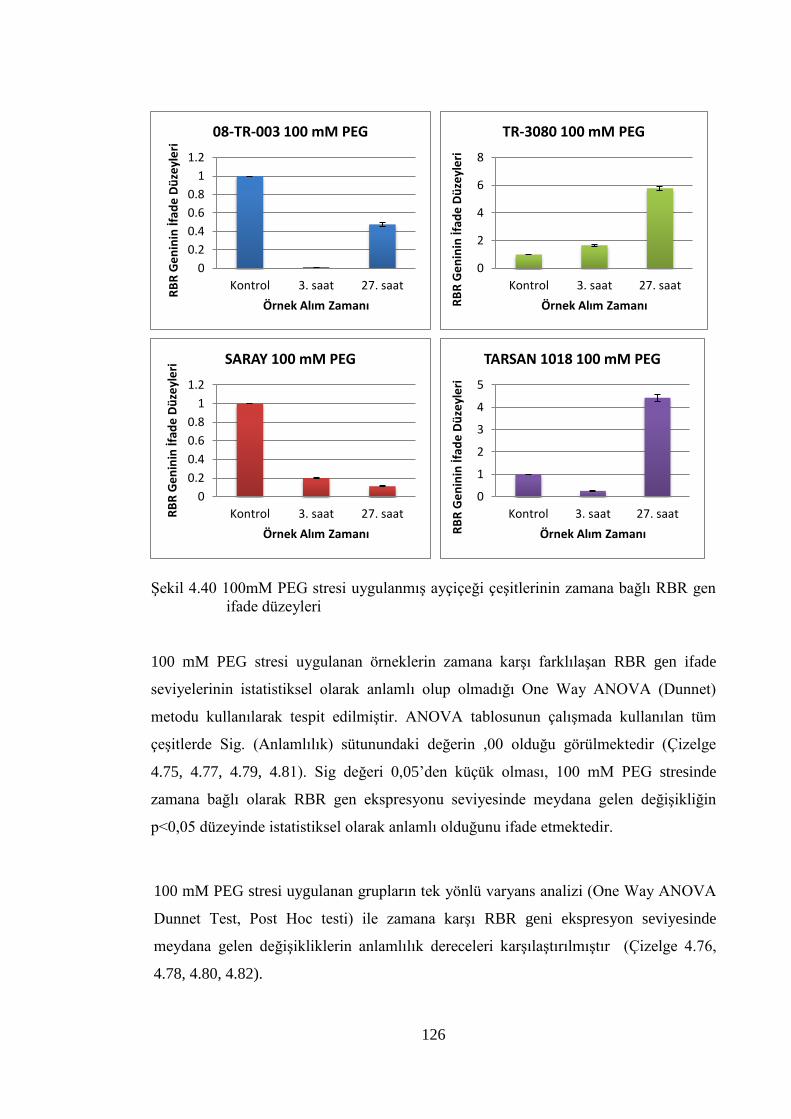
126
Şekil 4.40 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı RBR gen
ifade düzeyleri
100 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan RBR gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.75, 4.77, 4.79, 4.81). Sig değeri 0,05’den küçük olması, 100 mM PEG stresinde
zamana bağlı olarak RBR gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
100 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı RBR geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.76,
4.78, 4.80, 4.82).
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
08-TR-003 100 mM PEG
0
2
4
6
8
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 100 mM PEG
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
SARAY 100 mM PEG
0
1
2
3
4
5
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018 100 mM PEG
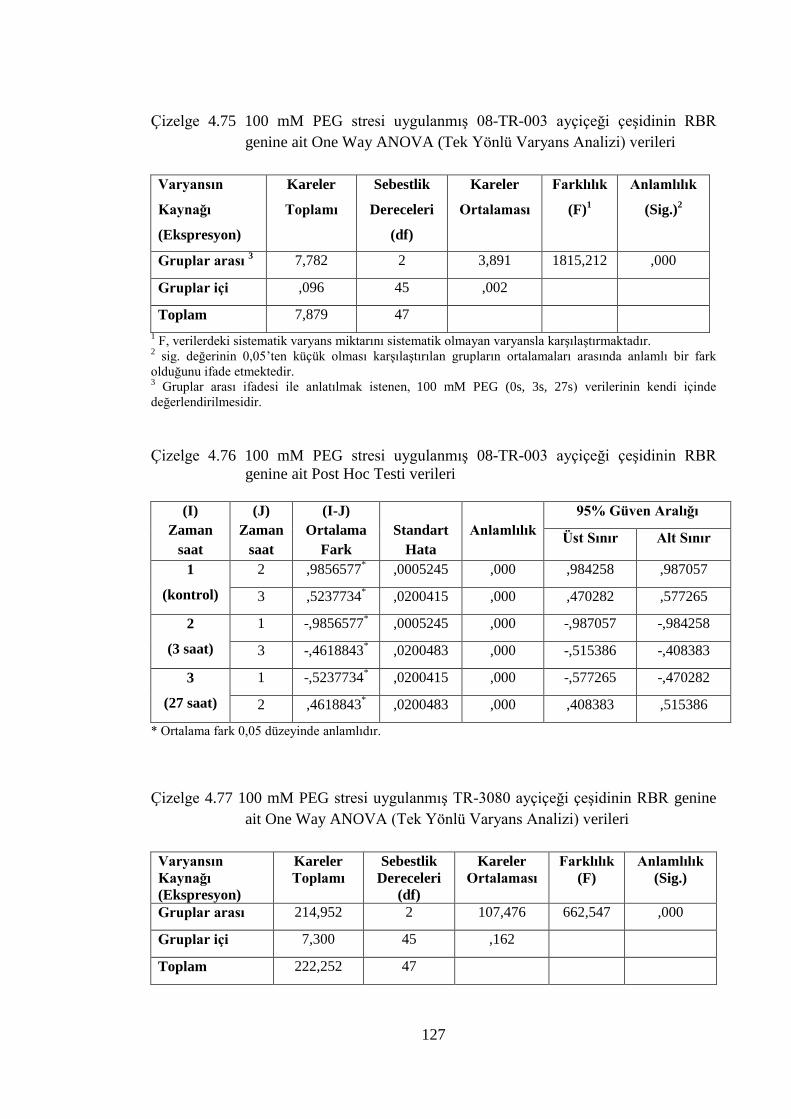
127
Çizelge 4.75 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 7,782 2 3,891 1815,212 ,000
Gruplar içi ,096 45 ,002
Toplam 7,879 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.76 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,9856577* ,0005245 ,000 ,984258 ,987057
3 ,5237734* ,0200415 ,000 ,470282 ,577265
2
(3 saat)
1 -,9856577* ,0005245 ,000 -,987057 -,984258
3 -,4618843* ,0200483 ,000 -,515386 -,408383
3
(27 saat)
1 -,5237734* ,0200415 ,000 -,577265 -,470282
2 ,4618843* ,0200483 ,000 ,408383 ,515386
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.77 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 214,952 2 107,476 662,547 ,000
Gruplar içi 7,300 45 ,162
Toplam 222,252 47
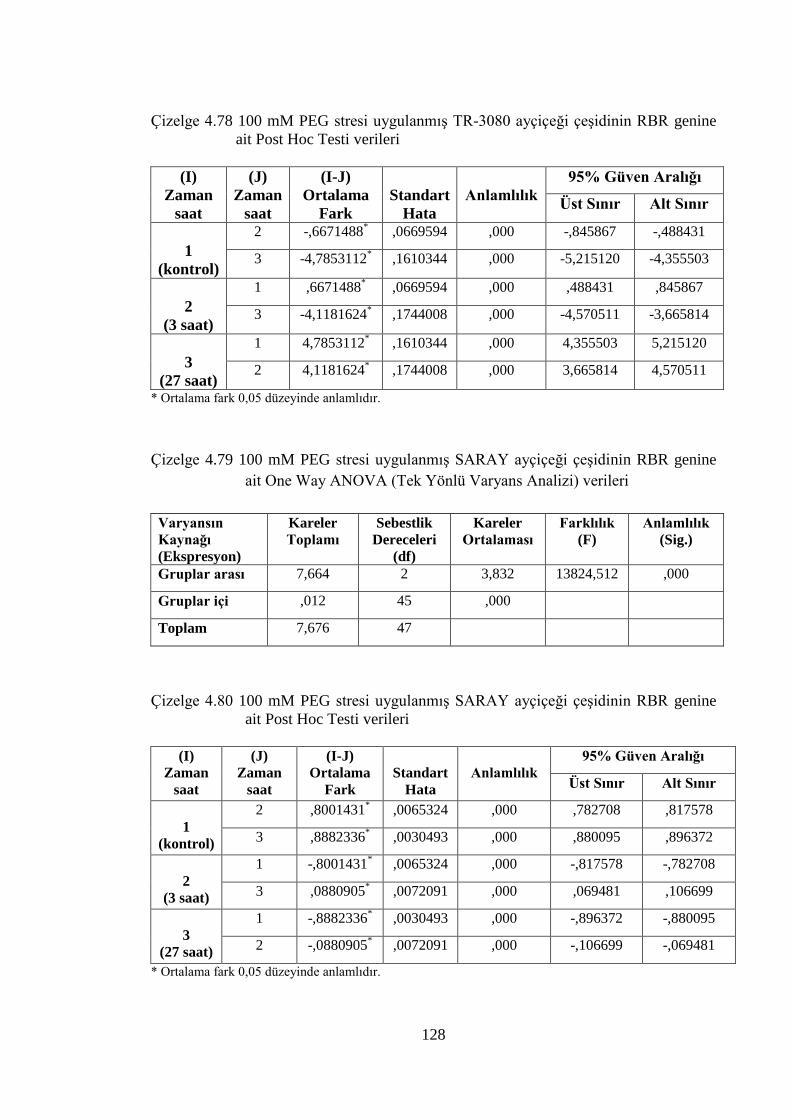
128
Çizelge 4.78 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,6671488* ,0669594 ,000 -,845867 -,488431
3 -4,7853112* ,1610344 ,000 -5,215120 -4,355503
2
(3 saat)
1 ,6671488* ,0669594 ,000 ,488431 ,845867
3 -4,1181624* ,1744008 ,000 -4,570511 -3,665814
3
(27 saat)
1 4,7853112* ,1610344 ,000 4,355503 5,215120
2 4,1181624* ,1744008 ,000 3,665814 4,570511
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.79 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 7,664 2 3,832 13824,512 ,000
Gruplar içi ,012 45 ,000
Toplam 7,676 47
Çizelge 4.80 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8001431* ,0065324 ,000 ,782708 ,817578
3 ,8882336* ,0030493 ,000 ,880095 ,896372
2
(3 saat)
1 -,8001431* ,0065324 ,000 -,817578 -,782708
3 ,0880905* ,0072091 ,000 ,069481 ,106699
3
(27 saat)
1 -,8882336* ,0030493 ,000 -,896372 -,880095
2 -,0880905* ,0072091 ,000 -,106699 -,069481
* Ortalama fark 0,05 düzeyinde anlamlıdır.
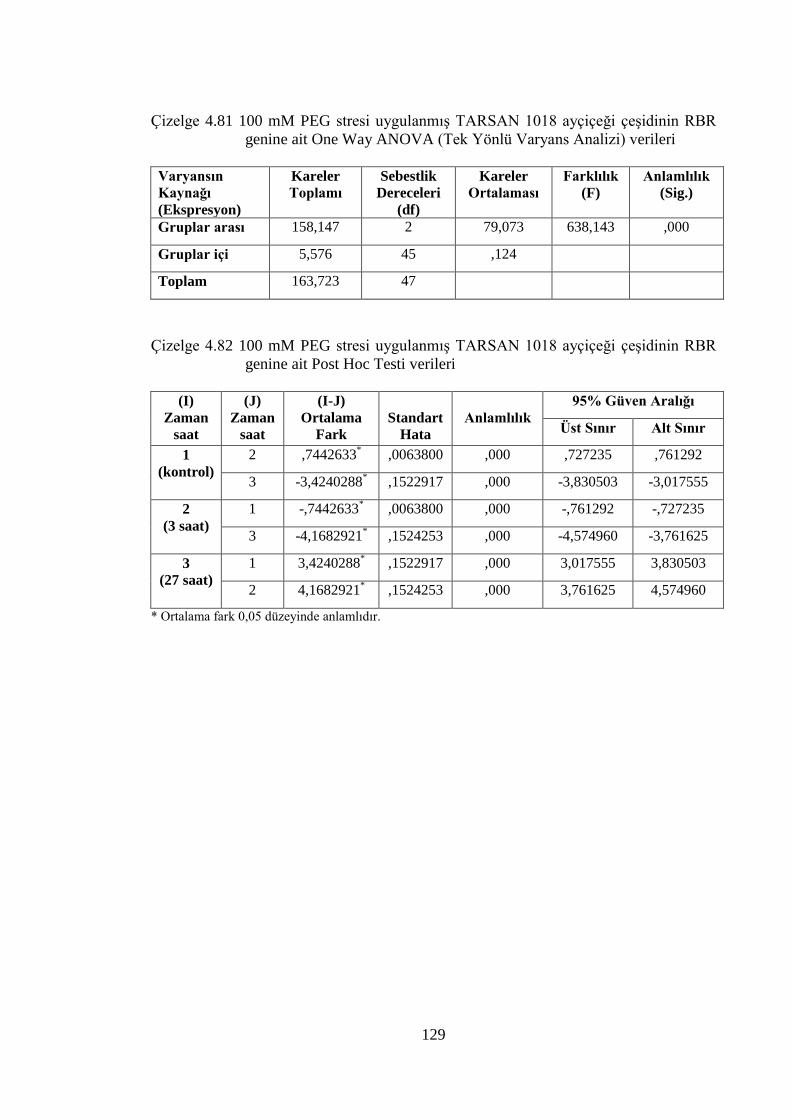
129
Çizelge 4.81 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 158,147 2 79,073 638,143 ,000
Gruplar içi 5,576 45 ,124
Toplam 163,723 47
Çizelge 4.82 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,7442633* ,0063800 ,000 ,727235 ,761292
3 -3,4240288* ,1522917 ,000 -3,830503 -3,017555
2
(3 saat)
1 -,7442633* ,0063800 ,000 -,761292 -,727235
3 -4,1682921* ,1524253 ,000 -4,574960 -3,761625
3
(27 saat)
1 3,4240288* ,1522917 ,000 3,017555 3,830503
2 4,1682921* ,1524253 ,000 3,761625 4,574960
* Ortalama fark 0,05 düzeyinde anlamlıdır.
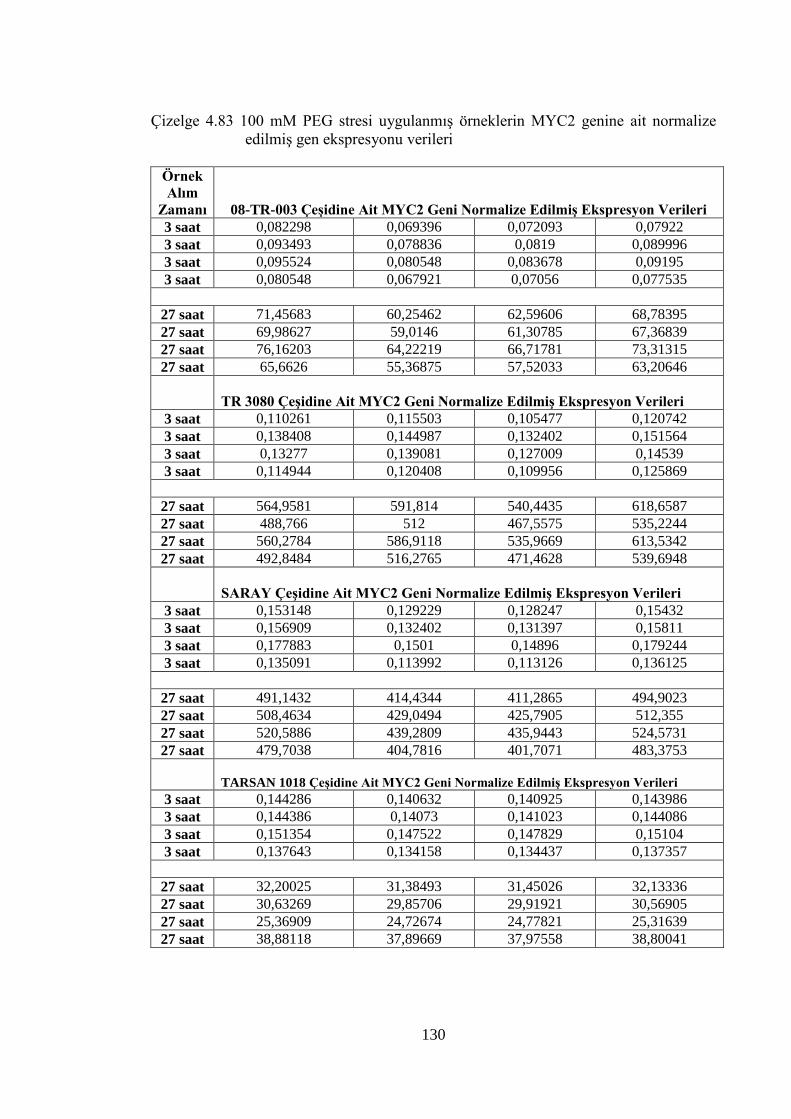
130
Çizelge 4.83 100 mM PEG stresi uygulanmış örneklerin MYC2 genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,082298 0,069396 0,072093 0,07922
3 saat 0,093493 0,078836 0,0819 0,089996
3 saat 0,095524 0,080548 0,083678 0,09195
3 saat 0,080548 0,067921 0,07056 0,077535
27 saat 71,45683 60,25462 62,59606 68,78395
27 saat 69,98627 59,0146 61,30785 67,36839
27 saat 76,16203 64,22219 66,71781 73,31315
27 saat 65,6626 55,36875 57,52033 63,20646
TR 3080 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,110261 0,115503 0,105477 0,120742
3 saat 0,138408 0,144987 0,132402 0,151564
3 saat 0,13277 0,139081 0,127009 0,14539
3 saat 0,114944 0,120408 0,109956 0,125869
27 saat 564,9581 591,814 540,4435 618,6587
27 saat 488,766 512 467,5575 535,2244
27 saat 560,2784 586,9118 535,9669 613,5342
27 saat 492,8484 516,2765 471,4628 539,6948
SARAY Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,153148 0,129229 0,128247 0,15432
3 saat 0,156909 0,132402 0,131397 0,15811
3 saat 0,177883 0,1501 0,14896 0,179244
3 saat 0,135091 0,113992 0,113126 0,136125
27 saat 491,1432 414,4344 411,2865 494,9023
27 saat 508,4634 429,0494 425,7905 512,355
27 saat 520,5886 439,2809 435,9443 524,5731
27 saat 479,7038 404,7816 401,7071 483,3753
TARSAN 1018 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,144286 0,140632 0,140925 0,143986
3 saat 0,144386 0,14073 0,141023 0,144086
3 saat 0,151354 0,147522 0,147829 0,15104
3 saat 0,137643 0,134158 0,134437 0,137357
27 saat 32,20025 31,38493 31,45026 32,13336
27 saat 30,63269 29,85706 29,91921 30,56905
27 saat 25,36909 24,72674 24,77821 25,31639
27 saat 38,88118 37,89669 37,97558 38,80041
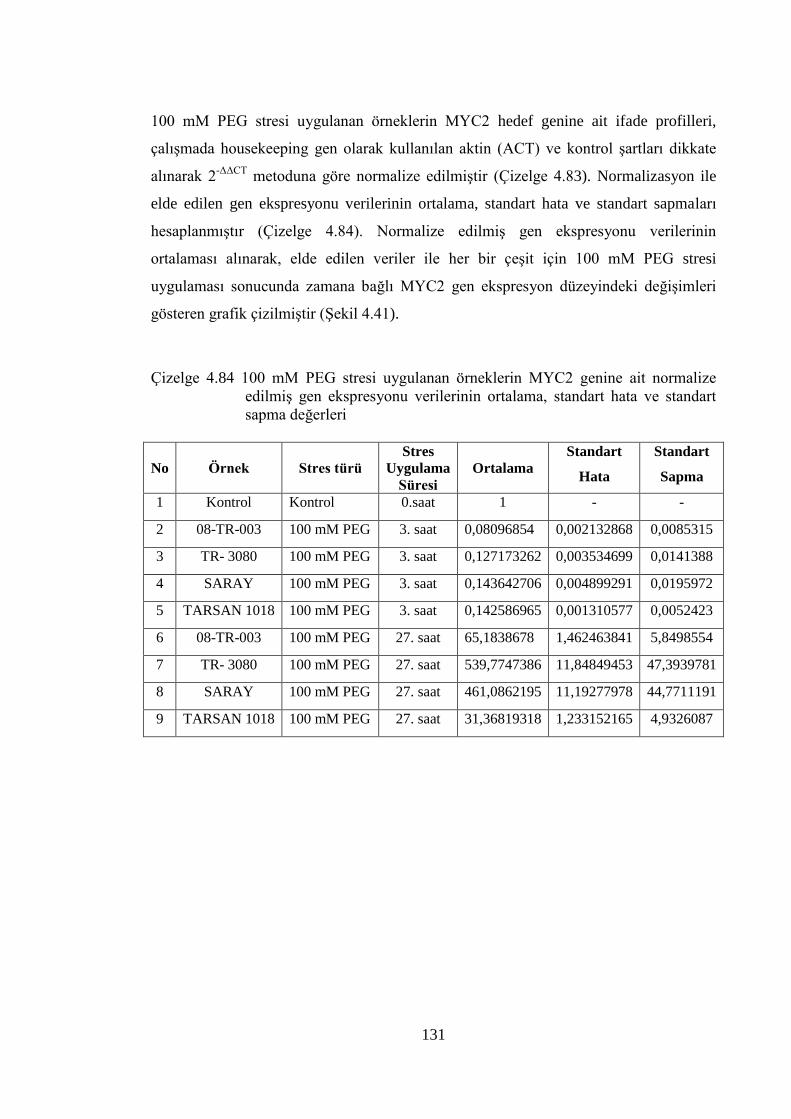
131
100 mM PEG stresi uygulanan örneklerin MYC2 hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.83). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.84). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 100 mM PEG stresi
uygulaması sonucunda zamana bağlı MYC2 gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.41).
Çizelge 4.84 100 mM PEG stresi uygulanan örneklerin MYC2 genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 100 mM PEG 3. saat 0,08096854 0,002132868 0,0085315
3 TR- 3080 100 mM PEG 3. saat 0,127173262 0,003534699 0,0141388
4 SARAY 100 mM PEG 3. saat 0,143642706 0,004899291 0,0195972
5 TARSAN 1018 100 mM PEG 3. saat 0,142586965 0,001310577 0,0052423
6 08-TR-003 100 mM PEG 27. saat 65,1838678 1,462463841 5,8498554
7 TR- 3080 100 mM PEG 27. saat 539,7747386 11,84849453 47,3939781
8 SARAY 100 mM PEG 27. saat 461,0862195 11,19277978 44,7711191
9 TARSAN 1018 100 mM PEG 27. saat 31,36819318 1,233152165 4,9326087
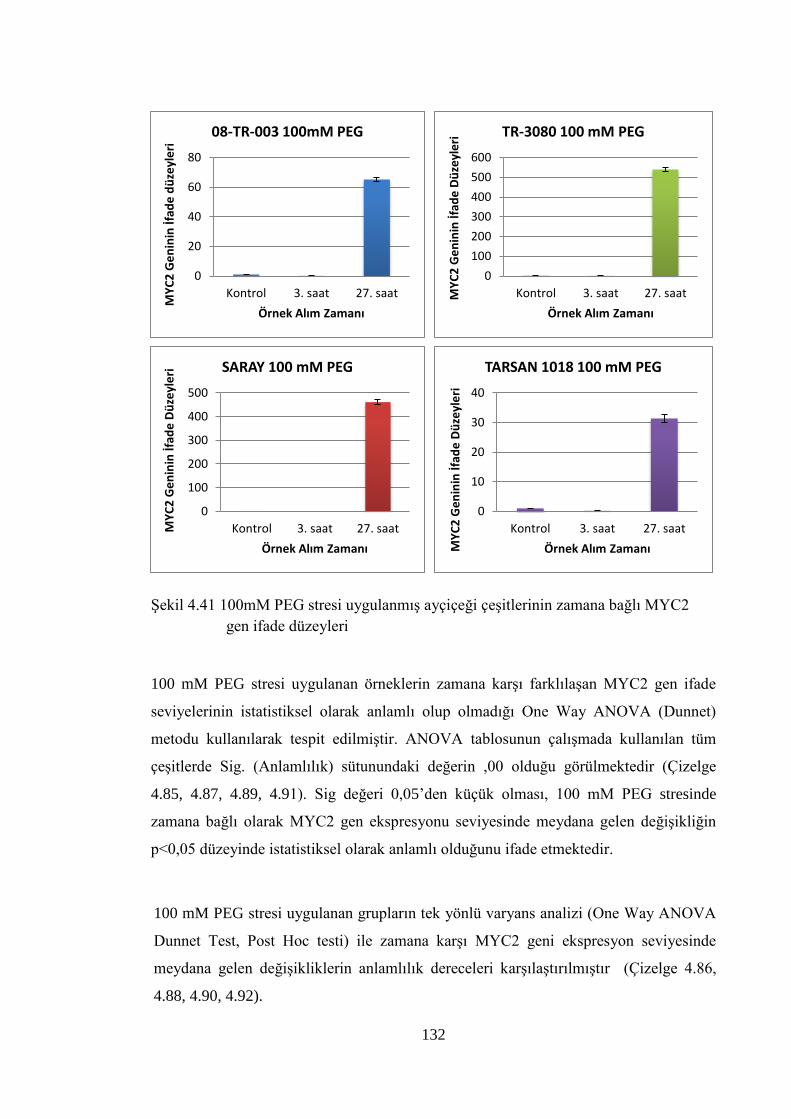
132
Şekil 4.41 100mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı MYC2
gen ifade düzeyleri
100 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan MYC2 gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.85, 4.87, 4.89, 4.91). Sig değeri 0,05’den küçük olması, 100 mM PEG stresinde
zamana bağlı olarak MYC2 gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
100 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.86,
4.88, 4.90, 4.92).
0
20
40
60
80
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 100mM PEG
0
100
200
300
400
500
600
Kontrol 3. saat 27. saatMY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 100 mM PEG
0
100
200
300
400
500
Kontrol 3. saat 27. saatMY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
SARAY 100 mM PEG
0
10
20
30
40
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 100 mM PEG
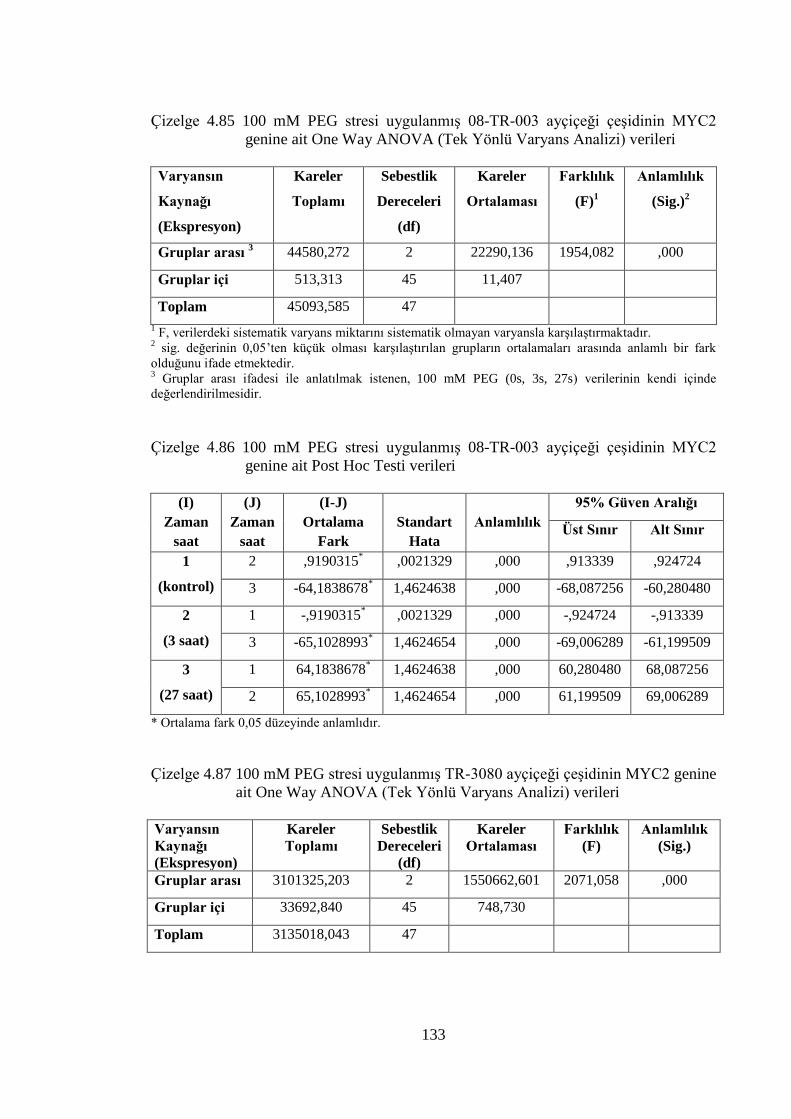
133
Çizelge 4.85 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 44580,272 2 22290,136 1954,082 ,000
Gruplar içi 513,313 45 11,407
Toplam 45093,585 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
100 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.86 100 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,9190315* ,0021329 ,000 ,913339 ,924724
3 -64,1838678* 1,4624638 ,000 -68,087256 -60,280480
2
(3 saat)
1 -,9190315* ,0021329 ,000 -,924724 -,913339
3 -65,1028993* 1,4624654 ,000 -69,006289 -61,199509
3
(27 saat)
1 64,1838678* 1,4624638 ,000 60,280480 68,087256
2 65,1028993* 1,4624654 ,000 61,199509 69,006289
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.87 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2 genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 3101325,203 2 1550662,601 2071,058 ,000
Gruplar içi 33692,840 45 748,730
Toplam 3135018,043 47
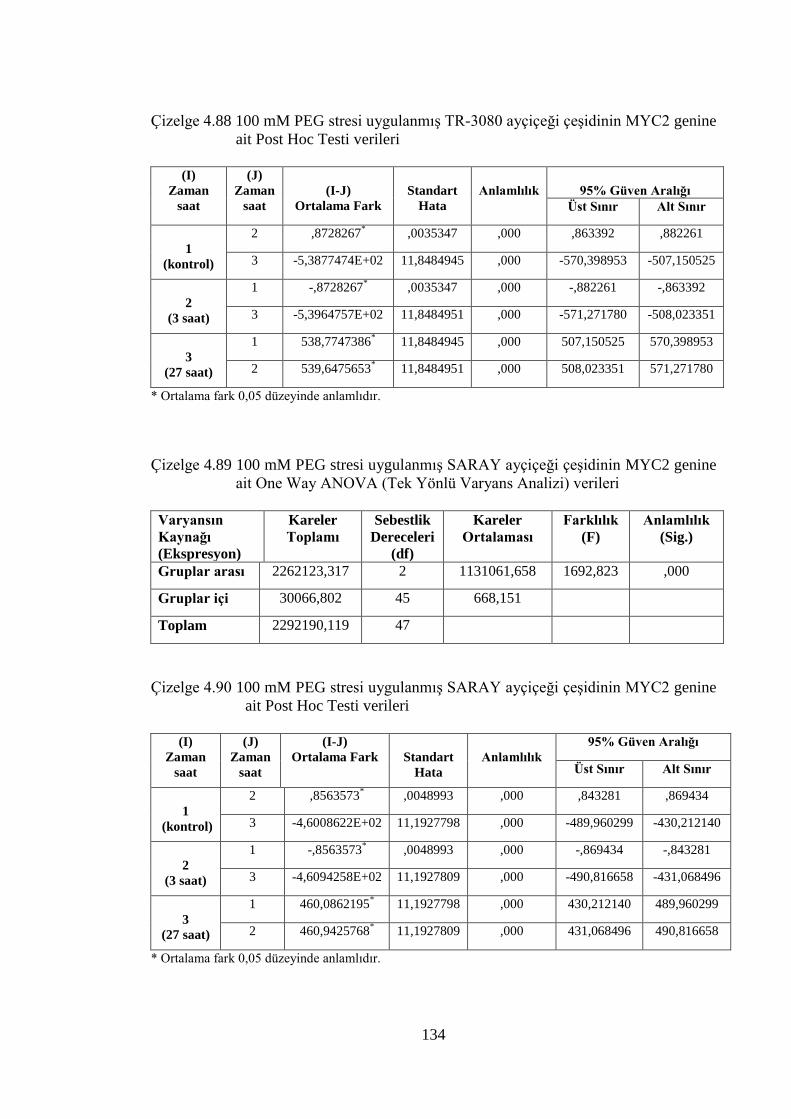
134
Çizelge 4.88 100 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2 genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8728267* ,0035347 ,000 ,863392 ,882261
3 -5,3877474E+02 11,8484945 ,000 -570,398953 -507,150525
2
(3 saat)
1 -,8728267* ,0035347 ,000 -,882261 -,863392
3 -5,3964757E+02 11,8484951 ,000 -571,271780 -508,023351
3
(27 saat)
1 538,7747386* 11,8484945 ,000 507,150525 570,398953
2 539,6475653* 11,8484951 ,000 508,023351 571,271780
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.89 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 2262123,317 2 1131061,658 1692,823 ,000
Gruplar içi 30066,802 45 668,151
Toplam 2292190,119 47
Çizelge 4.90 100 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8563573* ,0048993 ,000 ,843281 ,869434
3 -4,6008622E+02 11,1927798 ,000 -489,960299 -430,212140
2
(3 saat)
1 -,8563573* ,0048993 ,000 -,869434 -,843281
3 -4,6094258E+02 11,1927809 ,000 -490,816658 -431,068496
3
(27 saat)
1 460,0862195* 11,1927798 ,000 430,212140 489,960299
2 460,9425768* 11,1927809 ,000 431,068496 490,816658
* Ortalama fark 0,05 düzeyinde anlamlıdır.
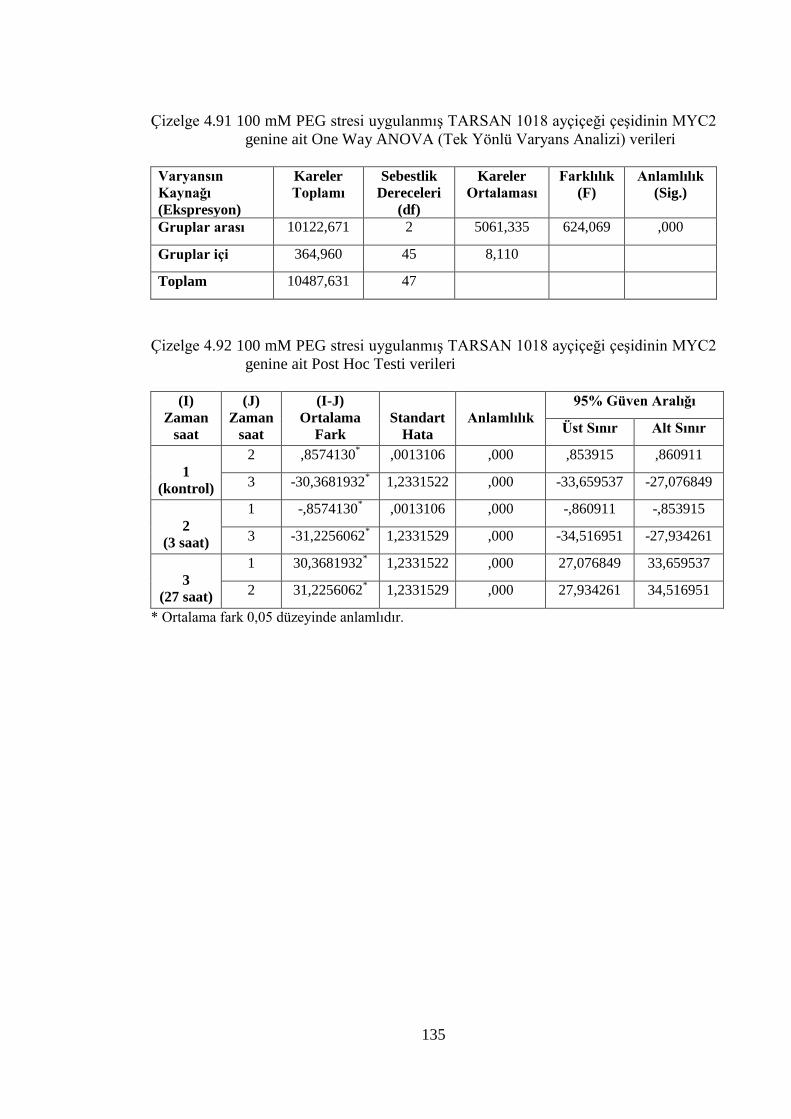
135
Çizelge 4.91 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 10122,671 2 5061,335 624,069 ,000
Gruplar içi 364,960 45 8,110
Toplam 10487,631 47
Çizelge 4.92 100 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8574130* ,0013106 ,000 ,853915 ,860911
3 -30,3681932* 1,2331522 ,000 -33,659537 -27,076849
2
(3 saat)
1 -,8574130* ,0013106 ,000 -,860911 -,853915
3 -31,2256062* 1,2331529 ,000 -34,516951 -27,934261
3
(27 saat)
1 30,3681932* 1,2331522 ,000 27,076849 33,659537
2 31,2256062* 1,2331529 ,000 27,934261 34,516951
* Ortalama fark 0,05 düzeyinde anlamlıdır.
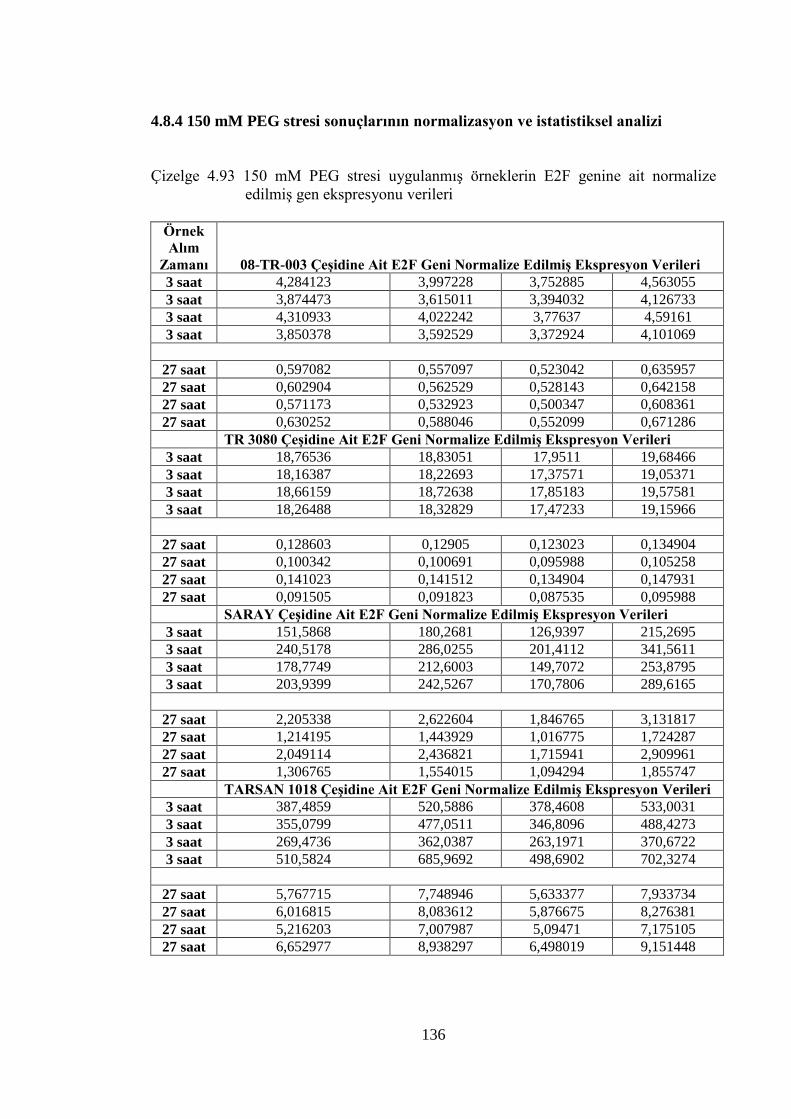
136
4.8.4 150 mM PEG stresi sonuçlarının normalizasyon ve istatistiksel analizi
Çizelge 4.93 150 mM PEG stresi uygulanmış örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 4,284123 3,997228 3,752885 4,563055
3 saat 3,874473 3,615011 3,394032 4,126733
3 saat 4,310933 4,022242 3,77637 4,59161
3 saat 3,850378 3,592529 3,372924 4,101069
27 saat 0,597082 0,557097 0,523042 0,635957
27 saat 0,602904 0,562529 0,528143 0,642158
27 saat 0,571173 0,532923 0,500347 0,608361
27 saat 0,630252 0,588046 0,552099 0,671286
TR 3080 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 18,76536 18,83051 17,9511 19,68466
3 saat 18,16387 18,22693 17,37571 19,05371
3 saat 18,66159 18,72638 17,85183 19,57581
3 saat 18,26488 18,32829 17,47233 19,15966
27 saat 0,128603 0,12905 0,123023 0,134904
27 saat 0,100342 0,100691 0,095988 0,105258
27 saat 0,141023 0,141512 0,134904 0,147931
27 saat 0,091505 0,091823 0,087535 0,095988
SARAY Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 151,5868 180,2681 126,9397 215,2695
3 saat 240,5178 286,0255 201,4112 341,5611
3 saat 178,7749 212,6003 149,7072 253,8795
3 saat 203,9399 242,5267 170,7806 289,6165
27 saat 2,205338 2,622604 1,846765 3,131817
27 saat 1,214195 1,443929 1,016775 1,724287
27 saat 2,049114 2,436821 1,715941 2,909961
27 saat 1,306765 1,554015 1,094294 1,855747
TARSAN 1018 Çeşidine Ait E2F Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 387,4859 520,5886 378,4608 533,0031
3 saat 355,0799 477,0511 346,8096 488,4273
3 saat 269,4736 362,0387 263,1971 370,6722
3 saat 510,5824 685,9692 498,6902 702,3274
27 saat 5,767715 7,748946 5,633377 7,933734
27 saat 6,016815 8,083612 5,876675 8,276381
27 saat 5,216203 7,007987 5,09471 7,175105
27 saat 6,652977 8,938297 6,498019 9,151448

137
150 mM PEG stresi uygulanan örneklerin E2F hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.93). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.94). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM PEG stresi
uygulaması sonucunda zamana bağlı E2F gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.42).
Çizelge 4.94 150 mM PEG stresi uygulanan örneklerin E2F genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM PEG 3. saat 3,951599637 0,09259287 0,3703715
3 TR- 3080 150 mM PEG 3. saat 18,50578793 0,169224686 0,6768987
4 SARAY 150 mM PEG 3. saat 215,3378283 14,48769148 57,9507659
5 TARSAN 1018 150 mM PEG 3. saat 446,8660674 32,24349114 128,9739645
6 08-TR-003 150 mM PEG 27. saat 0,581462414 0,012193699 0,0487748
7 TR- 3080 150 mM PEG 27. saat 0,115629985 0,005314393 0,0212576
8 SARAY 150 mM PEG 27. saat 1,883022941 0,159521949 0,6380878
9 TARSAN 1018 150 mM PEG 27. saat 6,941999938 0,325307121 1,3012285
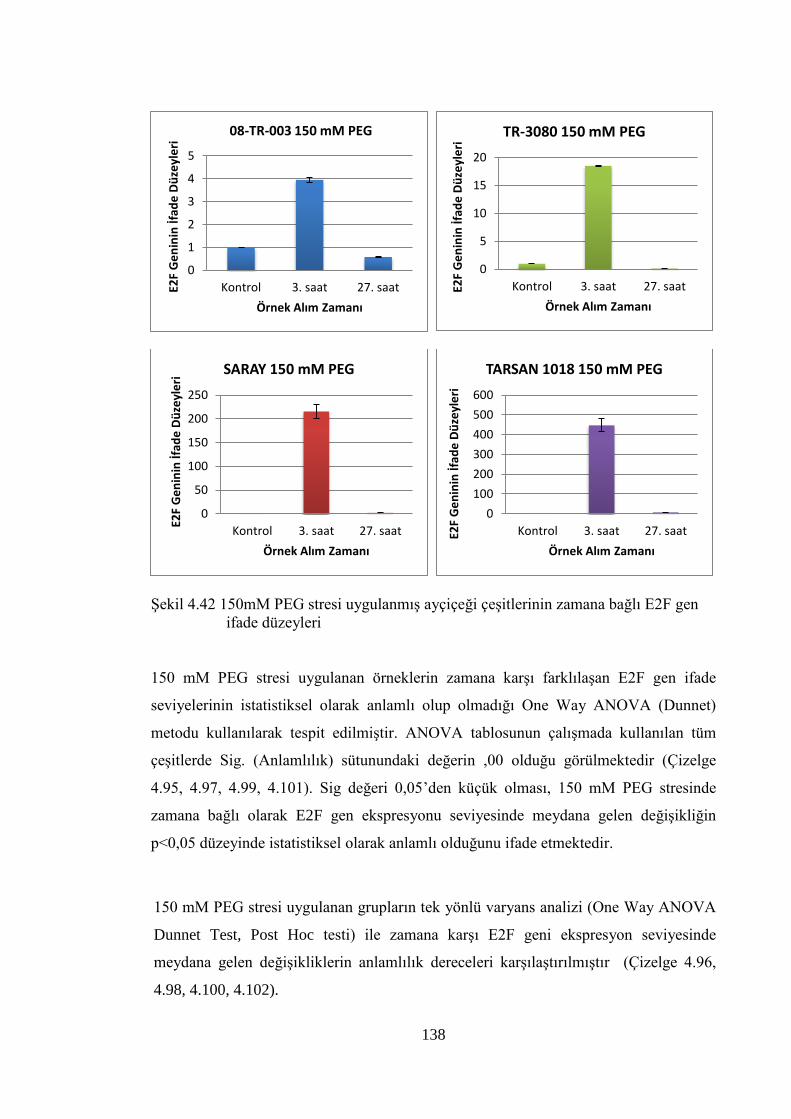
138
Şekil 4.42 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı E2F gen
ifade düzeyleri
150 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.95, 4.97, 4.99, 4.101). Sig değeri 0,05’den küçük olması, 150 mM PEG stresinde
zamana bağlı olarak E2F gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
150 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı E2F geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.96,
4.98, 4.100, 4.102).
0
1
2
3
4
5
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 150 mM PEG
0
5
10
15
20
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 150 mM PEG
0
50
100
150
200
250
Kontrol 3. saat 27. saat
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
SARAY 150 mM PEG
0
100
200
300
400
500
600
Kontrol 3. saat 27. saatE2F
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 150 mM PEG

139
Çizelge 4.95 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 107,973 2 53,987 1160,551 ,000
Gruplar içi 2,093 45 ,047
Toplam 110,066 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
150 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.96 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -2,9515996* ,0925929 ,000 -3,198735 -2,704465
3 ,4185376* ,0121937 ,000 ,385992 ,451083
2
(3 saat)
1 2,9515996* ,0925929 ,000 2,704465 3,198735
3 3,3701372* ,0933923 ,000 3,121824 3,618451
3
(27 saat)
1 -,4185376* ,0121937 ,000 -,451083 -,385992
2 -3,3701372* ,0933923 ,000 -3,618451 -3,121824
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.97 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 3442,307 2 1721,154 11258,107 ,000
Gruplar içi 6,880 45 ,153
Toplam 3449,187 47
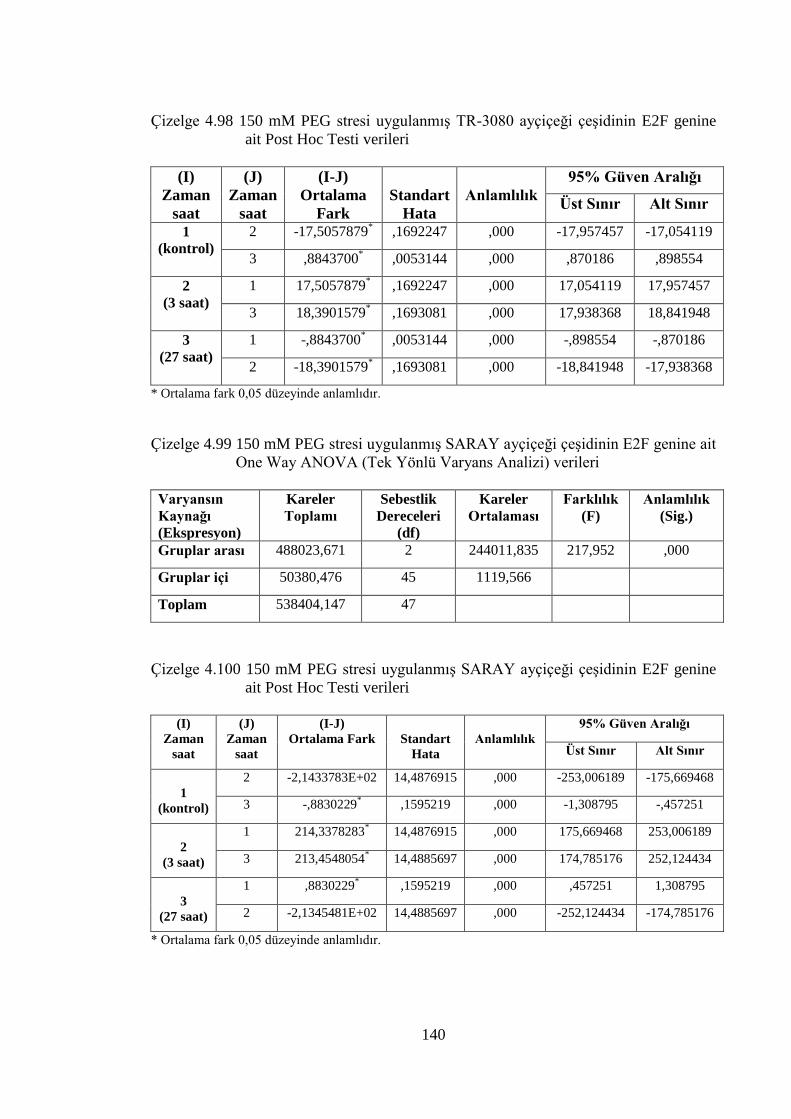
140
Çizelge 4.98 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -17,5057879* ,1692247 ,000 -17,957457 -17,054119
3 ,8843700* ,0053144 ,000 ,870186 ,898554
2
(3 saat)
1 17,5057879* ,1692247 ,000 17,054119 17,957457
3 18,3901579* ,1693081 ,000 17,938368 18,841948
3
(27 saat)
1 -,8843700* ,0053144 ,000 -,898554 -,870186
2 -18,3901579* ,1693081 ,000 -18,841948 -17,938368
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.99 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 488023,671 2 244011,835 217,952 ,000
Gruplar içi 50380,476 45 1119,566
Toplam 538404,147 47
Çizelge 4.100 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -2,1433783E+02 14,4876915 ,000 -253,006189 -175,669468
3 -,8830229* ,1595219 ,000 -1,308795 -,457251
2
(3 saat)
1 214,3378283* 14,4876915 ,000 175,669468 253,006189
3 213,4548054* 14,4885697 ,000 174,785176 252,124434
3
(27 saat)
1 ,8830229* ,1595219 ,000 ,457251 1,308795
2 -2,1345481E+02 14,4885697 ,000 -252,124434 -174,785176
* Ortalama fark 0,05 düzeyinde anlamlıdır.
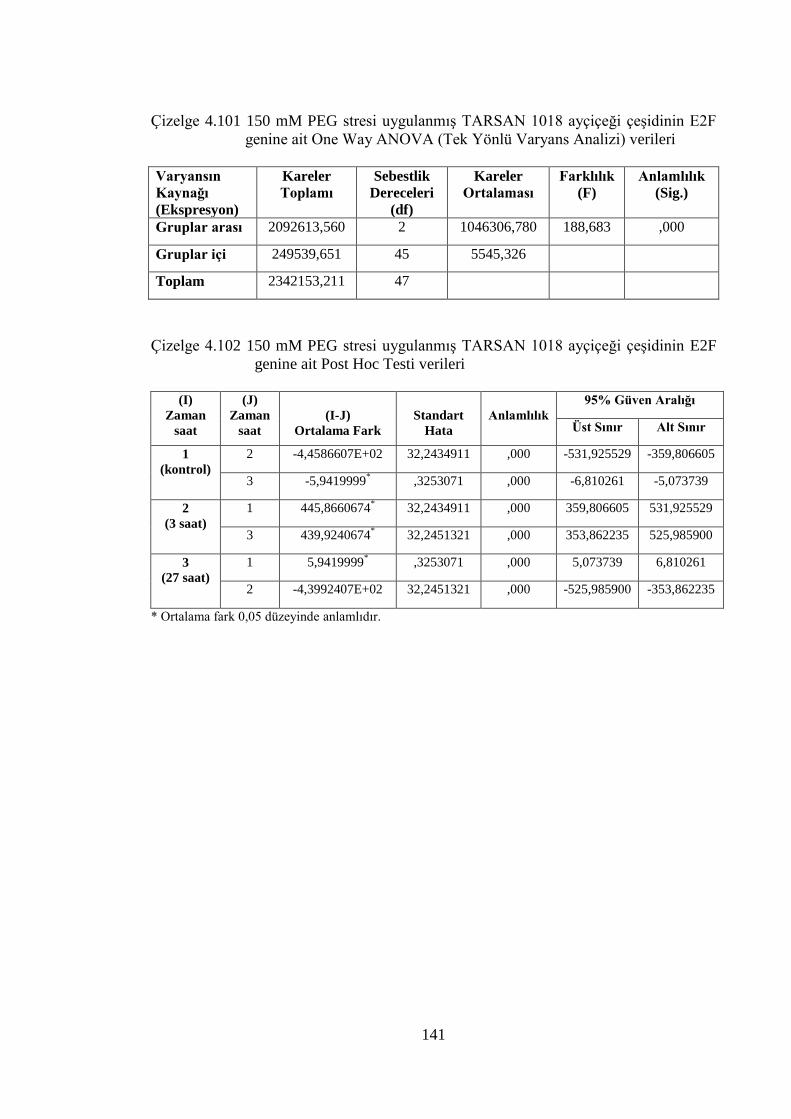
141
Çizelge 4.101 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 2092613,560 2 1046306,780 188,683 ,000
Gruplar içi 249539,651 45 5545,326
Toplam 2342153,211 47
Çizelge 4.102 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -4,4586607E+02 32,2434911 ,000 -531,925529 -359,806605
3 -5,9419999* ,3253071 ,000 -6,810261 -5,073739
2
(3 saat)
1 445,8660674* 32,2434911 ,000 359,806605 531,925529
3 439,9240674* 32,2451321 ,000 353,862235 525,985900
3
(27 saat)
1 5,9419999* ,3253071 ,000 5,073739 6,810261
2 -4,3992407E+02 32,2451321 ,000 -525,985900 -353,862235
* Ortalama fark 0,05 düzeyinde anlamlıdır.
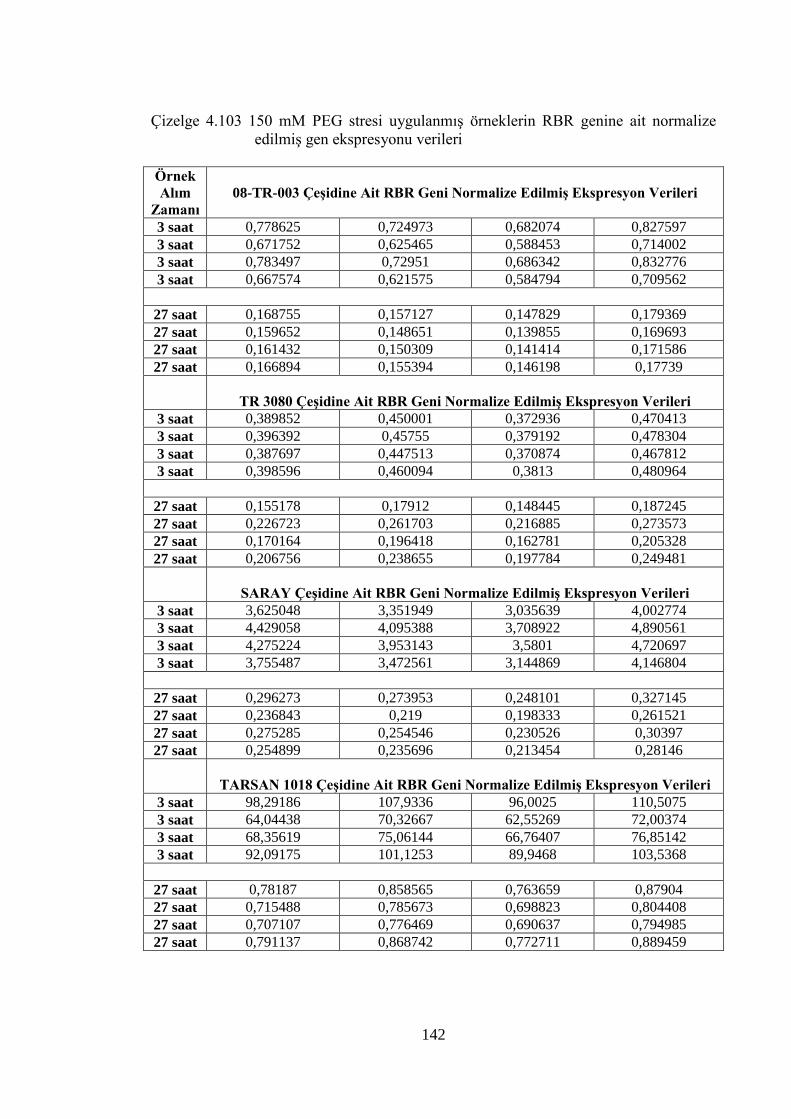
142
Çizelge 4.103 150 mM PEG stresi uygulanmış örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek
Alım
Zamanı
08-TR-003 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,778625 0,724973 0,682074 0,827597
3 saat 0,671752 0,625465 0,588453 0,714002
3 saat 0,783497 0,72951 0,686342 0,832776
3 saat 0,667574 0,621575 0,584794 0,709562
27 saat 0,168755 0,157127 0,147829 0,179369
27 saat 0,159652 0,148651 0,139855 0,169693
27 saat 0,161432 0,150309 0,141414 0,171586
27 saat 0,166894 0,155394 0,146198 0,17739
TR 3080 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,389852 0,450001 0,372936 0,470413
3 saat 0,396392 0,45755 0,379192 0,478304
3 saat 0,387697 0,447513 0,370874 0,467812
3 saat 0,398596 0,460094 0,3813 0,480964
27 saat 0,155178 0,17912 0,148445 0,187245
27 saat 0,226723 0,261703 0,216885 0,273573
27 saat 0,170164 0,196418 0,162781 0,205328
27 saat 0,206756 0,238655 0,197784 0,249481
SARAY Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 3,625048 3,351949 3,035639 4,002774
3 saat 4,429058 4,095388 3,708922 4,890561
3 saat 4,275224 3,953143 3,5801 4,720697
3 saat 3,755487 3,472561 3,144869 4,146804
27 saat 0,296273 0,273953 0,248101 0,327145
27 saat 0,236843 0,219 0,198333 0,261521
27 saat 0,275285 0,254546 0,230526 0,30397
27 saat 0,254899 0,235696 0,213454 0,28146
TARSAN 1018 Çeşidine Ait RBR Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 98,29186 107,9336 96,0025 110,5075
3 saat 64,04438 70,32667 62,55269 72,00374
3 saat 68,35619 75,06144 66,76407 76,85142
3 saat 92,09175 101,1253 89,9468 103,5368
27 saat 0,78187 0,858565 0,763659 0,87904
27 saat 0,715488 0,785673 0,698823 0,804408
27 saat 0,707107 0,776469 0,690637 0,794985
27 saat 0,791137 0,868742 0,772711 0,889459
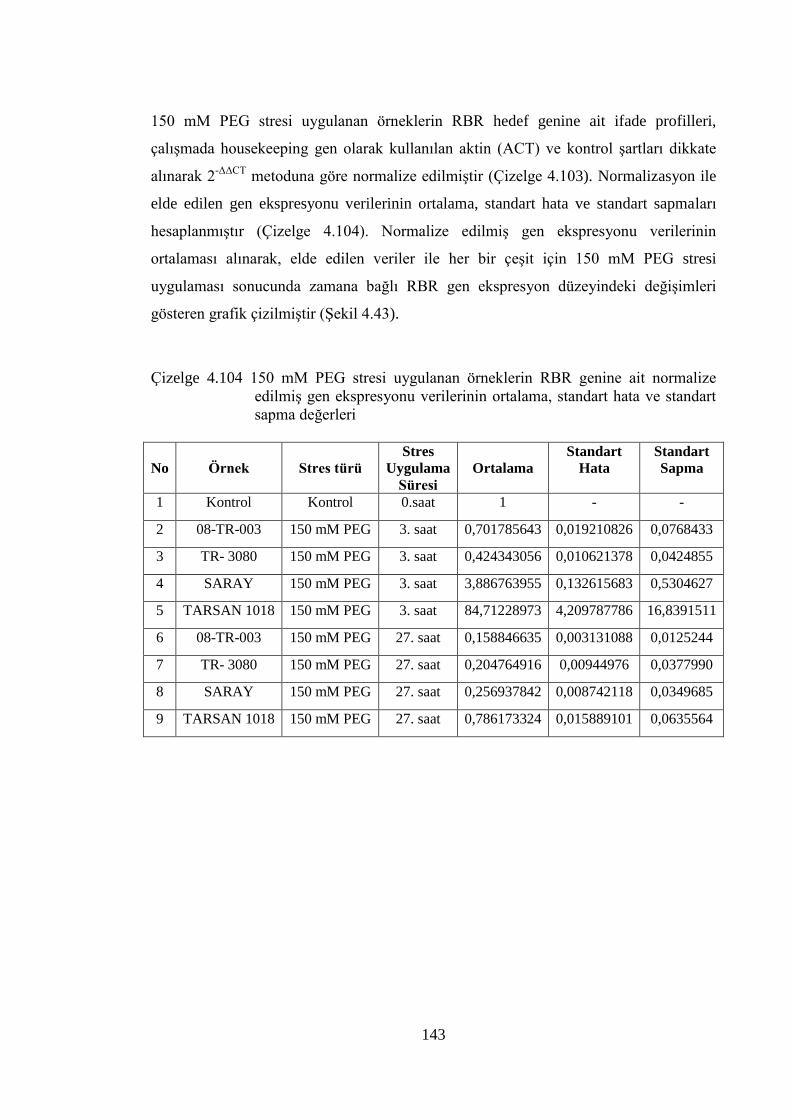
143
150 mM PEG stresi uygulanan örneklerin RBR hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.103). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.104). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM PEG stresi
uygulaması sonucunda zamana bağlı RBR gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.43).
Çizelge 4.104 150 mM PEG stresi uygulanan örneklerin RBR genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM PEG 3. saat 0,701785643 0,019210826 0,0768433
3 TR- 3080 150 mM PEG 3. saat 0,424343056 0,010621378 0,0424855
4 SARAY 150 mM PEG 3. saat 3,886763955 0,132615683 0,5304627
5 TARSAN 1018 150 mM PEG 3. saat 84,71228973 4,209787786 16,8391511
6 08-TR-003 150 mM PEG 27. saat 0,158846635 0,003131088 0,0125244
7 TR- 3080 150 mM PEG 27. saat 0,204764916 0,00944976 0,0377990
8 SARAY 150 mM PEG 27. saat 0,256937842 0,008742118 0,0349685
9 TARSAN 1018 150 mM PEG 27. saat 0,786173324 0,015889101 0,0635564
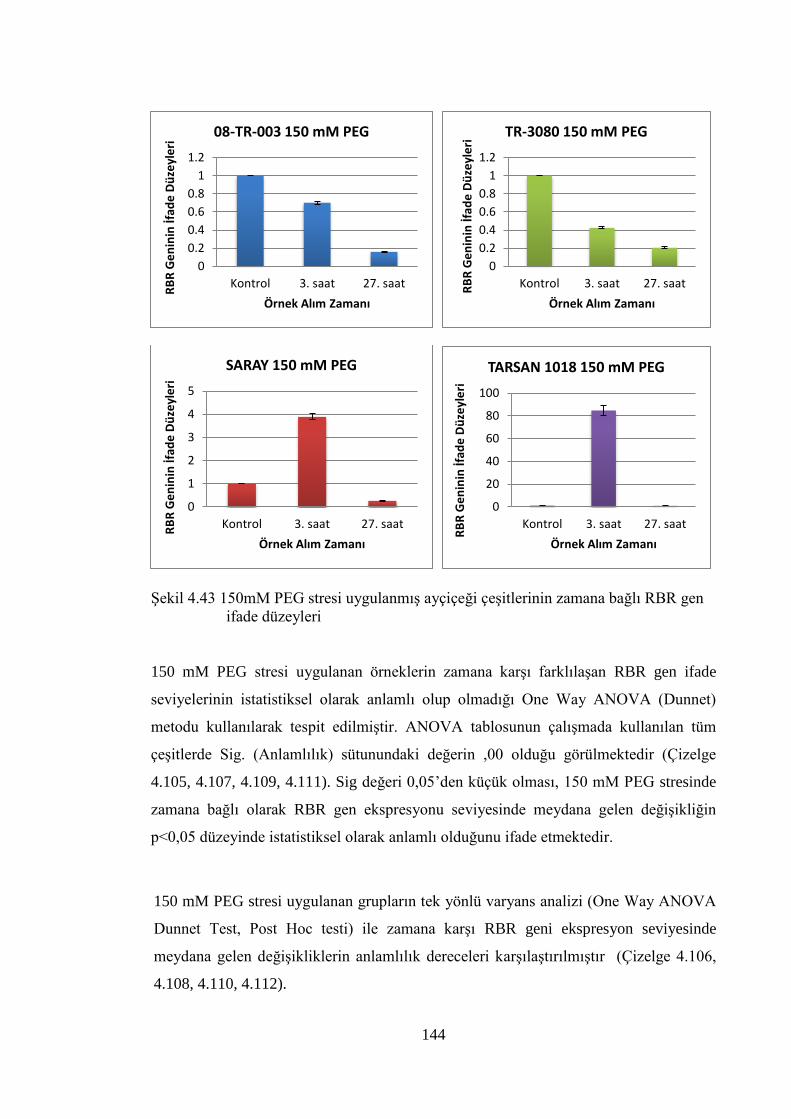
144
Şekil 4.43 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı RBR gen
ifade düzeyleri
150 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan RBR gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.105, 4.107, 4.109, 4.111). Sig değeri 0,05’den küçük olması, 150 mM PEG stresinde
zamana bağlı olarak RBR gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
150 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı RBR geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.106,
4.108, 4.110, 4.112).
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
08-TR-003 150 mM PEG
0
0.2
0.4
0.6
0.8
1
1.2
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TR-3080 150 mM PEG
0
1
2
3
4
5
Kontrol 3. saat 27. saatRB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
SARAY 150 mM PEG
0
20
40
60
80
100
Kontrol 3. saat 27. saat
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018 150 mM PEG
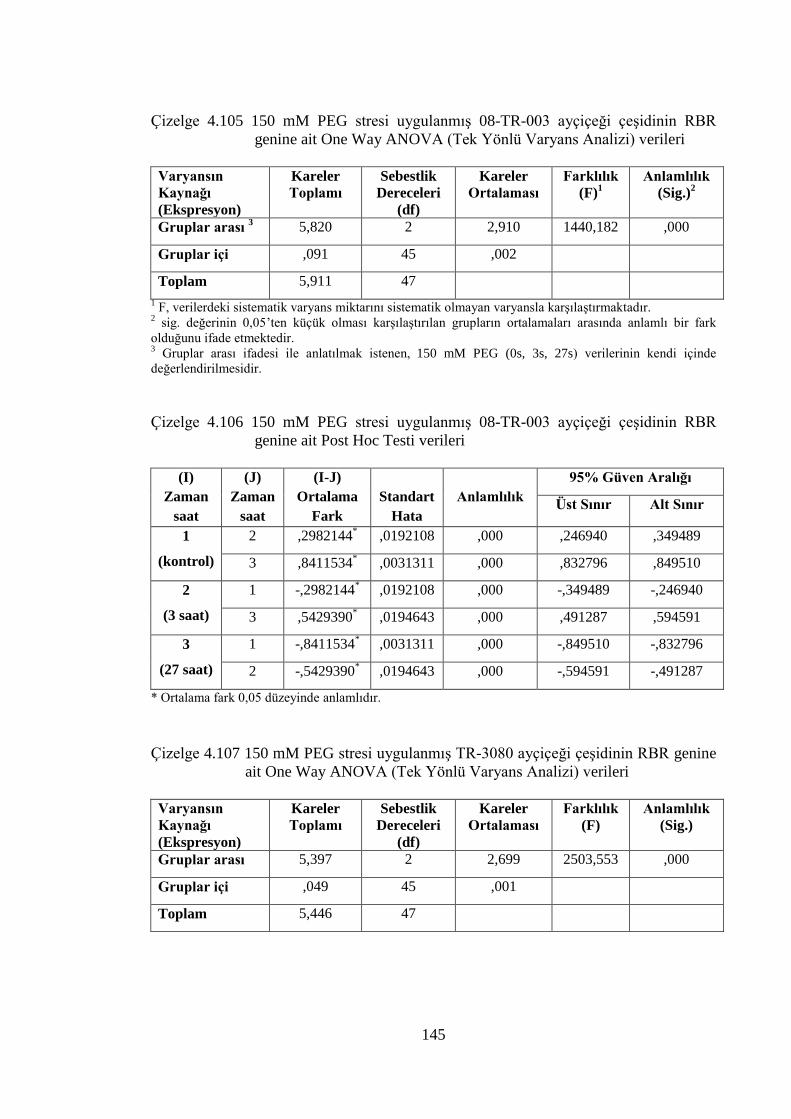
145
Çizelge 4.105 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 5,820 2 2,910 1440,182 ,000
Gruplar içi ,091 45 ,002
Toplam 5,911 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
150 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.106 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,2982144* ,0192108 ,000 ,246940 ,349489
3 ,8411534* ,0031311 ,000 ,832796 ,849510
2
(3 saat)
1 -,2982144* ,0192108 ,000 -,349489 -,246940
3 ,5429390* ,0194643 ,000 ,491287 ,594591
3
(27 saat)
1 -,8411534* ,0031311 ,000 -,849510 -,832796
2 -,5429390* ,0194643 ,000 -,594591 -,491287
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.107 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 5,397 2 2,699 2503,553 ,000
Gruplar içi ,049 45 ,001
Toplam 5,446 47
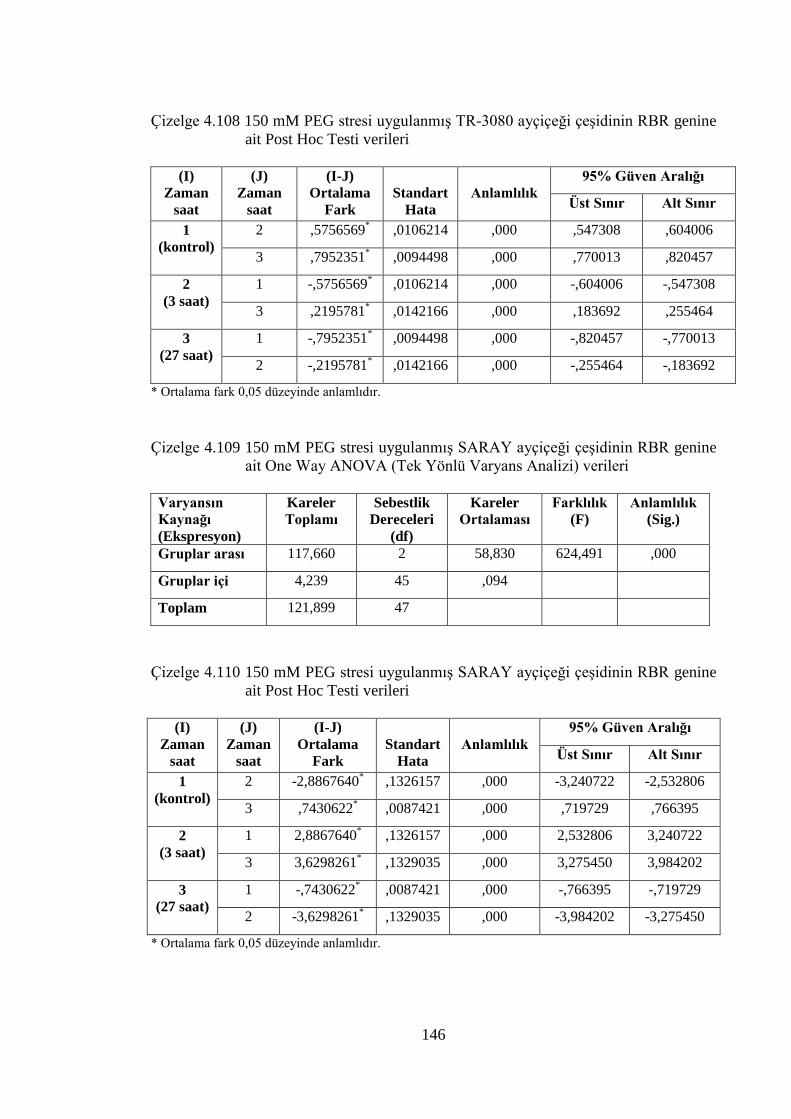
146
Çizelge 4.108 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,5756569* ,0106214 ,000 ,547308 ,604006
3 ,7952351* ,0094498 ,000 ,770013 ,820457
2
(3 saat)
1 -,5756569* ,0106214 ,000 -,604006 -,547308
3 ,2195781* ,0142166 ,000 ,183692 ,255464
3
(27 saat)
1 -,7952351* ,0094498 ,000 -,820457 -,770013
2 -,2195781* ,0142166 ,000 -,255464 -,183692
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.109 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 117,660 2 58,830 624,491 ,000
Gruplar içi 4,239 45 ,094
Toplam 121,899 47
Çizelge 4.110 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -2,8867640* ,1326157 ,000 -3,240722 -2,532806
3 ,7430622* ,0087421 ,000 ,719729 ,766395
2
(3 saat)
1 2,8867640* ,1326157 ,000 2,532806 3,240722
3 3,6298261* ,1329035 ,000 3,275450 3,984202
3
(27 saat)
1 -,7430622* ,0087421 ,000 -,766395 -,719729
2 -3,6298261* ,1329035 ,000 -3,984202 -3,275450
* Ortalama fark 0,05 düzeyinde anlamlıdır.
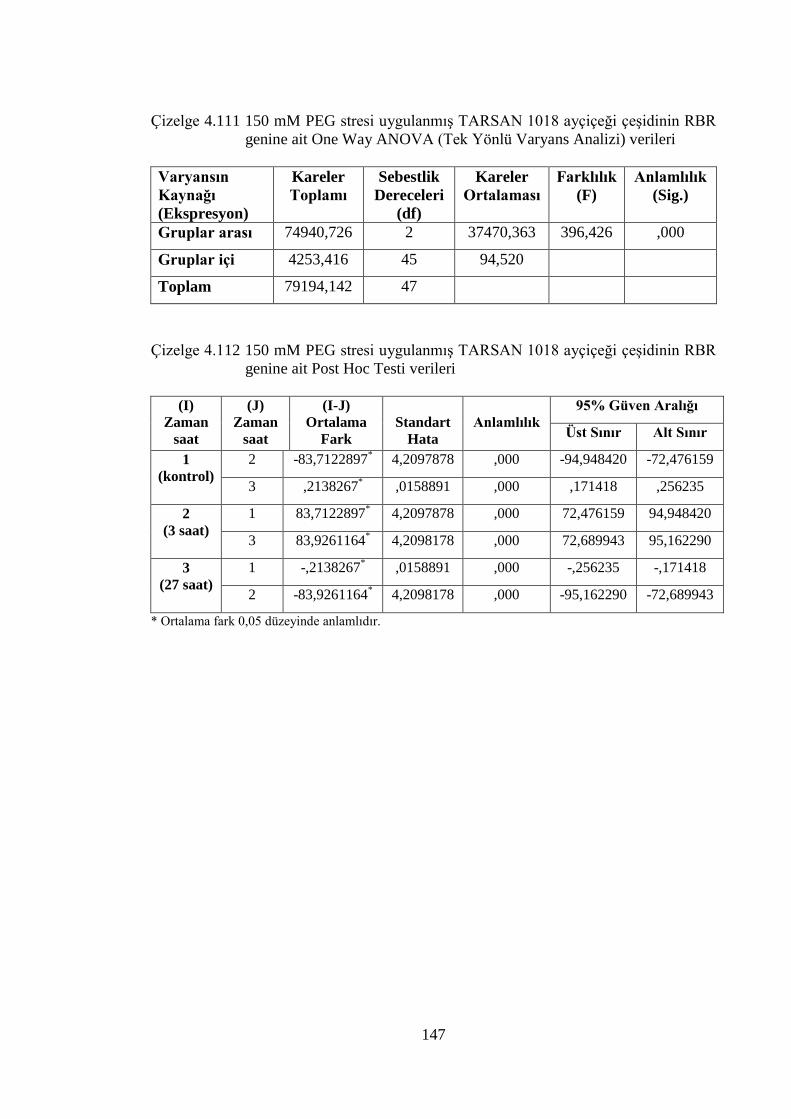
147
Çizelge 4.111 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 74940,726 2 37470,363 396,426 ,000
Gruplar içi 4253,416 45 94,520
Toplam 79194,142 47
Çizelge 4.112 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -83,7122897* 4,2097878 ,000 -94,948420 -72,476159
3 ,2138267* ,0158891 ,000 ,171418 ,256235
2
(3 saat)
1 83,7122897* 4,2097878 ,000 72,476159 94,948420
3 83,9261164* 4,2098178 ,000 72,689943 95,162290
3
(27 saat)
1 -,2138267* ,0158891 ,000 -,256235 -,171418
2 -83,9261164* 4,2098178 ,000 -95,162290 -72,689943
* Ortalama fark 0,05 düzeyinde anlamlıdır.
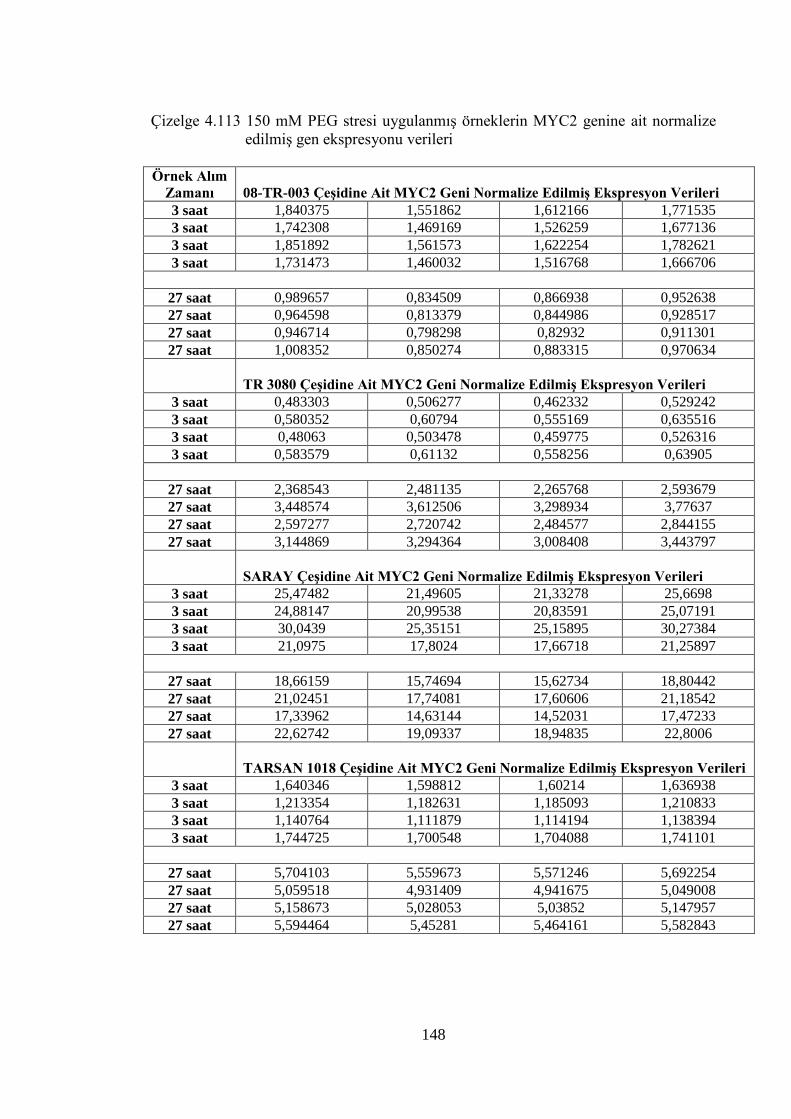
148
Çizelge 4.113 150 mM PEG stresi uygulanmış örneklerin MYC2 genine ait normalize
edilmiş gen ekspresyonu verileri
Örnek Alım
Zamanı
08-TR-003 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,840375 1,551862 1,612166 1,771535
3 saat 1,742308 1,469169 1,526259 1,677136
3 saat 1,851892 1,561573 1,622254 1,782621
3 saat 1,731473 1,460032 1,516768 1,666706
27 saat 0,989657 0,834509 0,866938 0,952638
27 saat 0,964598 0,813379 0,844986 0,928517
27 saat 0,946714 0,798298 0,82932 0,911301
27 saat 1,008352 0,850274 0,883315 0,970634
TR 3080 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 0,483303 0,506277 0,462332 0,529242
3 saat 0,580352 0,60794 0,555169 0,635516
3 saat 0,48063 0,503478 0,459775 0,526316
3 saat 0,583579 0,61132 0,558256 0,63905
27 saat 2,368543 2,481135 2,265768 2,593679
27 saat 3,448574 3,612506 3,298934 3,77637
27 saat 2,597277 2,720742 2,484577 2,844155
27 saat 3,144869 3,294364 3,008408 3,443797
SARAY Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 25,47482 21,49605 21,33278 25,6698
3 saat 24,88147 20,99538 20,83591 25,07191
3 saat 30,0439 25,35151 25,15895 30,27384
3 saat 21,0975 17,8024 17,66718 21,25897
27 saat 18,66159 15,74694 15,62734 18,80442
27 saat 21,02451 17,74081 17,60606 21,18542
27 saat 17,33962 14,63144 14,52031 17,47233
27 saat 22,62742 19,09337 18,94835 22,8006
TARSAN 1018 Çeşidine Ait MYC2 Geni Normalize Edilmiş Ekspresyon Verileri
3 saat 1,640346 1,598812 1,60214 1,636938
3 saat 1,213354 1,182631 1,185093 1,210833
3 saat 1,140764 1,111879 1,114194 1,138394
3 saat 1,744725 1,700548 1,704088 1,741101
27 saat 5,704103 5,559673 5,571246 5,692254
27 saat 5,059518 4,931409 4,941675 5,049008
27 saat 5,158673 5,028053 5,03852 5,147957
27 saat 5,594464 5,45281 5,464161 5,582843
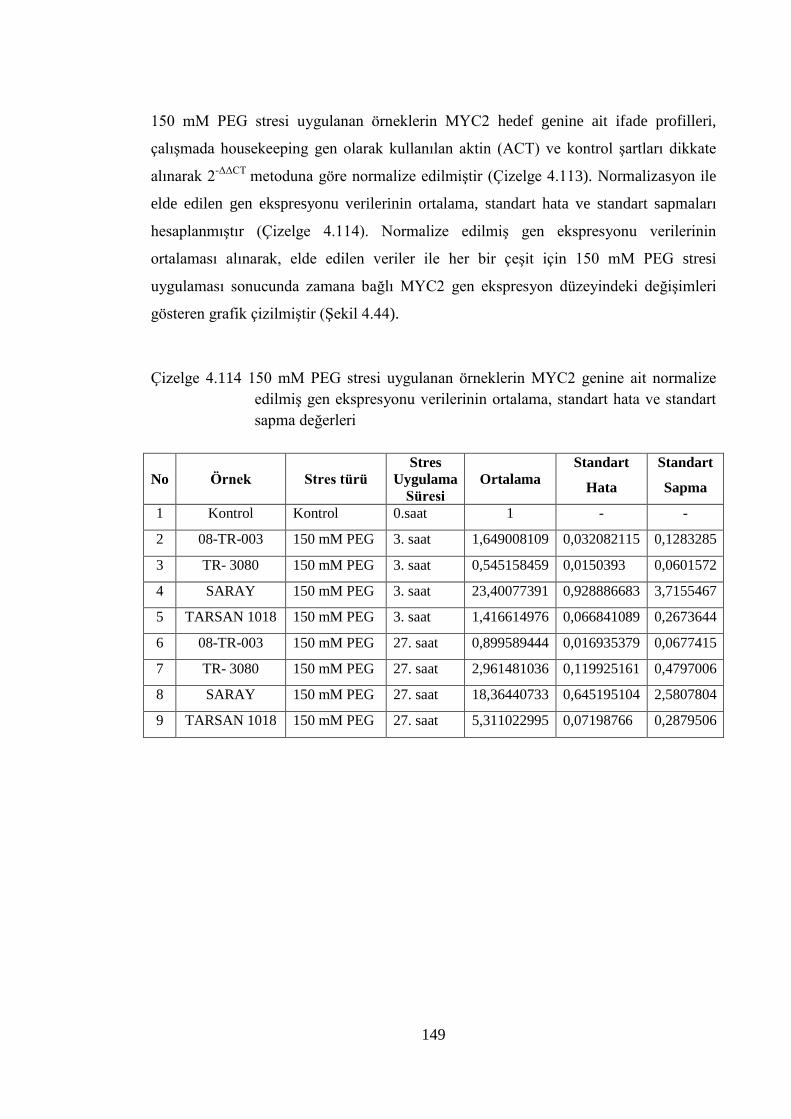
149
150 mM PEG stresi uygulanan örneklerin MYC2 hedef genine ait ifade profilleri,
çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol şartları dikkate
alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.113). Normalizasyon ile
elde edilen gen ekspresyonu verilerinin ortalama, standart hata ve standart sapmaları
hesaplanmıştır (Çizelge 4.114). Normalize edilmiş gen ekspresyonu verilerinin
ortalaması alınarak, elde edilen veriler ile her bir çeşit için 150 mM PEG stresi
uygulaması sonucunda zamana bağlı MYC2 gen ekspresyon düzeyindeki değişimleri
gösteren grafik çizilmiştir (Şekil 4.44).
Çizelge 4.114 150 mM PEG stresi uygulanan örneklerin MYC2 genine ait normalize
edilmiş gen ekspresyonu verilerinin ortalama, standart hata ve standart
sapma değerleri
No
Örnek
Stres türü
Stres
Uygulama
Süresi
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 0.saat 1 - -
2 08-TR-003 150 mM PEG 3. saat 1,649008109 0,032082115 0,1283285
3 TR- 3080 150 mM PEG 3. saat 0,545158459 0,0150393 0,0601572
4 SARAY 150 mM PEG 3. saat 23,40077391 0,928886683 3,7155467
5 TARSAN 1018 150 mM PEG 3. saat 1,416614976 0,066841089 0,2673644
6 08-TR-003 150 mM PEG 27. saat 0,899589444 0,016935379 0,0677415
7 TR- 3080 150 mM PEG 27. saat 2,961481036 0,119925161 0,4797006
8 SARAY 150 mM PEG 27. saat 18,36440733 0,645195104 2,5807804
9 TARSAN 1018 150 mM PEG 27. saat 5,311022995 0,07198766 0,2879506
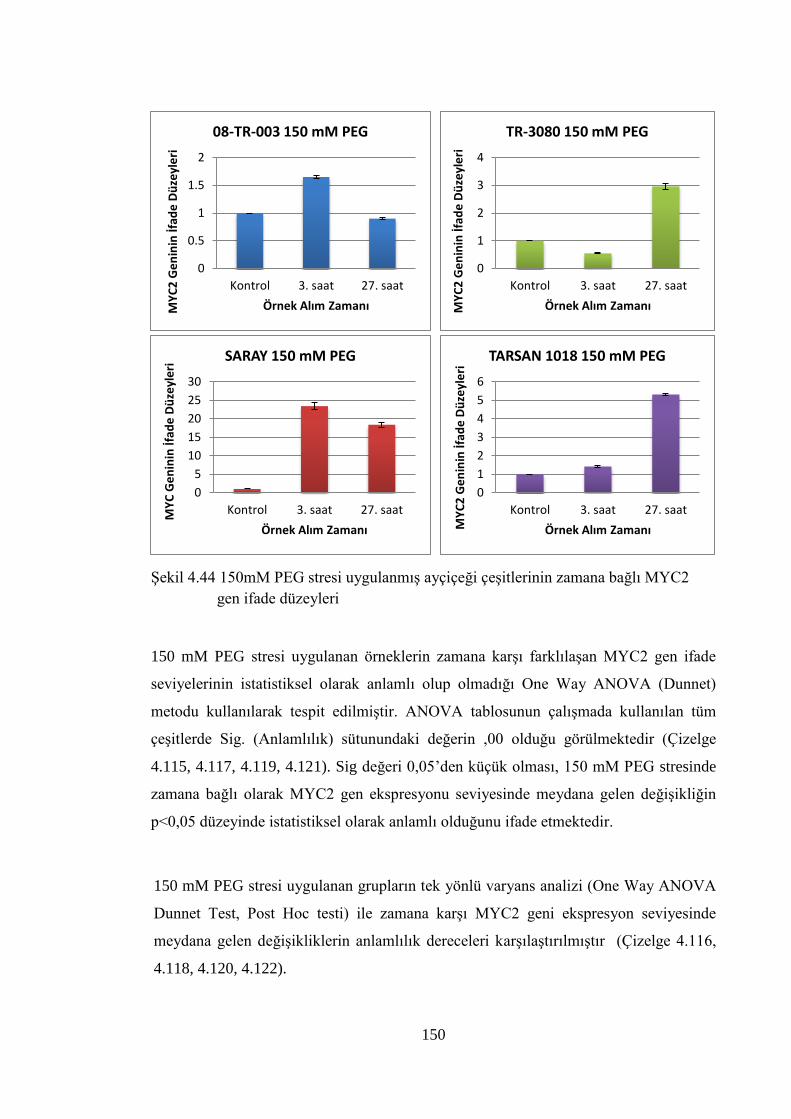
150
Şekil 4.44 150mM PEG stresi uygulanmış ayçiçeği çeşitlerinin zamana bağlı MYC2
gen ifade düzeyleri
150 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan MYC2 gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.115, 4.117, 4.119, 4.121). Sig değeri 0,05’den küçük olması, 150 mM PEG stresinde
zamana bağlı olarak MYC2 gen ekspresyonu seviyesinde meydana gelen değişikliğin
p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu ifade etmektedir.
150 mM PEG stresi uygulanan grupların tek yönlü varyans analizi (One Way ANOVA
Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni ekspresyon seviyesinde
meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır (Çizelge 4.116,
4.118, 4.120, 4.122).
0
0.5
1
1.5
2
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
08-TR-003 150 mM PEG
0
1
2
3
4
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TR-3080 150 mM PEG
0
5
10
15
20
25
30
Kontrol 3. saat 27. saatMY
C G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
SARAY 150 mM PEG
0
1
2
3
4
5
6
Kontrol 3. saat 27. saat
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
Örnek Alım Zamanı
TARSAN 1018 150 mM PEG
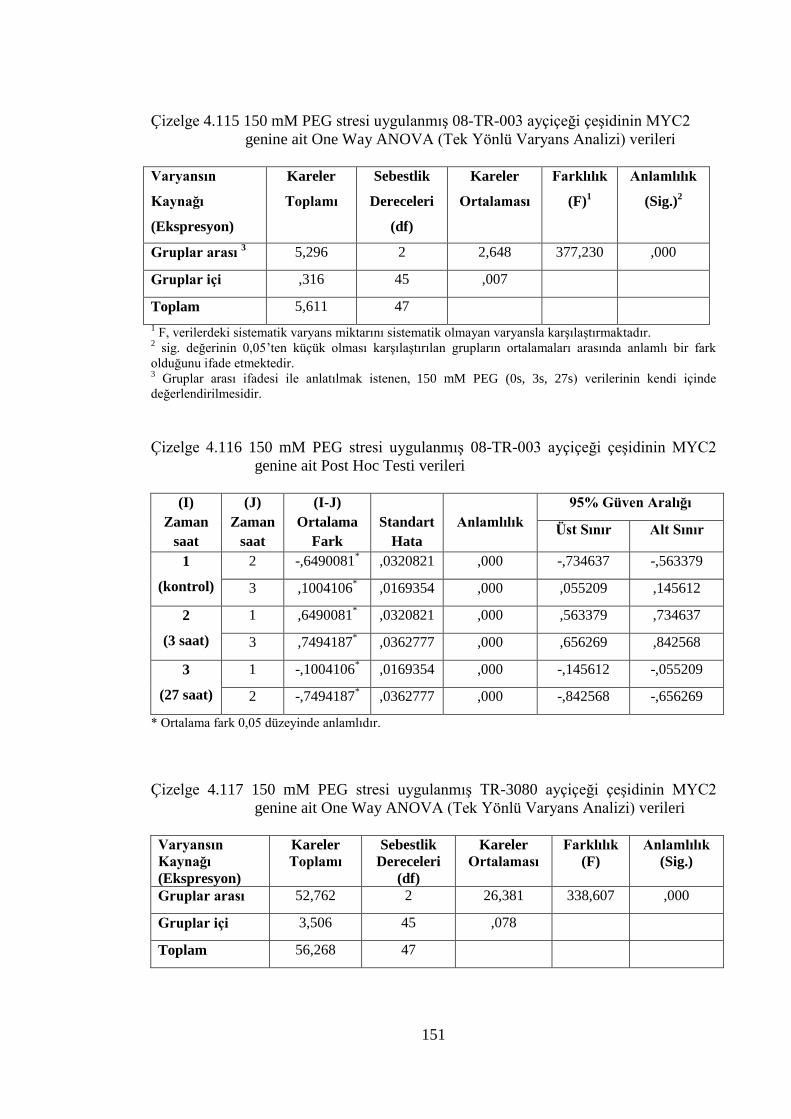
151
Çizelge 4.115 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)1
Anlamlılık
(Sig.)2
Gruplar arası 3 5,296 2 2,648 377,230 ,000
Gruplar içi ,316 45 ,007
Toplam 5,611 47
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
150 mM PEG (0s, 3s, 27s) verilerinin kendi içinde
değerlendirilmesidir.
Çizelge 4.116 150 mM PEG stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,6490081* ,0320821 ,000 -,734637 -,563379
3 ,1004106* ,0169354 ,000 ,055209 ,145612
2
(3 saat)
1 ,6490081* ,0320821 ,000 ,563379 ,734637
3 ,7494187* ,0362777 ,000 ,656269 ,842568
3
(27 saat)
1 -,1004106* ,0169354 ,000 -,145612 -,055209
2 -,7494187* ,0362777 ,000 -,842568 -,656269
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.117 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 52,762 2 26,381 338,607 ,000
Gruplar içi 3,506 45 ,078
Toplam 56,268 47
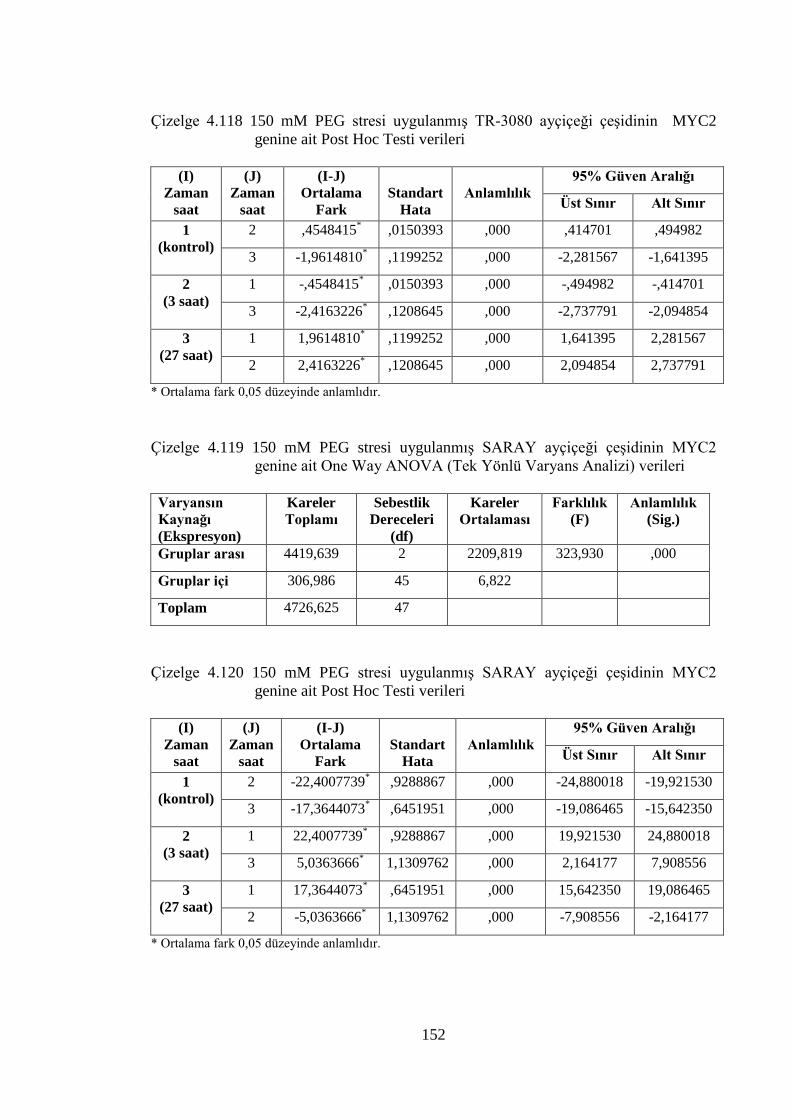
152
Çizelge 4.118 150 mM PEG stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,4548415* ,0150393 ,000 ,414701 ,494982
3 -1,9614810* ,1199252 ,000 -2,281567 -1,641395
2
(3 saat)
1 -,4548415* ,0150393 ,000 -,494982 -,414701
3 -2,4163226* ,1208645 ,000 -2,737791 -2,094854
3
(27 saat)
1 1,9614810* ,1199252 ,000 1,641395 2,281567
2 2,4163226* ,1208645 ,000 2,094854 2,737791
* Ortalama fark 0,05 düzeyinde anlamlıdır.
Çizelge 4.119 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 4419,639 2 2209,819 323,930 ,000
Gruplar içi 306,986 45 6,822
Toplam 4726,625 47
Çizelge 4.120 150 mM PEG stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -22,4007739* ,9288867 ,000 -24,880018 -19,921530
3 -17,3644073* ,6451951 ,000 -19,086465 -15,642350
2
(3 saat)
1 22,4007739* ,9288867 ,000 19,921530 24,880018
3 5,0363666* 1,1309762 ,000 2,164177 7,908556
3
(27 saat)
1 17,3644073* ,6451951 ,000 15,642350 19,086465
2 -5,0363666* 1,1309762 ,000 -7,908556 -2,164177
* Ortalama fark 0,05 düzeyinde anlamlıdır.
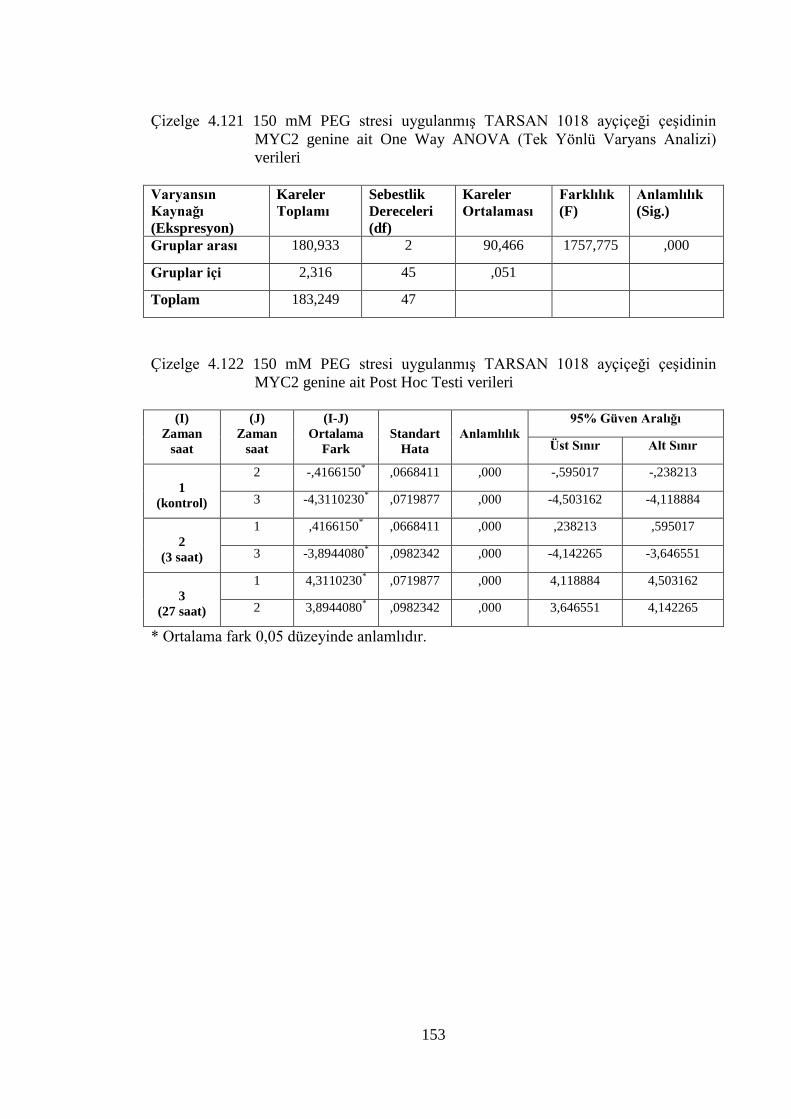
153
Çizelge 4.121 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait One Way ANOVA (Tek Yönlü Varyans Analizi)
verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 180,933 2 90,466 1757,775 ,000
Gruplar içi 2,316 45 ,051
Toplam 183,249 47
Çizelge 4.122 150 mM PEG stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin
MYC2 genine ait Post Hoc Testi verileri
(I)
Zaman
saat
(J)
Zaman
saat
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,4166150* ,0668411 ,000 -,595017 -,238213
3 -4,3110230* ,0719877 ,000 -4,503162 -4,118884
2
(3 saat)
1 ,4166150* ,0668411 ,000 ,238213 ,595017
3 -3,8944080* ,0982342 ,000 -4,142265 -3,646551
3
(27 saat)
1 4,3110230* ,0719877 ,000 4,118884 4,503162
2 3,8944080* ,0982342 ,000 3,646551 4,142265
* Ortalama fark 0,05 düzeyinde anlamlıdır.
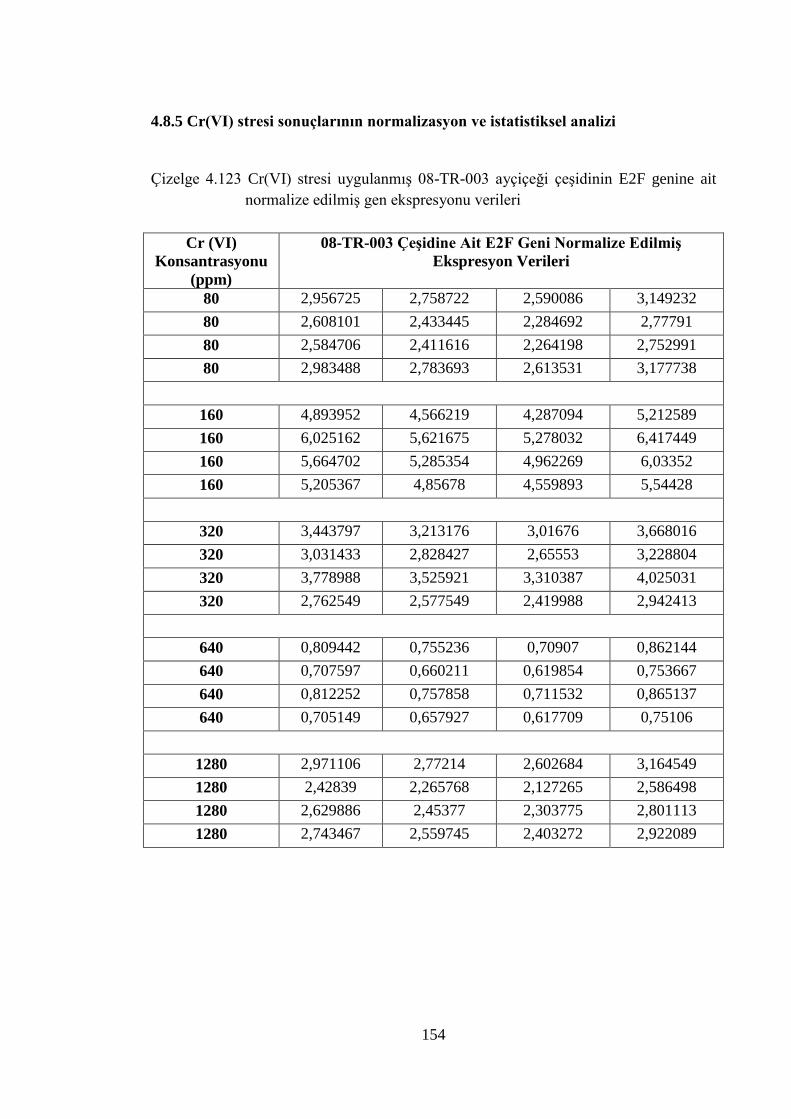
154
4.8.5 Cr(VI) stresi sonuçlarının normalizasyon ve istatistiksel analizi
Çizelge 4.123 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
08-TR-003 Çeşidine Ait E2F Geni Normalize Edilmiş
Ekspresyon Verileri
80 2,956725 2,758722 2,590086 3,149232
80 2,608101 2,433445 2,284692 2,77791
80 2,584706 2,411616 2,264198 2,752991
80 2,983488 2,783693 2,613531 3,177738
160 4,893952 4,566219 4,287094 5,212589
160 6,025162 5,621675 5,278032 6,417449
160 5,664702 5,285354 4,962269 6,03352
160 5,205367 4,85678 4,559893 5,54428
320 3,443797 3,213176 3,01676 3,668016
320 3,031433 2,828427 2,65553 3,228804
320 3,778988 3,525921 3,310387 4,025031
320 2,762549 2,577549 2,419988 2,942413
640 0,809442 0,755236 0,70907 0,862144
640 0,707597 0,660211 0,619854 0,753667
640 0,812252 0,757858 0,711532 0,865137
640 0,705149 0,657927 0,617709 0,75106
1280 2,971106 2,77214 2,602684 3,164549
1280 2,42839 2,265768 2,127265 2,586498
1280 2,629886 2,45377 2,303775 2,801113
1280 2,743467 2,559745 2,403272 2,922089
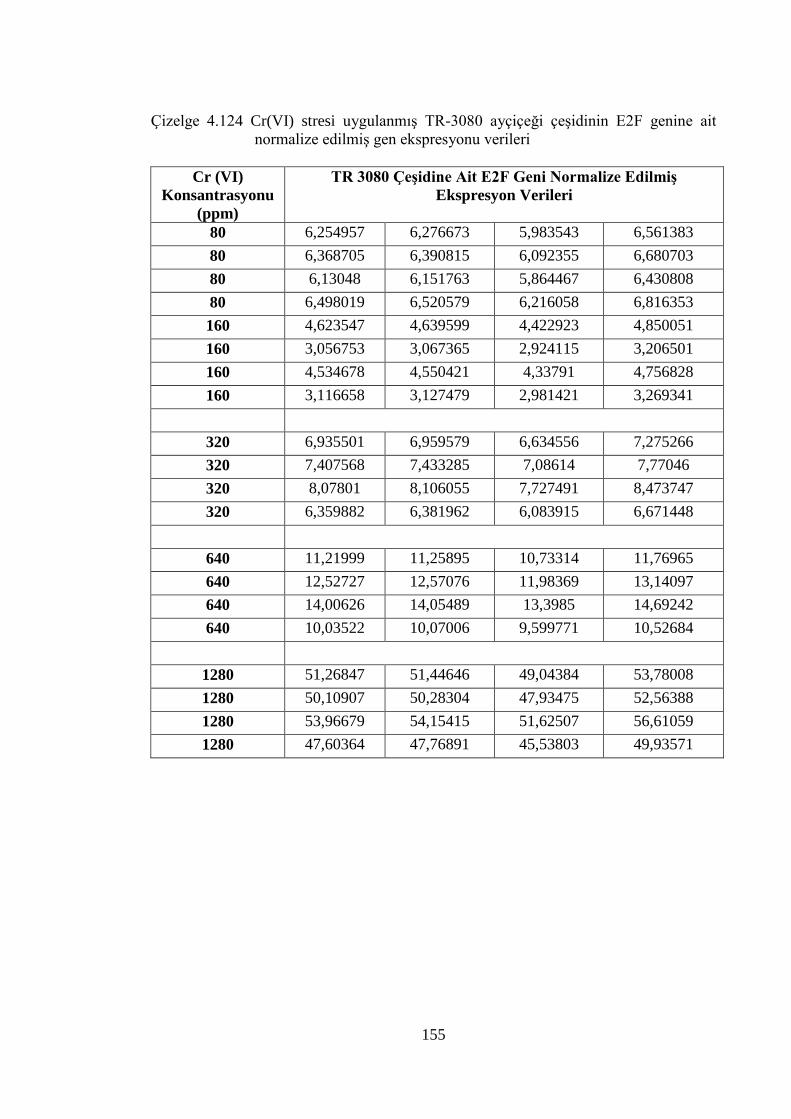
155
Çizelge 4.124 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TR 3080 Çeşidine Ait E2F Geni Normalize Edilmiş
Ekspresyon Verileri
80 6,254957 6,276673 5,983543 6,561383
80 6,368705 6,390815 6,092355 6,680703
80 6,13048 6,151763 5,864467 6,430808
80 6,498019 6,520579 6,216058 6,816353
160 4,623547 4,639599 4,422923 4,850051
160 3,056753 3,067365 2,924115 3,206501
160 4,534678 4,550421 4,33791 4,756828
160 3,116658 3,127479 2,981421 3,269341
320 6,935501 6,959579 6,634556 7,275266
320 7,407568 7,433285 7,08614 7,77046
320 8,07801 8,106055 7,727491 8,473747
320 6,359882 6,381962 6,083915 6,671448
640 11,21999 11,25895 10,73314 11,76965
640 12,52727 12,57076 11,98369 13,14097
640 14,00626 14,05489 13,3985 14,69242
640 10,03522 10,07006 9,599771 10,52684
1280 51,26847 51,44646 49,04384 53,78008
1280 50,10907 50,28304 47,93475 52,56388
1280 53,96679 54,15415 51,62507 56,61059
1280 47,60364 47,76891 45,53803 49,93571
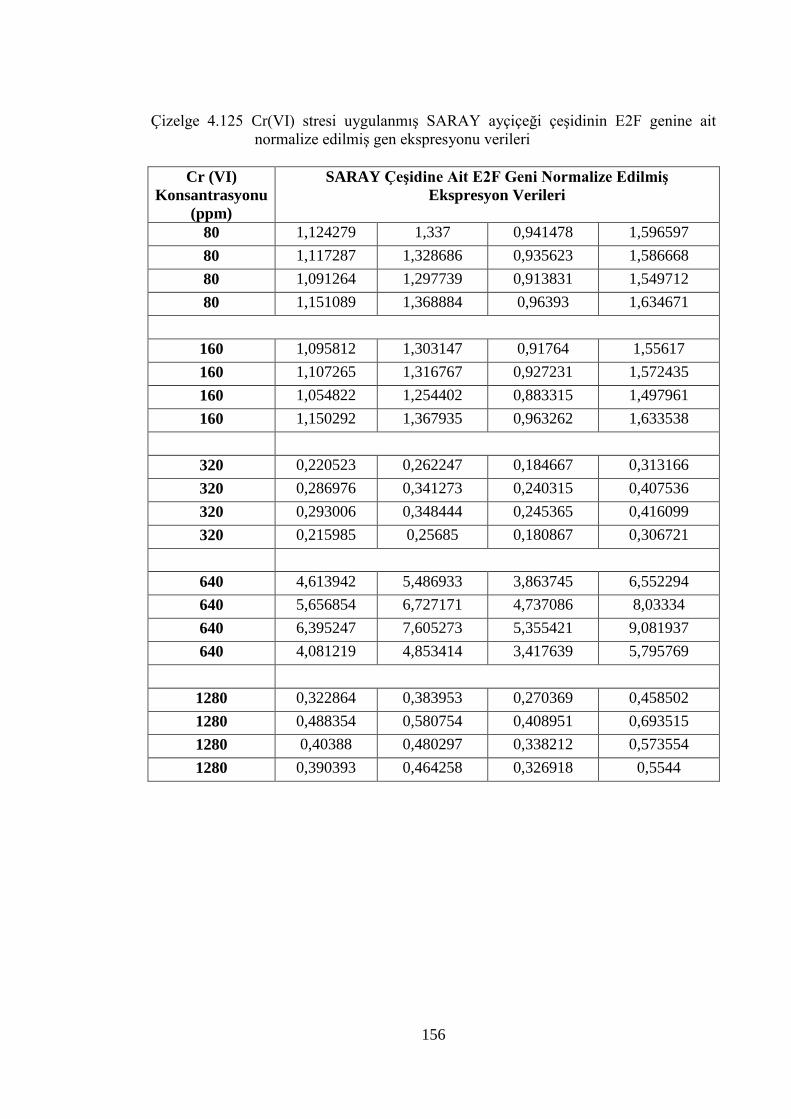
156
Çizelge 4.125 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
SARAY Çeşidine Ait E2F Geni Normalize Edilmiş
Ekspresyon Verileri
80 1,124279 1,337 0,941478 1,596597
80 1,117287 1,328686 0,935623 1,586668
80 1,091264 1,297739 0,913831 1,549712
80 1,151089 1,368884 0,96393 1,634671
160 1,095812 1,303147 0,91764 1,55617
160 1,107265 1,316767 0,927231 1,572435
160 1,054822 1,254402 0,883315 1,497961
160 1,150292 1,367935 0,963262 1,633538
320 0,220523 0,262247 0,184667 0,313166
320 0,286976 0,341273 0,240315 0,407536
320 0,293006 0,348444 0,245365 0,416099
320 0,215985 0,25685 0,180867 0,306721
640 4,613942 5,486933 3,863745 6,552294
640 5,656854 6,727171 4,737086 8,03334
640 6,395247 7,605273 5,355421 9,081937
640 4,081219 4,853414 3,417639 5,795769
1280 0,322864 0,383953 0,270369 0,458502
1280 0,488354 0,580754 0,408951 0,693515
1280 0,40388 0,480297 0,338212 0,573554
1280 0,390393 0,464258 0,326918 0,5544
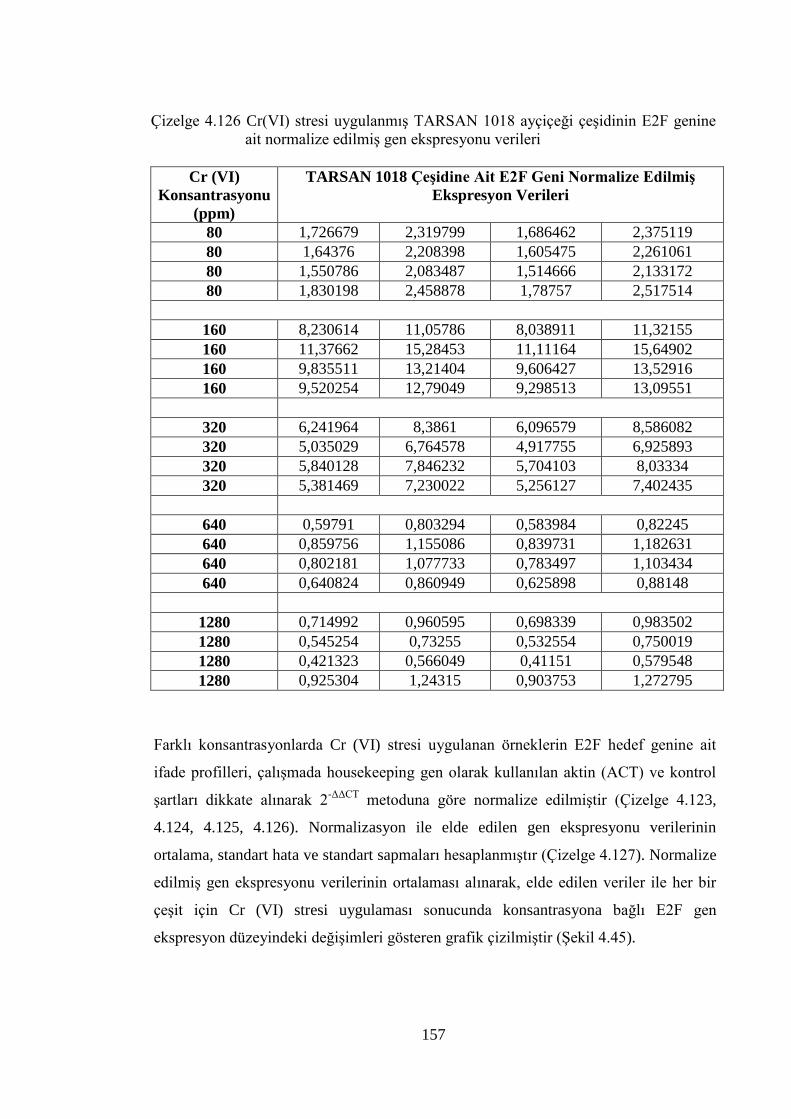
157
Çizelge 4.126 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F genine
ait normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TARSAN 1018 Çeşidine Ait E2F Geni Normalize Edilmiş
Ekspresyon Verileri
80 1,726679 2,319799 1,686462 2,375119
80 1,64376 2,208398 1,605475 2,261061
80 1,550786 2,083487 1,514666 2,133172
80 1,830198 2,458878 1,78757 2,517514
160 8,230614 11,05786 8,038911 11,32155
160 11,37662 15,28453 11,11164 15,64902
160 9,835511 13,21404 9,606427 13,52916
160 9,520254 12,79049 9,298513 13,09551
320 6,241964 8,3861 6,096579 8,586082
320 5,035029 6,764578 4,917755 6,925893
320 5,840128 7,846232 5,704103 8,03334
320 5,381469 7,230022 5,256127 7,402435
640 0,59791 0,803294 0,583984 0,82245
640 0,859756 1,155086 0,839731 1,182631
640 0,802181 1,077733 0,783497 1,103434
640 0,640824 0,860949 0,625898 0,88148
1280 0,714992 0,960595 0,698339 0,983502
1280 0,545254 0,73255 0,532554 0,750019
1280 0,421323 0,566049 0,41151 0,579548
1280 0,925304 1,24315 0,903753 1,272795
Farklı konsantrasyonlarda Cr (VI) stresi uygulanan örneklerin E2F hedef genine ait
ifade profilleri, çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol
şartları dikkate alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.123,
4.124, 4.125, 4.126). Normalizasyon ile elde edilen gen ekspresyonu verilerinin
ortalama, standart hata ve standart sapmaları hesaplanmıştır (Çizelge 4.127). Normalize
edilmiş gen ekspresyonu verilerinin ortalaması alınarak, elde edilen veriler ile her bir
çeşit için Cr (VI) stresi uygulaması sonucunda konsantrasyona bağlı E2F gen
ekspresyon düzeyindeki değişimleri gösteren grafik çizilmiştir (Şekil 4.45).
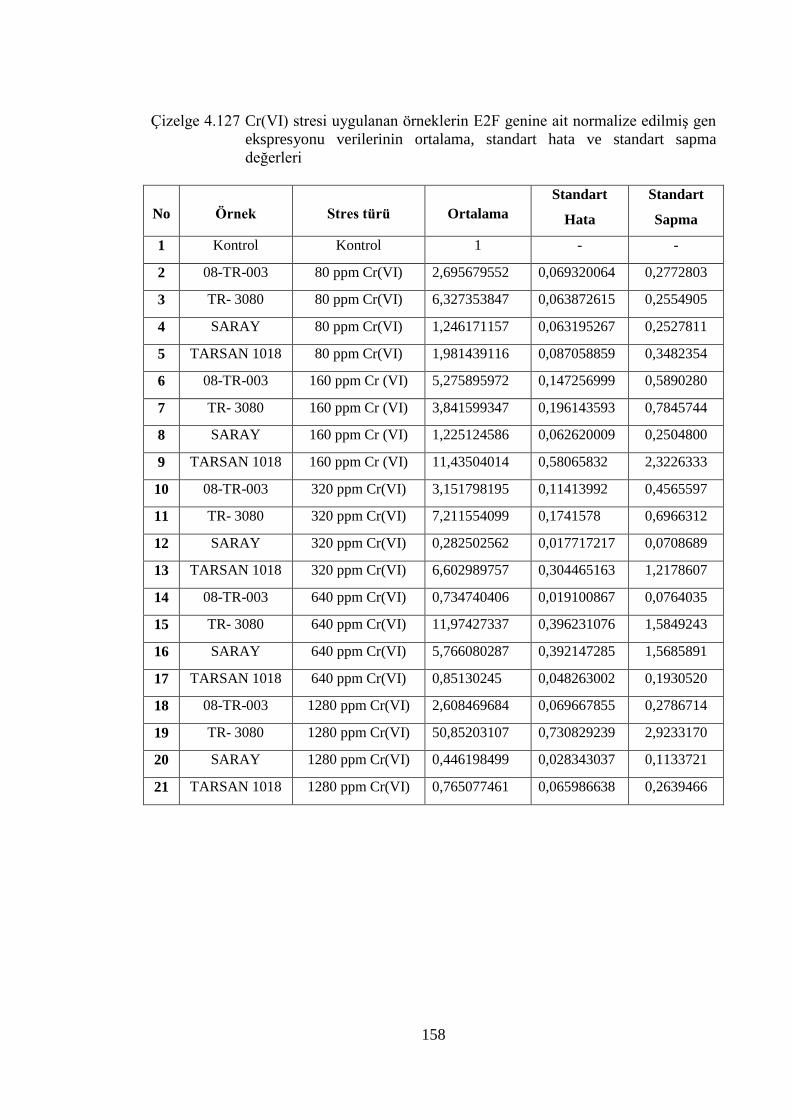
158
Çizelge 4.127 Cr(VI) stresi uygulanan örneklerin E2F genine ait normalize edilmiş gen
ekspresyonu verilerinin ortalama, standart hata ve standart sapma
değerleri
No
Örnek
Stres türü
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 1 - -
2 08-TR-003 80 ppm Cr(VI) 2,695679552 0,069320064 0,2772803
3 TR- 3080 80 ppm Cr(VI) 6,327353847 0,063872615 0,2554905
4 SARAY 80 ppm Cr(VI) 1,246171157 0,063195267 0,2527811
5 TARSAN 1018 80 ppm Cr(VI) 1,981439116 0,087058859 0,3482354
6 08-TR-003 160 ppm Cr (VI) 5,275895972 0,147256999 0,5890280
7 TR- 3080 160 ppm Cr (VI) 3,841599347 0,196143593 0,7845744
8 SARAY 160 ppm Cr (VI) 1,225124586 0,062620009 0,2504800
9 TARSAN 1018 160 ppm Cr (VI) 11,43504014 0,58065832 2,3226333
10 08-TR-003 320 ppm Cr(VI) 3,151798195 0,11413992 0,4565597
11 TR- 3080 320 ppm Cr(VI) 7,211554099 0,1741578 0,6966312
12 SARAY 320 ppm Cr(VI) 0,282502562 0,017717217 0,0708689
13 TARSAN 1018 320 ppm Cr(VI) 6,602989757 0,304465163 1,2178607
14 08-TR-003 640 ppm Cr(VI) 0,734740406 0,019100867 0,0764035
15 TR- 3080 640 ppm Cr(VI) 11,97427337 0,396231076 1,5849243
16 SARAY 640 ppm Cr(VI) 5,766080287 0,392147285 1,5685891
17 TARSAN 1018 640 ppm Cr(VI) 0,85130245 0,048263002 0,1930520
18 08-TR-003 1280 ppm Cr(VI) 2,608469684 0,069667855 0,2786714
19 TR- 3080 1280 ppm Cr(VI) 50,85203107 0,730829239 2,9233170
20 SARAY 1280 ppm Cr(VI) 0,446198499 0,028343037 0,1133721
21 TARSAN 1018 1280 ppm Cr(VI) 0,765077461 0,065986638 0,2639466
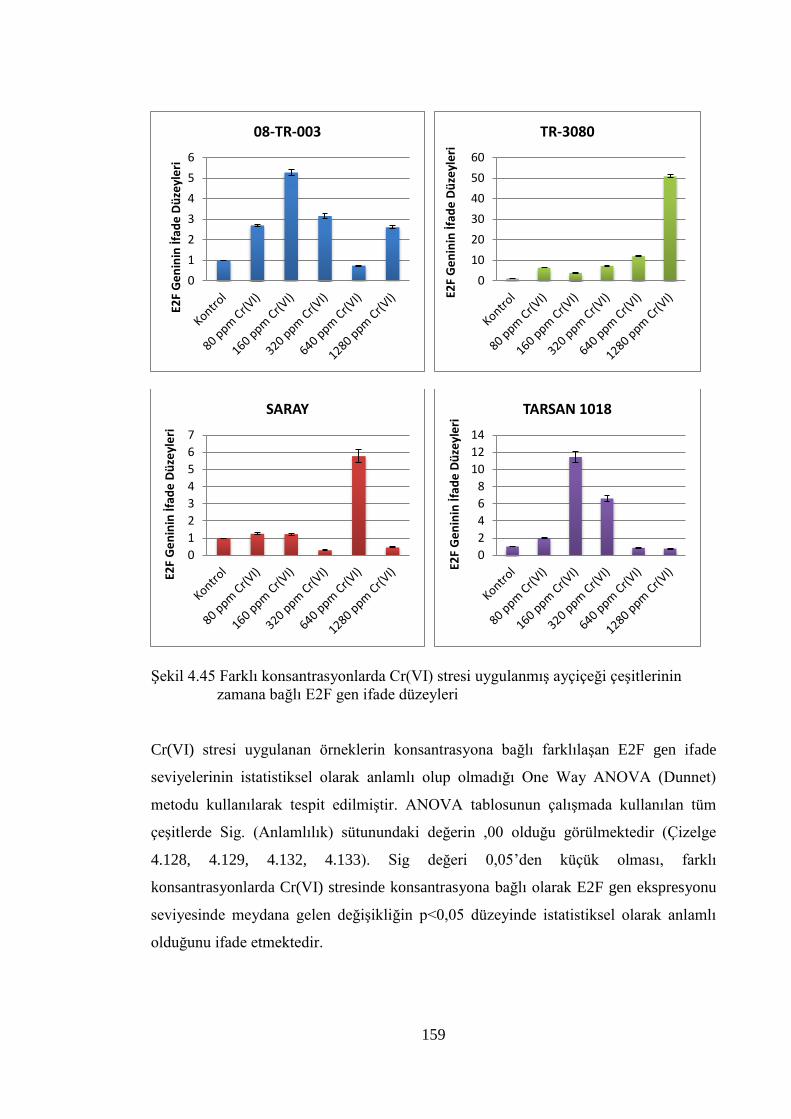
159
Şekil 4.45 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği çeşitlerinin
zamana bağlı E2F gen ifade düzeyleri
Cr(VI) stresi uygulanan örneklerin konsantrasyona bağlı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.128, 4.129, 4.132, 4.133). Sig değeri 0,05’den küçük olması, farklı
konsantrasyonlarda Cr(VI) stresinde konsantrasyona bağlı olarak E2F gen ekspresyonu
seviyesinde meydana gelen değişikliğin p<0,05 düzeyinde istatistiksel olarak anlamlı
olduğunu ifade etmektedir.
0
1
2
3
4
5
6
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
08-TR-003
0
10
20
30
40
50
60
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
TR-3080
0
1
2
3
4
5
6
7
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
SARAY
0
2
4
6
8
10
12
14
E2F
Ge
nin
in İf
ade
Dü
zeyl
eri
TARSAN 1018
160
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan grupların tek yönlü varyans analizi
(One Way ANOVA Dunnet Test, Post Hoc testi) ile konsantrasyona karşı E2F geni
ekspresyon seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri
karşılaştırılmıştır (Çizelge 4.130, 4.131, 4.134, 4.135).
Çizelge 4.128 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 216,166 5 43,233 362,400 ,000
Gruplar içi 10,737 90 ,119
Toplam 226,902 95
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
Cr(VI) (80 ppm, 160 ppm, 320 ppm, 640 ppm, 1280 ppm)
verilerinin kendi içinde değerlendirilmesidir.
Çizelge 4.129 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine ait One
Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 27808,337 5 5561,667 2729,900 ,000
Gruplar içi 183,358 90 2,037
Toplam 27991,695 95
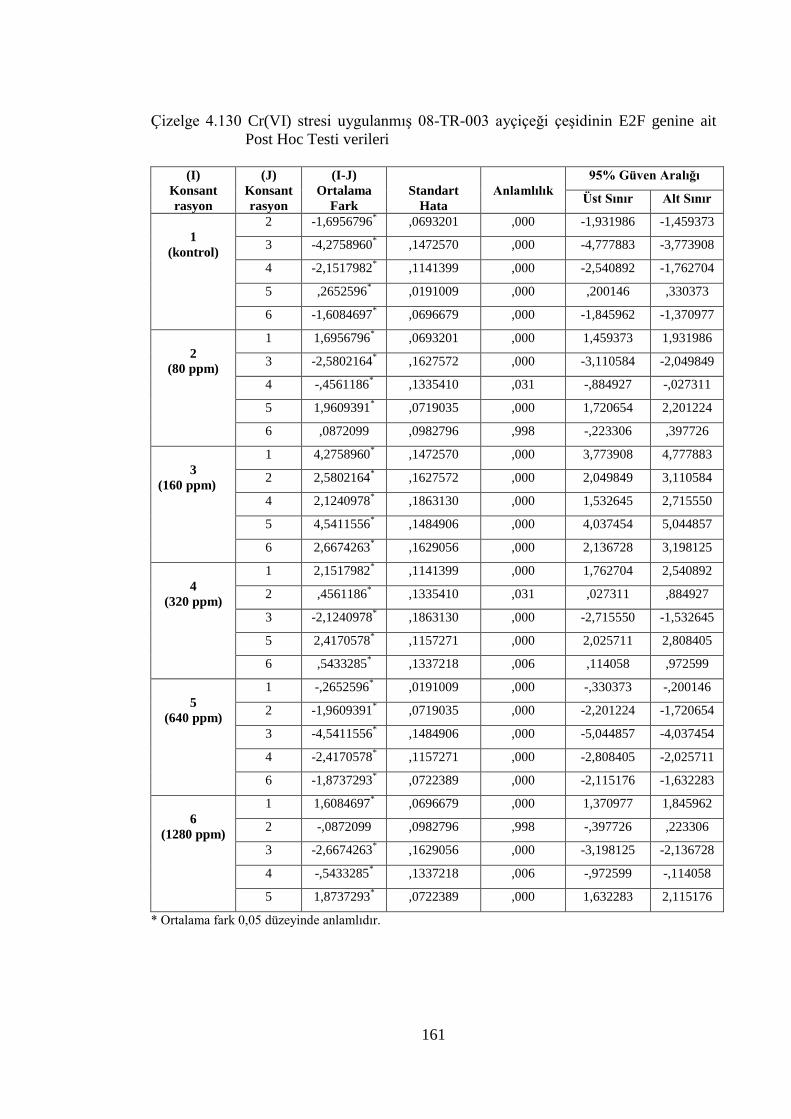
161
Çizelge 4.130 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin E2F genine ait
Post Hoc Testi verileri
(I)
Konsant
rasyon
(J)
Konsant
rasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -1,6956796* ,0693201 ,000 -1,931986 -1,459373
3 -4,2758960* ,1472570 ,000 -4,777883 -3,773908
4 -2,1517982* ,1141399 ,000 -2,540892 -1,762704
5 ,2652596* ,0191009 ,000 ,200146 ,330373
6 -1,6084697* ,0696679 ,000 -1,845962 -1,370977
2
(80 ppm)
1 1,6956796* ,0693201 ,000 1,459373 1,931986
3 -2,5802164* ,1627572 ,000 -3,110584 -2,049849
4 -,4561186* ,1335410 ,031 -,884927 -,027311
5 1,9609391* ,0719035 ,000 1,720654 2,201224
6 ,0872099 ,0982796 ,998 -,223306 ,397726
3
(160 ppm)
1 4,2758960* ,1472570 ,000 3,773908 4,777883
2 2,5802164* ,1627572 ,000 2,049849 3,110584
4 2,1240978* ,1863130 ,000 1,532645 2,715550
5 4,5411556* ,1484906 ,000 4,037454 5,044857
6 2,6674263* ,1629056 ,000 2,136728 3,198125
4
(320 ppm)
1 2,1517982* ,1141399 ,000 1,762704 2,540892
2 ,4561186* ,1335410 ,031 ,027311 ,884927
3 -2,1240978* ,1863130 ,000 -2,715550 -1,532645
5 2,4170578* ,1157271 ,000 2,025711 2,808405
6 ,5433285* ,1337218 ,006 ,114058 ,972599
5
(640 ppm)
1 -,2652596* ,0191009 ,000 -,330373 -,200146
2 -1,9609391* ,0719035 ,000 -2,201224 -1,720654
3 -4,5411556* ,1484906 ,000 -5,044857 -4,037454
4 -2,4170578* ,1157271 ,000 -2,808405 -2,025711
6 -1,8737293* ,0722389 ,000 -2,115176 -1,632283
6
(1280 ppm)
1 1,6084697* ,0696679 ,000 1,370977 1,845962
2 -,0872099 ,0982796 ,998 -,397726 ,223306
3 -2,6674263* ,1629056 ,000 -3,198125 -2,136728
4 -,5433285* ,1337218 ,006 -,972599 -,114058
5 1,8737293* ,0722389 ,000 1,632283 2,115176
* Ortalama fark 0,05 düzeyinde anlamlıdır.
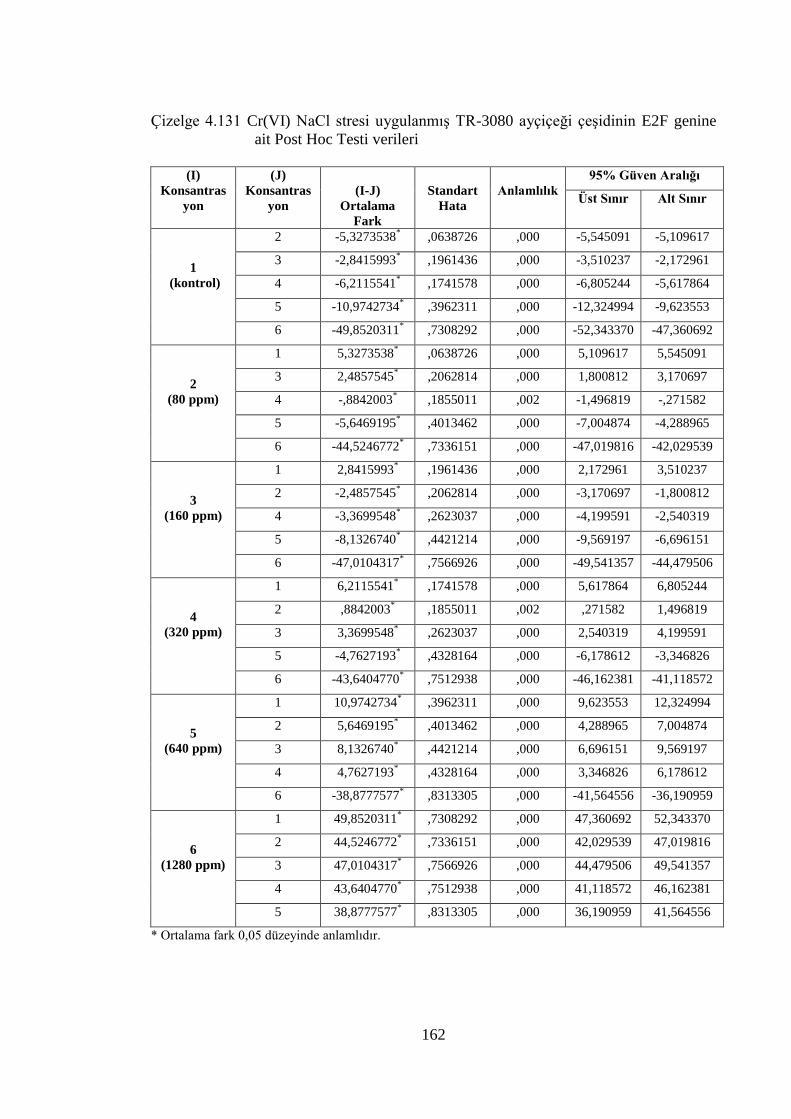
162
Çizelge 4.131 Cr(VI) NaCl stresi uygulanmış TR-3080 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsantras
yon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -5,3273538* ,0638726 ,000 -5,545091 -5,109617
3 -2,8415993* ,1961436 ,000 -3,510237 -2,172961
4 -6,2115541* ,1741578 ,000 -6,805244 -5,617864
5 -10,9742734* ,3962311 ,000 -12,324994 -9,623553
6 -49,8520311* ,7308292 ,000 -52,343370 -47,360692
2
(80 ppm)
1 5,3273538* ,0638726 ,000 5,109617 5,545091
3 2,4857545* ,2062814 ,000 1,800812 3,170697
4 -,8842003* ,1855011 ,002 -1,496819 -,271582
5 -5,6469195* ,4013462 ,000 -7,004874 -4,288965
6 -44,5246772* ,7336151 ,000 -47,019816 -42,029539
3
(160 ppm)
1 2,8415993* ,1961436 ,000 2,172961 3,510237
2 -2,4857545* ,2062814 ,000 -3,170697 -1,800812
4 -3,3699548* ,2623037 ,000 -4,199591 -2,540319
5 -8,1326740* ,4421214 ,000 -9,569197 -6,696151
6 -47,0104317* ,7566926 ,000 -49,541357 -44,479506
4
(320 ppm)
1 6,2115541* ,1741578 ,000 5,617864 6,805244
2 ,8842003* ,1855011 ,002 ,271582 1,496819
3 3,3699548* ,2623037 ,000 2,540319 4,199591
5 -4,7627193* ,4328164 ,000 -6,178612 -3,346826
6 -43,6404770* ,7512938 ,000 -46,162381 -41,118572
5
(640 ppm)
1 10,9742734* ,3962311 ,000 9,623553 12,324994
2 5,6469195* ,4013462 ,000 4,288965 7,004874
3 8,1326740* ,4421214 ,000 6,696151 9,569197
4 4,7627193* ,4328164 ,000 3,346826 6,178612
6 -38,8777577* ,8313305 ,000 -41,564556 -36,190959
6
(1280 ppm)
1 49,8520311* ,7308292 ,000 47,360692 52,343370
2 44,5246772* ,7336151 ,000 42,029539 47,019816
3 47,0104317* ,7566926 ,000 44,479506 49,541357
4 43,6404770* ,7512938 ,000 41,118572 46,162381
5 38,8777577* ,8313305 ,000 36,190959 41,564556
* Ortalama fark 0,05 düzeyinde anlamlıdır.
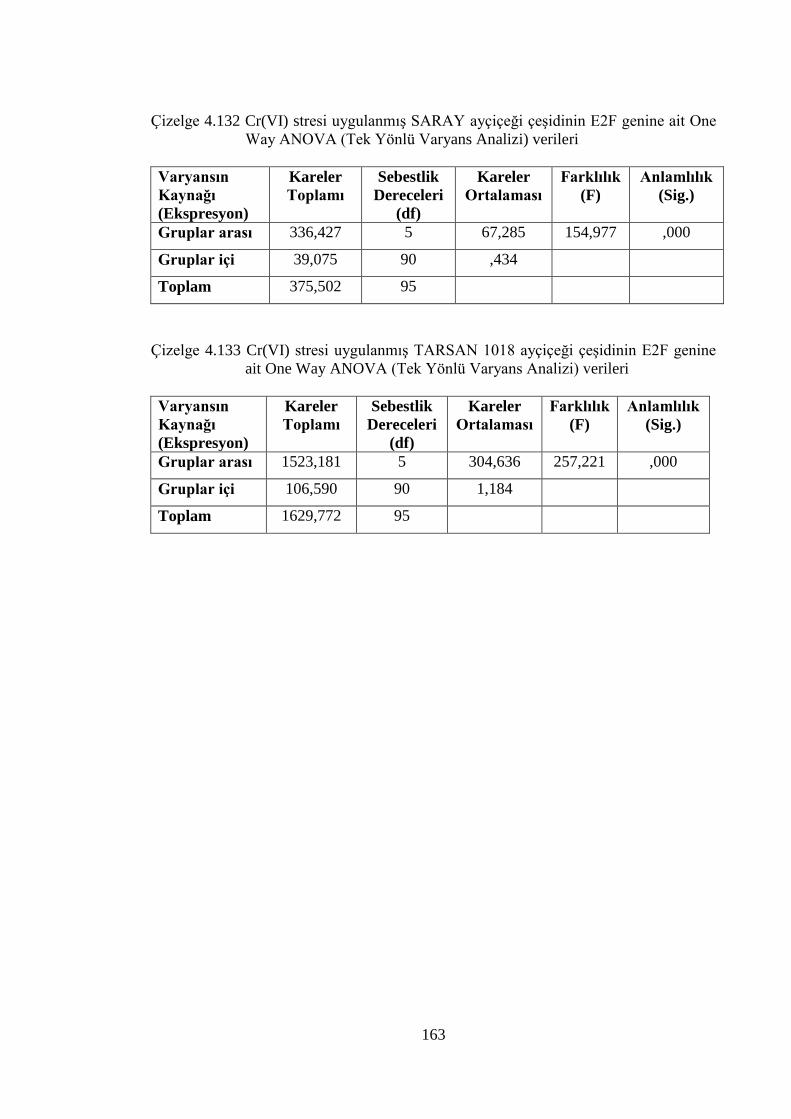
163
Çizelge 4.132 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait One
Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 336,427 5 67,285 154,977 ,000
Gruplar içi 39,075 90 ,434
Toplam 375,502 95
Çizelge 4.133 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 1523,181 5 304,636 257,221 ,000
Gruplar içi 106,590 90 1,184
Toplam 1629,772 95
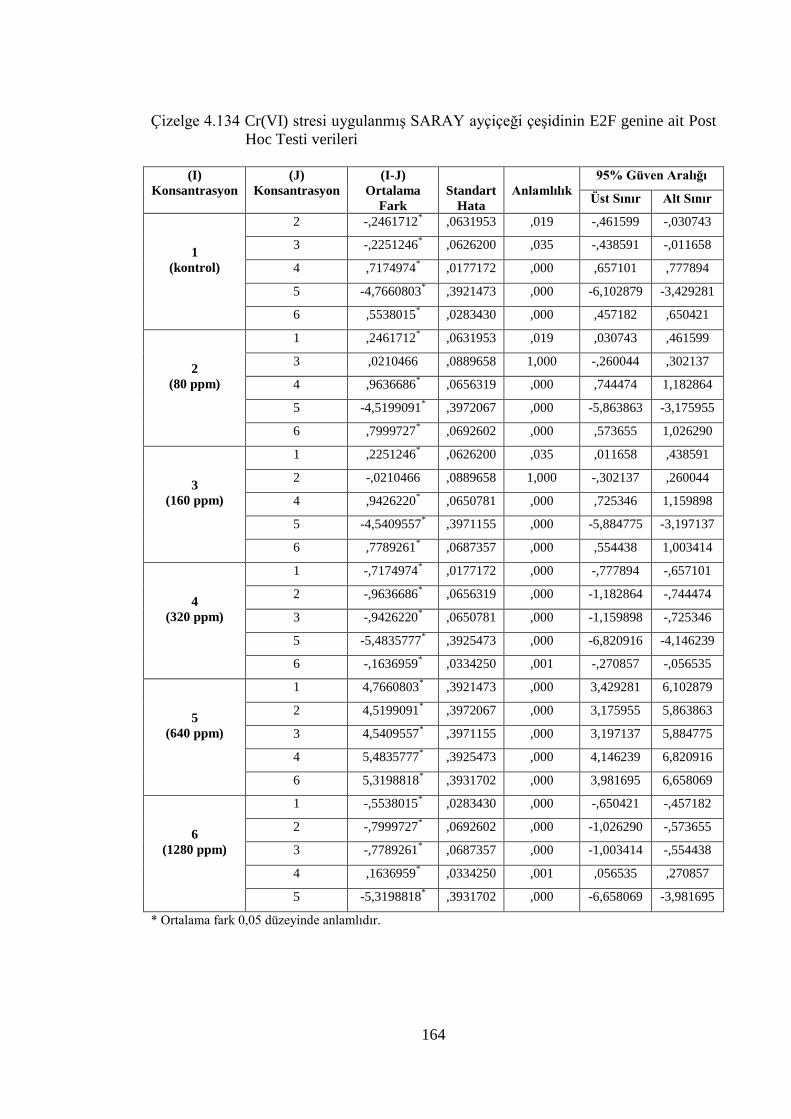
164
Çizelge 4.134 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin E2F genine ait Post
Hoc Testi verileri
(I)
Konsantrasyon
(J)
Konsantrasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,2461712* ,0631953 ,019 -,461599 -,030743
3 -,2251246* ,0626200 ,035 -,438591 -,011658
4 ,7174974* ,0177172 ,000 ,657101 ,777894
5 -4,7660803* ,3921473 ,000 -6,102879 -3,429281
6 ,5538015* ,0283430 ,000 ,457182 ,650421
2
(80 ppm)
1 ,2461712* ,0631953 ,019 ,030743 ,461599
3 ,0210466 ,0889658 1,000 -,260044 ,302137
4 ,9636686* ,0656319 ,000 ,744474 1,182864
5 -4,5199091* ,3972067 ,000 -5,863863 -3,175955
6 ,7999727* ,0692602 ,000 ,573655 1,026290
3
(160 ppm)
1 ,2251246* ,0626200 ,035 ,011658 ,438591
2 -,0210466 ,0889658 1,000 -,302137 ,260044
4 ,9426220* ,0650781 ,000 ,725346 1,159898
5 -4,5409557* ,3971155 ,000 -5,884775 -3,197137
6 ,7789261* ,0687357 ,000 ,554438 1,003414
4
(320 ppm)
1 -,7174974* ,0177172 ,000 -,777894 -,657101
2 -,9636686* ,0656319 ,000 -1,182864 -,744474
3 -,9426220* ,0650781 ,000 -1,159898 -,725346
5 -5,4835777* ,3925473 ,000 -6,820916 -4,146239
6 -,1636959* ,0334250 ,001 -,270857 -,056535
5
(640 ppm)
1 4,7660803* ,3921473 ,000 3,429281 6,102879
2 4,5199091* ,3972067 ,000 3,175955 5,863863
3 4,5409557* ,3971155 ,000 3,197137 5,884775
4 5,4835777* ,3925473 ,000 4,146239 6,820916
6 5,3198818* ,3931702 ,000 3,981695 6,658069
6
(1280 ppm)
1 -,5538015* ,0283430 ,000 -,650421 -,457182
2 -,7999727* ,0692602 ,000 -1,026290 -,573655
3 -,7789261* ,0687357 ,000 -1,003414 -,554438
4 ,1636959* ,0334250 ,001 ,056535 ,270857
5 -5,3198818* ,3931702 ,000 -6,658069 -3,981695
* Ortalama fark 0,05 düzeyinde anlamlıdır.
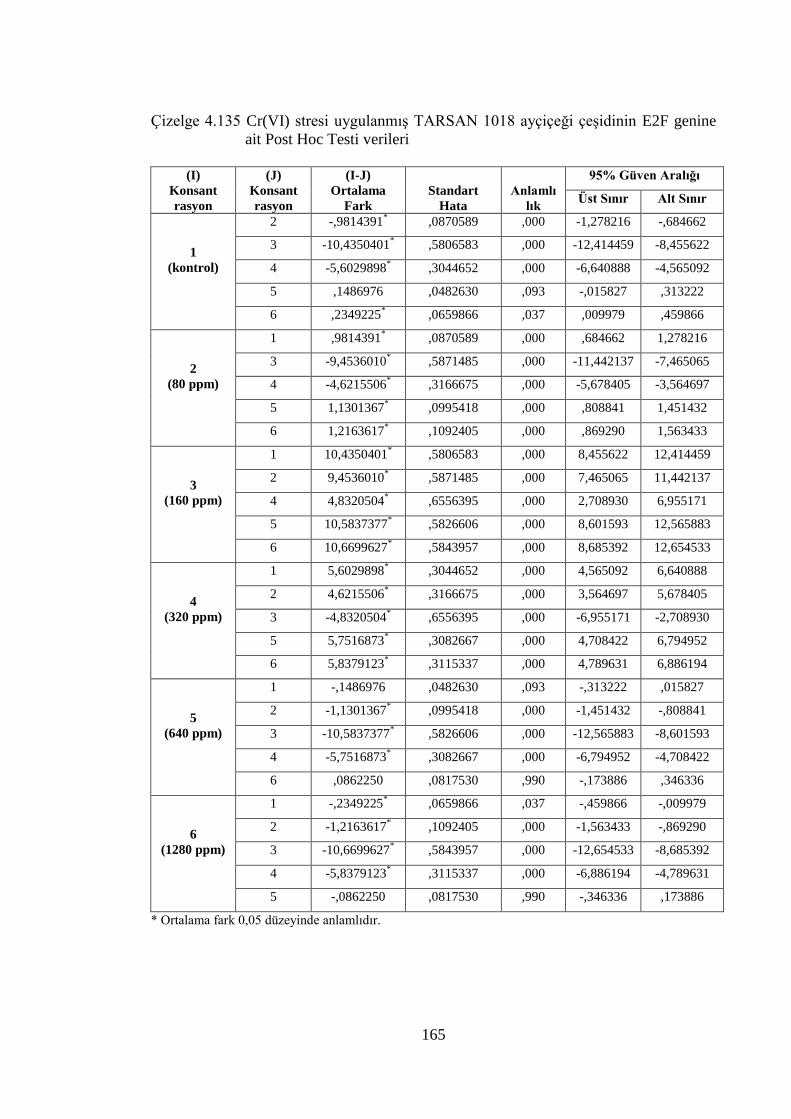
165
Çizelge 4.135 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin E2F genine
ait Post Hoc Testi verileri
(I)
Konsant
rasyon
(J)
Konsant
rasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlı
lık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -,9814391* ,0870589 ,000 -1,278216 -,684662
3 -10,4350401* ,5806583 ,000 -12,414459 -8,455622
4 -5,6029898* ,3044652 ,000 -6,640888 -4,565092
5 ,1486976 ,0482630 ,093 -,015827 ,313222
6 ,2349225* ,0659866 ,037 ,009979 ,459866
2
(80 ppm)
1 ,9814391* ,0870589 ,000 ,684662 1,278216
3 -9,4536010* ,5871485 ,000 -11,442137 -7,465065
4 -4,6215506* ,3166675 ,000 -5,678405 -3,564697
5 1,1301367* ,0995418 ,000 ,808841 1,451432
6 1,2163617* ,1092405 ,000 ,869290 1,563433
3
(160 ppm)
1 10,4350401* ,5806583 ,000 8,455622 12,414459
2 9,4536010* ,5871485 ,000 7,465065 11,442137
4 4,8320504* ,6556395 ,000 2,708930 6,955171
5 10,5837377* ,5826606 ,000 8,601593 12,565883
6 10,6699627* ,5843957 ,000 8,685392 12,654533
4
(320 ppm)
1 5,6029898* ,3044652 ,000 4,565092 6,640888
2 4,6215506* ,3166675 ,000 3,564697 5,678405
3 -4,8320504* ,6556395 ,000 -6,955171 -2,708930
5 5,7516873* ,3082667 ,000 4,708422 6,794952
6 5,8379123* ,3115337 ,000 4,789631 6,886194
5
(640 ppm)
1 -,1486976 ,0482630 ,093 -,313222 ,015827
2 -1,1301367* ,0995418 ,000 -1,451432 -,808841
3 -10,5837377* ,5826606 ,000 -12,565883 -8,601593
4 -5,7516873* ,3082667 ,000 -6,794952 -4,708422
6 ,0862250 ,0817530 ,990 -,173886 ,346336
6
(1280 ppm)
1 -,2349225* ,0659866 ,037 -,459866 -,009979
2 -1,2163617* ,1092405 ,000 -1,563433 -,869290
3 -10,6699627* ,5843957 ,000 -12,654533 -8,685392
4 -5,8379123* ,3115337 ,000 -6,886194 -4,789631
5 -,0862250 ,0817530 ,990 -,346336 ,173886
* Ortalama fark 0,05 düzeyinde anlamlıdır.
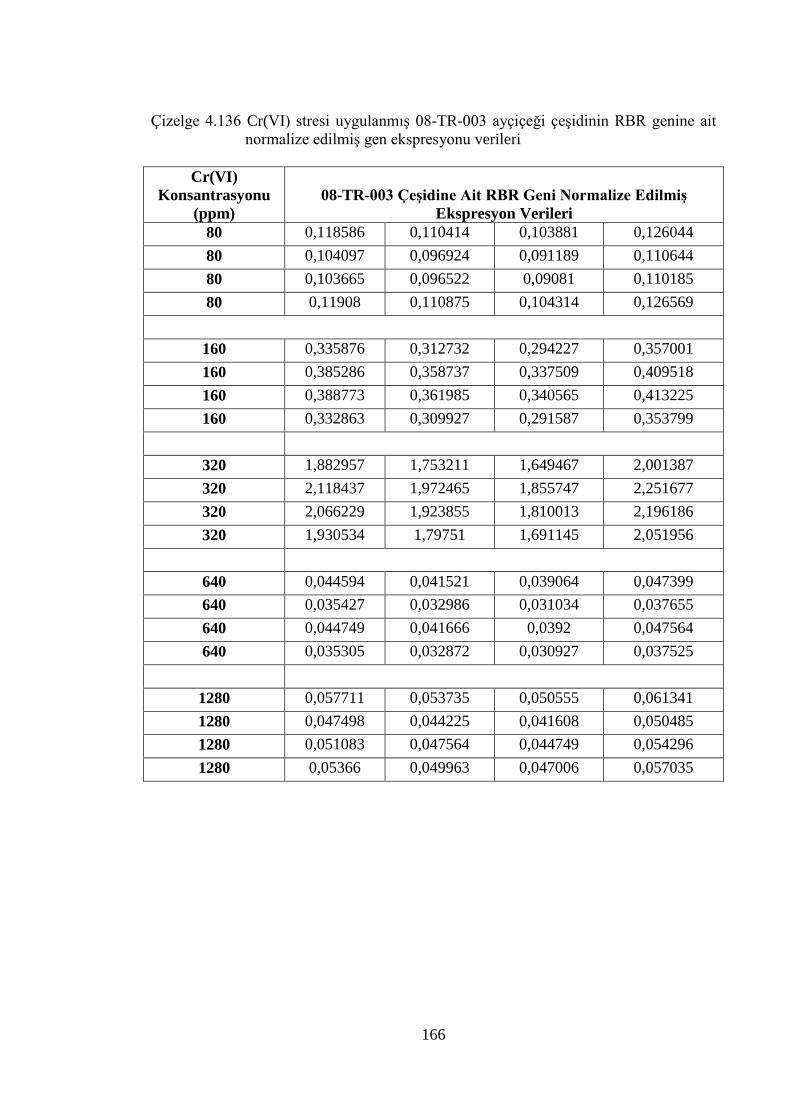
166
Çizelge 4.136 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine ait
normalize edilmiş gen ekspresyonu verileri
Cr(VI)
Konsantrasyonu
(ppm)
08-TR-003 Çeşidine Ait RBR Geni Normalize Edilmiş
Ekspresyon Verileri
80 0,118586 0,110414 0,103881 0,126044
80 0,104097 0,096924 0,091189 0,110644
80 0,103665 0,096522 0,09081 0,110185
80 0,11908 0,110875 0,104314 0,126569
160 0,335876 0,312732 0,294227 0,357001
160 0,385286 0,358737 0,337509 0,409518
160 0,388773 0,361985 0,340565 0,413225
160 0,332863 0,309927 0,291587 0,353799
320 1,882957 1,753211 1,649467 2,001387
320 2,118437 1,972465 1,855747 2,251677
320 2,066229 1,923855 1,810013 2,196186
320 1,930534 1,79751 1,691145 2,051956
640 0,044594 0,041521 0,039064 0,047399
640 0,035427 0,032986 0,031034 0,037655
640 0,044749 0,041666 0,0392 0,047564
640 0,035305 0,032872 0,030927 0,037525
1280 0,057711 0,053735 0,050555 0,061341
1280 0,047498 0,044225 0,041608 0,050485
1280 0,051083 0,047564 0,044749 0,054296
1280 0,05366 0,049963 0,047006 0,057035
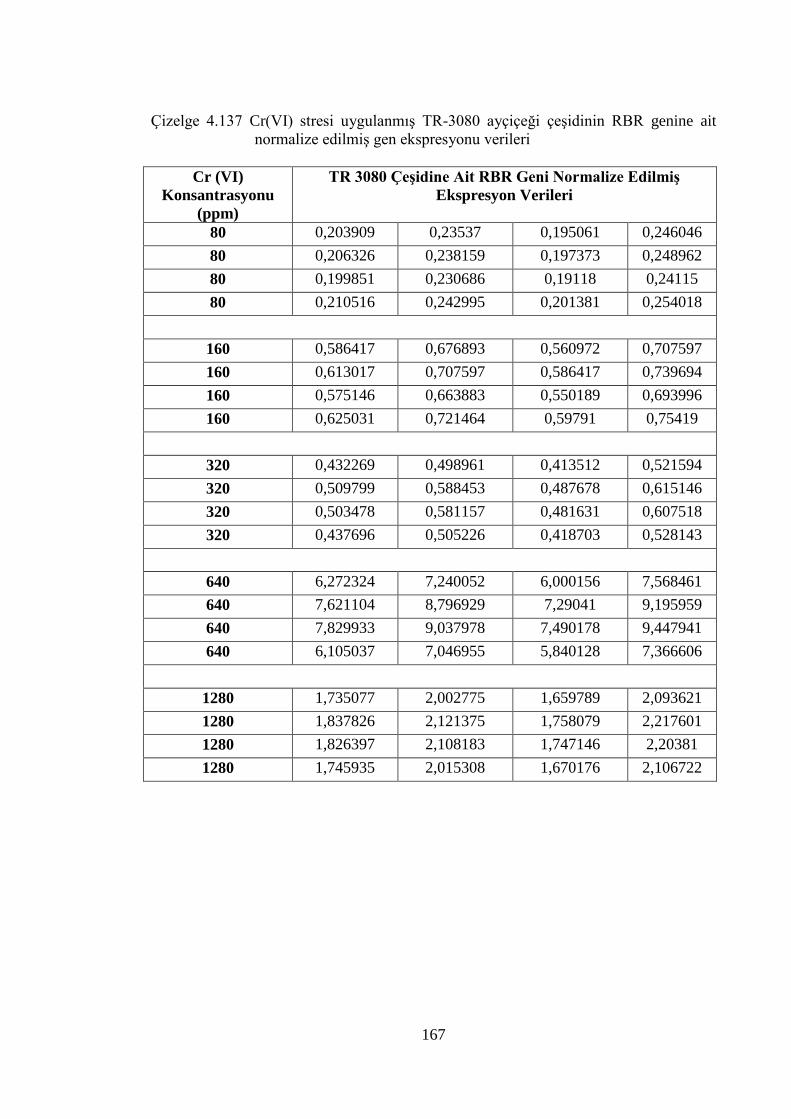
167
Çizelge 4.137 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TR 3080 Çeşidine Ait RBR Geni Normalize Edilmiş
Ekspresyon Verileri
80 0,203909 0,23537 0,195061 0,246046
80 0,206326 0,238159 0,197373 0,248962
80 0,199851 0,230686 0,19118 0,24115
80 0,210516 0,242995 0,201381 0,254018
160 0,586417 0,676893 0,560972 0,707597
160 0,613017 0,707597 0,586417 0,739694
160 0,575146 0,663883 0,550189 0,693996
160 0,625031 0,721464 0,59791 0,75419
320 0,432269 0,498961 0,413512 0,521594
320 0,509799 0,588453 0,487678 0,615146
320 0,503478 0,581157 0,481631 0,607518
320 0,437696 0,505226 0,418703 0,528143
640 6,272324 7,240052 6,000156 7,568461
640 7,621104 8,796929 7,29041 9,195959
640 7,829933 9,037978 7,490178 9,447941
640 6,105037 7,046955 5,840128 7,366606
1280 1,735077 2,002775 1,659789 2,093621
1280 1,837826 2,121375 1,758079 2,217601
1280 1,826397 2,108183 1,747146 2,20381
1280 1,745935 2,015308 1,670176 2,106722
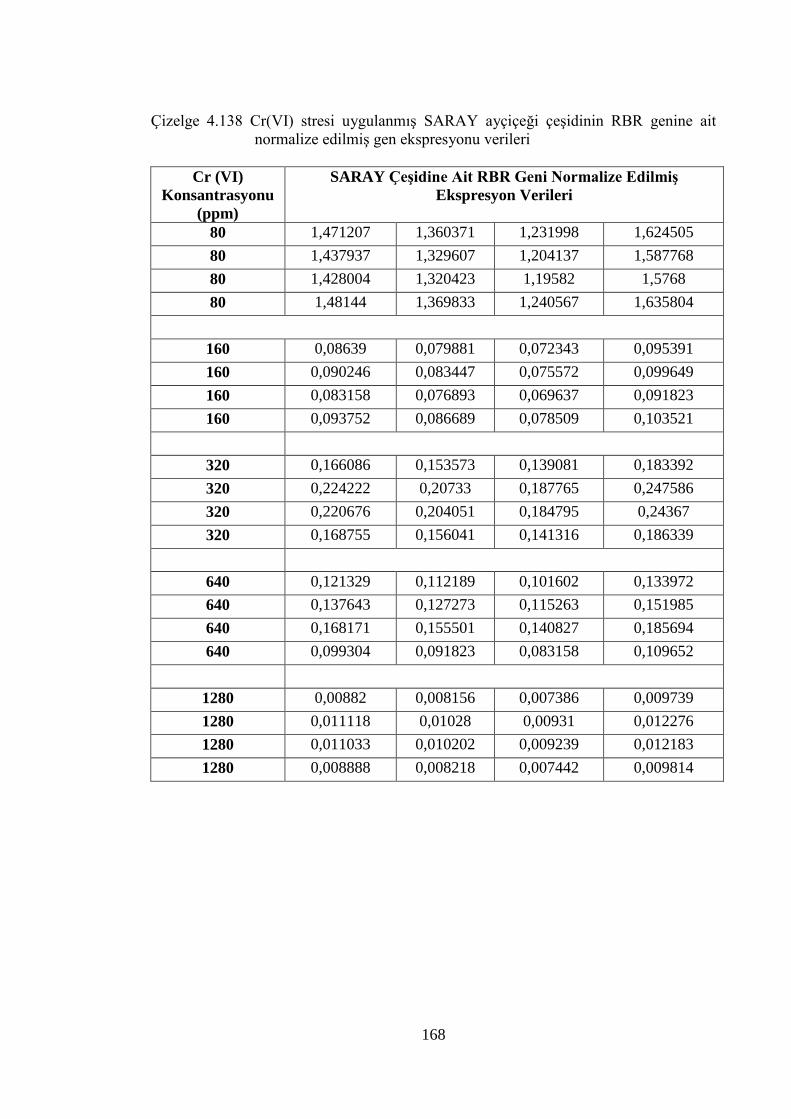
168
Çizelge 4.138 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
SARAY Çeşidine Ait RBR Geni Normalize Edilmiş
Ekspresyon Verileri
80 1,471207 1,360371 1,231998 1,624505
80 1,437937 1,329607 1,204137 1,587768
80 1,428004 1,320423 1,19582 1,5768
80 1,48144 1,369833 1,240567 1,635804
160 0,08639 0,079881 0,072343 0,095391
160 0,090246 0,083447 0,075572 0,099649
160 0,083158 0,076893 0,069637 0,091823
160 0,093752 0,086689 0,078509 0,103521
320 0,166086 0,153573 0,139081 0,183392
320 0,224222 0,20733 0,187765 0,247586
320 0,220676 0,204051 0,184795 0,24367
320 0,168755 0,156041 0,141316 0,186339
640 0,121329 0,112189 0,101602 0,133972
640 0,137643 0,127273 0,115263 0,151985
640 0,168171 0,155501 0,140827 0,185694
640 0,099304 0,091823 0,083158 0,109652
1280 0,00882 0,008156 0,007386 0,009739
1280 0,011118 0,01028 0,00931 0,012276
1280 0,011033 0,010202 0,009239 0,012183
1280 0,008888 0,008218 0,007442 0,009814
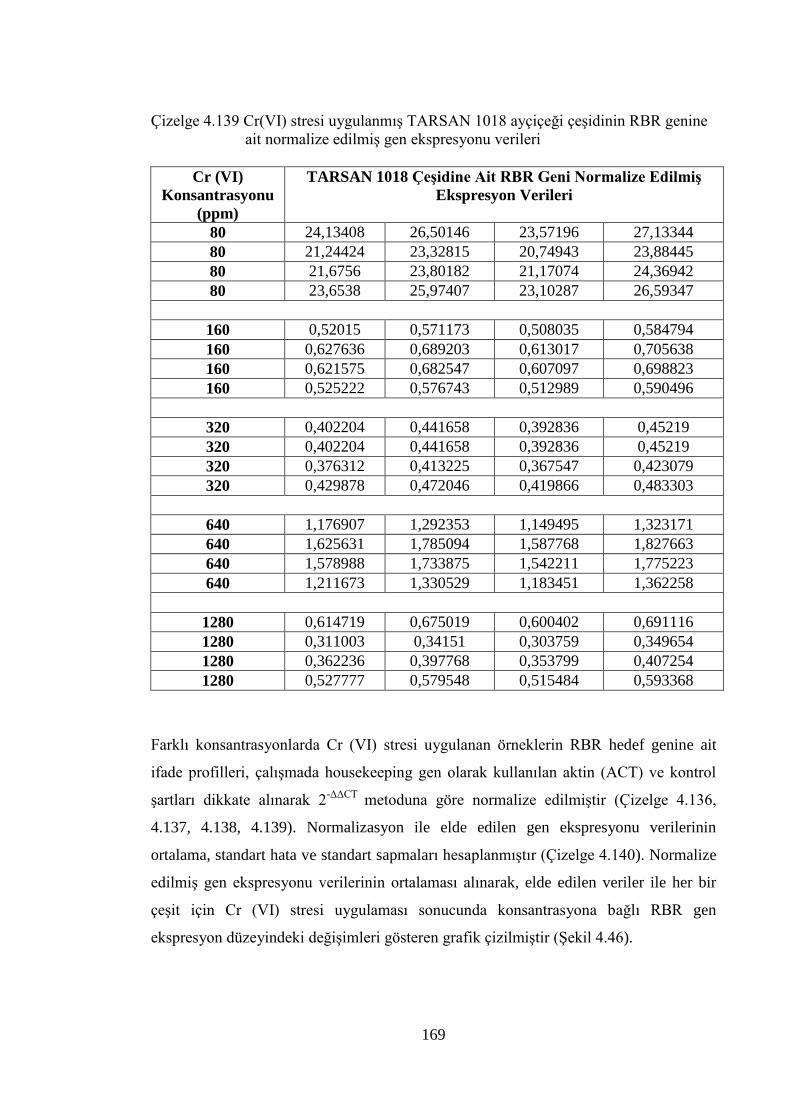
169
Çizelge 4.139 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR genine
ait normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TARSAN 1018 Çeşidine Ait RBR Geni Normalize Edilmiş
Ekspresyon Verileri
80 24,13408 26,50146 23,57196 27,13344
80 21,24424 23,32815 20,74943 23,88445
80 21,6756 23,80182 21,17074 24,36942
80 23,6538 25,97407 23,10287 26,59347
160 0,52015 0,571173 0,508035 0,584794
160 0,627636 0,689203 0,613017 0,705638
160 0,621575 0,682547 0,607097 0,698823
160 0,525222 0,576743 0,512989 0,590496
320 0,402204 0,441658 0,392836 0,45219
320 0,402204 0,441658 0,392836 0,45219
320 0,376312 0,413225 0,367547 0,423079
320 0,429878 0,472046 0,419866 0,483303
640 1,176907 1,292353 1,149495 1,323171
640 1,625631 1,785094 1,587768 1,827663
640 1,578988 1,733875 1,542211 1,775223
640 1,211673 1,330529 1,183451 1,362258
1280 0,614719 0,675019 0,600402 0,691116
1280 0,311003 0,34151 0,303759 0,349654
1280 0,362236 0,397768 0,353799 0,407254
1280 0,527777 0,579548 0,515484 0,593368
Farklı konsantrasyonlarda Cr (VI) stresi uygulanan örneklerin RBR hedef genine ait
ifade profilleri, çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol
şartları dikkate alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.136,
4.137, 4.138, 4.139). Normalizasyon ile elde edilen gen ekspresyonu verilerinin
ortalama, standart hata ve standart sapmaları hesaplanmıştır (Çizelge 4.140). Normalize
edilmiş gen ekspresyonu verilerinin ortalaması alınarak, elde edilen veriler ile her bir
çeşit için Cr (VI) stresi uygulaması sonucunda konsantrasyona bağlı RBR gen
ekspresyon düzeyindeki değişimleri gösteren grafik çizilmiştir (Şekil 4.46).
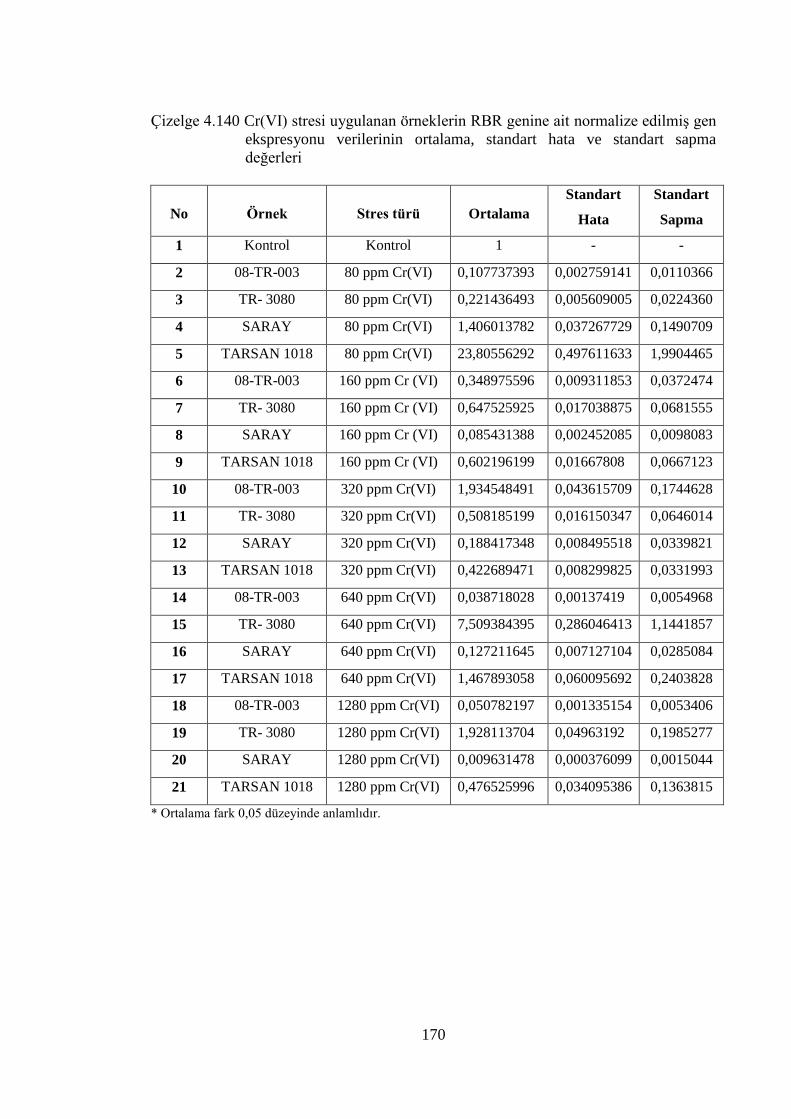
170
Çizelge 4.140 Cr(VI) stresi uygulanan örneklerin RBR genine ait normalize edilmiş gen
ekspresyonu verilerinin ortalama, standart hata ve standart sapma
değerleri
No
Örnek
Stres türü
Ortalama
Standart
Hata
Standart
Sapma
1 Kontrol Kontrol 1 - -
2 08-TR-003 80 ppm Cr(VI) 0,107737393 0,002759141 0,0110366
3 TR- 3080 80 ppm Cr(VI) 0,221436493 0,005609005 0,0224360
4 SARAY 80 ppm Cr(VI) 1,406013782 0,037267729 0,1490709
5 TARSAN 1018 80 ppm Cr(VI) 23,80556292 0,497611633 1,9904465
6 08-TR-003 160 ppm Cr (VI) 0,348975596 0,009311853 0,0372474
7 TR- 3080 160 ppm Cr (VI) 0,647525925 0,017038875 0,0681555
8 SARAY 160 ppm Cr (VI) 0,085431388 0,002452085 0,0098083
9 TARSAN 1018 160 ppm Cr (VI) 0,602196199 0,01667808 0,0667123
10 08-TR-003 320 ppm Cr(VI) 1,934548491 0,043615709 0,1744628
11 TR- 3080 320 ppm Cr(VI) 0,508185199 0,016150347 0,0646014
12 SARAY 320 ppm Cr(VI) 0,188417348 0,008495518 0,0339821
13 TARSAN 1018 320 ppm Cr(VI) 0,422689471 0,008299825 0,0331993
14 08-TR-003 640 ppm Cr(VI) 0,038718028 0,00137419 0,0054968
15 TR- 3080 640 ppm Cr(VI) 7,509384395 0,286046413 1,1441857
16 SARAY 640 ppm Cr(VI) 0,127211645 0,007127104 0,0285084
17 TARSAN 1018 640 ppm Cr(VI) 1,467893058 0,060095692 0,2403828
18 08-TR-003 1280 ppm Cr(VI) 0,050782197 0,001335154 0,0053406
19 TR- 3080 1280 ppm Cr(VI) 1,928113704 0,04963192 0,1985277
20 SARAY 1280 ppm Cr(VI) 0,009631478 0,000376099 0,0015044
21 TARSAN 1018 1280 ppm Cr(VI) 0,476525996 0,034095386 0,1363815
* Ortalama fark 0,05 düzeyinde anlamlıdır.
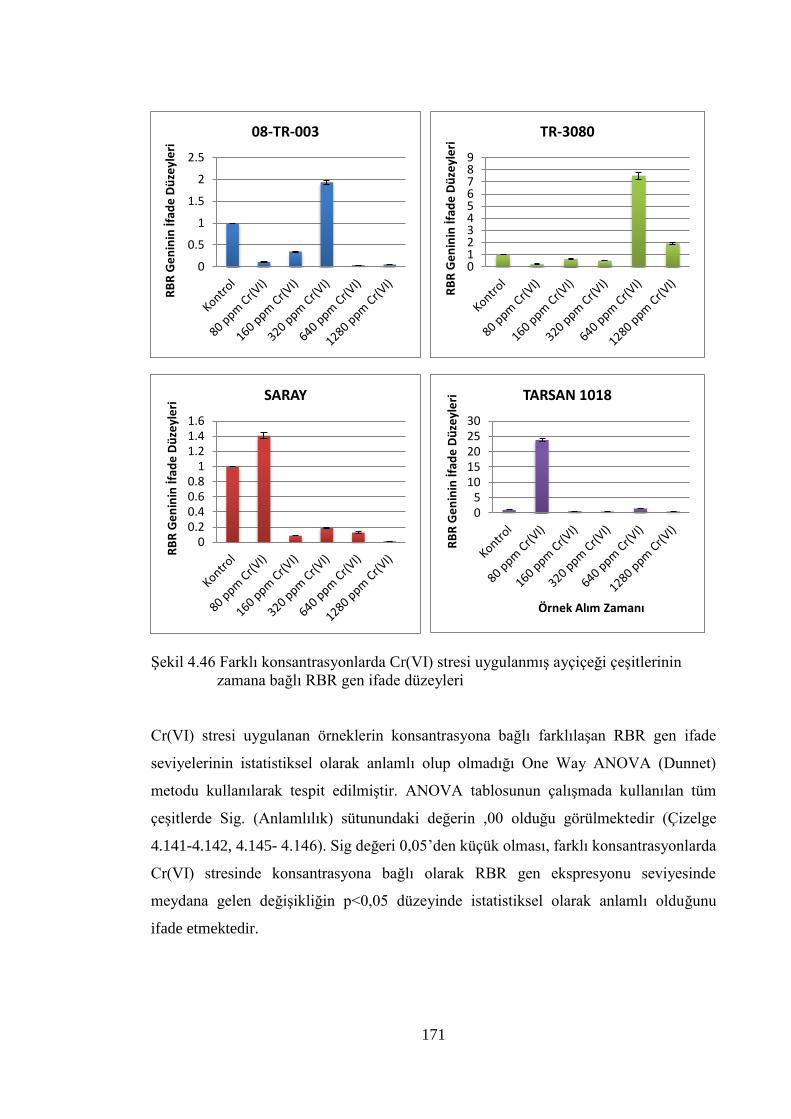
171
Şekil 4.46 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği çeşitlerinin
zamana bağlı RBR gen ifade düzeyleri
Cr(VI) stresi uygulanan örneklerin konsantrasyona bağlı farklılaşan RBR gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.141-4.142, 4.145- 4.146). Sig değeri 0,05’den küçük olması, farklı konsantrasyonlarda
Cr(VI) stresinde konsantrasyona bağlı olarak RBR gen ekspresyonu seviyesinde
meydana gelen değişikliğin p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu
ifade etmektedir.
0
0.5
1
1.5
2
2.5R
BR
Ge
nin
in İf
ade
Dü
zeyl
eri
08-TR-003
0123456789
RB
R G
en
inin
İfad
e D
üze
yle
ri TR-3080
00.20.40.60.8
11.21.41.6
RB
R G
en
inin
İfad
e D
üze
yle
ri SARAY
05
1015202530
RB
R G
en
inin
İfad
e D
üze
yle
ri
Örnek Alım Zamanı
TARSAN 1018
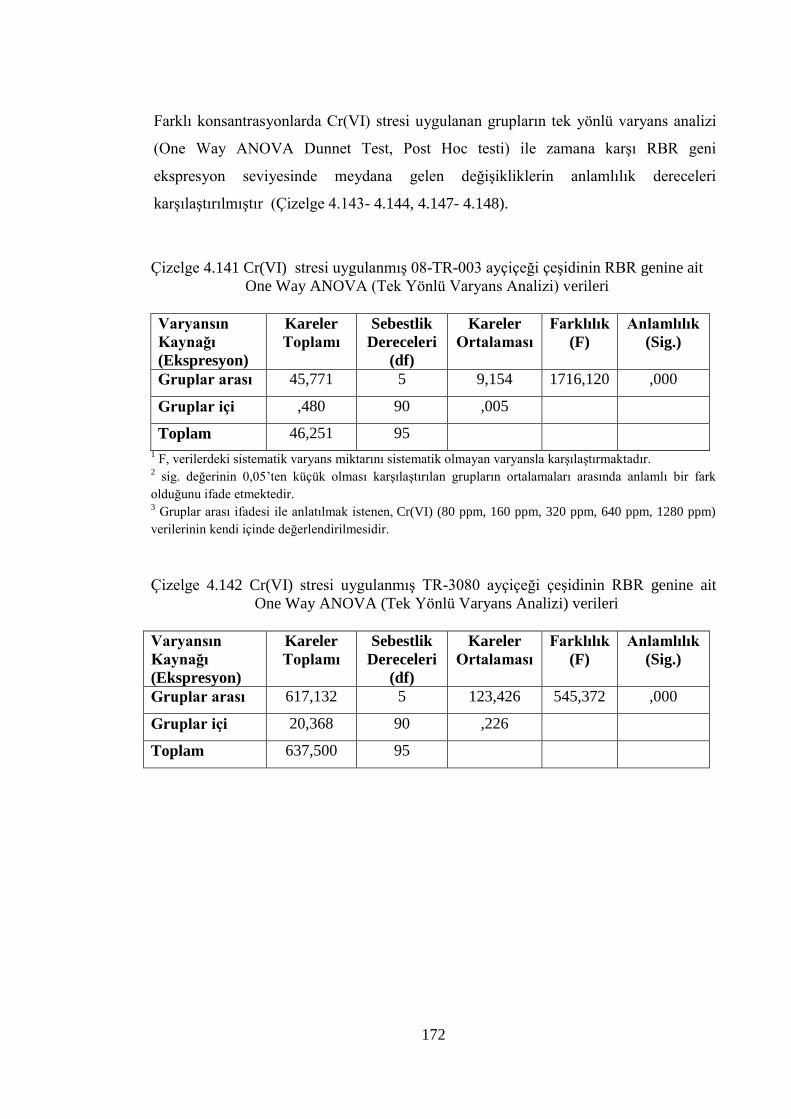
172
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan grupların tek yönlü varyans analizi
(One Way ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı RBR geni
ekspresyon seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri
karşılaştırılmıştır (Çizelge 4.143- 4.144, 4.147- 4.148).
Çizelge 4.141 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 45,771 5 9,154 1716,120 ,000
Gruplar içi ,480 90 ,005
Toplam 46,251 95
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
Cr(VI) (80 ppm, 160 ppm, 320 ppm, 640 ppm, 1280 ppm)
verilerinin kendi içinde değerlendirilmesidir.
Çizelge 4.142 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 617,132 5 123,426 545,372 ,000
Gruplar içi 20,368 90 ,226
Toplam 637,500 95
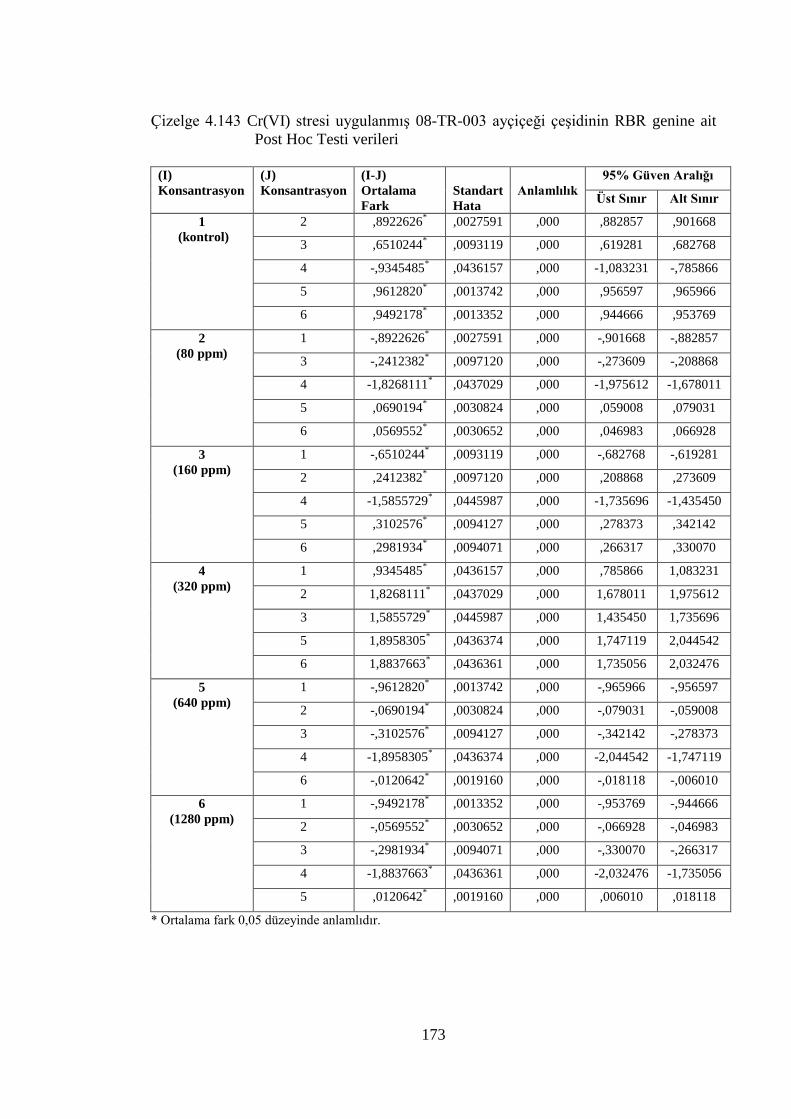
173
Çizelge 4.143 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin RBR genine ait
Post Hoc Testi verileri
(I)
Konsantrasyon
(J)
Konsantrasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8922626* ,0027591 ,000 ,882857 ,901668
3 ,6510244* ,0093119 ,000 ,619281 ,682768
4 -,9345485* ,0436157 ,000 -1,083231 -,785866
5 ,9612820* ,0013742 ,000 ,956597 ,965966
6 ,9492178* ,0013352 ,000 ,944666 ,953769
2
(80 ppm)
1 -,8922626* ,0027591 ,000 -,901668 -,882857
3 -,2412382* ,0097120 ,000 -,273609 -,208868
4 -1,8268111* ,0437029 ,000 -1,975612 -1,678011
5 ,0690194* ,0030824 ,000 ,059008 ,079031
6 ,0569552* ,0030652 ,000 ,046983 ,066928
3
(160 ppm)
1 -,6510244* ,0093119 ,000 -,682768 -,619281
2 ,2412382* ,0097120 ,000 ,208868 ,273609
4 -1,5855729* ,0445987 ,000 -1,735696 -1,435450
5 ,3102576* ,0094127 ,000 ,278373 ,342142
6 ,2981934* ,0094071 ,000 ,266317 ,330070
4
(320 ppm)
1 ,9345485* ,0436157 ,000 ,785866 1,083231
2 1,8268111* ,0437029 ,000 1,678011 1,975612
3 1,5855729* ,0445987 ,000 1,435450 1,735696
5 1,8958305* ,0436374 ,000 1,747119 2,044542
6 1,8837663* ,0436361 ,000 1,735056 2,032476
5
(640 ppm)
1 -,9612820* ,0013742 ,000 -,965966 -,956597
2 -,0690194* ,0030824 ,000 -,079031 -,059008
3 -,3102576* ,0094127 ,000 -,342142 -,278373
4 -1,8958305* ,0436374 ,000 -2,044542 -1,747119
6 -,0120642* ,0019160 ,000 -,018118 -,006010
6
(1280 ppm)
1 -,9492178* ,0013352 ,000 -,953769 -,944666
2 -,0569552* ,0030652 ,000 -,066928 -,046983
3 -,2981934* ,0094071 ,000 -,330070 -,266317
4 -1,8837663* ,0436361 ,000 -2,032476 -1,735056
5 ,0120642* ,0019160 ,000 ,006010 ,018118
* Ortalama fark 0,05 düzeyinde anlamlıdır.
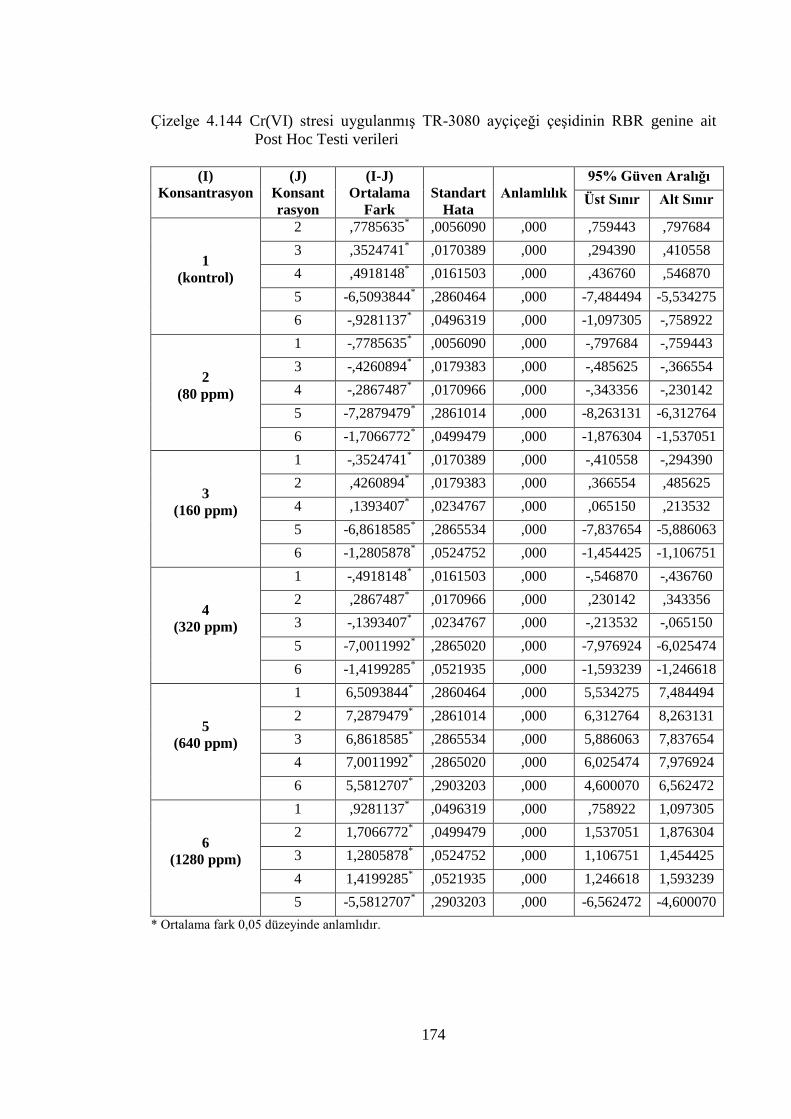
174
Çizelge 4.144 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin RBR genine ait
Post Hoc Testi verileri
(I)
Konsantrasyon
(J)
Konsant
rasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,7785635* ,0056090 ,000 ,759443 ,797684
3 ,3524741* ,0170389 ,000 ,294390 ,410558
4 ,4918148* ,0161503 ,000 ,436760 ,546870
5 -6,5093844* ,2860464 ,000 -7,484494 -5,534275
6 -,9281137* ,0496319 ,000 -1,097305 -,758922
2
(80 ppm)
1 -,7785635* ,0056090 ,000 -,797684 -,759443
3 -,4260894* ,0179383 ,000 -,485625 -,366554
4 -,2867487* ,0170966 ,000 -,343356 -,230142
5 -7,2879479* ,2861014 ,000 -8,263131 -6,312764
6 -1,7066772* ,0499479 ,000 -1,876304 -1,537051
3
(160 ppm)
1 -,3524741* ,0170389 ,000 -,410558 -,294390
2 ,4260894* ,0179383 ,000 ,366554 ,485625
4 ,1393407* ,0234767 ,000 ,065150 ,213532
5 -6,8618585* ,2865534 ,000 -7,837654 -5,886063
6 -1,2805878* ,0524752 ,000 -1,454425 -1,106751
4
(320 ppm)
1 -,4918148* ,0161503 ,000 -,546870 -,436760
2 ,2867487* ,0170966 ,000 ,230142 ,343356
3 -,1393407* ,0234767 ,000 -,213532 -,065150
5 -7,0011992* ,2865020 ,000 -7,976924 -6,025474
6 -1,4199285* ,0521935 ,000 -1,593239 -1,246618
5
(640 ppm)
1 6,5093844* ,2860464 ,000 5,534275 7,484494
2 7,2879479* ,2861014 ,000 6,312764 8,263131
3 6,8618585* ,2865534 ,000 5,886063 7,837654
4 7,0011992* ,2865020 ,000 6,025474 7,976924
6 5,5812707* ,2903203 ,000 4,600070 6,562472
6
(1280 ppm)
1 ,9281137* ,0496319 ,000 ,758922 1,097305
2 1,7066772* ,0499479 ,000 1,537051 1,876304
3 1,2805878* ,0524752 ,000 1,106751 1,454425
4 1,4199285* ,0521935 ,000 1,246618 1,593239
5 -5,5812707* ,2903203 ,000 -6,562472 -4,600070
* Ortalama fark 0,05 düzeyinde anlamlıdır.
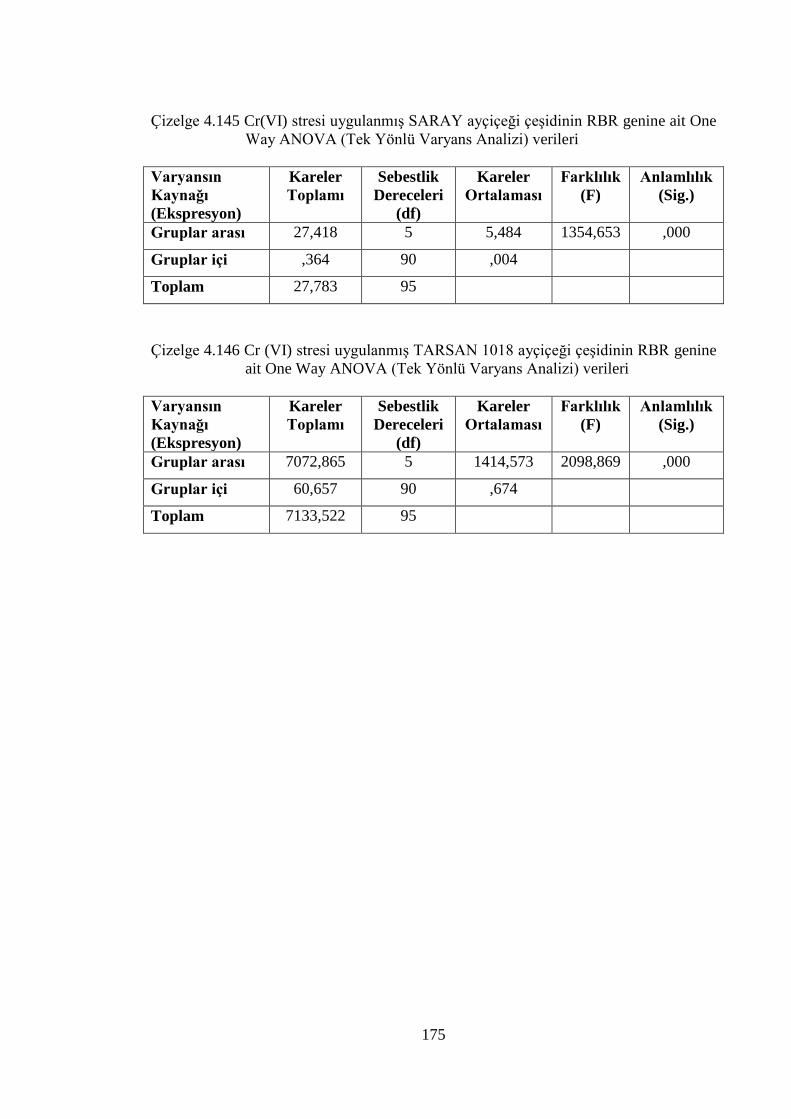
175
Çizelge 4.145 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine ait One
Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 27,418 5 5,484 1354,653 ,000
Gruplar içi ,364 90 ,004
Toplam 27,783 95
Çizelge 4.146 Cr (VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR genine
ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 7072,865 5 1414,573 2098,869 ,000
Gruplar içi 60,657 90 ,674
Toplam 7133,522 95
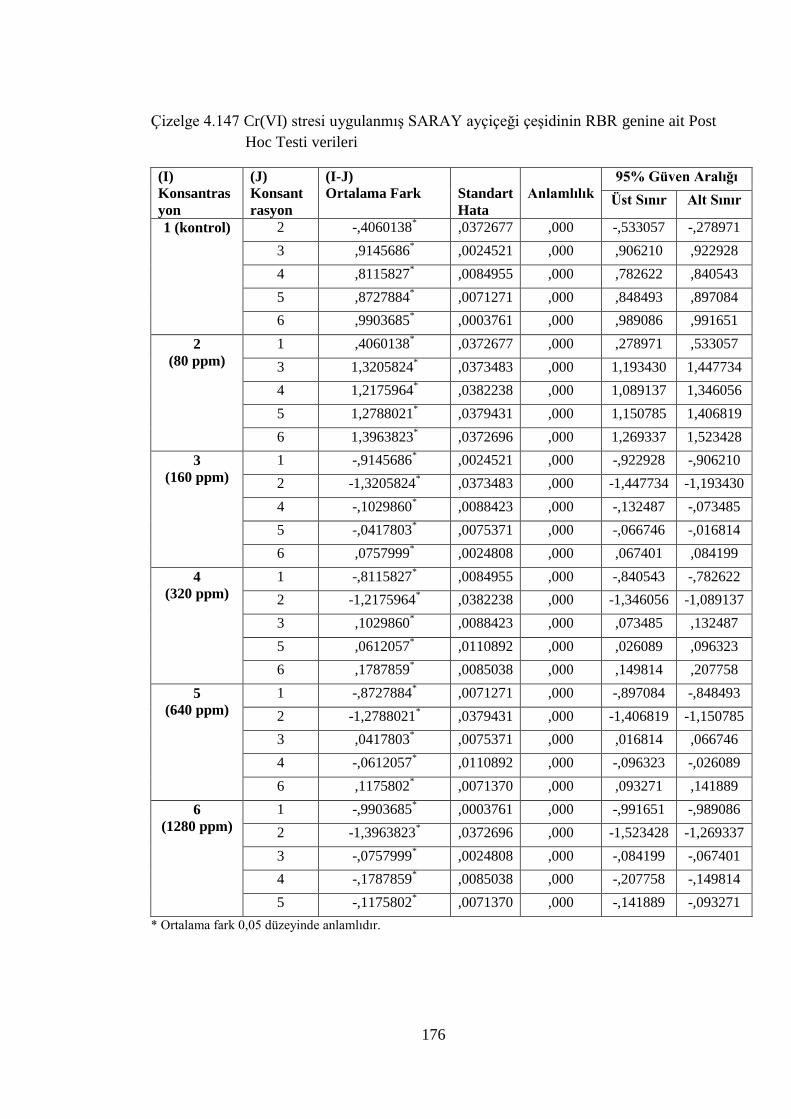
176
Çizelge 4.147 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin RBR genine ait Post
Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsant
rasyon
(I-J)
Ortalama Fark
Standart
Hata
Anlamlılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1 (kontrol) 2 -,4060138* ,0372677 ,000 -,533057 -,278971
3 ,9145686* ,0024521 ,000 ,906210 ,922928
4 ,8115827* ,0084955 ,000 ,782622 ,840543
5 ,8727884* ,0071271 ,000 ,848493 ,897084
6 ,9903685* ,0003761 ,000 ,989086 ,991651
2
(80 ppm)
1 ,4060138* ,0372677 ,000 ,278971 ,533057
3 1,3205824* ,0373483 ,000 1,193430 1,447734
4 1,2175964* ,0382238 ,000 1,089137 1,346056
5 1,2788021* ,0379431 ,000 1,150785 1,406819
6 1,3963823* ,0372696 ,000 1,269337 1,523428
3
(160 ppm)
1 -,9145686* ,0024521 ,000 -,922928 -,906210
2 -1,3205824* ,0373483 ,000 -1,447734 -1,193430
4 -,1029860* ,0088423 ,000 -,132487 -,073485
5 -,0417803* ,0075371 ,000 -,066746 -,016814
6 ,0757999* ,0024808 ,000 ,067401 ,084199
4
(320 ppm)
1 -,8115827* ,0084955 ,000 -,840543 -,782622
2 -1,2175964* ,0382238 ,000 -1,346056 -1,089137
3 ,1029860* ,0088423 ,000 ,073485 ,132487
5 ,0612057* ,0110892 ,000 ,026089 ,096323
6 ,1787859* ,0085038 ,000 ,149814 ,207758
5
(640 ppm)
1 -,8727884* ,0071271 ,000 -,897084 -,848493
2 -1,2788021* ,0379431 ,000 -1,406819 -1,150785
3 ,0417803* ,0075371 ,000 ,016814 ,066746
4 -,0612057* ,0110892 ,000 -,096323 -,026089
6 ,1175802* ,0071370 ,000 ,093271 ,141889
6
(1280 ppm)
1 -,9903685* ,0003761 ,000 -,991651 -,989086
2 -1,3963823* ,0372696 ,000 -1,523428 -1,269337
3 -,0757999* ,0024808 ,000 -,084199 -,067401
4 -,1787859* ,0085038 ,000 -,207758 -,149814
5 -,1175802* ,0071370 ,000 -,141889 -,093271
* Ortalama fark 0,05 düzeyinde anlamlıdır.
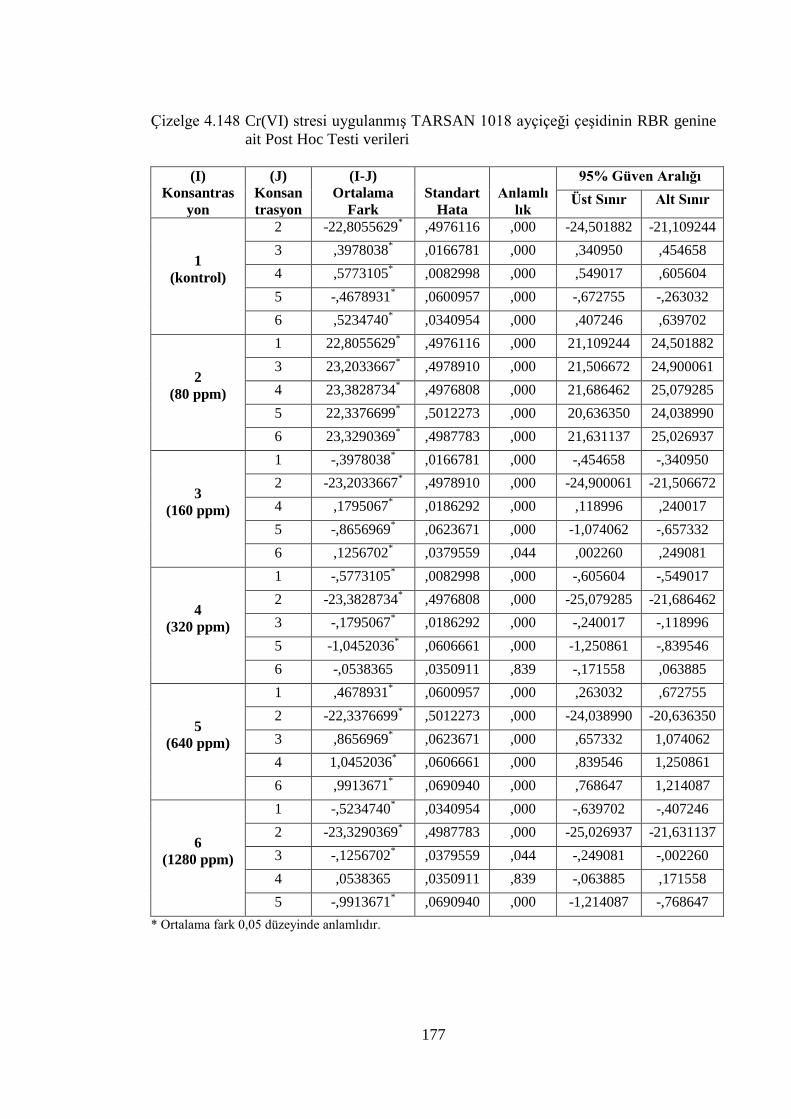
177
Çizelge 4.148 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin RBR genine
ait Post Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsan
trasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlı
lık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 -22,8055629* ,4976116 ,000 -24,501882 -21,109244
3 ,3978038* ,0166781 ,000 ,340950 ,454658
4 ,5773105* ,0082998 ,000 ,549017 ,605604
5 -,4678931* ,0600957 ,000 -,672755 -,263032
6 ,5234740* ,0340954 ,000 ,407246 ,639702
2
(80 ppm)
1 22,8055629* ,4976116 ,000 21,109244 24,501882
3 23,2033667* ,4978910 ,000 21,506672 24,900061
4 23,3828734* ,4976808 ,000 21,686462 25,079285
5 22,3376699* ,5012273 ,000 20,636350 24,038990
6 23,3290369* ,4987783 ,000 21,631137 25,026937
3
(160 ppm)
1 -,3978038* ,0166781 ,000 -,454658 -,340950
2 -23,2033667* ,4978910 ,000 -24,900061 -21,506672
4 ,1795067* ,0186292 ,000 ,118996 ,240017
5 -,8656969* ,0623671 ,000 -1,074062 -,657332
6 ,1256702* ,0379559 ,044 ,002260 ,249081
4
(320 ppm)
1 -,5773105* ,0082998 ,000 -,605604 -,549017
2 -23,3828734* ,4976808 ,000 -25,079285 -21,686462
3 -,1795067* ,0186292 ,000 -,240017 -,118996
5 -1,0452036* ,0606661 ,000 -1,250861 -,839546
6 -,0538365 ,0350911 ,839 -,171558 ,063885
5
(640 ppm)
1 ,4678931* ,0600957 ,000 ,263032 ,672755
2 -22,3376699* ,5012273 ,000 -24,038990 -20,636350
3 ,8656969* ,0623671 ,000 ,657332 1,074062
4 1,0452036* ,0606661 ,000 ,839546 1,250861
6 ,9913671* ,0690940 ,000 ,768647 1,214087
6
(1280 ppm)
1 -,5234740* ,0340954 ,000 -,639702 -,407246
2 -23,3290369* ,4987783 ,000 -25,026937 -21,631137
3 -,1256702* ,0379559 ,044 -,249081 -,002260
4 ,0538365 ,0350911 ,839 -,063885 ,171558
5 -,9913671* ,0690940 ,000 -1,214087 -,768647
* Ortalama fark 0,05 düzeyinde anlamlıdır.
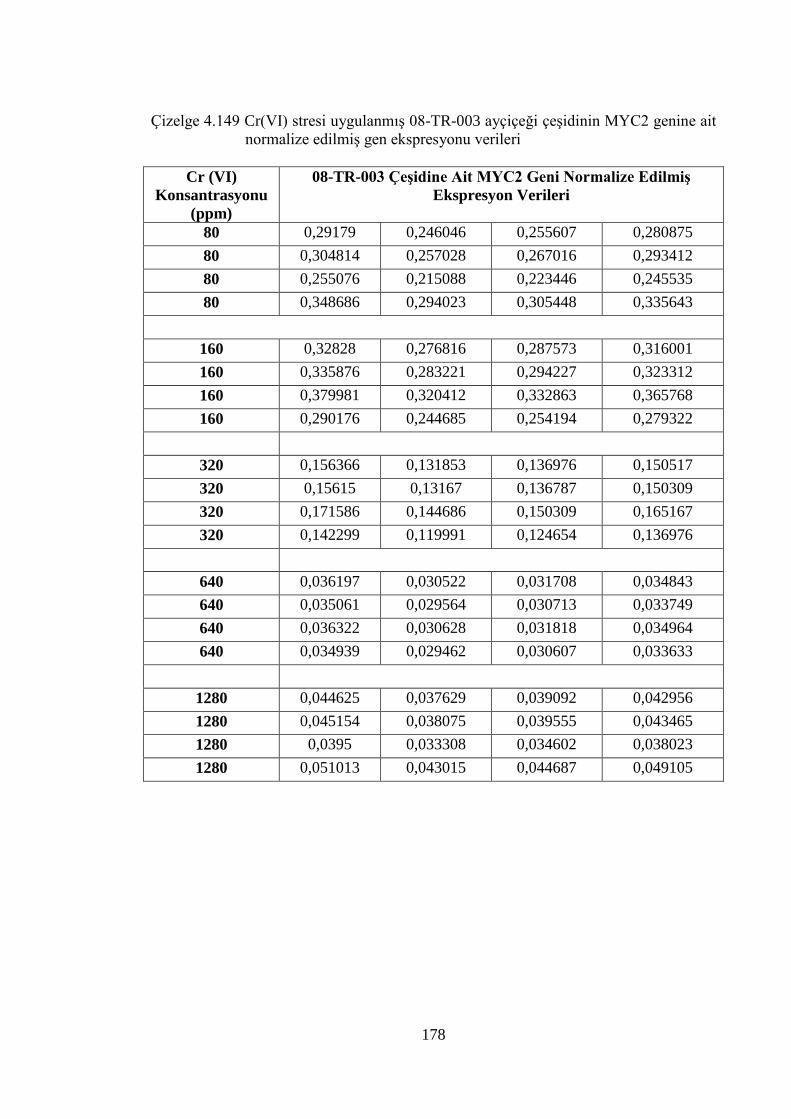
178
Çizelge 4.149 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
08-TR-003 Çeşidine Ait MYC2 Geni Normalize Edilmiş
Ekspresyon Verileri
80 0,29179 0,246046 0,255607 0,280875
80 0,304814 0,257028 0,267016 0,293412
80 0,255076 0,215088 0,223446 0,245535
80 0,348686 0,294023 0,305448 0,335643
160 0,32828 0,276816 0,287573 0,316001
160 0,335876 0,283221 0,294227 0,323312
160 0,379981 0,320412 0,332863 0,365768
160 0,290176 0,244685 0,254194 0,279322
320 0,156366 0,131853 0,136976 0,150517
320 0,15615 0,13167 0,136787 0,150309
320 0,171586 0,144686 0,150309 0,165167
320 0,142299 0,119991 0,124654 0,136976
640 0,036197 0,030522 0,031708 0,034843
640 0,035061 0,029564 0,030713 0,033749
640 0,036322 0,030628 0,031818 0,034964
640 0,034939 0,029462 0,030607 0,033633
1280 0,044625 0,037629 0,039092 0,042956
1280 0,045154 0,038075 0,039555 0,043465
1280 0,0395 0,033308 0,034602 0,038023
1280 0,051013 0,043015 0,044687 0,049105
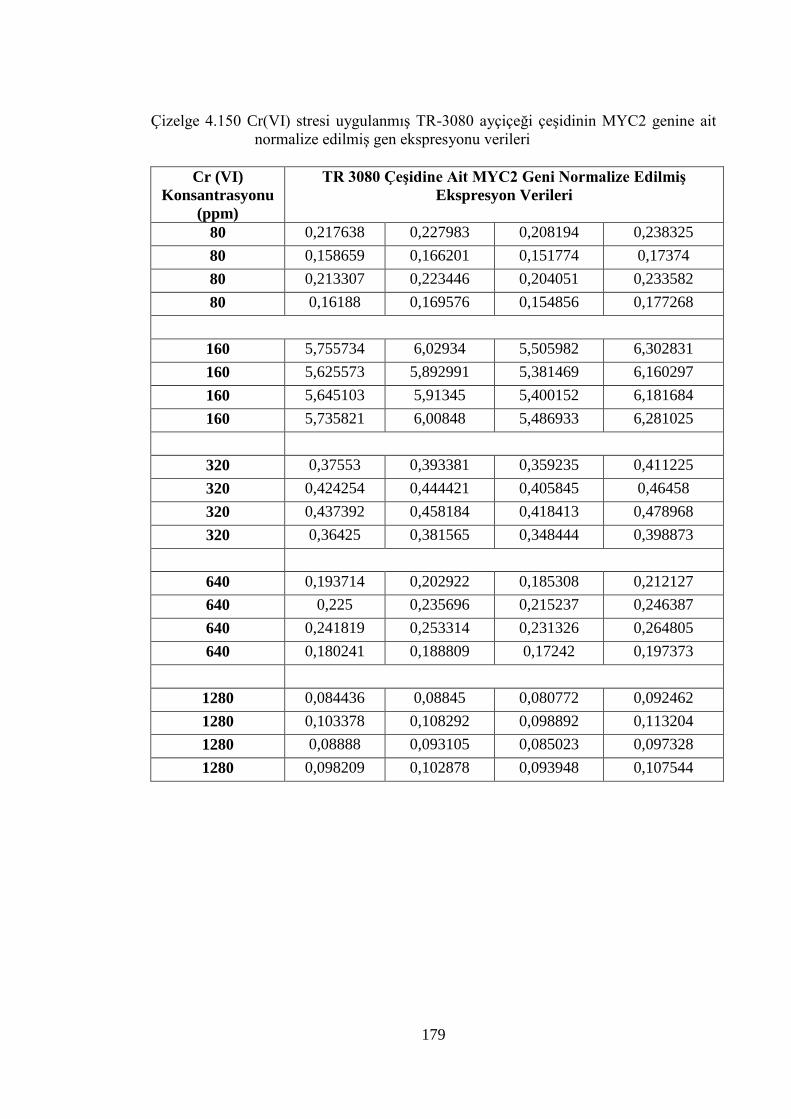
179
Çizelge 4.150 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TR 3080 Çeşidine Ait MYC2 Geni Normalize Edilmiş
Ekspresyon Verileri
80 0,217638 0,227983 0,208194 0,238325
80 0,158659 0,166201 0,151774 0,17374
80 0,213307 0,223446 0,204051 0,233582
80 0,16188 0,169576 0,154856 0,177268
160 5,755734 6,02934 5,505982 6,302831
160 5,625573 5,892991 5,381469 6,160297
160 5,645103 5,91345 5,400152 6,181684
160 5,735821 6,00848 5,486933 6,281025
320 0,37553 0,393381 0,359235 0,411225
320 0,424254 0,444421 0,405845 0,46458
320 0,437392 0,458184 0,418413 0,478968
320 0,36425 0,381565 0,348444 0,398873
640 0,193714 0,202922 0,185308 0,212127
640 0,225 0,235696 0,215237 0,246387
640 0,241819 0,253314 0,231326 0,264805
640 0,180241 0,188809 0,17242 0,197373
1280 0,084436 0,08845 0,080772 0,092462
1280 0,103378 0,108292 0,098892 0,113204
1280 0,08888 0,093105 0,085023 0,097328
1280 0,098209 0,102878 0,093948 0,107544
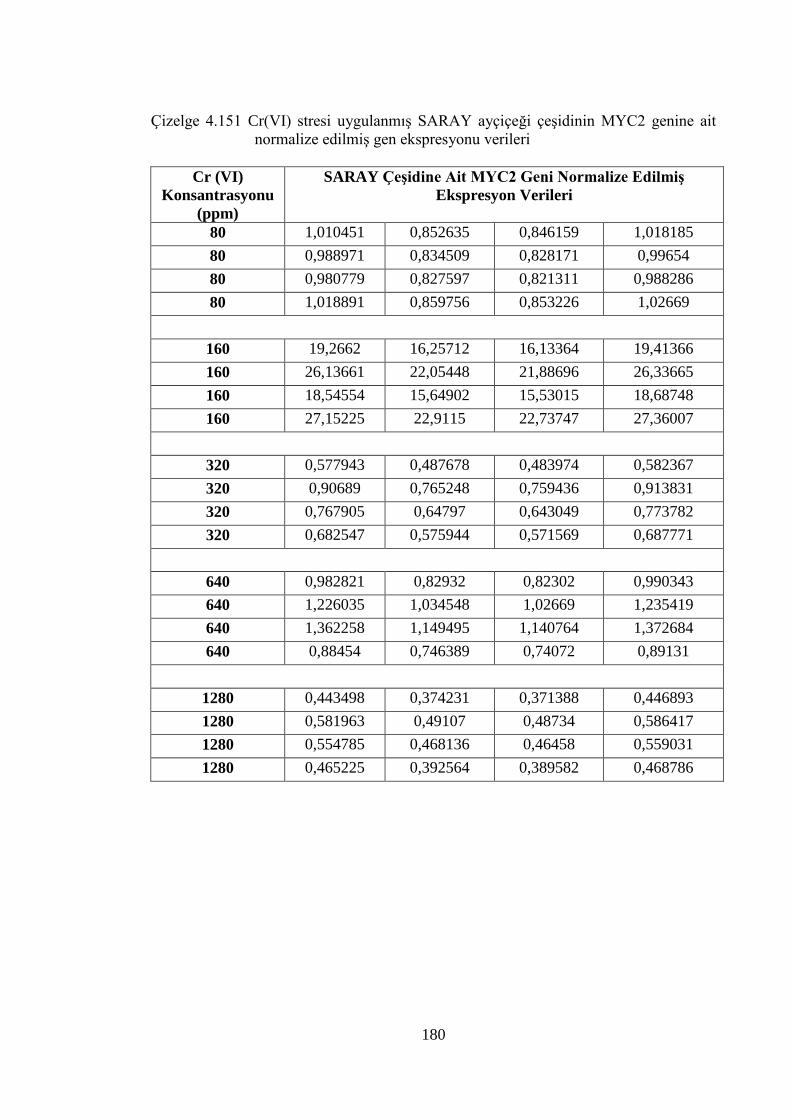
180
Çizelge 4.151 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine ait
normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
SARAY Çeşidine Ait MYC2 Geni Normalize Edilmiş
Ekspresyon Verileri
80 1,010451 0,852635 0,846159 1,018185
80 0,988971 0,834509 0,828171 0,99654
80 0,980779 0,827597 0,821311 0,988286
80 1,018891 0,859756 0,853226 1,02669
160 19,2662 16,25712 16,13364 19,41366
160 26,13661 22,05448 21,88696 26,33665
160 18,54554 15,64902 15,53015 18,68748
160 27,15225 22,9115 22,73747 27,36007
320 0,577943 0,487678 0,483974 0,582367
320 0,90689 0,765248 0,759436 0,913831
320 0,767905 0,64797 0,643049 0,773782
320 0,682547 0,575944 0,571569 0,687771
640 0,982821 0,82932 0,82302 0,990343
640 1,226035 1,034548 1,02669 1,235419
640 1,362258 1,149495 1,140764 1,372684
640 0,88454 0,746389 0,74072 0,89131
1280 0,443498 0,374231 0,371388 0,446893
1280 0,581963 0,49107 0,48734 0,586417
1280 0,554785 0,468136 0,46458 0,559031
1280 0,465225 0,392564 0,389582 0,468786
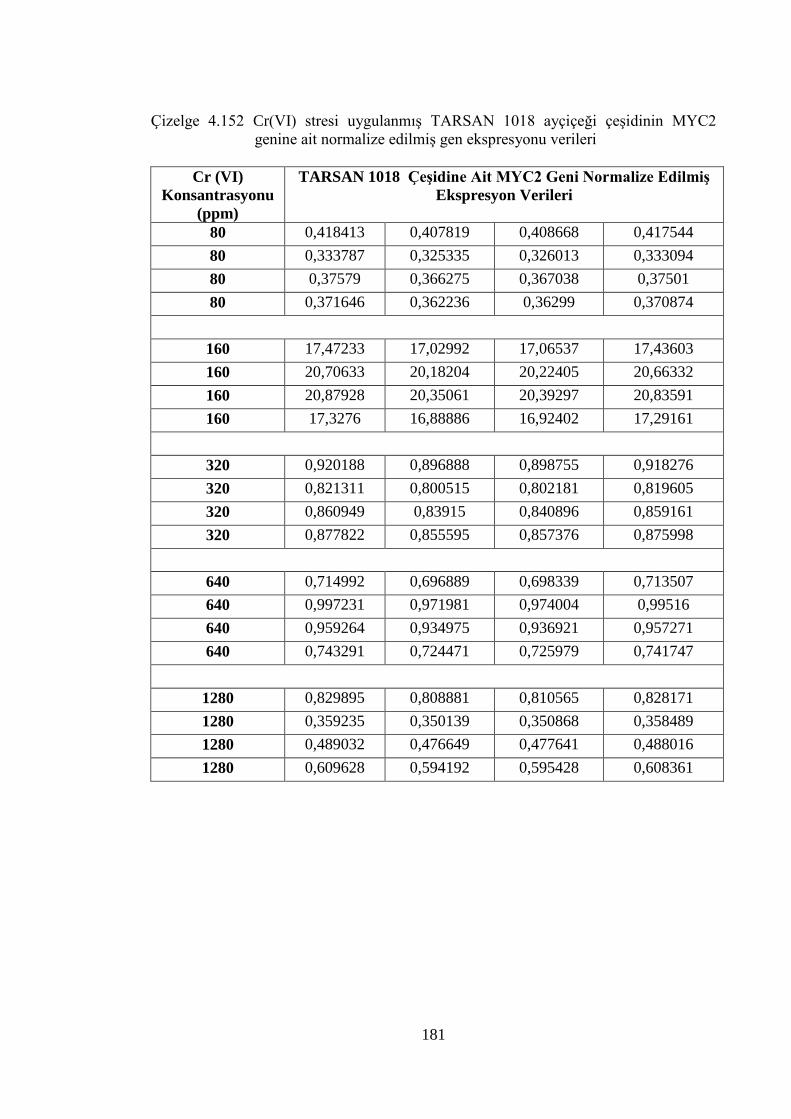
181
Çizelge 4.152 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait normalize edilmiş gen ekspresyonu verileri
Cr (VI)
Konsantrasyonu
(ppm)
TARSAN 1018 Çeşidine Ait MYC2 Geni Normalize Edilmiş
Ekspresyon Verileri
80 0,418413 0,407819 0,408668 0,417544
80 0,333787 0,325335 0,326013 0,333094
80 0,37579 0,366275 0,367038 0,37501
80 0,371646 0,362236 0,36299 0,370874
160 17,47233 17,02992 17,06537 17,43603
160 20,70633 20,18204 20,22405 20,66332
160 20,87928 20,35061 20,39297 20,83591
160 17,3276 16,88886 16,92402 17,29161
320 0,920188 0,896888 0,898755 0,918276
320 0,821311 0,800515 0,802181 0,819605
320 0,860949 0,83915 0,840896 0,859161
320 0,877822 0,855595 0,857376 0,875998
640 0,714992 0,696889 0,698339 0,713507
640 0,997231 0,971981 0,974004 0,99516
640 0,959264 0,934975 0,936921 0,957271
640 0,743291 0,724471 0,725979 0,741747
1280 0,829895 0,808881 0,810565 0,828171
1280 0,359235 0,350139 0,350868 0,358489
1280 0,489032 0,476649 0,477641 0,488016
1280 0,609628 0,594192 0,595428 0,608361
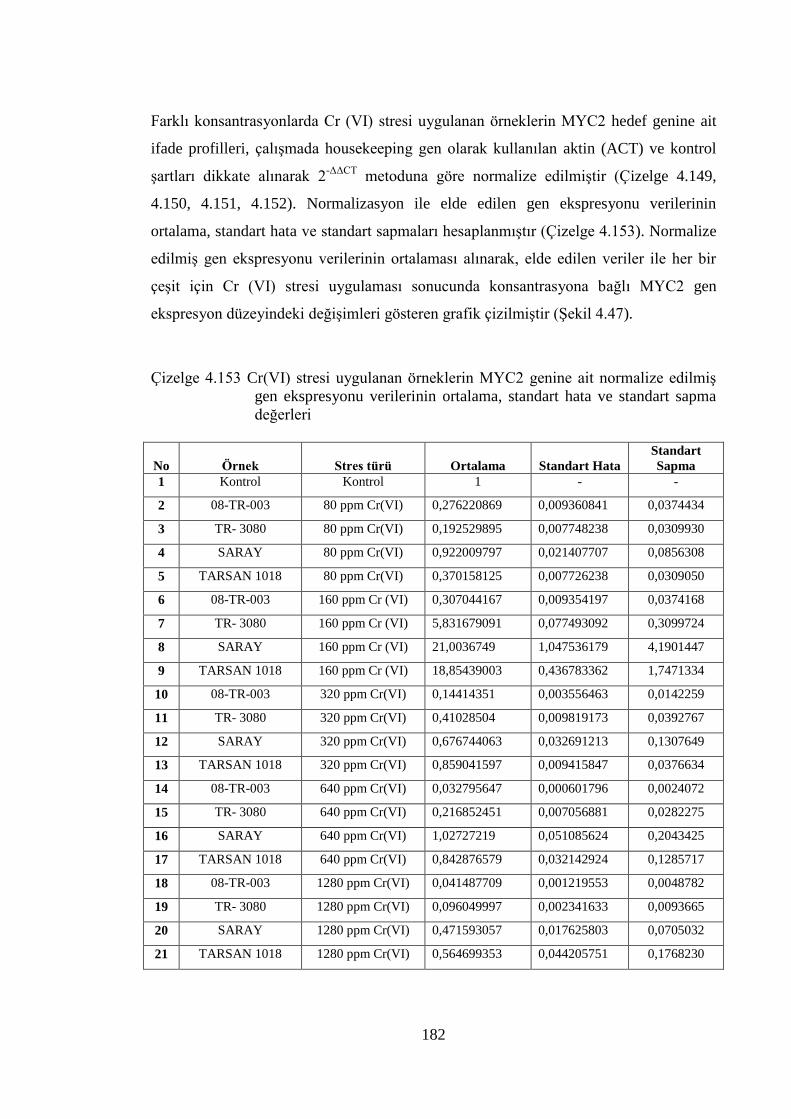
182
Farklı konsantrasyonlarda Cr (VI) stresi uygulanan örneklerin MYC2 hedef genine ait
ifade profilleri, çalışmada housekeeping gen olarak kullanılan aktin (ACT) ve kontrol
şartları dikkate alınarak 2-ΔΔCT
metoduna göre normalize edilmiştir (Çizelge 4.149,
4.150, 4.151, 4.152). Normalizasyon ile elde edilen gen ekspresyonu verilerinin
ortalama, standart hata ve standart sapmaları hesaplanmıştır (Çizelge 4.153). Normalize
edilmiş gen ekspresyonu verilerinin ortalaması alınarak, elde edilen veriler ile her bir
çeşit için Cr (VI) stresi uygulaması sonucunda konsantrasyona bağlı MYC2 gen
ekspresyon düzeyindeki değişimleri gösteren grafik çizilmiştir (Şekil 4.47).
Çizelge 4.153 Cr(VI) stresi uygulanan örneklerin MYC2 genine ait normalize edilmiş
gen ekspresyonu verilerinin ortalama, standart hata ve standart sapma
değerleri
No
Örnek
Stres türü
Ortalama
Standart Hata
Standart
Sapma
1 Kontrol Kontrol 1 - -
2 08-TR-003 80 ppm Cr(VI) 0,276220869 0,009360841 0,0374434
3 TR- 3080 80 ppm Cr(VI) 0,192529895 0,007748238 0,0309930
4 SARAY 80 ppm Cr(VI) 0,922009797 0,021407707 0,0856308
5 TARSAN 1018 80 ppm Cr(VI) 0,370158125 0,007726238 0,0309050
6 08-TR-003 160 ppm Cr (VI) 0,307044167 0,009354197 0,0374168
7 TR- 3080 160 ppm Cr (VI) 5,831679091 0,077493092 0,3099724
8 SARAY 160 ppm Cr (VI) 21,0036749 1,047536179 4,1901447
9 TARSAN 1018 160 ppm Cr (VI) 18,85439003 0,436783362 1,7471334
10 08-TR-003 320 ppm Cr(VI) 0,14414351 0,003556463 0,0142259
11 TR- 3080 320 ppm Cr(VI) 0,41028504 0,009819173 0,0392767
12 SARAY 320 ppm Cr(VI) 0,676744063 0,032691213 0,1307649
13 TARSAN 1018 320 ppm Cr(VI) 0,859041597 0,009415847 0,0376634
14 08-TR-003 640 ppm Cr(VI) 0,032795647 0,000601796 0,0024072
15 TR- 3080 640 ppm Cr(VI) 0,216852451 0,007056881 0,0282275
16 SARAY 640 ppm Cr(VI) 1,02727219 0,051085624 0,2043425
17 TARSAN 1018 640 ppm Cr(VI) 0,842876579 0,032142924 0,1285717
18 08-TR-003 1280 ppm Cr(VI) 0,041487709 0,001219553 0,0048782
19 TR- 3080 1280 ppm Cr(VI) 0,096049997 0,002341633 0,0093665
20 SARAY 1280 ppm Cr(VI) 0,471593057 0,017625803 0,0705032
21 TARSAN 1018 1280 ppm Cr(VI) 0,564699353 0,044205751 0,1768230
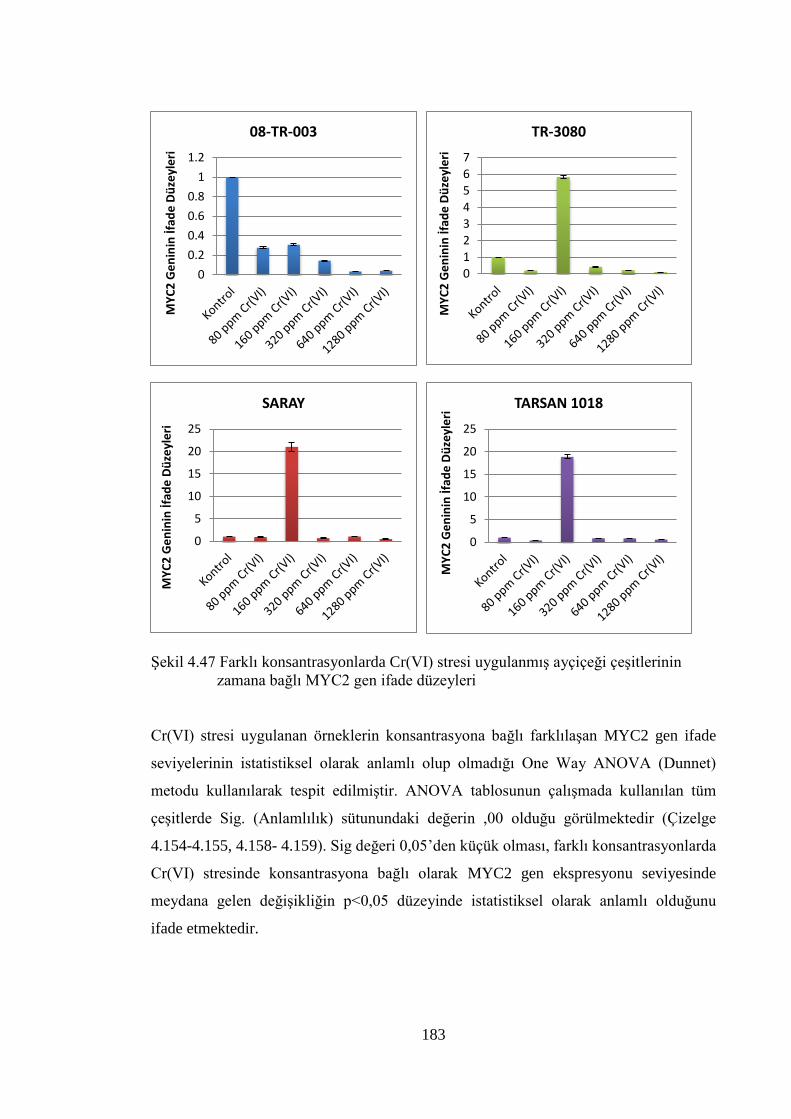
183
Şekil 4.47 Farklı konsantrasyonlarda Cr(VI) stresi uygulanmış ayçiçeği çeşitlerinin
zamana bağlı MYC2 gen ifade düzeyleri
Cr(VI) stresi uygulanan örneklerin konsantrasyona bağlı farklılaşan MYC2 gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiştir. ANOVA tablosunun çalışmada kullanılan tüm
çeşitlerde Sig. (Anlamlılık) sütunundaki değerin ,00 olduğu görülmektedir (Çizelge
4.154-4.155, 4.158- 4.159). Sig değeri 0,05’den küçük olması, farklı konsantrasyonlarda
Cr(VI) stresinde konsantrasyona bağlı olarak MYC2 gen ekspresyonu seviyesinde
meydana gelen değişikliğin p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu
ifade etmektedir.
0
0.2
0.4
0.6
0.8
1
1.2M
YC
2 G
en
inin
İfad
e D
üze
yle
ri
08-TR-003
0
1
2
3
4
5
6
7
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
TR-3080
0
5
10
15
20
25
MY
C2
Ge
nin
in İf
ade
Dü
zeyl
eri
SARAY
0
5
10
15
20
25M
YC
2 G
en
inin
İfad
e D
üze
yle
ri TARSAN 1018
184
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan grupların tek yönlü varyans analizi
(One Way ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni
ekspresyon seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri
karşılaştırılmıştır (Çizelge 4.156-4.157, 4.160- 4.161).
Çizelge 4.154 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2 genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 10,450 5 2,090 4133,222 ,000
Gruplar içi ,046 90 ,001
Toplam 10,496 95
1 F, verilerdeki sistematik varyans miktarını sistematik olmayan varyansla karşılaştırmaktadır.
2 sig. değerinin 0,05’ten küçük olması karşılaştırılan grupların ortalamaları arasında anlamlı bir fark
olduğunu ifade etmektedir. 3 Gruplar arası ifadesi ile anlatılmak istenen,
Cr(VI) (80 ppm, 160 ppm, 320 ppm, 640 ppm, 1280 ppm)
verilerinin kendi içinde değerlendirilmesidir.
Çizelge 4.155 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2 genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 404,313 5 80,863 4877,575 ,000
Gruplar içi 1,492 90 ,017
Toplam 405,805 95
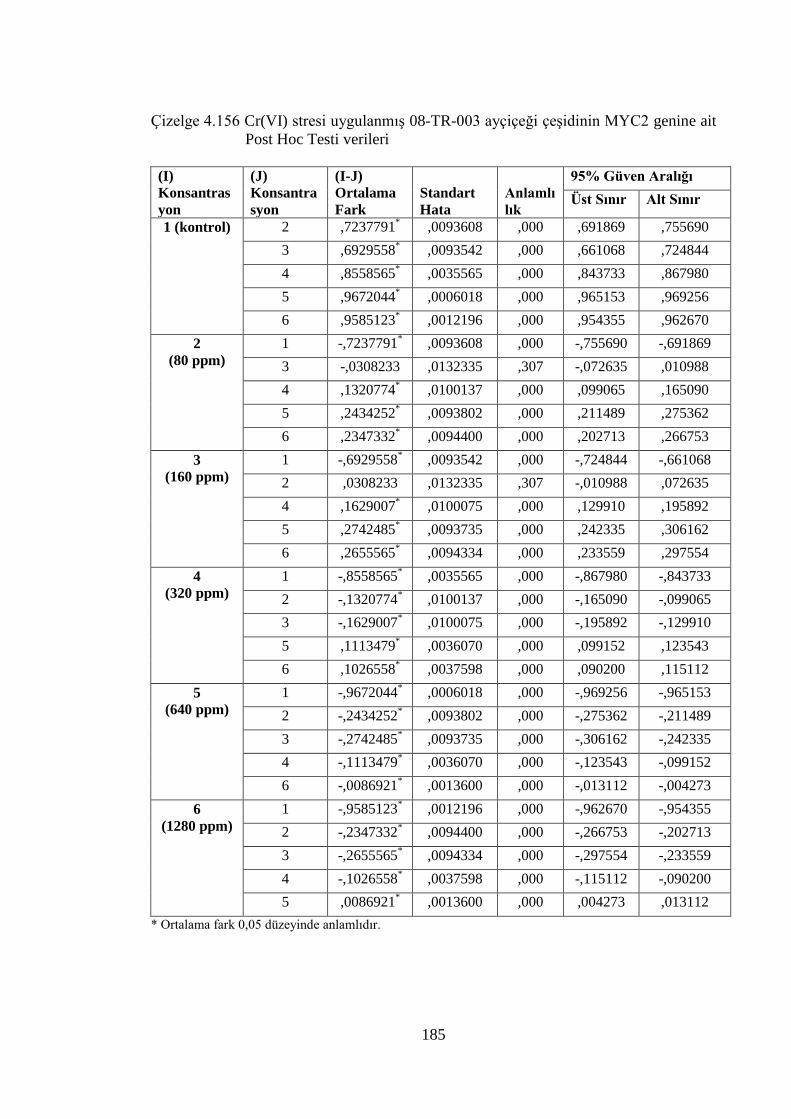
185
Çizelge 4.156 Cr(VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinin MYC2 genine ait
Post Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsantra
syon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlı
lık
95% Güven Aralığı
Üst Sınır Alt Sınır
1 (kontrol) 2 ,7237791* ,0093608 ,000 ,691869 ,755690
3 ,6929558* ,0093542 ,000 ,661068 ,724844
4 ,8558565* ,0035565 ,000 ,843733 ,867980
5 ,9672044* ,0006018 ,000 ,965153 ,969256
6 ,9585123* ,0012196 ,000 ,954355 ,962670
2
(80 ppm)
1 -,7237791* ,0093608 ,000 -,755690 -,691869
3 -,0308233 ,0132335 ,307 -,072635 ,010988
4 ,1320774* ,0100137 ,000 ,099065 ,165090
5 ,2434252* ,0093802 ,000 ,211489 ,275362
6 ,2347332* ,0094400 ,000 ,202713 ,266753
3
(160 ppm)
1 -,6929558* ,0093542 ,000 -,724844 -,661068
2 ,0308233 ,0132335 ,307 -,010988 ,072635
4 ,1629007* ,0100075 ,000 ,129910 ,195892
5 ,2742485* ,0093735 ,000 ,242335 ,306162
6 ,2655565* ,0094334 ,000 ,233559 ,297554
4
(320 ppm)
1 -,8558565* ,0035565 ,000 -,867980 -,843733
2 -,1320774* ,0100137 ,000 -,165090 -,099065
3 -,1629007* ,0100075 ,000 -,195892 -,129910
5 ,1113479* ,0036070 ,000 ,099152 ,123543
6 ,1026558* ,0037598 ,000 ,090200 ,115112
5
(640 ppm)
1 -,9672044* ,0006018 ,000 -,969256 -,965153
2 -,2434252* ,0093802 ,000 -,275362 -,211489
3 -,2742485* ,0093735 ,000 -,306162 -,242335
4 -,1113479* ,0036070 ,000 -,123543 -,099152
6 -,0086921* ,0013600 ,000 -,013112 -,004273
6
(1280 ppm)
1 -,9585123* ,0012196 ,000 -,962670 -,954355
2 -,2347332* ,0094400 ,000 -,266753 -,202713
3 -,2655565* ,0094334 ,000 -,297554 -,233559
4 -,1026558* ,0037598 ,000 -,115112 -,090200
5 ,0086921* ,0013600 ,000 ,004273 ,013112
* Ortalama fark 0,05 düzeyinde anlamlıdır.
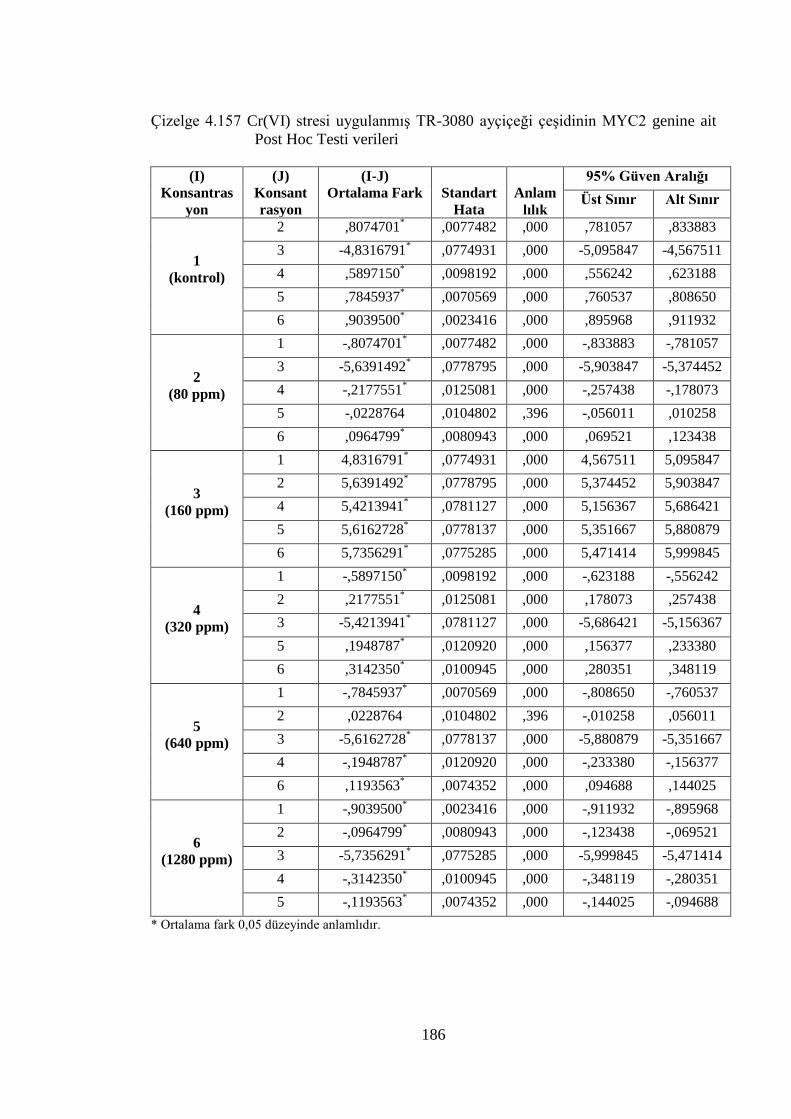
186
Çizelge 4.157 Cr(VI) stresi uygulanmış TR-3080 ayçiçeği çeşidinin MYC2 genine ait
Post Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsant
rasyon
(I-J)
Ortalama Fark
Standart
Hata
Anlam
lılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,8074701* ,0077482 ,000 ,781057 ,833883
3 -4,8316791* ,0774931 ,000 -5,095847 -4,567511
4 ,5897150* ,0098192 ,000 ,556242 ,623188
5 ,7845937* ,0070569 ,000 ,760537 ,808650
6 ,9039500* ,0023416 ,000 ,895968 ,911932
2
(80 ppm)
1 -,8074701* ,0077482 ,000 -,833883 -,781057
3 -5,6391492* ,0778795 ,000 -5,903847 -5,374452
4 -,2177551* ,0125081 ,000 -,257438 -,178073
5 -,0228764 ,0104802 ,396 -,056011 ,010258
6 ,0964799* ,0080943 ,000 ,069521 ,123438
3
(160 ppm)
1 4,8316791* ,0774931 ,000 4,567511 5,095847
2 5,6391492* ,0778795 ,000 5,374452 5,903847
4 5,4213941* ,0781127 ,000 5,156367 5,686421
5 5,6162728* ,0778137 ,000 5,351667 5,880879
6 5,7356291* ,0775285 ,000 5,471414 5,999845
4
(320 ppm)
1 -,5897150* ,0098192 ,000 -,623188 -,556242
2 ,2177551* ,0125081 ,000 ,178073 ,257438
3 -5,4213941* ,0781127 ,000 -5,686421 -5,156367
5 ,1948787* ,0120920 ,000 ,156377 ,233380
6 ,3142350* ,0100945 ,000 ,280351 ,348119
5
(640 ppm)
1 -,7845937* ,0070569 ,000 -,808650 -,760537
2 ,0228764 ,0104802 ,396 -,010258 ,056011
3 -5,6162728* ,0778137 ,000 -5,880879 -5,351667
4 -,1948787* ,0120920 ,000 -,233380 -,156377
6 ,1193563* ,0074352 ,000 ,094688 ,144025
6
(1280 ppm)
1 -,9039500* ,0023416 ,000 -,911932 -,895968
2 -,0964799* ,0080943 ,000 -,123438 -,069521
3 -5,7356291* ,0775285 ,000 -5,999845 -5,471414
4 -,3142350* ,0100945 ,000 -,348119 -,280351
5 -,1193563* ,0074352 ,000 -,144025 -,094688
* Ortalama fark 0,05 düzeyinde anlamlıdır.
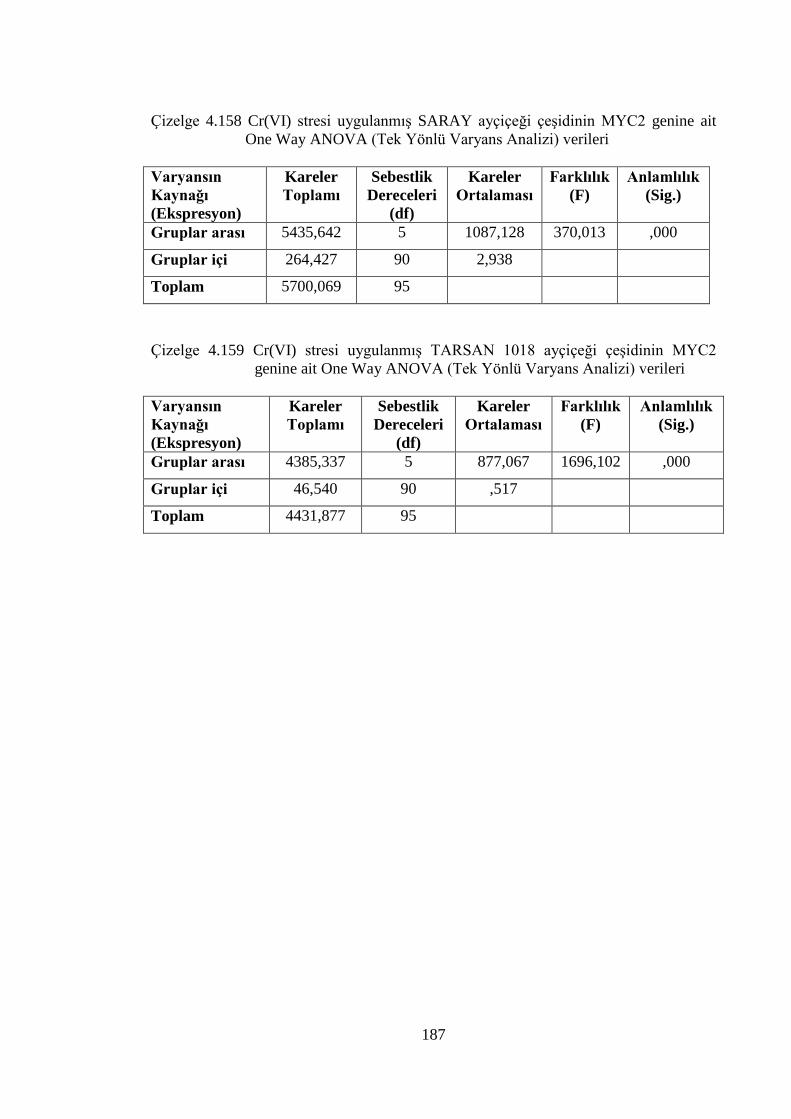
187
Çizelge 4.158 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine ait
One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 5435,642 5 1087,128 370,013 ,000
Gruplar içi 264,427 90 2,938
Toplam 5700,069 95
Çizelge 4.159 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait One Way ANOVA (Tek Yönlü Varyans Analizi) verileri
Varyansın
Kaynağı
(Ekspresyon)
Kareler
Toplamı
Sebestlik
Dereceleri
(df)
Kareler
Ortalaması
Farklılık
(F)
Anlamlılık
(Sig.)
Gruplar arası 4385,337 5 877,067 1696,102 ,000
Gruplar içi 46,540 90 ,517
Toplam 4431,877 95
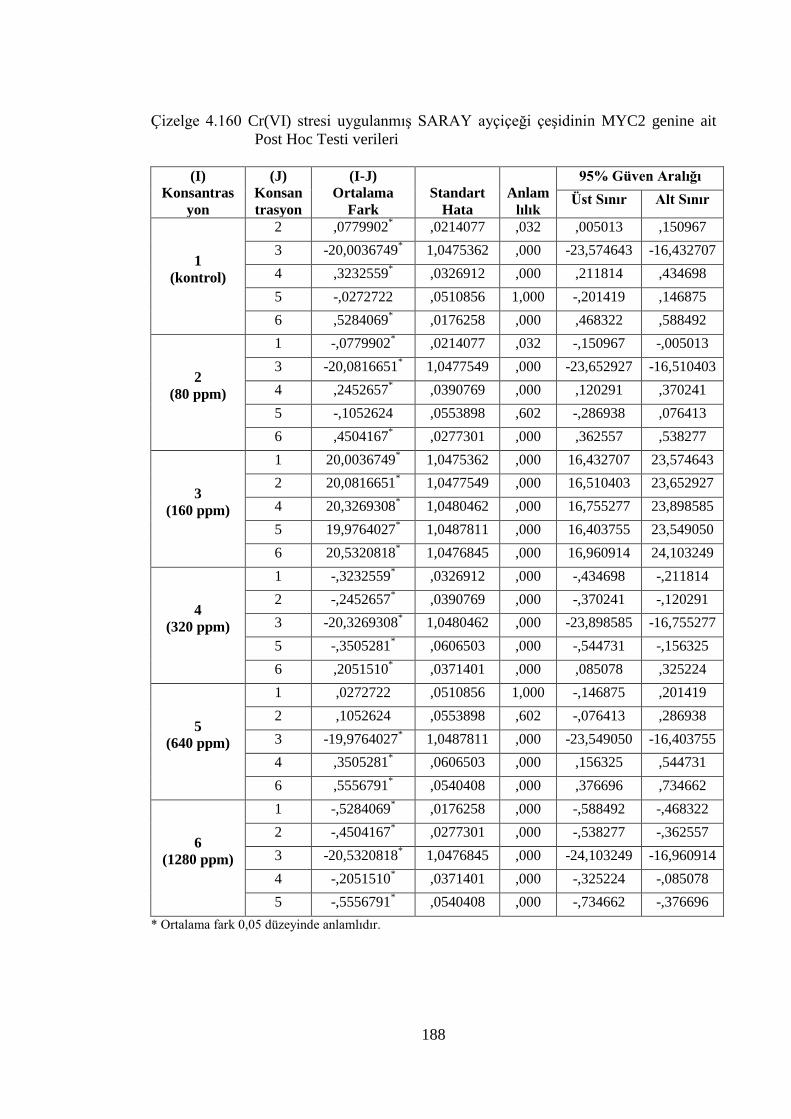
188
Çizelge 4.160 Cr(VI) stresi uygulanmış SARAY ayçiçeği çeşidinin MYC2 genine ait
Post Hoc Testi verileri
(I)
Konsantras
yon
(J)
Konsan
trasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlam
lılık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,0779902* ,0214077 ,032 ,005013 ,150967
3 -20,0036749* 1,0475362 ,000 -23,574643 -16,432707
4 ,3232559* ,0326912 ,000 ,211814 ,434698
5 -,0272722 ,0510856 1,000 -,201419 ,146875
6 ,5284069* ,0176258 ,000 ,468322 ,588492
2
(80 ppm)
1 -,0779902* ,0214077 ,032 -,150967 -,005013
3 -20,0816651* 1,0477549 ,000 -23,652927 -16,510403
4 ,2452657* ,0390769 ,000 ,120291 ,370241
5 -,1052624 ,0553898 ,602 -,286938 ,076413
6 ,4504167* ,0277301 ,000 ,362557 ,538277
3
(160 ppm)
1 20,0036749* 1,0475362 ,000 16,432707 23,574643
2 20,0816651* 1,0477549 ,000 16,510403 23,652927
4 20,3269308* 1,0480462 ,000 16,755277 23,898585
5 19,9764027* 1,0487811 ,000 16,403755 23,549050
6 20,5320818* 1,0476845 ,000 16,960914 24,103249
4
(320 ppm)
1 -,3232559* ,0326912 ,000 -,434698 -,211814
2 -,2452657* ,0390769 ,000 -,370241 -,120291
3 -20,3269308* 1,0480462 ,000 -23,898585 -16,755277
5 -,3505281* ,0606503 ,000 -,544731 -,156325
6 ,2051510* ,0371401 ,000 ,085078 ,325224
5
(640 ppm)
1 ,0272722 ,0510856 1,000 -,146875 ,201419
2 ,1052624 ,0553898 ,602 -,076413 ,286938
3 -19,9764027* 1,0487811 ,000 -23,549050 -16,403755
4 ,3505281* ,0606503 ,000 ,156325 ,544731
6 ,5556791* ,0540408 ,000 ,376696 ,734662
6
(1280 ppm)
1 -,5284069* ,0176258 ,000 -,588492 -,468322
2 -,4504167* ,0277301 ,000 -,538277 -,362557
3 -20,5320818* 1,0476845 ,000 -24,103249 -16,960914
4 -,2051510* ,0371401 ,000 -,325224 -,085078
5 -,5556791* ,0540408 ,000 -,734662 -,376696
* Ortalama fark 0,05 düzeyinde anlamlıdır.
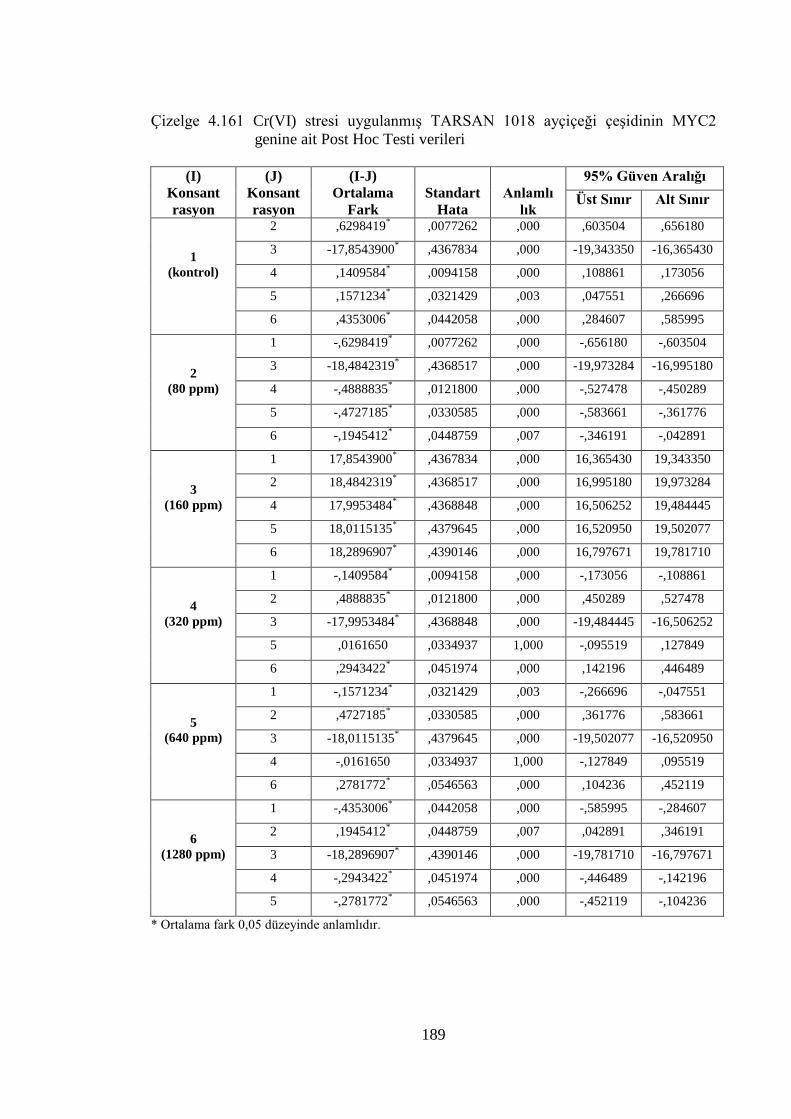
189
Çizelge 4.161 Cr(VI) stresi uygulanmış TARSAN 1018 ayçiçeği çeşidinin MYC2
genine ait Post Hoc Testi verileri
(I)
Konsant
rasyon
(J)
Konsant
rasyon
(I-J)
Ortalama
Fark
Standart
Hata
Anlamlı
lık
95% Güven Aralığı
Üst Sınır Alt Sınır
1
(kontrol)
2 ,6298419* ,0077262 ,000 ,603504 ,656180
3 -17,8543900* ,4367834 ,000 -19,343350 -16,365430
4 ,1409584* ,0094158 ,000 ,108861 ,173056
5 ,1571234* ,0321429 ,003 ,047551 ,266696
6 ,4353006* ,0442058 ,000 ,284607 ,585995
2
(80 ppm)
1 -,6298419* ,0077262 ,000 -,656180 -,603504
3 -18,4842319* ,4368517 ,000 -19,973284 -16,995180
4 -,4888835* ,0121800 ,000 -,527478 -,450289
5 -,4727185* ,0330585 ,000 -,583661 -,361776
6 -,1945412* ,0448759 ,007 -,346191 -,042891
3
(160 ppm)
1 17,8543900* ,4367834 ,000 16,365430 19,343350
2 18,4842319* ,4368517 ,000 16,995180 19,973284
4 17,9953484* ,4368848 ,000 16,506252 19,484445
5 18,0115135* ,4379645 ,000 16,520950 19,502077
6 18,2896907* ,4390146 ,000 16,797671 19,781710
4
(320 ppm)
1 -,1409584* ,0094158 ,000 -,173056 -,108861
2 ,4888835* ,0121800 ,000 ,450289 ,527478
3 -17,9953484* ,4368848 ,000 -19,484445 -16,506252
5 ,0161650 ,0334937 1,000 -,095519 ,127849
6 ,2943422* ,0451974 ,000 ,142196 ,446489
5
(640 ppm)
1 -,1571234* ,0321429 ,003 -,266696 -,047551
2 ,4727185* ,0330585 ,000 ,361776 ,583661
3 -18,0115135* ,4379645 ,000 -19,502077 -16,520950
4 -,0161650 ,0334937 1,000 -,127849 ,095519
6 ,2781772* ,0546563 ,000 ,104236 ,452119
6
(1280 ppm)
1 -,4353006* ,0442058 ,000 -,585995 -,284607
2 ,1945412* ,0448759 ,007 ,042891 ,346191
3 -18,2896907* ,4390146 ,000 -19,781710 -16,797671
4 -,2943422* ,0451974 ,000 -,446489 -,142196
5 -,2781772* ,0546563 ,000 -,452119 -,104236
* Ortalama fark 0,05 düzeyinde anlamlıdır.

190
5. TARTIŞMA VE SONUÇ
Bu çalışmada, 0, 3, 27 saat boyunca 100 ve 150 mM NaCl, 100 ve 150 mM PEG ve 24
saat boyunca farklı konsantrasyonlarda (0, 80 ppm, 160 ppm, 320 ppm ve 1280 ppm)
Cr(VI) stresi uygulanmış 08-TR-003, TR-3080, SARAY ve TARSAN 1018 ayçiçeği
çeşitlerinde, bitkilerde hücre döngüsü kontrolünde önemli roller üstlenen, dolayısıyla
hücre büyüme ve gelişiminin temel mekanizmalarını etkileyen Retinoblastoma ilişkili
protein (RBR), Myelocytomatosis (Myc) ve E2F genlerinin ifade seviyelerindeki
farklılıklar kantitatif Real Time PCR ile belirlenerek, ayçiçeğinin ilgili stres koşullarına
karşı olan savunma mekanizmaları hakkında bilgi edinilmeye çalışılmıştır. Günümüzde
moleküler biyoloji teknikleri oldukça gelişmiştir. Bu gelişmeler genom ve proteom
seviyesinde çalışmalar yapmamıza olanak sağlamaktadır. Herhangi bir organizmada
çeşitli koşullarda (hastalık, strese maruz kalma vb.) belirli genlere ait anlatım
profillerinin çıkarılmasında kullanılan en yaygın teknolojilerden biri kantitatif eş
zamanlı PCR yöntemidir. Real-time PCR teknolojisi ile araştırılan genlere ait transkript
seviyelerindeki değişimler oldukça doğru bir biçimde saptanabilmekte, sonuç alma
süresi önemli derecede kısalmakta ve personel hata payı en aza düşürülmektedir
(Shipley 2006). Çalışmada aynı zamanda bitkilerde stresin öncelikli etkilerinden biri
olarak gösterilen lipid peroksidasyonu sırasında oluşan reaktif bir metabolit ürün olan
malondialdehid (MDA) içeriği MDA analizi ile, stres faktörüne ve zamana bağlı olarak
ayçiçeği çeşitlerindeki total protein miktarındaki değişim de Bradford yöntemiyle tayin
edilmiştir.
5.1 Total Protein Miktarının Belirlenmesi (Bradford Yöntemi)
Abiyotik stresler arasında yer alan ozmotik streslerden kuraklık ve tuzluluk, bitkilerde
verim azalmasıyla sonuçlanan, çeşitli biyokimyasal, fizyolojik ve metabolik
değişikliklere neden olur (Xiong ve Zhu 2002).
Bitki büyümesinde olumsuz etkilere sahip olan tuzluluk bitki metabolizmasını
etkilemekte ve büyüme, gelişme ve gen ekspresyonunda önemli modifikasyonlara neden

191
olmaktadır. Bu modifikasyonlar bazı metabolitlerin birikimine veya tüketimine ve bazı
hücresel proteinlerin miktarında değişimlere neden olabilmektedir (Kong vd. 2005).
Tuz stresine maruz kalan pek çok bitki, ozmotik dengeyi sağlamak için hücrelerinde
düşük moleküler ağırlıklı çeşitli organik maddeleri yüksek konsantrasyonlarda
biriktirmektedir. Farklı bitki türlerinde tuzlulukla teşvik edilmiş çok sayıda protein
belirlenmiştir. Bu proteinleri, sadece tuzluluk şartlarında biriktirilen tuz stresi
proteinleri ve diğer abiyotik faktörlerce biriktirilen stresle ilgili proteinler olarak iki
gruba ayırmak mümkündür (Ashraf ve Haris 2004). Tuzlu koşullar altındaki bitkilerde
biriken proteinler, azot kaynağı olarak kullanılabilmekte ve ozmotik düzenlemede rol
oynayabilmektedir. Bu proteinler tuz stresine yanıt olarak yeni sentezlenebilir veya
düşük konsantrasyonda temel düzeyde mevcut olabilir (Pareek-Singla ve Grover 1997).
Tuzluluk koşulları altında uyarılan proteinlerin sitoplazmik proteinler oldukları ve
hücrelerin sitoplazmik akışkanlığında değişimlere neden olduğu bilinmektedir
(Hasegawa vd. 2000). Arpa, ayçiçeği, çeltik gibi bitkilerin tuza toleranslı genotiplerinin
daha yüksek çözünebilir protein içeriğine sahip oldukları gözlenmiştir (Parvaiz ve
Satyawati 2008).
Agastian vd. (2000) yaptıkları çalışmada, dut çeşitlerinde düşük tuz stresinde çözünür
protein miktarının arttığını, yüksek tuz stresinde azaldığını bildirmişlerdir. Ashraf ve
Fatima (1995) aspir bitkisinin tuza toleranslı ve tuz duyarlı çeşitlerinde yaprak çözünür
proteinlerinde önemli ölçüde farklılık olmadığını, tuzluluk cevabında çözünür protein
içeriğinde azalma olduğunu rapor etmişlerdir.
Alamgir ve Ali (1999)’nin Oryza sativa, Gadallah (1999)’ın Vicia faba, Wang ve Nil
(2000)’in Amaranthus tricolor, Parida vd. (2002)’nin Bruguiera parviflora ile yaptıkları
çalışmalarda tuzluluk koşullarında çözünebilir protein miktarında azalma olduğunu
tespit etmişlerdir. Quintero vd. (1996)’nin Arabidopsis thaliana ile yaptıkları
çalışmalarda tuzluluk koşullarında çözünebilir protein miktarında artış olduğunu rapor
etmişlerdir. Khedr vd. (2003)’nin kum zambağı (Pancratium maritimum) ile yaptıkları

192
çalışmada düşük tuzluluk ortamında total protein miktarında artış olduğunu, yüksek
tuzluluk ortamında ise total protein miktarında azalma olduğunu rapor etmişlerdir.
Huang vd. (2014), sodyum klorür konsantrasyonlarının hibrid rami bitkisi üzerinde,
fotosentez, antioksidan enzimler, büyüme ve lif verimi üzerine etkilerini araştırdıkları
çalışmada, bitkilere 10 ve 20 gün süreyle, farklı konsantrasyonlarda (0, 2, 4, 6 ve 8 g
NaCl/kg kuru toprak) NaCl stresi uygulamışlardır. 10 gün boyunca strese maruz kalan
bitkilerde, 20 güne kıyasla total protein miktarının daha fazla olduğu görülse de artan
NaCl konsantrasyonuyla beraber her iki zaman diliminde de total protein miktarında
azalma olduğunu tespit etmişlerdir.
Tuzlu koşullarda büyüyen bitkilerde, tuz stresi poliribozomların kaybolmasına ve
dolayısıyla protein sentez ve seviyesinin azalmasına neden olmaktadır (Artlip ve
Funkhouser 1995).
Farklı bitki türlerinde tuz stresi altında protein seviyesindeki azalma; protein sentezinde
ve amino asitlerin kullanılabilirliğinde azalma, amino asitler ve protein sentezini
kapsayan enzimlerin denatürasyonuna bağlıdır (Popova vd. 1995).
Tuzlu koşullarda artan protein seviyesi ise, proteinlerin yeni oluşumları kadar önceden
mevcut proteinlerin artan sentezinden de kaynaklanabilir (Dubey ve Rani 1989).
NaCl stresine karşı miktarı artan protein(ler)in türlerin savunma mekanizmasını
oluşturabileceği, aynı proteinlerin miktarının azalması durumunda ise stresin yeterince
üstesinden gelemeyeceği ve proteinin azalması veya kaybolmasının bir genotipin
duyarlılığı ile ilişkili olabileceği ileri sürülmüştür (Yıldız ve Terzi 2008).
Ashraf ve Tufail (1995) ayçiçeği bitkisinde tuzluluk tolerans değişimini araştırdıkları
çalışmada, tuza toleranslı çeşitlerde, tuza hassas çeşitlere göre çözünebilir protein
seviyesinin daha yüksek olduğunu bildirmişlerdir.

193
Wang vd. (2014), Kandelia candel köklerinde, antioksidan savunma sistemi üzerine tuz
stresinin etkilerini araştırdıkları çalışmalarında, bitkilere 0, 150, 300 ve 450 mmol/L tuz
stresi uygulamışlardır. Tuz konsantrasyonu ve işlem süresi artması ile Kandelia candel
kökü toplam protein içeriğinin azaldığını rapor etmişlerdir.
Çalışmamızda, 100 mM ve 150 mM tuz (NaCl) stresi uygulaması sonucu, 08-TR-003,
TR-3080 ve SARAY ayçiçeği çeşitlerinde kontrol örnekleriyle kıyaslandığında 3. saatte
total protein miktarında azalma görülürken, 27. saatte 3. saate göre bir artış gözlenmiştir
(Şekil 4.19). TARSAN 1018 ayçiçeği çeşidinde ise, 100 mM tuz stresi uygulaması
sonucu 3. saatte kontrole göre artış, 27. saatte ise hem kontrole, hem de 3. saate göre
azalma gözlenmiştir. Aynı çeşide, 150 mM tuz stresi uygulaması sonucu 3. ve 27.
saatlerde alınan örneklerde total protein miktarında stres uygulama zamanına bağlı
olarak artış gözlenmiştir (Şekil 4.20). Elde ettiğimiz sonuçlar, literatürde yer alan
çalışmalar doğrultusunda değerlendirildiğinde, 100mM ve 150 mM tuz stresinin kısa
süreli uygulanmasının 08-TR-003, TR-3080 ve SARAY çeşitleri üzerinde bir etki
oluşturduğu, stresin uygulama süresindeki artışla beraber protein seviyesinde meydana
gelen artışın bitkilerde stres tolerans mekanizmalarını aktif hale getirmiş olabileceği
sonucuna varılabilir. TARSAN 1018 çeşidinde 100 mM NaCl stresinde, protein
seviyesinde 3. saatte gözlenen artış, bu çeşidin diğerlerine kıyasla tuz stresine karşı
tolerans mekanizmalarını daha erken aktif hale getirdiği ve dolayısıyla daha toleranslı
olduğunu gösterebilir.
Kuraklık zararı sadece su kaybından değil, aynı zamanda protein kaybından da meydana
gelir. Kuraklık stresi ile bitkide protein metabolizmasında, proteinlerin parçalanması ve
protein sentezinin azalması şeklinde görülen bozulmalar meydana gelmektedir. Kuraklık
stresi altında uyarılan genler; hücreleri su yokluğundan yalnızca önemli metabolik
proteinlerin üretimi yoluyla korumak üzere fonksiyon göstermez, aynı zamanda
kuraklık stresi cevabında sinyal iletim genlerinin regülasyonunda da görev alır
(Shinozaki vd. 1996).

194
100 mM PEG (kuraklık) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinde total protein
miktarında 3. saatte kontrole göre bir azalma görülürken, 27. saatte 3. saate göre anlamlı
bir değişme gözlenmemiştir (Şekil 4.21). 150 mM PEG uygulaması sonucu aynı çeşitte
kontrole göre örnek alım zamanına bağlı olarak azalma görülmüştür. 100 mM PEG
stresi uygulaması sonucu, TR-3080 ayçiçeği çeşidinde total protein miktarında 3. saatte
kontrole göre artma, 27.saatte 3. saate göre azalma görülmektedir. 150 mM PEG stresi
uygulamasında ise total protein miktarında 3. saatte kontrole göre azalma, 27. saatte 3.
saate göre artış olduğu görülmektedir (Şekil 4.22). 100 mM ve 150 mM PEG stresi
uygulaması sonucunda SARAY ve TARSAN 1018 ayçiçeği çeşitlerinde 3. saatte
kontrole göre azalma, 27. saatte 3. saate göre artış gözlenmiştir. 100 mM ve 150 mM
PEG stresi uygulaması çalışılan çeşitlerde genel olarak aynı tepkilere sebep olmuştur.
100 mM ve 150 mM PEG stresinde, en yüksek çözünebilir protein miktarı sırasıyla, TR-
3080 ve TARSAN 1018 çeşidinde gözlenmiştir. Bu çeşitlerde kuraklığa toleransın daha
yüksek olduğu söylenebilir.
Sinha vd. (2005), Pistia stratiotes L. bitkisiyle yaptıkları çalışmada, 10μM Cr stresinin
48 saat uygulamasında görülen artışın dışında, tüm uygulama sürelerinde, Cr
konsantrasyonunda artışla beraber total protein miktarında azalma görülmüştür.
Rai vd. (2004), Ocimum tenuiflorum L. ile yaptıkları çalışmada, farklı
konsantrasyonlardaki (10, 20, 50 ve 100 μM) Cr stresi, farklı sürelerde (24, 48 ve 72
saat) uygulanmıştır. Genel olarak artan Cr konsantrasyonu ve sürelere bağlı olarak
protein miktarında azalma gözlenmiştir.
Rai vd. (2014), Catharanthus roseus’un Rosea ve Alba çeşitleri ile yaptıkları çalışmada,
30 gün süresince farklı konsantrasyonlarda (0, 10, 50 ve 100 μM) Cr stresi uygulaması
sonucunda, Rosea çeşidinde artan konsantrasyona bağlı protein miktarında kademeli
azalma görülmüştür. Alba çeşidinde ise en belirgin azalma 100 μM Cr stresi
uygulamasında gözlenmiştir.

195
Farklı konsantrasyonlarda Cr (VI) stresi uygulanmış 08-TR-003 ayçiçeği çeşidinde artan
krom konsantrasyonlarıyla total protein miktarında azalmalar olduğu görülmektedir.
Kontrol örneğiyle kıyaslandığında 640 ppm ve 1280 ppm Cr(VI) stresi uygulamasının
total protein miktarında önemli azalmalara sebep olduğu görülmektedir.
TR-3080 ayçiçeği çeşidinde 80 ppm Cr (VI) uygulamasının kontrole göre total protein
miktarında azalmaya sebep olurken, 160 ppm Cr (VI) uygulamasının 80 ppm
konsantrasyona göre bir miktar total protein miktarını arttırdığı görülmektedir. 160
ppm’den itibaren artan Cr konsantrasyonuna bağlı olarak total protein miktarında
azalma görülmektedir.
SARAY ayçiçeği çeşidinde 80 ppm Cr uygulamasının kontrole göre total protein
miktarını azalttığı gözlenirken 80, 160, 320 ve 640 ppm uygulamanın total protein
miktarında anlamlı bir değişime sebep olmadığı görülmektedir. 1280 ppm de ise anlamlı
azalma olduğu görülmektedir. TARSAN 1018 çeşidinde 80 ppm Cr uygulamasının
kontrole göre total protein miktarını azalttığı gözlenirken, 160ppm uygulamanın 80
ppm’e göre total protein miktarında anlamlı bir değişime sebep olmadığı görülmektedir.
320 ppm’ den itibaren artan Cr konsantrasyonuna bağlı total protein miktarında azalma
görülmektedir. Tüm çeşitlerde en anlamlı azalış 1280 ppm uygulaması sonucu
gözlenmiştir( Şekil 4.23).
Cr (VI) stresi uygulaması sonucu, total protein miktarında gözlenen değişimler, konuyla
ilgili daha önce yapılan çalışmaları (Rai vd. (2004, 2014), Sinha vd. (2005), Wang vd.
(2014)) destekler niteliktedir. Artan Cr (VI) konsantrasyonuyla beraber, total protein
miktarında bazı iniş çıkışlar görülse de, genel olarak azalma eğiliminde olduğu
gözlenmiştir. Kromun belli konsantrasyonlarına kadar, protein seviyesinin belli
seviyede tutulması, stres yanıt mekanizmalarının çalıştırıldığını, özellikle 640 ppm ve
1280 ppm’de gözlenen anlamlı azalmanın hücrelerin apoptoza girmesinden
kaynaklanabileceği düşünülmektedir.

196
5.2 Lipid Peroksidasyonunun Belirlenmesi (MDA Analizi)
Malondialdehit (MDA) içeriği değişimi, lipid peroksidasyonunu ölçmek için önemli bir
göstergedir. MDA içeriğinin yüksek olması, bitki hücre zarlarının oksidasyon
derecesinin yüksek olduğunu ve hasarın büyüklüğünü ifade eder (Cheruth vd. 2009).
Farklı çalışmalarda gösterildiği gibi, düşük MDA düzeyleri, tuz toleransı açısından
önemli olabilir. Tuz stresine maruz bırakılmış, tuza toleranslı arpa (Liang vd. 2003) ve
tütünde (Ruiz vd. 2005) kontrollere göre MDA düzeylerinin daha düşük olduğu
bulunmuştur.
Sergio vd. (2012), tuz stresinin yabani hindiba (Cichorium intybus L.)’da büyüme
parametreleri, enzimatik antioksidan sistem ve lipit peroksidasyonu üzerine etkilerini
araştırdıkları çalışmalarında, yaprakta gözlenen MDA düzeyinin köklerden daha fazla
olduğu, 100 mM ve 200 mM NaCl uygulamasının, hem yaprakta hem köklerde artan
NaCl konsantrasyonuna bağlı olarak MDA düzeyinde azalmaya sebep olduğunu
gözlemlemişlerdir.
Ellouzia vd. (2011), tuz stresinin Cakile maritima ve Arabidopsis thaliana’nın
fizyolojik ve oksidatif durumu üzerindeki etkilerini araştırdıkları çalışmalarında, 400
mM NaCl uygulamasının, Cakile maritima’da 4. saatte MDA düzeyini düşürdüğü, 16
ve 24 saat uygulamaların MDA düzeyini arttırdığı, 72 saat uygulamanın ise MDA
düzeyini düşürdüğünü gözlemişlerdir. Arabidopsis thaliana’da ise 100 mM NaCl
uygulamasının zamana bağlı MDA düzeyinde artışa sebep olduğunu gözlemişlerdir. Her
iki türde de Malondialdehid konsantrasyonları 24 saat tuz stresinde maksimum
değerlere ulaşmıştır.
Mittova vd. (2004) yaptıkları çalışmada, tuz toleranslı yabani domates Lycopersicon
pennellii köklerinde artan tuz stresine tepki olarak MDA düzeyinde bir azalma meydana
geldiğini bulmuşlardır. Yabani ve tuza toleranslı domates türlerinde tuz stresine cevabın

197
farklı olduğunu ve yabani türlerin tuz stresine yanıt olarak antioksidan sistemin artan
regülasyonunda daha başarılı olduklarını bulmuşlardır.
Ebrahimian ve Bybordi (2012) ayçiçeği ile yaptıkları çalışmada, askorbik asit
uygulamasının, lipid peroksidasyonu ve enzim aktivitesini önemli derecede azalttığını
bulmuşlardır. Öte yandan, tuzluluk ve askorbik asit arasındaki etkileşimin,
malondialdehit ve antioksidan enzim aktivitesi üzerinde önemli olduğu ve tuzluluk
stresi altında askorbik asit uygulamasının malondialdehit üretim ve enzim aktivitesini
önemli ölçüde düşürdüğünü gözlemlemişlerdir. Sadece 100, 200 ve 300 mM tuz stresi
uygulaması sonucu, artan NaCl konsantrasyonlarına bağlı MDA düzeyinin de arttığı
bulunmuştur. Tuz stresiyle beraber 4 mM askorbik asit uygulamasının artan NaCl
stresine bağlı MDA düzeyini arttırdığını ancak, MDA seviyelerinin askorbik asit
uygulanmayıp sadece tuz stresi uygulanan örneklere kıyasla daha düşük düzeyde olduğu
ifade etmişlerdir.
Wang vd. (2014) Kandelia candel köklerinde, antioksidan savunma sistemi üzerinde tuz
stresinin etkilerini araştırdıkları çalışmalarında, tuz konsantrasyonu ve işlem süresi
artması ile Kandelia candel kökü toplam protein içeriğinin azaldığını rapor etmişlerdir.
Ayrıca, CAT aktivitesi, SOD aktivitesi, ASA içeriği ve MDA içeriğinin, tuz
konsantrasyonu artışı ile azaldığını ve işlem süresi artışı ile iniş çıkışlar görülse de,
ASA ve MDA içeriğinin 6. saatte en üst düzeye ulaştığını ifade etmişlerdir. Belirli tuz
konsantrasyonunun (<300 mmol/L) Kandelia candel fide büyümesi için gerekli olduğu
sonucuna varmışlardır. Yüksek tuz stresinin erken aşamasında, Kandelia candel’in tuz
kaynaklı oksidadif strese karşı antioksidan savunma sistemini hızlı bir şekilde aktif hale
getirmesinin, plazma zarında oksidatif stresin zararını azaltmış, yüksek tuz stresine karşı
savunmada Kandelia candel için etkili bir yöntem olduğu sonucuna varmışlardır.
Miljuš-Djukić vd. (2013) üç bezelye türünde (Pisum arvense, P. sativum ve P. fulvum)
farklı konsantrasyonlarda tuz (75, 120, 150 ve 200 mM NaCl) stresinin antioksidan
savunma ve miRNA ifade değişimindeki etkisini araştırdıkları çalışmalarında, P.
arvense sürgünlerindeki, MDA seviyesi, 120 mM NaCl stresi altında önemli ölçüde (2,7

198
kat daha yüksek) artmıştır. P. sativum sürgünlerinde de 120 mM NaCI stresinde MDA
üretimi maksimum değere ulaşmış, ancak bu değer P. arvense ile karşılaştırıldığında
altında kalmıştır. Her iki genotipte de 150 ve 200 mM seviyesinde, MDA düzeyinin
kontrol grubu bitkilerinden daha düşük seviyeye inme sebebinin doku nekrozunu
düşündürdüğünü ifade etmişlerdir.
Çalışmamızda, 100 mM tuz (NaCl) stresi uygulamasında, 08-TR-003, TR-3080,
SARAY ve TARSAN 1018 ayçiçeği çeşitlerinde örnek alım zamanına bağlı olarak
MDA düzeyinde kademeli azalma görülmüştür. TARSAN 1018, 27. saat sonunda diğer
çeşitlerle karşılaştırıldığında MDA düzeyinde en fazla azalmanın gözlendiği çeşittir
(Şekil 4.14).
150 mM tuz (NaCl) stresi uygulaması, 08-TR-003, TR-3080 ve SARAY ayçiçeği
çeşitlerinde MDA düzeylerinde, örnek alım zamanına bağlı olarak kademeli azalmaya
sebep olurken, TARSAN 1018 çeşidinde, 3. saatte kontrole göre azalmaya, 27. saatte,
3. saate göre artışa sebep olmuştur (Şekil 4.15).
Çalışmada kullandığımız çeşitler üzerine bu konuda daha önce yapılmış bir çalışma
bulunmamaktadır. Literatürde yer alan diğer çalışmalara (Liang vd. 2003, Sergio vd.
2012, Mittova vd. 2004, Wang vd. 2014, Ruiz vd. 2005) göre değerlendirildiğinde, 100
mM ve 150 mM tuz stresi uygulaması sonucu MDA seviyesinde gözlenen düşüş,
bitkilerde stres toleransında, oksidadif strese karşı antioksidan savunma sisteminin hızlı
bir şekilde devreye sokularak, plazma zarında lipit peroksidasyonun önlendiğini ve tüm
çeşitlerin tuz stresine karşı toleranslı olduğunu gösterebilir.
Çevresel streslerde, MDA artışı, plasmalemma ve organel zarlarında lipid
peroksidasyonu veya hasar değerlendirilmesi için bir belirteç olarak kabul edilir. Lipit
peroksidasyonu, örneğin SOD, APX, GPX ve CAT artışı gibi antioksidan enzimlerin
aktivitesi ile ilişkilidir. Oksidatif stres toleransı gelişir ve MDA azaltılır. Soleimanzadeh
vd. (2010), potasyum düzeylerinin, kuraklık stresi altındaki ayçiçeğinde (Helianthus
annuus L.) antioksidan enzimler ve MDA içeriği üzerine etkisini araştırdıkları

199
çalışmada, bitkilerde MDA miktarının kuraklık stresi artışı ile arttığını, ancak potasyum
düzeylerinin artması ile azaldığını ortaya koymuşlardır. Deney verilerine göre, yüksek
kuraklık stresinde, MDA konsantrasyonu artışı, SOD ve GPX veya CAT’ın düşük
aktivitesine bağlı düşük potasyum düzeyleri, oksidatif stres hasarı için kritik bir
faktördür.
Vassilevska-Ivanova vd. (2014) ayçiçeği (Helianthus annuus L) genotiplerinde, PEG
kaynaklı su stresinin etkilerini araştırdıkları çalışmada, kullanılan her iki genotipte de
kuraklık stresinin PEG 6000 aracılığıyla, -0.4 ve -0.6 MPa ozmotik potansiyelinde
uygulanması MDA düzeyini önemli derecede etkilenmemiştir. Lipid peroksidasyon
seviyesinde aşamalı artış -8 MPa ozmotik potansiyelinde gözlenirken, istatistiksel stres
bağımlı artış -0.1 MPa’da gözlenmiştir.
Mirzaee vd. (2013) Brassica napus L. ile yaptıkları çalışmada, PEG uygulamasının,
kullanılan her iki çeşitte, kök ve sürgünlerde lipit peroksidasyon ürünü olan MDA
miktarını arttırdığı, Hyola 308 çeşidinde, MDA artış oranının SLM046 çeşidinden daha
yüksek olduğu bulunmuştur. Bunun yanında, kuraklık stresi SLM046 çeşidinin
köklerinde MDA içeriğinde önemli bir etkiye sebep olmadığı gözlenmiştir. SLM046
çeşidinin daha yüksek su stresi toleransı gösterdiği sonucuna varılmıştır.
Baloğlu vd. (2012), iki ayçiçeği (Helianthus annuus) çeşidinde (Aydın ve Musala) PEG
aracılığı ile kuraklık stresi altında antioksidan ve fizyolojik yanıtları araştırdıkları
çalışmalarında, artan kuraklık stresinin hücre zarlarında lipid peroksidasyonunun son
ürünleri olan MDA içeriklerinin her iki çeşitte de sürgünde önemli ölçüde arttığını
gözlemlemişlerdir. Sert kuraklık stresi koşullarının Aydın çeşidinin sürgünlerinde, daha
propagatif etkiye sahip olduğunu, her iki çeşitte de köklerde MDA seviyesinin kuraklık
stresinden etkilenmediğini ortaya koymuşlardır. Köklerde MDA düzeyinin
etkilenmemesi, düşük lipid peroksidasyona karşılık geldiği ve bu dokuların oksidatif
hasara karşı daha iyi koruma gösterebilecekleri sonucuna varılmıştır.

200
100 mM kuraklık (PEG) stresi uygulaması, 08-TR-003, TR-3080 ve SARAY ayçiçeği
çeşitlerinde MDA düzeylerinde uzayan stres uygulama zamanıyla beraber azalmalara
sebep olurken, TARSAN 1018 çeşidinde 3. saatte kontrole göre azalmaya, 27. saatte 3.
saate göre artışa sebep olmuştur (Şekil 4.16).
150 mM kuraklık (PEG) stresi uygulaması, tüm çeşitlerde stres uygulama zamanında
artışla beraber, MDA düzeyinde azalmaya sebep olmuştur (Şekil 4.17).
Çalışmada, 100 mM ve 150 mM PEG stresi uygulaması sonucu MDA seviyesinde tuz
stresiyle benzer sonuçlar elde edilmiş, stres tolerans mekanizmaları sayesinde kuraklık
stresinin de bitkilerde plazma zarları üzerine olumsuz etki göstermesi önlenmiştir.
Sonuçlar, ayçiçeği çeşitlerinin, tuz stresinin yanı sıra kuraklık stresine karşı da toleranslı
olduğunu göstermektedir.
Rai vd. (2014), Catharanthus roseus’un Rosea ve Alba çeşitleri ile yaptıkları çalışmada,
30 gün süresince farklı konsantrasyonlarda (0, 10, 50 ve 100 μM) Cr stresi uygulaması
sonucunda, artan konsantrasyona bağlı MDA düzeyinde artış gözlemlemişlerdir. Rosea
çeşidinde artan konsantrasyona bağlı protein miktarında kademeli azalma görülmüştür.
Alba çeşidinde ise en belirgin azalma 100 μM Cr stresi uygulamasında gözlenmiştir.
Maiti vd. (2012), mısır bitkisinde Cr stresi yanıtını araştırdıkları çalışmada, Sartaj ve
Deccan çeşitlerine, farklı konsantrasyonlarda (0, 50, 100, 200 ve 300 μM) Cr stresi
uygulamış ve konsantrasyondaki artışla beraber MDA düzeyinde de artış olduğunu
gözlemlemişlerdir. En belirgin artışın her iki çeşitte de 300 μM Cr stresi uygulamasında
olduğunu ifade etmişlerdir.
Sinha vd. (2005), Pistia stratiotes L. bitkisinde Cr kaynaklı lipid peroksidasyonun,
antioksidanlar ve antioksidan enzimler üzerine rolünü araştırdıkları çalışmada, metal
konsantrasyonun ve maruz kalma süresindeki artışla beraber hem kök, hem de
yapraklarda, MDA içeriğinde bir artış gözlendiğini bildirmişlerdir. 10μM Cr stresinin
48 saat uygulamasında görülen artış dışında, tüm uygulama sürelerinde, Cr
konsantrasyonlarında artışla, total protein miktarında azalma görülmüştür.

201
Shanker ve Pathmanabhan (2004) yaptıkları çalışmada, farklı konsantrasyonlarda (50
ve 100 mM), Cr(III) ve Cr(VI) stresi uygulaması sonucu, sorgum kök ve yapraklarında
malondialdehit (MDA) değişimini incelemişlerdir. Yaprak ve kökte bulunan MDA
düzeyleri karşılaştırıldığında köklerdeki miktarın daha fazla olduğunu
gözlemlemişlerdir. Hem Cr(III) hem de Cr(VI)’nın artan konsantrasyonlarına bağlı
MDA düzeyinde de artış görülmüştür. Cr(VI), Cr(III) ile kıyaslandığında Cr(VI)’nın
MDA düzeylerinde daha fazla artışa sebep olduğu bildirilmiştir.
Rai vd. (2004), Ocimum tenuiflorum L. ile yaptıkları çalışmada, farklı
konsantrasyonlardaki (10, 20, 50 ve 100 μM) Cr stresi, farklı sürelerde (24, 48 ve 72
saat) uygulanmıştır. Genel olarak artan Cr konsantrasyonu ve sürelere bağlı olarak
MDA düzeyinde artış, protein miktarında azalma gözlenmiştir. Cr birikimi, MDA
üretimi vasıtasıyla, serbest radikallerin aşırı üretimini teşvik eder. Ayrıca membran
lipitlerinin peroksidasyonu, membran bütünlüğünün bozulduğunu gösteren, potasyum
sızıntısıyla sonuçlanır. Yüksek MDA üretimi ile birlikte zar geçirgenliğinin krom
kaynaklı kaybı Vallisneria spiralis’da da gözlenmiştir (Vajpayee vd. 2001).
Çalışmamızda, Cr(VI) stresi uygulamasının tüm çeşitlerde 80 ppm uygulamada,
kontrole göre MDA düzeyinde azalmaya sebep olduğu, farklı Cr konsantrasyonlarında
(80ppm, 160 ppm, 320 ppm, 640 ppm, 1280 ppm) MDA düzeyinde artış azalışlar
görülse de genel olarak artan Cr(VI) konsantrasyonuna bağlı olarak MDA düzeyinde
artışın olduğu görülmektedir (Şekil 4.18). 80 ppm Cr uygulamasının, tüm çeşitlerde
antioksidan savunma sistemlerini aktive ettiği, bunun üzerindeki uygulamalarda MDA
seviyesinde gözlenen artışın stres karşısında savunma sistemlerinin yetersiz kaldığını ve
lipit peroksidasyonun gerçekleştiğini göstermektedir. Artan Cr konsantrasyonlarıyla
total protein miktarlarında görülen azalma da bu sonuçları destekler niteliktedir.
5.3 E2F Geni İfade Seviyesinin Belirlenmesi
Hücre döngüsünü düzenleyen yolağın ikinci bileşeni, E2F’ler olarak adlandırılan
transkripsiyon faktörleri ailesidir. E2F’lerin DP heterodimerizasyon domaini içermesi,

202
DP proteinleri ile birleşmeyi sağlar ve E2F-DP transkripsiyon kompleksinin
aktivasyonuyla sonuçlanır. Bu kompleks, hücre döngüsü ilerlemesinde rol oynayan
genlerin ekspresyonunu uyarır (Desvoyes vd. 2006). E2F’ler ayrıca, transkripsiyonel
aktiviteyi bloke etmek için Rb proteininin bağlandığı bağlanma alanları içerir.
Arabidopsis thaliana’da altı E2F (E2Fa, E2Fb, E2Fc, DEL1, DEL2 ve DEL3), iki DP
proteini (DPa ve DPb) ve bir tane Rb homoloğu (RBR1) tanımlanmıştır (Desvoyes vd.
2006, Hirano vd. 2008).
S fazındaki spesifik genlerin ekspresyonunu kontrol eden E2F transkripsiyon faktörleri
G1 evresindeki genleri susturmak için RBR ile birleşir. Geç G1 evresinde,
CYCD/CDKA, RBR’yi fosforile eder, E2F’yi serbest bırakır ve S-fazı genlerini aktive
eder (De Veylder vd. 2007).
Vandepoele vd. (2005) yaptıkları çalışmada, E2Fa ve DPa transkripsiyon faktörlerinin
Arabidopsis thaliana fidelerinde ektopik ifade genlerinin transkripsiyonel düzeyde
promotor bölgelerini uyardığı tespit etmişlerdir. E2F hedef genlerinin G1 ve S fazları
boyunca özel olarak ifade edildiğini ve hareketsiz hücrelerin yeniden aktive edilerek
hücre döngüsüne girdiğini göstermişlerdir.
.
AtE2Fa ve AtDPa aşırı üretimi ekstra postembriyonik hücre bölünmesini uyarır. Hücre
döngüsünden çıkışın gecikmesindeki farklılaşma ekstra hücre bölünmeleri gözlenmesini
açıklayabilir (De Veylder vd. 2002). AtE2Fa ve AtDPa’nın aşırı ekspresyonu ayrıca S
fazına yeniden girmek için farklılaşmış yaprak hücrelerinin üretimine sebep olur
(Rossignol vd. 2002).
E2Fa ve E2Fb; DNA replikasyonu, DNA tamiri ve kromatin korunması için gerekli
genleri de içeren, bunların muhtemel hedef genlerinde G1’den S fazına geçişi teşvik
eden transkripsiyonel aktivatörler olarak görev alırlar (Ramirez-Parra vd. 2003, Vlieghe
vd. 2003, Vandepoele vd. 2005, Naouar vd. 2009).

203
5.3.1 100 mM ve 150 mM NaCl stresi ile E2F seviyesinde meydana gelen değişim
100 mM NaCl stresi uygulanmış ayçiçeği çeşitlerinde zamana bağlı E2F gen ifade
düzeylerindeki değişim kontrole göre incelendiğinde farklılıklar oluştuğu gözlenmiştir.
08-TR-003 ve TR-3080 çeşitlerinde 3. saatte, kontrole göre E2F gen ifade düzeyinde
artış olduğu, 27. saatte, 3. saate göre azalma olduğu, ancak ifade düzeyinin kontrolle
kıyaslandığında, kontrolde gözlenen ifade düzeyinin üzerinde olduğu görülmektedir.
E2F geni ifade düzeyinin en fazla olduğu 3. saatteki gen ifade seviyeleri iki çeşit
arasında kıyaslandığında TR-3080 çeşidinde, 08-TR-003 çeşidine göre yaklaşık 3 kat
fazla olduğu görülmektedir (Şekil 4.33).
08-TR-003 ve TR-3080 çeşitlerinde, değişen E2F ifade seviyeleri değerlendirildiğinde
kısa süreli 100 mM NaCl stresi uygulamasının hücre döngüsü ilerlemesini uyardığı,
süre uzadığı durumda ilerlemenin baskılandığı söylenebilir. Stresin uygulandığı her iki
zamanda, E2F ifade seviyesinin kontrolde gözlenen seviyenin üzerinde olması,
çeşitlerin tuz toleransına sahip olduğunun göstergesi olabilir.
100 mM NaCl stresi uygulanmış, SARAY ve TARSAN 1018 çeşitlerinde E2F gen ifade
düzeyinde zamana bağlı artış olduğu gözlenmiştir. E2F gen ifade düzeyinde en fazla
artışın görüldüğü 27. saatteki ifade düzeylerin SARAY ve TARSAN 1018 çeşitleri
arasında kıyaslandığında, TARSAN 1018 çeşidinde kontrole göre artışın daha fazla
(yaklaşık 33 kat) olduğu görülmektedir (Şekil 4.33).
100 mM tuz stresi uygulanmış tüm çeşitlerde, kontrolle karşılaştırıldığında, zamana
bağlı E2F gen ifade seviyelerindeki değişim istatistiksel olarak p<0.05 düzeyinde
anlamlıdır.
SARAY ve TARSAN 1018 çeşitlerinde, 100mM NaCl stresinin uygulanma süreleriyle
paralel artış gösteren E2F ifade seviyesinin, G1’den S fazına geçişi teşvik ettiği
söylenebilir. TARSAN 1018 çeşidinde 27. saatte gözlenen artışın SARAY çeşidinden

204
fazla olması, tuz stresine karşı SARAY çeşidinden daha dayanıklı olduğunu
gösterebilir.
150 mM tuz stresi uygulanmış TR-3080, SARAY ve TARSAN 1018 ayçiçeği
çeşitlerinde, E2F gen ifade seviyelerinde zamana bağlı artış olduğu gözlenmiştir. 27.
saatte bu artışın üst düzeyde olduğu ve TR-3080 çeşidinde diğer çeşitlerle
kıyaslandığında kontrole göre artışın en fazla ( yaklaşık 858 kat) olduğu gözlenmiştir.
TARSAN 1018 çeşidinde 27. saatteki gen ifade seviyesinin, kontrole göre 200 kat,
SARAY çeşidinde ise yaklaşık 6 kat arttığı görülmektedir (Şekil 4.36).
150 mM tuz stresi uygulanmış TR-3080, SARAY ve TARSAN 1018 ayçiçeği
çeşitlerinde, kontrolle karşılaştırıldığında, zamana bağlı E2F gen ifade seviyelerindeki
değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı olduğu görülmektedir.
150 mM tuz stresi uygulanmış 08-TR-003 çeşidinde ise E2F geni ifade düzeyi en
yüksek değerine 3. saatte ulaşmıştır. 3. saatteki ifade düzeyinde kontrolle
kıyaslandığında yaklaşık 23 kat artış olduğu gözlenmiş, 27. saatte ifade düzeyi, kontrol
seviyesine gerilemiştir. İfade düzeylerindeki bu değişim istatistiksel olarak
değerlendirildiğinde, kontrole göre 3. saatte gözlenen değişimin p<0.05 düzeyinde
anlamlıdır. 27. saatteki ifade düzeyindeki değişim, 3. saate göre anlamlı, kontrole göre
anlamsızdır (Çizelge 4.36).
150 mM NaCl stresi uygulaması sonucu, 27. saatte E2F gen ifade seviyesinde gözlenen
bu artışın sebebi, bitkilerde stres yanıt mekanizmalarının aktifleştirildiği ve bu stresin
hücre döngüsü ilerlemesi üzerine olumsuz etki oluşturmasının engellendiği şeklinde
açıklanabilir. TR-3080 ve TARSAN 1018 çeşitlerinde gözlenen ifade seviyesinin, diğer
çeşitlerin üstünde olması, bu çeşitlerin tuz sresine karşı diğerlerinden daha toleraslı
olduğunu gösterebilir.

205
5.3.2 100 mM ve 150 mM PEG stresi ile E2F seviyesinde meydana gelen değişim
100 mM PEG stresi uygulanmış ayçiçeği çeşitlerinde E2F gen ifade düzeyi kontrolle
kıyaslandığında 3. saatte en yüksek seviyeye ulaşmış, 27. saatte ise ifade seviyesi
gerilemiştir. TR-3080 çeşidinde 3. saatteki E2F ifade düzeyinde kontrole göre artışın
diğer çeşitlerle kıyaslandığında en yüksek (yaklaşık 149 kat) olduğu görülmektedir. 3.
saatteki ifade düzeyinde kontrole göre olan bu artış SARAY çeşidinde yaklaşık 107 kat,
TARSAN 1018 çeşidinde yaklaşık 83 kat olarak gözlenmiştir. 08-TR-003 çeşidinde ise
3. saatte kontrole göre artışın yaklaşık %50 olduğu görülmektedir.
100 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiş ve meydana gelen değişikliğin p<0,05 düzeyinde
istatistiksel olarak anlamlı olduğunu görülmüştür (Çizelge 4.65, 4.67, 4.69, 4.71).
100 mM PEG stresi uygulanan 08-TR-003, TR-3080 ve TARSAN 1018 çeşitlerinde tek
yönlü varyans analizi (One Way ANOVA Dunnet Test, Post Hoc testi) sonuçlarına göre
3. ve 27. saatlerde E2F düzeyindeki değişimin istatistiksel olarak p<0.05 düzeyinde
anlamlı olduğu görülmektedir (Çizelge 4.66, 4.68, 4.72). SARAY çeşidinde ise 3. saatte
kontrole göre gözlenen değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı olduğu,
27. saatte kontrole göre olan değişimin p>0,05 düzeyinde anlamsız olduğu
görülmektedir (Çizelge 4.70).
150 mM PEG stresi uygulanmış ayçiçeği çeşitlerinde E2F gen ifade düzeyinin kontrolle
kıyaslandığında 3. saatte en yüksek seviyeye ulaştığı, 27. saatte ise ifade seviyesinde 3.
saatte göre azalma olduğu görülmektedir. Çeşitler arasında 46,866 değeriyle 3. saatte
kontrole göre en fazla artış TARSAN 1018’de gözlenmiştir. 3. saatte, 215,34 değeriyle
SARAY, 18,506 değeriyle TR-3080 ve 3,952 değeriyle 08-TR-003 çeşidinde kontrole
göre artış gözlenmiştir. 08-TR-003 ve TR-3080 çeşitlerinde 27 saat süreyle 150 mM
PEG stresi uygulaması, E2F gen ifade seviyelerinin kontrolün de altına inmesine sebep
olmuştur (Şekil 4.42).

206
150 mM PEG stresi uygulanan örneklerin zamana karşı farklılaşan E2F gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiş ve meydana gelen değişikliğin p<0,05 düzeyinde
istatistiksel olarak anlamlı olduğunu görülmüştür (Çizelge 4.95, 4.97, 4.99, 4.101).
150 mM PEG stresi uygulanan tüm ayçiçeği çeşitlerinde tek yönlü varyans analizi (One
Way ANOVA Dunnet Test, Post Hoc testi) sonuçlarına göre 3. ve 27. saatlerde E2F
düzeyindeki değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı olduğu
görülmektedir (Çizelge 4.96, 4.98, 4.100, 4.102).
100 mM ve 150 mM PEG stresi uygulaması sonucu, E2F ifade seviyesinde anlamlı
artış 3. saatte gözlenmiştir. Kuraklık stresinin kısa süreli uygulanmasının hücre döngü
ilerleyişini aktive ettiği, uzayan sürelerde E2F seviyesinde gözlenen azalma nedeniyle
baskılandığı söylenebilir. 08-TR-003 çeşidinde 3. saatte gözlenen E2F seviyesinin diğer
çeşitlerin çok altında kalması, kuraklık stresine karşı daha duyarlı olduğunun göstergesi
olabilir.
5.3.2 Cr(VI) stresi ile E2F gen ifade seviyesinde meydana gelen değişim
Cr(VI) stresi uygulamasının çalışılan çeşitlerde artan Cr konsantrasyonuna bağlı E2F
gen ifade düzeylerinde birbirlerinden farklı değişimlere sebep olduğu gözlenmiştir. 08-
TR-003 çeşidinde 80ppm ve 160 ppm Cr(VI) uygulamasının E2F gen ifade düzeyinde
kontrole göre kademeli artışa sebep olduğu görülmektedir. 320 ppm ve 640 ppm’de,
160 ppm’ deki seviyesine göre kademeli azalma olduğu, 1280 ppm’de tekrar artma
eğilimine geçtiği görülmektedir.
TR-3080 ayçiçeği çeşidinde, E2F gen ifade düzeyinde 80 ppm Cr(VI) uygulamasının
kontrole göre bir miktar artışa, 160 ppm Cr(VI) uygulamasının, 80 ppm’e göre
azalmaya sebep olduğu görülmektedir. 160 ppm’den itibaren 1280 ppm’e kadar artan
Cr konsantrasyonuna paralel olarak artışa sebep olduğu görülmektedir. 1280 ppm’de

207
ifade seviyesi kontrole göre yaklaşık olarak 51 kat artmış ve en yüksek seviyeye
ulaşmıştır.
SARAY ayçiçeği çeşidinde E2F gen ifade düzeyinde, 80 ppm ve 160 ppm Cr(VI)
uygulamasının, kontrole göre önemli bir değişime sebep olmadığı, 320 ppm ve 1280
ppm Cr(VI) uygulamasının ise kontrole göre azalmaya sebep olduğu görülmektedir.
640 ppm Cr(VI) uygulaması E2F gen ifade düzeyini yaklaşık olarak 6 kat arttırmış ve
en yüksek seviyeye ulaştırmıştır.
TARSAN 1018 ayçiçeği çeşidinde 80 ppm, 160 ppm ve 320 ppm Cr(VI) uygulaması,
E2F gen ifade düzeyinde kontrole göre artışa sebep olurken, 640ppm ve 1280 ppm
Cr(VI) uygulaması gen ifade düzeyinde kontrole göre önemli bir değişime sebep
olmamıştır. En yüksek E2F gen ifade düzeyi sırasıyla 160 ppm ve 320 ppm Cr(VI)
uygulamasında gözlenmiştir.
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan örneklerin artan Cr(VI)
konsantrasyonuna karşı farklılaşan E2F gen ifade seviyelerinin One Way ANOVA
(Dunnet) metodu ile p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu
görülmektedir (Çizelge 4.128, 4.129, 4.132, 4.133).
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan 08-TR-003, TR-3080 ve SARAY
çeşitlerinde tek yönlü varyans analizi (One Way ANOVA Dunnet Test, Post Hoc testi)
ile konsantrasyona karşı E2F geni ekspresyon seviyesinde meydana gelen değişimin
istatistiksel olarak p<0.05 düzeyinde anlamlı olduğu görülmektedir (Çizelge 4.130,
4.131, 4.134). TARSAN 1018 çeşidinde, 640 ppm Cr(VI) uygulamasının kontrole göre
E2F gen ifade düzeyinde meydana getirdiği değişim istatistiki olarak p>0,05 düzeyinde
anlamsız olup, diğer konsantrasyonlarda kontrole göre meydana gelen değişimler
p<0.05 düzeyinde anlamlıdır (Çizelge 4.135).
Cr(VI) stresi uygulaması sonucu, E2F geni ifade düzeylerinde gözlenen değişimler,
E2F’nin hücre döngüsündeki rolü doğrultusunda değerlendirildiğinde, özellikle SARAY
çeşidine 640 ppm ve TR-3080 çeşidine 1280 ppm Cr(VI) uygulamasında gen ifade

208
düzeyinde görülen artış anlamlı olmayıp, bu çeşitlerde Cr stresinin hücrede farklı
düzenlenmelere sebep olabileceğini düşündürmektedir. 08-TR-003 ve TARSAN 1018
çeşitlerinde Cr konsantrasyonundaki artışla beraber E2F ifade seviyesinin önce artma,
daha sonra azalma eğiliminde olması, bitki savunma mekanizmaları sayesinde bu
çeşitlerin belli konsantrasyona kadar Cr stresini tolere edebildiği, ancak konsantrasyon
artışıyla beraber stres etkilerinin üstesinden gelemeyip hücrelerin apoptozise gittiği
söylenebilir.
5.4 RBR Geni İfade Seviyesinin Belirlenmesi
Retinoblastoma ilişkili (RBR) proteinler, insan tümör baskılayıcı pRB’nin bitki
homologlarıdır. Hayvan karşıtlarıyla benzer olarak hücre döngüsü düzenlenmesinde ve
farklılaşmasında rol oynamaktadırlar. pRB’nin bitki homologları olan RBR ilk olarak
mısırda (Zea mays) keşfedilmiştir (Grafi vd. 1996).
Çok hücreli organizmaların gelişimi hücreler arası, hücre içi ve özellikle bitkilerde dış
çevresel sinyaller ile koordine edilmektedir. Herhangi bir sinyal gelişimi etkilese de, tek
hücre her zaman hücre döngüsünde ilerlemek zorundadır. Hücre döngüsünde diğer
aşamaya geçme ya da geri dönüşümlü ya da dönüşümsüz döngüden çıkma kararını
vermek zorundadır. pRB, genellikle hücre döngüsü girişinin kontrol edilmesi ve hücre
döngüsü ilerlemesinin sonraki kontrol noktalarıyla ilişkili bulunmuştur. Bir kontrol
noktası, hücre boyutu, DNA replikasyonu ve kromozom ayrımı gibi önceki olayların
tamamlandığı durumda işlemin başlatılmasını sağlar (Elledge vd. 1996).
pRB’nin bugün bilinen fonksiyonları üç gup altında toplanabilir:
(i) hücre döngüsü ilerlemesinin düzenlenmesi,
(ii) kromatin-düzenleyiciler aracılığıyla transkripsiyonun düzenlenmesi,
(iii) hücre kaderinin kararları için gerekli olan transkripsiyon faktörleri ile
etkileşimi sonucu farklılaşmaya teşvik etmesi.

209
Hücre döngüsü ilerlemesinin düzenlenmesinin klasik örneği E2F ve DP transkripsiyon
faktörleri ile pRB’nin etkileşimidir. E2F ve DP’nin oluşturduğu heterodimer, kendi
promotorlarındaki, spesifik E2F bağlanma sekansları ile genlerin ekspresyonunu aktive
eder. Genlerin bir çoğu, S-fazı sırasında gerekli olan DNA replikasyon düzeneğindeki
proteinler gibi proteinleri kodlar. Hipofosforile pRB’nin E2F’ye bağlanması, onun
aktivitesini inhibe eder. Mitojenik uyaranlar üzerine, siklin / CDK kompleksleri, pRB’yi
fosforile eder. Bu E2F/DP kompleksinin ayrılmasına ve ardından gen ekspresyonunun
aktivasyonunu sağlar.
Kromatin-düzenleyiciler aracılığıyla transkripsiyonun düzenlenmesinde, pRB,
memelilerde PRC2 (Polycomb Repressive Complex 2) kompleksinin alt birimi olan
RbAp48 ile etkileşime girer. RBR’nin, RbAp48’in bitki homologu olan MSI1 ile
etkileşimi ispat edilmesine (Ach vd. 1997), tanımlama için gerekli görülmesine (Jullien
vd. 2008) rağmen, pRB veya RBR’nin PRC2 aktivitesine doğrudan katkısını gösteren
örnekler şaşırtıcı olarak azdır. Kromatin birleşmesi ve histon deasetilasyon
komplekslerinde varlığı domates ve Arabidopsis’te RbAp48’in bitki homoloğunun
varlığı ve Rb’ye bağlandığı tanımlanmıştır (Ach vd. 1997).
Hipo asetillenmiş histonlar, inaktif kromatin ile ilişkili olmasından dolayı, bitki Rb
homologları, kromatin değişikliklerini yönlendirerek E2F regüle genlerin
ekspresyonunu olumsuz yönde düzenlerler. Bitkilerde çimlenme sırasında H3K27me3
aracılığıyla RBR tarafından bir gelişim programının baskılanması, heterotrofikten,
ototrofik fide büyümesine geçiş için gereklidir (Gutzat vd. 2011).
Memelilerde pRB’nin hücre kaderinin belirleyen faktörler içinde yer alan proteinlerle
etkileşimi gösterilse de bitkisel RBR’nin bu proteinlerle doğrudan etkileşimi henüz
rapor edilmemiştir. Farklı proteinlerle etkileşimini içeren pRB ve RBR’nin çoklu
fonksiyonları evrimsel süreçte korunmuştur. Bitkilerde RBR’nin ortaya çıkan
fonksiyonlarını tanımlamak, yüksek bitkilerde kök hücre nişi ve gelişimsel programların
düzenlenmesi için tek hücreli alg hücre boyutu kontrolüne uzanan evrimsel bakış açısı
sağlar.

210
Mısır endosperminde, RNAi tarafından RBR1’in aşağı-regülasyonu, hücre ölümünü
arttırır ve hem mitotik, hem de endoreduplikasyon hücre döngülerini uyarır. Ayrıca
depo protein içeriği ve çekirdek ağırlığı etkilenmemesine rağmen, DNA içeriği,
RBR1’in aşağı regülasyonu ile %43 artar (Sabelli vd. 2013). Endoreduplikasyonda,
mitoz olmadan, hücrede kromozomal DNA’nın replikasyonu, yüksek kromozom
sayıları seviyesiyle sonuçlanır (Lee vd. 2009).
Kök apikal meristem içinde RBR’nin lokal azalması, kök hücre sayısınının artmasına
neden olur (Wildwater vd. 2005).
Gutzat vd. (2011), Arabidopsis thaliana ile yaptıkları çalışmada, RBR’nin sükrozla ters
yönde hareket ettiği, embriyonik genleri baskıladığı ve hücre döngüsünü negatif yönde
düzenlediği sonucuna varmışlardır.
Borghi vd. (2010) retinoblastomanın Arabidopsis thaliana’da hücre döngüsünün anahtar
düzenleyicisi olduğu ve hücre döngüsü, proliferasyonu ve gelişiminde önemli roller
üstlendiğini ifade etmişlerdir.
Park vd. (2005) Nicotiana benthamiana’da, Rb’nın hücre döngüsündeki düzenleme
rolünü virüs indüklü gen susturma ile inhibe ederek açıklamışlardır. Gen susturma
sonucu yaprak ve gövde hücrelerinde farklılaşmanın geciktiği, hücre proliferasyonunun
uzadığı görülmüştür. Çiçek oluşumunda da gecikmeler gözlenmiştir.
RBR / E2F yolu, çoğalan ve farklılaşmış hücrelerin her ikisinde de gen ekspresyonunun
düzenlenmesinde önemli bir rol oynar. RBR, E2F düzenleyici genleri, E2F
transaktivasyon domeinini maskeleyerek ve / veya histon deasetilaz ve kromatin
yeniden yapılanma faktörlerini işe karıştırarak baskılar. Mısır Rpd3 histon deasetilaz,
transkripsiyonu baskılamak için, RbAp1 / RbAp48 / MSI1 kromatin birleştirme faktörü
(CAF-1) ‘in bir bileşenin yardımıyla, RBR ile etkileşime girer (Rossi vd. 2003).

211
5.4.1 100 mM ve 150 mM NaCl stresi ile RBR seviyesinde meydana gelen değişim
100 mM NaCl stresi uygulamasının zamana bağlı RBR gen ifade düzeyleri değişiminde
ayçiçeği çeşitleri arasında farklılıklar olduğu gözlenmiştir. 08-TR-003, TR-3080 ve
SARAY ayçiçeği çeşitlerinde 3. ve 27. saatlerde gözlenen RBR gen ifade düzeyinin
kontrole göre azaldığı görülmektedir.
08-TR-003 çeşidinde örnek alım zamanındaki uzamayla beraber gen ifade düzeyi
kademeli azalmıştır. TR-3080 çeşidinde 3. saat ve 27. saat süresince 100 mM NaCl
stresi uygulamasının RBR gen ifade düzeyinde önemli bir değişime sebep olmadığı
görülmektedir. SARAY çeşidinde gen ifade düzeyindeki en düşük değerin 3. saatte
olduğu görülürken, 27. saatte kontrol seviyesinin çok altında kalsa da 3. saatle
kıyaslandığında bir miktar artış olduğu görülmektedir. TARSAN 1018 çeşidinde diğer
çeşitlerden farklı olarak RBR gen ifade düzeyi kontrole göre yaklaşık 7 kat artarak 3.
saatte en yüksek düzeyine ulaşmıştır. 27. saatte ise 3. saatle karşılaştırıldığında azalma
görülse de kontrol değerinin üzerindedir (Şekil 4.34).
100 mM tuz stresi uygulanmış ayçiçeği çeşitlerinde, kontrolle karşılaştırıldığında,
zamana bağlı RBR gen ifade seviyelerindeki değişimin istatistiksel olarak p<0.05
düzeyinde anlamlı olduğu görülmektedir.
100 mM tuz stresi uygulamasının, TR-3080 ayçiçeği çeşidinde RBR gen ifade
düzeyinde 3. ve 27. saatlerde kontrole göre olan değişimin istatistiksel olarak p<0.05
düzeyinde anlamlı olduğu, 27. saatteki seviyenin 3. saate göre değişimi ise istatistiki
olarak p>0,05 düzeyinde anlamsızdır (Çizelge 4.18).
08-TR-003, TR-3080 ve SARAY çeşitlerinde 100 mM NaCl stresi uygulamasının RBR
geni ifade seviyesini kontrole göre azaltması sebebiyle, ortamda belli miktarda tuzun
bulunmasının hücre döngüsü ilerleme sürecini aktive ettiği söylenebilir. TARSAN
1018 çeşidinde gen ifade düzeyinde 3. saatte gözlenen artış tuz stresinden bir miktar
etkilendiğini gösterse de, 27. saatte RBR seviyesinin düşmesi stres tolerans

212
mekanizmalarının devreye sokulduğunu ve sürecin ilerleyişinin sağlandığını
gösterebilir.
150 mM NaCl stresi uygulaması TR-3080 ve TARSAN 1018 çeşitlerinde RBR gen
ifade düzeylerinde 3. ve 27. saatlerde kontrolle karşılaştırıldığında zamana bağlı artış
olduğu gözlenmiştir. Her iki çeşitte de en yüksek ifade düzeyi 27. saatte gözlenirken,
TR-3080 çeşidinde kontrolle kıyaslandığında artışın daha fazla olduğu görülmektedir.
SARAY çeşidinde ise zamana bağlı azalma görülmüştür. 08-TR-003 çeşidinde en
yüksek RBR gen ifade düzeyi kontrole göre yaklaşık 3 kat artışla beraber 3. saatte
gözlenmiştir. 27. saatte ise hem kontrole, hem de 3. saate göre azalma görülmektedir
(Şekil 4.37).
150 mM tuz stresi uygulanmış ayçiçeği çeşitlerinde, kontrole göre zamana bağlı RBR
gen ifade seviyelerindeki değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı
olduğu görülmektedir.
150 mM NaCl uygulaması sonucu, RBR gen ifade seviyelerindeki değişim
değerlendirildiğinde, TR-3080 ve TARSAN 1018 çeşitlerinde, stresin bu çeşitler
üzerinde etkili olduğunu ve hücre döngüsünün baskılanmasına sebep olduğunu, 08-TR-
003 çeşidinin stres tolerans mekanizmalarının aktivasyonu ile tuz stresinin etkilerini
ortadan kaldırdığını, SARAY çeşidinin tuza toleransının yüksek olduğunu gösterebilir.
5.4.2 100 mM ve 150 mM PEG stresi ile RBR seviyesinde meydana gelen değişim
100 mM PEG stresi uygulaması sonucu ayçiçeği çeşitlerinde RBR gen ifade seviyesinde
birbirinden farklı değişimler ortaya çıkmıştır. 08-TR-003 çeşidinde 3. saatte, ifade
düzeyinde kontrole göre bir azalma görülürken, 27. saatte, 3. saate göre bir artış
görülmüş, ancak ifade seviyesi kontrolün altında kalmıştır.

213
TARSAN 1018 çeşidinde ise aynı şekilde 3. saatte kontrole göre azalma görülmüş, 27.
saatte ifade düzeyinde bir artış görülmüş ve 08-TR-003 çeşidinden farklı olarak kontrol
seviyesinin üzerine çıkmıştır.
TR-3080 ve SARAY çeşitlerinde RBR gen ifade seviyeleri bu stres koşullarına birbirine
zıt şekilde tepki vermişlerdir. TR-3080 çeşidinde zamana bağlı gen ifade seviyesinde
artış gözlenirken, SARAY çeşidinde zamana bağlı azalma görülmüştür (Şekil 4.40).
100 mM PEG stresi uygulanmış ayçiçeği çeşitlerinde, kontrole göre zamana bağlı RBR
gen ifade seviyelerindeki değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı
olduğu görülmektedir.
100 mM PEG stresi uygulaması, TR-3080 ve TARSAN 1018 ayçiçeği çeşitlerinde RBR
gen ifadesinde kontrole göre gözlenen artışla beraber hücre döngüsü ilerleyişinin
baskılanmış olabileceği söylenebilir. 08-TR-003 ve SARAY çeşitlerinde ise stres
uygulaması hücre döngü ilerleyişini aktive etmiş olabilir.
150 mM PEG stresi uygulaması sonucu 08-TR-003 ve TR-3080 çeşitlerinde RBR gen
ifade düzeyinde zamana bağlı azalma görülmüştür. SARAY ve TARSAN 1018
çeşitlerinde ise 3. saatte kontrole göre bir artış görülmüş, 27. saatte kontrol seviyesinin
bile altına indiği gözlenmiştir. TARSAN 1018 çeşidinde 3. saatteki artışın daha belirgin
olduğu (kontrole göre yaklaşık 85 kat) görülmektedir (Şekil 4.43).
150 mM PEG stresi uygulanmış tüm çeşitlerde, kontrolle karşılaştırıldığında, zamana
bağlı RBR gen ifade seviyelerindeki değişim istatistiksel olarak p<0.05 düzeyinde
anlamlıdır.
150 mM PEG uygulaması 08-TR-003 ve TR-3080 çeşitlerinde hücre bölünmesinin
artışına sebep olmuş olabilir. SARAY ve TARSAN 1018 çeşitlerinde ise hücre
bölünmesi kısa süreli stres uygulanmasında baskılansa da, bitki savunma
mekanizmalarının devreye sokularak yeniden hücre döngüsünün aktivasyonu sağlanmış
olabilir.

214
5.4.3 Cr (VI) stresi ile RBR seviyesinde meydana gelen değişim
Cr(VI) stresi uygulamasında ayçiçeği çeşitlerinde farklı konsantrasyonların RBR gen
ifade düzeylerinde artışa sebep olduğu görülmektedir. 08-TR-003 çeşidinde, 80 ppm
Cr(VI) uygulaması, kontrole göre RBR gen ifade düzeyinde düşüşe sebep olmuş, 160
ppm ve 320 ppm konsantrasyonlarında uygulama ise örneklerde gen ifade düzeyini
arttırmıştır. 320 ppm Cr(VI) konsantrasyonunda en yüksek ifade düzeyi gözlenmiştir.
640 ppm ve 1280 ppm uygulamada ise ifade seviyesi düşerek, kontrolün bile altında
inmiştir.
TR-3080 çeşidinde, kontrolden 320 ppm’e kadar Cr(VI) uygulaması RBR gen ifade
düzeyinde önemli bir değişime sebep olmazken, 640 ppm uygulama ifade düzeyini
oldukça arttırmış ve en yüksek düzeye ulaşmasına sebep olmuştur. 1280 ppm
uygulamada ifade seviyesi gerilemiştir.
SARAY çeşidinde 80 ppm Cr(VI) uygulaması, RBR gen ifade düzeyinin kontrole göre
artışına sebep olmuş, bunun üzerindeki konsantrasyonlardaki uygulamalarda ifade
düzeyinde dalgalanmalar görülse de 80 ppm Cr(VI) stresi uygulanan örnekler ve
kontrole kıyasla ifade düzeyi çok altında kalmıştır.
TARSAN 1018 çeşidinde 80 ppm Cr (VI) uygulaması, ifade düzeyinin kontrole göre
yaklaşık 24 kat artışına sebep olmuş ve ifade düzeyinin diğer uygulamalar arasında en
yüksek seviyeye ulaşmasını sağlamıştır. Diğer uygulanan konsantrasyonlarda, ifade
düzeylerinde dalgalanmalar görülse de kontrole göre önemli bir değişim gözlenmemiştir
(Şekil 4.46).
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan örneklerin artan Cr(VI)
konsantrasyonuna karşı farklılaşan RBR gen ifade seviyelerinin One Way ANOVA
(Dunnet) metodu ile p<0,05 düzeyinde istatistiksel olarak anlamlı olduğu görülmektedir
(Çizelge 4.141 - 4.142, 4.145 - 4.146).

215
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan ayçiçeği çeşitlerinde tek yönlü
varyans analizi (One Way ANOVA Dunnet Test, Post Hoc testi) ile konsantrasyona
karşı RBR geni ekspresyon seviyesinde meydana gelen değişimin istatistiksel olarak
p<0.05 düzeyinde anlamlı olduğu görülmektedir (Çizelge 4.143 - 4.144, 4.147 - 4.148).
TARSAN 1018 çeşidinde, 1280 ppm Cr(VI) uygulamasının kontrole göre RBR gen
ifade düzeyinde meydana getirdiği değişim istatistiki olarak p<0.05 düzeyinde
anlamlıyken, 320 ppm Cr(VI) uygulamasına göre p>0,05 düzeyinde anlamsızdır
(Çizelge 4.148).
Vuosku vd. (2015), sarıçam zigotik embriyogenezinde, hücre ölümü süreci ile ilişkili
olan katalaz ve retinoblastoma ilişkili protein genlerinin ifadelerini araştırdıkları
çalışmalarında, tüm doku ve gelişimsel aşamalarda, otofaji ilgili ATG5, katalaz ve
retinoblastoma ilişkili protein (RBR)leri kodlayan genlerin ekspresyonları ile gen
ifadesi ve hücre ölüm programı arasında bir bağlantı olduğunu göstermişlerdir. Nişasta
birikimi ve düşük RBR ekspresyonu, embriyoyu çevreleyen bölgedeki (ESR)
megagametofit hücrelerinin karakteristiği iken, güçlü RBR ifadesi bitki tohum taslağı
katmanlarında geç embriyo gelişiminde gözlenmiştir. ERS’de düşük RBR ifadesi
bulunması, sarıçam megagametophyte hücrelerinde hücre ölümü öncesinde, RBR’nin
aşağı regülasyonunu düşündürdüğü, tohum taslağı katmanlarındaki hücre ölümünün,
yoğun RBR ifadesiyle hasarlı DNA’nın replikasyonunu önleyen ve hücre döngüsünün
durmasına yol açan oksidatif stresle bağlantılı olabileceği sonucuna varmışlardır.
Cr(VI) uygulamasının oluşturduğu stres her bir çeşitte farklı konsantrasyonlarda RBR
seviyesinde artışa sebep olmuştur. Bu artış Vuosku vd. (2015) yaptıkları çalışmanın
sonuçlarıyla benzer olarak oksidatif stres kaynaklı olabilir.

216
5.5 MYC2 Geni İfade Seviyesinin Belirlenmesi
Myelocytomatosis proteinleri, hem bitki hem de hayvanlarda bulunan farklı görevlere
sahip transkripsiyon faktörleridir. bHLH domaini içeren transkripsiyon faktörleri alt gen
ailesini temsil eder (Toledo-Ortiz vd. 2003).
bHLH proteinleri, ağırlıklı olarak antosiyanin biyosentezi, fitokrom sinyalizasyonu,
tohum globulin ifadesi, meyve açılması, karpel ve epidermal gelişme işleviyle
karakterize edilmektedir (Buck ve Atchley 2003).
Myc hücre döngüsü sırasında G1 fazında ortaya çıkmaya başlar ve G1/S kontrol
noktasındaki çeşitli anahtar hücre döngüsü genlerini hedef alır. c-Myc, G1/S kontrol
noktası için uygun proteindir ve mitotik uyarı oluşmadığı durumlarda S fazındaki
hücreleri olumlu yönde uyarır (Eilers 1999).
Bitki hormonu absisik asit (ABA), kuraklık ve tuz stresine yanıtı içeren, çeşitli
fizyolojik süreçlere aracılık eder. ABA, suyun yetersiz olduğu şartlarda üretilir ve stoma
kapanmasına ve tuz ve kuraklık stresine toleransa neden olur. Günümüze kadar kuraklık
sonucu uyarılabilen genler üzerine yapılan çalışmaların çoğu, bu genlerin ABA ile
uyarıldığını ortaya koymuştur (Bray 1997). Kuraklık stresinin, ABA üretimini
tetiklediği görülmektedir. Abe vd. (2003), yaptıkları çalışmada, her iki AtMYC2 ve
AtMYB2 proteinin, bitkilerde kuraklık stresi altında ABA ile uyarılabilir gen
ekspresyonunda transkripsiyonel aktivatörler olarak işlev gösterdiğini bildirmişlerdir.
AtMYC2 ve AtMYB2 genleri bitkisel dokuda olduğu gibi, tohumlarda da ifade edilir
(Abe vd. 1997) ve transgenik bitkilerde AtMYB2 ve /veya AtMYC2 proteinlerinin fazla
sentezlenmesi hem vejetatif doku, hem de tohumlarda, önemli ABA aşırı duyarlılığını
göstermektedir. Bu bulgular, her iki AtMYC2 ve AtMYB2 proteinin, bitkisel dokuda
olduğu gibi köklerde de ABA sinyal transdüksiyonunda, transkripsiyonel aktivatörler
olarak rol oynadığını göstermektedir.

217
5.5.1 100 mM ve 150 mM NaCl stresi ile MYC2 seviyesinde meydana gelen değişim
100 mM NaCl stresi uygulaması, 08-TR-003 çeşidinde, 3. saatte MYC2 gen ifade
düzeyinin kontrole göre düşmesine sebep olmuştur. 27 saat uygulama, 3. saate göre
ifade düzeyini arttırmış olmasına rağmen ifade düzeyi yine de kontrolün altında
kalmıştır.
TR-3080 çeşidinde 100 mM NaCl stresi uygulaması, MYC2 gen ifade düzeyinde
zamana bağlı düşüşe sebep olurken, 27.saatteki düşüşün kontrole göre daha belirgin
olduğu görülmektedir.
SARAY çeşidinde, 100 mM NaCl stresi uygulaması, MYC2 gen ifade düzeyinde
zamana bağlı artışa sebep olmuştur. 3. saatte, kontrole göre gözlenen artışın daha
belirgin olduğu gözlenirken, 3. ve 27. saatlerdeki ifade düzeyinde önemli bir değişim
olmadığı görülmektedir.
TARSAN 1018 çeşidinde, ifade düzeyinde, 3. saatte kontrole göre önemli derecede artış
olduğu (yaklaşık 7 kat) görülmektedir. 27. saatte, gen ifade düzeyinde gerileme
görülmüş ve kontrol seviyesinin bile altına düşmüştür (Şekil 4.35).
100 mM NaCl stresi uygulanan örneklerin zamana karşı farklılaşan MYC2 gen ifade
seviyelerinin istatistiksel olarak anlamlı olup olmadığı One Way ANOVA (Dunnet)
metodu kullanılarak tespit edilmiş ve meydana gelen değişikliğin p<0,05 düzeyinde
istatistiksel olarak anlamlı olduğunu görülmüştür (Çizelge 4.25, 4.27, 4.29, 4.31).
100 mM tuz (NaCl) stresi uygulanan grupların tek yönlü varyans analizi (One Way
ANOVA Dunnet Test, Post Hoc testi) ile zamana karşı MYC2 geni ekspresyon
seviyesinde meydana gelen değişikliklerin anlamlılık dereceleri karşılaştırılmıştır
(Çizelge 4.26, 4.28, 4.30, 4.32).

218
08-TR-003 ve TARSAN 1018 çeşitlerinde 100 mM NaCl uygulamasının kontrole ve
diğer örnek alım zamanlarının birbirine göre kıyaslanmasında, MYC2 gen ifade
düzeyinde meydana gelen değişimin p<0,05 düzeyinde istatistiksel olarak anlamlı
olduğu görülmüştür (Çizelge 4.26, 4.32).
TR-3080 çeşidinde, 3. saatteki MYC2 gen ifade düzeyinin kontrole göre değişimi
p>0,05 düzeyinde istatistiksel olarak anlamsızken, 27. saatteki gen ifade düzeyinin
kontrole göre değişimi p<0,05 düzeyinde istatistiksel olarak anlamlıdır (Çizelge 4.28).
SARAY çeşidinde ise 3. ve 27. saatlerde kontrole göre ifade düzeyindeki değişim
p<0,05 düzeyinde istatistiksel olarak anlamlıyken, 27. saatteki ifadenin 3.saate göre
değişimi p>0,05 düzeyinde istatistiksel olarak anlamsızdır (Çizelge 4.30).
08-TR-003 ve TR-3080 çeşitlerinde 100 mM tuz uygulamasında, MYC2 seviyesinde
kontrole göre gözlenen azalma, hücre döngüsünde G1 fazından S fazına geçişini bloke
etmiş olabilir. SARAY çeşidinde stres uygulama süreleriyle birlikte gen ifade
düzeyinde görülen artış, bitki tolerans mekanizmalarının çalıştırıldığı ve hücre döngü
ilerleyişinin aktive edildiğini gösterebilir. TARSAN 1018 çeşidinde gen ifade
düzeyinde gözlenen değişim sonucu, uygulamanın erken saatlerinde hücre döngüsü
ilerleyişinin aktive edildiği, uzayan uygulamalarda tuz stresine karşı savunma
mekanizmalarının aktivasyonuyla hücre döngüsü ilerleyişinin bloke edildiği
söylenebilir.
150 mM NaCl stresi uygulaması, 08-TR-003 çeşidinde 3. saatte MYC2 gen ifade
seviyesinde kontrole göre azalmaya sebep olmuştur. 27 saat uygulamada, 3. saatle
MYC2 gen ifade düzeyinde anlamlı bir değişime sebep olmamıştır. İstatistiksel
analizler de bu durumu desteklemekte, 3. ve 27. saatlerde gen ifade düzeyindeki
değişim birbirlerine göre değerlendirildiğinde, p>0,05 düzeyinde istatistiksel olarak
anlamsız olduğu görülmektedir.

219
150 mM NaCl stresi uygulaması TR-3080, SARAY ve TARSAN 1018 çeşitlerinde 3.
saatte kontrole göre artışa, 27. saatte azalmaya sebep olmuştur. MYC2 gen ifade
düzeyinin en fazla olduğu 3. saatler bu üç çeşit arasında değerlendirildiğinde SARAY
çeşidinde ifade seviyesinin diğerlerinden daha fazla olduğu görülmektedir (Şekil 4.38).
TR-3080, SARAY ve TARSAN 1018 çeşitlerinde 150 mM NaCl uygulamasının
kontrole ve diğer örnek alım zamanlarının birbirine göre kıyaslanmasında, MYC2 gen
ifade düzeyinde meydana gelen değişimin p<0,05 düzeyinde istatistiksel olarak anlamlı
olduğunu görülmüştür(Çizelge 4.58, 4.60, 4.62).
Son bulgular tuz stresini düzenlemede ABA ve jasmonate (JA) arasında bazı kanıtlar
olduğunu göstermektedir. (Shinozaki and Yamaguchi-Shinozaki 2007). JA, JAZ
(Jasmonatla ZIM-domain) transkripsiyon faktörlerinin modülasyonuyla gelişme ve bitki
savunma yanıtlarında yer almaktadır. MYC2 transkripsiyon faktörleri sıklıkla JA, ABA
ve tuz stresinin gen ekspresyonunu düzenlemek için kullanılır. JA, MYC2’yi direk
baskılayarak fonksiyon gösteren JAZ’ın proteolizini uyarır, bu sayede downstream
hedef gen ifadelerinin aktivasyonu için MYC2 transkripsiyon faktörlerini sağlar (Kazan
ve Manners 2012).
Ji vd. (2012)’de yaptıkları çalışmada, NaCl, osmotik ve ABA stres koşulları altında
MYC genlerinin rolünü araştırmak için Tamarix hispida’dan dokuz MYC geni
klonlamışlardır. Bu çalışmanın sonuçları MYC’lerin tuz ve ozmotik stres toleransında
rol oynadığını ve ABA sinyal transdüksiyon yolu tarafından kontrol edildiğini
göstermektedir.
MYC2 jasmonik asit (JA), absisik asit, oksin, etilen ve gibberellik asit gibi, çoklu sinyal
yollarına müdahale eder (Kazan ve Manners 2012, Gangappa vd. 2013b).
Başlangıçta JIN1 / JAI1 (jasmonat duyarsız 1) olarak izole edilen MYC2/ZBF1, JA-
kaynaklı genlerin iki grubunun ekspresyonunu farklı şekillerde düzenler. Birinci grupta
yer alan, patojenlere karşı savunma yanıtlarında rol oynayan genler, MYC2 ile

220
baskılanır. İkinci gruba dahil edilen, yaralanmada JA-aracılığı ile sistemik yanıtlarda
rol oynayan genler, MYC2 tarafından aktive edilir. Bu nedenle, MYC2, JA sinyal
yollarının hem pozitif, hem de negatif düzenleyici olarak davranır. MYC2, büyüme
evresinde, karanlıkta ve ışığın çeşitli dalga boylarında ifade edilir ve absisik asit ve JA-
aracılıklı uyarım MYC2 ekspresyonunu daha da arttırır (Abe vd. 2003, Anderson vd.
2004, Lorenzo vd. 2004, Yadav vd. 2005). MYC2 birikiminin, düşük düzeyleri, JA
yokluğunda olduğu bildirilmiştir (Zhai vd. 2013).
MYC2 seviyesinde tuz stresine bağlı gözlenen artışın sebebi, MYC2 ve ABA
arasındaki ilişkinin araştırıldığı çalışmalardan (Abe vd. (1997, 2003), Kazan ve
Manners (2012), Ji vd. (2012) Anderson vd. (2004), Lorenzo vd. (2004), Yadav vd.
(2005), Zhai vd. (2013)) elde edilen sonuçlarla benzer olarak tuz toleransında ABA ve
JA tarafından uyarılabilir gen ekspresyonunda transkripsiyonel aktivatörler olarak rol
alması olabilir.
150 mM tuz uygulamasının 08-TR-003 çeşidinde, 100 mM tuz uygulamasına göre
MYC2 gen ifade düzeyini daha çok azalttığı ve bu durumun hücre döngü ilerleyişinin
daha çok baskılanmasına sebep olduğu söylenebilir. TR-3080, SARAY ve TARSAN
1018 çeşitlerinde stresin kısa süreli uygulamasının hücre bölünmesini uyardığı, uzayan
sürelerin baskıladığı söylenebilir.
5.5.2 100 mM ve 150 mM PEG stresi ile MYC2 seviyesinde meydana gelen değişim
100 mM PEG stresi uygulaması, çalışılan tüm çeşitlerde 3. saatte kontrole göre MYC2
gen ifade düzeyinde önemli bir değişime sebep olmazken, 27. saatte ifade düzeyinde
önemli artışlar olduğu gözlenmiştir. En yüksek ifade seviyesi 27. saatte kontrole göre
yaklaşık 540 kat (539,7747386) artışla TR-3080 çeşidinde görülürken, ikinci sırada
yaklaşık 461 kat artışla (461,0862195) SARAY çeşidinde gözlenmiştir (Şekil 4.41).
100 mM PEG stresi uygulamasının kontrole ve diğer örnek alım zamanlarının birbirine
göre kıyaslanmasında, MYC2 gen ifade düzeyinde meydana gelen değişimin p<0,05

221
düzeyinde istatistiksel olarak anlamlı olduğunu görülmüştür (Çizelge 4.86, 4.88, 4.90,
4.92).
Sonuçlar, tüm çeşitlerde 100 mM PEG stresinin 3 saat süreyle uygulanmasının hücre
döngüsü ilerlemesi üzerine etkisinin olmadığının, 27 saat uygulamada gen ifadesinde
görülen artışın kuraklık stresine karşı toleranslı olduklarının göstergesi olabileceği
düşünülmektedir.
150 mM PEG stresi uygulaması, 08-TR-003 ve SARAY çeşitlerinde, MYC2 gen ifade
düzeyinde 3. saatte kontrole göre artışa, 27. saatte, 3. saate göre azalmaya sebep
olmuştur. İfade düzeyinde 3. saatte kontrole göre en belirgin yükseliş, yaklaşık 18 kat
(18,36440733) artışla, SARAY çeşidinde görülmüştür.
150 mM PEG stresi uygulaması, TR-3080 çeşidinde MYC2 gen ifade düzeyinde 3.
saatte kontrole göre bir miktar azalmaya, 27. saatte ise 3. saat ve kontrole göre artışa
sebep olmuştur. TARSAN 1018 çeşidinde ise zamana bağlı gen ifade seviyesinde artış
görülürken, en belirgin artış 27. saatte gözlenmiştir (Şekil 4.44).
150 mM PEG stresi uygulamasının kontrole ve diğer örnek alım zamanlarının birbirine
göre kıyaslanmasında, MYC2 gen ifade düzeyinde meydana gelen değişimin p<0,05
düzeyinde istatistiksel olarak anlamlı olduğunu görülmüştür (Çizelge 4.116, 4.118,
4.120, 4.122).
150 mM PEG stresi uygulaması sonucu MYC2 gen ifadesi seviyesindeki değişimler
değerlendirildiğinde, 08-TR-003 ve SARAY çeşitlerinde kısa süreli stres
uygulamalarının hücre bölünmesini aktive ettiği, uzayan sürelerde stres uygulamasının
bölünmeyi bloke ettiği söylenebilir.

222
5.5.3 Cr (VI) stresi ile MYC2 seviyesinde meydana gelen değişim
Cr(VI) stresi uygulaması sonucu, artan Cr(VI) konsantrasyonlarıyla birlikte 08-TR-003
çeşidinde, MYC2 gen ifade düzeyinin, dalgalanmalar görülse de kontrole göre azalma
eğiliminde olduğu, 640 ppm ve 1280 ppm uygulamalarda ifade seviyesinin en alt
seviyede olduğu görülmektedir. TR-3080, SARAY ve TARSAN 1018 çeşitlerinde en
yüksek ifade düzeyi 160 ppm Cr(VI) uygulamasında görülmüştür. 160 ppm dışındaki
konsantrasyonlarda kontrole göre MYC2 gen ifade düzeyinde önemli bir değişim
görülmemiştir (Şekil 4.47).
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan örneklerin artan Cr(VI)
konsantrasyonuna karşı farklılaşan MYC2 gen ifade seviyelerinin One Way ANOVA
(Dunnet) metodu ile p<0,05 düzeyinde istatistiksel olarak anlamlı olduğunu
görülmektedir (Çizelge 4.154, 4.155, 4.158, 4.159).
Farklı konsantrasyonlarda Cr(VI) stresi uygulanan 08-TR-003, TR-3080 ve TARSAN
1018 çeşitlerinde tek yönlü varyans analizi (One Way ANOVA Dunnet Test, Post Hoc
testi) ile konsantrasyona karşı MYC2 geni ekspresyon seviyesinde meydana gelen
değişimin istatistiksel olarak p<0.05 düzeyinde anlamlı olduğu görülmektedir (Çizelge
4.156, 4.157, 4.161). SARAY çeşidinde, 640 ppm Cr(VI) uygulamasının kontrole göre
MYC2 gen ifade düzeyinde meydana getirdiği değişim (1,000) istatistiki olarak p>0,05
düzeyinde anlamsız olup, diğer konsantrasyonlarda kontrole göre meydana gelen
değişimler p<0.05 düzeyinde anlamlıdır (Çizelge 4.160).
160 ppm Cr(VI) stresi uygulaması sonucu, TR-3080, SARAY ve TARSAN 1018
çeşitlerinde MYC2 seviyesinde gözlenen artış, antioksidan savunma sistemlerinin
aktivasyonuyla açıklanabilir. Artan konsantrasyonlarda ekspresyon seviyesindeki
gerilemenin, hücrenin apoptozisinden kaynaklanabileceği düşünülmektedir. 08-TR-003
çeşidinde uygulanan Cr konsantrasyonlarındaki artışla beraber hücre bölünmesi
baskılanmış olabilir.

223
6. SONUÇ
Önemli bir tarım bitkisi olan ayçiçeğinin hücre büyüme ve gelişiminin temel
mekanizmalarını etkileyen Retinoblastoma (Rb), Myelocytomatosis (Myc) ve E2F
genleri ile ilgili böyle bir çalışma daha önce yapılmamış olup, ayçiçeğinin stres
koşullarına göstermiş olduğu tepkinin genetik mekanizması henüz aydınlatılmamıştır.
Bu çalışma ile belli abiyotik stres faktörleri varlığında hücre döngüsünde önemli
yolaklarda yer alan bu genlerin ekspresyonlarındaki değişimlerin tespit edilmesiyle
hücre döngüsü düzenleme mekanizmalarının açığa çıkarılmasına katkı sağlanacak
veriler elde edilmiştir.
Literatürde bu gen bölgeleriyle yapılmış benzer çalışmalar bulunmadığından,
çalışmadan elde edilen veriler, genlerin hücre döngüsündeki rolleri esas alınarak
değerlendirilmiştir. Çalışılan abiyotik stres koşullarında gen ifadelerindeki değişimler
bitkilerin genel olarak tuz ve kuraklık stresine karşı toleranslı oldukları konusunda ön
bilgi vermektedir. Total protein miktarları ve MDA seviyeleri de bu sonuçları destekler
niteliktedir. Ayrıca, Cr(VI) stresinin özellikle 640 ppm ve 1280 ppm uygulamalarının
bitkiler üzerindeki olumsuz etkilere sebep olduğu ve gen ifade seviyelerinde önemli
değişmelere yol açtığı görülmüştür. Total protein miktarları ve MDA seviyeleri de bu
uygulamaların hücreyi apoptozise götürmüş olabileceğini düşündürmektedir.
Hücre döngüsü, bitkilerde birçok faktör tarafından kontrol edilen önemli bir süreçtir.
Sadece belli genlerde meydana gelen ifade farklılıklarının değerlendirilmesi, hücre
döngüsünün gidişatı konusunda yorum yapmak için yeterli olmayabilir. Ancak bu
konuda yapılacak çalışmalar için yol gösterici olacağı düşünülmektedir. Genlerin
görevleri anlaşıldıkça gen aktarımı veya gen susturma ile yeni transgeniklere ortam
sağlanacaktır. Ülkemizde ve dünyada geniş kullanım alanı olan ve dolayısıyla yüksek
ekonomik değere sahip ayçiçeğinde, bitkilerin yaygın olarak maruz kaldığı bu abiyotik
stres faktörlerinin etkilediği hücre döngüsü genlerinin aydınlatılması, sadece istenilen
özelliklere sahip yeni ayçiçeği transgeniklerinin oluşturulmasına değil, aynı zamanda
ekonomik değeri yüksek birçok tarım bitkisinde de aynı uygulamaların yapılabilmesine
olanak sağlayacaktır.

224
KAYNAKLAR
Abe, H., Yamaguchi-Shinozaki, K., Urao, T., Iwasaki, T., Hosokawa, D. and Shinozaki,
K. 1997. Role of Arabidopsis MYC and MYB homologs in drought- and
abscisic acid-regulated gene expression. Plant Cell 9; 1859–1868.
Abe, H., Urao, T., Ito, T., Seki, M., Shinozaki, K. and Yamaguchi-Shinozaki, K. 2003.
Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional
activators in abscisic acid signaling. Plant Cell, 15; 63–78.
Ach, R.A., Taranto, P. and Gruissem, W. 1997. A conserved family of WD-40 proteins
binds to the retinoblastoma protein in both plants and animals. Plant Cell, 9;
1595–1606.
Adams, M.R., Sears, R., Nuckolls, F., Leone, G. and Nevins, J.R. 2000. Complex
transcriptional regulatory mechanisms control ex- pression of the E2F3 locus.
Molecular and Cellular Biology, 20; 3633–9.
Agastian, P., Kingsley, S.J. and Vivekanandan, M. 2000. Effect of salinity on
photosynthesis and biochemical characteristics in mulberry genotypes.
Photosynthetica, 38; 287–290.
Alamgir, A.N.M. and Ali, M.Y. 1999. Effect of salinity on leaf pigments, sugar and
protein concentrations and chloroplast ATPAase activity of rice (Oryza sativa
L.). Bangladesh J. Bot., 28; 145–149.
Alberts, B. 2008. Hücrenin Moleküler Biyolojisi. Türkiye Bilimler Akademisi, 985,
Ankara.
Anastassopoulou, J. 2003. Magnesium-DNA interactions and the possible relation of
magnesium to carcinogenesis. Irradiation and free radicals, 42(1); 79.
Anderson, J.P., Badruzsaufari, E., Schenk, P.M., Manners, J.M., Desmond, O.J., Ehlert,
C., Maclean, D.J., Ebert, P.R. and Kazan, K. 2004. Antagonistic interaction
between abscisic acid and jasmonateethylene signaling pathways modulates
defense gene expression and disease resistance in Arabidopsis. Plant Cell, 16;
3460–3479.
Anonim. 2015. Web Sitesi: http://arastirma.tarim.gov.tr/ ttae/Sayfalar/Detay.aspx?
SayfaId=92. Erişim Tarihi: 10.08.2015.
Anonim. 2015. Web Sitesi: http://arastirma.tarim.gov.tr/ttae/Sayfalar/Detay.aspx?
SayfaId=96. Erişim Tarihi: 10.08.2015.
Anonim. 2015. Web Sitesi: http://www.betaziraat.com.tr/Tr/tarla-bitkileri/aycicegi.
Erişim Tarihi: 10.08.2015.

225
Anonim. 2015. Web Sitesi: http://koop.gtb.gov.tr/data/53319ccf487c8eb1e43d
7298/2014%20Ay%C3%A7i%C3%A7e%C4%9Fi%20Raporu.pdf. T.C.
Gümrük ve Ticaret Bakanlığı Koperatifçilik Genel Müdürlüğü, 2014 Ayçiçeği
Raporu. Erişim Tarihi: 12.08.2015.
Anonim. 2015. Web Sitesi: https://biruni.tuik.gov.tr/bitkiselapp/bitkisel.zul. Erişim
Tarihi: 12.10.2015.
Anonymous. 2015. Web Sitesi: http://arastirma.tarim.gov.tr/ttae/Sayfalar/EN/Detay
.aspx? SayfaId=130. Erişim Tarihi: 10.08.2015.
Arabi, A,. Wu, S., Ridderstrale, K., Bierhoff, H., Shiue, C., Fatyol, K., Fahlen, S.,
Hydbring, P., Söderberg, O., Grummt, I., Larsson, L.G. and Wright, A.P. 2005.
c-Myc associates with ribosomal DNA and activates RNA polymerase I
transcription. Nat Cell Biol, 7; 303-10.
Artlip, T.S. and Funkhouser, E.A. 1995. Protein Synthetic Responses to Environmental
Stresses. M. Pessarakli (Ed.), Handbook of Plant and Crop Physiology, pp. 627–
644. Marcel Dekker, New York.
Ashraf, M. 1994. Breeding for salinity tolerance in plants. Critjical Reviews in Plant
Sciences, 13(1); 17-42.
Ashraf, M. and Fatima H. 1995. Responses of some salt tolerant and salt sensitive lines
of safflower (Carthamus tinctorius L.). Acta Physiol. Plant, 17; 61–71.
Ashraf, M. and Tufail, M. 1995. Variation in salinity tolerance in sunflower (Helianthus
annuus L.). J. Agron. Soil Sci., 174; 351–362.
Ashraf, M. and Haris, P.J.C. 2004. Potential biochemical indicators of salinity tolerance
in plants. Plant Science, 166; 3-16.
Baloğlu, M. C., Kavas, M., Aydın, G. Öktem, H. A. and Yücel, A.M. 2012.
Antioxidative and physiological responses of two sunflower (Helianthus
annuus) cultivars under PEG-mediated drought stress. Turk J Bot., 36; 707-714.
Blum, A. 1986. Breeding Crop Varieties for Stress Environments. Critical Reviews in
Plant Sciences, 2; 199-237.
Boniotti, M.B. and Gutierrez, C. 2001. A cell-cycle-regulated kinase activity
phosphorylates plant retinoblastoma protein and contains, in Arabidopsis, a
CDKA/cyclin D complex. Plant J., 28; 341–350.
Borghi, L., Gutzat, R., Fütterer, J., Laizet, Y., Hennig, L. and Wilhelm Gruissem, W.
2010. Arabidopsis RETINOBLASTOMA-RELATED Is Required for Stem Cell
Maintenance, Cell Differentiation, and Lateral Organ Production. The Plant Cell,
22; 1792–1811.

226
Bradford, M.M. 1976. Rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal.
Biochem., 72; 248–254.
Bray, E.A. 1997. Plant responses to water deficit. Trends Plant Sci., 2; 48-54.
Bray, E.A., Bailey-Serres, J. and Weretilnyk, E. 2000. Responses to abiotic stress. In.
Buchanan BB, Gruissem W, Jones RL, eds. Biochemistry and molecularbiology
of plants. Rokwille: American Society of Plant Physiologists, 1158-1203.
Bray, E.A. 2007. Molecular and Physiological Responses to Water-Deficit Stres. In:
Advances in Molecular Breeding Toward Drought. M.A. Jenks et al. (eds.).
Springer, Netherlands.
Buck, M.J. and Atchley, W.R. 2003. Phylogenetic analysis of plant basic helix-loop-
helix proteins. J Mol Evol., 56; 742-750.
Cheruth, A.J., Ksouri, R., Ragupathi, G., Paramasivam, M., Jallali, I., Hameed, J.A.,
Zhao, C.X., Shao, H.B. and Rajaram, P. 2009. Antioxidant defense responses:
physiological plasticity in higher plants under abiotic constraints. Acta Physiol
Plant, 31; 427-436.
Chinnusamy, V., Schumaker, K. and Zhu, J.K. 2004. Molecular genetic perspectives on
cross-talk and specificity in abiotic stress signalling in plants. J Exp Bot., 55;
225–236.
Cornic, G., 2000. “Drought stress inhibits photosynthesis by decreasing stomatal
aperture not by affecting ATP synthesis.” Trends in Plant Science5: 187–188.
Costas C., Desvoyes, B. and Gutierrez, C. 2011. A chromatin perspective of plant cell
cycle progression. Biochimica et Biophysica Acta, 1809; 379–387.
Day, S., Kaya, D. ve Kolsarıcı, Ö. 2008. Bazı Çerezlik Ayçiçeği (Helianthus annuus L.)
Genotiplerinin Çimlenmesi Üzerine NaCl Konsantrasyonlarının Etkileri. Tarım
Bilimleri Dergisi, 14 (3); 230-236.
de Jager S.M., Scofield S., Huntley R.P., Robinson A.S., den Boer, B.G. and Murray,
J.A. 2009. Dissecting regulatory pathways of G1/S control in Arabidopsis:
Common and distinct targets of CYCD3;1, E2Fa and E2Fc. Plant Mol. Biol., 71;
345–365.
De Veylder, L., Beeckman, T., Beemster, G.T.S., Engler, J.A., Ormenese, S., Maes, S.,
Naudts, M., Schueren, E.V.D., Jacqmard, A., Engler, G. and Inzé, D. 2002.
Control of proliferation, endoreduplication and differentiation by the
Arabidopsis E2Fa–DPa transcription factor. EMBO J., 21; 1360–1368.
De Veylder, L., Joube`s, J. and Inze, D. 2003. Plant cell cycle transitions. Curr Opin
Plant Biol., 6; 536–543.

227
De Veylder, L., Beeckman, T. and Inzé, D. 2007. The ins and outs of the plant cell
cycle. Molecular Cell Bıology, 8; 655-665.
del Pozo, J.C., Boniotti, MB. and Gutierrez, C. 2002. Arabidopsis E2Fc functions in cell
division and is degraded by the ubiquitin-SCF(AtSKP2) pathway in response to
light. Plant Cell, 14; 3057–3071.
del Pozo, J.C., Diaz-Trivino, S., Cisneros, N. and Gutierrez, C. 2006. The balance
between cell division and endoreplication depends on E2FC-DPB, transcription
factors regulated by the ubiquitin-SCFSKP2A pathway in Arabidopsis. Plant
Cell, 18; 2224–2235.
Desvoyes, B., Elena, R.P., Xie, Q., Chua, N.H. and Gutierrez, C. 2006. Cell type-
specific role of the retinoblastoma/E2F pathway during Arabidopsis leaf
development. Plant Physiol., 140; 67-80.
Dewitte, W. and Murray, JAH. 2003. The plant cell cycle. Annu Rev Plant Biol., 54;
235–264.
Dixit, V., Pandey, V. and Shyam, R. 2002. Chromium Ions Inactivate Electron
Transport and Enhance Superoxide Generation in vivo in Pea (Pisum sativum L.
cv. Azad) Root Mitochondria. Plant Cell Environ., 25; 687-690.
Dubey, R.S. and Rani, M. 1989. Influence of NaCl salinity on growth and metabolic
status of protein amino acids in rice seedling. J. Agron. Crop Sci., 162; 97–106.
Dudits, D., Ábrahám, E., Miskolczi, P., Ayaydin, F., Bilgin, M. and Horváth, G.V.
2011. Cell-cycle control as a target for calcium, hormonal and developmental
signals: the role of phosphorylation in the retinoblastoma-centred pathway.
Annals of Botany, 107; 1193–1202.
Dunnet, C.W. 1955. A multiple comparison procedure for comparing several treatments
with a control. Journal of the American Statistical Association, 50; 1096-1121.
Ebel, C., Mariconti, L. and Gruissem, W. 2004. Plant retinoblastoma homologues
control nuclear proliferation in the female gametophyte. Nature, 429; 776–780.
Ebrahimian, E. and Bybordi, A. 2012. Effect of salinity, salicylic acid, silicium and
ascorbic acid on lipid peroxidation, antioxidant enzyme activity and fatty acid
content of sunflower. African Journal of Agricultural Research, 7(25); 3685-
3694.
Eilers, M. 1999. Control of cell proliferation by Myc family genes. Mol Cells, 9; 1-6.
Elledge, S.J. 1996. Cell cycle checkpoints: preventing an identity crisis. Science (New
York, N.Y.), 274; 1664–1672.

228
Ellouzia, H., Hameda, K.B., Celab, J., e-Boschb, S. M. and Abdellya, C. 2011. Early
effects of salt stress on the physiological and oxidative status of Cakile maritima
(halophyte) and Arabidopsis thaliana (glycophyte). Physiologia Plantarum, 142;
128–143.
Foolad, M. 2007. Current Status of Breeding Tomatoes for Salt and Drought Tolerance,
In: Advances in Molecular Breeding Toward Drought. M.A. Jenks et al. (eds.).
Springer, Netherlands.
Foyer, C.H. and Noctor, G. 2000. Oxygen processing in photosynthesis: regulation and
signalling. New Phytol., 146; 359–388.
Gadallah, M.A.A. 1999. Effects of proline and glycinebetaine on Vicia faba responses
to salt stress. Biol. Plant., 42; 249–257.
Gangappa, S.N., Srivastava, A.K., Maurya, J.P., Ram, H. and Chattopadhyay, S. 2013b.
Z-box binding transcription factors (ZBFs): a new class of transcription factors
in Arabidopsis seedling development. Mol. Plant, 6; 1758–1768.
Gomez-Roman, N., Grandori, C., Eisenman, R.N. and White, R.J. 2003. Direct
activation of RNA polymerase III transcription by c-Myc. Nature, 421; 290–4.
Grafi, G., Burnett, R.J., Helentjaris, T., Larkins, B.A., DeCaprio, J.A., Sellers, W.R. and
Kaelin, W.G. 1996. A maize cDNA encoding a member of the retinoblastoma
protein family: involvement in endoreduplication. Proc. Natl. Acad. Sci. U.S.A.
93, 8962–8967.
Gutzat, R., Borghi, L., Fütterer, J., Bischof, S., Laizet, Y., Hennig, L., Feil, R., Lunn, J.
and Gruissem, W. 2011. RETINOBLASTOMA-RELATED PROTEIN controls
the transition to autotrophic plant development. Development (Cambridge,
England), 138; 2977–2986.
Hasegawa, P.M., Bressan, R.A., Zhu, J.K. and Bohnert, H.J. 2000. Plant cellular and
molecular responses to high salinity. Annual Review of Plant Physiology and
Plant Molecular Biology, 51; 463–499.
Heiser, C.B. 1955. The origin and development of cultivated sunflower. Am. Biol.
Teacher, 17; 161-167.
Helin, K. 1998. Regulation of cell proliferation by the E2F transcription factors. Curr
Opin Genet Dev., 8; 28–35.
Hichri, I., Heppel, S.C., Pillet, J., Léon, C., Czemmel, S., Delrot, S., Lauvergeat, V. and
Bogs, J. 2010. The basic helix–loop–helix transcription factor MYC1 is involved
in the regulation of the flavonoid biosynthesis pathway in grapevine. Molecular
Plant, 3; 509-523.

229
Hirano, H., Harashima, H., Shinmyo, A. and Sekine, M. 2008. Arabidopsis
RETINOBLASTOMA RELATED PROTEIN 1 is involved in G1 phase cell
cycle arrest caused by sucrose starvation. Plant Mol. Biol. 66:259-275.
Hoagland, D.R. and Arnon, D.I. 1938. The Water Culture Method for Growing Plants
Without Soil. Circ. Calif. Agr. Exp. Sta., 347-461.
Hodges, D.M., DeLong, J.M., Forney, C.F. and Prange, R.K. 1999. Improving the
thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in
plant tissues containing anthocyanin and other interfering compounds. Planta,
207; 604–611.
Huang, C., Wei, G., Jie, Y., Wang, L., Zhou, H., Ran, C., Huang, Z., Jia, H. And
Anjum, S.A. 2014. Effects of concentrations of sodium chloride on
photosynthesis, antioxidative enzymes, growth and fiber yield of hybrid ramie.
Plant Physiology and Biochemistry, 76; 86-93.
Inze´, D. 2005. Green light for the cell cycle. EMBO J., 24; 657–662.
Ji, X., Wang, Y. and Liu, G. 2012. Expression Analysis of MYC Genes from Tamarix
hispida in Response to Different Abiotic Stresses. Int. J. Mol. Sci., 13; 1300-
1313.
Jullien, P.E., Mosquna, A., Ingouff, M., Sakata, T., Ohad, N. and Berger, F. 2008.
Retinoblastoma and Its Binding Partner MSI1 Control Imprinting in
Arabidopsis. PLoS Biol., 6(8); e194. doi:10.1371/journal.pbio.0060194
Kabata-Pendias, A. and Pendias, H. 2001. Trace elements in soils and plants. 3rd edn.
CRC Press Inc, Boca Raton, FL, p 413.
Karuppanapandian, T. and Manoharan, K. 2008. Uptake and Translocation of Tri- and
Hexa-Valent Chromium and Their Effects on Black Gram (Vigna mungo L.
Hepper cv. Co4) Roots. J. Plant Biol., 51, 192-201.
Katerji, N., Van Hoorn, J.W., Hamdy, A. and Mastrorıllı, M. 2004. Comparison of Corn
Yield Response to Plant Water Stress Caused by Salinity and By Drought.
Agricultural Water Management, 65; 95–101.
Katz, S.A. and Salem, H. 1994. The biological and environmental chemistry of
chromium. New York, NY: VCH Publishers.
Kaya, Y. 2004. Ayçiçeği biyoteknolojisinde son gelişmeler ve ıslahında kullanım
olanakları. Trakya Univ J Sci, 5(2); 141-147.
Kaya, M.D., Okçu, G., Atak, M., Çıkılı, Y. and Kolsarıcı, Ö. 2006. Seed Treatments to
Overcome Salt and Drought Stress During Germination in 334 Sunflower
(Helianthus annuus L.). European Journal of Agronomy, 24 (4); 291-295.

230
Kaya, A. and Yigit, E. 2014. The physiological and biochemical effects of salicylic acid
on sunflowers (Helianthus annuus) exposedto flurochloridone. Ecotoxicology
and Environmental Safety, 106; 232–238.
Kazan, K., and Manners, J.M. 2012. JAZ repressors and the orchestration of
phytohormone crosstalk. Trends Plant Sci., 17; 22–31.
Khedr, A.H.A., Abbas, M.A., Wahid, A.A.A., Quick, W.P. and Abogadallah, G.M.
2003. Proline induces the expression of salt stress responsive proteins and may
improve the adaptation of Pancratium maritimum L. to salt stress. J. Exp. Bot.,
54; 2553–2562.
Knight, J.A., Pleper, R.K. and Macellon, L. 1988. Specifity of the thiobarbituric acid
reaction: İts use in studies of lipid peroxidation. Clinical Chemistry, 34; 2433-
2438.
Komaki, S. and Sugimoto, K. 2012. Control of the Plant Cell Cycle by Developmental
and Environmental Cues. Plant Cell Physiol, 53(6); 953–964.
Kong, K., Daduang, S., Wongkham, C.H., Bun–Nag, S., Kosittrakun, M. and
Theerakulpisut, P. 2005. Protein profiles in response to salt stress in leaf sheaths
of rice seedlings. Sci. Asia, 31; 403–408.
Kurzbauer, M.T. and Schlögelhofer, P. 2011. Retinoblastoma protein goes green: the
role of RBR in Arabidopsis meiosis. The EMBO Journal, 30; 631–633.
Lammens, T., Boudolf, V., Kheibarshekan, L., Zalmas, L.P., Gaamouche, T., Maes, S.,
Vanstraelen, M., Kondorosi, E., La Thangue, NB, Govaerts, W., Inzé, D. and De
Veylder, L. 2008. Atypical E2F activity restrains APC/CCCS52A2 function
obligatory for endocycle onset. Proc Natl Acad Sci USA, 105; 14721– 14726.
Lavia, P. and Jansen-Dürr, P. 1999. E2F target genes and cell-cycle checkpoint control.
Bioessays, 21; 221–230.
Lawlor, D.W. 2002. Limitation to photosynthesis in water-stressed leaves:stoma vs.
metabolism and the role of ATP. Annals of Botany, 89; 871-885.
Lawlor, D.W. and Cornic, G. 2002. Photosynthetic carbon assimilation and associated
metabolism in relation to water deficits in higher plants. Plant Cell and
Environment, 25; 275–294.
Lee, H.O., Davidson, J.M. and Duronio, R.J. 2009. Endoreplication: polyploidy with
purpose. Genes Dev., 23(21); 2461–77.
Lees, C.B. 1965. Sunflower Hortic, 43; 24-25.

231
Leone, G., DeGregori, J., Sears, R., Jakoi, L. and Nevins, J.R. 1997. Myc and Ras
collaborate in inducing accumulation of active cyclin E/Cdk2 and E2F. Nature,
387; 422– 6.
Leone, G., Sears, R., Huang, E., Rempel, R., Nuckolls, F., Park, C.H., Giangrande, P.,
Wu, L., Saavedra, H.I., Field, S.J., Thompson, M.A., Yang,H., Fujiwara, Y.,
Greenberg, M.E., Orkin, S., Smith, C. and Nevins, J.R. 2001. Myc requires
distinct E2F activities to induce S phase and apoptosis. Mol Cell, 8; 105-13.
Liang, Y., Chen, Q., Liu, Q., Zhang, W. and Ding, R. 2003. Exogenous silicon (Si)
increases antioxidant enzyme activity and reduces lipid peroxidation in roots of
salt-stressed barley (Hordeum vulgare L.). J Plant Physiol., 160(10); 1157–1164.
Linnaeus, C. 1753. Species Plantarum, Tomus II: 904.
Livak, K.J. and Schmittgen, T.D. 2001. Analysis of Relative Gene Expression Data
Using Real- Time Quantitative PCR and the 2-ΔΔCT
Method. METHODS, 25;
402–408. doi:10.1006/meth.2001.1262.
Lorenzo, O., Chico, J., Sánches-Serrano, J. and Solano, R. 2004. JASMONATE-
INSENSITIVE1 encodes a MYC transcription factor essetioal to discriminate
between different jasmonate-regulated defense responses in Arabidopsis. Plant
Cell, 16; 1938–1950.
Magyar, Z., De Veylder, L., Atanassova, A., Bako, L., Inze, D. and Bogrea, L. 2005.
The Role of the Arabidopsis E2FB Transcription Factor in Regulating Auxin-
Dependent Cell Division. The Plant Cell, 17; 2527–254.
Mahajan, S. and Tuteja, N. 2005. Cold, salinity and drought stress: an overview.
Archives of Biochemistry and Biophysics, 444; 139-158.
Maiti, S., Ghosh, N., Mandal, C., Das, K., Dey, N., Adak, M.K. and Braz. J., 2012.
Responses of the maize plant to chromium stress with reference to antioxidation
activity Plant Physiol., 24(3); 203-212.
Mariconti, L., Pellegrini, B., Cantoni, R., Stevens, R., Bergounioux, C., Cella, R. and
Albani, D. 2002. The E2F family of transcription factors from Arabidopsis
thaliana. Novel and conserved components of the retinoblastoma/E2F pathway
in plants. J Biol Chem., 277; 9911–9919.
Marschner, H. 1995. Mineral Nutrition of Higher Plants. Academic Press, 657-680.
Meijer, M. and Murray, J.A.H. 2001. Cell cycle controls and the development of plant
form. Current Opinion in Plant Biology, 4; 44–49.
Menges, M., Pavesi, G., Morandini, P., Bögre, L. and Murray, J.A. 2007. Genomic
organization and evolutionary conservation of plant D-type cyclins. Plant
Physiology, 145; 1558–1576.

232
Miljuš-Djukić, J., Stanisavljević, N., Radović, S., Jovanović, Z., Mikić, A. and
Maksimović, V. 2013. Differential response of three contrasting pea (Pisum
arvense, P. sativum and P. fulvum) species to salt stress: assessment of variation
in antioxidative defence and miRNA expression. AJCS 7(13):2145-2153
ISSN:1835-2707.
Mirzaee, M., Moieni, A. and Ghanati F., 2013. Effects of Drought Stress on the Lipid
Peroxidation and Antioxidant Enzyme Activities in Two Canola (Brassica napus
L.) Cultivars. J. Agr. Sci. Tech., 15; 593-602.
Mittova, V., Guy, M., Tal, M. and Volokita, M. 2004. Salinity upregulates the
antioxidative system in root mitochondria and peroxisomes of the wild salt-
tolerant tomato species Lycopersicon pennellii. J Exp Bot., 55; 1105–1113.
Moussa, H.R., Abdel-Aziz, S.M., 2008. Comparative Response of Drought Tolerant and
Drought Sensitive Maize Genotypes to Water Stres. Australian Journal of Crop
Science, 1(1); 31-36.
Nakata, M., Mitsuda, N., Herde, M., J.K. Koo, A., Moreno, J. E., Suzuki, K, Howe, G.
A. and Ohme-Takagi, M. 2013. A bHLH-Type Transcription Factor, ABA-
INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED
MYC2-LIKE1, Acts as a Repressor to Negatively Regulate Jasmonate Signaling
in Arabidopsis. The Plant Cell, 25; 1641–1656.
Naouar, N., Vandepoele, K., Lammens, T., Casneuf, T., Zeller, G., van Hummelen, P.,
Weigel, D., Rätsch, G., Inzé, D., Kuiper, M., De Veylder, L. and Vuylsteke, M.
2009. Quantitative RNA expression analysis with Affymetrix Tiling 1.0R arrays
identifies new E2F target genes. Plant J., 57; 184–194.
Oren, R., Phillips, N., Ewers, B.E., Pataki, D.E. and Megonigal, J.P. 1999. Sap-flux-
scaled transpiration responses to light, vapor pressure deficit, and leaf area
reduction in a flooded Taxodium distichum forest. Tree Physiology, 19; 337–
347.
Panda, S.K. and Khan, M.H. 2003. Antioxidant efficiency in rice (Oryza sativa L.)
leaves under heavy metal toxicity. J Plant Biol., 30; 23–29.
Pandey, S.K. 2008. Germination and seedling growth of field pea (Pisum sativum)
Malviya Matar-15 (HUDP-15) and Pusa Prabhat (DDR-23) under varying level
of copper and chromium. J Am Sci., 4(3); 28–40.
Pareek-Singla, S.L. and Grover, A. 1997. Salt responsive proteins/genes in crop plants.
In: Jaiwal P.K., Singh R.P., Gulati A. (eds.): Strategies for Improving Salt
Tolerance in Higher Plants. Oxford and IBH Publishing Co., New Delhi.
Parida, A.K., Das, A.B. and Das, P. 2002. NaCl stress causes changes in photosynthetic
pigments, proteins and other metabolic components in the leaves of a true

233
mangrove, Bruguiera parviflora, in hydroponic cultures. J. Plant Biol., 45; 28–
36.
Park, J.A., Ahn, J.W.,Kim,Y.K., Kim, S.J., Kim, J.K., Kim, W.T. and Pai, H.S. 2005.
Retinoblastoma protein regulates cell proliferation, differentiation, and
endoreduplication in plants. Plant J., 42; 153–163.
Parvaiz, A. and Satyawati, S. 2008. Salt stress and phyto-biochemical responses of
plants-a review. Plant Soil Environment, 54 (3); 89-99.
Polit, J.T., Kaźmierczak, A. and Walczak-Drzewiecka, A. 2012. Cell cycle-dependent
phosphorylation of pRb-like proteinin root meristem cells of Vicia faba.
Protoplasma, 249; 131–137.
Popova, L.P., Stoinova, Z.G. and Maslenkova, L.T. 1995. Involvement of abscisic acid
in photosynthetic process in Hordeum vulgare L. during salinity stress. Plant
Growth Regul., 14; 211–218.
Pugnaire, F.I., Endolz, L.S. and Pardos, J. 1994. Constraints by water stress on plant
growth, in Handbook of Plant and Crop Stressed. Pessarakli M., editor. (New
York, NY: Marcel Dekker Inc; ), 247–259.
Quintero, F.J., Garciadeblas, B. and Rodriguez-Navarro, A. 1996. The SAL1 gene of
Arabidopsis, encoding an enzyme with 3’(2’),5’-bisphosphate nucleotidase and
inositol polyphosphate 1-phosphatase activities, increases salt tolerance in yeast.
Plant Cell, 8; 529–537.
Radziejwoski, A., Vlieghe, K., Lammens, T., Berckmans, B., Maes, S., Jansen, M.,
Knappe, C., Albert, A. Seidlitz, H.K., Bahnweg, G., Inze, D. and De Veylder, L.
2011. Atypical E2F activity coordinates PHR1 photolyase gene transcription
with endoreduplication onset. The EMBO Journal, 30; 355–363.
Rai, V., Vajpayee, P., Singh, S.N. and Mehrotra, S. 2004. Effect of chromium
accumulation on photosynthetic pigments, oxidative stress defense system,
nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L.
Plant Science, 167; 1159–1169.
Rai, V., Tandon, P.K. and Khatoon, S. 2014. Effect of Chromium on Antioxidant
Potential of Catharanthus roseus Varieties and Production of Their Anticancer
Alkaloids: Vincristine and Vinblastine. BioMed Research International. Volume
2014, Article ID 934182, 10 pages. http://dx.doi.org/10.1155/2014/934182.
Ramirez-Parra, E., Fründt, C. and Gutierrez, C. 2003. A genome-wide identification of
E2F-regulated genes in Arabidopsis. Plant J., 33; 801–811.
Ramirez-Parra, E., Lopez-Matas, M.A., Frundt, C. And Gutierrez C. 2004. Role of an
atypical E2F transcription factor in the control of Arabidopsis cell growth and
differentiation. Plant Cell, 16; 2350–2363.

234
Rıo, D.D., Stewart, A.J. and Pellegrını, N. 2005. A Review of Recent Studies on
Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress.
Nutrition, Metabolism & Cardiovascular Disease, 15; 316-328.
Roscoe, J.T. 1975. Fundemental research statistics for the behavioral sciences. New
York: Holt, Rinehart and Winston, Inc. 63.
Rossi, V., Locatelli, S., Lanzanova, C., Boniotti, M.B., Varotto, S., Pipal, A., Goralik-
Schramel, M., Lusser, A., Gatz, C., Gutierrez, C. and Motto, M. 2003. A maize
histone deacetylase and retinoblastoma-related protein physically interact and
cooperate in repressing gene transcription. Plant Molecular Biology, 51(3); 401-
413.
Rossignol, P., Stevens, R., Perennes, C., Jasinski, S., Cella, R., Tremousaygue, D. and
Bergounioux, C. 2002. AtE2F-a and AtDP-a, members of the E2F family of
transcription factors, induce Arabidopsis leaf cells to re-enter S phase. Molecular
Genetics and Genomics, 266; 995–1003.
Ruiz, J.M., Blasco, B., Rivero, R.M. and Romero, L. 2005. Nicotine-free and salt
tolerant tobacco plants obtained by grafting to salinityresistant rootstocks of
tomato. Physiol Plant 124; 465–475.
Sabelli, P.A., Liu, Y., Dante, R.A., Lizarraga, L.E., Nguyen, H.N., Brown, S.W.,
Klingler, J.P., Yu, J., LaBrant, E., Layton, T.M., Feldman, M. and Larkins, B.A.
2013. Control of cell proliferation, endoreduplication, cell size, and cell death by
the retinoblastoma-related pathway in maize endosperm. Proc Natl Acad Sci U S
A.110(19):E1827–36.
Santos, C.V. 2004. Regulation of chlorophyll biosynthesis and degradation by salt stress
in sunflower leaves. Sci. Hortic., 103; 93–99.
Scofield, S., Jones, A. and Murray, A.H.J. 2014. The plant cell cycle in context.
Journal of Experimental Botany, 65(10); 2557–2562.
Sears, R., Ohtani, K., and Nevins, J.R. 1997. Identification of positively and negatively
acting elements regulating expression of the E2F2 gene in response to cell
growth signals. Mol. Cell. Biol, 17; 5227–35.
Semelczi-Kovacs, A. 1975. Acclimatization and dissemination of the sunflower in
Europe. Acta. Ethnogr. Acad. Sci Hung., 24; 47-88.
Sergio, L., De Paola, A., Cantore, V., Pieralice, M., Cascarano, N.A., Bianco, V. V., Di
Venere, D. 2012. Effect of salt stress on growth parameters, enzymatic
antioxidant system, and lipid peroxidation in wild chicory (Cichorium intybus
L.) Acta Physiol Plant, 34; 2349–2358.

235
Shanker, A.K. and Pathmanabhan, G. 2004. Speciation dependant antioxidative
response in roots and leaves of Sorghum (Sorghum bicolor (L) Moench cv CO
27) under Cr(III) and Cr(VI) stress. Plant Soil., 265; 141– 51.
Shanker, A.K., Cervantes, C., Loza-Tavera, H. and Avudainayagam, S. 2005.
Chromium toxicity in plants. Environ Int., 31; 739–753.
Sharma, D.C., Chatterjee, C. and Sharma, C.P. 1995. Chromium accumulation by barley
seedlings (Hordeum vulgare L.). J Exp Bot., 25; 241–25.
Shinozaki, K. and Yamaguchi-Shinozaki K. 1996. Molecular responses to drought and
cold stress. Curr Opin Biotechnol., 7; 161-167.
Shipley, G.L. 2006. An introduction to real-time PCR. In: Real-Time PCR, Dorak M,.
T. (ed). Taylor and Francis Group, New York, 1-31.
Sinha, S., Saxena, R. and Singh, S. 2005. Chromium induced lipid peroxidation in the
plants of Pistia stratiotes L.: role of antioxidants and antioxidant enzymes.
Chemosphere, 58; 595–604.
Soleimanzadeh, H., Habibi, D., Ardakani, M.R. Paknejad, F. and Rejali, F. 2010. Effect
of Potassium Levels on Antioxidant Enzymes and Malondialdehyde Content
under Drought Stress in Sunflower (Helianthus annuus L.) American Journal of
Agricultural and Biological Sciences 5 (1); 56-61,ISSN 1557-4989.
Sozzani, R., Maggio, C., Giordo, R., Umana, E., Ascencio-Ibanez, J.T., Hanley-
Bowdoin, L., Bergounioux, C., Cella, R. and Albani, D. 2010. The E2FD/DEL2
factor is a component of a regulatory network controlling cell proliferation and
development in Arabidopsis. Plant Mol Biol., 72; 381–395.
Sönmez, F.K., Kömüscü, A.Ü., Erkan, A. and Turgu, E. 2005. An Analysis of Spatial
and Temporal Dimension of Drought Vulnerability in Turkey Using the
Standardized Precipitation Index. Natural Hazards, 35; 243–264.
Süzer, S. 2014. Ekolojik Ayçiçeği Tarımı. Trakya Tarımsal Araştırma Enstitüsü
Müdürlüğü Web sayfası: http://arastirma.tarim.gov.tr/ttae/Sayfalar/
Detay.aspx?SayfaId=56. Son erişim tarihi:11.09.2015.
Szabolcs, J. 1994. Soils and salinization. pp.3- 11. In: M. Pessarakli (Ed.), Handbook of
Plant and Crop Stress, Marcel Dekker, New York.
Theophanides, T. 1981. Fourier Transform Infrared Spectra of Calf Thymus DNA and
its Reactions with Anticancer Drug cis-platin”, Appl. Spectrosc., 35, 461.
Toledo-Ortiz, G., Huq, E. and Quail, P.H. 2003. The Arabidopsis basic/helix-loop-helix
transcription factor family. Plant Cell, 15; 1749–1770.
Vajpayee, P., Rai, U.N., Ali, M.B., Tripathi, R.D., Yadav, V., Sinha, S. 2001.
Chromium-Induced Physiologic Changes in Vallisneria spiralis L. and Its Role

236
in Phytoremediation of Tannery Effluent. Bulletin of Environmental
Contamination and Toxicology, 67(2); 246-256.
Van Verk, M.C., Gatz, C. and Linthorst, H.J.M. 2009. Transcriptional regulation of
plant defense responses. Adv. Bot. Res., 51; 397–438.
Vandepoele, K., Vlieghe, K., Florquin, K., Hennig, L., Beemster, G.T.S., Gruissem, W.,
Van de Peer, Y., Inze, D. and Lieven De Veylder , L., 2005. Genome-Wide
Identification of Potential Plant E2F Target Genes Plant Physiology, 139; 316–
328.
Vassilevska-Ivanova, R., Shtereva, L., Kraptchev, B. and Karceva, T. 2014. Response
of sunflower (Helianthus annuus L)cgenotypes to PEG-mediated water stress.
Cent. Eur. J. Biol., 9(12); 1206-1214.
Vlieghe, K., Vuylsteke, M., Florquin, K., Rombauts, S., Maes, S., Ormenese, S., Van
Hummelen, P., Van de Peer, Y., Inze, D. and De Veylder, L. 2003. Microarray
analysis of E2Fa-DPa-overexpressing plants reveals changes in the expression
levels of genes involved in DNA replication, cell wall biosynthesis, and nitrogen
assimilation. J Cell Sci., 116; 4249–4259.
Vuosku, J., Sutela, S., Kestilä, J., Jokela, A., Sarjala, T. and Häggman, H. 2015.
Expression of catalase and retinoblastoma-related protein genes associates with
cell death processes in Scots pine zygotic embryogenesis. BMC Plant Biology.
15; 88.
Wang, Y. and Nil, N. 2000. Changes in chlorophyll, ribulose biphosphate carboxylase-
oxygenase, glycine betaine content, photosynthesis and transpiration in
Amaranthus tricolor leaves during salt stress. J. Hortic. Sci. Biotechnol., 75;
623–627.
Wang, H., Xiao, X., Yang, M., Gao, Z., Zang, J., Fu, X. and Yin-hua Chen, Y., 2014.
Effects of salt stress on antioxidant defense system in the root of Kandelia
candel. Botanical Studies, 55-57.
West, G., Inze, D. and Beemster G.T.S. 2004. Cell Cycle Modulation in the Response of
the Primary Root of Arabidopsis to Salt Stress. Plant Physiology, 135; 1050–
1058.
Whiting, A.F. 1939. Ethnobotany of the Hopi. Museum of Northern Arizona Bulletin
No.15, Flagstaff: Northern Arizona Society of Science and Art.
Wildwater, M., Campilho, A., Perez-Perez, J. M., Heidstra, R., Blilou, I., Korthout, H.,
Chatterjee, J., Mariconti, L., Gruissem, W. and Scheres, B. 2005. The
RETINOBLASTOMA-RELATED gene regulates stem cell maintenance in
Arabidopsis roots. Cell, 123; 1337-1349.

237
Xiong, L. and Zhu, J.K. 2002. Molecular and genetic aspects of plant responses to
osmotic stress. Plant Cell Environ., 25; 131–139.
Yadav, V., Mallappa, C., Gangappa, S.N., Bhatia, S. and Chattopadhyay, S. 2005. A
basic helix-loop-helix transcription factor in Arabidopsis,MYC2, acts as a
repressor of blue light-mediated photomorphogenic growth. Plant Cell, 17;
1953–1966.
Yarsan, E. 1998. Lipid Peroksidasyon Olayı ve Önlenmesine Yönelik Uygulamalar. Y.
Y. U Vet. Fak. Derg., 9 (1-2); 89-95.
Yıldız, M. and Terzi, H. 2008. Effects of NaCl on protein profiles of tetraploid and
hexaploid wheat species and their diploid wild progenitors. Plant Soil Environ.
54; 227–233.
Yu, M.H. 2005. Environmental toxicology: Biological and health effects of pollutants.
CRC pres, Boca Raton.
Zayed, A.M. and Terry, N., 2003. Chromium in the Environment: Factors Affecting
Biological Remediation. Plant Soil, 249; 139-156.
Zeid, I.M., 2001. Responses of Phaseolus vulgaris to Chromium and Cobalt Treatments.
Biol. Plant., 44; 111-115.
Zhai, Q., Yan, L., Tan, D., Chen, R., Sun, J., Gao, L., Dong, M.Q., Wang, Y., and Li, C.
2013. Phosphorylation-coupled proteolysis of the transcription factor MYC2 is
important for jasmonate-signaled plant immunity. PLoS Genet. 9; e1003422.
doi: 10.1371/journal.pgen.

238
ÖZGEÇMİŞ
Adı Soyadı : Esin BAŞARAN
Doğum Yeri : Bulgaristan
Doğum Tarih : 01.06.1983
Medeni Hali : Bekar
Yabancı Dili : İngilizce
Eğitim Durumu (Kurum ve Yıl)
Lise : Yenimahalle Gaziçiftliği Lisesi (Yabancı Dil Ağırlıklı) (2001)
Lisans : Ankara Üniversitesi Fen Fakültesi Biyoloji Bölümü (2006)
Yüksek Lisans : Ankara Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
(Şubat 2007 – Şubat 2010 )
Çalıştığı Kurum/Kurumlar ve Yıl
Başkent Üniversitesi, 2012 – Devam
Yayınları (SCI ve diğer)
Uluslararası hakemli dergilerde yayımlanan makaleler
1. Vardar, Ç., Basaran, E., Cansaran-Duman, D. and Aras, S. 2014. Air-quality
biomonitoring: Assessment of genotoxicity of air pollution in the Province of Kayseri
(Central Anatolia) by use of the lichen Pseudevernia furfuracea (L.) Zopf and amplified
fragment-length polymorphism markers. Mutation Research/Genetic Toxicology and
Environmental Mutagenesis, 759; 43-50.
2. Soydam Aydın, S., Basaran, E., Cansaran Duman, D. and Aras, S. 2013. Genotoxic
effect of cadmium in okra seedlings: comperative investigation with population
parameters and molecular markers. Journal of Environmental Biology / Academy of
Environmental Biology, 34(6); 985-990.

239
Ulusal hakemli dergilerde yayımlanan makaleler
1. Basaran, E., Cansaran Duman, D., Büyük, İ. and Aras, S. 2015. Identification of
some Lecidea, Porpidia and Lecidella species (lichen-forming ascomycetes) distributed
in Turkey by sequence analysis of rDNA ITS region. Türk Hijyen ve Deneysel Biyoloji
Dergisi, 72(1); 45-58.
2. Basaran, E., Aras, S. ve Cansaran-Duman, D. 2010. Genomik, Proteomik,
Matabolomik Kavramlarına Genel Bakış ve Uygulama Alanları. Türk Hijyen ve
Deneysel Biyoloji Dergisi, 67(2); 85-96.
Kitaplar
Aras, S., Cansaran Duman, D., Vardar, Ç. and Basaran, E. 2011. Comparative Analysis
of Bioindicator and Genotoxicity Indicator Capacity of Lichens Exposed to Air
Pollution. In: Prof. Anca Moldoveanu editors. Air Pollution - New Developments.
China: InTech, 205-226.
Burslar
1. 2211 Tübitak Yurt İçi Doktora Bursu
10. 2010 – 02. 2015
2. Tübitak Proje Bursu
Proje No: 109T046
‘Kayseri İli Yöresinde Kirliliğe Maruz Bırakılmış Pseudevernia furfuracea (L.) Zopf.
Liken türünde Atomik Absorpsiyon Spektroskopisi ile Ağır Metal Analizi ve Hava
Kirliliğinin Genotoksisitesinin Moleküler Markörlerle Değerlendirilmesi.’
06. 2009 - 05. 2010







