
A functional evaluation of ansa cervicalis nervetransfer for unilateral vocal cord paralysis: Futuredirections for laryngeal reinnervationDAVID C. GREEN, MD, GERALD S. BERKE, MD, and MICHAEL C. GRAVES, MD, Los Angeles, California
There are a variety of methods for treating unilateral vocal cord paralysis. but to datethere are few objective studies that evaluate the functional results of nerve transferfrom the ansa cervlcalls. Six dogs underwent unilateral recurrent laryngeal nerve sec.tlon with Immediate reanastamosls to the sternothyroid branch of the ansa cervlcalls.After 5 to 6 months. measurements of vocal efficiency and acoustic parameters. vi.deolaryngoscopy. vldeostroboscopy. and evoked electromyography were performed.Identical measurements were made In eight control dogs during normal electricallyInduced phonation and a simulated unilateral recurrent laryngeal nerve paralysis.Histologic analysis of both vocalls muscles. recurrent laryngeal nerves. ansa cervlcalls.and the ansa·recurrent laryngeal nerve anastamosls site was performed. Evidence ofreinnervation was found In all of the animals that underwent nerve transfer. The vocalefficiency and acoustic quality after ansa cervlcalls nerve transfer were dependenton the degree of electrical stimUlation from the transferred nerve to the relnnervatedcord during phonation. In the absence of electrical stimulation to the nerve transfer.physiologic vocal cord motion could not be elicited from the relnnervated cord. (OTOLARYNGOL HEAD NECK SURG 1991;104:453.)
There are a variety of methods for treating unilateralvocal cord paralysis. These include Teflon injection, I
thyroplasty,' arytenoid adduction," and nerve" andnerve-muscle pedicle transfer.' A review of the literature reveals a lack of consensus regarding the optimalprocedure for voice improvement.6
Crumley et al. 4 have recommended nerve transferfrom the ansa hypoglossi to the recurrent laryngealnerve (RLN) as a treatment for unilateral vocal cordparalysis. This procedure was performed on five patients and the voice was thought to be superior to thatachieved with Teflon injection because of the restorationof normal stiffness, mass, and symmetry of the cord.Spectral analysis was improved postoperatively," andstroboscopic examination revealed synchronous mucosal waves. This technique required an open procedure, but did not expose or manipulate the larynx. Inaddition, Crumley et al." postulated that the denervated
From the Division of Head and Neck Surgery, UCLA School ofMedicine.
Supported by VA Merit Review Research Fund.Received for publication July 3D, 1990; accepted Aug. 29, 1990.Reprint requests: David C. Green, MD. 10992 Ashton Ave.,
Apt. 304, Los Angeles, CA 90024.23/1125135
sternothyroid muscle medialized the thyroid lamina andvocal cord. Tucker has also described a technique forlaryngeal reinnervation that uses an ansa hypoglossineuromuscular pedicle passed through a window in thethyroid cartilage.
The literature on laryngeal reinnervation is extensive.I An excellent review of the subject was recently pub
lished by Rice." The idea goes back to 1909, whenHorsley" first reported the repair of a recurrent laryngealnerve after transection by a gunshot wound. He reportedcomplete return of function in 3 months. In 1926, Blalock and Crowe" described some return of motion without atrophy of the cord after anastamosis of the distalsegment of the RLN to either the ansa hypoglossi (endto-end), the phrenic, or into a slit in the opposite RLN.Histologic studies demonstrated regrowth of axons afteranastarnosis in all three cases. Frazier and Mosser'!anastomosed the RLN to the ansa hypoglossi in 10patients with paralysis present from 8 months to 11years. They found improvement in 60% overall but littleimprovement in patients with paralysis for more than6 years,
In contrast to these early encouraging results, Hoover," in 1953, stated that attempts at restoration ofrecurrent laryngeal function by nerve anastamosis hadbeen unsuccessful. This situation was clarified in 1963
453

... GREENeta!.
by Siribodi et al.," who found a return of electromyographic activity after repair of the laryngeal nerve, butno functional movement. They proposed that aberrantregeneration of adductor and abductor fibers accountedfor the lack of function. This finding is now termedlaryngeal synkinesis."
Although Doyle et al. IS reported almost completereturn of function after anastamosis of the RLN in bothdogs and human beings, most reports have been consistent with the findings of Siribodi et al." Gordon andMcCabel6 found that after section and immediate repairof the RLN, only adductor function returned. Theyattributed this to the fact that adductor muscles outnumbered abductor muscles. Boles and Fritzell 17 foundpolyphasic potentials by means of electromyography(EMG) 3 to 6 months after RLN section and anastamosis in dogs with little motion in the reinnervatedcord. In contrast, in the crush injuries they studied,recovery took place in approximately 2 months withnormal cord movement. They concluded that surgicalrepair of the RLN could possibly return tone and someadduction to the cord and improve the glottis as a phonator and a protective sphincter. Tashiro" found EMOevidence of recovery but no definite motion after repairof the RLN. Murakami and Kirchner" confirmed thepresence of simultaneous activation of the adductorsand abductors after repair of the RLN by EMO. Thissupported the idea of misdirection in axon regrowth.Satoh et al. 1O performed direct muscle stimulation andEMO studies and thought that impaired muscle function, in addition to axon misdirection, contributed topoor function of the reinnervated laryngeal muscles.Ded021 found a small degree of adduction in the reinnervated cord after RLN anastamosis. He believed theadduction resulted from the action of the cricothyroidmuscle.
As reinnervation of the larynx began to emerge as apotential treatment for unilateral vocal cord paralysis,controversy arose as to which nerve transfer would givethe best results. Crumley et al." recommended theansa cervicalis branch to the sternothyroid branch forseveral reasons. This branch to the sternothyroid is located very near the RLN. Although the pattern of sternothyroid activation is predominantly inspiratory, itfires throughout the respiratory cycle, giving a certaindegree of continuous tone to the vocalis. Also, it isthought that the subsequent paralysis of the sternothyroid branch relaxes tension on the thyroid cartilageand further medializes the paralyzed cord. However,some investigators have argued against the use of thesternothyroid branch because the pattern of activation
OtoIaryngoJogyHead and Neck SUrgery
from this nerve may result in cord adduction duringinspiration rather than phonation." Marie et al. 23 found,after ansa-adductor branch transfer in the canine model,that during extended obstructive dyspnea, there wasadduction of the cord, which was abolished after sectioning of the ansa. In this same canine model, thereinnervated cord failed to reach midline during whining, demonstrating a lack of ansa stimulation duringphonation.
Crumley" has advised against using the ansa'sbranch to the thyrohyoid despite its being the mostactive strap muscle during phonation, with peak activityduring expiration. His reluctance stems from the requirement of a cable graft to reach the RLN stump andthe resulting paralysis of the thyrohyoid, eliminatingthe adductor function of this muscle.
The superior laryngeal nerve (SLN) has been proposed by Rice" to be ideal for nerve transfer to improvephonation after a unilateral vocal fold paralysis. Thisnerve is active during phonation and anatomically closeto the RLN. Woodsen2S found that during normal respiration in human beings, the cricothyroid muscle isactive predominantly during inspiration. This patternofactivation may be ofconcern because it is conjecturedthat inspiratory activation of the thyroarytenoid maycause airway obstruction. Rice" and Crumley" haveperformed a SLN-adductor branch anastamosis in dogs,which demonstrated spontaneous adduction of the cordwith coughing or whining. However, Crumley" believes that use of the superior laryngeal nerve is "robbing Peter to pay Paul" in that this would eliminate thenormal adductive effect of the cricothyroid muscle.
Crumley" has also counseled against the use of theproximal RLN in RLN-RLN anastamosis. His concernsare again related to the possibility ofairway obstructionduring inspiration as a result of hypertrophy and medialbulging of the thyroarytenoid from RLN synkinesis. 13
Although Tashiro," Sirbodhi et al., 13 Murakami andKirchner," Sato et al.,20 Veda et al.," and Iwamura"have shown activation of the adductors during inspiration, none of these studies has demonstrated respiratory obstruction with dyspnea from a unilateral RLNRLN anastamosis in the presence of a normal oppositecord. Iwamura" stated "such minor paradoxical adduction of the reinnervated vocal cord appears to be ofno great importance, since the inspiratory adductionlasts for only a fraction of a second." As mentionedpreviously. the RLN·RLN anastamosis may not be theonly "offender" in this regard. Marie et aI.23 reportedparadoxical inspiratory adduction of the reinnervatedcord by means of the ansa branch to the sternothyroid.

Volume 104 Number 4April 1991
A recent" clinical case report of a unilateral RLN-RLNanastamosis resulted in an excellent voice without airway obstruction.
Another issue is the duration of time that a nervetransfer can still be used after vocal cord paralysis.Janecka," by means of the rat siatic nerve, found thatsevered axons will not regrow when the nerve segmenthas lost contact with the target muscle. How long denervated muscle can provide this trophic response isnot known. Some authors obtain an EMG from anypatient who has been paralyzed for more than 2 years.Patients with absence of muscle activity on a thyroarytenoid EMG are then treated with Teflon injection.
Although the voice after nerve transfer has been reported as excellent, there are some disadvantages. Thepatient must undergo general anesthesia and an opensurgical procedure. An intact distal stump of the RLNmust be present. The patient must wait approximately3 to 4 months before there is improvement from theprocedure. Gelfoam injection into the paralyzed cordmay improve glottal closure during the reinnervationperiod.
Few reports have used objective voice measurementsto study the results of nerve transfer for unilateral paralysis of the vocal folds. Vocal efficiency is an objective measure of the voice that was first studied by vanden Berg" in 1956. He defined the efficiency of voiceas the ratio of the acoustic power of the voice to theSUbglottic power. The subglottic air power can be estimated as the product of the mean glottic air flow rateand the subglottic pressure. Clinically, vocal efficiencyhas been shown to decrease with some forms of laryngeal disease, such as invasive carcinoma and vocal cordparalysis.F-" Ueda et al. 27 studied acoustic intensity.subglottic pressure, and flow rate during induced whining in dogs after a combination SLN-SLN and RLNRLN anastamosis. The vocal efficiency after reinnervation in this canine study was significantly less thannormal.
Although vocal efficiency is useful as a functionalmeasure of voice, it may not correspond with vocalquality. The voice may be quite harsh with a normalvocal efficiency. Also. vocal efficiency does not indicate the degree of control the patient has over theglottis. 34
To measure voice quality, a number of acoustic measures have been used clinically. These include jitter.shimmer. and signal-to-noise ratio. Jitter is defined asthe fluctuation in the time interval between successivepeaks of the fundamental frequency. Shimmer is thecycle-to-cycle variation in the amplitudes of the peaks.
Future directions for laryngeal reinnervation 411
Signal-to-noise ratio is the ratio of the sound energyin the acoustic signal to the background noise.35.36
Lieberman" was the first to report an increased jitterin pathologic phonation, and Zyski et al.38 found increased jitter and shimmer in patients with laryngealtumors and unilateral vocal cord paralysis. Efforts havebeen made to use these measures as screening devicesfor pathologic conditions of the larynx" and to document the results of laryngeal surgery."
This study used an in vivo canine model to evaluatethe functional results of nerve transfer from the ansacervicalis to the RLN as a treatment for unilateralvocal cord paralysis. Measurements of vocal efficiency, acoustic analysis, videolaryngoscopy, videostroboscopy, and evoked EMG were performed afteransa cervicalis nerve transfer and compared with a control group of animals during normal phonation and asimulated RLN paralysis. Tissue samples were takenfrom both vocalis muscles, RLNs, ansa cervicalis, andthe ansa-RLN anastamosis site for histologic analysis.
METHODS AND MATERIALSExperimental Design
The experimental group consisted of six mongreldogs (25 kg each) that underwent a unilateral RLNsection with immediate reanastamosis to the sternothyroid branch of the ansa cervicalis under general anesthesia. One dog had a reaction to the anesthetic anddied before the evaluation. After 5 to 6 months, measurements for vocal efficiency and acoustic analysis(jitter, shimmer, and signal-to-noise ratio) were madeon the remaining five experimental dogs, in addition tovideolaryngoscopy and videostroboscopy. In the experimental group, vocal efficiency and acoustic measures were made, with and without electrical stimulation of the ansa cervicalis nerve transfer. In addition,measurements of vocal efficiency and acoustic analysis(jitter, shimmer, and signal-to-noise ratio) were madeon a group of eight control dogs during "normal" electrically stimulated phonation and a simulated RLN paralysis. In the experimental group, evoked EMG wasperformed to measure the nerve conduction velocitiesand the response amplitudes of the normal and reinnervated vocal folds. Tissue samples were taken of bothvocalis muscles, RLNs, ansa cervicalis, and the ansaRLN anastamosis site for histologic comparison.
Surgical Technique In the Experimental GroupEach animal was premedicated with acepromazine
and then underwent general anesthesia with endotracheal intubation. The ventral neck was shaved and pre-

.Q6 GREEN at oJ.
Vent ,lotor~
"'IICROPHONE
SOUND LEVEL METER
"'IULTICHANNElSTORAGEOSCILLOSCOPE
PRESSURETRANSDUCER
OtolaryngologyHead and Neck Surgery
DIREC TCOMPUTER IZEDDIGITIZ ATION
.r-:-J\-
Fig. 1. Schematic representation of experimental setup for In vivo canine model of phonatlon otteransa cervlcalls nerve transfer.
pared with povidone iodine. The animal was placed inthe supine position on a table in the operating room,and a midline neck incision was made from the midthyroid cartilage to 4 cm below the cricoid. The dissection was carried down to the trachea, and theRLN and the ansa cervicalis (ansa) were identified bilaterally. On the basis of the proximity of the ansa andthe RLN, one side was chosen for reanastamosis . Atapproximately 5 cm from the cricothyroid joint, theRLN was sectioned sharply with a no. 15 blade. Theproximal end of the RLN was ligated with 2-0 silksutures. The distal end was then anastamosed end-toend to the ansa cervicalis. The anastamosis was performed with four 10-0 nylon sutures according to standard microsurgical technique . The site of the ansa-RLNanastomosis was marked with a loose 2-0 silk sutureplaced I em away from the anastamosis . The woundwas closed in three layers with 3-0 VicryI. Six monthspostoperatively, the laryngeal phonatory characteristicswere evaluated by means of the in vivo canine modelof phonation ." Humane animal care was assured incompliance with The Principles of Laboratory AnimalCare. formulated by the National Society for MedicalResearch, and the Guide for the Care and Use of Lab-
oratory Animals. prepared by the National Academy ofSciences and published by the National Institutes ofHealth (NIH publication No. 80-23, revised 1978).
In Vivo Canine Phonation Model
Each dog in the control and experimental groups waspremedicated with intramuscular acepromazine. Intravenous thiopental was administered to a level of cornealanesthesia , and additional thiopental was used tomaintain this level of anesthes ia throughout the experiment.
The animal was placed in the supine position on theoperating table, and a midline incision was made toexpose the trachea from the hyoid to the sternal notch(Fig . I). In the animals undergoing experimental nervetransfer, the RLN on one side and the RLN-ansa anastamosis on the opposite side were identified and preserved. In the control group of animals, both RLNswere identified. For both control and experimentalgroups, the superior laryngeal nerves were identifiedalong their course to the cricothyroid muscles . A lowtracheotomy was performed at the level of the suprasternal notch, through which an endotracheal tube waspassed to allow ventilator-assisted respiration. A second

Volume 104 Number 4April 1991
tracheotomy was performed in a more superior location,through which a cuffed endotracheal tube was passedin a rostral direction and positioned with the tip 10 embelow the vocal folds. The cuff was inflated to just sealthe trachea. Humidified heated air was passed throughthis rostral endotracheal tube from a compressed airtank. Flow was controlled with a valve and measuredwith a Gilmont flowmeter (model FI500, Gilmont Instruments, Great Neck, N.Y.). The air flow was humidified and heated by bubbling it through 5 em ofheated water so that the temperature of the air was37° C when measured at the glottic outlet. A l-cmbutton was used to suspend the epiglottis from a fixedpoint to provide direct visualization of the larynxthrough the oral cavity.
A l-cm segment of each superior laryngeal nerve(external branch) was isolated, and Harvard miniatureelectrodes (Harvard Apparatus Inc., Millis, Mass.)were applied around each nerve. Harvard electrodeswere also applied to the RLN on one side and the ansacervicalis on the opposite side in the animals undergoing nerve transfer or both RLNs in the control animals. Care was taken to place the electrode on the ansacervicalis 2 em proximal to the anastamosis site in theanimals undergoing nerve transfer, to avoid currentspread to the distal RLN. The electrodes were theninsulated from surrounding tissue. The cut proximalRLN on the side of the anastamosis in the animalsundergoing nerve transfer was also dissected out andstimulated with a supramaximal current of 5 mA toverify an absence of vocal cord movement or EMGstimulation. Two Model S2LH constant-current nervestimulators (WR Medical Electronics Co., St. Paul,Minn.) were used to stimulate the RLN, ansa cervicalis,and superior laryngeal nerves independently. Thesenerves were stimulated at 70 to 80 Hz stimulus frequency, with 0.5 to 2.0 mA intensity for 1.5-msecduration. Phonation was produced with an air flow of318 to 523 cc/sec, applied through the larynx by therostral endotracheal tube.
Acoustic MeasuresAcoustic measurements were obtained for the ex
perimental nerve transfer group, both with and withoutstimulation of the ansa nerve transfer. Acoustic measurements for the control group were obtained duringsimulated normal phonation and RLN paralysis. Normal phonation required stimulation of all four laryngealnerves. A unilateral RLN paralysis was simulated bynot stimulating one RLN. The sound level measurements were made with a I-inch Quest condenser microphone placed 30 em from and level with the glotticoutlet. The microphone was directed 90 degrees from
Future directions for laryngeal reinnervation ••7
the direction of the sound source. The sound level measurements were made in decibels, with a Quest soundlevel meter on the C scale. The acoustic signal was alsodigitized, after C scale filtering at 20 kHz, and storedon the hard disk of a personal computer.
Subglottic pressure was measured with a MillarMikro-Tip catheter pressure transducer (model No.SPC-330, size 3F, Millar Instruments, Inc., Houston,Texas) passed rostrally through the superior tracheotomy. It was placed 5 em below the glottis. This signalwas low-pass filtered at 3 kHz, digitized at 20 kHz,and stored in a personal computer. Because of the variation in subglottic pressure during phonation, the peakpressures attained during the glottic cycle were used.These peaks were identified by means of a commercially available software package for the PC system("C-Speech," Paul Milenkovic, University of Wisconsin, Madison, Wis.).36 The pressure transducer was calibrated before each experiment against a mercury manometer at 3r C.
Vocal efficiency was calculated as the ratio of theacoustic power of the voice to the subglottic power.The total acoustic power was calculated according tothe method of Koyama et al.," in which total soundpower = 2 rPc
2/Poc. This formula applies for a sound
power radiating with no known direction into ahemisphere of area (2 r'), a distance (r) away fromthe source. The product Po (the density of the medium)and c (the velocity of propagation) is the specificacoustic impedance of the medium, which is 41.1dynes/sec/em' in air at 200 C. The term P, is the rootmean square sound pressure in dynes/em' at the distance r from the sound source. The subglottic powerwas calculated as the product of the flow rate times thepeak subglottic pressure.
Acoustic analysis of the digitized acoustic signal wasperformed with a commercial software program ("CSpeech," Paul Milenkovic, University of Wisconsin,Madison, Wis.). Jitter, shimmer, and signal-to-noiseratio were calculated on approximately 0.3 second ofstable phonation from each trial. The background noisein the experimental quarters was 35 dB lower than thevalues measured with the C scale. To normalize forvarying fundamental frequency, jitter was calculated asa fraction of the period of the fundamental frequency.
For each dog in either the experimental or the controlgroup, the jitter, shimmer, signal-to-noise ratio, andvocal efficiency for that dog were calculated from themean of five trials of phonation. A mean value for theentire experimental group, with and without ansa stimulation for jitter, shimmer, signal-to-noise ratio, andvocal efficiency, was calculated by taking the averageacross all five experimental dogs. A mean value for the

.51 GREEN et 01.
control group during simulated normal phonation andRLN paralysis for jitter, shimmer, signal-to-noise ratio,and vocal efficiency was calculated by taking the average across all eight control dogs.
VldeoltrobolcoPY and VldeolaryngolcoPY
Videolaryngoscopy was performed during singlenerve stimulation of the RLN on the normal side oransa cervicaJis on the nerve transfer side. The currentstimulus was increased, starting from zero, to observethe effect of stimulation from each nerve separately.The image was detected by a Jed-Med CCD (chargecoupled device) video camera (Model 70-5110, JedMed, St. Louis, Mo.) and a Sony V-matic videocassetterecorder (VO-5850. Park Ridge. N.J.). Videolaryngoscopy was also performed under a lighter plane ofanesthesia to detect the degree of spontaneous motionof the cords during laryngeal stimulation with a cottonswab and obstructive respiratory dyspnea.
Videostroboscopy was performed, both with andwithout stimulation of the ansa cervicalis, in the animals undergoing nerve transfer, and with and withoutstimulation of one RLN in the normal control animals,while the other three laryngeal nerves were beingstimulated. For stroboscopic imaging of the larynx.a Bruel & Kjaer laryngostrobe unit (model 4914,Bruel & Kjaer, Orange. Calif.) was used. The stroboscope was connected to a Storz zero-degree telescopevia a fluid-filled light cable. The video images wereanalyzed frame-by-frame with the videorecordingunit.
Evoked EMG
Conventional spontaneous EMG recordings were initially made during anesthesia. to detect fibrillation potentials in the thyroarytenoid muscle. Evoked EMG wasthen performed in the nerve transfer experimental animals by stimulating the ansa cervicalis nerve transferor the normal RLN on the opposite side with a single0.5 msec pulse of sufficient voltage through a Harvardminiature electrode. to obtain an adequate response.The response was detected with a bipolar needle placedtransorally into the ipsilateral thyroarytenoid muscle.The signal was digitized at 20 kHz and stored in apersonal computer. A conduction velocity for eachnerve was calculated by measuring the latency of theresponse in milliseconds between two points along thenerve. The time difference in the latency between thestimulated points divided by the distance between thesepoints gave the conduction velocity between thesepoints. In the experimental animals, the conduction velocity was measured on the normal side between two
OtolaryngologyHead and Neck Surgery
points along the distal RLN. On the nerve transfer side,conduction velocity was measured between a pointproximal and distal to the ansa-Rl.N anastamosis. Inaddition, the amplitude of the evoked EMG was recorded in millivolts .as a measure of the amount ofdepolarizing muscle.
Histologic AnaIY.'1
The larynx of each animal undergoing nerve transferwas removed. divided into right and left halves, andplaced in 10% .formalin. A 3-mm thick cross-sectionof each vocalis muscle was taken midway between theanterior commissure and the tip of the vocalis processfor staining with hematoxylin and eosin.
A l-cm segment of the RLN was removed at a siteI cm proximal to the cricothyroid joint bilaterally andat 5 cm proximal to the cricothyroid joint on the sidenot operated on. A l-cm segment of ansa cervicaliswas taken I em proximal to the anastamosis on the sideoperated on and at a similar site on the ansa of the sidenot operated on. The site of the ansa-RLN anastamosiswas also removed. All five nerve specimens were fixedin 4% paraformaldehyde-0.4% glutaraldehyde. Theanastamosis site was embedded in parlodion and sectioned longitudinally. The remaining specimens werestained with osmium. embedded in Eponl Araldite(Electron Microscopy Sciences, Fort Washington, Pa.)and thin sections (0.5 "",m) were cut transversely.
Statistical Ana'y."
A paired t test was used to compare mean changesfrom ansa stimulation to nonstimulation within the experimental group (N = 5) for vocal efficiency, jitter,shimmer, and signal-to-noise ratio. Two group t testswere used to compare the mean difference in thesemeasurements between the experimental group and thecontrol group (N = 8). For the vocal efficiency measurement, the t test was computed on the log values.Between-dog comparisons by means of Mann-Whitneyand Wilcoxon rank-sum criteria instead of the t test didnot change any conclusions.
RESULTSNerve and Vocal Cord Histologic Findings
There was little sign of atrophy in the reinnervatedthyroarytenoid muscle when compared to the normalside. The average diameter of both the reinnervated andnormal thyroarytenoid muscles was identical at 7.3 mm.Figure 2 shows the lack of atrophy in the reinnervatedcord when compared to normal.
Examination of the RLN on the side of the nervetransfer showed regenerated axons as seen in Fig. 3.

Volume 104 Number 4April 1991
A
B
Future directions for laryngeal reinnervation 419
Fig. 2. Cross section of A, normal cord and B, reinnervated cord.
VldeolaryngolcoPY of Groll Movement
Increasing electrical stimulation of the normal RLNdemonstrated increasing ipsilateral vocalis musclebulging and arytenoid adduction in all of the dogs.The threshold for vocalis contraction on the normal sidewas between 0.05 and 0.19 rnA. With increasing stimulation of the ansa cervicalis on the side of the nervetransfer, increasing ipsilateral vocalis contraction wasseen in all dogs. In four of the dogs, this vocalis contraction was associated with arytenoid adduction, but
in one animal there was distinct arytenoid abductionwith vocalis contraction. The threshold for vocalis contraction from ansa stimulation was between 0.01 and0.2 rnA.
During obstructive respiratory dyspnea, under a lightplane of anesthesia, after sectioning both superior laryngeal nerves, the normal cord was found to abductwith inspiratory efforts in all dogs. The reinnervatedcord did not adduct or abduct to any degree during thistime in any subject.

<t6O GREEN at 01.
A
,
r
otolaryngologyHead and Neck Surgery
B
Fig. 3. Cross sectionof A, normal RLN and B, reinnervated RLN.
-
During stimulation of coughing under a light planeof anesthesia. the normal cord adducted but the reinnervated cord showed no gross movement in any of theanimals.
VldeoltrobolCoPY
The results of videostroboscopy are summarized inTable I. Videostroboscopy of normal phonation in thecontrol animals showed glottal dynamics similar to

Volume 104 Number 4April1991 Future directions for laryngeal reinnervation 461
Normal Cord
Reinnervated Cord
25
20
15
mV10
5
o2 3 4
Dog Number5 Mean
Fig. 4. Amplitude of response to evoked EMG in millivolts.
those of phonation in the animals undergoing nervetransfer during stimulation of the ansa cervicalis nervetransfer. There was complete glottic closure, two-mass(upper and lower margin) motion of the mucosa on bothcords, and complete glottic symmetry. Videostroboscopy without stimulation of the nerve transfer in theanimals undergoing nerve transfer resulted in a picturesimilar to that of a simulated unilateral vocal cord paralysis. There was a mild posterior glottic chink incompetence and one-mass motion of the paralyzed cord.Without stimulation of the ansa cervicalis nerve transfer, the reinnervated cord stroboscopically appearedto track greater lateral excursions on opening thanthe normal cord, which remained primarily in themidline.
Evoked EMGand Conduction Velocity
At 5 to 6 months after RLN section and nerve transfer, no fibrillation potentials were seen in any of thereinnervated vocal cords, indicating some degree ofreinnervation in all subjects. Figure 4 shows the amplitude in millivolts of the evoked EMG. The amplitudeof the response of the reinnervated side was between5% and 155% of the normal side with an average of66% for the five subjects. By measuring the delay inthe EMG response to the evoked stimuli at two separatestimulation points on the nerve, a conduction velocitywas obtained for the normal and nerve transfer side.Table 2 shows these conduction velocities. For the normal side, the conduction velocity varied from 34.6 to50 m/sec, with an average of 42.2 m/sec. For thereinnervated side, the conduction velocity varied from
Table 1. Summary of stroboscopic analysis
Glottic Mucosal CordAnimal group closure motion motion
Control groupNormal Complete Two-mass Symmetric
bilaterallyParalysis Incomplete; One-mass Asymmetric
posterior paralyzedglottic cordleak
Nerve transfergroupAnsa stimu- Complete Two-mass Symmetric
lated bilaterallyAnsa not Incomplete; One-mass Asymmetric
stimu- posterior paralyzedlated glottic cord
leak
Table 2. Conduction veloclfy (m/sec):
Experimental group
Normal Ansa cervlcaUsDog RLN nerve tranafer
1 50.0 18.0
2 34.6 3563 42.9 13.04 37.6 37.5
5 45.8 50.0
MEAN 42.2 30.8
SD 6.2 151
RLN. Recurrent laryngeal nerve.
462 GREEN et al.
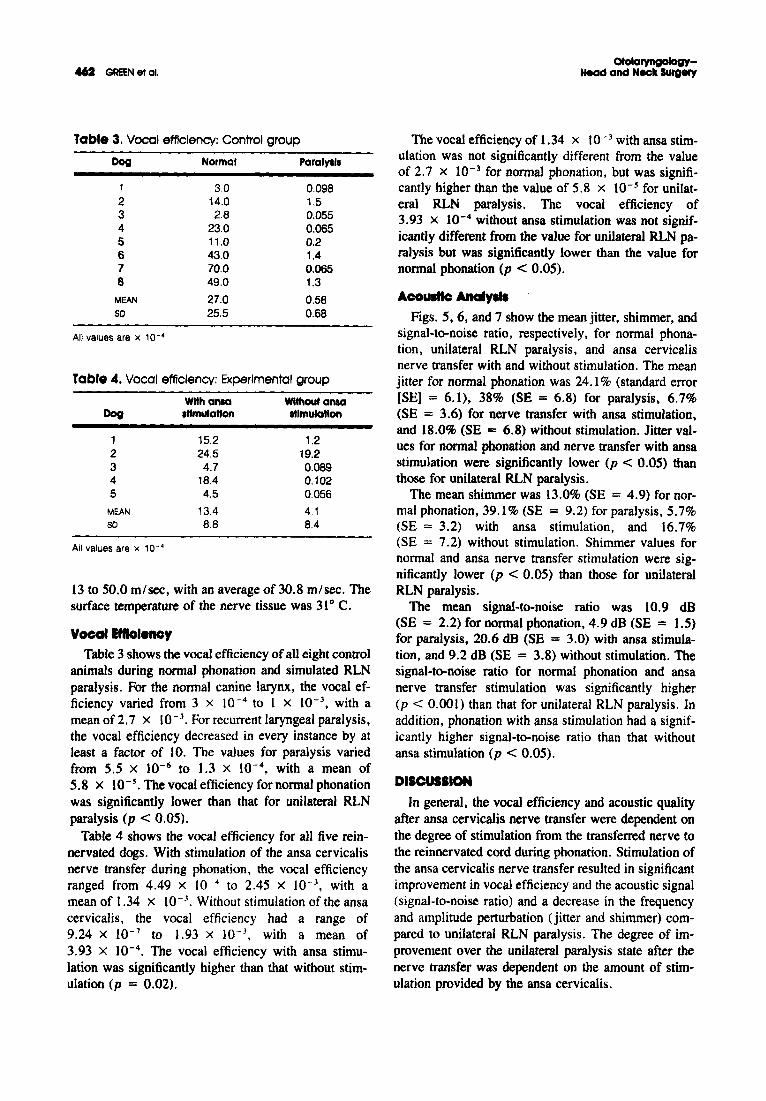
Tabl. 3. Vocal efficiency: Control group
Dog Normal Paralyal.
1 30 0.0982 14.0 1.53 2.8 0.0554 23.0 0,0655 11.0 0.26 43.0 1,47 70.0 0,0658 49.0 1.3
MEAN 27.0 0,58so 25,S 0.68
All values are x 10- 4
Tabl. 4. Vocal efficiency: Experimental group
Wllhansa Without ansaDog Illmulallon .tlmulatlon
1 15.2 1.22 24.5 19.23 4.7 0.0894 18.4 0.1025 4.5 0.056
MEAN 13.4 4.1so 8.8 8.4
All values are x 10- 4
13 to 50.0 m/sec, with an average of 30.8 m/sec. Thesurface temperature of the nerve tissue was 31° C.
YocallfftolenoyTable 3 shows the vocal efficiency of all eight control
animals during normal phonation and simulated RLNparalysis. For the normal canine larynx, the vocal efficiency varied from 3 x 10- 4 to I x 10-3, with amean of2.7 x 10- 3
• For recurrent laryngeal paralysis,the vocal efficiency decreased in every instance by atleast a factor of 10. The values for paralysis variedfrom 5.5 x 10- 6 to 1.3 x 10- 4
, with a mean of5.8 x 10- s. The vocal efficiency for normal phonationwas significantly lower than that for unilateral RLNparalysis (p < 0.05).
Table 4 shows the vocal efficiency for all five reinnervated dogs. With stimulation of the ansa cervicalisnerve transfer during phonation, the vocal efficiencyranged from 4.49 x 10- 4 to 2.45 X 10- 3
, with amean of 1.34 x 10- 3
• Without stimulation of the ansacervicalis, the vocal efficiency had a range of9.24 x 10- 7 to 1.93 X 10- 3
, with a mean of3.93 x 10- 4 • The vocal efficiency with ansa stimulation was significantly higher than that without stimulation (p = 0.02).
OtolaryngologyHead and Neck SUrgery
The vocal efficiency of 1.34 x 10-3 with ansa stimulation was not significantly different from the valueof 2.7 x 10-3 for normal phonation, but was significantly higher than the value of 5.8 x lO- s for unilateral RLN paralysis. The vocal efficiency of3.93 x 10-4 without ansa stimulation was not significantly different from the value for unilateral RLN paralysis but was significantly lower than the value fornormal phonation (p < 0.05).
Acoustic Analy."Figs. 5, 6, and 7 show the mean jitter, shimmer, and
signal-to-noise ratio, respectively, for normal phonation, unilateral RLN paralysis, and ansa cervicalisnerve transfer with and without stimulation. The meanjitter for normal phonation was 24.1% (standard error[SE] = 6.1), 38% (SE = 6.8) for paralysis, 6.7%(SE = 3.6) for nerve transfer with ansa stimulation,and 18.0% (SE = 6.8) without stimulation. Jitter values for normal phonation and nerve transfer with ansastimulation were significantly lower (p < 0.05) thanthose for unilateral RLN paralysis.
The mean shimmer was 13.0% (SE = 4.9) for normal phonation, 39.1 % (SE = 9.2) for paralysis, 5.7%(SE = 3.2) with ansa stimulation, and 16.7%(SE = 7.2) without stimulation. Shimmer values fornormal and ansa nerve transfer stimulation were significantly lower (p < 0.05) than those for unilateralRLN paralysis.
The mean signal-to-noise ratio was 10.9 dB(SE = 2.2) for normal phonation, 4.9 dB (SE = 1.5)for paralysis, 20.6 dB (SE = 3.0) with ansa stimulation, and 9.2 dB (SE = 3.8) without stimulation. Thesignal-to-noise ratio for normal phonation and ansanerve transfer stimulation was significantly higher(p < 0.001) than that for unilateral RLN paralysis. Inaddition, phonation with ansa stimulation had a significantly higher signal-to-noise ratio than that withoutansa stimulation (p < 0.05).
DISCUSSION
In general, the vocal efficiency and acoustic qualityafter ansa cervicalis nerve transfer were dependent onthe degree of stimulation from the transferred nerve tothe reinnervated cord during phonation. Stimulation ofthe ansa cervicalis nerve transfer resulted in significantimprovement in vocal efficiency and the acoustic signal(signal-to-noise ratio) and a decrease in the frequencyand amplitude perturbation (jitter and shimmer) compared to unilateral RLN paralysis. The degree of improvement over the unilateral paralysis state after thenerve transfer was dependent on the amount of stimulation provided by the ansa cervicalis.

Volume 104 Number 4April 1991 Future directions for laryngeal reinnervation 463
--i
Without AnsaStimulation
With AnsaStimulation
40
35
30
25
% 20
15
10
5
0 --+-
Normal Paralysis
Fig . 5. Mean jitter for norma l phonation and unilatera l RLN paralysis in control group (N = 8). andwith and without stimulation of ansa cervica lis nerve transfer in experimental group (N = 5).
Normal ParalysisWith AnsaStimulation
Without AnsaStimulatio n
Fig. 6. Mean shimmer for normal phonation and unilaterol RLN paralysis In control group (N = 8).and with and without stimulation of ansa cervlcous nerve transfer In experimental group (N = 5).
It is generally accepted that the bulk of the vocalcord adductor muscles outweighs that of the abductors.For this reason , random synkinesis usually results inadduction during stimulation of the reinnervated nerve.In four of the reinnervated dogs, stimulation of the ansacervicalis nerve transfer produced vocalis contractionand arytenoid adduction ; however, one dog demonstrated vocalis contraction and arytenoid abduct ion.During phonation in the dog with the arytenoid abduction, the normal cord and arytenoid crossed the midlineto meet the reinnervated cord, thereby completing glottic closure during laryngeal nerve stimulation . AI-
though the posterior glottis was shifted to the reinnervated side in this dog, there was symmetric vocal iscontraction and two-mass mucosal motion in bothcords. It is interesting to speculate what pattern of reinnervation led to this abduction of the cord with vocaliscontraction . The evoked thyroarytenoid EMG on thereinnervated side in this dog was among the lowest. at30% of the normal side. This finding lends support tothe notion that there was a relative lack of reinnervationof the lateral cricoarytenoid and vocalis and strong reinnervation of the posterior cricoarytenoid. This findingof cord abduction with reinnervation of the RLN stump

... GREEN at al.
25
20
15
db10
5
0
Normal ParalysisWith AnsaStimulation
OfolaryngologyHead and Neck SUrgery
--i
Without AnsaStimulation
Fig. 7. Mean signal-tD-noise ratio for normal phonation. unilateral RLN paralysis in control group(N = 8). and with and without stimulation of ansa cervicalis nerve transfer in experimental group(N = 5).
by the ansa cervicalis has not been previously describedin human beings.
During stimulation of all the laryngeal nervesincluding the ansa cervicalis nerve transfervideostroboscopy appeared normal in all five of thereinnervated dogs with complete glottic closure, twomass mucosal motion, and symmetry. Absence of nervetransfer stimulation during stimulation of the other threelaryngeal nerves provided the stroboscopic picture ofan acute unilateral RLN paralysis with a mild posteriorglottic chink incompetence, one-mass motion of theparalyzed cord, and greater lateral excursions of thereinnervated cord than the normal cord.
In the past, other studies have relied on the isolatedpresence of vocal cord mobility on laryngoscopy orEMG evidence to prove reinnervation. However, theseprocedures may not be sufficient evidence of reinnervation because of-the action of the other intrinsic andextrinsic laryngeal muscles and the ability of neighboring nerves to reinnervate adjacent paralyzed muscles." This study used the combination of evoked EMGfrom direct stimulation of the transferred nerve, observed motion from direct nerve stimulation, histologicevidence of preserved vocalis muscle bulk, and axonalregrowth into the RLN stump as evidence of a successful nerve transfer.
The amount of reinnervated muscle in the thyroarytenoid was estimated by the amplitude of depolarization on the evoked EMG. In human beings, Peytzet al.44 found the peak-to-peak amplitude of the evokedthyroarytenoid muscle action potential to be 6.8 ± 0.7
mY, and Steiss and Marshall" obtained values of 1.2to 26.0 mV in the dog. The values of 5.5 to 20.0 mVobtained in this study are in agreement with the findingsof Steiss and Marshall." Of the five dogs tested in thisstudy, all showed some degree of reinnervation byevoked EMG, with an average amplitude of 66% of thenormal side. Possible reasons for partial reinnervationafter nerve transfer include axonal escape from theanastamosis, tension or infection of the anastamosis,and devascularization of the nerves after dissection.One of the dogs had an evoked EMG in the reinnervatedthyroarytenoid that was 156% of the normal side. Thismay have been a result of synkinesis within the reinnervated thyroarytenoid. Intramuscular synkinesis maytend to simultaneously depolarize a greater muscle bulkthan the normal side, which would depolarize the samemuscle bulk, but over a longer period of time. Althoughthe absolute bulk of depolarized muscle, as calculatedby the integral under the depolarization curve, may bethe same on both sides, the peak-to-peak amplitude forthe reinnervated cord may be higher.
Motor nerve conduction velocities are closely relatedto the fastest conducting fibers." These fibers are thosewith the largest diameter. The maximal conduction velocity increases by approximately 6 m/sec/ umincreasein fiber diameter. Braund et al." calculated the conduction velocity of the RLN to be 54 to 60 m/sec basedon the largest fiber diameter size of 9 to 10 urn in theRLN. Several studies have measured the conductionvelocity of the RLN directly with evoked EMG.Atkins" found a symmetric conduction velocity of 65

Volume 104 Number 4Aprt11991
m/sec in the proximal RLN and 30 m/sec in the distalRLN. This slowing of conduction in the distal portionof the nerve has also been found by Shin and Rabuzzi.49
Their study also found a faster conduction on the leftin the proximal portion of the RLN. Steiss andMarshall" found the mean normal canine conductionvelocity to be approximately 56 m/sec bilaterally.These values are in agreement with the conductionvelocity of 42.2 m/ sec found in this study for the normal RLN. The RLN conduction velocity has been measured in human beings and found to be 29 m/sec bilaterally."
In this study, the reinnervated side had a conductionvelocity of 30.8 m/sec on average. This slowing of theconduction velocity by approximately 10 m/sec couldbe the result of either a decrease in the axon fiber diameter by 1 to 2 ....m or a relative absence of myelinafter 6 months of regrowth.
The thyroarytenoid muscle is composed of 95% fasttwitch type 2 muscle fiberscompared to 5% slow-twitchtype I fibers." Rice and Cooper" studied the contractileproperties of the canine thyroarytenoid muscle reinnervated from the ansa cervicalis. They found that thetwitch contraction times increased by 23% to 60% overthe normal side. They attributed this change to adaptation of the muscle to the different nature of the motorneuron innervating it with a change in the fiber type.The thyrohyoid and sternothyroid have contractiontimes of 50 msec compared to 12.5 msec for the thyroarytenoid. In two of the three dogs studied by Riceand Copper.50 the twitch was weaker than on the operated side on. However, they did not study fatigability.
In this study, the vocal efficiency (the ratio of acoustic to subglottic power) and the resulting acoustics afteransa nerve transfer were much improved over the paralysis state if there was stimulation of the ansa cervicalis nerve transfer. To what degree the ansa-RLNtransfer is capable of providing this necessary degreeof stimulation during phonation in the human beingsis unclear. Fata et al." found in the canine that asternothyroid nerve-muscle pedicle, when implantedinto the posterior cricoarytenoid, could reinnervatebut not spontaneously activate this muscle, evenduring prolonged occlusion of the airway. GrundfestBroniatowski et al." found that reinnervated strap muscles do not re-establish proprioception. Loss of proprioception may have an effect on the ability of thecentral nervous system to control the fine motor movements of a reinnervated muscle. To obtain muscle contraction, Broniatowski et al." used direct electronicstimulation of the ansa cervicalis nerve-muscle pedicle.This was done to contract an implanted posterior cri-
Futuredlrecttons for laryngeal reinnervation 461
coarytenoid muscle as treatment for bilateral vocal cordparalysis.
What future role the nerve transfer procedure willplay in the treatment of unilateral vocal cord paralysisis evolving. Without physiologic arytenoid adductionand thyroarytenoid contraction, a nerve transfer mayserve to merely maintain the bulk and tone of the paralyzed cord. On the other hand, the choice of appropriate nerve for transfer may produce physiologic arytenoid adduction with vocalis contraction. This wouldresult in improved vocal efficiency and acoustics duringphonation. The optimal nerve for transfer in the humanto provide this necessary degree of stimulation shouldbe the subject of future research.
CONCLUSIONIn this study, the vocal efficiency (the ratio of acous
tic to subglottic power) and acoustics (jitter, shimmer,and signal-to-noise ratio) after ansa-RLN nerve transferfor unilateral recurrent laryngeal paralysis were significantly improved with stimulation of the ansa-RLNanastamosis (as opposed to no stimulation). In the absence of stimulation. physiologic movement could notbe elicited from the reinnervated cord. To what degreethe ansa-RLN transfer is capable of providing this necessary degree of stimulation during phonation in humanbeings is unclear. The optimal nerve for transfer to theRLN. to rehabilitate laryngeal function, should be thesubject of future studies.
We wish to thank Jeff Gombein, DrPH. for his assistancein statistical analysis.
REFERENCES
I. Arnold GE. Vocal rehabilitation of paralytic dysphonia. VIll.Phoniatric methods of vocal compensation. Arch Otolaryngol1962;76:76-83.
2. Isshiki N, Okamura H, Ishikawa T. Thyroplasty type I (lateralcompression) for dysphonia due to vocal cord paralysis or atrophy. Acta Otolaryngol (Stockh) 1975;80:465-73.
3. Isshiki N. Tanabe M. Sawada M. Arytenoid adduction for unilateral vocal cord paralysis. Arch Otolaryngol 1978;104:555-8.
4. Crumley RL. Izdebski K. McMicken B. Nerve transfer versusTeflon injection for vocal cord paralysis: a comparison. laryngoscope 1988;98:1200-4.
5. Tucker H. Laryngeal reinnervation for unilateral vocal cord paralysis. Ann Otol Rl\inol Laryngol 1981;90:457-9.
6. Von Leden H. Abitbol A, Bouchayer M. Hirano M. Tucker H.Phonosurgery. J Voice 1989:3:175-82.
7. Crumley RL. Izdebski K. Voicequality following laryngeal reinnervation by ansa hypoglossi transfer. Laryngoscope 1986;96:611-6.
8. Rice DH. Laryngeal reinnervation. Laryngoscope 1982;92:104959.
9. Horsley J. Suture of the recurrent laryngeal nerve with report ofa case. Trans South Surg Gynecol Assoc 1909;22:161-7.

4166 GREEN 9t 01.
10. Blalock A, Crowe SJ. The recurrent laryngeal nerves in dogs:experimental studies. Arch Surg 1926;12:95-116.
11. Frazier CH. Mosser WB. Treatment of recurrent laryngeal nerveparalysis by nerve anastomosis. Surg Gynecol Obstet 1926;43:134-9.
12. Hoover WB. Surgical procedures for the relief of symptoms ofparalysis of the recurrent layngeal nerves. Surg Clin North Am1953;33:879·85.
13. Siribodhi C. Sunderland W, Atkins JP. et al. Electromyographicstudies of laryngeal paralysis and regeneration of laryngeal motornerves in dogs. Laryngoscope 1963;73:148-63.
14. Crumley RL. Laryngeal synkinesis: its significance to the laryngologist. Ann Otol Rhinal Laryngol 1988;97:87-91.
IS. Doyle PJ. Brummett RE. Everts EC. Results of surgical sectionand repair of the recurrent laryngeal nerve. Laryngoscope1967;77:1245-54.
16. Gordon JH. McCabe BF. The effect of accurate neurorrhaphyon reinnervation and return of laryngeal function. Laryngoscope1968;78:236-59.
17. Boles R, Fritzell B. Injury and repair of the recurrent laryngealnerve in dogs. Laryngoscope 1969;79:1405-18.
18. Tashiro T. Experimental studies on the reinnervation of the larynx after accurate neurorrhaphy. Laryngoscope 1972;82:22S-36.
19. Murakami Y. Kirchner JA. Vocal cord abduction by regeneratedrecurrent laryngeal nerve. Arch Ololaryngol 1971;94:64-8.
20. Sato I. Harvey JE. Ogura JH. Impairment of function of theintrinsic laryngeal muscles after regeneration of the recurrentlaryngeal nerve. Laryngoscope 1974;84:53-66.
21. Dedo HH. Electromyographic and visual evaluation of recurrentlaryngeal nerve anastamosis in dogs. Ann Otol Rhinol Laryngol1971;80:664-8.
22. Ellenbogen BG. Gerber TG. Coon RL. Toohill RJ. Accessorymuscle activity and respiration. OrolARYNGOL HEAD NECK SURD1981;89:360-7S.
23. Marie IP. Dehesdin D. Ducastelle T. Senant I. Selective reinnervation of the abductor and adductor muscles of the caninelarynx after recurrent laryngeal nerve paralysis. Ann Otol RhinolLaryngol 1989;98:530·6.
24. Armstrong B. Smith DI. Function of certain neck muscles duringthe respiratory cycle. Am] Physiol 1955;182:599-600.
25. Woodson GE. Respiratory activity of the cricothyroid muscle inconscious humans. Laryngoscope 1990;100:49-53.
26. Crumley RL. Selective reinnervation of vocal cord adductors inunilateral vocal cord paralysis. Ann Otol Rhinal Laryngol1984;93:351-6.
27. UedaN. Ohyama M. Harvey JE. Mogi G. Ogura IH. Subglotticpressure and induced live voices of dogs with normal, reinnervated and paralyzed larynges. II. Comparison of voice functionof dogs with normal and reinnervated larynges. Laryngoscope1971;81:74-86.
28. Iwamura S. Functioning remobilization of the paralyzed vocalcord in dogs. Arch Ololaryngol 1973;100:122-9.
29. Green DC. Ward PW. Management of the divided recurrentlaryngeal nerve. Laryngoscope 1990;100:779-82.
30. Janecka IP. Peripheral nerve regeneration: an experimental study.Laryngoscope 1987;97:942-50.
31. Van den Berg JW. Direct and indirect determination of the meansubglottic pressure. Folia Phoniatr (Basel) 1956;8:1-24.
32. Iwata S. Aerodynamic aspects for phonation in normal and pathologic larynges. In: Fujimura O. ed. Vocal physiology: Voiceproduction. Mechanisms and functions. New York: Raven Press.Ltd. 1988:423-3 I.
33. Tanaka S. Gould W. Vocal efficiency and aerodynamic aspectsin voice disorders. Ann 0101 Rhinal Laryngol 1985;94:29-33.
OtolaryngologyHeadand Neck SUrg8ly
34. Tanaka S, Tanabe M. Glottal adjustp'lent for regulating vocalefficiency. Acta Otolaryngol (Stockh) 1986;102:315-24.
35. Ludlow C. Coulter D. Gentges F. The differential sensitivityof frequency perturbation to laryngeal neoplasms and neuropathologies. In: Bless OM. Abbs IH. eds. Vocal fold physiology:Contemporary research and clinical issues. San Diego. Calif.:College-Hill Press. 1983:381-92.
36. Milenkovic P. Least mean squares measures of voice perturbation. J Speech Hearing Res I987;30:S29-38.
37. Lieberman P. Some acoustic measurements of the fundamentalperiodicity of normal and pathologic larynges. J Acoust Soc Am1963;35:344-53.
38. Zyski B. Bull G. McDonald W. Johns M. Perturbation analysisof normal and pathologic larynges. Folia Phoniatr (Basel)1984;36:190-8.
39. Iwata S. Periodicities of pitch perturbations in normal and pathologic larynges. Laryngoscope 1972;87:87-95.
40. McDonald W. Zyski B. Johns M, Bull G. Adjunctive use ofperturbation analysis for objective assessment of laryngeal surgery. Proceedings of the Fifth Annual Symposium on ComputerApplications in Medical Care (Institute of Electrical and Electronic Engineers. 198\).
41. Berke G. Moore D. Gerratt B. Hanson D. Bell T. Natividad M.The effect of recurrent laryngeal nerve stimulation on phonationin an in vivo canine model. Laryngoscope 1989;99:977-82.
42. Koyama T. Harvey IE. Ogura IH. Mechanics of voice production. III. Efficiency of voice production. Laryngoscope 1971;81:210-7.
43. van Harreveld A. Re-innervation of denervated muscle fibers byadjacent functioning motor units. Am J Physiol 1945;144:47793.
44. Peytz F. Rasmussen H. Buchthal F. Conduction time and velocityin human recurrent laryngeal nerves. Dan Moo Bull 1965;12:12S-7.
45. Steiss IE. Marshall AE. Electromyographic evaluation of conduction time and velocity of the recurrent laryngeal nerve ofclinically normal dogs. Am] Vet Res 1988;49:1533-6.
46. Hursh JB. Conduction velocity and diameter of nerve fibers. AmJ Physiol 1939;127:131-9.
47. Braund KG, Streiss JE, Marshall AE, Mehta JR. Amling KA.Morphologic and morphometric studies of the intrinsic laryngealmuscles in clinically normal adult dogs. Am J Vet Res1988;49:2105-10.
48. Atkins IP. An electromyographic study of recurrent laryngealnerve conduction and its clinical application. Laryngoscope1973;83:796-807.
49. Shin T, Rabuzzi D. Conduction studies of the canine recurrentlaryngeal nerve. Laryngoscope 1971;81:586-96.
50. Rice DH, Cooper DS. Contractile properties of the canine thyroarytenoid muscle reinnervated from the ansa cervicalis. AnnOtol Rhinol Laryngol 1989;98:153-6.
51. Fata II. Malmgren LT.Gacek RR. Dum R. Woo P. Histochemicalstudy of posterior cricoarytenoid muscle reinnervation by anerve-muscle pedicle in the cat. Ann Otol Rhinal Laryngol1987;96:469-87.
52. Grundfest-Broniatowski S. Broniatowski M. Davies CR. er al.An artificial myotatic reflex: a potential avenue to fine motorcontrol. OTOLARYNGOL HEAD NECK SURD 1989;101:621-8.
53. Broniatowski M. Kaneko S, Jacobs G, Nose Y, Tucker HM.Laryngeal pacemaker. II. Electronic pacing of reinnervated posterior cricoarytenoid muscles in the canine. Laryngoscope1985;95:1194-8.







