zebrafish sex differentiation and gonad development after exposure to 17α-ethinylestradiol,...
TRANSCRIPT

Accepted Manuscript
Title: Zebrafish sex differentiation and gonad developmentafter exposure to 17�-ethinylestradiol, fadrozole and theirbinary mixture: A stereological study
Author: Ana Luzio Sandra M. Monteiro Sofia Garcia-SantosEduardo Rocha Antonio A. Fontaınhas-Fernandes Ana M.Coimbra
PII: S0166-445X(15)30020-5DOI: http://dx.doi.org/doi:10.1016/j.aquatox.2015.07.015Reference: AQTOX 4166
To appear in: Aquatic Toxicology
Received date: 19-5-2015Revised date: 22-7-2015Accepted date: 24-7-2015
Please cite this article as: Luzio, Ana, Monteiro, Sandra M., Garcia-Santos, Sofia,Rocha, Eduardo, Fontainhas-Fernandes, Antonio A., Coimbra, Ana M., Zebrafish sexdifferentiation and gonad development after exposure to 17rmalpha-ethinylestradiol,fadrozole and their binary mixture: A stereological study.Aquatic Toxicologyhttp://dx.doi.org/10.1016/j.aquatox.2015.07.015
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

Zebrafish sex differentiation and gonad development after exposure to 17α-ethinylestradiol, fadrozole and their binary mixture: a
stereological study
Ana Luzio 1,a, Sandra M. Monteiro1, Sofia Garcia-Santos1, Eduardo Rocha2,3, António A. Fontaínhas-Fernandes1, Ana M. Coimbra 1,a
1 Centre for the Research and Technology of Agro-Environmental and Biological Sciences, CITAB, Departamento de Biologia e Ambiente
(DeBA), Escola de Ciências da Vida e Ambiente (ECVA), University of Trás-os-Montes and Alto Douro, UTAD, Quinta de Prados, 5000-801
Vila Real, Portugal, www.utad.pt.
2 Laboratory of Histology and Embryology, Department of Microscopy, Institute of Biomedical Sciences Abel Salazar (ICBAS), University of
Porto (U. Porto), Portugal
3 Histomorphology, Physiopathology and Applied Toxicology Group, Interdisciplinary Centre of Marine and Environmental Research
(CIIMAR), CIMAR Associated Laboratory (CIMAR LA), University of Porto (U. Porto), Portugal
a Corresponding authors. Tel.: +351 259 350 236
E-mail address: [email protected] (A. Luzio), [email protected] (A.M. Coimbra).

Key words: Gonad, sex differentiation, 17α-ethinylestradiol, fadrozole, histology, Danio rerio.
Highlights
• Not all zebrafish males develop a “juvenile ovary” during gonad differentiation.
• EE2 exposure enhanced both zebrafish growth and gonad development.
• Fadrozole affected zebrafish sexual development, inducing masculinization.
• The EE2-Fad mixture allowed identifying sex dependent roles of steroid hormones.
Abstract
Current knowledge on zebrafish (Danio rerio) sex determination suggests that this trait has a polygenic genetic basis, although environmental
factors, such as endocrine disrupting chemicals (EDC), may also be involved in modeling or disturbing the species sex differentiation and
development.

This study aimed to assess how sex steroids imbalance triggers impact on sex differentiation and gonad development in zebrafish. Fish where
exposed to an estrogen (EE2, i.e. 17α-ethinylestradiol, 4 ng/L), to an inhibitor of estrogen synthesis (Fad, i.e. fadrozole, 50 μg/L) or to their
binary mixture (Mix-EE2+ Fad, 4 ng/L + 50 μg/L), from 2 hours to 60 days post-fertilization (dpf). Afterwards, a quantitative (stereological)
analysis using light microscopy, based on systematic sampling, was made at 35 and 60 dpf, to identify alterations on gonad differentiation and
development. During the sex differentiation period, our histological data showed that not all zebrafish males develop a “juvenile ovary”,
contrarily to what is currently taken for granted. Furthermore, the stereological analysis suggests that EE2 alone enhanced both zebrafish growth
and gonad development. On the other hand, exposure to Fad affected the sexual development in zebrafish, inducing masculinization of the
specimens, with some degree of intersex observed in males. In addition, the binary mixture allowed identifying sex-dependent roles of steroid
hormones in the general growth and gonad development of zebrafish, with estrogens acting as growth promoters in females and being essential
for ovary development. Data further support that sex-specific and single EDC impact studies are important, but clearly not sufficient to
understand what may occur in the environment.
1. Introduction

Zebrafish (Danio rerio) has been classified as an undifferentiated gonochoristic species (Yamamoto, 1969), with all individuals developing an
immature ovarian tissue (Uchida et al., 2002) prior to the differentiation into mature ovaries or testes (Maack and Segner, 2003; Takahashi, 1977;
Wang et al., 2007). However, recent studies have shown that this may not always be true (Hsiao and Tsai, 2003; Wang et al., 2007).
Over the past decades, research reports on zebrafish sexual determination (SD) mechanisms have been divergent. According to current
knowledge, SD is apparently controlled by genetic factors, with secondary influences from environmental or even social factors (Devlin and
Nagahama, 2002; Godwin et al., 2003; Liew et al., 2012; Orban et al., 2009; Zhang et al., 2008). In this species, the SD seems to be a complex
trait with a polygenic genetic basis (Anderson et al., 2012; Liew et al., 2012; Liew and Orban, 2013), which is corroborated by the identification
of several loci that affect this mechanism distributed throughout the zebrafish genome (Bradley et al., 2011; Orban et al., 2009). In addition,
several environmental factors are known to perturb SD in zebrafish, such as temperature (Sfakianakis et al., 2012; Uchida et al., 2004), food
(Lawrence et al., 2008), or environmental contaminants, like endocrine disrupting chemicals (EDC) (Hill and Janz, 2003; Soares et al., 2009).
In fish, EDC exposure during critical development periods can impair a normal gonadal development (Scholz and Kluver, 2009; Strussmann and
Nakamura, 2002) and alter sex phenotypes (Fenske et al., 2005; Uchida et al., 2004). Furthermore, both estrogens and androgens, present at
specific developmental stages, are crucial for gonad development (Baroiller et al., 1999; Leet et al., 2011) and exert effects on general growth
(Malison et al., 1985; Malison et al., 1988; Örn et al., 2003). Zebrafish exhibit a sexual dimorphic growth pattern, with females achieving larger
sizes than males (Santos et al., 2008).

The natural estrogen 17ß-estradiol is a sex steroid, and aromatase is a key enzyme for its synthesis, both are essential for the normal ovarian
differentiation and development (Fenske and Segner, 2004; Guiguen et al., 2010). The key role of 17ß-estradiol in ovarian differentiation is
supported by the fact that the inhibition of its synthesis triggers partial or complete masculinization of genotypic females (Fenske and Segner,
2004; Navarro-Martin et al., 2009; Piferrer et al., 1994). In fact, exposure to EDC, such as 17α-ethinylestradiol (EE2) and fadrozole (Fad),
induces sex steroid imbalances, the first as a 17ß-estradiol agonist and the second as an aromatase inhibitor (Fenske and Segner, 2004; Orban et
al., 2009). In fish, previous studies have shown that EE2 and other compounds with estrogenic activity are able to induce a feminization response
(Fenske et al., 2005; Lange et al., 2012; Larsen et al., 2008) and body weight gain (Malison et al., 1985; Malison et al., 1988; Örn et al., 2003;
van der Ven et al., 2007). On the other hand, Fad exposure has been shown to cause sex reversal of genetic females into phenotypic males both in
zebrafish (Andersen et al., 2004; Fenske and Segner, 2004; Uchida et al., 2004) and in other fish species (Ankley et al., 2002; Babiak et al., 2012;
Navarro-Martin et al., 2009). Several studies have quantified potential alterations in gonad morphology of zebrafish after EDC exposure
(Madureira et al., 2011; Silva et al., 2012; Soares et al., 2009). However, the evaluation performed has been done on adult individuals and
information of what happens during the critical period of gonad differentiation is still lacking.
Thus, the purpose of the present study was to fill the identified gap of knowledge by investigating how chemicals known to evoke sex steroid
imbalance impact sex differentiation and gonad development in zebrafish. In order to evaluate this impact, zebrafish were exposed from the
embryonic to the adult stage to a potent synthetic estrogen (EE2), to an inhibitor of estrogen synthesis (Fad) and to their binary mixture (Mix). To

identify alterations on gonad development and differentiation, changes on the relative and absolute volumes of gonad germ cells were assessed
using stereological tools. The study was performed during the gonad differentiation period and in adult fish. In addition, the impact on sex ratio
and biometric parameters were also determined.
2. Materials and methods
2.1 Chemicals
17α-Ethinylestradiol (EE2) (≥ 98% purity), and fadrozole (Fad) (≥ 98% purity) were purchased from Sigma (St. Louis, MO, USA) and were
diluted in ethanol. Working solutions were prepared in MilliQ water and all, including the control water, had in the experimental system an
insignificant final ethanol volume (0.000002%).
2.2 Zebrafish maintenance
Adult wild-type Danio rerio obtained from local suppliers were used as breeding stocks and kept, during a 30 days period of acclimatization, in
120 L aquaria, in a recirculated system with dechlorinated and aerated water. Standardized conditions were maintained: water quality was
monitored and kept optimal (pH 7.7±0.2; hardness 3–4 dGH; 6±1 mg/L of dissolved oxygen; 376 µS/cm of conductivity; 0.08±0.06mg/L of
ammonium and 0.01±0.01mg/L of nitrite). The water was mechanically and biologically filtered continuously (thermo-controlled filter, Eheim,

Germany), aerated and kept at 28 ± 1 ◦C; the photoperiod was kept at 14:10 (light:dark cycle). Fish were fed three times a day with a standard
diet (Westerfield, 2000). The experimental procedures were performed in agreement with European (Directive 2010/63/EU) and Portuguese
(Directive 113/2013) legislations for the correct use of laboratorial animals.
In the afternoon before breeding, several mass crosses (4 females: 8 males) were randomly housed in cages — with a net bottom cover with glass
marbles within a 30 L aquarium under the same water and photoperiod conditions as the stock — and fed with live brine shrimp. In the following
day, breeding fish were removed 1.5 h after the beginning of the light period and the eggs were collected and rinsed several times in system water
to remove debris. Fertilized eggs were randomly allocated to experimental aquaria.
2.3 Exposure conditions
Throughout the experimental period, zebrafish were under a 14:10 h (light:dark) photoperiod and water physicochemical parameters were
measured weekly, in order to maintain the above mentioned values, with the exception of temperature that was checked on daily basis.
Exposures were performed using a flow-through system with slight modifications from the experimental setup described by Soares et al. (2009).
The water (8.4 mL/min) and EE2, Fad and Mix solutions (0.067 mL/min) flows were maintained by peristaltic pumps (ISM 144, ISMATEC) and
mixed together, in a mixing chamber, before entering by gravity the continuously aerated aquaria. The use of peristaltic pumps to precisely

control water and contaminant flows was implemented to increase the accuracy of dosing over time. Furthermore, each working stock solution
was prepared daily.
Four exposure conditions were set up, in duplicate: an experimental control, EE2 (4 ng/L), Fad (50 μg/L) and Mix (4 ng/L EE2 + 50 μg/L Fad).
The selected concentration of EE2 is environmentally relevant, as in surface waters the majority of the detected EE2 concentrations range from
non-detectable to 5 ng/L (Belfroid et al., 1999; Cargouet et al., 2004; Rocha et al., 2012; Rocha et al., 2013). The Fad concentration used (50
μg/L) was selected based on previous studies conducted in zebrafish (Andersen et al., 2004) and other fish species (Ankley et al., 2002; Zhang et
al., 2008), where masculinization induction was reported.
Before the beginning of the exposure period, the aquaria were equilibrated with the solutions for 15 days. The experiment was initiated by
randomly allocating 500 fertilized eggs with proximally 2 h post-fertilization (hpf) in 5 L aquaria until 15 dpf, these were placed within the
corresponding 30 L aquarium, and afterwards zebrafish were allocated in the 30 L aquaria. According to the methodologies previously described
by Soares et al., 2009, daily feeding was initiated at 6 dpf with two meals of a commercial fish diet Tetramin (Tetra, Melle, Germany)
supplemented with one meal of live brine shrimp (Artemia spp., Ocean Nutrition; Salt Creek Salt Lake City, UT, USA) per day; at 9 dpf this was
changed to two brine shrimp meals and one Tetramin meal per day, which was maintained until the end of exposure. The food particle size and
quantity was adjusted throughout the experiment based on the age and number of fish remaining in the aquarium as previously described by
Carvalho et al. (2006).

Mortality was recorded daily during the entire exposure period, and dead eggs/individuals were removed. At 35 and 60 dpf, 30 zebrafish per
treatment (15 of each duplicate) were randomly sampled, anaesthetized in a buffered solution of tricaine methanesulfonate (MS-222 100 mg/mL;
Finquel, Redmond WA, USA), measured, weighed and immediately euthanized. Their bodies were collected to buffered formaldehyde (Panreac,
Barcelona, Spain), for sex examination and stereological analysis.
2.4 Histological preparation
Fixed fish were processed for histological examination by light microscopy. After fixation (48 h), the tissues were dehydrated through a series of
graded ethanol (Fisher Scientific, U.K.) solutions (70–100%), cleared in xylene (Fisher Scientific, U.K.), and impregnated in paraffin Histosec®
pastilles (Merck, Darmstadt, Germany) in a semi-enclosed tissue processor (Tissue-Tek II model 4634). Finally, the tissues were embedded in
paraffin in a modular tissue embedding center (Leica EG 1160). Ten zebrafish per treatment, 5 per replicate, were randomly sampled and each
gonad was entirely sectioned into thin sections (3 µm thick) in a rotary microtome (Leica RM 2135). The sagittal sections were systematically
sampled during sectioning so that at least a minimum of 15 sections per animal were analyzed. The remaining zebrafish, 20 per treatment, were
sectioned (3 µm thick) for sex identification. To improve section adhesion, the sampled sections were mounted in (3-aminopropyl)triethoxysilane
(APES) (Sigma–Aldrich, Germany) coated slides, then stained with hematoxylin-eosin (H&E stain) before being coverslipped.

2.5 Stereological analysis
To determine the volume density (VV) of the components of the zebrafish gonad, a stereological approach was performed according to previously
described methodologies (Madureira et al., 2011). The VV is defined as the percentage of the total volume, of a well defined reference space,
occupied by any given component within it. According to this definition, the estimate VV of the different identified cellular elements, within the
gonad (the present reference space), was made by a classical stereological technique based on point counting (Freere and Weibel, 1968) using the
following formula:
VV(structure, reference) = [P(structure) × 100] ÷ [k × P(reference)]
where P(structure) is the number of test points within each structural component, P(reference) is the total number of test points lying over the
reference space (gonad), and k is the ratio between the number of points on the grid used for the structure of interest and for the reference space.
Counting was done using an inverted microscope (IX 51, Olympus, Japan) equipped with a digital camera (Color View III, Olympus, Hamburg,
Germany), connected to a 17’’ PC monitor (Dell), and in images captured with the Olympus Cell 2.6 software. The first field of vision was
randomly selected and the following fields were systematically sampled by stepwise movements of the stage in the x- and y-directions, spaced
according to the gonad size, in order to count all components in each ovary or testis section area observed: 430 µm × 330 µm in females and 220
µm × 170 µm in males. The stereological test grids used for estimating the VV of each compartment had two sets of points, which were
previously established depending on the frequency and size of the targeted compartment, and were super-imposed on the live image of the

monitor. Point counting was made in the systematically sampled fields working with the 20× (females) and the 40× (males) Olympus UIS-2
objective lens (Olympus Co., Ltd., Tokyo, Japan).
For the ovary, a 1:9 grid was used, with a total of 9 coarse points targeting the late/mature oocyte compartment, the atretic mass compartment and
the reference space, and 81 points targeting the remaining compartments. For the testis, a 1:4 grid was designed and the VV of spermatozoa,
spermatocytes, lumen, and reference space were evaluated via a 16 point subset, and the other compartments with a denser 64 point subset.
The different stages considered during oogenesis and spermatogenesis were distinguished based on morphological and physiological criteria
described previously (Leal et al., 2009; Madureira et al., 2011; Selman et al., 1993; Silva et al., 2012).
Briefly, at 35 dpf, common to both sex, undifferentiated gonocytes were identified - cells with rather homogeneous, slightly basophilic
karyoplasm, and containing one or more nucleoli in the large nucleus. Regarding the ovary, five follicular stages of maturation were identified
and counted (Fig. 1): Oogonium - high ratio nucleus/cytoplasm; cytoplasm weakly acidophilic; acidophilic nuclear membrane and nucleolus;
Primary growth stage - enlarged basophilic cytoplasm; pale nucleus with many nucleoli distributed along the nuclear membrane; Cortical
alveolus stage - presence of the cortical alveoli in the cytoplasm, first in the peripheral zone and after in all cytoplasm; irregular nuclei shape
with nucleoli attached to inner border of nuclear membrane; Vitellogenic stage - cytoplasm with yolk bodies which are heterogeneous in
appearance; migration of the nucleus to the animal pole; zona radiata prominent and granulosa-theca cells easily identified; Mature stage -

oocyte filled with yolk bodies; small area occupied by the cytoplasm; oocyte layers folded irregularly. Atretic (degenerated) follicles and
interstitial connective tissue were also counted.
Regarding the testis, also five cells populations were identified as follows (Fig. 2): Spermatogonium - the largest of the spermatogenic cells,
displaying acidophilic cytoplasm with a relatively larger nucleus containing poorly condensed chromatin and one or two compact nucleoli; Early
and late spermatocyte - early spermatocytes are larger than late spermatocytes, with thread-like or condensed chromatin, respectively, and with
relatively smaller cytoplasm with no affinity for dye; Spermatid - dense nuclei and narrow rims of eosinophilic cytoplasm; and Spermatozoon -
dark, round nuclei and minimal or no apparent cytoplasm.
The absolute gonad volume [V(gonad)] (µm3), for both ovaries and testes, was estimated by the Cavalieri principle, based on the formula
(Gundersen et al., 1988):
V(gonad) = ∑ A × t
where (A) (µm2) is the area of one section observed and (t) (µm) is the thickness of each section; nominally 3 µm, based on a standardized
sectioning procedure across all fish. The measurements of the individual areas were done using a digital image analysis software (Digimizer
version 4.2.0.0, MedCalc Software, Mariakerke, Belgium).
The absolute volumes (µm3) of all the gonadal structures indicated above were then estimated based on the formula:
V(structure) = V(gonad) × VV(structure, gonad)

For estimating the gonad weight (GW), it was assumed that the gonad density is ≈1 mg/mm3 for both sex (Silva et al., 2012). Accordingly, GW
(mg) ≈ V(gonad) (µm3).
The remaining fish ovary and testis sections were examined under light microscopy, to identify the sex, gonad development, and the presence of
pathological alterations.
2.6 Statistical analysis
To identify differences in zebrafish sex ratio between control (1:1) and each treatment group was applied a Chi-square analysis, using the
GraphPad Prism 6.00 software (GraphPad Software, San Diego, CA, USA). Biometric parameters and stereological data were analyzed using
SigmaStat software (Systat). After testing for ANOVA assumptions (homogeneity of variances, with the Levene's mean test, and normality, with
the Kolmogorov-Smirnov test, statistical differences between different EDC exposures in gonads were evaluated by one-way factorial ANOVA,
followed by the post hoc Student-Newman-Keuls test. As zebrafish have dimorphic growth, biometric parameters statistical differences where
determined by a two-way ANOVA, in this case both gender and EDC exposures effects where used as independent variables. In the specific
cases where the parametric ANOVA assumptions failed and when the observed values were equal to zero, data were submitted to a non-
parametric ANOVA, using Kruskal-Wallis test (for multiple comparisons), and to the Mann-Whitney rank sum test (for pairwise comparisons).
The non-parametric results were alike those of the parametric ANOVA analysis, strengthening the confidence in the significance of the

differences. Significant differences were considered for p<0.05. Zeros were used when a specific compartment was not seen in a particular group
or time, but there was a chance of being present in the different conditions; such zeros were included in the statistical analysis. Gonad
compartments that are never present, at any time, in a specific gonad type were presented as non-determined (N.D.) and were excluded from the
statistical analysis.
3. Results
3.1 Biometric parameters
During the 60 days experimental period the mortality did not exceed 20% in any group and no differences were observed between the exposure
groups. Body weight, length, gonad weight and gonadosomatic index (GSI) were determined in both sexes at 35 and 60 dpf (Table 1).
The body weight of zebrafish females was found to be increasing more in the group exposed to EE2 than in control group (p<0.05) at 35 dpf. At
60 dpf, the presence of EE2 clearly induced an increment of female body weight, observed both in the EE2 (p<0.001) and in the Mix (p<0.05)
exposed groups. Regarding zebrafish males, an increase of body weight was also observed in the group exposed to EE2, but only at 60 dpf
(p<0.001). A similar pattern was observed for total length, although the differences between EE2-exposed females and remaining females at 35
dpf did not reach the significance levels (p>0.05). Differences between genders, for both weight (p<0.05) and length (p<0.05), were observed at
35 dpf in the group exposed to EE2 and at 60 dpf in the group exposed to the Mix.

Concerning ovary weight, at 35 dpf, this did not follow the slight increment observed for total weight of females exposed to EE2, as the control
group was the one presenting a trend for highest mean value. Particularly, the females from Fad (p<0.05) group showed lower values than control
fish; the Mix was between control and Fad (p>0.05). At 60 dpf, both testis (p<0.05) and ovary (p<0.01) weights of fish exposed to EE2 followed
the trend of body weight gain. Likewise, females from the Mix-exposed group presented heavier ovaries than controls (p<0.05). Differences
between ovary and testis weight were observed in the groups control and EE2 (p<0.001), at 35 dpf, and EE2 and Mix (p<0.01), at 60 dpf.
A significant reduction of the GSI (p<0.05) at 35 dpf was observed in zebrafish females exposed to all EDC groups. However, this was not
observed at 60 dpf. This biometric parameter presented always higher values in female than in male fish.
3.2 Histological analysis and gonad development classification
The histological analysis performed to identify the phenotypic sex at 35 dpf, showed the existence of two process of gonad development in male
juvenile zebrafish, as previously described by Wang et al. (2007). Accordingly, fish in differentiation were classified in four categories: females
and type I, II and III males (Fig. 3). In males, the gonad differentiation process in “type II and III” involved the transformation of the “juvenile
ovary” into testis, whereas in “type I” there was no evidence of the “juvenile ovary” being ever formed. The gonads of “type III” males showed a
typical ovarian structure, similar to that of the females. However, “type III” males presented larger ovarian cavities (p<0.05) and a considerably

smaller size than females. These were he main criteria used in the present work to differentiate females from “type III” males (Figs. 3C, 3D and
Table 2).
Gonads of “type II” males, more advanced in the differentiation process, presented partially masculine and feminine structures (Figs. 3E, 3F and
Table 2). Oocyte degeneration was evidenced by the appearance of red staining eosinophilic cells and vacuolated oocytes (Fig. 3F). The presence
of degenerating oocytes and the proliferation of stroma cells, mixed sometimes with early spermatocytes, indicated that the gonads were
transforming from the “juvenile ovary” into testis (Figs. 3C to 3F). At last, males with “type I” gonads contained only one or two oocytes, in at
least one section, but there was no evidence of a “juvenile ovary” ever being formed before transformation (Figs. 3G, 3H and Table 2). This type
of males compared to “type II/III” showed a rise in the relative volumes of stroma somatic cells (p<0.05), while the volumetric density of
primary growth (p<0.001) and of interstitial tissue (p<0.01) were significantly decreased (Table 2). In addition, some males that already had
completed the differentiation process were identified in the groups exposed to EE2 (40%) and to the Mix (27%). However, none of these fish
were among the 10 randomly selected individuals with totally sectioned gonads.
3.3 Sex ratio

The Chi-squared test was performed to determine significant alterations in the proportion of each sex in the different treatments. The control
group, both at 35 and 60 dpf, presented a 1:1 proportion of male to female phenotypic sex ratio (Fig. 4). The EE2 group also presented a similar
proportion, at both times of exposure. However, the number of females was slightly (but not significantly) higher at 60 dpf.
In contrast, both Fad and Mix exposures induced zebrafish masculinization, with the corresponding exposed groups showing significant increases
in the number of males (p<0.0001). Both Fad groups were composed of 90% of males, while the Mix groups presented 87% and 90% of male
individuals, at 35 and 60 dpf, respectively (Fig. 4).
At 35 dpf, testes were classified in four categories, type I, II, III and differentiated (Fig. 5). The results show the presence of differentiated testis
only in the groups exposed to EE2 and to the Mix, indicating an advance on this organ maturation. The group exposed to Fad presented
percentages of each male type similar to the control group. Overall, whatever the group, the percentage values of “type II” males were always the
lowest.
3.4 Stereological analysis
For stereological analyses, “type II and III” males were assessed as a single group. The histological examination of serial gonad sections, from
control and treatment groups, revealed normal testicular and ovarian organization. In this work, and because fish were still in development and
this is not a synchronized process, differences in the maturation within the same group and treatment substantially increased the standard

deviations (SD) of the means. To illustrate variability, at 35 dpf, in the “type I” males (Table 2 and Suppl. 1), individuals with advanced and
delayed maturation were both observed.
At 35 dpf, the absolute ovary volume of fish exposed to EE2 did not follow the increase observed in fish weight (Suppl. 1). At that time the
control group was the one that already presented a trend for highest absolute ovary volume (p>0.05). In males, increments of the absolute
testicular volume were observed upon exposure to EE2; however, the increment was only in “type II/III” (p<0.01). In addition, the absolute
testicular volume increased on “type II/III” males exposed to the Mix (p<0.05) (Suppl. 1). Both females and “type I” males exposed to EE2
showed an advance on gonad development, presenting lowest relative volumes of undifferentiated gonocytes (p<0.05) (Table 2).
At 60 dpf, the exposure to EE2 promoted zebrafish growth and gonad development; this was observed both in females and in males and was
reflected by an increase of the absolute gonad volumes of the exposed fish (p<0.001) (Figs. 6, 7 and Suppl. 2, 3). In females, this fact was mainly
due to increases in the relative volumes of advanced oocyte maturation stages, such as cortical alveoli, vitellogenic and mature oocytes, while the
volumetric density of primary oocyte decreased (Fig. 6). In males, the absolute volumes of the germ cell types selected for quantification were
significantly increased in the EE2 group (Fig. 7 and Suppl. 3). Despite this, the corresponding relative volumes of these cells were not different
from control (Fig. 7 and Suppl. 3). The increment of absolute testes volumes detected in this group was thus the result of general fish and gonad
growth (p<0.001).

In contrast, females exposed to Fad showed a delay in the gonad development (Fig. 8 and Suppl. 2). In this case, ovaries were mainly composed
of cells in early gametogenic stages, and oocytes in advanced maturation were absent (Figs. 6, 8 and Suppl. 2). The relative volumes of ovary
cells also revealed a high percentage of primary growth, a low presence of cortical alveoli and the absence of vitellogenic and mature oocytes
(Fig. 6).
The results show opposite effects of EE2 and Fad exposures, with advance of ovary development and delay of ovary maturation being,
respectively, observed (Fig. 8). These opposite effects were reflected by significant differences on the relative volumes of primary oocytes and of
oogonia (p<0.05) (Fig. 6). In males exposed to Fad, singly or in co-exposure, a slight decrease was observed on the absolute volume of the testis
(p>0.05) (Suppl. 3).
Exposure to the Mix also promoted zebrafish ovary growth (Fig. 8) and, as in the EE2 group, advanced follicle maturation stages were observed
with a rise in the relative volumes of both cortical alveoli and vitellogenic oocytes (Fig. 6). In contrast, the exposure of males to the Mix did not
promote testicular maturation, as both the absolute and the respective relative volumes of the various germ cell types were present in amounts
similar to control male (Fig. 7 and Suppl. 3). In addition, in two individuals of this group the relative volumes of spermatozoa were close to zero.
Concerning pathological alterations of gonads, none were observed in individuals from the control and EE2 exposed groups. However, at 60 dpf,
one male from the Fad and another from the Mix exposed groups had primary oocytes in some of the testicular sections (Fig.9). The relative
volumes of primary oocytes in these fish were 3.45% and 0.40%, and the absolute volumes 1.34 x10-3 mm3 and 0.15 x10-3 mm3, respectively. The

simultaneous presence of male and female gonadal tissue in a gonochoristic species, known as an intersex condition, was observed in 20% of the
males whose gonad was totally sectioned and which were exposed to the aromatase inhibitor. As the relative volumes of primary oocytes
observed on totally sectioned testes was very low, they are highly unlikely to be observed in random gonad sections. Consequently, it is possible
that some of the remaining males may also present an intersex condition.
Furthermore, at 60 dpf, one of the zebrafish females exposed to Fad (Fig. 10A) and a male from the Mix group (Fig. 10C) revealed a delay in
gonad maturation, presenting the gonads and the relative volumes of each tissue similar to the ones observed in 35 dpf individuals (Fig. 10B and
10D). This female had a gonad similar to a “juvenile ovary”, and presented a body weight (48.08 mg) and length (18.16 cm) two times lower
when compared to other females from the same group (Fig. 10A). The “juvenile ovary” of this female had an absolute volume of 9.15 x10-3 mm3,
which was two times lower than the volume presented by females from the same group at 35 dpf. Regarding the male from the Mix group, it
presented a gonad structure and an absolute volume of 3.09 x10-3 mm3 similar to a “type I” male gonad at 35 dpf. Still, this male body weight
(95.74 mg) and length (22.72 cm) were within the values observed in 60 dpf males from the same group (Fig. 10C).
4. Discussion
Fish exposure to EDC is widely known for inducing many effects on the reproductive system, ranging from changes in gene expression to
alterations in the morphological level. However, how these can interfere in critical early periods such as gonad differentiation is still poorly

understood. In addition, learning the effects of a single EDC may not be sufficient for understanding what is really occurring in the environment
(Rocha et al., 2012; Silva et al., 2012; Urbatzka et al., 2012), as, among others, estrogenic and androgenic compounds co-exist in aquatic
ecosystems (Leusch et al., 2006; Soto et al., 2004).
This is the first study to apply stereological methods to zebrafish gonad during the sexual differentiation period, allowing both the identification
and quantification of alterations on the gonadal development caused by the selected EDC exposures; and further showing that they do interfere
not only with early development but also with the final sexual differentiation of zebrafish, both when used alone and in co-exposure.
During the “juvenile ovary” stage, the histological analysis and classification of gonads showed the existence of two processes of gonad
development in juvenile males. This has been first described on a stable transgenic zebrafish (vas::egfp), with male individuals being classified in
three categories according to the intensity of the reporter expression of the enhanced green fluorescence protein (EGFP), and to the morphology
of the gonad during the differentiation period (Wang et al., 2007). In fact, the proportion observed for each category in the control group was
similar to the results of Wang et al. (2007). Taken together, the results of that pioneering work and ours question the established concept that all
zebrafish males go through a “juvenile ovary” stage (Maack and Segner, 2003; Takahashi, 1977), as both show that a large proportion of
zebrafish males (type I) during that transient “juvenile ovary” stage only develop an insignificant number of primary oocytes.
During the sex differentiation period, at 35 dpf, in the groups exposed to EE2 and to the Mix, some males had already completed the testicular
differentiation process. In contrast, females only showed a decline on the relative volume of undifferentiated gonocytes in the EE2 group,

indicating that exposure to estrogens may accelerate the process of gonad maturation. This effect of EE2 was evident at 60 dpf, where females of
the EE2 and Mix groups and males exposed to EE2 showed both higher absolute volumes of the gonads and of the more advanced stages of the
germ cell lines. While in females this can be related to the relative volume increase of later maturation stages, in males this effect of gonad
growth was only reflected in the absolute testicular volume. The increase in gonad size and the acceleration of differentiation (male and female),
as the presence of later maturation stages in the ovary, were paralleled by a general increase in the growth of individuals.
Several studies have reported that exposures to estrogens, such as 17ß-estradiol and EE2, promote body weight gain in zebrafish (Larsen et al.,
2008; Örn et al., 2003; van der Ven et al., 2007) and other fish species such as three-spined stickleback (Gasterosteus aculeatus) (Bell, 2004),
European eel (Anguilla anguilla) (Degani, 1986; Tzchori et al., 2004), yellow perch (Perca flavescens) (Goetz et al., 2009; Malison et al., 1985;
Malison et al., 1988) and goldfish (Carassius auratus auratus) (Zou et al., 1997). It is worth noting that the dual action of estrogens depends on
the concentrations that animals are exposed to, reflecting their role as hormones and the different feedback mechanisms. In this vein, whereas
zebrafish exposed to 100 ng/L EE2 showed a significantly reduced growth (Silva et al., 2012; Versonnen and Janssen, 2004), the opposite
occurred when zebrafish were exposed to low EE2 concentrations, viz. 2, 5, and 10 ng/L (Örn et al., 2003), and 0.05, 0.5, and 5 ng/L (Larsen et
al., 2008). Increased growth was also observed in the present study after 60 dpf at low EE2 concentrations. The same kinds of impacts over
zebrafish growth as a result of low exposure concentrations were also seen with 17β-estradiol (van der Ven et al., 2007). The increased growth
rates resulting from estrogen exposure are most related with the role of these hormones in growth regulation through affecting the growth

hormone/insulin-like growth factor system (GH/IGF) (Goetz et al., 2009; Malison et al., 1985; Malison et al., 1988; Örn et al., 2003; Roberts et
al., 2004; Zou et al., 1997). Estrogen modulates metabolism primarily through the insulin-like growth factor (IGF) pathway, which in turn,
through the stimulation of amino acid and lipid metabolism, influences growth (Roberts et al., 2004). The present study showed that EE2
stimulated both gonad development and somatic growth, as indicated by the positive effects on body and gonad weight and total length,
corroborating the growth-promoting potential of this compound, at least under certain concentrations and exposure conditions (Malison et al.,
1985; Malison et al., 1988; Örn et al., 2003; van der Ven et al., 2007; Zou et al., 1997).
The sex ratio observed in the EE2 group was identical to the one in the control group, and expected for zebrafish reared under normal laboratory
conditions, theoretically 1:1 male to female (Liew and Orban, 2013; Santos et al., 2006). Likewise, EE2 studies where zebrafish were also
exposed to similar (Baumann et al., 2014; Soares et al. 2009) and higher (Islinger et al., 2003; Örn et al., 2003) concentrations, showed no
alteration of the sex ratio. On the other hand, the concentration of Fad used herein induced the masculinization of zebrafish, even when fish were
co-exposed with EE2. Similar results of female zebrafish masculinization were observed when this species was exposed to Fad during the sex
differentiation period, both through diet (Fenske and Segner, 2004; Uchida et al., 2004) and water (Andersen et al., 2004). In fact, at 60 dpf, the
few fish that remained as females after Fad exposure showed a predominance of primary oocytes over the later stages of oocyte maturation. This
is in accordance with previous studies, where Fad exposures also induced a delay of the maturity of ovaries (Andersen et al., 2004; Ankley et al.,
2002; Olmstead et al., 2009). These results indicate that estrogens are essential for normal ovarian differentiation and development (Fenske and

Segner, 2004; Guiguen et al., 2010), and that the EE2 concentration used in this study was not enough to induce the correct ovary development; at
a time when aromatase was inhibited and potentially decreased natural estrogen levels.
Moreover, incidence of intersex, at 60 dpf, was documented histologically in this work. The most frequently reported manifestation of intersex is
the presence of single or multiple oocytes which normally grow into the testis (Bahamonde et al., 2013; Nolan et al., 2001; Randak et al., 2009)
which is in agreement with previous occurrences reported both in zebrafish (Brion et al., 2004; Hill and Janz, 2003; Örn et al., 2003) and in
studies with similar treatments and other fish species (Afonso et al., 2001; Gao et al., 2010; Peranandam et al., 2014). This result indicates that
Fad exposure, although potentiating masculinization, may at the concentration used not completely transform females into males, as some of the
gonads still present oocytes, or the exposure time was not sufficient to masculinize all females as they have an asynchronous gonad maturation.
In zebrafish males exposed to this aromatase inhibitor, also a slight decrease on the absolute volume of testes was observed, even in co-exposed
fish. In general, male fish are more sensitive to EDC and are thus more affected than females (Tejeda-Vera et al., 2007) with a decrease in testis
size a reported effect (Bahamonde et al., 2013; Randak et al., 2009). This decrease may result from the activity of Fad as aromatase inhibitor,
leading to declines on estradiol production (Afonso et al., 2001; Ankley et al., 2002; Panter et al., 2004) and consequently its circulating
concentration (Villeneuve et al., 2009; Zhang et al., 2008). This is due to a down-regulation of GH protein levels and consequently affects gonad
growth of zebrafish (Andersen et al., 2004). The present results suggest that for zebrafish growth and sexual maturation estrogen levels are more
relevant than androgens levels. This conclusion is supported by the fact that both processes were enhanced by the increase in estrogens by EE2

exposure and not affected, or even slightly depressed, when there was a condition for the potential decline on female sex steroids, towards an
increase on androgens levels, logically induced by Fad aromatase inhibition.
Few studies report the combined effects of EDC mixtures with antagonistic effects on zebrafish sex differentiation (Micael et al., 2007; Örn et
al., 2003; Santos et al., 2006). In the Mix, Fad still induced zebrafish masculinization, promoted intersex in testis and neutralized the body and
gonad weight gain response to EE2. While EE2 delayed the maturation of testis in males, lowering the relative volume of spermatozoa, in the few
females (10%) that were not masculinized in Mix group, it acted as growth and maturation promoter as shown by the rise of biometric parameters
and of the relative volumes of advanced oocyte maturation stages. This clearly shows that estrogens and androgens have different targets and
roles triggering distinct effects depending on the individual sex, and raises new questions regarding: 1) how natural fish populations may respond
to EDC mixtures, and 2) whether or not gender differences may turn one of the sexes more susceptible to endocrine disruption.
In summary, this is the first study to used unbiased stereological methods to identify structural effects of model waterborne EDC on zebrafish
gonad during and after the sex differentiation period. The quantification of absolute volume of the gonad compartments showed to be a more
sensible method to evaluate effects in comparison to assessing only relative volumes. In fact, the present work showed that the most prominent
effect was a global increase in almost all gonad compartments, instead of alterations on the proportions of each one.
Furthermore, it was shown that estrogens are not only essential for the zebrafish ovary development, but they also have an important role as
growth promoters. Moreover, the Fad promotion of androgen levels in zebrafish induced masculinization, as with the addition of the estrogenic

compound the effect was not reversed. Clearly, studying the effects of single EDC are not be sufficient to understand what is really occurring in
the environment. Moreover, as the EDC had sex-dependent targets and responses, further studies on xenoestrogens and xenoandrogens actions
should always include both sexes.
Acknowledgments
This work was supported by the Portuguese Foundation for Science and Technology through the project grant PTCD/CTV/102453/2008
(FCOMP-01-0124 FEDER-009527) and the PhD grant SFRH/BD/44794/2008 attributed to Ana Luzio.
Bibliography
Afonso, L.O.B., Wassermann, G.J., De Oliveira, R.T., 2001. Sex reversal in Nile tilapia (Oreochromis niloticus) using a nonsteroidal aromatase
inhibitor. J Exp Zool 290, 177-181.
Andersen, L., Holbech, H., Gessbo, A., Norrgren, L., Petersen, G.I., 2003. Effects of exposure to 17 alpha-ethinylestradiol during early
development on sexual differentiation and induction of vitellogenin in zebrafish (Danio rerio). Comp Biochem Physiol C 134, 365-374.

Andersen, L., Kinnberg, K., Holbech, H., Korsgaard, B., Bjerregaard, P., 2004. Evaluation of a 40 day assay for testing endocrine disrupters:
Effects of an anti-estrogen and an aromatase inhibitor on sex ratio and vitellogenin concentrations in juvenile zebrafish (Danio rerio). Fish
Physiol Biochem 30, 257-266.
Anderson, J.L., Mari, A.R., Braasch, I., Amores, A., Hohenlohe, P., Batzel, P., Postlethwait, J.H., 2012. Multiple sex-associated regions and a
putative sex chromosome in zebrafish revealed by RAD mapping and population genomics. PLoS ONE 7(7): e40701.
Ankley, G.T., Kahl, M.D., Jensen, K.M., Hornung, M.W., Korte, J.J., Makynen, E.A., Leino, R.L., 2002. Evaluation of the aromatase inhibitor
fadrozole in a short-term reproduction assay with the fathead minnow (Pimephales promelas). Toxicol Sci 67, 121-130.
Babiak, J., Babiak, I., van Nes, S., Harboe, T., Haugen, T., Norberg, B., 2012. Induced sex reversal using an aromatase inhibitor, Fadrozole, in
Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 324, 276-280.
Bahamonde, P.A., Munkittrick, K.R., Martyniuk, C.J., 2013. Intersex in teleost fish: Are we distinguishing endocrine disruption from natural
phenomena? Gen Comp Endocrinol 192, 25-35.
Baroiller, J.F., Guiguen, Y., Fostier, A., 1999. Endocrine and environmental aspects of sex differentiation in fish. Cell Mol Life Sci 55, 910-931.
Baumann, L., Knorr, S., Keiter, S., Rehberger, K., Volz, S., Schiller, V., Fenske, M., Holbech, H., Segner, H., Braunbeck, T., 2014. Reversibility
of endocrine disruption in zebrafish (Danio rerio) after discontinued exposure to the estrogen 17 alpha-ethinylestradiol. Toxicol Appl Pharmacol
278, 230-237.

Belfroid, A.C., Van der Horst, A., Vethaak, A.D., Schafer, A.J., Rijs, G.B.J., Wegener, J., Cofino, W.P., 1999. Analysis and occurrence of
estrogenic hormones and their glucuronides in surface water and waste water in the Netherlands. Sci Total Environ 225, 101-108.
Bell, A.M., 2004. An endocrine disrupter increases growth and risky behavior in threespined stickleback (Gasterosteus aculeatus). Horm Behav
45, 108-114.
Bradley, K.M., Breyer, J.P., Melville, D.B., Broman, K.W., Knapik, E.W., Smith, J.R., 2011. An SNP-Based linkage map for zebrafish reveals
sex determination loci. G3 (Bethesda) 1, 3-9.
Brion, F., Tyler, C.R., Palazzi, X., Laillet, B., Porcher, J.M., Garric, J., Flammarion, P., 2004. Impacts of 17 beta-estradiol, including
environmentally relevant concentrations, on reproduction after exposure during embryo-larval-, juvenile- and adult-life stages in zebrafish
(Danio rerio). Aquat Toxicol 68, 193-217.
Cargouet, M., Perdiz, D., Mouatassim-Souali, A., Tamisier-Karolak, S., Levi, Y., 2004. Assessment of river contamination by estrogenic
compounds in Paris area (France). Sci Total Environ 324, 55-66.
Carvalho, A.P., Araújo, L., Santos, M.M., 2006. Rearing zebrafish (Danio rerio) larvae
without live food: evaluation of a commercial, a practical and a purified starter diet on larval performance. Aquaculture Res. 37, 1107–1111.
Degani, G., 1986. Effect of dietary 17-Beta-Estradiol and 17-Alpha-Methyltestosterone on growth and body composition of slow-growing Elvers
(Anguilla anguilla L.). Comp Biochem Physiol A 85, 243-247.

Devlin, R.H., Nagahama, Y., 2002. Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental
influences. Aquaculture 208, 191-364.
Fenske, M., Maack, G., Schafers, C., Segner, H., 2005. An environmentally relevant concentration of estrogen induces arrest of male gonad
development in zebrafish, Danio rerio. Environ Toxicol Chem 24, 1088-1098.
Fenske, M., Segner, H., 2004. Aromatase modulation alters gonadal differentiation in developing zebrafish (Danio rerio). Aquat Toxicol 67, 105-
126.
Freere, R.H., Weibel, E.R., 1967. Stereologic techniques in microscopy. J Royal Microsc Soc 87, 25–34.
Gao, Z.X., Wang, H.P., Wallat, G., Yao, H., Rapp, D., O'Bryant, P., MacDonald, R., Wang, W.M., 2010. Effects of a nonsteroidal aromatase
inhibitor on gonadal differentiation of bluegill sunfish Lepomis macrochirus. Aquacult Res 41, 1282-1289.
Godwin, J., Luckenbach, J.A., Borski, R.J., 2003. Ecology meets endocrinology: environmental sex determination in fishes. Evol Dev 5, 40-49.
Goetz, F.W., Rise, M.L., Rise, M., Goetz, G.W., Binkowski, F., Shepherd, B.S., 2009. Stimulation of growth and changes in the hepatic
transcriptome by 17 β-estradiol in the yellow perch (Perca flavescens). Physiol Genomics 38, 261-280.
Guiguen, Y., Fostier, A., Piferrer, F., Chang, C.F., 2010. Ovarian aromatase and estrogens: A pivotal role for gonadal sex differentiation and sex
change in fish. Gen Comp Endocrinol 165, 352-366.

Gundersen, H. J. G., Bendtesen, T. F., Korbo, L., Marcussen, N., Moller, A., Nielsen, K., Nyengaard, J. R. , Pakkenberg, B., Sorensen, F. B.,
Vesterby, A., West, M. J., 1988. Some new, simple and efficient stereological methods and their use in pathological research and diagnosis.
APMIS 96, 379-394.
Hill, R.L., Janz, D.M., 2003. Developmental estrogenic exposure in zebrafish (Danio rerio): I. Effects on sex ratio and breeding success. Aquat
Toxicol 63, 417-429.
Hsiao, C.D., Tsai, H.J., 2003. Transgenic zebrafish with fluorescent germ cell: a useful tool to visualize germ cell proliferation and juvenile
hermaphroditism in vivo. Dev Biol 262, 313-323.
Islinger, M., Willimski, D., Volkl, A., Braunbeck, T., 2003. Effects of 17a-ethinylestradiol on the expression of three estrogen-responsive genes
and cellular ultrastructure of liver and testes in male zebrafish. Aquat Toxicol 62, 85-103.
Lange, A., Katsu, Y., Miyagawa, S., Ogino, Y., Urushitani, H., Kobayashi, T., Hirai, T., Shears, J.A., Nagae, M., Yamamoto, J., Ohnishi, Y.,
Oka, T., Tatarazako, N., Ohta, Y., Tyler, C.R., Iguchi, T., 2012. Comparative responsiveness to natural and synthetic estrogens of fish species
commonly used in the laboratory and field monitoring. Aquat Toxicol 109, 250-258.
Larsen, M.G., Hansen, K.B., Henriksen, P.G., Baatrup, E., 2008. Male zebrafish (Danio rerio) courtship behaviour resists the feminising effects
of 17 alpha-ethinyloestradiol - morphological sexual characteristics do not. Aquat Toxicol 87, 234-244.

Lawrence, C., Ebersole, J.P., Kesseli, R.V., 2008. Rapid growth and out-crossing promote female development in zebrafish (Danio rerio).
Environ Biol Fish 81, 239-246.
Leal, M.C., Cardoso, E.R., Nobrega, R.H., Batlouni, S.R., Bogerd, J., Franca, L.R., Schulz, R.W., 2009. Histological and stereological evaluation
of zebrafish (Danio rerio) spermatogenesis with an emphasis on spermatogonial generations. Biol Reprod 81, 177-187.
Leet, J.K., Gall, H.E., Sepulveda, M.S., 2011. A review of studies on androgen and estrogen exposure in fish early life stages: effects on gene and
hormonal control of sexual differentiation. J Appl Toxicol 31, 379-398.
Leusch, F.D.L., Chapman, H.F., van den Heuvel, M.R., Tan, B.L.L., Gooneratne, S.R., Tremblay, L.A., 2006. Bioassay-derived androgenic and
estrogenic activity in municipal sewage in Australia and New Zealand. Ecotoxicol Environ Safe 65, 403-411.
Liew, W.C., Bartfai, R., Lim, Z.J., Sreenivasan, R., Siegfried, K.R., Orban, L., 2012. Polygenic sex determination system in zebrafish. PLoS
ONE 7(4): e34397.
Liew, W.C., Orban, L., 2013. Zebrafish sex: a complicated affair. Brief Funct Genomics 13:172-187.
Maack, G., Segner, H., 2003. Morphological development of the gonads in zebrafish. J Fish Biol 62, 895-906.
Madureira, T.V., Rocha, M.J., Cruzeiro, C., Galante, M.H., Monteiro, R.A.F., Rocha, E., 2011. The toxicity potential of pharmaceuticals found in
the Douro River estuary (Portugal): Assessing impacts on gonadal maturation with a histopathological and stereological study of zebrafish ovary
and testis after sub-acute exposures. Aquat Toxicol 105, 292-299.

Malison, J.A., Best, C.D., Kayes, T.B., Amundson, C.H., Wentworth, B.C., 1985. Hormonal growth promotion and evidence for a size-related
difference in response to estradiol-17-beta in Yellow Perch (Perca flavescens). Can J Fish Aquat Sci 42, 1627-1633.
Malison, J.A., Kayes, T.B., Wentworth, B.C., Amundson, C.H., 1988. Growth and feeding responses of male versus female Yellow Perch (Perca
flavescens) treated with estradiol-17β. Can J Fish Aquat Sci 45, 1942-1948.
Micael, J., Reis-Henriques, M.A., Carvalho, A.P., Santos, M.M., 2007. Genotoxic effects of binary mixtures of xenoandrogens (tributyltin,
triphenyltin) and a xenoestrogen (ethinylestradiol) in a partial life-cycle test with Zebrafish (Danio rerio). Environ Int 33, 1035-1039.
Navarro-Martin, L., Blazquez, M., Piferrer, F., 2009. Masculinization of the European sea bass (Dicentrarchus labrax) by treatment with an
androgen or aromatase inhibitor involves different gene expression and has distinct lasting effects on maturation. Gen Comp Endocrinol 160, 3-
11.
Nolan, M., Jobling, S., Brighty, G., Sumpter, J.P., Tyler, C.R., 2001. A histological description of intersexuality in the roach. J Fish Biol 58, 160-
176.
Olmstead, A.W., Kosian, P.A., Korte, J.J., Holcombe, G.W., Woodis, K.K., Degitz, S.J., 2009. Sex reversal of the amphibian, Xenopus
tropicalis, following larval exposure to an aromatase inhibitor. Aquat Toxicol 91, 143-150.
Orban, L., Sreenivasan, R., Olsson, P.E., 2009. Long and winding roads: Testis differentiation in zebrafish. Mol Cell Endocrinol 312, 35-41.

Örn, S., Holbech, H., Madsen, T.H., Norrgren, L., Petersen, G.I., 2003. Gonad development and vitellogenin production in zebrafish (Danio
rerio) exposed to ethinylestradiol and methyltestosterone. Aquat Toxicol 65, 397-411.
Panter, G.H., Hutchinson, T.H., Hurd, K.S., Sherren, A., Stanley, R.D., Tyler, C.R., 2004. Successful detection of (anti-) androgenic and
aromatase inhibitors in pre-spawning adult fathead minnows (Pimephales promelas) using easily measured endpoints of sexual development.
Aquat Toxicol 70, 11-21.
Pawlowski, S., van Aerle, R., Tyler, C.R., Braunbeck, T., 2004. Effects of 17 alpha-ethinylestradiol in a fathead minnow (Pimephales promelas)
gonadal recrudescence assay. Ecotoxicol Environ Safe 57, 330-345.
Peranandam, R., Palanisamy, I., Lourdaraj, A.V., Natesan, M., Vimalananthan, A.P., Thangaiyan, S., Perumal, A., Muthukalingan, K., 2014.
TBT Effects on the development of intersex (ovotestis) in female fresh water prawn Macrobrachium rosenbergii. Biomed Res Int, 412619.
Piferrer, F., Zanuy, S., Carrillo, M., Solar, I.I., Devlin, R.H., Donaldson, E.M., 1994. Brief treatment with an aromatase inhibitor during sex-
differentiation causes chromosomally female salmon to develop as normal, functional males. J Exp Zool 270, 255-262.
Randak, T., Zlabek, V., Pulkrabova, J., Kolarova, J., Kroupova, H., Siroka, Z., Velisek, J., Svobodova, Z., Hajslova, J., 2009. Effects of pollution
on chub in the River Elbe, Czech Republic. Ecotoxicol Environ Safe 72, 737-746.
Roberts, S., Barry, T., Malison, J., Goetz, F., 2004. Production of a recombinantly derived growth hormone antibody and the characterization of
growth hormone levels in yellow perch. Aquaculture 232, 591-602.

Rocha, M.J., Cruzeiro, C., Ferreira, C., Rocha, E., 2012. Occurrence of endocrine disruptor compounds in the estuary of the Iberian Douro River
and nearby Porto Coast (NW Portugal). Toxicol Environ Chem 94, 252-261.
Rocha, S., Domingues, V.F., Pinho, C., Fernandes, V.C., Delerue-Matos, C., Gameiro, P., Mansilha, C., 2013. Occurrence of bisphenol A,
estrone, 17β-estradiol and 17 α-ethinylestradiol in Portuguese rivers. Bull Environ Contam Toxicol 90, 73-78.
Santos, E.M., Kille, P., Workman, V.L., Paull, G.C., Tyler, C.R., 2008. Sexually dimorphic gene expression in the brains of mature zebrafish.
Comp Biochem Physiol A 149, 314-324.
Santos, M.M., Micael, M., Carvalho, A.P., Morabito, R., Booy, P., Massanisso, P., Lamoree, M., Reis-Henriques, M.A., 2006. Estrogens
counteract the masculinizing effect of tributyltin in zebrafish. Comp Biochem Physiol C 142, 151-155.
Scholz, S., Kluver, N., 2009. Effects of endocrine disrupters on sexual, gonadal development in fish. Sex Dev 3, 136-151.
Selman, K., Wallace, R.A., Sarka, A., Qi, X.P., 1993. Stages of oocyte development in the zebrafish, Brachydanio Rerio. J Morphol 218, 203-
224.
Sfakianakis, D.G., Leris, I., Mylonas, C.C., Kentouri, M., 2012. Temperature during early life determines sex in zebrafish, Danio rerio
(Hamilton, 1822). J Biol Res Thessalon 17, 68-73.

Silva, P., Rocha, M.J., Cruzeiro, C., Malhao, F., Reis, B., Urbatzka, R., Monteiro, R.A.F., Rocha, E., 2012. Testing the effects of ethinylestradiol
and of an environmentally relevant mixture of xenoestrogens as found in the Douro River (Portugal) on the maturation of fish gonads-A
stereological study using the zebrafish (Danio rerio) as model. Aquat Toxicol 124, 1-10.
Soares, J., Coimbra, A.M., Reis-Henriques, M.A., Monteiro, N.M., Vieira, M.N., Oliveira, J.M.A., Guedes-Dias, P., Fontainhas-Fernandes, A.,
Parra, S.S., Carvalho, A.P., Castro, L.F.C., Santos, M.M., 2009. Disruption of zebrafish (Danio rerio) embryonic development after full life-
cycle parental exposure to low levels of ethinylestradiol. Aquat Toxicol 95, 330-338.
Soto, A.M., Calabro, J.M., Prechtl, N.V., Yau, A.Y., Orlando, E.F., Daxenberger, A., Kolok, A.S., Guillette, L.J., le Bizec, B., Lange, I.G.,
Sonnenschein, C., 2004. Androgenic and estrogenic activity in water bodies receiving cattle feedlot effluent in eastern Nebraska, USA. Environ
Health Persp 112, 346-352.
Strussmann, C.A., Nakamura, M., 2002. Morphology, endocrinology, and environmental modulation of gonadal sex differentiation in teleost
fishes. Fish Physiol Biochem 26, 13-29.
Takahashi, H., 1977. Juvenile hermaphroditism in the zebrafish, Brachydanio rerio. Bull Fac Fish Hokkaido Univ 28, 57– 65.
Tejeda-Vera, R., Lopez-Lopez, E., Sedeno-Diaz, J.E., 2007. Biomarkers and bioindicators of the health condition of Ameca splendens and
Goodea atripinnis (Pisces: Goodeaidae) in the Ameca River, Mexico. Environ Int 33, 521-531.

Tzchori, I., Degani, G., Elisha, R., Eliyahu, R., Hurvitz, A., Vaya, J., Moav, B., 2004. The influence of phytoestrogens and oestradiol-17 beta on
growth and sex determination in the European eel (Anguilla anguilla). Aquacult Res 35, 1213-1219.
Uchida, D., Yamashita, M., Kitano, T., Iguchi, T., 2002. Oocyte apoptosis during the transition from ovary-like tissue to testes during sex
differentiation of juvenile zebrafish. J Exp Biol 205, 711-718.
Uchida, D., Yamashita, M., Kitano, T., Iguchi, T., 2004. An aromatase inhibitor or high water temperature induce oocyte apoptosis and depletion
of P450 aromatase activity in the gonads of genetic female zebrafish during sex-reversal. Comp Biochem Physiol A 137, 11-20.
Urbatzka, R., Rocha, E., Reis, B., Cruzeiro, C., Monteiro, R.A.F., Rocha, M.J., 2012. Effects of ethinylestradiol and of an environmentally
relevant mixture of xenoestrogens on steroidogenic gene expression and specific transcription factors in zebrafish. Environ Pollut 164, 28-35.
van der Ven, L.T.M., van den Brandhof, E.J., Vos, J.H., Wester, P.W., 2007. Effects of the estrogen agonist 17 β-estradiol and antagonist
tamoxifen in a partial life-cycle assay with zebrafish (Danio rerio). Environ Toxicol Chem 26, 380-380.
Versonnen, B.J., Arijs, K., Verslycke, T., Lema, W., Janssen, C.R., 2003. In vitro and in vivo estrogenicity and toxicity of o-, m-, and p-
dichlorobenzene. Environ Toxicol Chem 22, 329-335.
Versonnen, B.J., Janssen, C.R., 2004. Xenoestrogenic effects of ethinylestradiol in zebrafish (Danio rerio). Environ Toxicol 19, 198-206.

Villeneuve, D.L., Mueller, N.D., Martinovic, D., Makynen, E.A., Kahl, M.D., Jensen, K.M., Durhan, E.J., Cavallin, J.E., Bencic, D., Ankley,
G.T., 2009. Direct effects, compensation, and recovery in Female Fathead Minnows exposed to a model aromatase inhibitor. Environ Health
Persp 117, 624-631.
Wang, X.G., Bartfai, R., Sleptsova-Freidrich, I., Orban, L., 2007. The timing and extent of 'juvenile ovary' phase are highly variable during
zebrafish testis differentiation. J Fish Biol 70, 33-44.
Westerfield, M., 2000. The Zebrafish Book. A guide for the laboratory use of zebrafish (Danio rerio), 4th ed. University of Oregon Press,
Eugene.
Yamamoto, T., 1969. Sex differentiation, in: Hoar, W., Randall, D. (Eds.), Fish Physiology. Academic Press, pp. 117– 175.
Zhang, X.W., Hecker, M., Park, J.W., Tompsett, A.R., Jones, P.D., Newsted, J., Au, D.W.T., Kong, R., Wu, R.S.S., Giesy, J.P., 2008. Time-
dependent transcriptional profiles of genes of the hypothalamic-pituitary-gonadal axis in medaka (Oryzias Latipes) exposed to fadrozole and 17
beta-trenbolone. Environ Toxicol Chem 27, 2504-2511.
Zou, J.J., Trudeau, V.L., Cui, Z., Brechin, J., Mackenzie, K., Zhu, Z., Houlihan, D.F., Peter, R.E., 1997. Estradiol stimulates growth hormone
production in female goldfish. Gen Comp Endocrinol 106, 102-112.

Figure legends
Fig. 1. Different germ cell types of zebrafish (D. rerio) observed during the oogenesis: A – Undifferentiated gonocytes (arrows), these were
present in both genders at 35 dpf; B – Oogonium stage; C – Primary oocyte stage; D – Cortical alveolus stage; E – Vitellogenic stage; and F –
Mature stage.
Fig. 2. Different germ cell types of zebrafish (D. rerio) observed during the spermatogenic process: S – Spermatogonia; ES – Early
spermatocytes; LS – Late spermatocytes; SPs – spermatids; SZ – Spermatozoa; and L – Lumen.
Fig. 3. Zebrafish (D. rerio) sex differentiation at 35 dpf. Histological study of females (A, B) and of males showing the two differentiation
processes (C to H). The testis differentiation process in “type III/II” involved the transformation of the “juvenile ovary” into testis (C to F),
whereas in “type I” this structure was never formed before testis differentiation (G, H). Figures B, D, F and H are, respectively, magnifications of
A, C, E and G. PO – Primary oocyte; ES – Early spermatocytes; IT– Interstitial tissue and ST– Stroma.
Fig. 4. Zebrafish (D. rerio) sex ratios observed after exposures to 17α-ethinylestradiol (EE2: 4 ng/L); fadrozole (Fad: 50 μg/L) and 17α-
ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L), at 35 and 60 dpf. Data are expressed as the percent of males (M) and females (F)
(n=30/treatment). Statistical analysis was performed using the Chi-square test. *** represents highly significant differences to control
(p<0.0001).

Fig. 5. Percentages of the different gonad types of zebrafish (D. rerio) males observed after exposures to 17α-ethinylestradiol (EE2: 4 ng/L),
fadrozole (Fad: 50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L), at 35 dpf. Data are expressed as the percentage of males
with type I, II, III and differentiated gonads.
Fig. 6. Relative volumes of ovary compartments of zebrafish (D. rerio) after exposures to 17α-ethinylestradiol (EE2: 4 ng/L), fadrozole (Fad: 50
μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L), at 60 dpf. Values are presented as mean ± standard deviation (SD), n=5
(Control and EE2), n=3 (Fad and Mix, where only 3 females were identified at 60 dpf). Statistical analysis was carried out using a one-way
factorial ANOVA. Different lower case letters represent significant differences between treatments (p < 0.05) considering the same structural
compartment. Absence of superscript indicates no significant difference between treatments.
Fig. 7. Relative volumes of the testis compartments of zebrafish (D. rerio) after exposures to 17α-ethinylestradiol (EE2: 4 ng/L), fadrozole (Fad:
50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L), at 60 dpf. Values are presented as mean ± standard deviation (SD), n=5
(Control and EE2), n=7 (Fad and Mix). Statistical analysis was carried out using a one-way factorial ANOVA. Different lower case letters
represent significant differences between treatments (p < 0.05) considering the same structural compartment. Absence of superscript indicates no
significant difference between treatments.

Fig. 8. Zebrafish (D. rerio) ovary sections: A – Control; B – 17α-ethinylestradiol (EE2: 4 ng/L); C – fadrozole (Fad: 50 μg/L); and D – 17α-
ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L). EE2 stimulus enhanced zebrafish growth and gonad development (B, D); Fad stimulus
induced a delay in the gonad development (C). PO – primary oocyte; CAO – cortical-alveolar oocyte; and VO – vitellogenic oocyte.
Fig. 9. Zebrafish (D. rerio) testis sections of the two 60 dpf males where intersex was detected. The condition was identified by the presence of
primary oocytes (arrows). A – Male from fadrozole (Fad: 50 μg/L); and B – Male from 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L).
Fig. 10. Zebrafish (D. rerio) gonad sections (A and B) and relative volumes of gonad compartments (C, and D) of the two individuals that
showed abnormal development at 60 dpf: A and C — female exposed to Fad, showing a gonad similar to a “juvenile ovary”; B and D — male
exposed to the Mix, presenting a gonad similar to a “type I” male gonad. ST– Stroma S; PO – Primary oocyte; S – Spermatogonia; SZ –
Spermatozoa; ES – Early spermatocytes; and UG – Undifferentiated gonocyte.
Table 1 Zebrafish (D. rerio) female and male body weight (BW), total length (TL), gonad weight (GW), and gonadosomatic index (GSI), after exposures to 17α-ethinylestradiol (EE2: 4 ng/L), fadrozole (Fad: 50 μg/L), and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L+50 μg/L), at 35 and 60 dpf.

Values are presented as mean ±
standard deviation
(SD), n=30/treatment and age for BW and TL; n=10/treatment and age for GW and GSI.
Statistical analysis was performed using a two-way factorial ANOVA. Different lower case letters represent significant differences between treatments within gender (p < 0.05), and * represents significant differences between genders within treatments. Absence of superscript indicates no significant difference between treatments.
Table 2 Relative volumes (%) of zebrafish (D. rerio) ovary and testis germ cell compartments after exposures to 17α-ethinylestradiol (EE2: 4 ng/L), fadrozole (Fad: 50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L + 50 μg/L), at 35 dpf.
Age Group BW (mg) TL (cm) GW (µg) GSI (%)
Female Male Female Male Female Male Female Male
35 dpf Control 30.6±12.4 19.9±14.9 14.7±1.8 12.5±3.0 43.6±13.4a* 3.1±1.5 0.144±0.041a* 0.031±0.028 EE2 43.8±17.9* 26.6±10.61 16.0±2.0* 13.7±1.8 33.2±6.8ab* 7.8±4.6 0.072±0.034b* 0.026±0.016 Fad 24.6±5.9 20.4±13.9 13.8±1.0 12.8±2.8 19.2±10.1b 4.6±2.8 0.071±0.018b 0.028±0.019 Mix 31.5±10.7 32.7±13.9 14.2±2.1 14.4±2.2 25.5±16.5ab 10.1±11.3 0.076±0.016b 0.030±0.027 60 dpf Control 89.7±37.1a 88.2±39.4a 21.6±3.6a 21.1±3.4a 101.9±80.1a 45.8±18.9a 0.127±0.041* 0.042±0.012 EE2 175.5±43.7b 173.8±43.4b 27.2±2.2b 27.2±2.1b 228.9±743b* 118.0±23.2b 0.144±0.073* 0.079±0.003 Fad 89.1±14.3a 92.3±31.3a 22.3±1.4ac 21.9±2.6a 110.4±10.2a 33.1±11.4a 0.121±0.005* 0.033±0.013 Mix 163.2±22.8b* 90.5±33.6a 25.9±1.9bc* 22.0±2.5a 206.9±25.7b* 36.4±31.2a 0.126±0.021* 0.046±0.049
Sex Group Stroma Interstitial Tissue
Undifferentiated gonocyte
Primary oocyte
Early spermatocyte
Late spermatocyte
Control 1.37±1.49 48.51±5.24 2.25±1.53a 48.33±6.47 N.D. N.D. EE2 0.59±0.67 53.18±6.72 0.16±0.24b 46.06±6.35 N.D. N.D. Fad 2.46±3.48 60.59±13.56 1.26±0.16ab 35.73±17.14 N.D. N.D.
Female
Mix 0.21±0.30 55.50±2.13 1.18±0.23ab 43.11±2.06 N.D. N.D. Male
Control 3.05±1.48* 65.12±14.55* 1.23±1.74 27.68±13.65* 2.91±4.12 0 EE2 3.11±1.14 63.59±2.09 5.19±0.82 23.32±1.39 4.79±1.26 0 Fad 2.90±1.80 60.32±9.84 1.02±2.03 31.49±7.94 4.27±5.04 0
“type III/II”
Mix 3.46±4.89 56.24±3.72 5.02±4.92 29.41±14.40 5.87±8.31 0
Control 46.85±2.28 29.73±5.61 14.39±4.57a 0.11±0.16 8.92±12.62 0 EE2 35.98±40.93 21.13±5.56 2.49±2.27b 1.65±1.46 22.20±21.28 16.55±14.70 Fad 11.34±12.53 35.59±15.91 10.05±8.09ab 0.36±0.62 24.86±21.63 17.80±15.77
“type I”
Mix 50.03±19.52 25.04±10.03 8.11±4.17ab 5.54±5.92 9.45±10.80 1.83±3.18

Values are presented as mean ± standard deviation (SD), n=10/treatment. Statistical analysis was performed using a one-way factorial ANOVA. Different lower case letters represent significant differences between treatments within gender (p < 0.05), and * represents significant differences between males “type III/II” and “type I” within the control group. Absence of superscript indicates no significant difference between treatments. Non-determined data (N.D.).
Supplementary 1 Absolute volumes (x10-3 mm3) of ovary and testis compartments of zebrafish (D. rerio) after exposures to 17α-ethinylestradiol (EE2: 4 ng/L), fadrozole (Fad: 50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L+50 μg/L), at 35 dpf.
Sex Group Gonad Stroma Interstitial Tissue
Undifferentiated gonocyte
Primary oocyte
Early spermatocyte
Late spermatocyte
Control 43.63±13.45a 0.28±0.34 21.40±7.70 0.94±0.76 21.00±6.83a N.D. N.D. EE2 33.24±6.75ab 0.19±0.20 17.61±3.85 0.05±0.06 15.40±4.47ab N.D. N.D. Fad 19.20±10.07b 0.30±0.42 10.95±3.50 0.23±0.09 7.72±6.89b N.D. N.D.
Female
Mix 25.52±16.51ab 0.03±0.04 14.59±10.00 0.07±0.09 10.83±6.59ab N.D. N.D. Male
Control 3.05±2.27a 0.07±0.02 1.82±1.03a 0.02±0.02a 0.99±1.04a 0.13±0.19 0.00 EE2 15.76±1.43bc 0.50±0.22 10.74±1.62b 0.92±0.35b 3.82±0.74ab 0.74±0.13 0.00 Fad 6.99±1.18b 0.19±0.17 4.00±0.92c 0.09±0.02a 2.48±0.59a 0.21±0.36 0.00
“type III/II”
Mix 21.86±13.11c 0.43±0.61 12.54±8.18b 0.77±0.42b 7.37±7.00b 0.74±0.10 0.00
Control 3.13±1.85 1.44±0.79ab 0.88±0.37ab 0.40±0.12 <0.01 0.39±0.56 0.00a EE2 5.49±0.49 1.90±2.00a 1.20±0.42a 0.14±0.14 0.09±0.07 1.3±1.2 0.94±0.83b Fad 2.14±0.44 0.21±0.20b 0.71±0.16ab 0.20±0.12 <0.01 0.59±0.51 0.43±0.39ab
“type I”
Mix 2.21±0.99 1.2±0.09ab 0.48±0.04b 0.16±0.07 0.15±0.21 0.15±0.15 0.03±0.04a

Values are presented as mean ± standard deviation (SD), n=10/treatment. Statistical analysis was performed using a one-way factorial ANOVA. Different lower case letters represent significant differences between treatments within gender (p < 0.05). Absence of superscript indicates no significant difference between treatments. Non-determined data (N.D.).
Supplementary 2 Absol
ute volu
mes (x10-3 mm3) of ovary and ovary compartments of zebrafish (D. rerio) after exposures to 17α-ethinylestradiol (EE2: 4 ng/L); fadrozole (Fad: 50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L+50 μg/L), at 60 dpf.
Values are presented as mean ± standard deviation (SD), n=5/Control, n=5/EE2, n=3/Fad and n=3/Mix (n is different because Fad and Mix only present 3 females at 60 dpf). Statistical analysis was performed using a one-way factorial ANOVA. Different lower case letters represent significant differences between treatments (p < 0.05). Absence of superscript indicates no significant difference between treatments.
Supplementary 3 Absolute volumes (x10-3 mm3) of testis and testis compartments of zebrafish (D. rerio) after exposures to 17α-ethinylestradiol (EE2: 4 ng/L); fadrozole (Fad: 50 μg/L) and 17α-ethinylestradiol + fadrozole (Mix: 4 ng/L+50 μg/L), at 60 dpf.
Group Ovary Oogonium Primary oocyte Cortical alveolus Vitellogenic Mature Atretic Interstitial tissue
Control 101.86±80.05a 0.10±0.10 43.43±21.49 11.26±22.05ab 13.58±23.43a 0.00 0.29±0.65a 33.20±23.88a EE2 228.88±74.34b 0.26±0.15 61.03±36.34 29.99±4.56a 53.26±34.39b 9.52±12.49 0.23±0.39a 74.02±15.23b Fad 110.44±10.17a 0.13±0.01 73.92±4.75 4.09±3.14b 0.00a 0.00 <0.01a 32.16±2.29a Mix 206.92±25.71b 0.26±0.02 96.11±19.48 32.74±9.89a 25.46±12.26ab 0.00 2.10±1.14b 50.24±7.45ab
Group Testis Spermatogonium Early spermatocyte Late spermatocyte Spermatid Spermatozoon Lumen Interstitial tissue
Control 45.76±18.90a 0.67±0.05ab 4.21±0.91a 15.40±7.22a 3.50±1.10a 13.80±7.71a 2.65±0.58a 5.53±1.82a EE2 118.02±23.17b 1.42±0.85b 11.22±5.39b 50.44±11.78b 8.58±2.22b 23.38±4.83b 7.36±0.86b 15.62±1.25b Fad 33.07±11.44a 0.35±0.14a 3.39±1.65a 13.02±4.43a 1.83±0.63a 6.65±4.25a 2.43±1.40a 5.12±2.24a Mix 36.36±31.24a 0.47±0.27a 4.79±2.17a 15.44±14.24a 1.92±2.32a 6.35±7.03a 2.22±2.23a 5.14±3.62a

Values are presented as mean ± standard deviation (SD), n=5/Control, n=5/EE2, n=7/Fad and n=7/Mix. Statistical analysis was performed using a one-way factorial ANOVA. Different lower case letters represent significant differences between treatments (p < 0.05). Absence of superscript indicates no significant difference between treatments.

Figure 1 .

Figure 10 .

Figure 2 .

Figure 3 .


Figure 4 .


Figure 5 .

Figure 6 .
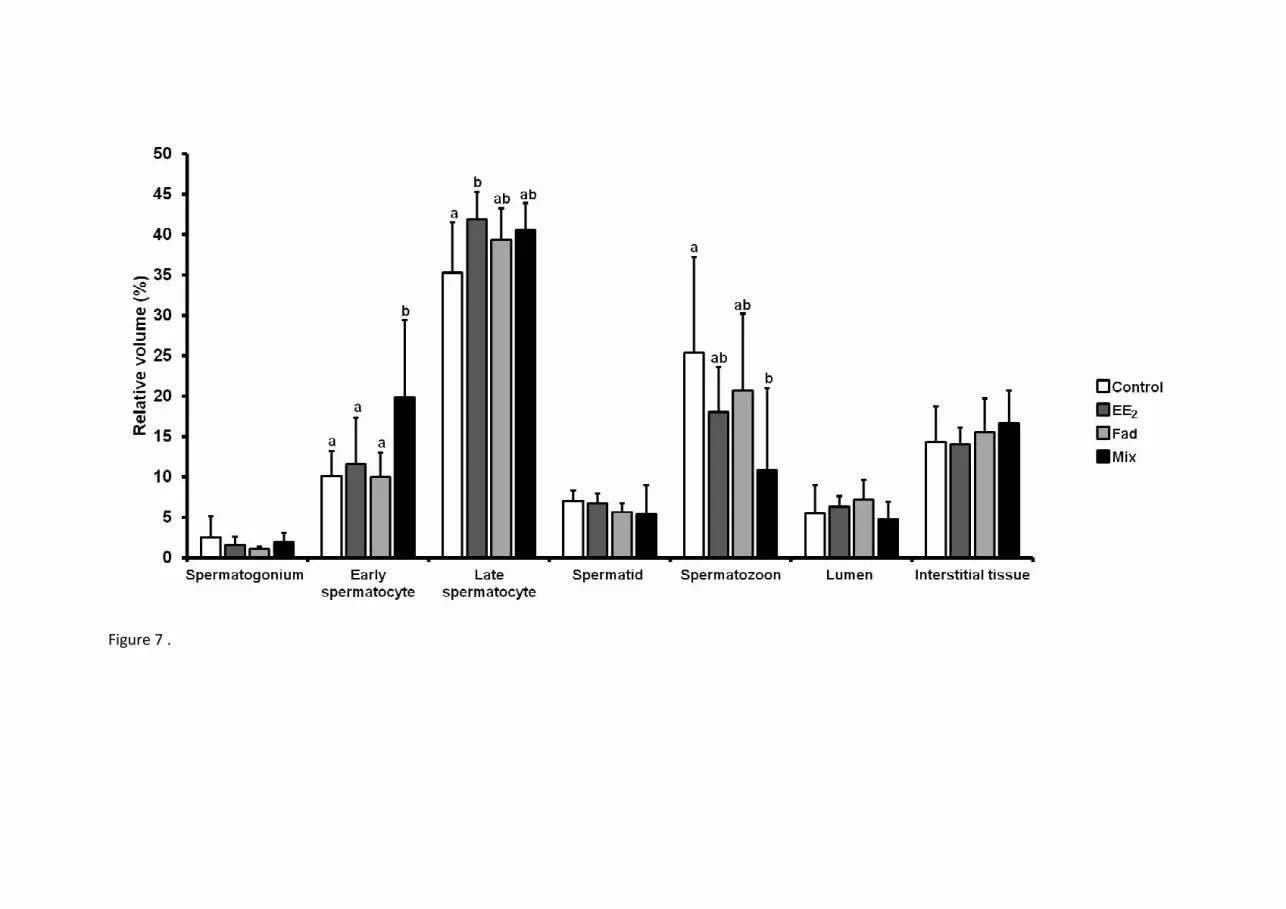
Figure 7 .

Figure 8 .

Figure 9 .