wd40 repeat proteins striatin and s/g2 nuclear autoantigen are members of a novel family of...
TRANSCRIPT

WD40 Repeat Proteins Striatin and S/G2 Nuclear Autoantigen AreMembers of a Novel Family of Calmodulin-binding Proteins ThatAssociate with Protein Phosphatase 2A*
Carlos S. Moreno‡, Susan Park§,¶, Kasey Nelson‡, Danita Ashby‡, Frantisek Hubalek‡,William S. Lane∥, and David C. Pallas‡,**
‡Department of Biochemistry, Emory University School of Medicine, Atlanta, Georgia 30322§Division of Cellular and Molecular Biology, Dana-Farber Cancer Institute, Boston, Massachusetts02115∥Harvard Microchemistry Facility, Harvard University, Cambridge, Massachusetts 02138
AbstractProtein phosphatase 2A (PP2A) is a multifunctional serine/threonine phosphatase that is critical tomany cellular processes including development, neuronal signaling, cell cycle regulation, and viraltransformation. PP2A has been implicated in Ca2+-dependent signaling pathways, but how PP2Ais targeted to these pathways is not understood. We have identified two calmodulin (CaM)-bindingproteins that form stable complexes with the PP2A A/C heterodimer and may represent a novelfamily of PP2A B-type subunits. These two proteins, striatin and S/G2 nuclear autoantigen(SG2NA), are highly related WD40 repeat proteins of previously unknown function and distinctsubcellular localizations. Striatin has been reported to associate with the postsynaptic densities ofneurons, whereas SG2NA has been reported to be a nuclear protein expressed primarily during theS and G2 phases of the cell cycle. We show that SG2NA, like striatin, binds to CaM in a Ca2+-dependent manner. In addition to CaM and PP2A, several unidentified proteins stably associatewith the striatin-PP2A and SG2NA-PP2A complexes. Thus, one mechanism of targeting andorganizing PP2A with components of Ca2+-dependent signaling pathways may be through themolecular scaffolding proteins striatin and SG2NA.
PP2A,1 an essential serine/threonine protein phosphatase found in all eukaryotic cells,regulates a wide variety of important cellular events, including DNA replication,transcription, translation, development, neuronal signaling and progression of the cell cycle(for reviews see Refs. 1–3). The PP2A heterotrimer consists of a catalytic (C) subunit, astructural (A) subunit, and a regulatory (B-type) subunit (4). Although relatively few C- andA-type subunits have been identified, multiple B-type subunits exist, including B (or B55),B′ (or B56), and B″ (or PR72/130) classes (5–9). To enable utilization of this phosphatasefor numerous substrates in different pathways, PP2A is regulated at multiple levels,
*This work was supported by National Institutes of Health Grant CA57327.
© 2000 by The American Society for Biochemistry and Molecular Biology, Inc.**To whom correspondence should be addressed: Dept. of Biochemistry, Emory University School of Medicine, 1510 Clifton Rd.,Atlanta, GA 30322. Tel.: 404-727-5620; Fax: 404-727-3231; [email protected].¶Present address: Dept. of Surgical Research, Children’s Hospital, 320 Longwood Ave., Boston, MA 02115.
The terms of this arrangement have been reviewed and approved by Emory University in accordance with its conflict of interestpolicies.1The abbreviations used are: PP2A, protein phosphatase 2A; SG2NA, S/G2 nuclear autoantigen; CaM, calmodulin; 2D, two-dimensional; mAb, monoclonal antibody; CaMKII/IV, CaM-dependent kinase II/IV; HA, hemagglutinin; MT, middle tumor antigen;MS, mass spectrometry; KLH, keyhole limpet hemocyanin; PAGE, polyacrylamide gel electrophoresis; wt, wild type.
NIH Public AccessAuthor ManuscriptJ Biol Chem. Author manuscript; available in PMC 2012 November 23.
Published in final edited form as:J Biol Chem. 2000 February 25; 275(8): 5257–5263.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

including covalent modifications, interaction with inhibitory proteins and lipids, andassociation with the various B-type subunits. For example, B′ subunits were recently shownto target PP2A to the adenomatous polyposis coli tumor suppressor scaffolding protein,physically associating PP2A with specific substrates and thus regulating Wnt-β-cateninsignaling (10).
PP2A has also been shown to form complexes with CaM-dependent kinase IV (CaMKIV)(11), suggesting a role for PP2A in Ca2+-dependent signaling. This possibility is furthersupported by patch clamp experiments with both neuronal (12) and smooth muscle cells (13)that have used both okadaic acid and recombinant PP2A C subunit to implicate PP2A in theregulation of calcium-activated potassium channels and L-type Ca2+ channels (14).
To better understand how PP2A is targeted to various microenvironments and signaltransduction pathways within the cell, we have looked for additional PP2A targetingsubunits. Here we report the identification of two PP2A-associated proteins that mayrepresent a novel family of B-type subunits. These two proteins contain WD40 repeats andbind to CaM in a calcium-dependent manner. One member of this family, striatin, islocalized to the post-synaptic densities of neuronal dendrites (15), whereas the other,SG2NA, has been reported to be localized to the nucleus (16). Striatin-PP2A and SG2NA-PP2A complexes contain several additional unidentified proteins, suggesting that striatin andSG2NA may function as scaffolding proteins involved in Ca2+-dependent signaltransduction pathways.
EXPERIMENTAL PROCEDURESMetabolic Labeling and Immunoprecipitations
For metabolic labeling of NIH3T3 cells with methionine, subconfluent dishes of cells werelabeled for 4–6 h with 0.25 mCi/ml [35S]methionine in methionine-free Dulbecco’smodified Eagle’s medium supplemented with dialyzed 0.5% fetal calf serum. Cells werewashed twice with phosphate-buffered saline and once with IP wash buffer (0.135 M NaCl,1% glycerol, 20 mM Tris, pH 8.0) and then were lysed in 1 ml of IP lysis buffer (1% NonidetP-40, 0.135 M NaCl, 1% glycerol, 20 mM Tris, pH 8.0, 0.03 units/ml aprotinin, and 1–2 mM
phenylmethylsulfonyl fluoride) while rocking for 20 min at 4 °C. Lysates were cleared at13,000 × g for 10 min at 4 °C and then incubated at 4 °C for 90 min while rocking in 1.5-mlEppendorf tubes with protein A-Sepharose and the appropriate antisera. Immune complexeswere precipitated by centrifugation for 1 min at 700 × g, and the supernatants were removed.Immune complexes were washed twice with 1 ml of IP lysis buffer and three times with 1ml of phosphate-buffered saline. Two-dimensional gel electrophoresis was performed asdescribed previously (17), and proteins were transferred to nitrocellulose membranes.Autoradiographs were performed with Biomax MS Film (Kodak).
Preparative ImmunopurificationUsing affinity-purified AR-1 antisera, samples were immunopurified from 40 15-cm dishesof polyomavirus MT-transformed NIH3T3 cells as described (17) except that the IP lysisbuffer contained 20% Triton and 1% glycerol. For preparative immunoprecipitations, PP2Acomplexes were immunoaffinity purified using the 1d6 anti-C subunit monoclonal antibody(mAb) chemically cross-linked to protein A-Sepharose as described previously (18) exceptthat 10 15-cm dishes of cells were used for one batch purification and whole immunecomplexes were analyzed on two-dimensional (2D) gels (17). Control immunopurificationswere performed with 7-34-1 mAb chemically cross-linked to protein A-Sepharose using five15-cm dishes of cells. Proteins were visualized with Coomassie Brilliant Blue R250 (Bio-Rad).
Moreno et al. Page 2
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Ion Trap Mass SpectroscopySpecific spots at 93 and 110 kDa were subjected to in-gel reduction,carboxyamidomethylation, and tryptic digestion (Promega). Multiple peptide sequenceswere determined in a single run by microcapillary reverse-phase chromatography directlycoupled to a Finnigan LCQ quadrupole ion trap mass spectrometer. The ion trap wasprogrammed to acquire successive sets of three scan modes consisting of full scan MS overalternating ranges of 395–800 m/z or 800–1300 m/z, followed by two data-dependent scanson the most abundant ion in those full scans. These data-dependent scans allowed: 1) theautomatic acquisition of a high resolution (zoom) scan to determine charge state and exactmass and 2) MS/MS spectra for peptide sequence information. MS/MS spectra wereacquired with a relative collision energy of 30%, an isolation width of 2.5 daltons, anddynamic exclusion of ions from repeat analysis. Interpretation of the resulting MS/MSspectra of the peptides was facilitated by programs developed in the HarvardMicrochemistry Facility and by data base correlation with the algorithm SEQUEST (19, 20).
AntibodiesLasergene DNASTAR Protean software was utilized to identify highly hydrophilic andantigenic sequences for selection of peptide antigens. Rabbit polyclonal pan-B′ (AR-1),striatin, and SG2NA antisera were generated using keyhole limpet hemocyanin (KLH)-conjugated peptides as immunogens. Peptide DP47 (ELFDSEDPRERDFLKTC)corresponds to residues 194–209 of the human α-isoform of B′ (B56) (7) with an additionalcarboxyl-terminal cysteine for coupling to KLH. Peptide DP52(GESPKQKGQEIKRSSGDC) corresponds to residues 227–243 of SG2NA with a carboxyl-terminal cysteine for coupling. Peptide DP53 (SVGSPSRPSSSRLPEC) corresponds toresidues 373–387 of striatin with an added carboxyl-terminal cysteine for coupling. Peptideswere conjugated to KLH using the Imject maleimide KLH conjugation kit (Pierce)according to the manufacturer’s instructions. The methylation-sensitive PP2A C subunitmAb, 4b7, was generated to a 15-residue unmethylated carboxyl-terminal peptide with anadditional amino-terminal cysteine added for coupling to KLH (18).2 Anti-B″ (PR72/130)polyclonal antibodies were provided by Brian Hemmings. 3 A methylation-insensitive mAbto PP2A C subunit was obtained from Transduction Laboratories (Lexington, KY). Thecontrol mAb 7-34-1 (American Type Culture Collection) is directed against majorhistocompatibility complex class I swine leukocyte antigen.
Calmodulin-Sepharose PrecipitationsNIH3T3 cells were treated for 30 min with 1 µM ionomycin, washed twice with 8 ml of TNCbuffer (50 mM Tris, pH 7.5, 100 mM NaCl, 2 mM CaCl2) and once with 8 ml of IP wash buffercontaining 2 mM CaCl2, and lysed with 1 ml of IP lysis buffer containing 2 mM CaCl2.Untreated cells were washed twice with 8 ml of TNE buffer (50 mM Tris, pH 7.5, 100 mM
NaCl, 1 mM EGTA) and once with 8 ml of IP wash buffer containing 1 mM EGTA and thenlysed with 1 ml of IP lysis buffer containing 1 mM EGTA. Calmodulin-Sepharoseprecipitations were performed as for immunoprecipitations described above, except thatcomplexes from ionomycin-treated cells were washed in IP lysis buffer with 2 mM CaCl2 andTNC, and those from untreated cells were washed in IP lysis buffer with 1 mM EGTA andTNE.
2X. X. Yu, X. Du, R. E. Green, E. Ogris, O. Feng, L. Chou, and D. C. Pallas, manuscript in preparation.3B. Hemmings, unpublished observation.
Moreno et al. Page 3
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Cell CultureAll cells were cultured in Dulbecco’s modified Eagle’s medium, 10% bovine calf serum(Life Technologies, Inc.). The medium for NIH3T3 cells stably expressing hemagglutinin(HA)-tagged B subunit2 was supplemented with 100 µg/ml hygromycin. In the case of theGRE only, 36wt, 301Stop, and T304A cell lines (22), the medium was supplemented with100 µg/ml hygromycin and 25 µg/ml geneticin.
RESULTS AND DISCUSSIONIdentification of 110- and 93-kDa PP2A-associated Proteins as Striatin and SG2NA
In an effort to discover previously unknown members of the B′ family of PP2A subunits, apan-B′ subunit antibody termed AR-1 was generated against a sequence highly conserved inall known B′ subunits (Figs. 1A and 2A). AR-1 recognized known B′ subunits thatmigrated at 56 and 74 kDa, as well as three unknown proteins that migrated at 93, 110, and131 kDa (Fig. 1A). In a parallel attempt to identify PP2A-associated proteins, PP2Aimmunoprecipitations were prepared from [35S]methionine-labeled NIH3T3 cells using amonoclonal antibody (1d6) directed against the carboxyl terminus of the PP2A C subunit.Several specific spots were observed on 2D gels, including two at 93 and 110 kDa (Fig. 1B).To determine whether the 93- and 110-kDa proteins in 1d6 immunoprecipitates were thesame proteins detected in AR-1 immunoblots of whole cell lysates (Fig. 1A), 1d6immunoprecipitates were immunoblotted with AR-1 (Fig. 1C, lanes 1 and 2). The twoproteins that migrated at 93 and 110 kDa were specifically recognized by the AR-1 antisera,suggesting that they might be novel B′-type subunits.
To identify the 110-kDa protein, a large scale direct immunoprecipitation using the AR-1antisera was subjected to 2D gel electrophoresis, and peptide sequences were obtained byion trap mass spectrometry (Fig. 1D). These sequences corresponded to a previously clonedgene of unknown function, striatin, which was originally purified from rat brain (15).Striatin has been reported to bind CaM with a 40 nM KD in a calcium-dependent manner at ahalf-maximal Ca2+ concentration of 0.5 µM (23). Moreover, striatin contains two polybasicdomains that may facilitate association with the post-synaptic membrane (15).Immunolabeling has shown that striatin is excluded from neuronal axons but is foundthroughout dendrites and is abundant in the post-synaptic densities of neuronal dendriticspines (15). These data suggest that striatin targets PP2A to a cellular microenvironment inwhich it may play a role in the modulation of calcium-dependent neuronal signaling.Although striatin was originally described as a brain-specific protein, we have observedstriatin protein in murine NIH3T3 fibroblasts and human Jurkat T lymphocytes. We havealso detected striatin mRNA in human HeLa cervical cancer cells, and expressed sequencetags (ESTs) were found in the dBEST data base that represent partial striatin cDNAs fromhuman B lymphocytes, human heart, murine myotubules, and murine testis (data notshown). Thus, striatin is present in both dividing and nondividing cells and is much morewidely expressed than previously thought.
To identify the 93-kDa protein and obtain further peptide sequence from the 110-kDaprotein, large scale PP2A (1d6) immunoprecipitations were subjected to 2D gelelectrophoresis. Mass spectrometric sequencing revealed multiple additional peptides fromstriatin for the 110-kDa protein and identified the 93-kDa protein to be a highly relatedprotein, S/G2 nuclear autoantigen (SG2NA) (Fig. 1D). Little is known about SG2NA otherthan that it is localized to the nucleus, it contains WD40 repeats, and as assayed byimmunofluorescence, its expression appears to be cell cycle-regulated, peaking during the Sand G2 phases (16). Striatin and SG2NA bear little homology to the B, B′, or B″ subunits,raising the possibility that they might comprise a new family of PP2A B-type subunits. At
Moreno et al. Page 4
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

least one homolog of striatin and SG2NA exists in Caenorhabditis elegans (GenBank™accession no. CAA94873) (24), suggesting that this form of PP2A may play an importantrole in all metazoans. Although there is no obvious homolog for either of these proteins inyeast, potential WD40-containing open reading frames of unknown function with somehomology to striatin and SG2NA do exist in both Saccharomyces cerevisiae, (GenBank™accession no. CAA89144) and Schizosaccharomyces pombe (GenBank™ accession no.CAA21906).
Striatin and SG2NA Share a Conserved Epitope with B′ SubunitsBecause striatin and SG2NA showed no obvious homology to PP2A B′ subunits, it waspuzzling that they were recognized efficiently by the pan-B′ antiserum, AR-1. However, acareful comparison revealed that, whereas neither striatin nor SG2NA contains the preciseconsensus sequence used to generate the AR-1 antisera, they both contain sequences thatshare some homology with this motif at positions corresponding to striatin 277–294 and551–566 (Fig. 2B). To determine whether the AR-1 antisera recognized either or both ofthese sequences, two peptides were synthesized corresponding to these two sequences aswell as two control peptides corresponding to randomized sequences containing the sameamino acids. A dot blot (Fig. 2C) demonstrated that the AR-1 antisera recognizes theSTR277 peptide corresponding to striatin 277–294 but not the striatin 551–566 peptide orthe control peptides. The STR277 peptide was also tested to determine whether it couldblock immunoprecipitations with the AR-1 antisera (Fig. 2D). As expected, both theSTR277 peptide and the AR-1 peptide used to generate the AR-1 antisera were effective atblocking immunoprecipitation of both striatin and SG2NA, whereas the randomized controlpeptide did not block AR-1 immunoprecipitations. The conservation of the sequence (D/E)X2D(S/T)X(D/E)X1–2(R/K)EX(D/E)(F/Y)LXT between most B′ subunits and striatin andSG2NA suggests that it may be involved in interactions between these proteins and the A/Cheterodimer. Consistent with this hypothesis, AR-1 was able to immunoprecipitate striatinand SG2NA only in the presence of 20% Triton. Under these conditions, little or none of thePP2A A and C subunits are co-immunoprecipitated (data not shown), suggesting that 20%Triton may have dissociated striatin and SG2NA from the A/C heterodimer, revealing theAR-1-specific epitope.
To facilitate the investigation of striatin and SG2NA interactions with the A/C heterodimer,polyclonal antisera were raised to both proteins. The striatin antiserum was raised against apeptide antigen corresponding to amino acids 373–387 of human striatin. This sequence islocated within a basic domain that may be important for striatin association with the cellularmembrane (15). Although this region is 100% identical between mouse and human striatin,it is not found in SG2NA (Fig. 1D). The SG2NA antisera was raised against a peptideantigen corresponding to amino acids 227–243 of human SG2NA in a region that has littlehomology with striatin sequences. 1d6 immunoprecipitations of PP2A were probed with theanti-striatin and anti-SG2NA antisera, confirming that these antisera recognize the 110- and93-kDa proteins, respectively (Fig. 1C).
PP2A A and C Subunits Coimmunoprecipitate with Striatin and SG2NATo confirm that striatin and SG2NA form stable complexes with PP2A, striatin and SG2NAcomplexes were immunoprecipitated with the anti-striatin and anti-SG2NA antisera,respectively, analyzed by SDS-PAGE, and immunoblotted with a commercially obtainedmAb to PP2A C subunit (Fig. 3A). The C subunit was present in immunoprecipitations ofboth striatin and SG2NA but not in immunoprecipitations using preimmune sera from thesame rabbits. The presence of A subunit was similarly detected (Fig. 3A).
Moreno et al. Page 5
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Several lines of evidence support the observation that no other B-type subunits were presentin striatin-A/C or SG2NA-A/C complexes. First, 2D analysis of striatin and SG2NAimmune complexes labeled with [35S]methionine or stained with Coomassie Blue showedno spots that corresponded to the known migration positions of the B, B′, or B″ subunits(Fig. 4B and data not shown). Second, it has previously been shown (22) that the 1d6 mAbused to precipitate the striatin-A/C or SG2NA-A/C complexes does not immunoprecipitateany B subunit. Third, immunoblots of striatin and SG2NA immune complexes with theAR-1 antisera do not detect any other B′ subunits (Fig. 1C). Finally, immunoblots ofNIH3T3 whole cell lysates did not detect the presence of any B″ subunits (data not shown).
SG2NA Binds to CaM in a Calcium-dependent MannerBecause striatin previously has been shown (23) to bind to CaM in a calcium-dependentmanner in a region (amino acids 149–166) that is nearly identical with SG2NA, wehypothesized that SG2NA would also bind to CaM. Both striatin (data not shown) andSG2NA (Fig. 4A) were precipitated with CaM-Sepharose from NIH3T3 cells stimulatedwith the calcium ionophore ionomycin but not from untreated control cells, as expected.This is the first evidence that SG2NA binds to CaM in a calcium-dependent manner similarto striatin. However, immunoblots of striatin and SG2NA immunoprecipitations probed withanti-CaM antibodies failed to detect CaM in both treated and untreated cells (data notshown). One hypothesis for the failure to detect CaM in immunoblots of striatin-PP2A andSG2NA-PP2A complexes is that the immunoprecipitating antibodies to striatin and SG2NAmay sterically interfere with CaM binding.
Striatin and SG2NA Complexes Contain Phosphatase Activity That Is Sensitive to OkadaicAcid and Does Not Require Calcium
To test whether immunoprecipitated striatin/PP2A and SG2NA/PP2A complexes containedenzymatically active PP2A, phosphatase assays were performed using 32P-labeledphosphorylase a as a substrate, and sensitivity to the PP2A inhibitor okadaic acid wasmeasured. Immunoprecipitations using antisera to striatin, SG2NA, and C subunit (4e1) allcontained significant phosphatase activity (Fig. 3B). The measured activity was largelyinhibited by 2 nM okadaic acid, as expected for PP2A. These assays were performed in thepresence of 1 mM EGTA, indicating that calcium is not required for the activity of striatin-PP2A and SG2NA-PP2A complexes. The addition of 2 mM calcium to these phosphatasereactions had a variable but slightly inhibitory effect on PP2A activity (data not shown).
Striatin and SG2NA Activate A/C Heterodimers toward cdc2-phoshorylated Histone H1Different B-type subunits have been shown to differentially activate the PP2A A/Cheterodimer toward different substrates. For example, B subunit is the only known B-typesubunit reported to activate A/C heterodimers toward cdc2-phosphorylated histone H1substrate (50–100-fold) (25–27). To determine whether the presence of striatin and SG2NAin PP2A complexes modulates the activity of A/C heterodimers, radiolabeled cdc2-phosphorylated histone H1 substrate was used to compare the activity of striatin and SG2NAcomplexes with other forms of PP2A. Immunoprecipitations were prepared from NIH3T3cells that stably express HA-tagged C subunit (36wt cells) using two different mAbs to thecarboxyl terminus of C subunit (1d6 and 4e1) and one mAb to the amino-terminal epitopetag (12CA5). Approximately 10–30% of 12CA5 complexes prepared from 36wt cellscontain B subunit, whereas approximately 5% of 1d6 complexes contain striatin and SG2NA(data not shown); 4e1 complexes contain C subunit alone and A/C heterodimers (18).Striatin and SG2NA complexes immunoprecipitated with anti-striatin and anti-SG2NAantisera, respectively, were highly activated toward histone H1 substrate compared withPP2A in 4e1 and 1d6 immune complexes (Fig. 3C). PP2A immunoprecipitated by 12CA5had similar (within 2-fold) activity to striatin and SG2NA complexes. Based on our estimate
Moreno et al. Page 6
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

of the percentage of 12CA5 complexes containing B subunit, we would estimate that striatinand SG2NA activate A/C heterodimers less than B subunit toward histone H1. Similarexperiments using NIH3T3 cells stably expressing HA-tagged B subunit2 confirmed thishypothesis (Fig. 3D), with striatin and SG2NA complexes approximately 35 and 29% asactive, respectively, as B subunit complexes against cdc2 phosphorylation sites. The highlevel of observed histone H1 phosphatase activity suggests that striatin and SG2NA are notbound to the A/C heterodimers via the catalytic site.
Chemiluminescence quantitation of immunoblots containing both lysates andimmunoprecipitations indicated that approximately 0.1–0.5% of the total C subunit presentin lysates was immunoprecipitated with the striatin and SG2NA antisera (data not shown).This finding contrasts with the observation that 5% of 1d6 immunoprecipitates containstriatin and SG2NA. Potential explanations for this difference are: 1) not all of the striatinand SG2NA present in lysates was immunoprecipitated; 2) the striatin and SG2NA antiseramay partially destabilize complex formation with the A/C heterodimer; or 3) that 1d6 maybind preferentially to striatin/PP2A and SG2NA/PP2A complexes.
Multiple Additional Proteins Are Present in Striatin and SG2NA ComplexesThe observation that striatin and SG2NA bind to both CaM and PP2A and contain WD40repeats suggested that these proteins might function as molecular scaffolds for PP2Asignaling complexes. To test whether striatin and SG2NA interact with additional proteins,NIH3T3 cells were metabolically labeled with [35S]methionine, and whole cell lysates wereimmunoprecipitated with both the anti-striatin and anti-SG2NA antiseras.Immunoprecipitated complexes were subjected to 2D gel electrophoresis and A subunit, Csubunit, striatin, and SG2NA were detected in striatin and SG2NA immunoprecipitations,but not in pre-immune controls (Fig. 4B and data not shown). Immunoblots of SG2NAimmunoprecipitates with affinity-purified SG2NA antisera revealed multiple additionalimmune-specific bands, suggesting that other members of the SG2NA family may exist(Fig. 4B and data not shown). Consistent with a role for striatin and SG2NA as molecularscaffolds, several unidentified proteins were coimmunoprecipitated with striatin and SG2NA(Fig. 4B).
The Carboxyl Terminus of PP2A C Subunit Present in Striatin-PP2A and SG2NA-PP2AComplexes Is Highly Methylated
We have previously suggested that the methylation state of the C subunit might regulate theassociation of A/C heterodimers with B-type subunits (22). To determine the methylationstate of the C subunit in striatin-PP2A and SG2NA-PP2A complexes, a portion of the Csubunit in striatin and SG2NA immunoprecipitations was subjected to demethylation bybase treatment. Both untreated and demethylated samples were analyzed by immunoblotwith a methylation-sensitive mAb (Fig. 5A) that recognizes only the demethylated Csubunit.2 The level of C subunit detected by the methylation-sensitive antibody wassubstantially enhanced in base-treated samples relative to untreated controls, andchemiluminescence quantitation determined that more than 90% of the C subunits associatedwith striatin and SG2NA are methylated. This result indicates that C subunit methylationdoes not prevent striatin and SG2NA association with the A/C heterodimer, but it does notindicate whether methylation is needed for formation of striatin-PP2A and SG2NA-PP2Acomplexes.
Deletion of the Carboxyl Terminus of PP2A C Subunit Does Not Prevent Striatin-PP2A andSG2NA-PP2A Complex Formation
Although the C subunit carboxyl terminus is essential for formation of PP2A heterotrimerscontaining the cellular B subunit, it is not required for formation of heterotrimers containing
Moreno et al. Page 7
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

the viral B-type subunit, polyoma virus middle tumor antigen (MT) (22). To investigate theimportance of the C subunit carboxyl terminus for the formation of A/C/striatin and A/C/SG2NA heterotrimers, striatin and SG2NA immunoprecipitations were performed from celllines expressing HA-tagged C subunit mutants (22). The final nine amino acids that containsites of both phosphorylation (Tyr-307) (28) and methylation (Leu-309) (29) are deleted inone of these mutants (301Stop). The 301Stop mutant was co-immunoprecipitated withstriatin and SG2NA at least as efficiently as the wt C subunit (Fig. 5B). These data indicatethat striatin and SG2NA interact with the core A/C heterodimer in a fundamentally differentmanner than the B subunit, behaving more like polyoma virus MT than B subunit in theirbinding to the A/C heterodimer. Thus, the association of striatin and SG2NA with the A/Cheterodimer is probably not directly affected by the covalent modification of the C subunitcarboxyl terminus. However, the association of striatin and SG2NA with the A/Cheterodimer could be indirectly affected by modifications that influence competition for theA/C heterodimer by altering the affinity of other B-type subunits. Consistent with thispossibility, the C subunit mutant, T304A, which has been shown to have an increasedaffinity for B subunits (22), was reduced 5-fold in its ability to form complexes with striatinand SG2NA compared with the wild-type HA-tagged C subunit (36wt, Fig. 5C).
Taken together, the following observations all strongly suggest that striatin and SG2NArepresent a novel family of B-type subunits: 1) striatin and SG2NA complex with the PP2AA/C heterodimer; 2) these complexes contain okadaic acid-sensitive, PP2A-like phosphataseactivity; 3) striatin and SG2NA share a conserved epitope with B′ subunits; 4) no known B-type subunits can be observed in these complexes; 5) unlike B subunits, the association ofstriatin and SG2NA with the A/C heterodimer is independent of the C subunit carboxylterminus, yet they can activate the A/C heterodimer toward cdc-2 phosphorylated histoneH1; and 6) their relative binding to wt and T304A C subunits is opposite that of B subunit(implying that B subunit may even compete with them for binding to the T304A mutantform of the A/C heterodimer). However, we cannot exclude the formal possibility that someother undiscovered B-type subunit that is not recognized by any of the B, B′, or B″ antiseracould be present in these complexes. Our conclusion that striatin and SG2NA may bemembers of a new class of B-type subunits would be further strengthened by additionalevidence of competition with other B-type subunits or demonstration of direct interactionbetween bacterially expressed recombinant striatin and SG2NA and recombinant PP2A Asubunit or A/C heterodimer. Should further evidence conclusively demonstrate that striatinand SG2NA are members of a novel family of B-type subunits, we propose that they bedesignated the B‴ (or B93/110) family.
The finding that striatin and SG2NA form stable complexes with PP2A and might representa novel (B‴) family of PP2A subunits is the first description of a function for these highlyrelated WD40 repeat proteins. The fact that striatin is highly abundant in post-synapticmembranes, whereas SG2NA appears to be targeted to the nucleus, provides yet anothermechanism for the localization of PP2A to different cellular microenvironments. Theobservation that SG2NA, as well as striatin, binds to CaM in a calcium-dependent mannerindicates that these proteins probably link PP2A to calcium-dependent signaling pathwaysand cellular events. Although PP2A complexes with CaMKIV have been detected by cross-linking (11), it is not yet known whether this interaction is direct or requires a molecularscaffold. Although we were not able to detect CaMKII or CaMKIV in immunoblots ofstriatin or SG2NA immunoprecipitations, we have detected kinase activity in theseimmunoprecipitates (data not shown). Furthermore, the large number of stably associatedproteins observed in SG2NA immunoprecipitations suggests that striatin and SG2NAfunction as molecular scaffolds for the interactions of PP2A with large signal transductioncomplexes. The identification of additional cellular components of these complexes willprovide new insights into the cellular function of striatin, SG2NA, and PP2A.
Moreno et al. Page 8
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

AcknowledgmentsThe authors thank Brian Lang, Michael Sanford, and Russell Thomas for technical assistance; Renee Robinson,Dan Kirby, and Kerry Pierce of the Harvard Microchemistry Facility for their expertise in high pressure liquidchromatography and mass spectrometry; and Dr. Brian Hemmings for the gift of anti-B″ (PR72/130) polyclonalantisera. Under agreements between Upstate Biotechnology Inc. and Emory University and Calbiochem and EmoryUniversity, David Pallas is entitled to a share of sales royalty received by the University from these companies. Inaddition, this same author serves as a consultant to Upstate Biotechnology Inc.
REFERENCES1. Mumby MC, Walter G. Physiol. Rev. 1993; 73:673–699. [PubMed: 8415923]
2. Hopkin K. J. NIH Res. 1995; 7:27–30.
3. Goldberg Y. Biochem. Pharmacol. 1999; 57:321–328. [PubMed: 9933020]
4. Cohen P. Annu. Rev. Biochem. 1989; 58:453–508. [PubMed: 2549856]
5. Tanabe O, Nagase T, Murakami T, Nozaki H, Usui H, Nishito Y, Hayashi H, Kagamiyama H,Takeda M. FEBS Lett. 1996; 379:107–111. [PubMed: 8566219]
6. Zolnierowicz S, Csortos C, Bondor J, Verin A, Mumby MC, DePaoli-Roach AA. Biochemistry.1994; 33:11858–11867. [PubMed: 7918404]
7. McCright B, Virshup DM. J. Biol. Chem. 1995; 270:26123–26128. [PubMed: 7592815]
8. Tehrani MA, Mumby MC, Kamibayashi C. J. Biol. Chem. 1996; 271:5164–5170. [PubMed:8617797]
9. McCright B, Rivers AM, Audlin S, Virshup DM. J. Biol. Chem. 1996; 271:22081–22089. [PubMed:8703017]
10. Seeling JM, Miller JR, Gil R, Moon RT, White R, Virshup DM. Science. 1999; 283:2089–2091.[PubMed: 10092233]
11. Westphal RS, Anderson KA, Means AR, Wadzinski BE. Science. 1998; 280:1258–1261.[PubMed: 9596578]
12. Tian L, Knaus HG, Shipston MJ. J. Biol. Chem. 1998; 273:13531–13536. [PubMed: 9593688]
13. Zhou XB, Ruth P, Schlossmann J, Hofmann F, Korth M. J. Biol. Chem. 1996; 271:19760–19767.[PubMed: 8702682]
14. Groschner K, Schuhmann K, Mieskes G, Baumgartner W, Romanin C. Biochem. J. 1996;318:513–517. [PubMed: 8809040]
15. Castets F, Bartoli M, Barnier JV, Baillat G, Salin P, Moqrich A, Bourgeois JP, Denizot F, RougonG, Calothy G, Monneron A. J. Cell Biol. 1996; 134:1051–1062. [PubMed: 8769426]
16. Muro Y, Chan EK, Landberg G, Tan EM. Biochem. Biophys. Res. Commun. 1995; 207:1029–1037. [PubMed: 7864889]
17. Pallas DC, Shahrik LK, Martin BL, Jaspers S, Miller TB, Brautigan DL, Roberts TM. Cell. 1990;60:167–176. [PubMed: 2153055]
18. Ogris E, Du X, Nelson KC, Mak EK, Yu XX, Lane WS, Pallas DC. J. Biol. Chem. 1999;274:14382–14391. [PubMed: 10318862]
19. Chittum HS, Lane WS, Carlson BA, Roller PP, Lung FD, Lee BJ, Hatfield DL. Biochemistry.1998; 37:10866–10870. [PubMed: 9692979]
20. Eng JK, McCormick AL, Yates JR III. J. Am. Soc. Mass Spectrom. 1994; 5:976–989.
21. Turowski P, Fernandez A, Favre B, Lamb NJ, Hemmings BA. J. Cell Biol. 1995; 129:397–410.[PubMed: 7721943]
22. Ogris E, Gibson DM, Pallas DC. Oncogene. 1997; 15:911–917. [PubMed: 9285686]
23. Bartoli M, Monneron A, Ladant D. J. Biol. Chem. 1998; 273:22248–22253. [PubMed: 9712839]
24. Wilson R, Ainscough R, Anderson K, Baynes C, Berks M, Bonfield J, Burton J, Connell M,Copsey T, Cooper J, Coulson A, Craxton M, Dear S, Du Z, Durbin R. Nature. 1994; 368:32–38.[PubMed: 7906398]
25. Ferrigno P, Langan TA, Cohen P. Mol. Biol. Cell. 1993; 4:669–677. [PubMed: 8400454]
26. Sola MM, Langan T, Cohen P. Biochim. Biophys. Acta. 1991; 1094:211–216. [PubMed: 1654119]
Moreno et al. Page 9
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

27. Agostinis P, Derua R, Sarno S, Goris J, Merlevede W. Eur. J. Biochem. 1992; 205:241–248.[PubMed: 1313364]
28. Chen J, Martin BL, Brautigan DL. Science. 1992; 257:1261–1264. [PubMed: 1325671]
29. Lee J, Stock J. J. Biol. Chem. 1993; 268:19192–19195. [PubMed: 8396127]
Moreno et al. Page 10
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Fig. 1. Identification of 110- and 93-kDa PP2A-associated proteins as striatin and SG2NAA, immunoblot of NIH3T3 lysate using affinity-purified AR-1 antisera (AR-1) and affinity-purified AR-1 antisera pre-blocked with 100 µg/ml of AR-1 peptide (AR-1 + peptide).Proteins that are recognized by the AR-1 antisera and their apparent molecular mass areindicated. Although only the top portion of the gel is shown, no lower molecular weightbands were visible. Although the 74-kDa protein is barely visible here, it has been observedrepeatedly. The intensity of the 131- and 74-kDa bands can vary from cell line to cell lineand blot to blot. B, autoradiograph of 2D gel analysis of anti-PP2A C subunit (1d6 mAb;PP2A IP panel) and control (7-34-1 mAb; Control IP panel) immunoprecipitates (IP)prepared from 35S-labeled NIH3T3 cells. 93- and 110-kDa PP2A-associated proteins are
Moreno et al. Page 11
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

indicated. C, immunoblot of SDS-PAGE analysis of 7-34-1 (Control IP) and 1d6 (PP2A IP)immunoprecipitates probed with AR-1 antisera (AR-1), anti-striatin antisera (Striatin), andanti-SG2NA antisera (SG2NA). Ab, antibody. D, protein sequence alignment of striatin andSG2NA. Tryptic peptide sequences determined by ion trap mass spectrometry of the 93- and110-kDa proteins are underlined. Peptides used to generate polyclonal antisera to striatin andSG2NA are double underlined. Probable recognition sites for the AR-1 antisera are shown initalics.
Moreno et al. Page 12
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text
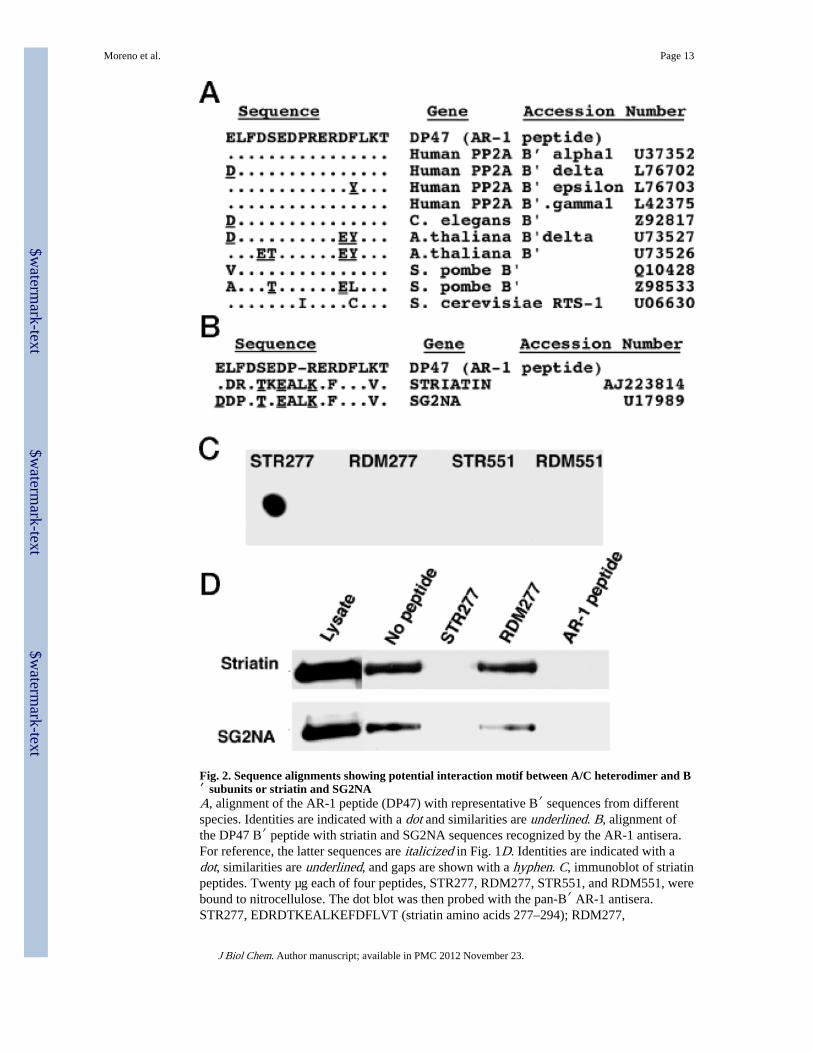
Fig. 2. Sequence alignments showing potential interaction motif between A/C heterodimer and B′ subunits or striatin and SG2NAA, alignment of the AR-1 peptide (DP47) with representative B′ sequences from differentspecies. Identities are indicated with a dot and similarities are underlined. B, alignment ofthe DP47 B′ peptide with striatin and SG2NA sequences recognized by the AR-1 antisera.For reference, the latter sequences are italicized in Fig. 1D. Identities are indicated with adot, similarities are underlined, and gaps are shown with a hyphen. C, immunoblot of striatinpeptides. Twenty µg each of four peptides, STR277, RDM277, STR551, and RDM551, werebound to nitrocellulose. The dot blot was then probed with the pan-B′ AR-1 antisera.STR277, EDRDTKEALKEFDFLVT (striatin amino acids 277–294); RDM277,
Moreno et al. Page 13
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

TLDELETREVFFKAKDD (randomized STR277 sequence); STR551,DPYDSYDPSVLRGPLL (striatin amino acids 551–566); RDM551,LYPRYVPPLDGLSSDD (randomized STR551 sequence). D, AR-1 immunoprecipitatesimmunoblotted with striatin and SG2NA antisera. AR-1 immunoprecipitations (lanes 2–5)were carried out in 20% Triton either with no peptide added or preincubated with 100 µg/mlof the STR277, RDM277, or AR-1 peptides, respectively. Each set of lanes is from the samegel, but the lanes were not all originally adjacent.
Moreno et al. Page 14
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Fig. 3. PP2A subunits and phosphatase activity in striatin and SG2NA immunoprecipitatesA, NIH3T3 whole cell lysate (Lysate) and striatin and SG2NA immunoprecipitates andpreimmune controls were simultaneously probed with a mixture of monoclonal antibodies toC subunit (Transduction Laboratories) and A subunit (4 g7 mAb; Ref. 17). These lanes werefrom the same gel but were not all originally adjacent and have been cut vertically. Althoughthe top and bottom portions of the blot are not shown, no other bands were visible. WhetherPP2A C subunit migrates as a doublet or as a single band varies from gel to gel. Thisbehavior on SDS-PAGE has been observed previously (22) and does not appear to be theresult of degradation. B, normalized phosphorylase a phosphatase activity of immunecomplexes prepared with 4e1 anti-PP2A C subunit (PP2A), striatin, and SG2NA antisera.
Moreno et al. Page 15
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Immunoprecipitates were prepared from approximately 3 mg of total protein and used tomeasure phosphatase activity. Phosphorylase a phosphatase assays were carried out usingthe Protein Phosphatase Assay System (Life Technologies, Inc.) according to themanufacturer’s instructions with the addition of 1 mM EGTA and the indicatedconcentrations of okadaic acid. Cpm released was first corrected by subtracting backgroundactivity obtained with pre-immune immunoprecipitations. Preimmune Cpm averaged 7 ± 5%of PP2A activity, 34 ± 13% of SG2NA activity, and 45 ± 15% of striatin activity. Theaverage immune-specific activity measured in the absence of okadaic acid was arbitrarily setto 100% for each immunoprecipitate. The effect of okadaic acid was measured bynormalizing Cpm released in the presence of okadaic acid against Cpm released in theabsence of okadaic acid. The averages and standard deviations of at least three independentexperiments are shown. C, normalized histone H1 phosphatase activity ofimmunoprecipitations prepared from NIH3T3 cells stably expressing HA-tagged PP2A Csubunit (HA-C subunit). Immunoprecipitations were performed using 12CA5 anti-HA-tagmAb; 1d6 and 4e1 anti-C subunit mAbs; and polyclonal antisera against striatin andSG2NA. The averages and standard deviations of at least three independent experiments areshown. Two-thirds of each immunoprecipitate prepared from approximately 3 mg of totalprotein was used to measure phosphatase activity as described previously (22), and one-thirdwas analyzed by SDS-PAGE and immunoblotted with commercial anti-C subunit mAbs(Transduction Laboratories). The amount of C subunit present in immunoblots of eachcomplex was quantitated using a chemiluminescence imager (Bio-Rad). Cpm released wasfirst corrected by subtracting background activity obtained with pre-immuneimmunoprecipitations. Specific activity (immune-specific Cpm released/chemiluminescencecounts) was then calculated, and the level of phosphatase specific activity was finallynormalized relative to the amount of specific activity present in 12CA5immunoprecipitations of HA-tagged PP2A C subunit to obtain the percent of HA-C subunitspecific activity. D, normalized histone H1 phosphatase activity of immunoprecipitationsprepared from NIH3T3 cells stably expressing HA-tagged PP2A B subunit (HA-B subunit).Immunoprecipitations were perfomed using 12CA5 anti-HA-tag mAb and striatin andSG2NA polyclonal antisera as described in C. The averages and standard deviations of atleast three independent experiments are shown. Calculations were performed as described inC, except specific activity was computed as a percent of HA-tagged PP2A B subunit specificactivity.
Moreno et al. Page 16
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Fig. 4. SG2NA/PP2A complexes contain multiple additional cellular proteinsA, SGN2A binds to CaM in a calcium-dependent manner. Whole cell lysates and CaM-Sepharose precipitates were prepared from NIH3T3 cells treated for 30 min with 1 µM
ionomycin (+) and untreated control cells (−). Lysates and precipitates were analyzed bySDS-PAGE and immunoblotted with affinity-purified SG2NA antisera. B, 2D gel analysesof preimmune and SG2NA immunoprecipitates (IP) prepared from 35S-labeled NIH3T3cells are shown. SG2NA and the A and C subunits of PP2A are indicated. Bracketed spotsmarked with asterisks are recognized by affinity-purified SG2NA antisera and may representother members of the SG2NA family or alternatively spliced forms of SG2NA. Additional,unidentified spots associated with SG2NA are indicated with arrowheads and brackets
Moreno et al. Page 17
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

without asterisks. The corresponding positions for all immune-specific spots are alsoindicated on the pre-immune exposure.
Moreno et al. Page 18
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

Fig. 5. PP2A C subunits associated with striatin and SG2NA are highly methylated, but Csubunit methylation is probably not required for formation of these complexesA, methylation level of striatin- and SG2NA-associated PP2A C subunits.Immunoprecipitates prepared with preimmune and immune striatin antisera and preimmuneand immune SG2NA antisera were each divided into two equal portions, and one of eachpair (+) was demethylated by treatment with 0.2 N NaOH on ice for 5 min (21). Base-treatedsamples were neutralized, and preneutralized base solution was added to untreated control(−) samples. All samples were then analyzed by SDS-PAGE and immunoblotting. Themembrane was probed first with 4b7 (Methylation Sensitive Ab panel), an anti-C subunitantibody that recognizes only unmethylated C subunits. Subsequently, the same membrane
Moreno et al. Page 19
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text

was probed with Transduction Laboratories anti-PP2A C subunit antibody (MethylationInsensitive Ab panel), which is insensitive to the methylation state of PP2A and thereforereveals the total C subunit in each lane. Ab, antibody. B, deletion of nine carboxyl-terminalresidues from the PP2A C subunit does not affect binding to striatin and SG2NA. Celllysates and striatin and SG2NA immunoprecipitates were prepared from polyomavirus MT-transformed NIH3T3 cell lines expressing empty vector (GRE only), HA-tagged wt Csubunit (36wt), or HA-tagged C subunit truncation mutant lacking nine carboxyl-terminalamino acids (301STOP). Samples were analyzed by SDS-PAGE and immunoblotted withanti-HA-tag antibody (16b12, BAbCO, Richmond, CA). Each set of lanes is from the samegel, but the lanes were not all originally adjacent. PP2A C subunit has been previouslyobserved to migrate sometimes as a doublet and sometimes as a singlet on SDS-PAGE (22).C, a single substitution of alanine for threonine 304 in the PP2A C subunit greatly reducescomplex formation with striatin and SG2NA. Cell lysates and striatin and SG2NAimmunoprecipitates (IP) were prepared from polyomavirus MT-transformed NIH3T3 celllines expressing empty vector (GRE only), HA-tagged wt C subunit (36wt), or HA-tagged Csubunit mutant (T304A). Samples were analyzed by SDS-PAGE and immunoblotted withanti-HA-tag antibody (16b12, BAbCO). Quantitation with a chemiluminescence imagersystem (Bio-Rad) indicated that T304A bound to striatin and SG2NA approximately 5-foldless efficiently than did 36wt. Each set of lanes is from the same gel, but the lanes were notall originally adjacent.
Moreno et al. Page 20
J Biol Chem. Author manuscript; available in PMC 2012 November 23.
$waterm
ark-text$w
atermark-text
$waterm
ark-text