unusual patterns of somatic embryogenesis in the domesticated carrot: developmental effects of...
TRANSCRIPT

Cell Differentiation, 21 (1987) 53-62 53 Elsevier Scientific Publishers Ireland, Ltd.
CDF 00429
Unusual patterns of somatic embryogenesis in the domesticated carrot: developmental effects
of exogenous auxins and auxin transport inhibitors
F. Mark Schiavone and Todd J. Cooke
Department of Botany, University of Maryland, College Park, MD 20742, U.S.A.
(Accepted 4 December 1986)
The effects of various exogenous auxins and polar auxin transport inhibitors on somatic embryogenesis in carrot cultures were investigated. Indole-3-acetic acid and 2,4-dichlorophenoxyacetic acid do not disrupt the sequence or the polarity of individual stages in embryo development, but tend to cause developing embryos to revert to undifferentiated callus, with increasing frequency in later embryo stages. The transport inhibitors, N-(1-naphthyl)phthalamic acid and 2,3,5-triiodobenzoic acid, block morphological transitions to the subsequent stage; for example, they cause the formation of enlarged globular and oblong embryos. Heart embryos in these treatments usually develop additional lateral growth axes. These results shed light on the role of auxin and its polar transport in somatic embryogenesis.
Somatic embryogenesis; Auxin; Polar transport; Carrot
Introduction
Somatic embryos are a more useful experimen- tal system than zygotic embryos for studying the control mechanisms of plant embryogenesis. For instance, it is possible to induce large numbers of somewhat synchronous somatic embryos in carrot cultures by removing exogenous supplied auxin from the cultural medium (for review, see Ragha- van, 1985). However, the mechanism of auxin action has remained enigmatic.
Correspondence address: F.M. Schiavone, Department of Botany, University of Maryland, College Park, MD 20742, U.S.A. Abbreviations: 2,4-D, 2,4-dichlorophenoxyacetic acid; 2,4,6-T, 2,4,6-trichlorophenoxyacetic acid; IAA, indole-3-acetic acid; NPA, N-(1-naphthyl)phthalamic acid; TIBA, 2,3,5-triiodo- benzoic acid.
We attempt in this paper to outline the roles that the endogenous auxin indole-3-acetic acid, and its analogues, play in the physiological regu- lation of somatic embryogenesis in carrot cultures. Since auxin added to the culture medium main- tains carrot callus in an undifferentiated state, and since the removal of this auxin initiates the em- bryogenic process, our first concern was the elucidation of auxin effects early in development. Secondly, we wanted to explore whether later stages of embryo development, which are char- acterized by the initiation and extension of the hypocotyl, cotyledons, and root, are sensitive to elevated auxin levels, or to auxin gradients within the embryo. Our experiments indicate that polar auxin transport is intimately involved in the ini- tiation and maintenance of polarized growth in developing embryos.
0045-6039/87/$03.50 © 1987 Elsevier Scientific Publishers Ireland, Ltd.

54
Materials and Methods
All procedures outlined below have been previ- ously described (Schiavone and Cooke, 1985), with the exception of experiments that involve sieved embryos raised in 3 ml volumes. Callus cultures were derived from the hypocotyls of carrot seed- lings (Daucus carota L. cv Danvers) and were maintained as liquid suspensions in Murashige and Skoog's (1962) medium supplemented with 5 #M 2,4-dichlorophenoxyacet ic acid (2,4-D) (MSA-medium). Somatic embryos were initiated by transferring sieved cell clusters between 43 and 117 #m in diameter to Murashige and Skoog's medium devoid of 2,4-D (MSE medium) at a concentration of 650 cell clusters/ml. In some experiments, indole-3-acetic acid (IAA), 2,4,6-tri- chlorophenoxyacetic acid (2,4,6-T) or 2,4-D was added to three replicate flasks at various times after the removal of MSA medium. Small aliquots were periodically drawn from each flask to de- termine the number /ml of cell clusters, different stages of developing embryos, and two types of organized structures of unusual morphology termed abnormal and abortive embryos. Abnormal embryos assumed a variety of atypical shapes such as additional growth axes or multiple cotyledons, but maintained a smooth protoderm or immature epidermis. In contrast, abortive embryos resem- bled recognized stages, but their irregular surfaces suggested that they were reverting back to callus. In addition, a sample of developing embryos was randomly selected from each aliquot for geometric analysis (see Schiavone and Cooke, 1985).
To study the effects of auxin analogues and transport inhibitors on developing embryos, the embryos were grown in MSE medium until a substantial number of heart embryos appeared in 10 to 11 days. The cultures were then passed through 380 ~tm, 117 #m, and 53 /tm sieves in series. The ll7-/xm sieve isolated mostly heart embryos while large cell dusters, globular em- bryos, and oblong embryos were retained on the 53/xm sieve. The embryos were further purified by swirling each fraction in a petri dish, which caused the heavier embryos to sediment in the dish center and lighter cell clusters to move to the dish pe- riphery (Giuliano et al., 1983). These purified frac-
tions were adjusted to a final concentration be- tween 80 and 120 globular or heart embryos /ml and were placed in 3 ml of MSE medium in six-well plastic dishes on a rotary shaker set at 90 rpm. IAA, 2,4-D, N-(1-naphthyl)phthalamic acid (NPA) or 2,3,5-triiodobenzoic acid (TIBA), were added to wells in three replicate treatments. Small aliquots were collected after 6 days for counting embryo stages and for geometric analysis.
Results
Since this paper attempts to characterize how auxin regulates the different stages of embryo development, it is necessary to first define the stages encountered in somatic embryogenesis. Careful geometric analysis (Schiavone and Cooke, 1985) indicates that developing carrot embryos pass through four distinct morphological stages, i.e., the globular, oblong, heart, and torpedo stages. The oblong stage is consistently seen as an inter- mediate step between the globular and heart stages, in which the future hypocotyl axis starts to elon- gate before the initiation of the cotyledons.
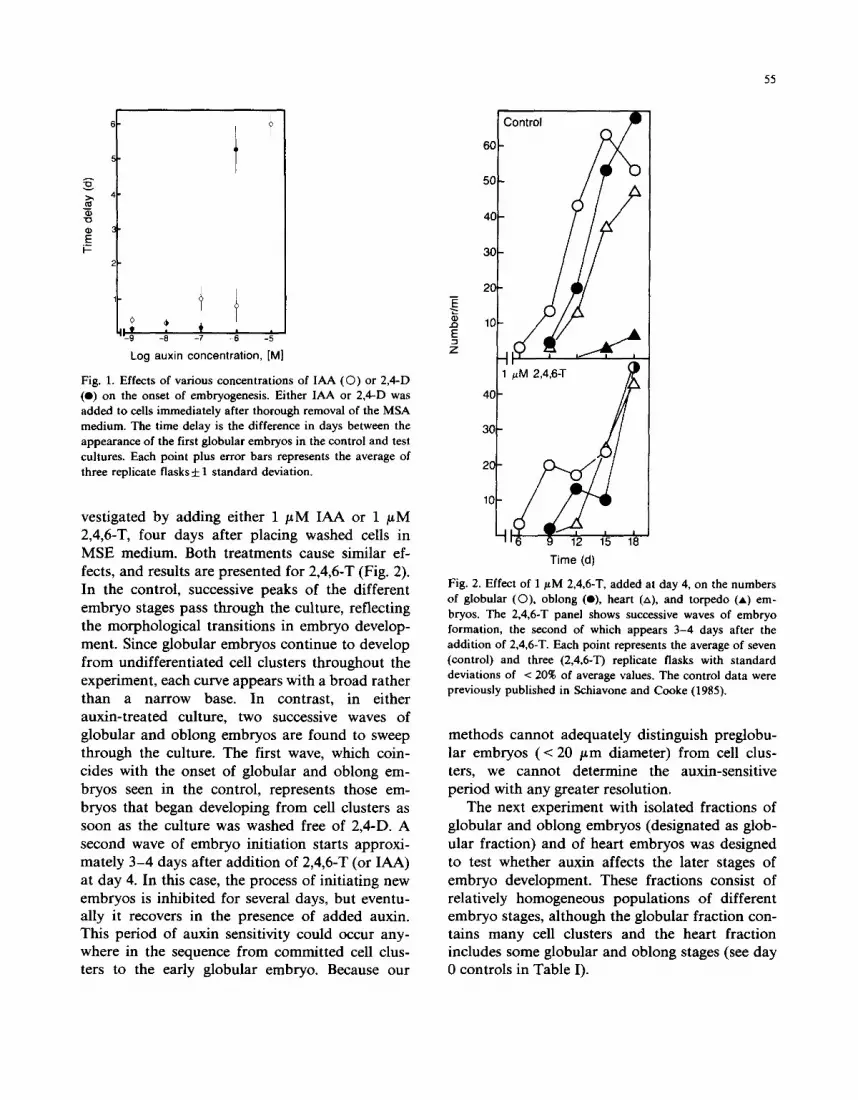
The first experiment was designed to determine the threshold concentration of auxin required to block embryogenesis. Thus, cell cultures were washed free of MSA medium with 2,4-D, and then embryo development was measured in MSE medium supplemented with either 2,4-D or IAA. The inhibition of embryogenesis could then be measured as a delay in the onset of globular embryos relative to control cultures (unsupple- mented MSE medium).
It is clear from Fig. 1 that both auxins effect sharp threshold responses. IAA causes an inhibi- tion of a day or less at low concentrations (1 nM to 1/~M), with a sharp rise to a 6-day delay at 10 /~M. Nanomolar 2,4-D concentrations cause de- lays of less than half a day, whereas a similar sharp rise in the delay occurs at a 10-fold lower concentration than IAA. No embryos are present 12 days after the addition of 10 / tM 2,4-D. More- over, another auxin, 2,4,6-T, has inhibitory effects very similar to those observed with 2,4-D (data not shown).
The nature of the time delay was further in-
° +
.!
Log auxin concentration, [M]
Fig. 1. Effects of various concentrations of IAA (0 ) or 2,4-D (e) on the onset of embryogenesis. Either IAA or 2,4-D was added to cells immediately after thorough removal of the MSA medium. The time delay is the difference in days between the appearance of the first globular embryos in the control and test cultures. Each point plus error bars represents the average of three replicate flasks d: 1 standard deviation.
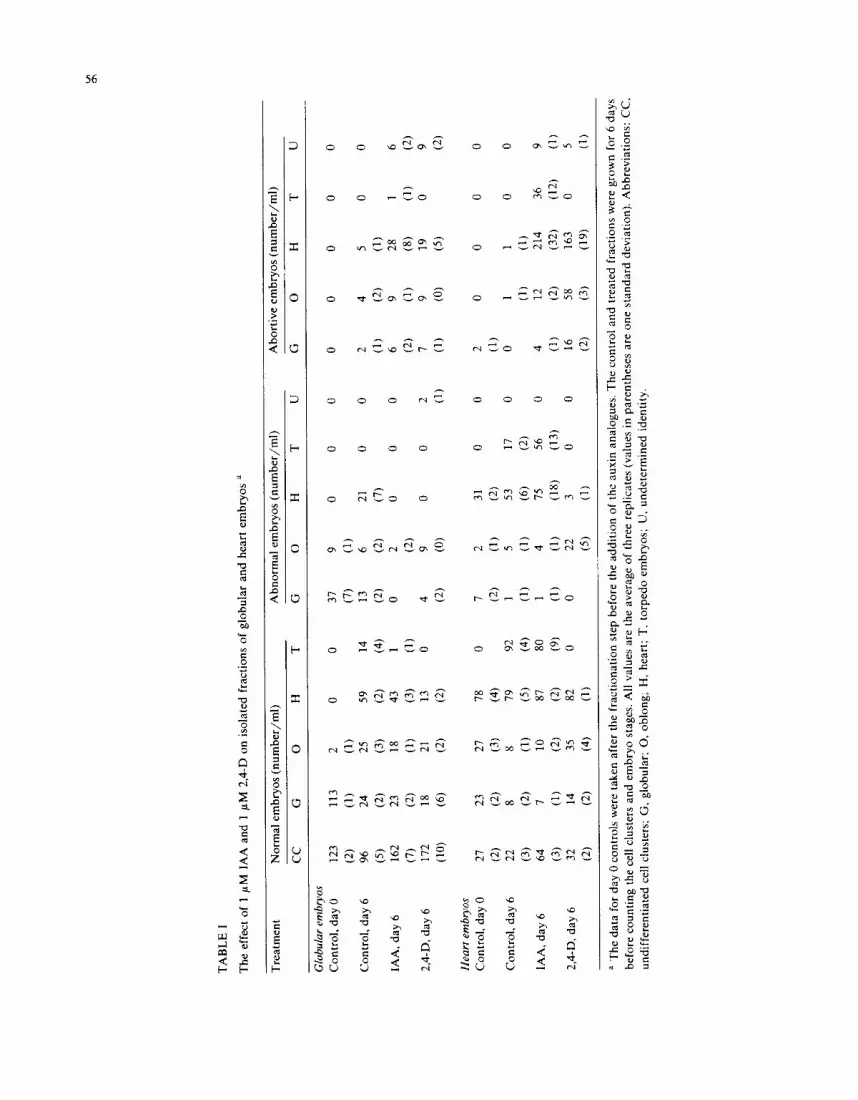
vestigated by adding either 1 ptM IAA or 1 #M 2,4,6-T, four days after placing washed cells in MSE medium. Both treatments cause similar ef- fects, and results are presented for 2,4,6-T (Fig. 2). In the control, successive peaks of the different embryo stages pass through the culture, reflecting the morphological transitions in embryo develop- ment. Since globular embryos continue to develop from undifferentiated cell clusters throughout the experiment, each curve appears with a broad rather than a narrow base. In contrast, in either auxin-treated culture, two successive waves of globular and oblong embryos are found to sweep through the culture. The first wave, which coin- cides with the onset of globular and oblong em- bryos seen in the control, represents those em- bryos that began developing from cell clusters as soon as the culture was washed free of 2,4-D. A second wave of embryo initiation starts approxi- mately 3-4 days after addition of 2,4,6-T (or IAA) at day 4. In this case, the process of initiating new embryos is inhibited for several days, but eventu- ally it recovers in the presence of added auxin. This period of auxin sensitivity could occur any- where in the sequence from committed cell clus- ters to the early globular embryo. Because our
55
..Q
E :3 z
60
50
4C
30
20
10
ontrol
F i b 40 1 #M 2,4,6-T / / ~
S 3O
. o
10
!
=6 9 12 15 18 Time (d)
Fig. 2. Effect of 1 #M 2,4,6-T, added at day 4, on the numbers of globular (O), oblong (O), heart (zx), and torpedo (=) em- bryos. The 2,4,6-T panel shows successive waves of embryo formation, the second of which appears 3-4 days after the addition of 2,4,6-T. Each point represents the average of seven (control) and three (2,4,6-T) replicate flasks with standard deviations of < 20% of average values. The control data were previously published in Schiavone and Cooke (1985).
methods cannot adequately distinguish preglobu- lar embryos (< 20 ttm diameter) from cell clus- ters, we cannot determine the auxin-sensitive period with any greater resolution.
The next experiment with isolated fractions of globular and oblong embryos (designated as glob- ular fraction) and of heart embryos was designed to test whether auxin affects the later stages of embryo development. These fractions consist of relatively homogeneous populations of different embryo stages, although the globular fraction con- tains many cell clusters and the heart fraction includes some globular and oblong stages (see day 0 controls in Table I).
56
O b
r ,
o
.2
o
<
0
m~mGo ~G
~ d
" 0 . 0 . = ,..-, . ~
. ~ <
o .=_ -~
~ N
N o ©
8 ~ " d
,.2 ° ~ .=

The effects of 1 /~M IAA and 1 /zM 2,4-D added to globular or heart populations are sum- marized in Table I. When IAA is added to globu- lar embryos, they progress to form heart embryos, but at numbers slightly lower than those of the control culture. Globular embryos treated with the same concentration of 2,4-D develop into fewer heart embryos. Simultaneously, the numbers of undifferentiated cell clusters and abortive heart embryos are much greater in both treatments than in the control.
Similar results are seen in the heart fraction (Table I). Here, 1 /~M IAA causes a marked increase in abortive embryos, and a slight increase in abnormal embryos, as compared with the con- trol cultures. One noteworthy difference is that many heart embryos treated with IAA continue to develop into torpedo embryos, with their numbers only slightly lower than the control, whereas 2,4- D-treated heart embryos are incapable of produc- ing torpedo embryos. Moreover, fewer abnormal heart embryos arise in 2,4-D-treated cultures, but these cultures also exhibit a large number of abor- tive heart embryos. It is not known whether most abortive or abnormal heart embryos result from various changes in initially normal heart embryos, or from younger embryos which start to undergo atypical development before they reach the heart stage.
Since the polar transport of auxin has been shown to occur in the mature embryos of several plant species (Greenwood and Goldsmith, 1970; Fry and Wangermann, 1976), it is conceivable that auxin transport might also help maintain the polarized growth of developing carrot embryos. This possibility was examined by using two differ- ent transport inhibitors, NPA and TIBA, which are thought to act via two independent mecha- nisms (Thomson et al., 1973). Initial experiments showed that neither NPA nor TIBA, in the range of 1 nM to 1/zM, delayed the timing or the rate of embryo initiation. These inhibitors at 1 /~M did cause a 40% decrease in the final fresh weight of cultures relative to controls, but this effect appears attributable to the slower growth of cell clusters. Higher inhibitor concentrations caused profound inhibition of cell growth and embryo formation.
Due to slight variations in the control cultures
57
in the two experiments, the results of NPA and TIBA added to the globular and heart fractions are presented separately in Tables II and III. Because the results of each treatment were so similar, the following text does not specifically mention either experiment. A typical progression for a population of globular embryos over 6 days is a decline in globular embryos with successive rises in oblong, heart, and torpedo embryos (Fig. 2, and day 6 controls of Tables II and III). When an auxin transport inhibitor is added to this popu- lation, very few oblong embryos continue to de- velop into heart embryos, while there is a marked accumulation of globular and oblong embryos. These apparent developmental blocks do not in- volve the formation of abortive embryos as seen in the IAA or 2,4-D experiments. Rather, when a polar auxin transport inhibitor blocks subsequent transitions in embryo formation, globular and ob- long embryos continue to increase in overall size but remain at the same developmental stage (see below).
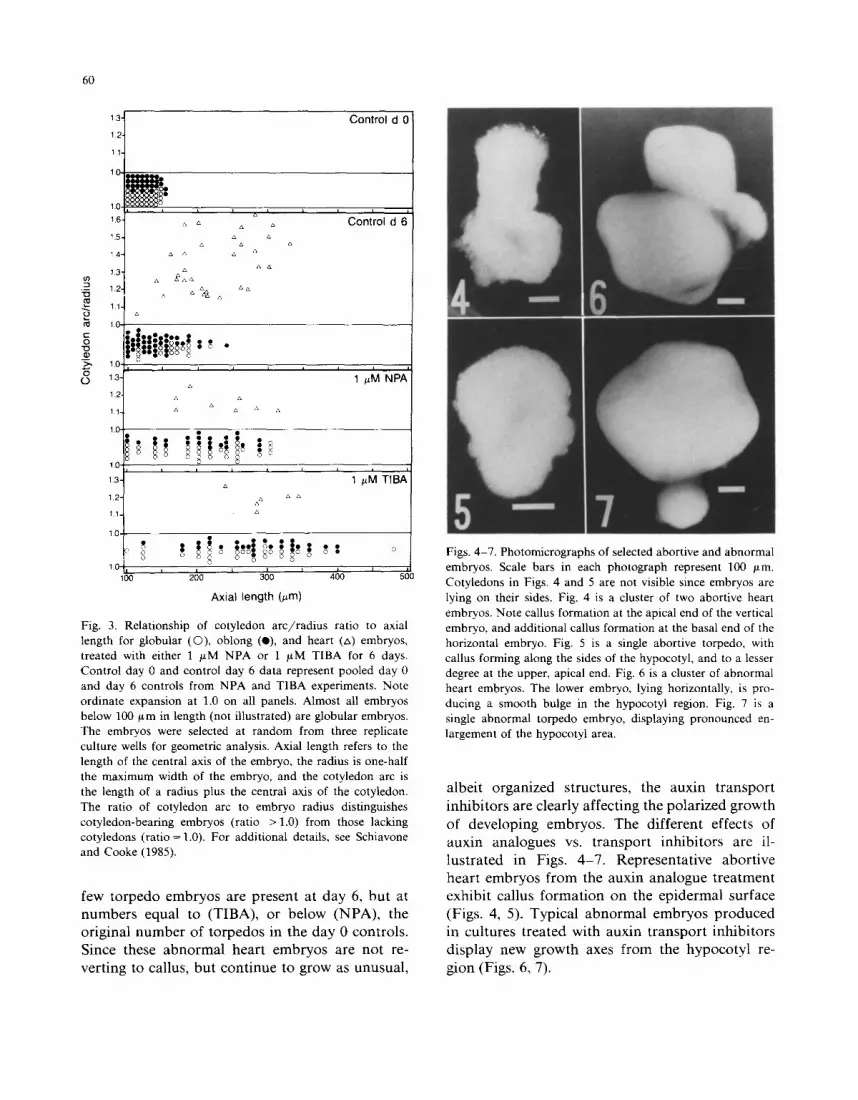
Fig. 3 applies a geometric method for measur- ing physical dimensions of somatic embryos (Schiavone and Cooke, 1985) to characterize the effects of NPA and TIBA on the globular fraction. The controls for NPA and TIBA are combined in Fig. 3, because no morphological differences in the embryos of each stage were observed between the two sets of control treatments. Whereas the typical axial lengths of late globular and oblong embryos are 80-140 and 100-200 /zm, respec- tively, the data demonstrate that the treatments cause globular and oblong embryos to reach axial lengths between 350 and 600 /~m. The few heart embryos appearing in the treatment may arise from a few oblong embryos which either are genuinely unsensitive to the transport inhibitors, or had already passed some physiologically sensi- tive point in their development prior to the ad- dition of NPA or TIBA. Nonetheless, the trans- port inhibitors cause globular embryos to continue spherical growth with a few eventually becoming oblong embryos, all of which remain in this stage.
When auxin transport inhibitors are added to heart populations, all embryos fail to reach the torpedo stage, but instead become abnormal em- bryos (Tables II and III). In both treatments, a

TA
BL
E I
I
The
eff
ect
of 1
~tM
TIB
A o
n is
olat
ed f
ract
ions
of
glob
ular
and
hea
rt e
mb
ryo
s "
Tre
atm
ent
No
rmal
em
bry
os
(nu
mb
er/m
l)
Ab
no
rmal
em
bry
os
(nu
mb
er/m
l)
CC
G
O
H
T
G
O
H
T
Ab
ort
ive
emb
ryo
s (n
um
ber
/ml)
U
G
O
H
T
U
Glo
bula
r em
bryo
s C
ontr
ol,
day
0 17
8 42
12
0
0 22
5
0 0
0 0
0 0
0 0
(11)
(2
) (1
) (3
) (2
) C
ontr
ol,
day
6 19
6 29
40
14
0 67
16
9
49
0 0
2 7
13
0 1
(6)
(2)
(2)
(7)
(8)
(2)
(2)
(12)
(1
) (2
) (3
) (1
) T
IBA
, da
y 6
223
75
58
26
0 13
19
0
0 0
7 10
0
0 0
(8)
(6)
(3)
(2)
(2)
(10)
(0
) (3
)
Hea
rt e
mbr
yos
Con
trol
, da
y 0
82
47
70
118
7 14
6
47
0 1
4 0
0 0
0 (6
) (2
) (3
) (8
) (1
) (3
) (2
) (2
) (1
) (2
) C
on
tro
l da
y 6
60
10
23
193
137
1 13
13
0 26
1
0 2
3 0
0 (3
) (1
) (1
) (7
) (6
) (1
) (3
) (1
1)
(2)
(1)
(2)
(2)
TIB
A,
day
6 93
20
47
10
0 7
20
109
300
23
14
0 0
10
0 0
(6)
(2)
(4)
(4)
(3)
(11)
(1
9)
(22)
(2
) (3
) (3
)
" M
any
of t
he g
lobu
lar
and
ob
lon
g e
mb
ryo
s in
the
tre
ated
cul
ture
s ar
e gr
eatl
y en
larg
ed r
elat
ive
to t
he t
ypic
al s
izes
of
cont
rol
embr
yos.
How
ever
, th
ese
emb
ryo
s w
ere
coun
ted
as b
eing
nor
mal
sin
ce t
hey
mai
ntai
n th
e sa
me
shap
e as
the
con
trol
em
bryo
s. F
or
addi
tion
al n
otes
, se
e th
e le
gend
to
Tab
le 1
.

TA
BL
E I
I1
The
eff
ect
of 1
#M
Tre
atm
ent
Glo
bula
r em
bryo
s C
ontr
ol,
day
0
Con
trol
, da
y 6
NP
A,
day
6
Hea
rt e
mbr
yos
Con
trol
, da
y 0
Con
trol
, da
y 6
NP
A,
day
6
NP
A o
n is
olat
ed f
ract
ions
of
glob
ular
and
hea
rt e
mbr
yos
a
Nor
mal
em
bryo
s (n
umbe
r/m
l)
Abn
orm
al e
mbr
yos
(num
ber/
ml)
CC
G
O
H
T
G
O
H
T
340
57
7 0
0 30
3
0 0
(7)
(5)
(2)
(3)
(2)
427
37
43
76
3 20
10
27
0
(15)
(2
) (2
) (1
3)
(2)
(3)
(3)
(3)
340
56
40
7 0
17
3 3
0 (2
0)
(4)
(6)
(4)
(3)
(2)
(2)
U
0 0 3 (2)
433
33
77
150
7 10
7
60
0 7
(30)
(5
) (2
) (1
5)
(2)
(3)
(3)
(3)
(3)
320
30
30
197
120
3 17
13
3 23
7
(17)
(7
) (3
) (1
7)
(15)
(2
) (3
) (1
1)
(2)
(2)
270
43
47
63
3 3
13
247
10
20
(4)
(2)
(11)
(5
) (1
) (2
) (2
) (9
) (2
) (4
)
Abo
rtiv
e em
bryo
s (n
umbe
r/m
l)
G
O
H
T
U
0 0
0 0
0
3 7
7 0
3 (2
) (2
) (2
) (2
) 0
10
3 0
0 (2
) (2
)
3 0
0 0
0 (2
) 0
3 3
0 0
(2)
(2)
7 0
7 0
0 (1
) (1
)
a F
ootn
ote
as i
n T
able
II.
60
1 3 ~ ~ 12
11
10
10
Control d 0
:5
g "I3
o
1.6.
1.5-
14-
13-
1,2-
11-
10
1.0. 1 31 1.24
1/01 1 0
13-
12-
11.
10
i i i i
A m
z~ a
zx zx
a
id-i-"'.! : .
i
i i i i i
zx
i i L i L
~x
zx
1 0 , I i L
100 200 300 t 4 0 0
Axial length (~m)
i i Control d 6
i i
1 /~M NPA
/ 4 TIBA'
o
J ,J 500
Fig. 3. Relationship of cotyledon arc / radius ratio to axial length for globular (C)), oblong (e), and heart (zx) embryos, treated with either 1 p~M NP A or 1 ~M TIBA for 6 days. Control day 0 and control day 6 data represent pooled day 0 and day 6 controls from NPA and TIBA experiments. Note ordinate expansion at 1.0 on all panels. Almost all embryos below 100 t~m in length (not illustrated) are globular embryos. The embryos were selected at random from three replicate culture wells for geometric analysis. Axial length refers to the length of the central axis of the embryo, the radius is one-half the maximum width of the embryo, and the cotyledon arc is the length of a radius plus the central axis of the cotyledon. The ratio of cotyledon arc to embryo radius distinguishes cotyledon-bearing embryos (ratio > 1.0) from those lacking cotyledons (ratio = 1.0). For additional details, see Schiavone and Cooke (1985).
few torpedo embryos are present at day 6, but at numbers equal to (TIBA), or below (NPA), the original number of torpedos in the day 0 controls. Since these abnormal heart embryos are not re- verting to callus, but continue to grow as unusual,
Figs. 4-7. Photomicrographs of selected abortive and abnormal embryos. Scale bars in each photograph represent 100 #m. Cotyledons in Figs. 4 and 5 are not visible since embryos are lying on their sides. Fig. 4 is a cluster of two abortive heart embryos. Note callus formation at the apical end of the vertical embryo, and additional callus formation at the basal end of the horizontal embryo. Fig. 5 is a single abortive torpedo, with callus forming along the sides of the hypocotyl, and to a lesser degree at the upper, apical end. Fig. 6 is a cluster of abnormal heart embryos. The lower embryo, lying horizontally, is pro- ducing a smooth bulge in the hypocotyl region. Fig. 7 is a single abnormal torpedo embryo, displaying pronounced en- largement of the hypocotyl area.
albeit organized structures, the auxin transport inhibitors are clearly affecting the polarized growth of developing embryos. The different effects of auxin analogues vs. transport inhibitors are il- lustrated in Figs. 4-7 . Representative abortive heart embryos from the auxin analogue treatment exhibit callus formation on the epidermal surface (Figs. 4, 5). Typical abnormal embryos produced in cultures treated with auxin transport inhibitors display new growth axes from the hypocotyl re- gion (Figs. 6, 7).

Discussion
The data presented in this paper support the concept that auxin regulates several different processes in somatic embryogenesis. These processes span much of the developmental history of the embryo, from the initiation of embryogene- sis to the maintenance of organization and polar- ity in heart and torpedo embryos. Considering the diversity of auxin effects, it is likely that auxin operates via several independent mechanisms. Starting with Reinert (1959) and Steward et al. (1964), numerous authors have observed that ex- ogenous auxins block the initiation of carrot em- bryogenesis in vitro. Although one report (New- comb and Wetherell, 1970) suggested that the weak auxin analogue 2,4,6-T promotes carrot em- bryogenesis in vitro, Fujimura and Komamine (1979) found that IAA, 2,4,6-T and a third ana- logue p-chlorophenoxyisobutyric acid are all ef- fective inhibitors of total embryogenesis. This ap- parent discrepancy was attributed to residual amounts of 2,4-D in the embryogenic medium used in the first study.
Preliminary observations of carrot callus raised in MSA medium suggest that endogenous free IAA has a concentration on the order of 1-10/~M (F.M. Schiavone and J.D. Cohen, unpublished results). Although the threshold concentration of exogenous auxin required to delay embryo forma- tion falls within this range (Fig. 1), this informa- tion provides no clue about the mechanism of auxin action, since either sensitive enzymatic reac- tions or internal gradients could be disrupted by exogenous applications. Furthermore, IAA and 2,4-D may form a variety of active and inactive conjugates, which make concentration data even more difficult to interpret.
The present paper demonstrates that the in- dividual stages of carrot embryos are sensitive to exogenous application of auxin analogues, such that treated embryos tend to exhibit the abortive morphology, i.e. they are directly reverting to un- differentiated callus (Table I). The embryos be- come increasingly sensitive to exogenous auxin as they progress to later stages. Nonetheless, auxin does not seem to directly cause any changes in the growth patterns of developing embryos, as re-
61
flected in the relatively low numbers of abnormal embryos.
The two inhibitors of polar auxin transport used in this work have profound effects on em- bryo development (Tables II and III; Fig. 3); both globular and oblong embryos are induced to grow to enormous sizes. Thus, it appears that the ini- tiation of polarized axial growth at the end of the globular stage, as well as the initiation of cotyle- don formation at the end of the oblong stage, involves the polar movement of auxin through the embryo. Similarly, large globular embryos are caused by treating embryogenic cultures with pro- line (Caligo et al., 1985).
In contrast, the transport inhibitors do not simply block the transitions of heart embryos to subsequent embryogenic stages, but rather cause the embryos to assume abnormal shapes. Most frequently, these abnormal heart embryos exhibit lateral bulges on their hypocotyls which appear to represent additional growth axes. Thus, the main- tenance of the polar axis of embryo growth in the heart stage seems to depend at least in part on polar auxin transport.
These effects of auxin analogues and transport inhibitors are interpreted in terms of the following model. Since auxin is basipetally transported from the shoot apex of most plants (Goldsmith, 1977), we assume that a similar transport can also occur in developing carrot embryos. This assumption is supported by earlier observations on mature em- bryos (Greenwood and Goldsmith, 1970; Fry and Wangermann, 1976), while the exact site of auxin production and the cells involved in polar trans- port are not known in the case of these embryos. It appears that globular embryos develop into oblong embryos via some mechanism which does not depend directly on the bulk concentration of auxin. Given that the globular-to-oblong transi- tion is affected by polar auxin transport inhibi- tors, it is conceivable that an embryo acquires the ability to transport auxin at the late globular stage. The resulting accumulation of auxin in the basal region causes these cells to elongate in the axial direction (see fig. 8 in Schiavone and Cooke, 1985), and thus the embryo assumes an oblong shape. Simultaneously, these basal cells may also become more sensitive to auxin; a gradient in

62
sensitivity would effect the same growth response as genuine basipetal transport. The initiation of cotyledons appears obligatorily dependent on polar auxin transport, because the transport in- hibitors mediate the formation of enlarged globu- lar and oblong embryos, with very few heart em- bryos arising in the culture. Maintaining polarized growth in the heart embryo also appears to require polar auxin transport; in addition, the embryos of this stage are especially sensitive to exogenous auxins. This enhanced sensitivity to both transport inhibitors and exogenous auxin may reflect the greater structural complexity of the heart embryo or a general increase in the embryo's sensitivity to localized auxin levels.
Acknowledgements
We wish to thank Dr. J.D. Cohen (Beltsville Agricultural Research Center) for his guidance and his gift of NPA. This work was supported in part by USDA Competitive Research Grant No. 83-CRCR-1255 to T.J. Cooke and R.H. Racusen. Scientific article number A-4519, Contribution number 7512 of the Maryland Agricultural Ex- periment Center, College Park, MD 20742, U.S.A.
References
Caligo, M.A., V. Nuti Ronchi and M. Nozzolini: Proline and serine affect polarity and development of carrot somatic embryos. Cell Differ. 17, 193-198 (1985).
Fry, S.C. and E. Wangermann: Polar transport of auxin through embryos. New Phytol. 77, 313-317 (1976).
Fujimura, T. and A. Komamine: Involvement of endogenous auxin in somatic embryogenesis in a carrot cell suspension culture. Z. Pflanzenphysiol. 95, 13-19 (1979).
Giuliano, G., D. Rosellini and M. Terzi: A new method for the purification of the different stages of carrot embryoids. Plant Cell Rep. 2, 216-218 (1983).
Goldsmith, M.H.M.: The polar transport of auxin. Annu. Rev. Plant Physiol. 28, 439-478 (1977).
Greenwood, M.S. and M.H.M. Goldsmith: Polar transport and accumulation of indole-3-acetic acid during root regenera- tion by Pinus lambertiana embryos. Planta 95, 297-313 (1970).
Murashige, T. and F. Skoog: A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol. Plant. 15, 473-497 (1962).
Newcomb, W. and D.F. Wetherell: The effects of 2A,6-trichlo- rophenoxyacetic acid on embryogenesis in wild carrot tis- sue cultures. Bot. Gaz. 131,242-245 (1970).
Raghavan, V.: Embryogenesis in Angiosperms: A Develop- mental and Experimental Study (Cambridge University Press, New York) 304 pp. (1985).
Reinert, J.: Uber die Kontrolle der Morphogenese und die Induktion von Adventivembryone an Gewebekulturen aus Karrotten. Planta 53, 318-333 (1959).
Schiavone, F.M. and T.J. Cooke: A geometric analysis of somatic embryo formation in carrot cell cultures. Can. J. Bot. 63, 1573-1578 (1985).
Steward, F .C , M.O. Mapes, A.E. Kent and R.D. Holsten: Growth and development of cultured plant cells. Science 143, 20-27 (1964).
Thomson, K-S., R. Hertel, S. Muller and J.E. Tavares: 1-N- Naphthylphthalamic acid and 2,3,5-triiodobenzoic acid: In vitro binding to particulate cell fractions and action on auxin transport in corn coleoptiles. Planta 109, 337-352 (1973).