isothermal calorimetry approach to evaluate tissue damage in carrot slices upon thermal processing
TRANSCRIPT

Journal of Food Engineering 65 (2004) 165–173
www.elsevier.com/locate/jfoodeng
Isothermal calorimetry approach to evaluate tissue damagein carrot slices upon thermal processing
Federico G�omez a,*, Romeo T. Toledo b, Lars Wads€o c, Vassilis Gekas a,d,Ingegerd Sj€oholm a
a Department of Food Engineering, Lund University, P.O. Box 124, SE-221 00 Lund, Swedenb Department of Food Science and Technology, The University of Georgia, Athens, GA 30602, USA
c Department of Building Materials, Lund University, P.O. Box 118, SE-221 00 Lund, Swedend Department of Environmental Engineering, Technical University of Crete, Chania, Greece
Received 24 July 2003; accepted 5 January 2004
Abstract
The purpose of this investigation was to test calorimetry as a means of quantifying cell damage occurring at various stages of
mild blanching treatments. Calorimetric measurements were carried out on cores taken from carrot slices blanched at 100 �C for 5–
45 s. The calorimetric data were corrected for the influence of wounding stress due to slicing and coring. The validity of calorimetry
to measure cell damage was assessed by simulating inactivation of cell metabolism based on bibliographic data on thermal inac-
tivation kinetics of plasma membrane Hþ-ATPase, used as a marker of overall cell membrane damage. Calorimetric data paralleled
the simulated decrease of actively metabolizing cells only in the first 20 s of blanching but diverged at higher blanching times. Our
results suggest that inactivation of enzymes responsible for quality-related changes during frozen storage (e.g. blanching at 100 �Cfor 20 s) may be accompanied by cell damage in about 70% of the tissue.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Calorimetry; Mild blanching; Cell damage; Hþ-ATPase
1. Introduction
In the frozen vegetable industry, blanching is used to
inactivate enzymes that cause off-flavour, changes in
colour and other chemical reactions during frozen
storage. Several blanching times and temperatures have
been reported for blanching carrot slices of various
thickness. These include treatments in boiling water (100
�C), 3–4 min (3–5 mm-thick slices) (Lee, Bourne, & VanBuren, 1979; Pr�estamo, Fuster, & Risue~no, 1998; Rah-
man, Henning, & Westcott, 1971) and 90 �C, 4 min
(Kidmose & Martens, 1999). These treatments were di-
rected towards the inactivation of peroxidase, however,
the use of this enzyme as an indicator of the blanching
process may result in undesirable quality changes (Kim
et al., 2001). The use of other enzymes as indicators of
the blanching process has also been suggested as a meanto minimise the heat treatment. Roy, Taylor, and Kra-
*Corresponding author. Tel.: +46-46-222-98-22; fax: +46-46-222-46
22.
E-mail address: [email protected] (F. G�omez).
0260-8774/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jfoodeng.2004.01.010
mer (2001) used lipoxygenase as a marker, obtaininggood retention of quality attributes after blanching 1-cm
carrot cubes at 100 �C for 0.58 min. These blanching
procedures, following a classical approach, are all based
on the average effect of the heat treatment in the sample.
Mathematical modelling for blanching optimization
has been directed towards a better understanding of
processes taking place during the heat treatment such as
turgor loss (De Belie, Herppich, & Baerdemaeker, 2000)and starch gelatinization (Verlinden & De Baerdemae-
ker, 1997). These models were based on a detailed
description of the temperature distribution in the sam-
ple, from the surface towards the centre. However, as
the thermal properties of the samples are usually taken
from the literature at a low, constant temperature, the
models failed to consider changes in these properties
during the heating process. Chang and Toledo (1990)reported a significant elevation of effective thermal dif-
fusivity as a result of internal convective heat transfer
due to penetration of heating fluid into the sample and
possible structural changes. Ignoring changes of thermal
conductivity with temperature would cause errors in the

Nomenclature
Subindex i slice section
Superscript k time interval
a thermal difussivity (m2 s�1)
b calibration factor of the line heat source
probe used for determination of thermalconductivity
� calibration coefficient of the calorimeter
(mWmV�1)
q density (kgm�3)
ln term in Eq. (6)
C fraction of thermal power contributed by
normal cell metabolic activity
Cd fraction of actively metabolizing cells (un-damaged and partially damaged) ¼ remained
activity of plasma membrane Hþ-ATPase
(1/s)
Cp specific heat (J kg�1 K�1)
I thermal conductivity probe heater input cur-
rent (A)
k thermal conductivity (Wm�1 K�1)
kd first order kinetics constant of inactivation ofplasma membrane Hþ-ATPase (1/s)
k0 kinetic constant of Arrhenius law ({}/s)
Ea activation energy (KJmol�1)
M mass of the sample (g)
n numerical index
P thermal power (mWg�1)
r variable distance from the centre axis of the
slab (m)R gas constant (8.314 J gmol�1 K�1)
Rk resistance/length of Constantan heating wire
for thermal conductivity measurement (X/m)
RS half-thickness of slab (m)
S slope of the linear portion of a plot temper-
ature vs. ln time
t time (s)
T temperature (�C)TM temperature of the blanching water (�C)T0 initial temperature (�C)VS voltage recorded from the sample in the cal-
orimeter (mV)
VBL voltage recorded from the base line in the
calorimeter (mV)
X thickness of slice (mm)
XW fraction of water contentW fraction of the total thermal power contri-
buted by the reaction to wounding stress
166 F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173
temperature profile calculations (Gratzek & Toledo,
1993) and in the proposed blanching models.
Starting from the surface of the heated sample,
blanching will lead to cell membrane disruption and
protein denaturation (Hansen, Afzal, Breidenbach, &
Criddle, 1994). Other processes such as turgor loss and
starch gelatinization will be a direct consequence of
membrane damage (Greve et al., 1994), influencingquality attributes such as firmness and crispness (De
Belie et al., 2000). Possibilities of mild blanching con-
ditions have been explored for preservation of tissue
integrity and quality improvement (Pala, 1983; Pizzo-
caro, Senesi, Querro, & Gasparoli, 1995). Mild blanch-
ing treatments directed towards avoidance of oxidative
phenomena in carrot lipids during frozen storage were
applied at temperatures as high as 100 �C for a fewseconds, a process designed to inactivate enzymes near
the surface of the carrot pieces. Samples kept in boiling
water for 20–30 s followed by immediate cooling in tap
water, appeared to exhibit fresh-like quality after frozen
storage (Pizzocaro et al., 1995), therefore, the process
may be potentially beneficial in minimising cellular
damage so that sensorial quality in the stored frozen
carrots is maximised. However, there are to ourknowledge no previous reports on how to measure the
extent of tissue damage during these mild blanching
treatments that can be used to set the optimal range of
the treatments.
Inactivation of cell metabolism, and hence, cell
damage due to high temperature injury has been as-
sessed using calorimetry (Hansen et al., 1994; Rank,
Breidenbach, Fontana, Hansen, & Criddle, 1991).
Calorimetric measures of the rate of heat production are
proportional to metabolic rates and provide a directindication of integrated metabolic responses such as
respiration and reaction to wounding stress (Criddle,
Breindenbach, & Hansen, 1991; Iversen, Wilhelmsen, &
Criddle, 1989). Calorimetry may be potentially useful to
quantify cell damage that occurs during processing
operations such as blanching. Increasing intensity of the
heat treatment would progressively reduce the number
of viable cells in the tissue and thus, result in a corre-sponding reduction in the rate of metabolic processes.
The purpose of this investigation has been to test
calorimetry as a means of quantifying the decrease of
cell viability in carrot tissue during processing and
hence, to evaluate tissue damage occurring at various
stages of the blanching treatment. A simulation was
carried out to calculate the extent of cell membrane
damage at different times of blanching at 100 �C basedon thermal inactivation kinetics of plasma membrane
Hþ-ATPase as a marker of overall membrane damage.

F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173 167
Plasma membrane Hþ-ATPase actively pumps protonsout of the cell creating pH and electrical potential dif-
ferences across the membrane. Many physiological
processes are driven by this electrochemical proton
gradient such as the transport of solutes into and out of
the cell. The enzyme also has an important role in reg-
ulating the cytoplasmic pH, and the pH gradient
developed over the membrane may also affect pH-sen-
sitive enzymes and plasma membrane channel proteins(Michelet & Bountry, 1995). Hþ-ATPase enzyme activ-
ity reflects both the physical and the physiological state
of the cell membranes (Gonz�alez-Mart�ınez, 2003; Stan-ley, 1991). Thus, by using thermal inactivation kinetics
of Hþ-ATPase, we attempted to have a marker of
overall membrane damage due to thermal process, as
membranes will lose their capacity to control the active
transport of ions and solutes into and out the cytoplasm(Gonz�alez-Mart�ınez, 2003).
2. Materials and methods
2.1. Raw material, handling and storage
Carrots (Daucus carota L. cv. Nerac), grown in the
south of Sweden, were used. The carrots, 400 kg, har-
vested in the last week of October 2001, were washed,
hydrocooled to about 3 �C and stored in one largepolyethylene plastic bag at 0 �C and approx. 100% rel-
ative humidity, as routinely practiced by the producer.
Medium-sized carrots (19.5 ± 1.0 cm in length, 3.0 ± 0.2
cm diameter) free from defects, were selected at the farm
storage facility and transported to our laboratory in
plastic bags kept on ice in insulated boxes. The carrots
were then taken out of the bags and placed in a closed,
refrigerated chamber at )0.2 �C and 100% relativehumidity until used.
2.2. Preparation of samples
Carrots were manually peeled, the top and tip re-
moved and the remainder was cut into slices. Slices 3.0–
3.2 cm diameter and 7 mm thick were selected and used
for the blanching experiments. After blanching, four tap
root cores 7 mm long and 11 mm in diameter were ex-cised longitudinally from the phloem parenchyma of
each slice with a cork borer. The cores were used for
isothermal calorimetry measurements and evaluation of
damaged cells with Trypan blue staining.
2.3. Blanching
Carrot slices was immersed in boiling water (100 �C)for a defined time and rapidly cooled in ice water to
minimise over-processing. Non-heated slices of the same
dimensions were used as control.
2.4. Isothermal calorimetry measurements on the
blanched samples
Isothermal calorimetry was performed with a proto-
type of the TAM Air isothermal calorimeter (Thermo-
metric AB, J€arf€alla, Sweden). This isothermal heat
conduction instrument contains eight twin calorimeters.
The instrument was placed inside an insulated holder that
was placed in a cooled incubator. This arrangement gaveenough temperature stability for the present measure-
ments. The samples were placed in 20 ml glass ampoules
sealed with teflon-coated rubber seals and aluminium
caps. Each calorimeter had its own reference which
consisted of a sealed 20 ml glass ampoule containing 4 ml
water. The eight calorimeters permitted eight simulta-
neous measurements of the rate of heat production.
Samples for calorimetry were four cores obtainedfrom the phloem parenchyma of raw and blanched
carrot slices. To perform the isothermal calorimetry
measurement, each sample was placed in a numbered
ampoule that was immediately weighed and sealed. In
this way samples consisting of a raw sample (control)
and seven other heat treated slices, were distributed in
eight ampoules. The sealed ampoules were immediately
placed in the calorimeters.The isothermal measurements were performed at 20
�C for a period of about 24 h. Baselines (BLs) were re-
corded before or after each measurement. The primary
output from the heat flow sensors in the calorimeters (a
voltage) was recorded by computer from the digitized
output of the calorimeters. The corresponding thermal
powers (heat production rates) were calculated by the
following equation:
P ¼ e�VS � VBL
Mð1Þ
were P (mWg�1) is the specific thermal power of the
carrot sample, e is the calibration coefficient of the cal-
orimeter (mWmV�1), VS is the voltage (mV) recorded
from the sample, VBL is the voltage (mV) recorded from
the base line and M is the mass (g) of the sample. Thecalibration coefficients were calculated from electrical
calibrations made at 20 �C.
2.5. Isothermal calorimetry measurements on the
blanched samples: influence of wounding on thermal power
When tissues are wounded, the cells at the site of such
a stress strengthen their walls by the secretion of addi-
tional structural components such as hydroxyproline-
rich glycoproteins, lignin or suberin (Satoh, Sturm,
Fujii, & Crispeels, 1992). To evaluate the metabolic heat
produced by these stressed cells near the cut surface andthus, estimate the contribution of wounding stress to the
total thermal power measured in the blanched samples,
a separate calorimetric experiment was carried out. Four

168 F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173
carrot root cores were submerged in boiling water forabout 2 s, a treatment that will damage only the cells
near the cut surface of the samples, and immediately
cooled in ice water. To perform the isothermal mea-
surement, the cores were placed in a numbered ampoule
which was immediately weighed and sealed. Four cores
of the same dimensions, taken from another slice of the
same carrot were kept raw as control and placed in a
second ampoule. The heat production rate from eachampoule was measured in a TAM Air instrument as
described previously for the blanched samples. The
difference in the thermal power between both samples,
as calculated with Eq. (1), will be a direct estimation of
the heat contributed by the cells near the cut surface.
The same experiment was repeated for each of the
blanching treatments. Four cores were obtained from
each of the blanched slices and placed in a calorimeterampoule. Four cores of the same dimensions, taken
from another slice, blanched for the same period of time,
were heated for about two additional seconds to damage
the cells from the surfaces that were not in direct contact
with the boiling water when the entire slice was blan-
ched, and placed in a second ampoule.
2.6. Evaluation of damaged cells using trypan blue
staining solution
Carrot root cores from the phloem parenchyma of
each slice used in the blanching experiment were treated
with Trypan blue staining solution (0.4%, Sigma-Al-
drich, USA). Trypan blue is a membrane-impermeabledye, which is not taken up by viable cells which have an
intact plasma membrane. Thus, damaged cells (non-
viable) stain and living cells (viable) do not (Danielson
et al., 2002). The cores were steeped 5 min in the staining
solution, washed with distilled water to remove excess of
dye and examined under a stereomicroscope (Olympus
SZ-CTV, Japan).
2.7. Determination of thermal and physical properties
The thermal conductivity, k (Wm�1 K�1), density, q(kgm�3) and moisture content were experimentally
determined and used to calculate heat penetration
through the slices during blanching treatments.The thermal conductivity was measured using a line
heat source probe as described by Gratzek and Toledo
(1993) with some modifications on the power supply and
measurement circuit. The probe was calibrated by
immersion in glycerine (k ¼ 0:2873 Wm�1 K�1) Carrots
with more or less uniform cylindrical shape were se-
lected, peeled and top and tip separated. The thermal
conductivity probe (with Teflon insulated Constantanheater wire and Teflon insulated Copper–Constantan
thermocouple wire enclosed in a 6.9 cm long, 1.27 mm
diameter stainless steel active probe section) was in-
serted along the axis of the 7.3 cm-long, cylindricalcarrot samples. The carrots, placed in a plastic mesh,
were carefully immersed in a thermostatic bath set at 20
�C. The temperature of both the surrounding fluid and
the sample were monitored until they were identical.
When the sample and surrounding water were at the
same temperature, the measurement was started. The
measurements were done by admitting a pre-set constant
current into the probe heating wire. The current levelwas determined by measurement of the voltage drop
across a high precision (0.05%) 1.0 ohm shunt resistor
with a digital voltimeter. A current level of 0.140 A was
used for all tests. The temperature signal was measured
at a sampling frequency of 200 Hz with a time averaging
each 0.15 s. Tests were usually run for 30 s. Time and
temperature information was recorded in a data acqui-
sition system (Fluke, USA).Thermal conductivity, k, was calculated from the
following (Gratzek & Toledo, 1993):
k ¼ b2I2Rk
4pSð2Þ
where IðAÞ is the probe heater input current, Rk (X/m) isthe resistance/length of Constantan heating wire, and bthe calibration factor determined during the calibration
of the line heat source probe with glycerin. The
numerator is multiplied by 2 because the wire doubled
back in the probe. S is the slope of the linear portion of a
plot DT vs. ln time. DT is the temperature rise from the
equilibrium temperature as recorded by the computer
from the output of the probe thermocouple.The change of thermal conductivity with increasing
temperature was measured. When the temperature of
the surrounding water was increased, the temperature of
the sample was monitored until equilibrium was
reached, at which time a new thermal conductivity
measurement was initiated. After a pause to re-establish
equilibrium, another measurement was made at the
same temperature. After completing measurements at aparticular temperature, the water was heated and the
procedure repeated at the next temperature level.
The density of carrot pieces was measured at 20 �Cwith an ultrapycnometer 1000 (Quantachrome, USA)
using helium as the fluid. Sample moisture was deter-
mined after drying the sample (ca 3 g fresh weight) for
18 h in a vacuum-oven (VWR Scientific Products, USA)
at 70 �C and 380 mm Hg of absolute pressure.
3. Results and discussion
3.1. The thermal power decreases as the intensity of the
blanching process increases
An example of the results from the calorimetric
measurements of raw and blanched carrots is shown in

F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173 169
Fig. 1a. Each time an ampoule was charged into acalorimeter there was an initial disturbance lasting for at
least 30 min. After that, the true voltage (mV) from the
samples was measured. In the unblanched sample (0 s),
the voltage increased and then reached a short plateau
after about 8 h, followed by a decrease that could be
attributed to the depletion of oxygen in the ampoule
(Criddle, Breidenbach, Lewis, Eatough, & Hansen,
1988). In the 5 and 10 s-blanched samples, the voltageincreased during the first 8 h of the measurement al-
though it did not increase as much as that of the raw
sample. The following plateau was fairly constant until
the experiment was interrupted. In the 20, 30 and 45 s-
blanched samples the voltage did not increase at the
beginning of the measurements and was fairly constant
during the time course of the experiment with a slight
tendency to increase in the last hours. The curve corre-sponding to the 45 s-blanched sample was fairly con-
stant and very close to that of the baseline. Interestingly,
there were remarkable differences in the voltage values
Fig. 1. (a) Calorimeter output of blanched carrot samples at 100 �C for
different time intervals. Blanching time is indicated next to each curve.
The experiment was interrupted after 24 h and the baseline (BL) was
recorded. (b) Relative thermal power (heat production rates) of carrots
with increasing blanching time. The thermal power, P , calculated with
Eq. (1) was normalized by dividing values with those of the raw carrot
(control). Means of three measurements are shown.
from the different samples, decreasing with increasingblanching time. The experiment was interrupted after 24
h by taking the samples out from the calorimeter. This
interruption caused a disturbance lasting for at least 30
min before the BL started to be recorded.
The constant voltage plateau of the curves was used
to calculate thermal power according to Eq. (1). Fig. 1b
describes the decrease in the thermal power of carrots
blanched at 100 �C for different time intervals.The thermal power of the carrot samples decreased
sharply during the first 20 s of blanching and then re-
ceded slowly with prolonged heating to reach a very low
value, 1.6% after 45 s. Since the thermal power is a
parameter related with metabolic activity of the cells in
the tissue (Criddle et al., 1991), our results suggests that
the amount of living cells by that time is very low or
null. In support of this idea, Fig. 2 shows that the carrottissue blanched 45 s exhibited complete staining with the
Trypan blue, indicating that viable cells are no longer
present. This result is in agreement with Greve et al.
(1994) who could not identify any living cells in 1 cm-
thick carrot slices blanched at 100 �C for 60 s using a
flourescein diacetate dye.
3.2. Estimation of the reaction to wounding stress
We could consider the thermal power values (P in
Fig. 1b) to be the result of the sum of the energy released
by two main physiological processes, that from the
‘‘normal’’ cell metabolic activity ðCÞ and that originat-
ing from wounding stress produced by the cells near the
cut surface of the cores ðW Þ:P ¼ W þ C ð3Þ
where W is the result of the experimental estimation ofthe heat contributed by the cells near the cut surface.
The contribution of wounding stress to the thermal
power is summarised in Table 1. The results show that
carrot metabolism is highly influenced by the wounding
reaction. In the raw carrot sample, the contribution of
the reaction to wounding stress to the total thermal
power was 52% and, surprisingly, in the 30 s-blanched
samples this contribution was 41%.
3.3. Simulation of cell damage during blanching treat-
ments
3.3.1. Kinetic parameters
Cell membrane damage upon heating has been re-
cently investigated by Gonz�alez-Mart�ınez (2003). The
authors used vesicles isolated from potato cell mem-
brane to test inactivation of plasma membrane Hþ-ATPase activity as a marker for cell membrane damage.
Eppendorf tubes containing isolated cell membranes in
suspension were heated at selected temperatures and

Table 1
Fractions of ‘‘normal’’ cellular metabolism, C and wounding stress, Wof the total thermal power (heat production rates), P (plotted in Fig.
1b), measured with isothermal calorimetry in cores taken from carrot
slices blanched at 100 �C for various time intervals
Blanch-
ing time
(s)
P (mW) C1 W a P �C(mW)
0 0.3908 0.484 0.516 0.189
5 0.2462 0.498 0.502 0.132
10 0.1911 0.552 0.448 0.105
20 0.0824 0.559 0.441 0.046
30 0.0469 0.595 0.405 0.028
45 0.007 1.000 – 0.007
a The values are the mean of duplicate determinations; variations
within duplicates were <5%.
Fig. 2. Front and side of cores obtained from carrot slices blanched at 100 �C for different times and stained with Trypan blue, as indicated in the
Materials and methods section. Damaged cells were stained (dark areas) while intact cells did not absorb the stain.
170 F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173
times and the Hþ-ATPase activity was measured after
the heat treatment to determine the inactivation kinetics
of the enzyme. The authors determined Hþ-ATPase
activity at 20 �C according to Palmgren and Sommarin(1989). In this method, ATP hydrolysis is coupled
enzymatically to the oxidation of NADH and the rate of
hydrolysis is measured as the decrease in absorbance of
NADH at 340 nm. The activity was determined after the
addition of 0.05% (w/v) polyoxyethylene-20-cetylether
(Brij 58) that produced 100% inside-out vesicles suitable
for the activity measurements.
The kinetics of inactivation of this enzyme showed tobe first order:
dCd
dt¼ �kdCd ð4Þ
The temperature dependence of the kinetics constant kdcould be expressed by two linear Arrhenius plots:
kd ¼ koe�EaRT ð5Þ
The first was valid in the range between 45 and 53 �Cand the second was valid in the range between 60 and 75
�C. The authors calculated 55 �C as a transition tem-
perature represented by the intercept of the two linear
plots. The kinetic parameters for inactivation of Hþ-
ATPase are summarised in Table 2. We here assume the
inactivation of Hþ-ATPase, an indicator of cell mem-brane damage, to be proportional to the decrease in cell
metabolic activity.
3.3.2. Calculation of temperature profile
The temperature distribution in the carrot slice dur-
ing the blanching treatments was calculated as follows.
The carrot slice, considered as an infinite slab, was
divided horizontally, from the surface to the centre, into
7 sections of 0.5 mm each. The change of temperature, T(�C) with time, tðsÞ, in the centre of each of the slabsections during blanching, was calculated with the fol-
lowing equation (Singh, 1992):
T � TMT0 � TM
¼X1n¼1
2
lnð�1Þnþ1
coslnrRS
exp
�� l2
n
atR2S
�ð6Þ
where RS (m) is the half-thickness of a slab and r (m) isthe variable distance from the centre axis, TM is the
temperature of the blanching water (100 �C), T0 is the
initial temperature (20 �C), ln is defined by

Table 2
Parameters of the first-order kinetics for inactivation of Hþ-ATPase in
plasma membranes isolated from potato tubers (adapted from
Gonz�alez-Mart�ınez (2003), Fig. 2)
Temperature (�C) kd (1/s)
45 0.070
49 0.110
53 0.244
60 0.372
38 0.372
75 0.507
F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173 171
ln ¼ ð2n� 1Þ p2
ð7Þ
a, the thermal difussivity (m2 s�1), is calculated by
a ¼ kqCp
ð8Þ
where the increase of the thermal conductivity, k, withtemperature (Fig. 3) was measured as
k ¼ 0:5464þ 0:0012T � 2:0� 10�6T 2 ðr2 ¼ 0:9838Þð9Þ
The measured density, q ¼ 1154 kgm�3 and the specific
heat, Cp ¼ 3736 J kg�1 K�1 as calculated with the mois-
ture content of the carrots according to Siebel (1918):
Cp ¼ 0:837þ 3348 Xw ð10Þ
The summatory term in Eq. (6) was repeated three
times.
93.7˚ C81.2˚ C69.4˚ C59.1˚ C50.8˚ C45.1˚ C42.1˚ C
Xi = 0.5 mm
˚
˚˚˚˚˚
100˚C
3.3.3. Decrease of cell metabolic activity during blanching
The effect of the temperature distribution in the car-
rot sample during blanching treatments on the decrease
of cell metabolic activity was investigated as follows.
The fraction of actively metabolizing cells (undam-aged and partially damaged) Cdi at the centre of each of
the slice sections i as a function of time (superscript k)was calculated by solving Eq. (4):
0.540
0.560
0.580
0.600
0.620
0.640
10 30 50 70 90
k (w
/m°K
)
Temperature (°C)
Fig. 3. Carrot thermal conductivity as a function of temperature.
Ckþ1di ¼ Ck
di � kT ki
d CkdiDt ð11Þ
where kd is a function of the temperature at the centre of
each element i at time k, according to Eq. (5). Fig. 4a
and b shows an example of the result of the calculation,showing both the values of temperature and the fraction
of actively metabolizing cells, Cdi in the centre of each of
the slice sections of thickness X after 20 s of blanching at
100 �C.The fraction of actively metabolizing cells, Cd , in the
whole slab is obtained by averaging all Cdi at each time
step:
Cd ¼2P
XiCdi
Xð12Þ
3.4. Fitting the calorimetric data to the simulated decrease
of cell metabolic activity
The values of thermal power P , plotted in Fig. 1b and
reported in Table 1, were multiplied by the fraction ofthe ‘‘normal’’ cell metabolic activity, C (Table 1), to
obtain the value of thermal power that excludes the heat
produced by the reaction to wounding stress of the cells
near the cut surface. The resulting values (P �C in Table
1) are plotted in Fig. 5, expressed as relative to the one
of the control. These values are fitted in the same figure
to the fraction of actively metabolizing cells, Cd .
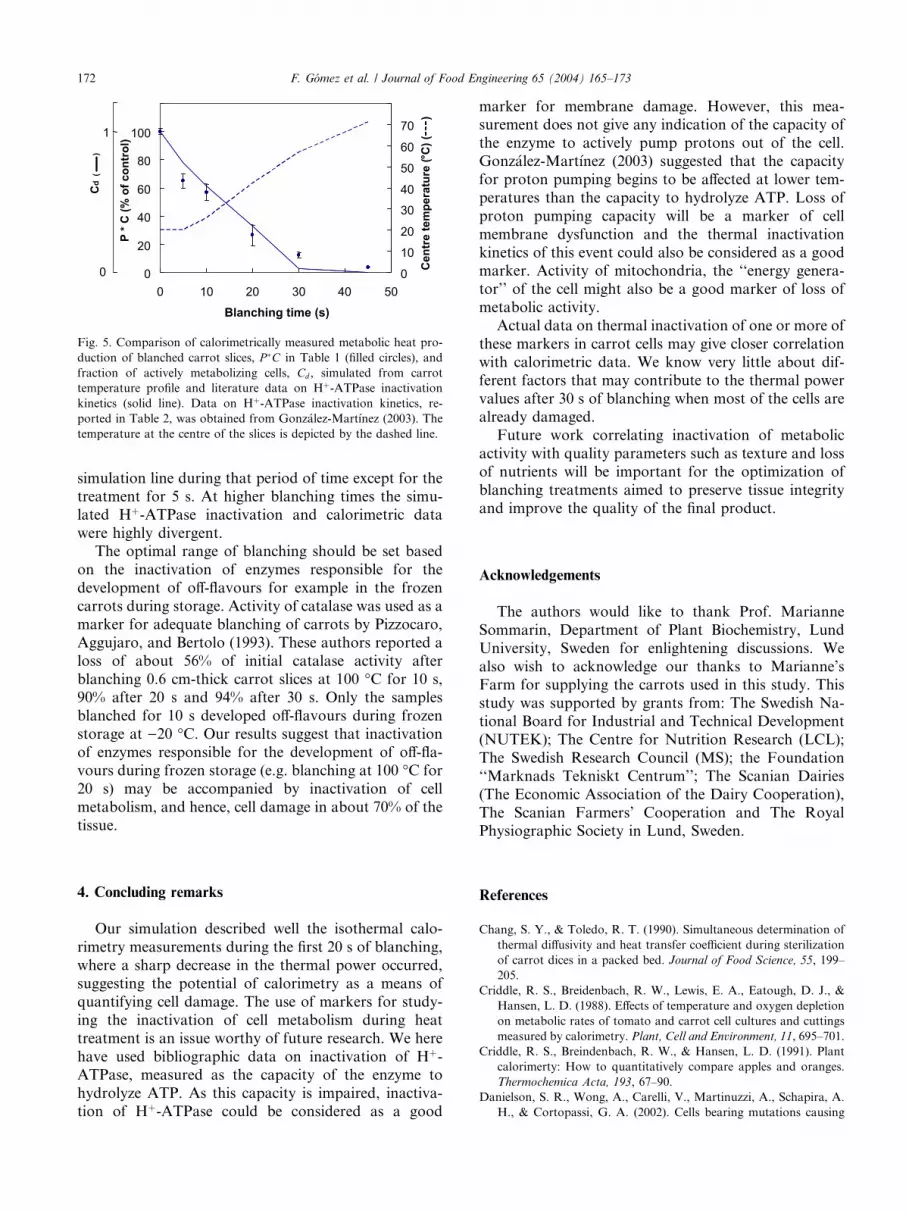
Fig. 5 shows that the decrease of cell metabolicactivity simulated using the inactivation kinetics of Hþ-
ATPase parallels the calorimetric data adequately for
the first 20 s of blanching. The error bars enclose the
Centre
Surface
0
00.0075
0.0956
0.53960.8144
0.9012
Centre
Xi = 0.5 mm
(b)
(a)
Fig. 4. (a) Schematic representation of the calculated temperature
distribution in a carrot slice after 20 s of blanching in water at 100 �C.(b) Schematic representation of the distribution of the fraction of ac-
tively metabolizing cells, Cdi, in a carrot slice after 20 s of blanching in
water at 100 �C (calculated from temperature distribution and thermal
inactivation kinetics of Hþ-ATPase).
0
20
40
60
80
100
0 10 20 30 40 50
Blanching time (s)
P * C
(% o
f con
trol
)
0
10
20
30
40
50
60
70
Cen
tre
tem
pera
ture
(°C
) (
)
Cd
(
)
0
1
Fig. 5. Comparison of calorimetrically measured metabolic heat pro-
duction of blanched carrot slices, P �C in Table 1 (filled circles), and
fraction of actively metabolizing cells, Cd , simulated from carrot
temperature profile and literature data on Hþ-ATPase inactivation
kinetics (solid line). Data on Hþ-ATPase inactivation kinetics, re-
ported in Table 2, was obtained from Gonz�alez-Mart�ınez (2003). The
temperature at the centre of the slices is depicted by the dashed line.
172 F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173
simulation line during that period of time except for the
treatment for 5 s. At higher blanching times the simu-
lated Hþ-ATPase inactivation and calorimetric data
were highly divergent.The optimal range of blanching should be set based
on the inactivation of enzymes responsible for the
development of off-flavours for example in the frozen
carrots during storage. Activity of catalase was used as a
marker for adequate blanching of carrots by Pizzocaro,
Aggujaro, and Bertolo (1993). These authors reported a
loss of about 56% of initial catalase activity after
blanching 0.6 cm-thick carrot slices at 100 �C for 10 s,90% after 20 s and 94% after 30 s. Only the samples
blanched for 10 s developed off-flavours during frozen
storage at )20 �C. Our results suggest that inactivation
of enzymes responsible for the development of off-fla-
vours during frozen storage (e.g. blanching at 100 �C for
20 s) may be accompanied by inactivation of cell
metabolism, and hence, cell damage in about 70% of the
tissue.
4. Concluding remarks
Our simulation described well the isothermal calo-
rimetry measurements during the first 20 s of blanching,
where a sharp decrease in the thermal power occurred,
suggesting the potential of calorimetry as a means of
quantifying cell damage. The use of markers for study-
ing the inactivation of cell metabolism during heat
treatment is an issue worthy of future research. We here
have used bibliographic data on inactivation of Hþ-ATPase, measured as the capacity of the enzyme to
hydrolyze ATP. As this capacity is impaired, inactiva-
tion of Hþ-ATPase could be considered as a good
marker for membrane damage. However, this mea-surement does not give any indication of the capacity of
the enzyme to actively pump protons out of the cell.
Gonz�alez-Mart�ınez (2003) suggested that the capacity
for proton pumping begins to be affected at lower tem-
peratures than the capacity to hydrolyze ATP. Loss of
proton pumping capacity will be a marker of cell
membrane dysfunction and the thermal inactivation
kinetics of this event could also be considered as a goodmarker. Activity of mitochondria, the ‘‘energy genera-
tor’’ of the cell might also be a good marker of loss of
metabolic activity.
Actual data on thermal inactivation of one or more of
these markers in carrot cells may give closer correlation
with calorimetric data. We know very little about dif-
ferent factors that may contribute to the thermal power
values after 30 s of blanching when most of the cells arealready damaged.
Future work correlating inactivation of metabolic
activity with quality parameters such as texture and loss
of nutrients will be important for the optimization of
blanching treatments aimed to preserve tissue integrity
and improve the quality of the final product.
Acknowledgements
The authors would like to thank Prof. Marianne
Sommarin, Department of Plant Biochemistry, Lund
University, Sweden for enlightening discussions. We
also wish to acknowledge our thanks to Marianne’sFarm for supplying the carrots used in this study. This
study was supported by grants from: The Swedish Na-
tional Board for Industrial and Technical Development
(NUTEK); The Centre for Nutrition Research (LCL);
The Swedish Research Council (MS); the Foundation
‘‘Marknads Tekniskt Centrum’’; The Scanian Dairies
(The Economic Association of the Dairy Cooperation),
The Scanian Farmers’ Cooperation and The RoyalPhysiographic Society in Lund, Sweden.
References
Chang, S. Y., & Toledo, R. T. (1990). Simultaneous determination of
thermal diffusivity and heat transfer coefficient during sterilization
of carrot dices in a packed bed. Journal of Food Science, 55, 199–
205.
Criddle, R. S., Breidenbach, R. W., Lewis, E. A., Eatough, D. J., &
Hansen, L. D. (1988). Effects of temperature and oxygen depletion
on metabolic rates of tomato and carrot cell cultures and cuttings
measured by calorimetry. Plant, Cell and Environment, 11, 695–701.
Criddle, R. S., Breindenbach, R. W., & Hansen, L. D. (1991). Plant
calorimerty: How to quantitatively compare apples and oranges.
Thermochemica Acta, 193, 67–90.
Danielson, S. R., Wong, A., Carelli, V., Martinuzzi, A., Schapira, A.
H., & Cortopassi, G. A. (2002). Cells bearing mutations causing

F. G�omez et al. / Journal of Food Engineering 65 (2004) 165–173 173
Leber’s hereditary optic neuropathy are sensitized to fas-induced
apoptosis. The Journal of Biological Chemistry, 277, 5810–5815.
De Belie, N., Herppich, W., & Baerdemaeker, J. (2000). Turgor
changes in red cabbage during mild heat treatment. Journal of
Plant Physiology, 157, 263–272.
Gonz�alez-Mart�ınez, G. (2003). Heat induced cell membrane injury of
vegetable tissues––an applied study on potatoes. Thesis, Food
Engineering Department, Lund University, Sweden.
Gratzek, J. P., & Toledo, R. T. (1993). Solid food thermal conductivity
determination at high temperatures. Journal of Food Science, 58,
908–913.
Greve, L. C., Shackel, K. A., Ahmadi, H., McArdle, R. N., Gohlke, J.
R., & Labavitch, J. M. (1994). Impact on heating on carrot
firmness: Contribution of cellular turgor. Journal of Agricultural
and Food Chemistry, 42, 2896–2899.
Hansen, L. D., Afzal, M., Breidenbach, R. W., & Criddle, R. S. (1994).
High- and low-temperature limits to growth of tomato cells.
Planta, 195, 1–9.
Iversen, E., Wilhelmsen, E., & Criddle, R. S. (1989). Calorimetric
examination of cut fresh pineapple metabolism. Journal of Food
Science, 54, 1246–1249.
Kidmose, U., & Martens, H. J. (1999). Changes in texture, micro-
structure and nutritional quality of carrot slices during blanching
and freezing. Journal of the Science of Food and Agriculture, 79,
1747–1753.
Kim, M. J, Oh, J. M., Cheon, S. H., Cheong, T. K., Lee, S. H., Choi, E.
U., Lee, H. G, Park, C. S., & Park, K. H. (2001). Thermal
inactivation kinetics and application of phospho- and galactolipid-
degrading enzymes for evaluation of quality changes in frozen
vegetables. Journal of Agricultural and Food Chemistry, 49, 2241–
2248.
Lee, C. Y., Bourne, M. C., & Van Buren, J. P. (1979). Effect of
blanching treatments on the firmness of carrots. Journal of Food
Science, 44, 615–616.
Michelet, B., & Bountry, M. (1995). The plasma membrane Hþ-
ATPase––a highly regulated enzyme with multiple physiological
functions. Plant Physiology, 108, 1–6.
Pala, M. (1983). Effect of different pretreatments on the quality of deep
frozen green beans and carrots. International Journal of Refriger-
ation, 6, 237–246.
Palmgren, M. G., & Sommarin, M. (1989). Lysophosphatidylcholine
stimulates ATP dependent proton accumu€olation in isolated oat
root plasma membrane vesicles. Plant Physiology, 90, 1009–
1014.
Pizzocaro, F., Aggujaro, R., & Bertolo, G. (1993). Kinetics of thermal
inactivation of oxidases in carrot disks during blanching. Rivista di
Scienzia dell’Alimentazione, 22, 279–285.
Pizzocaro, F., Senesi, E., Querro, O., & Gasparoli, A. (1995).
Blanching effect on carrots. Study of the lipids stability during
the frozen conservation. Industrie Alimentari, 34, 1265–1272.
Pr�estamo, G., Fuster, C., & Risue~no, M. C. (1998). Effects of
blanching and freezing on the structure of carrots cells and their
implications for food industry. Journal of the Science of Food and
Agriculture, 77, 223–229.
Rahman, A. R., Henning, W. L., & Westcott, D. E. (1971).
Histological and physical changes in carrots as affected by
blanching, cooking, freezing, freeze drying and compression.
Journal of Food Science, 36, 500–502.
Rank, D. R., Breidenbach, R. W., Fontana, A. J., Hansen, L. D., &
Criddle, R. S. (1991). Time-temperature responses of tomato cells
during high-and low-temperature inactivation. Planta, 185, 576–
582.
Roy, S. S., Taylor, T. A., & Kramer, H. L. (2001). Textural and
ultrastructural changes in carrot tissue as affected by blanching and
freezing. Journal of Food Science, 66, 176–180.
Satoh, S., Sturm, A., Fujii, T., & Crispeels, M. J. (1992). cDNA
cloning of an extracellular dermal glycoprotein of carrot and its
expression in response to wounding. Planta, 188, 432–438.
Siebel, J. E. (1918). Compend of mechanical refrigeration and engineer-
ing (9th ed.). Chicago: Nickerson and Collins.
Singh, R. P. (1992). Heating and cooling processes for foods. In D. R.
Heldman & D. B. Lund (Eds.), Handbook of food engineering (pp.
247–276). New York: Marcel Dekker.
Stanley, D. W. (1991). Biological membrane deterioration and
associated quality losses in food tissues. Critical Reviews in Food
Science and Nutrition, 30, 487–553.
Verlinden, B. E., & De Baerdemaeker, J. (1997). Modeling low
temperature blanched carrot firmness based on heat induced
processes and enzyme activity. Journal of Food Science, 62, 213–
229.