ultrastructure and immunocytochemistry of the neuroepithelial bodies in the lung of the tiger...
TRANSCRIPT

THE ANATOMICAL RECORD 234:419-431 (1992)
Ultrastructure and lmmunocytochemistry of the Neuroepithelial Bodies in the Lung of the Tiger Salamander, Ambystoma tigrinum
(U rodela, Am ph i bia) L. GONIAKOWSKA-WITALINSKA, J.M. LAUWERYNS, G. ZACCONE, S. FASULO,
AND G. TAGLIAFIERRO Department of comparative Anatomy, Jagielloonian University, Krakow, Poland (L.G. - W.);
Department of Histopathology, Catholic University of Leuven, Belgium (J.M.L.); Department of Animal Biology and Marine Ecology, University of Messina, Italy (G.Z.,
S.F); and Institute of Comparative Anatomy, University of Genoa, Italy (G.T.)
ABSTRACT Light and electron microscopy of the lungs of Ambystoma tigrinum (Urodela) revealed a relatively complex pattern of the neuroen- docrine (NE) cells. In the apical parts of smaller septa single NE cells not associated with nerve fibres were covered and surrounded by pneumo- cytes. The larger septa possessed small areas of ciliated epithelium, in which the NE cells were grouped in a form of neuroepithelial bodies (NEB) consisting of 3-5 cells and covered by goblet cells. NE cells possessed a large nucleus with patches of condensed chromatin, clear cytoplasm, and membrane-bound vesicles of variable morphology and size, containing an electron dense interior surrounded by a lucent space. The size of these dense core vesicles (DCV) ranged from 70-140 nm, while rarely the larger ones exhibited a diameter of 300-600 nm. In some NEB a second type of NE cells was observed for the first time in an amphibian species: these cells communicated with the air space and exhibited on their surface microvilli and a single modified cilium with a 8+ 1 microtubule arrangement. Their cytoplasm contained two types of DCV: dense core granules with a diam- eter of 140-260 nm and vesicles 320-700 nm in diameter with a moderately electron dense interior. The NEB were associated with intracorpuscular, sensory nerve terminals morphologically afferent and efferent. By immu- nocytochemistry, the NE cells revealed the presence of serotonin, met-en- kephalin, and leu-enkephalin. A paracrine and chemoreceptor role is pro- posed for NEB of Ambystoma tigrinum.
Key words: Neuroendocrine cells, Pulmonary epithelium, Salamander
Q 1992 Wiley-Liss, Inc.
The respiratory epithelium of many vertebrates re- veals the presence of solitary neuroendocrine (NE) cells (Frohlich, 1949; Lauweryns and Peuskens, 1969; Cutz and Conen, 1972) and/or groups of these cells associ- ated with nerve endings and termed neuroepithelial bodies (NEB) by Lauweryns and Peuskens (1972). The investigations of NE cells have been mostly focused on mammals (for review see Scheuermann, 1987; Sorokin and Hoyt, 19891, but data concerning other vertebrates such as birds (Cook and King, 1969; Wasano and Ya- mamoto, 1979; Walsch and MacLelland, 1974) and rep- tiles (Wasano and Yamamoto, 1976; Scheuermann et al., 1983; Pastor et al., 1987, 1989) are also available. The literature dealing with the morphology and im- munohistochemistry of NE cells in the lungs of am- phibians is rather scarce (Rogers and Haller, 1978, 1980; Wasano and Yamamoto, 1978; Goniakowska- Witalinska, 1981; Cutz et al., 1986; Goniakowska- Witalinska and Cutz, 1990; Goniakowska-Witalinska et al., 1990). As far as Urodela are concerned, solitary noninnervated NE cells were observed in the larval lung of Salamandra salamandra (Goniakowska- 0 1992 WILEY-LISS, INC.
Witalinska, 1982) and simple NE cells associated with efferent nerve endings were found in the ciliated epi- thelium of the newt Triturus alpestris lung. These cells were serotonin-negative (Goniakowska-Witalinska, 1980), although further studies revealed the occur- rence of bombesin in their cytoplasm (Cutz et al., 1986). Apart from solitary NE cells, Matsumura (1985) ob- served neuroepithelial bodies in the pulmonary epithe- lium of semiaquatic salamander Hynobius nebulosus, while Scheuermann et al. (1989) described single and grouped NE cells in the lungs of entirely aquatic Am- bystoma mexicanum. The NE cells described so far in the lungs of tailed amphibians corresponded with the NE cell type common for all the vertebrates studied,
Received May 1, 1991; accepted February 25, 1992. The ultrastructural data of this paper were presented at the Third
International Congress of Vertebrate Morphology, Antwerp 1989 and published as an abstract.
Address reprint requests to L. Goniakowska-Witalinska, Depart- ment of Comparative Anatomy, Jagiellonian University, R. Ingar- dena 6, 30-060 Krakow, Poland.

420 L. GONIAKOWSKA-WITALINSKA ET AL.
i.e., cells containing characteristic small, electron- dense granules. In this paper we report the occurrence of NEB in the lung of the terrestrial salamander Am- bystoma tigrinum; they have innervation atypical for amphibians and consist of two cell types: the usual cells with small dense core vesicles and an unusual type of NE cell with very large dense core vesicles and a mod- ified cilium protruding into the air space.
MATERIALS AND METHODS lmmunohistochemistry
Pieces of the lung tissue were fixed with 4% parafor- maldehyde buffered to pH 7.2 with 0.05 M phosphate for 12 h and rinsed in 0.1 M phosphate buffer a t + 4°C. The tissues were embedded in Paraplast. Sections of 5 p.m were placed on clear glass slides. These sections were subjected to the peroxidase-antiperoxidase method of Sternberger (1979) and the indirect fluores- cence technique of Coons et al. (1955). In order to re- duce the nonspecific binding of proteins, the sections were incubated with normal goat serum (NGS 1:30,30 min) diluted in phosphate-buffered saline (PBS) 0.1 M at pH 7.4. The PAP procedure was carried out in PBS containing 1% NGS. Dewaxed sections were rinsed in PBS, pretreated with 2% hydrogen peroxide in PBS, and incubated in a moist chamber at room temperature for 24 h with the antisera raised in rabbit against se- rotonin (INCSTAR, USA, diluted 1500 in PBS), syn- thetic met-benkephalin (Cambridge Research Bio- chemical Limited, CRB, England, diluted 1:200 in PBS), synthetic leu-5-enkephalin (CRB, England di- luted 1:200 in PBS), and amphibian bombesin (CRB, England diluted 1:400). Following a rinse in PBS (30 min), the sections were incubated in moist chamber for 1 h at room temperature with goat anti-rabbit gamma globulin conjugated with fluorescein isothiocyanate (FITC, Dakopatts, Denmark, diluted 1 : l O O in PBS) or with anti-rabbit IgG (H + L) peroxidase (Chemical Cre- dential ICN, Immunobiologicals, England, diluted 1: 100 in PBS) and rinsed in PBS. Peroxidase activity was detected by fresh solution of 3,3’ diaminobenzidine, 4 HC1 (DAB), 30 mg/100 ml, and H,02 (0.01%) in PBS (5 min); then the sections were dehydrated, and mounted in DPX. The specificity of the antisera were tested by absorption overnight a t + 4°C with different quanti- ties (10-50 p.g/ml of diluted antiserum) of their corre- sponding haptens. Additional control procedures in- cluded omission of the specific antiserum, or its substitution by a non-immune serum and the use of known positive tissue, e.g., mammalian nervous tissue and mammalian lung tissue.
Electron Microscopy Seven adult males and females (length 22 cm, weight
60 g) of Ambystoma tigrinum were purchased from a commercial supplier (Dr. W. De Rover, Herpetologisch Instituut and Dr. Ce Den Dolder, The Netherlands). The animals were kept for 3 weeks and fed with earth- worms. Before being sacrified they were anaesthetized with 0.01% MS 222. Lung tissue was immediately fixed in a cold mixture of two parts of 1% osmium tetroxide to one part 2.5% glutaraldehyde, cacodylate-buffered to pH 7.4, rinsed in cacodylate buffer, and quickly dehy- drated in ethanol to be embedded in Epon. Semithin sections were stained with toluidine blue and exam-
ined under the light microscope. Ultrathin serial sec- tions were mounted on Formvar-coated grids, stained with uranyl acetate and lead citrate, and examined in a Phillips CM 10 electron microscope at 60 kV.
RESULTS General Remarks
The lungs of Ambystoma tigrinum possess several septa of the first, second, and third order which divide the internal lung space into several irregular cham- bers. The apical part of the 1st and IInd order septa are thickened by the occurrence of smooth muscle cells and blood vessels. Both sides of the septa and their apical parts are covered by one type of pneumocytes charac- teristic for the lungs of amphibia. Only in the apical part of the first order septa small dispersed patches of ciliated cells are observed (Fig. la). Single neuroendo- crine (NE) cells occur intercalated and covered by pneumocytes in the apical part of the second order septa, while the first order septa reveal the presence of neuroepithelial bodies (NEB) surrounded and covered by goblet cells. In some semithin sections, two NEB can be found on the top and side of the apical part of the septum. The distance between the NEB base and the large vessel inside the septum ranges from 14 to 66 p.m.
lmmunohistochemistry Endocrine cells occur as single or in groups of two to
five cells. Immunostaining with antibody against sero- tonin showed a positive reaction (Fig. lb) in both single and clusters cells. Met-5-enkephalin immunostaining was observed in grouped cells only (Fig. lc), while sin- gle cells sometimes communicating with the air space were leu-5-enkephalin-positive (Fig. Id). Immuno- staining with bombesin antiserum was negative in all lung specimens, although control sections from mam- malian lungs (adult rats) showed a strong immunore- activity.
Electron Microscopy All neuroepithelial bodies (NEB) consist of 3-5
round or oval neuroendocrine (NE) cells, surrounded and covered exclusively by goblet cells (Fig. 2). Soli- tary NE cells, ultrastructurally identical with those of NEB, are intercalated between pneumocytes on smaller septa (Fig. 3). The size of NE cells ranges from 13 to 16 p.m. Both NEB and single NE cells are local- ized in the simple epithelium, being separated from the air space by a cytoplasmic layer (0.1 to 4.0 p.m thick) belonging to the surrounding cells (Figs. 2, 3, 10). In some cases this layer is discontinuous, allowing a di- rect communication of the NE cells with the air space. Some images even suggest that NE cells can be ex- pulsed towards the lung lumen (Fig. 10). The goblet cells surrounding NEB exhibit typical structure, al- though their basal regions may contain dense bodies and lamellar bodies characteristic for pneumocytes.
The NE cell possess a large nucleus with deep in- vaginations of the nuclear envelope and characteristic patches of condensed chromatin (Figs. 2, 3). The cyto- plasm contains numerous elongated mitochondria, multivesicular bodies, lysosomes, a centrosome located in the supranuclear region, RER and free ribosomes, numerous actin filaments, and Golgi complexes, mostly grouped in the basal part of the cell (Fig. 4a-c). Nu- merous dense core vesicles (DCV) occur throughout in

NEUROEPITHELIAL BODIES IN THE LUNG OF AMBYSTOMA 42 1
Fig. 1, a: Cross section through the first order septa covered in apical part by ciliated epithelium (black arrow) while the other parts of the septa are lined by respiratory epithelium (open arrow). A, air space; V, blood vessel, haematoxylin, eosin. X 370. b: Serotonin im- munostaining in two NE cells covered by a thin cytoplasmic layer of
surrounding cells. Immunoperoxidase staining. A, air space; ct, con- nective tissue. x 1,100, c: A cluster of met-5-enkephalin immunore- active NE cells close to the air space (A). Immunoperoxidase staining. X 1,100. d Leu-5-enkephalin immunoreactivity in a solitary NE cells. A, air space. Immunoperoxidase staining. x 1,100.
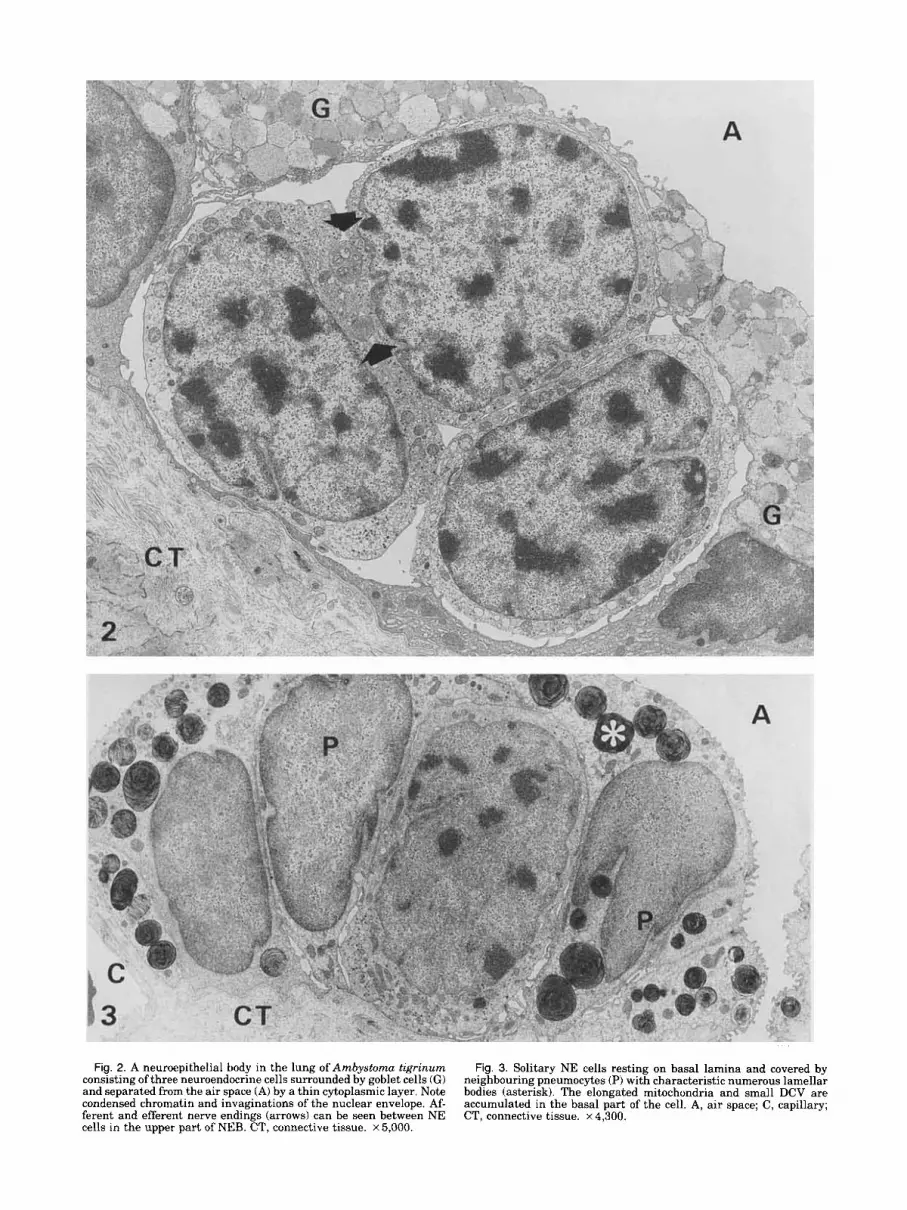
Fig. 2. A neuroepithelial body in the lung of Ambystoma tigrinum consisting of three neuroendocrine cells surrounded by goblet cells (GI and separated from the air space (A) by a thin cytoplasmic layer. Note condensed chromatin and invaginations of the nuclear envelope. Af- ferent and efferent nerve endings (arrows) can be seen between NE cells in the upper part of NEB. CT, connective tissue. ~5 ,000 .
Fig. 3. Solitary NE cells resting on basal lamina and covered by neighbouring pneumocytes (P) with characteristic numerous lamellar bodies (asterisk). The elongated mitochondria and small DCV are accumulated in the basal part of the cell. A, air space; C, capillary; CT, connective tissue. x 4,300.

NEUROEPITHELIAL BODIES IN THE LUNG O F AMBYSTOMA 423
the entire cytoplasm, though their majority may be observed in the basal cell region, adjacent to the un- derlying connective tissue (Figs. 4a-q 5, 6).
NEB and solitary NE cells usually remain in a direct contact with the basal lamina, although they may be separated in some cases from this lamina by a thin cytoplasmic layer belonging to the surrounding cells (Figs. 2,4b,c). NE cells are connected to the neighbour- ing cells and between each other by desmosomes (Fig. 4a) in which long tonofilaments are anchored.
Some NEB contain single cells of another type: larger and approximately 20 pm in length. They are located adjacent to type 1 cells on the one hand and to goblet or ciliated cells on the other, but never inbe- tween type 1 cells (Figs. 4b, 5, 6, 11). Some of type 2 cells (probably the mature ones) are pyramidal in shape and extend from the basal lamina to the air space. Their apical surface is small (approx. 2.0 km in diameter) and equipped with short microvilli and a sin- gle modified cilium a 8 + 1 arrangement of the axone- ma1 microtubules. The apical and subapical cytoplasm contains several small (50-70 nm in diameter) electron lucent vesicles and very rare dense core granules (Figs. 11, 12). The main ultrastructural difference between type 1 and type 2 cells also concerns the morphology of the dense core vesicles (Fig. 4a,c, 5, 6). In type 1 cells, both solitary and corpuscular, vesicles can be divided into several morphological forms, probably represent- ing different stages of maturation (Fig. 6): a) “open” vesicles 180 nm in diameter, with moderately dense content and incomplete limiting membrane, b) vesicles 150 nm in diameter with moderately dense core sur- rounded by a clear space 15-30 nm wide, c) electron dense core vesicles 70-140 nm in diameter, with a lu- cent halo 15-30 nm wide, and d) exceptionally rare large vesicles, 300-600 nm in diameter, with a mod- erately electron dense core surrounded by a lucent space 20 nm in width.
Type 2 cells possess only two vesicle forms (Figs. 4b, 5, 6), i.e., a) dense core vesicles, 140-260 nm in diam- eter, with a clear and 20 nm wide space and b) large vesicles, 320-700 nm in diameter, with a moderately electron dense core surrounded by a lucent halo 20-70 nm wide.
In all NE cells, mature DCV can be observed in close vicinity to the basal cell membrane (Figs. 4b, 5, 6, 7). The solitary NE cells are in a direct contact with the basal lamina only in a relatively small area (Fig. 31, whereas the cell membrane forms several invagina- tions (up to 0.5 pm long) containing DCV in different secretory phases. Coated vesicles can also be observed in this region of the cell (Fig. 7). The solitary NE cells are devoid of any detectable innervation, while the NEB are associated with sensory nerves containing morphologically afferent and efferent intraepithelial terminals (Figs. 2, 8a,b) located between both types of NE cells (Fig. 6), sometimes in the vicinity of the air space (Figs. 2, 12) as well as between NE cells and goblet cells. The main part of the innervation occurs in the basal region of NEB (Figs. 9,121, where apart from nerve endings some nonmyelinated axons can be ob- served in the connective tissue close to the basal lam- ina (Fig. 5). The afferent nerve endings (Fig. 8a) are more numerous and contain abundant mitochondria with long cristae, small vesicles, 40-100 nm in diam-
eter, glycogen particles, and rare concentric lamellated bodies. The efferent nerve endings (Fig. 8b) are char- acterized by the presence of numerous small lucent ves- icles, 40-90 nm in diameter, occasional larger vesicles and relatively scarce dense core vesicles. The synaptic junctions between NE cells and nerve terminals reveal electron dense layers associated with both synaptic membranes, separated by a 20 nm wide cleft, and a synaptic grid on the NE cell side (Fig. 9). Vesicles can be observed on both sides of the junction: electron lu- cent in the nerve endings and with dense core in the NE cell. Serial sections of the intraepithelial nerves reveal that both morphologically afferent and efferent types of endings can be formed by the same nerve fiber which branches a t some distance from the terminals (Fig. 9). The structure of the NEB and their innerva- tion is summarized on Figure 12.
DISCUSSION The NEB located in bronchopulmonary epithelium of
mammals usually communicate with the air space via NE cells extending to the lumen and possessing short microvilli on their surface (Cutz and Connen, 1972; Ericsson et al., 1972; Lauweryns et al., 1972; Hung et al., 1973; Hage et al., 1974; Cutz et al., 1975; Cutz, 1982; Hoyt et al., 1982; Hung, 1982; Sorokin et al., 1982, 1983; Lauweryns and Van Lommel, 1987). The “open” NEB type was also observed in the chicken (Walsh and McLelland, 1974) and turtle (Scheuermann et al., 1983).
As appears from previous studies, most amphibians possess “closed” NEB surrounded and covered by a thick layer of ciliated cells, e.g., in Rana nigroma- culata (Wasano and Yamamoto, 1978) or goblet cells, e.g., in Hynobius nebulosus (Matsumura, 1985). They may also be coated by a thin layer of pneumocytes and/or ciliated cells, as in Hyla arborea, Bornbina uariegata, Bufo bufo, Bufo uiridis, Bombina orientalis (Goniakowska-Witalinska, 1981; Goniakowska-Wita- linska and Cutz, 1990; Goniakowska-Witalinska et al., 1990) and in Ambystoma mexicanum (Scheuermann et al., 1989). The “open” type of amphibian NEB was re- ported exclusively in Bufo rnarinus (Rogers and Haller, 1978, 1980).
There are both “open and closed” NEB in the lung of Ambystoma tigrinum. The predominant morphological type consist of type 1 NE cells covered and separated from the air space by a thin cytoplasmic layer of goblet cells. Some NEB however contain type 2 NE cells which often communicate with the lung lumen, thus accounting for the “open” form of the entire body (Fig. 11).
In all amphibian species investigated so far, the NE cells of NEB tightly adhere to each other. Only in Born- bina uariegata they are separated by the cytoplasmic processes of the surrounding ciliated cells (Gonia- kowska-Witalinska and Cutz, 19901, as also observed in some birds (Cook and King, 1969) and mammals (Sorokin and Hoyt, 1989).
Apart from the NEB, solitary NE cells were de- scribed in the respiratory epithelium of mammals, birds, reptiles, and amphibians (for review see Scheuermann, 1987; Sorokin and Hoyt, 1989). In Urodela, such NE cells are mostly found in the ciliated epithelium, displaying different levels of specialization

Fig. 4. a: Part of type 1 NE cell with two multivesicular bodies (black arrows), elongated mitochondrium, several actin filaments (open arrow), free ribosomes scattering over the cytoplasm, small dense core vesicles, and a part of rough endoplasmic reticulum (arrow- head). Note the desmosome between NE cells and pneumocyte in left lower corner. x 49,000. b: Type 2 NE cells with both types of dense
core vesicles: large with moderately electron dense interior (asterisk) and several smaller ones with electron dense core. The goblet cell (GI, contain lamellar body. L, lysosome. x 31,800. c: Type 1 NE cell with Golgi complex (asterisk), and dense core vesicles in their vicinity. P, pneumocyte. x 48,000.

Fig, 5. Type 2 NE cell (11) located between goblet (G) and type 1 NE cells (I). Two types of DCV are seen. Larger DCV (arrows) with mod- erately electron dense interior are scattered between the smaller DCV with electron dense core. Note an afferent nerve (N) in the connective tissue near the basement membrane (BM). X 11,500.
Fig. 6. Two types of NE cells (I and 11) with afferent nerve endings (N) between them. Note the differences between the DCV in the two types of cells. Type 1 cell (I) contains open vesicles (black arrow), vesicles with moderately electron interior (arrowhead), and dense core vesicles (open arrow). Some DCV in type 2 cells are located near the cell membrane. CT, connective tissue; BM, basal membrane. x 38,000.

426 L. GONIAKOWSKA-WITALINSKA ET AL.
Fig. 7. Basal part of solitary NE cells with DCV a t different secre- tory stages. Coated vesicles (arrow) are also observed. BM, basement membrane. x 71,500.
Fig. 8. Nerve endings between type 1 NE cells. x 42,000. a: Mor- phologically afferent type with small lucent vesicles and mitochon- dria containing long cristae. b: Morphologically efferent type with numerous small lucent vesicles and some larger ones.
Fig. 9. Basal part of NEB with afferent nerve plexus and several intraepithelial nerve endings of morphologically afferent and efferent type. The cytoplasmic continuity between the upper efferent and lower afferent ending is evident (asterisk). The synapse (arrow) be- tween the cell membrane of type 1 cell and the afferent nerve ending reveals the presence of an electron dense grid on the NE cell side. x 41,000.
Fig. 10. The second order septum. Solitary NE cells separated from the air space (A) by a thin layer of pneumocyte (P) cytoplasm. CT, connective tissue. x 4,200.
Fig. l l . Apical part of type 2 NE cell with microvilli and atypical cilium containing 8 + 1 pattern of axonemal microtubules. Note small electron lucent vesicles (asterisk) and large dense core vesicle (arrow) of 230 nm in diameter. The NE cell is flanked by ciliated (CL) and goblet (G) cell. A, air space. x 38,300.

NEUROEPITHELIAL BODIES IN THE LUNG OF AMBYSTOMA 427
Figs. 9-1 1.

Fig.
12.
Sch
emat
ic r
epre
sent
atio
n of
ope
n ty
pe o
f ne
uroe
pith
elia
l bo
dies
in
the
lung
of
Am
byst
oma
tigri
num
. The
NE
B c
onsi
sts
of N
E c
ells
of t
ype
1 (I
) pos
sess
ing
smal
l den
se c
ore
vesi
cles
, NE
cel
l typ
e 2
(11)
wit
h la
rge
DC
V, a
nd in
trac
orpu
scul
ar n
erve
end
ings
of a
ffer
ent a
nd
effe
rent
typ
e (a
rrow
s). T
he m
ain
inne
rvat
ion
of N
EB
is
loca
lized
in
the
basa
l pa
rt,
whe
re
cyto
plas
mic
con
tinu
ity
betw
een
affe
rent
and
eff
eren
t ner
ve e
ndin
gs i
s se
en (
star
). N
EB
is
inte
rcal
ated
bet
wee
n go
blet
(G
) and
cili
ated
(CL)
cells
. A, a
ir sp
ace;
CT,
conn
ectiv
e tis
sue
wit
h ne
rve
prof
ile o
f af
fere
nt ty
pe.

NEUROEPITHELIAL BODIES IN THE LUNG OF AMBYSTOMA
TABLE 1. Diameter ranges of dense core vesicles from different types of NE cells in the lungs of tailed amvhibians
429
Species Diameter ranges (nm) References Hynobius nebulosus I. 46-69 Matsumura, 1985
Ambystoma mexicanum I. 58 Scheuermann et al., 1989 11. 80-110
11. 73 Salamandra salamandra I. 90-150 and 150-270 Goniakowska-Witalinska, 1982
11. 150-450 Triturus a fpestris I. 69-189 Goniakowska-Witalinska, 1980 Ambystoma tigrinum I. 70-140 and 300-600
11. 140-260 and 320-700
including cells without innervation in Hynobius nebu- losus (Matsumura, 1985), Ambystoma mexicanum (Scheuermann et al., 1989) and A. tigrinum; two types of NE cells in Salamandra salamandra and single cells with efferent innervation in Triturus alpestris (Gonia- kowska-Witalinska, 1980, 1982). Solitary NE cells de- void of innervation were also described in the lungs of an ancient fish Polypterus (Scheuermann and De Groodt-Lasseel, 1982), further studies having shown that these cells communicate with the air space (Zac- cone et al., 1989).
The secretory vesicles (DCV) found in the NE cells of tailed amphibians exhibit a considerable diversity in size (Table 1) and morphology. The occurrence in the same NE cells of open vesicles, as well as vesicles with moderately electron dense interior and dense core ves- icles suggest that these forms may represent different maturation stages of DCV. The DCV size (Table 1) ranges from the smallest observed in H. nebulosus and A. mexicanum to the largest occurring in A. tigrinum. Type 1 NE cells of A. tigrinum contain apart from small DCV, also occasionally large ones, similar to those described in NE cells of such anurans as Hyla, Bufo, and Bombina (Goniakowska-Witalinska, 1981, Goniakowska-Witalinska and Cutz, 1990). Type 2 NE cells of A. tigrinum are quite unusual and hitherto have not been described in the lungs of either amphib- ians or other vertebrates. They contain smaller DCV, comparable to those occurring in type 2 NE cells of S. salamandra, although revealing a more electron dense interior and wider lucent halo. Apart from lungs, type 2 cells in A . tigrinum were also observed in the gill epithelium of neotenic animals and were serotonin, met&enkephalin, and leu-5-enkephalin-positive (Go- niakowska-Witalinska et al., in preparation).
In the basal part of both solitary and NEB-associated NE cells of A. tigrinum, mature DCV are often seen in contact with the cell membrane, suggesting the release of their content to the basement membrane. In the sol- itary NE cells, the basal cell membrane forms invagi- nations which increase the secretory surface and DCV at different stages of secretion were observed in this region. A similar configuration of the basal part of NE cells was described in Polypterus (Zaccone et al., 1989).
In three species, a toad, Bufo marinus (Rogers and Haller, 1980), a turtle, Pseudemis scripta elegans (Scheuermann et al., 1983; Pastor et al., 19871, and Ambystoma tigrinum, some NE cells communicate with the air space via the apical surface equipped with
short microvilli and a single modified cilium. In the turtle, normal NE cell bear on their surface a cilium with 9 + 0 or 8+ 1 microtubules, while in A. tigrinum only type 2 cell communicating with the air space dis- play the presence of a cilium with 8 + 1 axonemal con- figuration. The apical cells in the lung of Bufo marinus are highly specialized and possess in their apical part numerous coated vesicles and a single 8+ 1-type cil- ium. The ultrastructure of these modified cilia suggests a sensory function for the cells bearing them (Sorokin, 1968; Afzelius, 1975). Rogers and Haller (1980) hypoth- esized that the apical cells in B. marinus serve as re- ceptors transducing stimuli to the basal NE cells. A recepto-sensory role was also postulated for NEB in the turtle (Scheuermann et al., 1983). Our results seem to agree with this supposition, although it should be noted that type 2 cells in Ambystoma are situated be- tween goblet and ciliated cells, their modified cilia be- ing embedded in the surface lining layer. Therefore it seems more plausible that these cells function as re- ceptors of local chemical changes as well as sensors monitoring the oxygen and carbon dioxide levels.
The investigations carried out so far point out that in amphibians the nerve endings are located in the basal part of NEB (Rogers and Haller, 1978, 1980; Wasano and Yamamoto, 1978; Goniakowska-Witalinska, 1981; Matsumura, 1985; Scheuermann et al., 1989; Gonia- kowska-Witalinska and Cutz, 1990; Goniakowska- Witalinska et al., 1990). The present study revealed a morphologically atypical innervation of NEB in A . ti- grinum: apart from the plexus of morphologically af- ferent and efferent nerve endings located in the basal portion, some endings of both types occur in the apical region of NEB, as well between NE cells alone as be- tween NE cells and the surrounding goblet cells (Fig. 11). A similar type of afferent innervation was found in rabbit NEB (Lauweryns and Van Lommel, 1986). Fur- ther analysis of serial sections revealed that both mor- phologically afferent and efferent nerve endings form very often a cytoplasmic continuity as previously re- ported by Lauweryns and Van Lommel(1987). The ul- trastructural features of the synaptic junctions associ- ated with NE cells suggest a transmission of stimuli from NE cell to the nerve ending, thus supporting the hypothetical neuroreceptor function of NEB.
Some attention should be also paid to the observa- tions concerning the life cycle of the NE cells in A. tigrinum. Both solitary and NEB-associated NE cells are initially separated from the air space by a cytoplas-

430 L. GONIAKOWSKA-WITALINSKA ET AL.
mic layer of pneumocytes and/or goblet cells. This layer becomes gradually thinner (Fig. 10) up to the point of disruption, whereupon the NE cells are extruding from the epithelium to the lung lumen. This phenomenon has not been so far observed in NE cells of other vertebrates.
The comparison of NE cells in the lungs of five spe- cies of tailed amphibia reveals some correlation be- tween the mode of life of animal and the structure of NEB. In entirely aquatic A. mexicanum exist single or grouped NE cells, but typical NEB are absent (Scheuermann et al., 1989). The semi-aquatic H. neb- ulosus possesses closed NEB and solitary NE cells (Matsumura, 1985). Terrestrial tiger salamander has relatively large oxygen requirement during its activity (Full et al., 1988) and the uptake of oxygen proceeds mainly via the lung (Whitford and Hutchison, 1965). In these animals the structure of open NEB is most com- plicated, with innervation similar to that of the mam- malian NEB.
The occurrence of serotonin was identified in many vertebrates (see Scheuerman, 1987; Sorokin and Hoyt, 1989), including several amphibian species (Rogers and Haller, 1978; Wasano and Yamamoto, 1978; Cutz et al., 1986; Scheuermann et al., 1989; Goniakowska- Witalinska et al., 1990). On the other hand, the pres- ence of enkephalins in NE cells ofAmbystoma tigrinum is also of interest. Met-5-enkephalin was found in mammals (Cutz et al., 1981; Lauweryns and Van Ranst, 1987) and the present study reports the first observation of this peptide in NE cells of amphibia. Among the investigated species of amphibia only sin- gle leu-enkephalin cells were observed in the lungs of Bombina uariegata (Cutz et al., 19861, as well as in the gill NE cells of A. tigrinum (Goniakowska-Witalinska et al., in preparation) and lamprey Lampetra japonica (Zaccone, personal communication). Co-occurrence of met-enkephalin and leu-enkephalin was also found in the NE cells of the gills of some teleost fish species (Zaccone et al., 1992). The present results reveal that single leu-enkephalin positive cells in the lung of A. tigrinum may correspond to type 2 NE cells, although it requires further immunocytochemical studies in EM.
In the course of ontogeny the air tract originates from the alimentary canal and the neuroendocrine cells occurring in the lungs are a part of gastroenteric paraneurons (Fujita et al., 1988). The role of NEB in the respiratory epithelium of vertebrates has not been fully elucidated so far, but the recent observations, es- pecially the occurrence of type 2 NE cell, suggest that in Ambystoma tigrinum NEB may serve as receptors sensitive to local chemical changes and are modulated by the central nervous system. They probably also have endocrine andlor paracrine functions.
ACKNOWLEDGMENTS We are grateful to R. Renward, K. Armee for tech-
nical and A. Van Dormael for photographical assis- tance. This investigation was supported by a grant number 93/90 UJ, Poland and a grant from M.U.R.S.T. Rome.
LITERATURE CITED Afzelius, B.A. 1975 Ultrastructure of cilia and flagella. In: Handbook
of Molecular Cytology. A. Lima-de-Faria, ed. North Holland Publ., Amsterdam, pp. 1219-1242.
Cook, R.D., and AS. King 1969 A neurite-receptor complex in the avian lung: Electron microscopical observations. Experientia, 25: 1162-1164.
Coons, A.H., E.H. Leduc, and J.M. Conelly 1955 Studies on antibody production. I. A method for the histochemical demonstration of specific antibody and its application to a study of hiperimmune rabbit. J . Exp. Med., 102:49-59.
Cutz, E., and P.E. Conen 1972 Endocrine-like cells in human fetal lungs: An electron microscopic study. Anat. Rec., 173:115-122.
Cutz, E., W. Chan, V. Wong, and P.E. Conen 1975 Ultrastructure and fluorescence histochemistry of endocrine (APUD-type) cells in tracheal mucosa of human and various animal species. Cell Tis- sue Res., 158:425-437.
Cutz, E., W. Chan, and N.S. Track 1981 Bombesin, calcitonin and leu-enkephalin immunoreactivity in endocrine cells of human lung. Experientia, 37:765-767.
Cutz, E. 1982 Neuroendocrine cells of the lung: An overview of mor- phologic characteristics and development. Exp. Lung Res., 3: 185-208.
Cutz, E., L. Goniakowska-Witalinska, and W. Chan 1986 An immu- nohistochemical study of regulatory peptides in lungs of amphib- ians. Cell Tissue Res., 244:227-233.
Ericsson, L.E., R. HakAnson, B. Larcon, C. Owman, and E. Sundler 1972 Fluorescence and electron microscopy of amine-storing en- terochromaffn-like cells in tracheal epithelium of mouse. Z. Zell- forsch., 124532-545,
Frohlich, F. 1949 Die “Helle Zelle” der Bronchialschleimhaut und ihre Beziehungen zum Problem der Chemoreceptoren. Frankf. 2. Pathol., 60517-559.
Fujita, T., T. Kanno, and S. Kobayashi 1988 Bronchopulmonary para- neuron. In: The Paraneuron. Springer Verlag, Tokyo, pp. 190- 198.
Full, R.J., B.D. Anderson, C.M. Finnerty, and M.E. Feder 1988 Exer- cising with and without lungs. I. The effects of metabolic cost maximal oxygen transport and body size on terrestrial locomotion in salamander species. J . Exp. Biol., 138:471-485.
Goniakowska-Witalinska, L. 1980 Endocrine-like cells in the lungs of the newt Triturus alpestris Laur. Cell Tissue Res., 210521-524.
Goniakowska-Witalinska, L. 1981 Neuroepithelial bodies in the lung of the tree frog Hyla arborea L. Cell Tissue Res., 21 7:435-441.
Goniakowska-Witalinska, L. 1982 Development of the larval lung of Salamandra salamandra L., A Scanning and transmission elec- tron microscopic study. Anat. Embryol., 164:113-137.
Goniakowska-Witalinska, L., and E. Cutz 1990 Ultrastructure of neu- roendocrine cells in the lungs of three anuran species. J . Mor-
Goniakowska-Witalinska, L., J.M. Lauweryns, and L. Van Ranst 1990 Neuroepithelial bodies in the lungs of Bombina orientalis (Boul.). In: Chemoreceptors and Chemoreceptor Reflexes. H. Acker, ed. Plenum Press, New York, pp. 111-117.
Goniakowska-Witalihska, L., G. Zaccone, and S. Fasulo 1992 Neu- roepithelial cells in the gills of the neotenic tiger salamander Ambystoma tigrinum (in preparation).
Hage, E. 1974 Histochemistry and fine structure of endocrine cells in foetal lungs of the rabbit, mouse and guinea pig. Cell Tissue Res., 149513-524.
Hoyt, R.F., S.P. Sorokin, and H. Feldman 1982 Small-granule (Neuro) endocrine cells in the infracardial lobe of hamster lung. Number, subtypes and distribution. Exp. Lung Res., 3~273-298.
Hung, K-S. 1982 Development of neuroepithelial bodies in pre- and postnatal mouse lung: Scanning electron microscopic study. Anat. Rec., 203:285-291.
Hung, K-S., M.S. Hertweck, J.D. Hardy, and C.G. Loosli 1973 Ultra- structure of nerves and associated cells in bronchiolar epithelium of mouse lung. J . Ultrastruct. Res., 43:426-437.
Lauweryns, J.M., M. Cokelaere, and P. Theunynck 1972 Neuroepi- thelial bodies in the respiratory mucosa of various mammals. 2. Zellforsch., 135569-592.
Lauweryns, J.M., and J.C. Peuskens 1969 Argyrophil (storing and amine producing?) cells in human infant airway epithelium. Life Sci., 8577-585.
Lauweryns, J.M., and J.C. Peuskens 1972 Neuroepithelial bodies (neuroreceptor or secretory organs?) in human infant bronchial and bronchiolar epithelium. Anat. Rec., 172:471-482.
Lauweryns, J.M., and A. Van Lommel 1986 Effect of various vagot- omy procedures in the reaction to hypoxia of rabbit neuroepithe- lial bodies: Modulation by intrapulmonary axon reflexes? Exp. Lung Res., llr319-339.
Lauweryns, J.M., and A. Van Lommel 1987 Ultrastructure of nerve endings and synaptic junctions in rabbit intrapulmonary neu-
phol., 203:l-9.

NEUROEPITHELIAL BODIES IN THE LUNG OF AMBYSTOMA 431
roepithelial bodies: A single and serial sections analysis. J. Anat., 151 :65-83.
Lauweryns, J.M., and L. Van Ranst 1987 Leu-7 immunoreactivity in human, monkey and pig bronchopulmonary neuroepithelial bod- ies and neuroendocrine cells. J . Histochem. Cytochem., 35t687- 690.
Matsumura, H. 1985 Electron microscopic studies of the lung of the salamander, Hynobius nebulosus. 111. A scanning and transmis- sion electron microscopic observations on neuroepithelial bodies. Okajimas Folia Anat. Jpn., 62:187-204.
Pastor, L.M., J . Ballesta, M.T. Castells, R. Perez-Tomas, J.A. Marin, and J.F. Madrid 1989 A microscopic study of the lung of Testudo graeca (Chelonia). J . Anat., 164:19-39.
Pastor, L.M., J. Ballesta, F. Hernandez, R. Perez-Tomas, A. Zuasti, and C. Ferrer 1987 A microscopic study of the tracheal epithe- lium of Testudo graeca and Pseudemis scripta elegans. J. Anat., 153:171-183.
Rogers, D.C., and C.J. Haller 1978 Innervation and cytochemistry of neuroepithelial bodies in the ciliated epithelium of the toad lung (Bufo marinus). Cell Tissue Res., 195:395-410.
Rogers, D.C., and C.J. Haller 1980 The ultrastructural characteristics of the apical cell in the neuroepithelial bodies of the toad lung (Bufo marinus). Cell Tissue Res., 209:485-498.
Scheuermann, D.W., M.H.A. De Groodt-Lasseel, C. Stilman, and M.L. Meisters 1983 A correlative light-, fluorescence and electron mi- croscopic study of neuroepithelial bodies in the lung of the red- eared turtle, Pseudemis scripta elegans. Cell Tissue Res., 234: 249-269.
Scheuermann, D.W. 1987 Morphology and cytochemistry of the endo- crine epithelial system in the lung. Internat. Rev. Cytol., 106: 35-88.
Scheuermann, D.W., D. Andriaensen, J.P. Timmermans, and M.H.A. De Groodt-Lasseel 1989 Neuroepithelial endocrine cells in the lung of Ambystoma mexicanum. Anat. Rec., 225:139-149.
Scheuermann, D.W., and M.H.A. De Groodt-Lasseel 1982 Monoam-
ine-containing granulated cells in the Polypterus lung. Verh. Anat. Ges., 76:301-302.
Sorokin, S.P. 1968 Reconstruction of centriole-formation on ciliogen- esis in mammalian lungs. J. Cell Sci., 3:207-230.
Sorokin, S.P., and R.F. Hoyt 1989 Neuroepithelial bodies and solitary small-granule cells. In: Lung Cell Biology. D. Massaro ed. M. Dekker Inc., New York, Basel, pp. 191-344.
Sorokin, S.P., R.F. Hoyt, and M. Grant 1982 Development of neuroep- ithelial bodies in fetal rabbit lungs. I. Appearance and functional maturation as demonstrated by high-resolution light microscopy and formaldehyde induced fluorescence. Exp. Lung Res., 3237- 259.
Sorokin, S.P., R.F. Hoyt, and A.D. Persall 1983 Comparative biology of small granule cells and neuroepithelial bodies in the respira- tory system: Short review. Am. Rev. Respir. Dis., 128:S26S31.
Sternberger, L.A. 1979 Immunocytochemistry. 2nd ED., J . Wiley and Sons, New York.
Walsh, C., and J. McLelland 1974 Granular “endocrine” cells in avian respiratory epithelia. Cell Tissue Res., 153269-276.
Wasano, K., and T. Yamamoto 1976 Granule-containing cells in the snake respiratory mucosa. Acta Anat. Nippon, 51 :299.
Wasano, K., and T. Yamamoto 1978 Monoamine-containing granu- lated cells in the frog lung. Cell Tissue Res., 193:201-209.
Wasano, K., and T. Yamamoto 1979 APUD-type recepto-secretory cells in the chicken lung. Cell Tissue Res., 201:197-205.
Whitford, W.G., and V.H. Hutchison 1965 Gas exchange in sala- manders. Physiol. Zool., 38228-242.
Zaccone, G., L. Goniakowska-Witalinska, J.M. Lauweryns, M. Fasulo, and G. Tagliafierro 1989 Fine structure and serotonin immuno- histochemistry of the neuroendocrine cells in the lungs of the bichirs Polypterus delhezi and P. ornatipinnis. Bas. Appl. His- tochem., 33:277-287.
Zaccone, G., L.M. Lauweryns, S. Fasulo, G. Tagliafierro, L. Ainis, and L. Licata 1992 Immunocytochemical localization of serotonin and neuropeptides in the neuroepithelial endocrine cells of teleost and lungfish gills. Acta Zool. Stockh., in press.