ultrastructure and gaba immunoreactivity in layers 8 and 9 of the optic tectum of xenopus laevis
TRANSCRIPT

European Journal of Neuroscience, Vol, 6, pp. 1567-1582, 1994 0 European Neuroscience Association
Ultrastructure and GABA lmmunoreactivity in Layers 8 and 9 of the Optic Tectum of Xenopus laevis
Krystyna Kielan Rybicka and Susan B. Udin Department of Physiology, State University of New York, Buffalo, NY 14214, USA
Key words: frogs, binocular vision, retinotectal projection, isthmotectal projection
Abstract
This study presents an ultrastructural analysis of layers 8 and 9 in the optic tectum of Xenopus laevis. Retinotectal axons were labelled with horseradish peroxidase and tectal cells were labelled with antibody to GABA. Four distinct axonal and dendritic structures were identified. GABA-negative axon terminals formed asymmetric synapses and were categorized as type a-1 (which included retinotectal axons), characterized by medium size synaptic vesicles and pale mitochondria, and type a-2 (non-retinotectal) with large vesicles and dense mitochondria. GABA-negative dendrites (type d) contained dense mitochondria, microtubules in the dendritic shafts, and dendritic spines devoid of microtubules. GABA-positive structures contained small synaptic vesicles and dense mitochondria. Some dendrites (type D) were not only postsynaptic but were also presynaptic elements, as defined by the presence of vesicles and distinct synaptic clefts with symmetric specializations. GABA-positive presynaptic structures were mostly located in vesicle-filled, bulbous extensions of dendritic shafts and usually terminated onto dendritic spines. Some type D dendrites were the middle element in serial synapses, with input from either GABA-positive or GABA-negative structures and output to GABA-negative structures. Retinotectal terminals were identified as one of the synaptic inputs to GABA-positive .processes. Glia were characterized by granular cytoplasm and large mitochondria, often displaying a crystalline matrix structure. These results indicate that GABA-positive neurons are a prominent component of circuitry in the superficial layers of the tectum of Xenopus and that, as in mammals, they participate in serial synaptic arrangements in which retinotectal axons are the first element. These arrangements are consistent with complex processing of visual input to the tectum and a central role for inhibitory processes in the shaping of tectal responses.
Introduction
This study is a part of ongoing investigations to determine the mechanisms underlying the activity-dependent matching of maps from the two eyes to the superficial layers of the optic tectum of Xenopus laevis. Some evidence about the ultrastucture of the frog tectum is available from studies on Rana frogs but a comparison and extension of those data to the somewhat different tectum of Xenopus is necessary. Four types of nerve terminals in the frog optic tectum were identified by ultrastuctural study with enucleation for identification of retinotectal axons in Rana esculenta (SzBkely et al., 1973) and confirmed by the use of horseradish peroxidase (HRP) labelling in R. pipiens (Hughes, 1990). In addition, antibodies to GABA have been used to characterize putative inhibitory neurons in Rana (Antal, 1991). In this study, we have used HRP filling of optic axons and antibodies to GABA to establish a better understanding of synaptic circuitry in Xenopus tectum, particularly with regard to identifying neuronal processes that may participate in the circuitry of binocular convergence.
Materials and methods
Labelling of retinotectal axons with HRP Four adult Xenopus laevis frogs were anaesthetized by injection of 0.04-0.06 ml of 2% tricaine methanesulphonate (ethyl-m-amino- benzoate; Sigma). The skin and connective tissue above the eye were cut to gain access to the optic nerve, which was transected behind the eye. A crystal of HRP (Type VI; Sigma) was applied to the site of the transection. The skin was sutured and the frog allowed to survive for 3 days.
Fixation Frogs were anaesthetized as described above. The skull was opened and the brain was removed and immersed in 2.5-5% glutaraldehyde and 0.8-2% paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4. Material was usually fixed for 3 h and washed in PB. Samples were embedded in gelatin-albumin and sections were cut on a vibratome in the parasagittal plane.
Correspondence to: Dr S . B. Udin, as above
Received I9 July, 1993, revised 31 January, 1994, accepted I1 May, 1994

1568 Xenopus tectal ultrastructure
HRP reaction Free-floating sections from the specimens injected with HRP were incubated with diaminobenzidine, cobaltous chloride and nickel ammonium sulphate (Sigma) according to the procedure of Adams (1981).
Anti-GA BA staining procedure for light microscopy Vibratome sections from four frogs were rinsed in Tris-buffered saline (TBS) [50 mM Tris, pH 7.5, with I % bovine serum albumin (BSA) and 0.9 NaCI] (3 X 5 min), incubated in 10% normal goat serum (NGS) with 0.9% BSA and 1% glycine in TBS/O.I% saponin for 1 h, rinsed for 10 min in TBS/O.I% saponin, incubated overnight in GABA antiserum (Incstar, USA) (1 : lOOO to 1:5000) in TBS/O.I% saponin with 1% NGS and 0.1% NaN3, rinsed in 1% NGS in TBS/ 0.1% saponin (3 X 5 min), incubated in biotinylated goat-anti rabbit IgG (1 drop in 10 ml) in 1% NGS in TBS/O.l% saponin (2 h) and reacted with the Vectastain ABC (Vector Laboratories) kit using diaminobenzidine (10 min).
General electron microscopy procedures Vibratome sections from 12 adult X.laevis frogs were postfixed in 1% Os04 in PB, usually processed with uranyl acetate en bloc, dehydrated with graded ethanols and acetone, and flat-embedded in either Spurr’s epoxy resin or Durcupan ACM. After polymerization, sections were photographed and re-embedded on inverted BEEM capsules filled with Epon-Araldite. Regions of interest were cut on an ultramicrotome and silver or gold sections were collected on copper grids for routine uranyl-lead staining and on nickel grids for immunolabelling.
Anti-GABA immunogold staining Two anti-GABA immunogold staining procedures were used. All reagents and washing solutions were stored in sterile bottles and filtered (Millipore size 0.22 pm) before use.
Anti-GABA irnmunogold procedure I (Somogyi and Soltkz, 1986) Material was embedded in Durcupan ACM. Sections were etched in 1% periodic acid (7-10 min), rinsed four times in double-distilled water, treated with freshly prepared sodium metaperiodate (7-10 rnin), and rinsed as above. Further processing with grids floating on drops of solution included: (i) Rinsing in TPBS (10 mM Tris-HCI, 10 mM PB, 0.7% NaCI, pH 7.6) (2 X 10 min). (ii) Incubation in 5% NGS in TPBS (15 min). (iii) Rinsing in 1 % NGS in TPBS (1-2 min). (iv) Incubation in polyclonal rabbit GABA antiserum (Incstar) diluted 1500 in 1% NGS in TPBS (2 h). (v) Rinsing in 1% NGS in TPBS (3 X 15 min). (vi) Incubation in goat- anti-rabbit IgG-colloidal gold (15 nm) diluted 1 : 10 in 0.1 M Tris, pH 7.4, 21% BSA, and 0.1% Tween 20 (2 h). Grids were dipped three times in double-distilled water and rinsed in double-distilled water for 10 min, dried and poststained with I % uranyl acetate in 50% ethanol and with lead citrate. This method produced only sparse labelling in our material. We therefore used the following method, which produced substantial deposits of gold particles, for our sub- sequent studies.
Anti-GABA immunogold procedure 2 (Phend et al., 1992) Material was embedded either in Durcupan ACM or in Spurr’s resin. Grids were covered using the Coat-Quick ‘G’ pen (Electron Microscopy Sciences, USA). Sections were processed by immersion of grids in droplets of solutions. The processing of grids involved:
(i) Immersion in 0.05 M Tris-HCI buffered saline-Triton X-100 (TBST), pH 7.6 (2 X 10 min). (ii) Incubation in polyclonal rabbit GABA antiserum (either Incstar or Arnell, USA) diluted 1500 in TBST in a moist chamber at room temperature (overnight). (iii) Rinsing in TBST, pH 7.6 (2 X 5 min). (iv) Incubation in TBST, pH 7.6 (30 min). ( v ) Incubation in TBST, pH 8.2 (5 rnin). (vi) Incubation in IgG-colloidal gold (15 nm) in TBST pH 8.2 (1:25) ( I h). (vii) Transfer to double-distilled water and rinsing in three beakers (30 times each). (viii) Drying and staining in 3% uranyl acetate in SO% ethanol, followed by lead citrate.
Controls were performed by the omission of GABA antibody and the incubation of grids in the respective buffer used as diluent, and by preincubation of GABA antibody with GABA/BSA conjugate with GABA concentration 500 pg/ml, kindly provided by Incstar. Mixtures of 40 and 80 p1 of conjugate per ml of diluted GABA antiserum were left overnight at 4°C and then were used for reaction and control in procedure 2.
Results
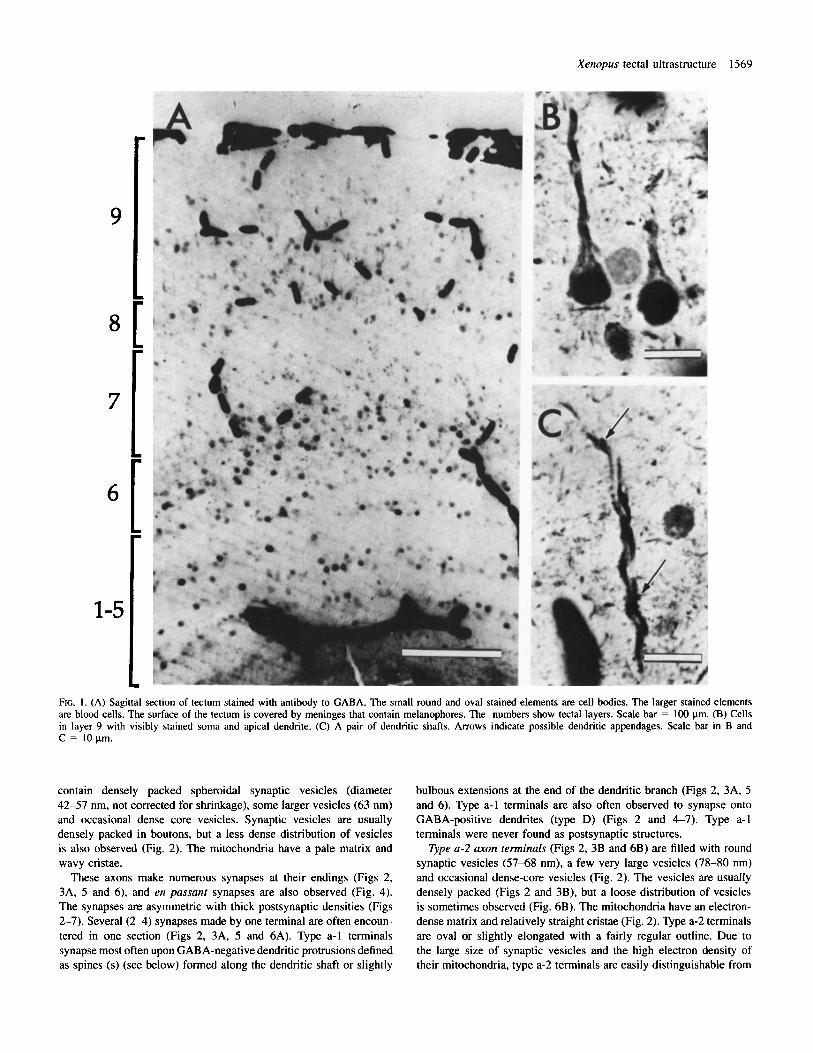
General structure of the superficial layers of the optic tectum Xenopus tectum is typical of anuran tecta in having alternating cell- dense and cell-sparse layers. These layers are numbered from 1 (the ependymoglial layer adjacent to the ventricle) to 9 (the superficial neuropil underlying the pia) (Fig. I ) . The focus of this study is layer 9, which contains the retinotectal and isthmotectal axons that relay input from the two eyes (Udin and Keating, 1981). Other axonal components include input from the ipsilateral nucleus isthmi (Udin and Keating, 1981) and a possible projection from the thalamus (Trachtenberg and Ingle, 1974). Some cell bodies reside within layer 9, including projection neurons with output to the nucleus isthmi (S. B. Udin, unpublished observations). Other cells include glia and neurons of unknown connectivity, possibly interneurons. Layer 9 also includes apical dendrites of cells in deeper layers, plus processes of ependymoglial cells (SzCkely and Lizk , 1976).
Layer 8 comprises sparsely distributed neurons interspersed with apical dendrites and glial processes arising from deeper layers. In addition some retinotectal and isthmotectal axons extend into layer 8 (Udin and Fisher, 1985).
Distribution of GABA-like immunoreactivity: light microscopy Sagittal sections of the tectum in which GABA-like immunoreactivity was visualized with a peroxidase stain reveal stained cell bodies in all cellular layers of the tectum (Fig. IA). Layer 9, the focus of our study, contains scattered cell bodies, most of which are stained, and a plexus of GABA-positive processes (Fig. 1 B, C). Radially-oriented processes can be identified in favourable sections as apical dendrites. Individual processes vary in darkness and diameter and occasionally display very short appendages (Fig. IC).
Ultrastructure GABA-negative neuronal structures In this paper, GABA-negative neuronal structures are labelled with small letters and GABA-positive with capital letters.
Type 0-1 axon terminals (Figs 2-7) are distributed throughout layers 8 and 9, in which they comprise more than SO% of the presynaptic structures, as judged from counts of synapses on a montage of 20 micrographs (20 000-24 OOOX) of layers 9 and 8. Anterograde labelling of retinotectal axons demonstrates that many of these axons originate in the retina (Figs 5, 6B and 7A). They
Xenopus tectal ultrastructure 1569
9
8
7
6
1-5
FIG. I . (A) Sagittal section of tectum stained with antibody to GABA. The small round and oval stained elements are cell bodies. The larger stained elements are blood cells. The surface of the tectum is covered by meninges that contain melanophores. The numbers show tectal layers. Scale bar = 100 pm. (B) Cells in layer 9 with visibly stained soma and apical dendrite. (C) A pair of dendritic shafts. Arrows indicate possible dendritic appendages. Scale bar in B and C = 10ym.
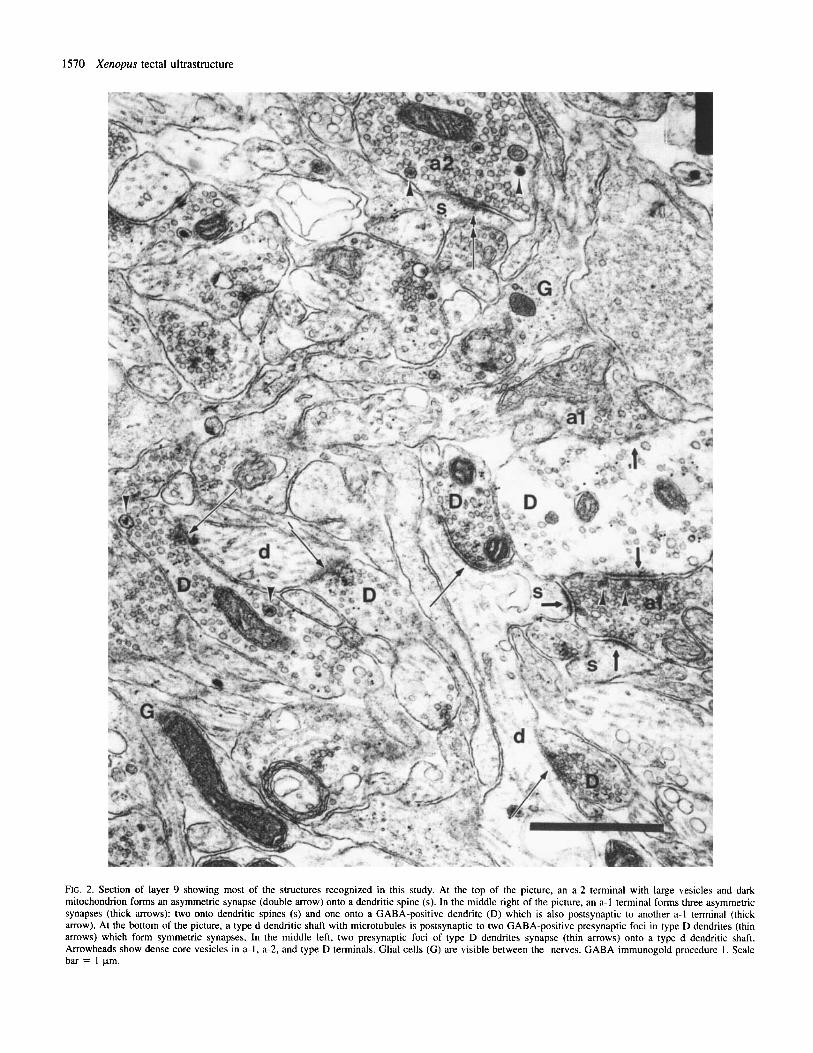
contain densely packed spheroidal synaptic vesicles (diameter 42-57 nm, not corrected for shrinkage), some larger vesicles (63 nm) and occasional dense core vesicles. Synaptic vesicles are usually densely packed in boutons, but a less dense distribution of vesicles is also observed (Fig. 2). The mitochondria have a pale matrix and wavy cristae.
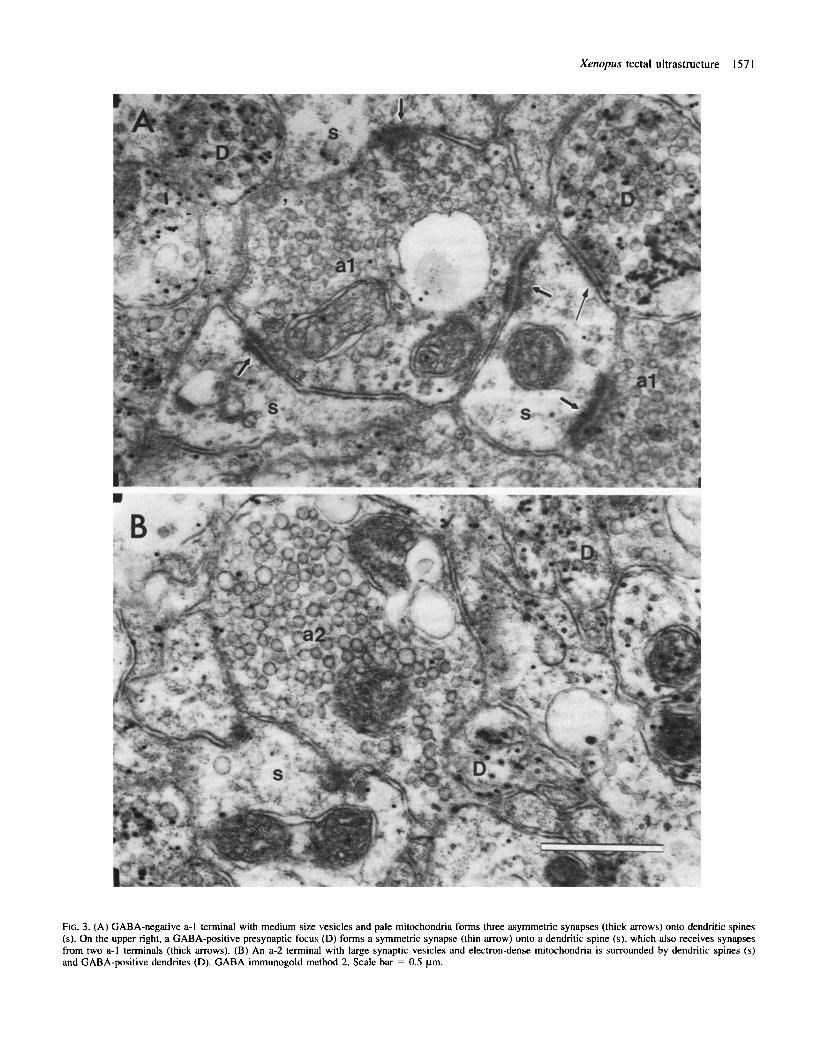
These axons make numerous synapses at their endings (Figs 2, 3A, 5 and 6), and en passant synapses are also observed (Fig. 4). The synapses are asymmetric with thick postsynaptic densities (Figs 2-7). Several ( 2 4 ) synapses made by one terminal are often encoun- tered in one section (Figs 2, 3A, 5 and 6A). Type a-I terminals synapse most often upon GABA-negative dendritic protrusions defined as spines (s) (see below) formed along the dendritic shaft or slightly
bulbous extensions at the end of the dendritic branch (Figs 2, 3A, 5 and 6). Type a-I terminals are also often observed to synapse onto GABA-positive dendrites (type D) (Figs 2 and 4-7). Type a-I terminals were never found as postsynaptic structures.
Type a-2 axon terminals (Figs 2, 3B and 6B) are filled with round synaptic vesicles ( 5 7 4 8 nm), a few very large vesicles (78-80 nm) and occasional dense-core vesicles (Fig. 2). The vesicles are usually densely packed (Figs 2 and 3B), but a loose distribution of vesicles is sometimes observed (Fig. 6B). The mitochondria have an electron- dense matrix and relatively straight cristae (Fig. 2). Type a-2 terminals are oval or slightly elongated with a fairly regular outline. Due to the large size of synaptic vesicles and the high electron density of their mitochondria, type a-2 terminals are easily distinguishable from
1570 Xenopus tectal ultrastructure
FIG. 2. Section of layer 9 showing most of the structures recognized in this study. At the top of the picture, an a-2 terminal with large vesicles and dark mitochondrion forms an asymmetric synapse (double arrow) onto a dendritic spine (s). In the middle right of the picture, an a-I terminal forms three asymmetric synapses (thick mows): two onto dendritic spines (s) and one onto a GABA-positive dendrite (D) which is also postsynaptic to another a-1 terminal (thick arrow). At the bottom of the picture, a type d dendritic shaft with microtubules is postsynaptic to two GABA-positive presynaptic foci in type D dendrites (thin mows) which form symmetric synapses. In the middle left, two presynaptic foci of type D dendrites synapse (thin arrows) onto a type d dendritic shaft. Arrowheads show dense core vesicles in a-I, a-2, and type D terminals. Glial cells ( G ) are visible between the nerves. GABA immunogold procedure 1 . Scale bar = 1 pm.
Xenopus tectal ultrastructure 157 1
FIG. 3. (A) GABA-negative a-I terminal with medium size vesicles and pale mitochondria forms three asymmetric synapses (thick arrows) onto dendritic spines (s). On the upper right, a GABA-positive presynaptic focus (D) forms a symmetric synapse (thin m o w ) onto a dendritic spine ( s ) , which also receives synapses from two a-I terminals (thick arrows). (B) An a-2 terminal with large synaptic vesicles and electron-dense mitochondria is surrounded by dendritic spines (s) and GABA-positive dendrites (D). GABA immunogold method 2. Scale bar = 0.5 pm.

1572 Xenopus tectal ultrastructure
type a-I (Figs 2 and 3). They are distributed throughout layers 8 and 9, being most common in the upper part of layer 9.
Type a-2 axons form asymmetric synapses (Figs 2 and 6B) similar to those made by type a-1 axons but they seem to make fewer synaptic contacts, as indicated by the fact that most a-2 terminals participate in only a single synapse in one section. Their synaptic targets are primarily GABA-negative dendritic spines (defined below), with fewer contacts with GABA-negative dendritic shafts and GABA- positive (type D) dendrites. Type a-2 axons were never found as postsynaptic elements.
Typed dendrites (Figs 2-7) are elongated, often branched, structures with relatively pale cytoplasm and numerous microtubules. Their mitochondria are usually dark with straight, often parallel, cristae, although less contrasted mitochondria with tubular cristae are also observed (Fig. 6A, inset). They bear dendritic protrusions along the dendritic shaft and slightly bulbous extensions at the end of dendritic branches (Figs 2 ,4 and 5) . These protrusions fit several of the criteria for spines presented by Peters et al. (1991); in particular, they contain irregular fluffy material, occasional mitochondria and no microtubules. On the other hand, they do not appear to contain the spine apparatus nor are spine necks commonly observed in this tissue. We refer to such profiles as spines despite their incomplete conformity with the classical description of spines. These spines are oval or elongated in cross-sections and are often arrayed in a group surrounding nerve terminals (Figs 2 and 3). They represent the most common postsynaptic element for all types of terminals.
In most sections, a type d dendrite is seen to receive only one synaptic input, but -10% of type d dendrites show two or three synapses in one section. When multiple synaptic inputs are observed, the presynaptic elements generally are of different classes, but no convergence of a-I and a-2 terminals onto the same profile was seen. Three synapses onto a type d dendrite are shown in Figure 2 (lower right) where an a-1 terminal synapses onto dendritic spine and two presynaptic foci of D dendrites form synapses onto the dendritic shaft. Figure 3A shows a dendritic spine postsynaptic to two a-I terminals and to one GABA-positive process.
GABA-negative cell bodies with dendritic extensions are observed in layer 6. They contain spheroid nuclei with electron-dense chromatin and a thin rim of the cytoplasm containing polyribosomes, rough endoplasmic reticulum, microtubules, and Golgi complexes.
GABA-positive neumnal structures Two immunogold procedures were employed (Somogyi and SoltCsz, 1986; Phend et al., 1992), and they yielded differential staining of the tectal neuropil, the method of Phend (Figs 3 and 5-8) producing more intense immunoreactivity than that of Somogyi and SoltCsz (Figs 2 and 4). A dense accumulation of gold particles appears in elements, which are thus defined as GABA-positive, while other structures are overlaid by very few gold particles and are therefore defined as GABA-negative structures. The immunogold particles are primarily associated with synaptic vesicles and mitochondria. All control sections with preabsorption of antibody by GABA/BSA or with omission of primary antibody produced entirely negative results.
All GABA-positive structures contain synaptic vesicles. In many cases, these structures can be identified as ‘vesicle bearing dendrites’ by virtue of the fact that single profiles contain both presynaptic and postsynaptic areas. We refer to presynaptic regions in dendrites as ‘presynaptic foci’.
Type D vesicle-bearing dendrites (Figs 2-7) contain microtubules, endoplasmic reticulum and occasional ribosomes. Small synaptic vesicles appear in dendritic shafts that contain microtubules (Figs 4, 6A and 7B,C). Similar vesicles are observed in microtubule-free
FIG. 4. ’ h o a-1 axons with en passanr asymmetrical synapses (thick arrows) onto type D dendrites. The upper dendrite (D) forms a symmetrical synapse (thin arrow) onto an invagination by a dendritic spine. Another dendritic spine (s) is visible in the lower centre. Arrowhead shows dense core vesicle. GABA immunogold method 1. Scale bar = 0.5 pm.
extensions of dendritic shafts (Figs 2, 3 and 5-7). The extensions often appear as terminal bulbs. Synaptic vesicles in the dendritic bulbs are generally round and small (40-50 nm) but a very few elongated vesicles are also present (Fig. 7C); there are also a few larger vesicles (60-68 nm) and occasional dense core vesicles (Figs 2 and 7A, C). The mitochondria usually have a dense matrix and a few, often parallel cristae (Figs 2 and 6A). Type D dendrites appear through the entire optic tectum, and they are densely distributed in all zones of layers 8 and 9.
Type D dendrites contain presynaptic areas defined as presynaptic foci. They are most common in bulbous extensions (Figs 2, 3 and 7A, thin arrows), but they also appear along the dendritic shafts as small accumulations of presynaptic vesicles (Figs 4 and 6A, B, thin arrows). The vesicles usually accumulate more densely in the bulbous extensions than along the dendritic shafts. The variation in vesicle distribution is distinctly visible on longitudinal sections of dendrites (Fig 2, left). All presynaptic foci form symmetric synapses onto dendrites, predominantly type s dendritic spines and, less often, GABA-negative dendritic shafts and GABA-positive dendrites. No
Xenopus tectal ultrastructure 1573
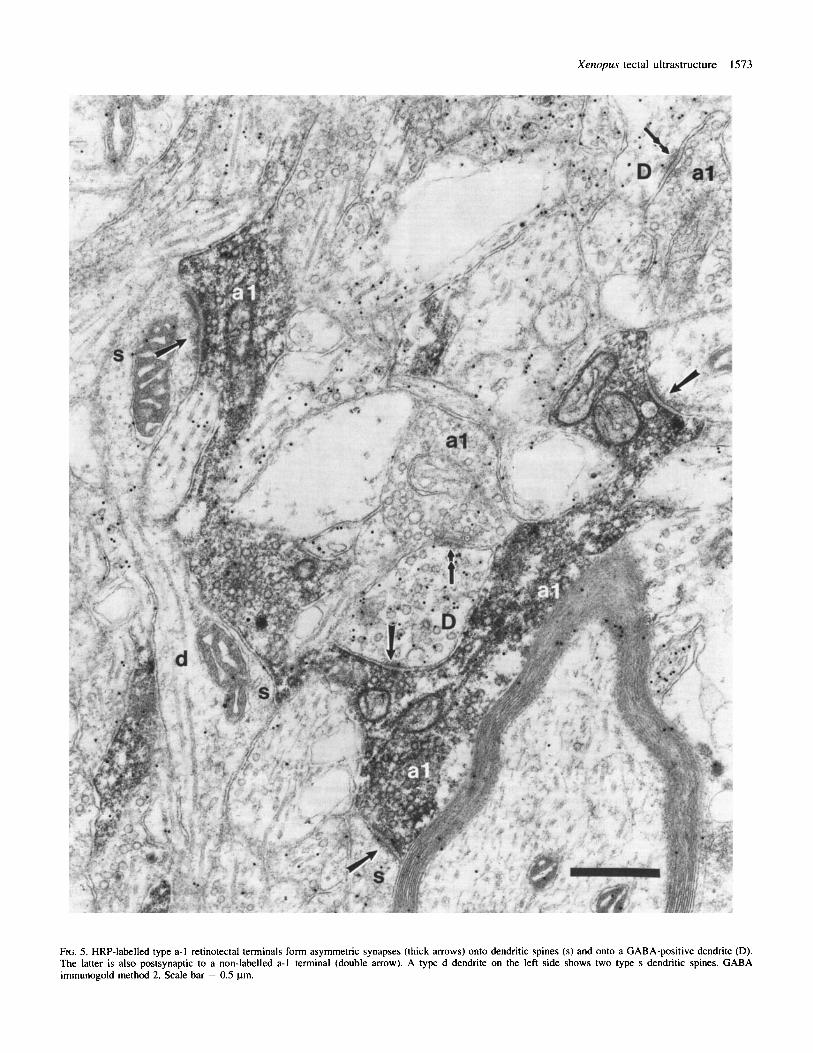
FIG. 5. HRP-labelled type a-1 retinotectal terminals form asymmetric synapses (thick arrows) onto dendritic spines (s) and onto a GABA-positive dendrite (D). The latter is also postsynaptic to a non-labelled a-I terminal (double arrow). A type d dendrite on the left side shows two type s dendritic spines. GABA immunogold method 2. Scale bar = 0.5 pm.
1574 Xenopus tectal ultrastructure
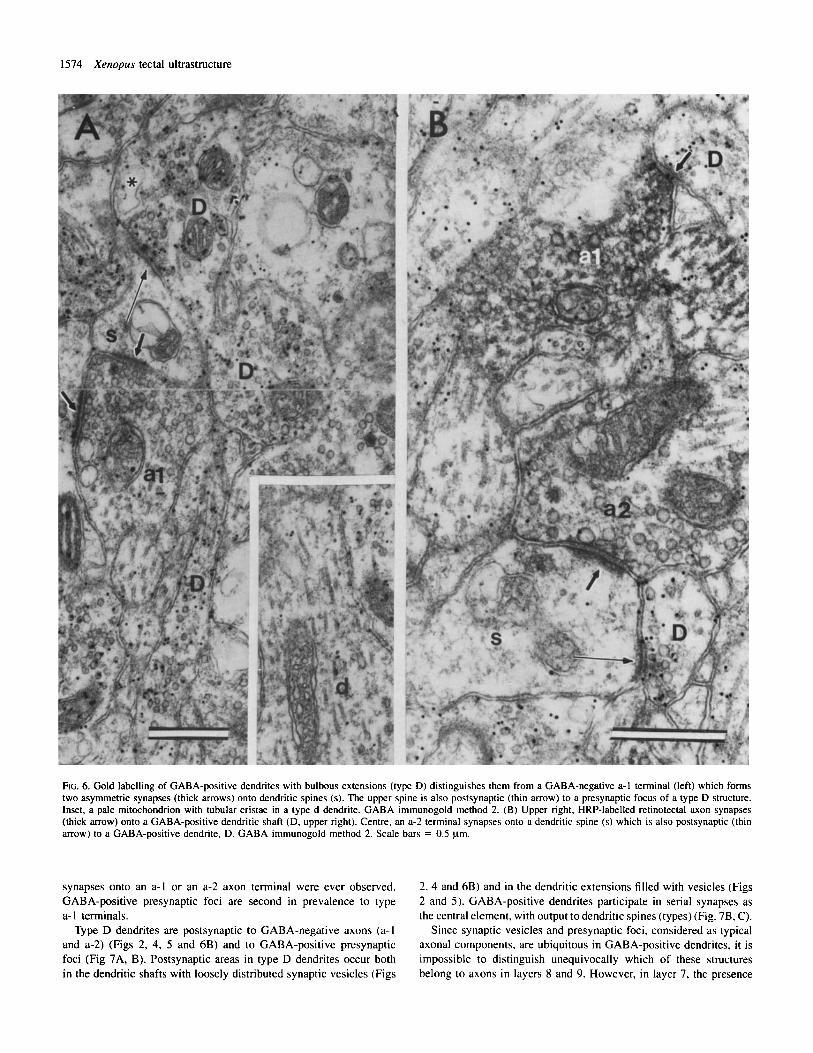
FIG. 6. Gold labelling of GABA-positive dendrites with bulbous extensions (type D) distinguishes them from a GABA-negative a-1 terminal (left) which forms two asymmetric synapses (thick arrows) onto dendritic spines (s). The upper spine is also postsynaptic (thin arrow) to a presynaptic focus of a type D structure. Inset, a pale mitochondrion with tubular cristae in a type d dendrite. GABA immunogold method 2. (B) Upper right, HRP-labelled retinotectal axon synapses (thick arrow) onto a GABA-positive dendritic shaft (D, upper right). Centre, an a-2 terminal synapses onto a dendritic spine (s) which is also postsynaptic (thin arrow) to a GABA-positive dendrite, D. GABA irnmunogold method 2. Scale bars = 0.5 prn.
synapses onto an a-I or an a-2 axon terminal were ever observed. GABA-positive presynaptic foci are second in prevalence to type a- 1 terminals.
Type D dendrites are postsynaptic to GABA-negative axons (a- I and a-2) (Figs 2, 4, 5 and 6B) and to GABA-positive presynaptic foci (Fig 7A, B). Postsynaptic areas in type D dendrites occur both in the dendritic shafts with loosely distributed synaptic vesicles (Figs
2, 4 and 6B) and in the dendritic extensions filled with vesicles (Figs 2 and 5). GABA-positive dendrites participate in serial synapses as the central element, with output to dendritic spines (types) (Fig. 7B, C).
Since synaptic vesicles and presynaptic foci, considered as typical axonal components, are ubiquitous in GABA-positive dendrites, it is impossible to distinguish unequivocally which of these structures belong to axons in layers 8 and 9. However, in layer 7, the presence
Xenopus tectal ultrastructure 1575
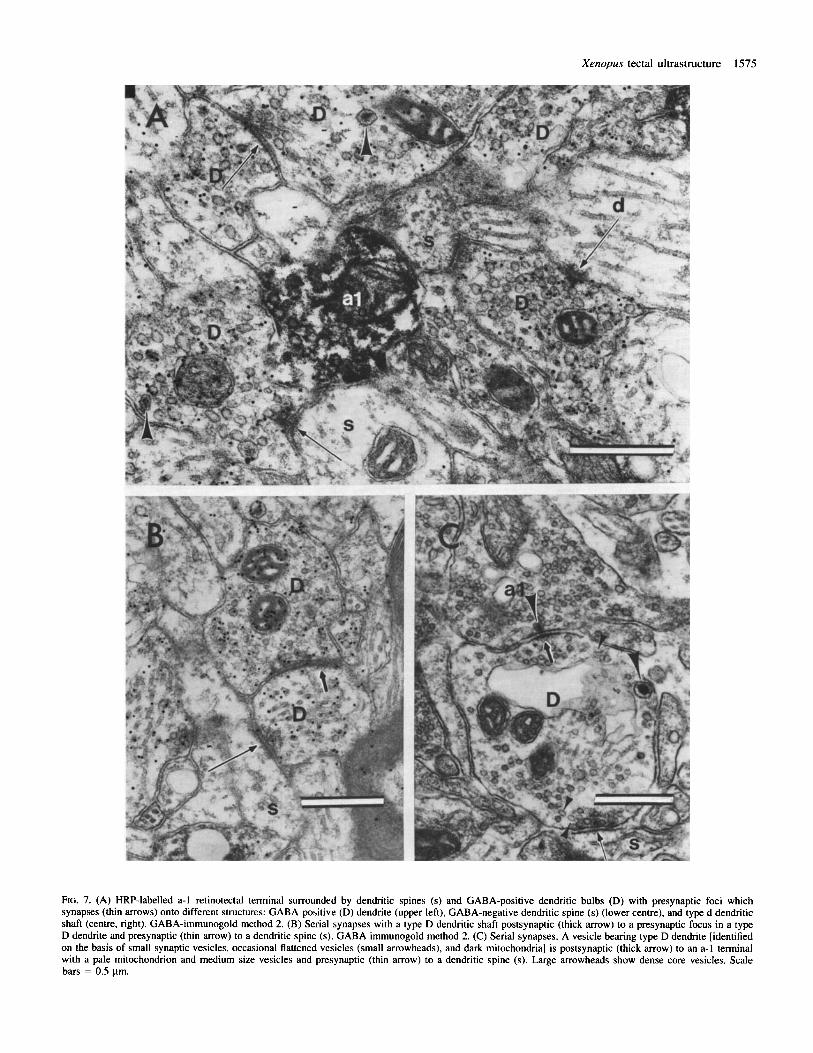
Ftc. 7. (A) HRP-labelled a-1 retinotectal terminal surrounded by dendritic spines (s) and GABA-positive dendritic bulbs (D) with presynaptic foci which synapses (thin arrows) onto different structures: GABA-positive (D) dendrite (upper left), GABA-negative dendritic spine (s) (lower centre), and type d dendritic shaft (centre, right). GABA-immunogold method 2. (B) Serial synapses with a type D dendritic shaft postsynaptic (thick arrow) to a presynaptic focus in a type D dendrite and presynaptic (thin arrow) to a dendritic spine (s). GABA immunogold method 2. (C) Serial synapses. A vesicle bearing type D dendrite [identified on the basis of small synaptic vesicles, occasional flattened vesicles (small arrowheads), and dark mitochondria] is postsynaptic (thick arrow) to an a-1 terminal with a pale mitochondrion and medium size vesicles and presynaptic (thin arrow) to a dendritic spine (s). Large arrowheads show dense core vesicles. Scale bars = 0.5 pm.
1576 Xenopus tectal ultrastructure
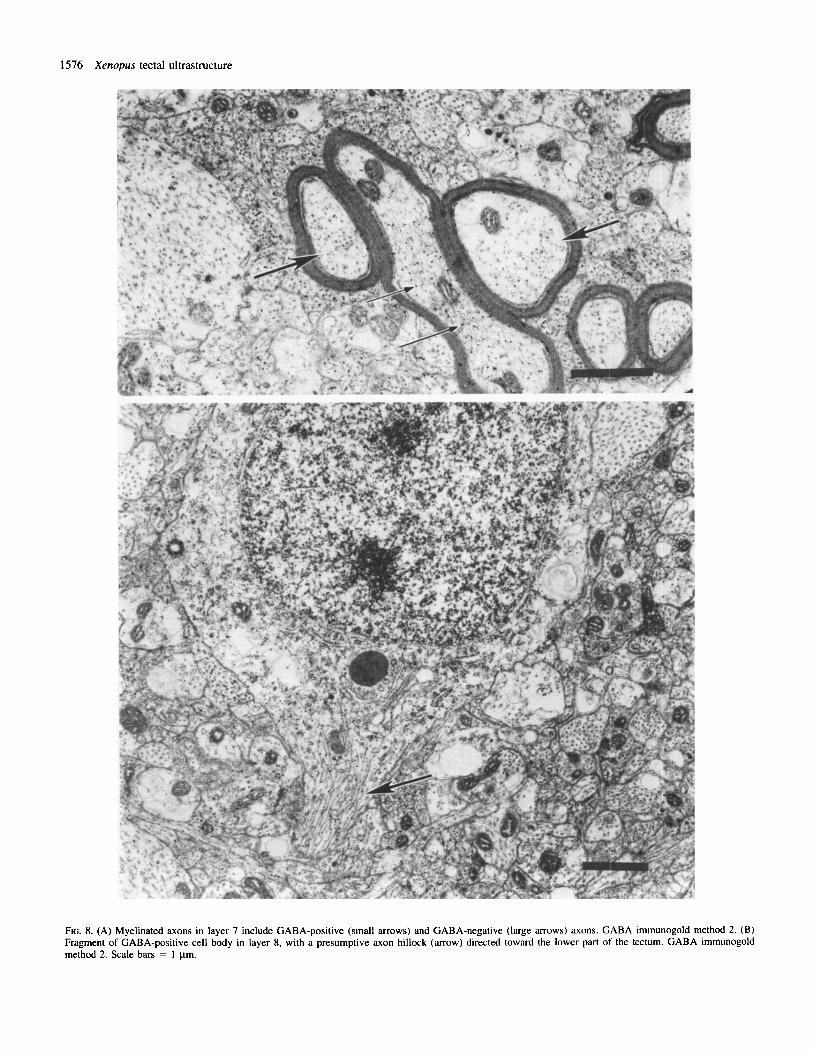
FIG. 8. (A) Myelinated axons in layer 7 include GABA-positive (small arrows) and GABA-negative (large arrows) axons. GABA immunogold method 2. (B) Fragment of GABA-positive cell body in layer 8, with a presumptive axon hillock (arrow) directed toward the lower part of the tectum. GABA immunogold method 2. Scale bars = 1 pm.

Xenopus tectal ultrastructure 1577
of myelin around GABA-positive profiles demonstrates the presence of axons (Fig. 8A).
GABA-positive cell bodies are common in layers 6 and 8. The typical cell bodies are relatively small, with large nuclei containing quite homogeneously distributed chromatin, often showing two nucleoli (Fig. 8B). The cytoplasm contains polyribosomes, rough endoplasmic reticulum, Golgi complexes, vesicles and microtubules. Mitochondria are usually compact, with dense matrix, but less dense mitochondria with the tubular cristae are also observed.
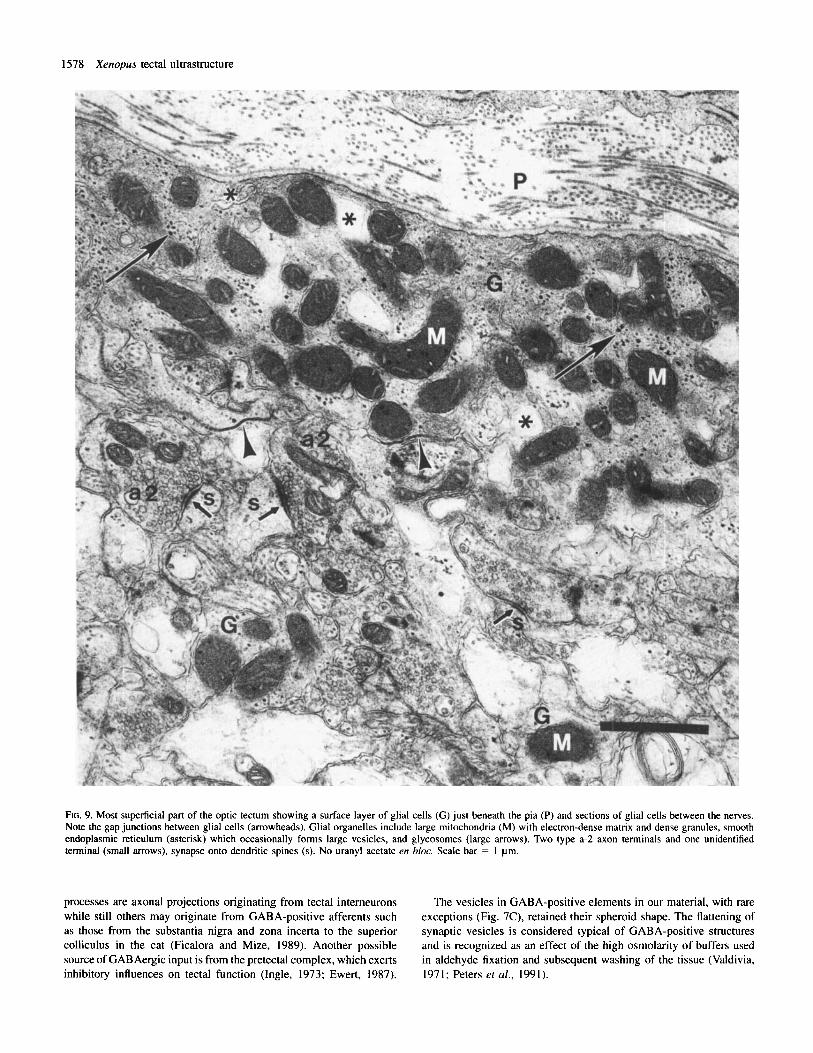
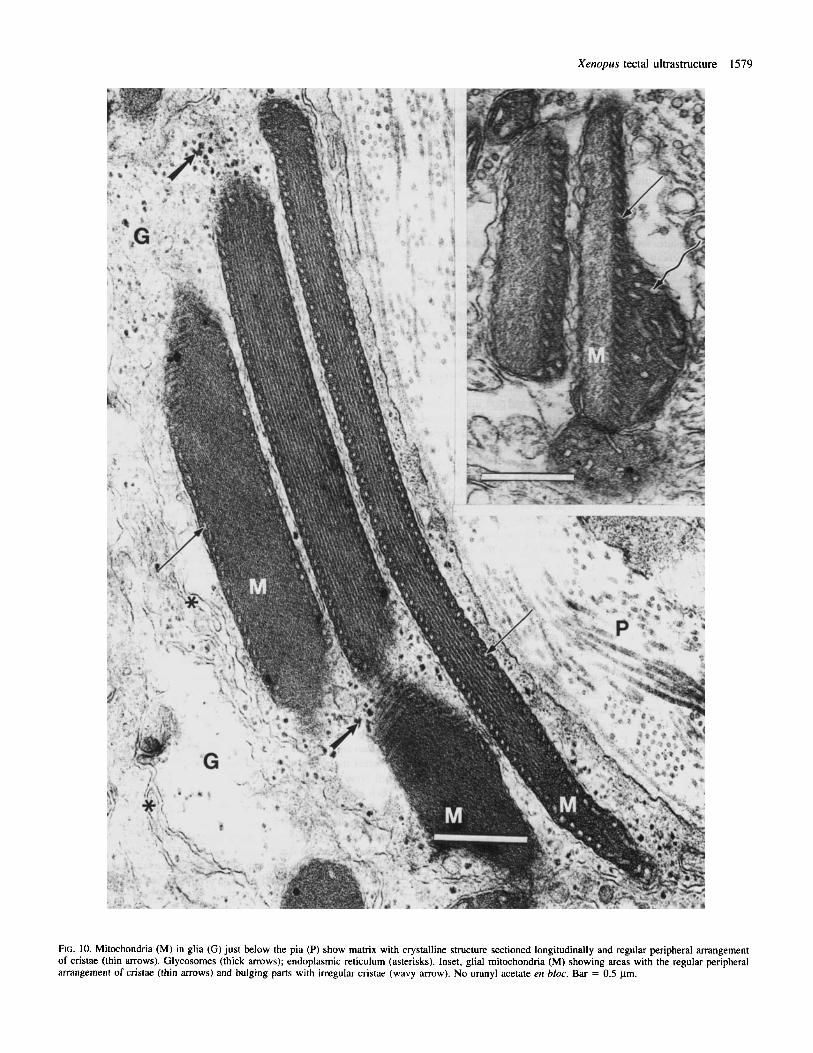
Glial cells Glial cell processes are the most easily identifiable elements in the tectum due to their morphology and to the specific structure, size and abundance of their mitochondria. Glial cytoplasm is moderately electron-dense and contains glycosomes (organelles composed of glycogen and their metabolic enzymes (Rybicka, 1979, 1981)) and membranes of smooth endoplasmic reticulum. The glycosomes are clearly distinguishable only in tissue processed without en bloc treatment with uranyl acetate (Figs 9 and 10). The endoplasmic reticulum often appears as a large, irregular vesicular structure (Fig. 9). Some mitochondria are very long and display a regular crystalline structure in their matrix and an extremely regular pattern of cristae (Fig. 10). Figure 10 (inset) indicates that this regular pattern may appear in some parts of the mitochondrion whereas other parts display the more typical irregularly arranged cristae. The crystalline structure is usually displayed by several mitochondria in every sample, although sections showing the regular pattern presented in Figure 10 are rare.
The surface of layer 9 just beneath the pia is lined with end-feet of ependymoglial cells (Fig. 9). Long gap junctions between glial cells are often observed (Fig. 9, arrowheads). Ependymoglial cells are judged to be GABA-negative, although some gold particles appear occasionally in glial mitochondria.
GABA-positive oligodendrocytes are found in layer 9. They are relatively small glial cells with electron-dense cytoplasm, rich in ribosomes.
Discussion Ultrastructure and GA BA immunoreactivity in superficial layers of Xenopus tectum Electron microscopic samples show GABA immunoreactivity in the majority of cells in layers 8 and 9. Also numerous in layer 6 are GABA-positive cells, although a large proportion of cells in this area remain GABA-negative.
In layer 9, most cell bodies are immunopositive. Little is known about the cells of layer 9 in Xenopus. Some of them project to the nucleus isthmi (S. B. Udin, unpublished observations), but it is unlikely that any of the GABA-positive cell bodies participate in this projection, since the nucleus isthmi is quite strikingly devoid of GABA immunoreactivity, even in sections in which high concentrations of antibody are employed (S. B. Udin, unpublished observations). It is possible that the axons of the stained cells in layer 9 and/or other layers contribute to the population of GABA-positive transverse axons in layer 7. The targets of these axons have yet to be determined.
We have identified four distinct ultrastructural types of neuronal structures in layers 8 and 9. These include two types of GABA- negative axon terminals, GABA-negative dendrites, and GABA-
positive dendrites with presynaptic foci. It is possible that some of the GABA-positive presynaptic structures are axonal rather than dendritic.
The axons classified as type a-I contain synaptic vesicles of medium size and pale mitochondria and participate in asymmetric synapses. These terminals synapse onto either GABA-positive (type D) or GABA-negative (type d) dendrites. Many of these axons are retinotectal axons, as demonstrated by labelling of retinal nerves with HRP. Light microscopic observation prior to embedding the sections revealed that HRP was not taken up by all retinotectal nerves which accounts for the appearance of HRP in only some of the a-I terminals.
Type a-2 axons have large synaptic vesicles and dark mitochondria. Like type a-I axons, they form asymmetric synapses. They are most common in the uppermost part of layer 9, and they synapse mostly onto type d dendrites and less often onto vesicle-bearing dendrites (type D). Type a-2 terminals remain unstained after labelling of the optic nerve with HRP; therefore, they are not retinotectal axons.
Type d dendrites and their spines constitute the majority of the dendritic population and are connected with all types of terminals in layer 9. The spines of type d dendrites appear to be the most common postsynaptic elements in the optic tectum of Xenopus. We have chosen to use the term spine to describe type d postsynaptic structures devoid of microtubules and with light, flocculent cytoplasm (Peters et al., 1991). No spine apparatus is visible in these profiles in Xenopus. A few fortuitous sections unequivocally demonstrate the prototypical stalk and bulb connected to microtubule-containing dendritic shafts. The morphological characteristics of these spines form the criteria for applying the term to other postsynaptic profiles in which continuity with a dendritic shaft has not been established. Due to the small stalk size, such connections are seldom observed in ultrathin sections. Neither light microscopic nor previous electron microscopic studies have demonstrated a prominent number of spines in frog tectum (Szkkely et al., 1973; Potter, 1969; Hughes, 1990). Cobalt labelling demonstrates that only a small proportion of the pyrifom cells of layer 6 are spiny (Antal et al., 1986). It is also possible that some spines are subregions of dendrites that have some of the morphological characteristics of classically defined spines but do not protrude enough to be recognized as such in the light microscope. It seems reasonable to assume that many type d dendrites belong to GABA-negative cell bodies common in layer 6.
A fourth type of structure is the GABA-positive vesicle-bearing dendrite (type D), indicating presynaptic elements defined as presyn- aptic foci. Because this presynaptic region is often clearly in continuity with processes which show dendritic morphology, we use the term ‘presynaptic focus’ rather than ‘axon’ or ‘dendrite’ to describe it. However, it is possible that some of these profiles are terminals of GABA-positive axons, which we were able to recognize unequivocally only in layer 7, where some are myelinated. The paucity of ribosomes in these layers of the tectum prevents us from using the presence or absence of ribosomes as a marker to distinguish axons from dendrites.
Many of the GABA-positive dendrites observed in the electron microscope undoubtedly correspond to the GABA-positive radial structures observed at the light microscopic level. The dendritic nature of these radial processes was sometimes established by virtue of their continuity with GABA-positive cell bodies. These structures are uneven in diameter and density of staining, perhaps because they contain the highest concentration of antigen at the presynaptic foci which we observed in GABA-positive structures with the electron microscope. Occasionally, very small spine-like protrusions were observed.
It is possible that some of the GABA-positive vesicle-bearing
1578 Xenopus tectal ultrastructure
FIG. 9. Most superficial part of the optic tectum showing a surface layer of glial cells ( G ) just beneath the pia (P) and sections of glial cells between the nerves. Note the gap junctions between glial cells (arrowheads). Glial organelles include large mitochondria (M) with electron-dense matrix and dense granules, smooth endoplasmic reticulum (asterisk) which occasionally forms large vesicles, and glycosomes (large arrows). Two type a-2 axon terminals and one unidentified terminal (small arrows), synapse onto dendritic spines (s). No uranyl acetate en bloc. Scale bar = 1 pm.
processes are axonal projections originating from tectal intemeurons while still others may originate from GABA-positive afferents such as those from the substantia nigra and zona incerta to the superior colliculus in the cat (Ficalora and Mize, 1989). Another possible source of GABAergic input is from the pretectal complex, which exerts inhibitory influences on tectal function (Ingle, 1973; Ewert, 1987).
The vesicles in GABA-positive elements in our material, with rare exceptions (Fig. 7C), retained their spheroid shape. The flattening of synaptic vesicles is considered typical of GABA-positive structures and is recognized as an effect of the high osmolarity of buffers used in aldehyde fixation and subsequent washing of the tissue (Valdivia, 1971; Peters ef al., 1991).
Xenopus tectal ultrastructure 1579
FIG. 10. Mitochondria (M) in glia (G) just below the pia (P) show matrix with crystalline structure sectioned longitudinally and regular peripheral arrangement of cristae (thin arrows). Glycosomes (thick arrows); endoplasmic reticulum (asterisks). Inset, glial mitochondria (M) showing areas with the regular peripheral arrangement of cristae (thin arrows) and bulging parts with irregular cristae (wavy arrow). No uranyl acetate en bloc. Bar = 0.5 pm.

1580 Xenopus tectal ultrastructure
Some of the glial cells contain mitochondria with a striking crystalline arrangement of their matrix. They are usually longer than other mitochondria and therefore could be considered as giant mitochondria described in different normal and pathological tissues (Ghadially, 1975). Although giant mitochondria often contain crystalline inclusions, the mitochondria found in Xenopus seem to show a crystalline structure of their matrix rather than inclusions. The functional correlates of this structure are not known.
Comparisons of Xenopus with Rana The basic structures found in Xenopus clearly correspond to the types found in the tecta of R. esculenta (SzCkely et al., 1973; SzCkely and Liz&, 1976) and R. pipiens (Hughes, 1990). The axons that we classify as type a-1 contain synaptic vesicles of medium size and pale mitochondria and participate in asymmetric synapses. These axons correspond to type 1 terminals in R. esculenta (SzCkely and Lh&, 1976). Many of these axons are retinotectal axons, as inferred from degeneration studies after enucleation in R. esculenta (SzCkely et al., 1973; Sztkely and L k k , 1976) and by labelling of retinal nerves with HRP in R. pipiens (Hughes, 1990), and in Xenopus in the present study.
Type a-2 axons have large synaptic vesicles and dark mitochondria. Like type a-1 axons, they form asymmetric synapses. These axons correspond to the type 2 axons described in R. esculenta (SzCkely et al., 1973) and to the terminals with round, large (RL) synaptic vesicles in R. pipiens (Hughes, 1990) although we find that vesicles are more densely packed than those described in Rana. They are most common in the uppermost part of layer 9, and they synapse mostly onto ‘pale dendrites’ (our type d) and less often onto the ‘vesicle-bearing dendrites’ (our type D). The fact that these axons do not originate from the retina is established by the fact that they are resistant to the enucleation procedure which led to the degeneration of the frog optic nerve (SzCkely et al., 1973) and remain unstained after labelling of the optic nerve with HRP in Rana (Hughes, 1990) as well as in Xenopus.
Type d dendrites, and particularly their poorly contrasted spines, clearly correspond to the ‘pale’ dendrites in R. esculenta, where they constitute the majority of the dendritic population and are connected with all types of terminals in layer 9 (SzCkely et al., 1973).
The presynaptic foci that we have identified in type D dendrites correspond to the type 3 terminals with small synaptic vesicles, electron-dense mitochondria, and symmetric synapses which were judged to be the dendritic spines of large, pear-shaped interneurons in R. esculenta (Sztkely et al., 1973; Sztkely and Liz&, 1976). Hughes (1990) classified them as the terminals bearing flattened vesicles (FM terminals) in R. pipiens. GABA-positive terminals often show flattened or pleomorphic synaptic vesicles (Van der Togt et al., 1991; Rio et al., 1992). The spheroid shape of these vesicles observed in Xenopus was also retained in the tectum of R. esculenta (SzCkely et al., 1973).
GABA-positive presynaptic structures are the second most numer- ous presynaptic profiles both in Rana and in Xenopus. Their prevalence is consistent with Antal’s estimate that one-third of tectal cells in Rana are GABA-positive (Antal, 1991).
Antal (I99 I), using GABA immunoreactivity to characterize the optic tectum of R. esculenta, defined two classes of GABA-positive dendrites, one type containing many microtubules and the other containing few microtubules. We have found only one identifiable type of GABA-positive dendrite, but we also see microtubule rich domains, classified as dendritic shafts, and microtubule-poor domains,
classified as dendritic extensions. Also, Antal distinguished two types of GABA-positive axon terminals while we have identified presynaptic foci in GABA-positive dendrites. As discussed above, it is extremely difficult to make an unequivocal identification of these profiles as belonging to axons or dendrites without employing additional techniques such as double-labelling.
Comparisons of Xenopus with other species Layers 8 and 9 of the frog tectum are comparable to the superficial layers of the mammalian superior colliculus (tectum) in being the layers that receive direct input from the retina and the nucleus isthmi [parabigeminal (Graybiel, 1978)l. There are, however, some prominent differences, notably the presence of a corticotectal projection in mammals and its absence in frogs. Also, most of the cells whose dendrites are found in layers 8 and 9 of the frog tectum have somata in deeper layers, while the cells with dendrites in superficial layers of the mammalian tectum have somata within those layers.
There are striking similarities, as well as some differences, between the synaptic arrangements that we report for Xenopus tectum and those described for the mammalian superior colliculus (Mize, 1988, 1992). GABA-positive somata and processes are numerous in the tectum and in the superior colliculus. In the latter, there are several identifiable morphological classes of GABA-positive cells. In Xenopus, our light microscopic staining does not clearly reveal difficult classes, but the fact that staining occurs in all cellular levels as well as in layer 9 implies that multiple classes of cells are GABA-positive in this species. Both colliculus and tectum contain GABA-positive myelinated axons.
In both mammals and Xenopus frogs, GABA-positive vesicle- containing dendrites are common, and it is often difficult or impossible to conclusively distinguish such dendrites from axons on the basis of ultrastructural characteristics. In both tectum and collic- ulus, presynaptic GABA-positive dendrites receive direct input from retinal terminals and participate in serial or triadic arrangements with output to other dendrites, some of which are GABA-positive. A possible dissimilarity between mammals and Xenopus is that presyn- aptic dendritic structures may be located on spines in mammals, while in Xenopus they seem to be associated with local protuberances of the dendrites. In both tectum and colliculus, the vesicles in presynaptic dendrites are small and generally have a round or ovoid shape rather than the flattened aspect more typically associated with inhibitory synapses (Mize, 1992). In both Xenopus and mammals, vesicles in some presynaptic dendrites are found as loose accumula- tions while in other presynaptic dendrites the vesicles are densely packed. In mammals, these two patterns are associated with granule neurons and horizontal cells, respectively. We have no direct evidence whether these two patterns of distribution are found in two distinct cell types in Xenopus. A third type of GABA-positive profile has been identified in mammals and contains flattened vesicles (Mize, 1992); we have not seen comparable processes in Xenopus.
The triadic arrangement, with GABA-positive processes that contain small vesicles located postsynaptic to retinal terminals and presynaptic to other dendrites, has been observed in tectum reptiles and birds (Streit et al., 1978; Rio et al., 1992). Comparable arrangements have been reported for mammalian lateral geniculate nucleus (Hendrickson et al., 1983).
Functional implications GABA-positive presynaptic structures are the second most numerous presynaptic profiles in Xenopus. Their prevalance underscores the

Xenopus tectal ultrastructure 158 1
importance of inhibitory circuitry in the tectum a s in most brain structures. The frequent convergence of GABA-positive and GABA- negative profiles onto the same GABA-negative postsynaptic element indicates that inhibitory synapses are appropriately positioned to oppose the action of excitatory inputs. The potential complexity of information processing is underscored by the fact that GABA-positive dendrites receive both GABA-positive and GABA-negative inputs and are involved in serial synaptic arrays. Moreover, there may be presynaptic as well a s postsynaptic GABA receptors, allowing not only inhibition of postsynaptic targets but also enhancement of net excitation by suppressing the release of GABA. Such circuitry may underlie the seemingly paradoxical enhancement of postsynaptic potentials by GABA in the tectum of Rana (Mazda et a/., 1990). although the more typical effect of GABA receptor blockers is to promote high levels of spontaneous activity.
GABAergic cells are likely to play a role in activity-dependent refinement of retinotectal and isthmotectal axons. There is substantial evidence that such refinement is dependent upon activation of N-methyh-aspartate (NMDA) receptors (Constantine-Paton et a/., 1990; Udin and Scherer, 1990). and inhibitory processes can influence the depolarization that is required for calcium fluxes through NMDA receptors (Nowak et al., 1984). Such interactions have been demon- strated in the hippocampus, in which activation of N M D A receptors is increased by the autosuppression of G A B A release; the net effect is to promote induction of long-term potentiation (Davies e ta / . , 1991; Mott and Lewis, 1991). Activity-dependent alterations in ocular dominance columns in the cat visual cortex are also influenced by experimental activation of G A B A receptors (Reiter and Stryker, 1988).
Acknowledgements
We thank Dr K. D. Phend for kind assistance with the immunogold labelling technique, Ms Julie Lakatos for her great help with preparation of photographs, Mr Jeff Spengler from Incstar for the supply of GABA/BSA conjugate, and Drs J. Murphy and D. Williams, and Ms L. Cannon-Murakami for comments on the manuscript. This work was supported by USPHS grant EY-03470 to S. B. U.
Abbreviations
BSA GABA HRP NGS PB TB S TB ST TPBS
bovine serum albumin y-aminobutyric acid horseradish peroxi dase normal goat serum phosphate buffer Tris-buffered saline Tris-HCI buffered saline with Triton X- 100 Tris-HCI phosphate buffered saline
References
Adams, J. C. (1981) Heavy metal intensification of DAB-based HRP reaction product. J . Histochem. Cytochem., 29, 775.
Antal. M. (1991) Distribution of GABA immunoreactivity in the optic tectum of the frog: A light and electron microscopic study. Neuroscience, 42,
Antal, M., Matsumoto, N. and Sztkely, G. (1986) Tectal neurons of the frog: intracellular recording and labeling with cobalt electrodes. J . Comp. Neurol.,
Constantine-Paton, M., Cline, H. T. and Debski, E. (1990) Patterned activity, synaptic convergence, and the NMDA receptor in developing visual pathways. Annu. Rev. Neurosci., 13, 129-154.
879-891.
246, 238-253.
Davies, C. H., Starkey, S. J., Pozza, M. F. and Collingridge, G. L. (1991) GABAB autoreceptors regulate the induction of LTP. Nature, 349, 60961 1.
Ewert, J.-P. (1 987) Neuroethology of releasing mechanisms: Prey-catching in toads. Behav. Brain Sci., 10, 337405.
Ficalora, A. S. and Mize, R. R. (1989) The neurons of the substantia nigra and zona incerta which project to the cat superior colliculus are GABA immunoreactive: A double-label study using GABA immunocytochemistry and lectin retrograde transport. Neuroscience, 29, 567-58 I .
Ghadially, F. N. (1975) Ultrastrucrural Pathology of the Cell. Butterworths, London.
Graybiel, A. M. (1978) A satellite system of the superior colliculus: the parabigeminal nucleus and its projections to the superficial collicular layers. Brain Res., 145, 365-374.
Hendrickson, A. E., Ogren, M. P., Vaughn, J. E., Barber, R. P. and Wu, J.-Y. ( I 983) Light and electron microscopic immunocytochemical localization of glutamic acid decarboxylase in monkey geniculate complex: Evidence for GABAergic neurons and synapses. J . Neurosci., 3, 1245-1262.
Hughes, T. E. (1990) A light- and electron-microscopic investigation of the optic tectum of the frog, Rana pipiens, 1: The retinal axons. Vis. Neurosci.,
Ingle, D. (1973) Disinhibition of tectal neurons by pretectal lesions in the frog. Science, 180, 422-424.
Mazda. G. Y., Nistri, A. and Sivilotti, L. (1990) The effect of GABA on the frog optic tectum is sensitive to ammonium and to penicillin. Euc J. Pharmacol., 179, 1 1 1-1 18.
Mize, R. R. ( 1 988) Immunocytochemical localization of gamma-aminobutyric acid (GABA) in the cat superior colliculus. J . Comp. Neurol., 267, 169-187.
Mize, R. R. (1992) The organization of GABAergic neurons in the mammalian superior colliculus. Prog. Bruin Res., 90, 2 19-248.
Mott, D. D. and Lewis, D. V. (1991) Facilitation of the induction of long- term potentiation by GABAB receptors. Science, 252, 17 18-1 720.
Nowak, L., Bregestovski, P., Ascher, P., Herbert, A. and Prochiantz, A. (1984) Magnesium gates glutamate-activated channels in mouse central neurones. Nature, 307, 462465.
Peters, A., Palay, S. L. and Webster, H. d. F. (1991) The Fine Structure of the Nervous System. Neurons and Their Supporting Cells. Oxford University Press, Oxford, UK.
Phend, K. D., Weinberg, R. J. and Rustioni, A. (1992) Techniques to optimize post-embedding single and double staining for amino acid neurotransmitters. J . Histochem. Cytochem., 40, 101 1-1020.
Potter, H. D. (1969) Structural characteristics of cell and fiber populations in the optic tectum of the frog (Rana catesbeiana). J. Comp. Neurol., 136, 203-232.
Reiter, H.-0. and Stryker, M. P. (1988) Neural plasticity without postsynaptic action potentials: Less active inputs become dominant when kitten visual cortical cells are pharmacologically inhibited. Proc. Natl. Acad. Sci. USA,
Rio, J.-P., Reperant, J., Ward, R., Miceli, D. and Medina, M. (1992) Evidence of GABA-immunopositive neurons in the dorsal part of the lateral geniculate nucleus of reptiles: morphological correlates with interneurons. Neuro- science, 47, 395-407.
Rybicka, K. (1979) Glycosomes (protein-glycogen complex) in the canine heart. Ultrastructure, histochemistry and changes induced by acidic treat- ment. Virchows Arch. B., 30, 335-347.
Rybicka, K. (1981) Binding of glycosomes to endoplasmic reticulum and to intermediate filaments in cardiac conduction fibers. J. Histochem. Cyrochem.,
Somogyi, P. and Soltesz, 1. (1986) Immunogold demonstration of GABA in synaptic terminals of intracellularly recorded, horseradish peroxidase-filled basket cells and clutch cells in the cat’s visual cortex. Neuroscience, 19,
Streit, P., Knecht, E., Reubi, J.-C., Hunt, S. P. and Cuenod, M. (1978) GABA- specific presynaptic dendrites in pigeon optic tectum: A high resolution autoradiographic study. Brain Res., 149, 204-210.
SzCkely, G. and LaZAr, G. (1976) Cellular and synaptic architecture of the optic tectum. In Llinis, R. and Precht, W. (eds), Frog Neurobiology. Springer-Verlag, New York, 407-434.
Sztkely, G., Stti16, G. and Liz&, G. (1973) Fine structure of the frog’s optic tectum: optic fiber termination layers. J. Hirnforsch., 14, 189-225.
Trachtenberg, M. C. and Ingle, D. (1974) Thalamo-tectal projections in the frog. Brain Res., 79, 419-430.
Udin, S. B. and Fisher, M. D. (1985) The development of the nucleus isthmi in Xenopus laevis: 1. Cell genesis and formation of connections with the
4,499-5 18.
85. 3623-3627.
29, 553-560.
105 I - I 065.

1582 Xenopus tectal ultrastructure
tecta. J. Comp. Neurol., 232, 25-35. Udin, S. B. and Keating, M. J. (1981) Plasticity in a central nervous pathway
in Xenopus: Anatomical changes in the isthmotectal projection after larval eye rotation. J. Comp. Neurol., 203, 575-594.
Udin, S. B. and Scherer, W. J. (1990) Restoration of the plasticity of binocular maps by NMDA after the critical period in Xenopus. Science, 249,669-672.
Valdivia, 0. (1971) Methods of fixation and the morphology of synaptic vesicles. J. Cotnp. Neurol., 142, 257-274.
Van der Togt, C., Cardozo, B. N. and Van der Want, J. (1991) Medial terminal nucleus terminals in the nucleus of the optic tract contain GABA: an electron microscopical study with immuntxytochemical double labelling of GABA and PHA-L. J. Coip. Neurol.. 312, 231-241.