simulated microgravity induce glutathione antioxidant pathwayin xenopus laevis embryos
TRANSCRIPT

Cell Biology International 33 (2009) 893e898www.elsevier.com/locate/cellbi
Short communication
Simulated microgravity induce glutathione antioxidant pathwayin Xenopus laevis embryos
Angela M. Rizzo*, Gigliola Montorfano, Manuela Negroni, Paola Corsetto, Patrizia Berselli,Paola Marciani, Stefania Zava, Bruno Berra
Dipartimento di Scienze Molecolari Applicate ai Biosistemi, DISMAB, Via D. Trentacoste 2, I-20134 Milano, Italy
Received 4 November 2008; revised 27 March 2009; accepted 24 April 2009
Abstract
Space flights cause a number of patho-physiological changes. Oxidative damage has been demonstrated in astronauts after space flights.Oxidative stress is due to an imbalance between production of oxidant and antioxidative defence. In embryos of Xenopus laevis, the glutathionesystem is an inducible antioxidant defence. For this reason, we investigated the effect of gravity deprivation on endogenous antioxidant enzymesin X. laevis embryos developed for 6 days in a Random Positioning Machine. The results show that glutathione content and the activity ofantioxidant enzymes increase in RPM embryos, suggesting the presence of a protective mechanism. An induction of antioxidant defence mightplay an important role for animals to adapt to micro-gravitational stress, possibly during actual space flights.� 2009 International Federation for Cell Biology. Published by Elsevier Ltd. All rights reserved.
Keywords: Simulated microgravity; Xenopus laevis embryos; Glutathione; Oxidative stress; Random positioning machine
1. Introduction
Oxygen dependent metabolism produces reactive oxygenspecies (ROS). Low levels of ROS play a beneficial role inprocesses such as defence against microorganisms and intra-cellular signalling. Nonetheless, ROS are a potential threat toimportant bio-molecules and metabolic functions. Indeedoxidative stress has been related to ageing and to a number ofchronic and acute diseases (for a review see Valko et al.,2007). In aerobic organisms, enzymatic and non-enzymaticactivities provide a defence against the accumulation of ROS.The health and survival of organisms is dependent on theregulation and balance of antioxidant activities.
The antioxidant defence of cells consists of enzymatic andnon-enzymatic antioxidants that operate together with repair-ing mechanisms (Thorpe et al., 2004; Hayes and Mc Lellan,1999; Yu, 1994). The enzymatic antioxidant system is
* Corresponding author. Tel.: þ39 025 031 5789; fax: þ39 025 031 5775.
E-mail address: [email protected] (A.M. Rizzo).
1065-6995/$ - see front matter � 2009 International Federation for Cell Biology.
doi:10.1016/j.cellbi.2009.04.015
composed mainly of peroxidases and catalases. A distinct typeof antioxidant defence is represented by heat shock proteins(Anderson, 1996). Other non-enzymatic antioxidants are alsoavailable, such as vitamin E, Vitamin C, carotenoids, flavo-noids, coenzyme Q10 and squalene.
Among endogenous antioxidants, reduced glutathione(GSH) plays a key role. GSH is an intracellular tripeptide thatreacts with ROS, reduces hydro-peroxides and breaks disulfidebridges formed by oxidants; its antioxidant action producesglutathione disulfide (GSSG) that can be either exported fromthe cell or reduced back to GSH by GSH reductase andNADPH. GSH also produces antioxidant molecules regener-ating ascorbate from dehydroascorbate and a-tocopherol fromtocopheroxyl radical (Franco et al., 2007).
Exposure to microgravity and radiations during spaceflights could be responsible for a number of changes in livingorganisms resembling modifications that occur in ageing. Infact human life in space and ageing are both characterized byincreased stress hormones, insulin resistance, anorexia, alteredmusculoskeletal system structure and function, and alteredinflammatory response and mitochondrial function with
Published by Elsevier Ltd. All rights reserved.

894 A.M. Rizzo et al. / Cell Biology International 33 (2009) 893e898
increased oxidative stress (Stein, 2002; Finkel and Holbrook,2000; Stein and Leskiw, 2000). Of note, increased oxidativestress and sensitivity to oxidants are two main aspects sharedby ageing and space environment, and several studies ondifferent species are consistent with a causal connectionbetween increased stress resistance and extended life span (Orrand Sohal, 1994).
Oxidative stress can be caused either by a geneticallydetermined ‘‘weakness’’ of the antioxidant defence, or by anincrease of the random damage caused by an over exposure toreactive oxygen metabolites (ROS). Since many types ofionizing radiations that are encountered on Earth, and more inspace, are capable of generating ROS and free radicals (Guanet al., 2006; Hollander et al., 1998; Lyng et al., 2001), radia-tion is a potential threat for long-term outer-space missions.Nonetheless many metabolic processes must be taken inaccount since the oxidative stress can be due to a number ofco-existing factors.
The role of microgravity on oxidative stress during spaceflights has not been resolved. Some authors have suggestedthat microgravity might enhance pro-oxidant processes, whileothers did not record synergic effects. Although there aremany discrepancies among the results, presumably due to thevariety of organisms that have been tested, the relevance ofsuch studies lies on the presumption that results could suggeststrategies to protect living organisms, perhaps also humans,when exposed to the space environment (Manti, 2006).
Xenopus laevis has been one of the first organisms used inspace and stratospheric balloon flights to study fertilizationand embryogenesis processes (Rizzo et al., 2007a,b). Our aimwas to investigate whether microgravity exposure of X. laevisembryos might influence antioxidant defences of the animalcontributing to enhance oxidative stress.
2. Materials and methods
2.1. Substrates and reactive for enzyme determinations
NAD(P)H, DTNB, GSH, GSSG, glutathione reductase andtert-butyl hydroperoxide were purchased from SigmaeAldrich(St. Louis, Missouri, USA).
2.2. Embryo culture
Embryos of X. laevis were obtained by in vitro fertilization(Rizzo et al., 1994). Briefly, Xenopus, (Rettili, Varese, Italy),were maintained in aquaria (Tecniplast, Varese, Italy).Females were injected with 700e800 I.U. of human chorionicgonadotropin in the evening, and the next day they were madeto lay eggs in 90 mm plastic Petri dishes. Eggs were imme-diately inseminated with sperm suspension obtained bymincing testes in 1e2 mL of cold DBT solution. Eggs wereselected, and the embryos with deformations were eliminated.Embryos were maintained in a thermostatic chamber at23� 0.5 �C for 1 day, and collected with large bore pipettes.
2.3. Microgravity simulation
Simulation of microgravity was achieved with a RandomPositioning Machine (RPM, Dutch, Space, Leiden, TheNetherlands). It is essentially a 3-axis clinostat, in which theweight vector is continually reoriented as in traditional cli-norotation, but with increased directional randomization(van Loon, 2007).
In the RPM, probes are fixed as close as possible to thecentre of 2 frames rotating one inside the other, driven byseparate motors. The rotation of each frame is random andautonomous under computer control. The low g conditions aremodelled by averaging the gravity vector via the independentrotation of the 2 frames. The outer frame rotated perpendicularto the inner frame, which caused the samples to moverandomly in 3 axes.
The speed of rotation was 60 �/s (about 10�3� g), anddirection and interval were set at random. The RPM wasaccommodated in a temperature-controlled incubator at 23 �C.In the experiments, both control and microgravity embryos(ageing 1 day) were housed in T25 vented cell culture flaskfull of Fetax medium (Rizzo et al., 1994). After 3 days ofincubation, the instrument was stopped for few minutes toallow medium changes. Control samples (1� g) were placedon the fixed base of the RPM, facing the same vibrations as therotating ones. At day 3 and 6, RPM and control embryos weresampled, frozen at �80 �C for enzymatic assays, or immedi-ately homogenized for total glutathione determination.
2.4. Enzymes assay
Embryos were homogenized on ice in H2O or 100 mM, pH7.0 K-phosphate buffer and centrifuged at 1000 �g for 10 minat 4 �C.; supernatants were assayed for protein contentaccording to Peterson (1977).
2.4.1. Superoxide dismutase (SOD) activity assayEnzyme activity was assayed using the method based on
NAD(P)H oxidation inhibition, according to Paoletti andMocali (1990). Briefly, the inhibition of NADPH oxidation bychemically generated superoxide was measured at 340 nm for20 min, in the presence of tissue extracts. The incubationmixture included: 800 ml of TDB (triethanolamine/diethanol-amine 100 mM, pH 7.4), 40 ml NADPH 7.5 mM, 25 mlEDTAeMnCl2 (100 mM/50 mM), and 100 ml of sample orblank. One unit of SOD activity is defined as the amount ofenzyme required to inhibit the rate of NADPH oxidation by50%.
2.4.2. Catalase (CAT) activity assaySupernatant enzyme activity was assayed by measuring the
consumption of H2O2 according to Aebi (1984). Theconsumption of hydrogen peroxide by the tissue extracts wasdetermined at 240 nm for 1 min at 30 �C. The incubationmixture included: 50 ml H2O2 200 mM, 100 mg of proteins oftissue extract, and Na-phosphate buffer (50 mM pH 7.0) toreach a final volume of 1 mL. One unit of CAT activity is

895A.M. Rizzo et al. / Cell Biology International 33 (2009) 893e898
defined as the amount of enzyme required to catalyse thedecomposition of 1 mmol H2O2 min�1.
2.4.3. Glutathione reductase (GR) activity assaySupernatant enzyme activity was assayed according to
Pinto et al. (1984). Briefly, GSSG reduction and NADPHconsumption were recorded at 340 nm. The incubationmixture included: 20 mL GSSG 125 mM, 11 ml NADPH11 mM, 100 mg of proteins of tissue extract, and K-phosphatebuffer (100 mM pH 7.0) to 1 mL final volume.
2.4.4. Glutathione peroxidase (GPx) activity assaySupernatant enzyme activity of selenium-dependent GPx
was assayed according to Prohaska and Ganther (1976) byfollowing the decrease in the absorbance at 340 nm for 3 min,which corresponds to the rate of GSH oxidation to GSSG inthe presence of NADPH and glutathione reductase. Theincubation mixture included: 20 ml GSH 100 mM, 10 mlNADPH 11 mM, GR 1 Unit, 10 ml terbutylhydroperoxide20 mM, 150 mg of proteins of tissue extract, and EDTA-Kphosphate buffer (3 mMe100 mM pH 7.0) to 1 mL finalvolume. One unit of GR or GPx activity is defined as theamount of enzyme required to catalyse the oxidation of1 mmol NADPH min�1.
2.5. Determination of the total glutathione (GSH)
Immediately after collection, embryos were homogenizedon ice in 5% metaphosphoric acid; the homogenate wascentrifuged at 5000 g for 10 min at 4 �C and the supernatantwas assayed according to Griffith (1985), with slight modifi-cations. Briefly, the sulfhydryl group of GSH, also generatedfrom GSSG by adding GR, reacts with DTNB (5,50-dithio-bis-2-nitrobenzoic acid) and produces a yellow-coloured 5-thio-2-nitrobenzoic acid (TNB). The rate of TNB production isdirectly proportional to the concentration of GSH in thesample. Measurement of the absorbance of TNB at 412 nmprovides an accurate estimation of the GSH level present in thesample.
Fig. 1. Malformations induced in Xenopus laevis embryos d
2.6. Determination of thiobarbituric acid reactivesubstances (TBARS)
Embryos were homogenized on ice in H2O; samples,standards (from 2.5 to 100 pmol TEP, 1,1-3,3 tetraetox-ipropane) and blank were assayed according to Wey et al.(1993). Induction of lipid peroxidation involved incubation ofextracts for 2 h at 30 �C with FeSO4 and ascorbic acid.TBARS were determined with a fluorescence spectropho-tometer (Carly Eclipse, Varian, CA USA) excitation wave-length: 517 nm, emission wavelength: 550 nm.
2.7. Statistical analysis
Differences between microgravity exposed and controlembryos, of the same age, were analyzed by Student’sunpaired t-test.
3. Results and discussion
Since space flights began many species have been studiedin microgravity conditions to increase our knowledge of theeffects of gravity deprivation and reducing human space flightrisks. Tadpoles, during space flights, suffered behavioural andmorphological modifications such as: swimming abnormalitiesand hyperextension of the tail (lordosis); which persisted forsome days after grounding, as observed in X. laevis tadpoleslaunched before hatching (Horn, 2006).
We used RPM to expose X. laevis embryos to a reducedgravity. Although RPM is a widely used microgravity simu-lator, cautions must be taken before ascribing effects shown inRPM experiments to microgravity (Hoson et al., 1997). RPMprinciples are not completely understood and even when everyprecaution has been taken, some aspects of RPM machinerysuch as fluid motion or mechanical stress can influence theinterpretation of the data (van Loon, 2007), and confirmatoryexperiments need to be carried in space flight conditions.
We found w10% of X. laevis embryos grown in RPMconditions displayed axial malformation (Fig. 1). Tail lordosis
eveloped for 6 days in simulated microgravity (RPM).
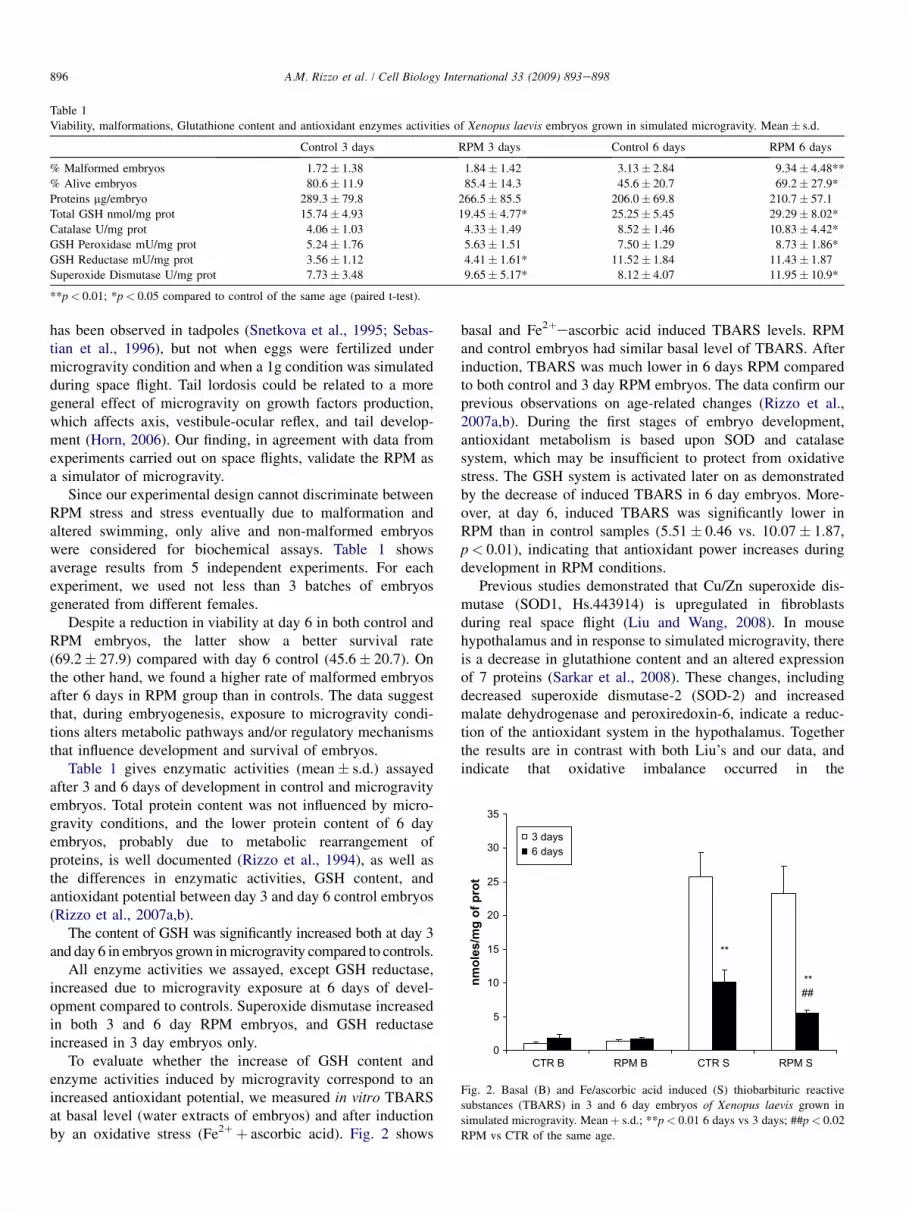
Table 1
Viability, malformations, Glutathione content and antioxidant enzymes activities of Xenopus laevis embryos grown in simulated microgravity. Mean� s.d.
Control 3 days RPM 3 days Control 6 days RPM 6 days
% Malformed embryos 1.72� 1.38 1.84� 1.42 3.13� 2.84 9.34� 4.48**
% Alive embryos 80.6� 11.9 85.4� 14.3 45.6� 20.7 69.2� 27.9*
Proteins mg/embryo 289.3� 79.8 266.5� 85.5 206.0� 69.8 210.7� 57.1
Total GSH nmol/mg prot 15.74� 4.93 19.45� 4.77* 25.25� 5.45 29.29� 8.02*
Catalase U/mg prot 4.06� 1.03 4.33� 1.49 8.52� 1.46 10.83� 4.42*
GSH Peroxidase mU/mg prot 5.24� 1.76 5.63� 1.51 7.50� 1.29 8.73� 1.86*
GSH Reductase mU/mg prot 3.56� 1.12 4.41� 1.61* 11.52� 1.84 11.43� 1.87
Superoxide Dismutase U/mg prot 7.73� 3.48 9.65� 5.17* 8.12� 4.07 11.95� 10.9*
**p< 0.01; *p< 0.05 compared to control of the same age (paired t-test).
0
5
10
15
20
25
30
35
CTR B RPM B CTR S RPM S
nm
oles/m
g o
f p
ro
t
3 days6 days
**
**##
Fig. 2. Basal (B) and Fe/ascorbic acid induced (S) thiobarbituric reactive
substances (TBARS) in 3 and 6 day embryos of Xenopus laevis grown in
simulated microgravity. Meanþ s.d.; **p< 0.01 6 days vs 3 days; ##p< 0.02
RPM vs CTR of the same age.
896 A.M. Rizzo et al. / Cell Biology International 33 (2009) 893e898
has been observed in tadpoles (Snetkova et al., 1995; Sebas-tian et al., 1996), but not when eggs were fertilized undermicrogravity condition and when a 1g condition was simulatedduring space flight. Tail lordosis could be related to a moregeneral effect of microgravity on growth factors production,which affects axis, vestibule-ocular reflex, and tail develop-ment (Horn, 2006). Our finding, in agreement with data fromexperiments carried out on space flights, validate the RPM asa simulator of microgravity.
Since our experimental design cannot discriminate betweenRPM stress and stress eventually due to malformation andaltered swimming, only alive and non-malformed embryoswere considered for biochemical assays. Table 1 showsaverage results from 5 independent experiments. For eachexperiment, we used not less than 3 batches of embryosgenerated from different females.
Despite a reduction in viability at day 6 in both control andRPM embryos, the latter show a better survival rate(69.2� 27.9) compared with day 6 control (45.6� 20.7). Onthe other hand, we found a higher rate of malformed embryosafter 6 days in RPM group than in controls. The data suggestthat, during embryogenesis, exposure to microgravity condi-tions alters metabolic pathways and/or regulatory mechanismsthat influence development and survival of embryos.
Table 1 gives enzymatic activities (mean� s.d.) assayedafter 3 and 6 days of development in control and microgravityembryos. Total protein content was not influenced by micro-gravity conditions, and the lower protein content of 6 dayembryos, probably due to metabolic rearrangement ofproteins, is well documented (Rizzo et al., 1994), as well asthe differences in enzymatic activities, GSH content, andantioxidant potential between day 3 and day 6 control embryos(Rizzo et al., 2007a,b).
The content of GSH was significantly increased both at day 3and day 6 in embryos grown in microgravity compared to controls.
All enzyme activities we assayed, except GSH reductase,increased due to microgravity exposure at 6 days of devel-opment compared to controls. Superoxide dismutase increasedin both 3 and 6 day RPM embryos, and GSH reductaseincreased in 3 day embryos only.
To evaluate whether the increase of GSH content andenzyme activities induced by microgravity correspond to anincreased antioxidant potential, we measured in vitro TBARSat basal level (water extracts of embryos) and after inductionby an oxidative stress (Fe2þþ ascorbic acid). Fig. 2 shows
basal and Fe2þeascorbic acid induced TBARS levels. RPMand control embryos had similar basal level of TBARS. Afterinduction, TBARS was much lower in 6 days RPM comparedto both control and 3 day RPM embryos. The data confirm ourprevious observations on age-related changes (Rizzo et al.,2007a,b). During the first stages of embryo development,antioxidant metabolism is based upon SOD and catalasesystem, which may be insufficient to protect from oxidativestress. The GSH system is activated later on as demonstratedby the decrease of induced TBARS in 6 day embryos. More-over, at day 6, induced TBARS was significantly lower inRPM than in control samples (5.51� 0.46 vs. 10.07� 1.87,p< 0.01), indicating that antioxidant power increases duringdevelopment in RPM conditions.
Previous studies demonstrated that Cu/Zn superoxide dis-mutase (SOD1, Hs.443914) is upregulated in fibroblastsduring real space flight (Liu and Wang, 2008). In mousehypothalamus and in response to simulated microgravity, thereis a decrease in glutathione content and an altered expressionof 7 proteins (Sarkar et al., 2008). These changes, includingdecreased superoxide dismutase-2 (SOD-2) and increasedmalate dehydrogenase and peroxiredoxin-6, indicate a reduc-tion of the antioxidant system in the hypothalamus. Togetherthe results are in contrast with both Liu’s and our data, andindicate that oxidative imbalance occurred in the

897A.M. Rizzo et al. / Cell Biology International 33 (2009) 893e898
hypothalamus in response to simulated microgravity. It ispossible that the observed effects on brain of suspended miceare related to stress per se, due to the head-down condition.
In contrast, in Anabaena cells, ROS increased undersimulated microgravity provided by a clinostat. Activities ofintracellular antioxidant enzymes, such as superoxide dis-mutase and catalase, were higher in microgravity than incontrol samples for the experimental time (7 days). However,the contents of glutathione decreased in comparison with thecontrol samples. These results indicate a protective mecha-nism that plays an important role for algal cells to adapt togravitational stress (Li et al., 2004). The same hypothesis canbe considered for X. laevis embryos, with an induction ofantioxidant enzymes related to GSH synthesis and utilization.
Lipid peroxidation (LP) and the activity of antioxidantenzyme superoxide dismutase have been studied in chloro-plasts of Pisum sativum under clinorotation (for 7 and 14days). In comparison with the control, increased LP levels inchloroplasts were established during both the terms of cli-norotation, and SOD activity rose in the chloroplasts of plantsclinorotated for 7 days (Baranenko, 2001).
Since there was no difference between basal controls (CTRB) and basal RPM (RPM B) samples, we did not find lipidperoxidation induced by microgravity, probably due to theearly activation of antioxidant systems of X.l. embryos,recorded after 3 days of microgravity exposure.
It is worth noting that manned space flight has been associ-ated with increased oxidative stress after return to 1 g (Stein andLeskiw, 2000). Effects on oxidative state are very relevant forlong missions lasting for several weeks, which might be due toan increased oxidative metabolism for muscle after return to 1 g,and/or to an adaptive mechanism of erythropoiesis during flightthat leads to an altered iron storage (Smith, 2002). In the longterm, high iron availability raises concern since iron promotesoxidative stress through ROS production via Fenton’s reaction.
Our results suggest that microgravity provided by RPMmight break the oxidative/antioxidative balance. Duringdevelopment, embryos have protective mechanisms thatincrease the total antioxidant system activity, and might playan important role to adapt cells to gravity deprivation. Nutri-tional countermeasures before, during and particularly afterspace flight might reinforce these systems and cooperate toreduce ROS negative side effects (Guan et al., 2004; Kennedyet al., 2004; Rabin et al., 2005).
Acknowledgments
Italian Space Agency (ASI) with ‘‘MoMa’’ Project,contract no. I/014/06/0 line ASSC to Angela M. Rizzo,granted this work.
References
Aebi H. Catalase in vitro. Meth Enzymol 1984;105:121e6.
Anderson D. Antioxidant defences against reactive oxygen species causing
genetic and other damages. Mutat Res 1996;350:103e8.
Baranenko VV. Effect of clinorotation on the intensity of lipid peroxidation
and superoxide dismutase activity in pea chloroplasts. Tsitologiia 2001;43:
1020e3 [in Russian].
Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing.
Nature 2000;408:239e47.
Franco R, Schoneveld OJ, Pappa A, Panayiotidis MI. The central role of
glutathione in pathophysiology of human diseases. Arch Physiol Biochem
2007;133:234e58.
Griffith OW. Glutathione and glutathione disulphide. In: Bergmeyer HU,
editor. Methods of enzymatic analysis, vol. VIII. New York: Academic
Press; 1985. p. 521e9.
Guan J, Wan XS, Zhou Z, Ware J, Donahue JJ, Biaglow JE, et al. Effects of
dietary supplements on space radiation-induced oxidative stress in Spra-
gue-Dawley rats. Radiat Res 2004;162:572e9.
Guan J, Stewart JH, Ware JH, Zhou Z, Donahue JJ, Kennedy AR. Effects of
dietary supplements on the space radiation-induced reduction in total
antioxidant status in CSA mice. Radiat Res 2006;165:373e8.
Hayes JD, Mc Lellan LI. Glutathione and glutathioneedependent enzymes
represent a co-ordinately regulated defence against oxidative stress. Free
Radic Res 1999;31:273e300.
Hollander J, Gore M, Fiebig R, Mazzeo R, Ohisi S, Ohno H, et al. Spaceflight
downregulates antioxidant defense systems in rat liver. Free Radic Biol
Med 1998;24:385e90.
Horn ER. Microgravity-induced modifications of the vestibuloocular reflex in
Xenopus laevis tadpoles are related to development and the occurrence of
tail lordosis. J Exp Biol 2006;209:2847e58.
Hoson T, Kamisaka S, Masuda Y, Yamashita M, Buchen B. Evaluation of the
three dimensional clinostat as a simulator of weightlessness. Planta 1997;
203:S187e97.
Kennedy AR, Ware JH, Guan J, Donahue JJ, Biaglow JE, Zhou Z, et al.
Selenomethionine protects against adverse biological effects induced by
space radiation. Free Radic Biol Med 2004;36:259e66.
Li GB, Liu YD, Wang GH, Song LR. Reactive oxygen species and antioxidant
enzymes activity of Anabaena sp. PCC 7120 (Cyanobacterium) under
simulated microgravity. Acta Astronaut 2004;55:953e7.
Liu Y, Wang E. Transcriptional analysis of normal human fibroblast responses to
microgravity stress. Genomics Proteomics Bioinformatics 2008;6:29e41.
Lyng FM, Seymour CB, Mothersill C. Oxidative stress in cells exposed to low
levels of ionizing radiation. Biochem Soc Trans 2001;29:350e3.
Manti L. Does reduced gravity alter cellular response to ionizing radiation?
Radiat Environ Biophys 2006;45:1e8.
Orr WC, Sohal RS. Extension of life-span by overexpression of superoxide dis-
mutase and catalase in Drosophila melanogaster. Science 1994;263:1128e30.
Paoletti F, Mocali A. Determination of superoxide dismutase activity by purely
chemical system based on NADPH oxidation. Meth Enzymol 1990;186:
209e20.
Peterson GM. A simplification of the protein assay method of Lowry et al.
which is more general applicable. Anal Biochem 1977;83:346e56.
Pinto C, Mata A, Lopez Barea J. Reversible inactivation of SaccharomycesCervisiae glutathione reductase under reducing conditions. Arch Biochem
Biophys 1984;228:1e12.
Prohaska JR, Ganther HE. Selenium and glutathione peroxidase in developing
rat. J Neurochem 1976;27:1379e87.
Rabin BM, Shukitt-Hale B, Joseph J, Todd P. Diet as a factor in behavioral
radiation protection following exposure to heavy particles. Gravit Space
Biol Bull 2005;18:71e7.
Rizzo AM, Gornati R, Galli C, Bernardini G, Berra B. Cholesterol, tri-
acylglicerols and phospholipids during Xenopus embryo development.
Cell Biol Int 1994;18:1085e90.
Rizzo AM, Rossi F, Zava S, Montorfano G, Adorni L, Cotronei V, et al.
Antioxidant metabolisms of Xenopus laevis embryos is affected by
stratospheric balloon flight. Cell Biol Int 2007a;31:716e23.
Rizzo AM, Montorfano G, Adorni L, Berra B. Antioxidant metabolism of
Xenopus laevis during the first days of development. Comp Biochem
Physiol B Biochem Mol Biol 2007b;146:94e100.
Sarkar P, Sarkar S, Ramesh V, Kim H, Barnes S, Kulkarni A, et al. Proteomic
analysis of mouse hypothalamus under simulated microgravity. Neuro-
chem Res 2008;33:2335e41.

898 A.M. Rizzo et al. / Cell Biology International 33 (2009) 893e898
Sebastian C, Esseling K, Horn E. Altered gravitational experience during early
periods of life affects the static vestibule-ocular reflex of tadpoles of the
southern clawned toad, Xenopus laevis. Exp Brain Res 1996;112:213e22.
Smith SM. Red blood cell and iron metabolism durinf space flight. Nutrition
2002;18:864e6.
Snetkova E, Chelnaya N, Serova L, Saveliev S, Chardanzova E, Pronych S,
et al. Effects of space flight on Xenopus laevis larval development. J Exp
Zool 1995;273:21e32.
Stein TP. Space flight and oxidative stress. Nutrition 2002;18:867e71.
Stein SJ, Leskiw MJ. Oxidant damage during and after space flight. Am J
Physiol Endocrinol Metabol 2000;278. pp. E375eE382.
Thorpe GW, Fong CS, Alic N, Higgins VJ, Dawes IW. Cells have distinct
mechanisms to maintain protection against different reactive oxygen
species: oxidative-stress-response genes. Proc Natl Acad Sci U S A 2004;
101:6564e9.
Valko M, Liebfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free
radicals and antioxidants in normal physiological functions and human
disease. Int J Biochem Cell Biol 2007;39:44e84.
van Loon JJWA. Some history and use of the random positioning
machine, RPM, in gravity related research. Adv Space Res 2007;39:
1161e5.
Wey HE, Pyron L, Woolery M. Essential fatty acid deficiency in cultured
human keratinocytes attenuates toxicity due to lipid peroxidation. Toxicol
Appl Pharmacol 1993;120:72e9.
Yu BP. Cellular defences against damage from reactive oxygen species.
Physiol Rev 1994;74:139e62.