tsuji-et-al-2013.pdf - smu
TRANSCRIPT

This article was downloaded by: [76.187.62.88]On: 16 May 2014, At: 23:11Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Vertebrate PaleontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ujvp20
The vertebrate fauna of the Upper Permian ofNiger—VII. Cranial anatomy and relationships ofBunostegos akokanensis (Pareiasauria)Linda A. Tsuji a , Christian A. Sidor a , J.- Sébastien Steyer b , Roger M. H. Smith c , Neil J.Tabor d & Oumarou Ide ea Burke Museum and Department of Biology , University of Washington , Seattle ,Washington , 98195 , U.S.A.b Bâtiment de Paléontologie, UMR 7207 CNRS, Département Histoire de la Terre, CP38 ,Muséum national d’Histoire naturelle , F-75005 , Paris , Francec Karoo Palaeontology , Iziko South African Museum , PO Box 61, Cape Town , 8000 , SouthAfricad Roy M. Huffington Department of Earth Sciences , Southern Methodist University , Dallas ,Texas , 75275 , U.S.A.e Institut de Recherches en Sciences Humaines , Université de Niamey , Republique du NigerPublished online: 25 Jun 2013.
To cite this article: Linda A. Tsuji , Christian A. Sidor , J.- Sébastien Steyer , Roger M. H. Smith , Neil J. Tabor & Oumarou Ide(2013) The vertebrate fauna of the Upper Permian of Niger—VII. Cranial anatomy and relationships of Bunostegos akokanensis(Pareiasauria), Journal of Vertebrate Paleontology, 33:4, 747-763, DOI: 10.1080/02724634.2013.739537
To link to this article: http://dx.doi.org/10.1080/02724634.2013.739537
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Vertebrate Paleontology 33(4):747–763, July 2013© 2013 by the Society of Vertebrate Paleontology
FEATURED ARTICLE
THE VERTEBRATE FAUNA OF THE UPPER PERMIAN OF NIGER—VII. CRANIAL ANATOMYAND RELATIONSHIPS OF BUNOSTEGOS AKOKANENSIS (PAREIASAURIA)
LINDA A. TSUJI,1 CHRISTIAN A. SIDOR,*,1 J.- SEBASTIEN STEYER,2 ROGER M. H. SMITH,3 NEIL J. TABOR,4
and OUMAROU IDE5
1Burke Museum and Department of Biology, University of Washington, Seattle, Washington 98195, U.S.A., [email protected],[email protected];
2Batiment de Paleontologie, UMR 7207 CNRS, Departement Histoire de la Terre, CP38, Museum national d’Histoire naturelle,F-75005, Paris, France, [email protected];
3Karoo Palaeontology, Iziko South African Museum, PO Box 61, Cape Town 8000, South Africa, [email protected];4Roy M. Huffington Department of Earth Sciences, Southern Methodist University, Dallas, Texas 75275, U.S.A., [email protected];
5Institut de Recherches en Sciences Humaines, Universite de Niamey, Republique du Niger, [email protected]
ABSTRACT—We describe newly recovered cranial material of Bunostegos akokanensis, a pareiasaurian reptile known fromthe Upper Permian Moradi Formation of northern Niger. Bunostegos is highly autapomorphic, with diagnostic cranial fea-tures including two or three hemispherical bosses located above and between the external nares; laterally projecting supraor-bital ‘horn’ formed by an enlarged postfrontal; large foramen present on ventral surface of postfrontal; and hemisphericalsupratemporal boss located at posterolateral corner of skull roof. We addressed the phylogenetic position of Bunostegos byincorporating it into a cladistic analysis of 29 parareptilian taxa (including all 21 currently valid pareiasaurs) and 127 cranialand postcranial characters. The results of this analysis place Bunostegos as more derived than middle Permian forms such asBradysaurus and as the sister taxon to the clade including Deltavjatia plus Velosauria. Certain characters, such as the patternof cranial ornamentation and the size and placement of the tabulars, appear to be more similar to more derived pareiasaurssuch as Elginia from Scotland and Arganaceras from Morocco, but the most parsimonious tree topology indicates that thesefeatures were evolved independently in the Nigerien form. The lack of both dicynodont herbivores and Glossopteris, com-bined with the presence of a giant herbivorous captorhinid, indicates a markedly different community structure in the Permianof Niger compared with those for contemporaneous southern Pangean basins (i.e., Karoo, Luangwa, Ruhuhu). The endemictetrapod fauna of Niger supports the theory that central Pangea was biogeographically isolated from the rest of the supercon-tinent by desert-like conditions during Late Permian times.
SUPPLEMENTAL DATA—Supplemental materials are available for this article for free at www.tandfonline.com/UJVP
INTRODUCTION
Pareiasaurs were a moderately diverse clade of parareptiles re-stricted to the middle and Upper Permian, but with a near-globaldistribution (Lee, 1997a). Historically, most of the group’s diver-sity has come from the Beaufort Group of South Africa (10 of21 named species; Lee, 1997a), but recent work has extended thegeographic range of African pareiasaurs northward (Sidor et al.,2003; Jalil and Janvier, 2005). Sidor et al. (2003) named Bunos-tegos akokanensis as the first pareiasaur from Niger on the basisof a weathered skull lacking lower jaws that was collected fromthe Upper Permian Moradi Formation in 2000. Despite the lackof surface detail, the holotype was clearly diagnosable as a newpareiasaur genus by virtue of its three enlarged supranarial bossesand paired supraorbital and supratemporal ‘horns.’
Additional fossils were collected from the Moradi Formationin 2003 and 2006 (Fig. 1). These include amphibian material de-scribed by Sidor et al. (2005b) as the edopoid, Saharastega mora-diensis, and the derived cochleosaurid, Nigerpeton ricqlesi (seealso Damiani et al., 2006, Steyer et al., 2006). Reptilian fossils in-cluded juveniles and subadults of the large captorhinid Moradis-aurus grandis (O’Keefe et al., 2005, 2006), as well as undescribed
*Corresponding author.
remains of a possible second, smaller captorhinid taxon (Sidoret al., 2005a). In addition, Smiley et al. (2008) described the firsttherapsid from the formation as an indeterminate rubidgine gor-gonopsid and noted that previous reports (Taquet, 1967, 1978;de Ricqles and Taquet, 1982) suggesting the presence of a dicyn-odont were in error.
Pareiasaur fossils were found to be relatively common in theMoradi Formation, with finds in 2003 and 2006 ranging fromisolated elements to partially articulated skeletons with axialand appendicular material. The collected elements span a widerange of body sizes, suggesting the possibility that more than onepareiasaur species coexisted in the Moradi fauna. Here we de-scribe cranial material referable to Bunostegos that provides crit-ical information on the phylogenetic position of this remarkabletaxon. Our cladistic analysis builds on that of Tsuji (2006, 2010, inpress) and results in phylogenetic support for an emerging pictureof central Pangean endemism during Late Permian times (Sidoret al., 2005b; Tabor et al., 2011).
Institutional Abbreviations—BP, Bernard Price Institute forPalaeontological Research, University of the Witwatersrand,Johannesburg; MB, Museum fur Naturkunde, Humboldt Uni-versitat zu Berlin, Berlin; MNHN, Museum national d’Histoirenaturelle, Paris; MNN, Musee National du Niger, Niamey; PIN,Paleontological Institute, Moscow; SMNS, Staatliches Museumfur Naturkunde, Stuttgart.
747
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

748 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
FIGURE 1. Geographic position of the study area and stratigraphy ofthe Permian Izegouandane Group of Niger. Filled area within countryoutline of Niger indicates mapped extent of the Izegouandane Group.Abbreviation: Fm., Formation.
BACKGROUND
Pareiasaurs are known exclusively from middle–Upper Per-mian rocks, with the bulk of the group’s diversity concen-trated in Africa. The most recent taxonomic revision recognizeseight genera from the Beaufort Group of South Africa (viz.Anthodon, Bradysaurus, Embrithosaurus, Nanoparia, Nochele-saurus, Pareiasaurus, Pareiasuchus, Pumiliopareia), but some ofthese genera are also known from Karoo-equivalent strata acrosssouthern Africa (Lee, 1997b). For example, Pareiasuchus is alsoknown from the upper Madumabisa Mudstone Formation ofZambia (Lee et al., 1997), and Anthodon and Pareiasaurus arerecorded from the Usili Formation of Tanzania (Sidor et al.,2010). Besides Bunostegos from Niger, other African pareiasaursinclude indeterminate material from Malawi (Huene, 1944; Lee,1997b) as well as the recently described Moroccan form, Ar-ganaceras vacanti (Jalil and Janvier, 2005). It should be notedthat Jalil and Janvier (2005) recognized two distinct morphs ofpostcranial material from the same level of the Ikakern Forma-tion as Arganaceras. However, they noted that none of this ma-terial could be unambiguously assigned to the named taxon (Jaliland Janvier, 2005).
The South American pareiasaur fossil record comes from theRio do Rasto Formation of Brazil and is restricted to a fewreasonably complete specimens. These bones were initially de-scribed as ‘Pareiasaurus’ americanus by Araujo (1985), but thistaxon was later transferred to a new genus, Provelosaurus, by Lee(1997b) in order to keep the genus Pareiasaurus monophyletic.The latter assignment was upheld by Cisneros et al. (2005).
Laurasian pareiasaurs are recorded from China (Sanchuan-saurus, Shansisaurus, and Shihtienfenia), Germany (Parasaurus),Russia (Deltavjatia, Scutosaurus, and Obirkovia), and Scotland(Elginia). Of these, the Russian forms are the best understood,each being known by several skulls and partial skeletons (Lee,2000; Tsuji, 2010, in press). Elginia, a highly autapomorphic form
known only from natural molds (Newton, 1893; Spencer and Lee,2000), has attracted recent attention because of its unique cra-nial anatomy and superficial similarity to Arganaceras (Jalil andJanvier, 2005).
MATERIAL
Our description is based primarily on three specimens. Thefirst, MNN MOR86, comprises a partial skull lacking its lowerjaws. The second specimen, MNN MOR28, consists of a skull thatis three-dimensionally preserved, but highly eroded on its dorsalsurface. The third specimen, MNN MOR47, preserves the brain-case including the sphenethmoid in left lateral view, along withpart of the palate. Five additional specimens including identifi-able cranial material are described where relevant.
The following specimens were studied for comparative pur-poses: MB R.939 (skull and anterior cervical vertebrae of Pareia-suchus peringueyi); MNHN ARG 518 (disarticulated cranial re-mains of Arganaceras vacanti); PIN 2212/6 (skull and lower jawof a subadult Deltavjatia vjatkensis); and SMNS 58317 (skull andanterior skeleton of a subadult Deltavjatia vjatkensis).
SYSTEMATIC PALEONTOLOGY
PAREIASAURIA Seeley, 1888BUNOSTEGOS AKOKANENSIS Sidor, Blackburn, and Gado,
2003(Figs. 2–9)
Holotype—MNN MOR72, ventrally eroded skull lackinglower jaws.
Referred Material—MNN MOR25, eroded braincase andpalate; MNN MOR28, weathered skull; MNN MOR35, fragmen-tary skull roof elements; MNN MOR37, partial skull includingsnout, interorbital region, and posterior margin of skull table;MNN MOR47, partial skull with braincase exposed in left lat-eral view and associated cervical vertebra; MNN MOR86, nearlycomplete skull, right scapulocoracoid, and two dorsal vertebrae;MNN MOR101, isolated left postfrontal, right scapulocoracoid,right innominate, and dorsal vertebra; MNN MOR104, isolatednasals, interorbital skull roof, right ulna, isolated neural spine,and isolated osteoderm.
Locality and Horizon—The specimens described here werecollected from several localities located approximately 20 kmwest of Arlit, Agadez Department, northern Niger (Fig. 1). Thelocalities are within 1 km of each other and all fall within theupper one-third of the Moradi Formation. Detailed locality dataare available to qualified researchers at the MNN or by contact-ing C.A.S. The Moradi Formation is typically considered to beLate Permian in age, although the data supporting such an as-signment are not overwhelming (Taquet, 1972, 1976; Sidor et al.,2005b; Tabor et al., 2011).
Revised Diagnosis—Medium-sized pareiasaur with two orthree hemispherical bosses located at the anterior end of thesnout; nasal with posterolateral tab-like process articulating withthe frontal; frontal and parietal lack central bosses; elongate, lat-erally projecting supraorbital ‘horn’ formed by postfrontal andoverhanging orbit in dorsal view; large foramen present on ven-tral surface of postfrontal; hemispherical supratemporal boss lo-cated at posterolateral corner of skull roof; supraorbital ‘horn’and supratemporal bosses with neck separating globular headfrom skull roof; postorbital extends more posteriorly than pari-etal on skull roof; pineal foramen equidistant from frontoparietaland parietal-postparietal sutures.
DESCRIPTION
Skull Roof
Unless noted, the following description is based on the mostcomplete skull (MNN MOR86; Figs. 2, 3), with reference made toa referred skull (MNN MOR28; Fig. 4) and the holotype (MNN
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
TSUJI ET AL.—PAREIASAUR FROM NIGER 749
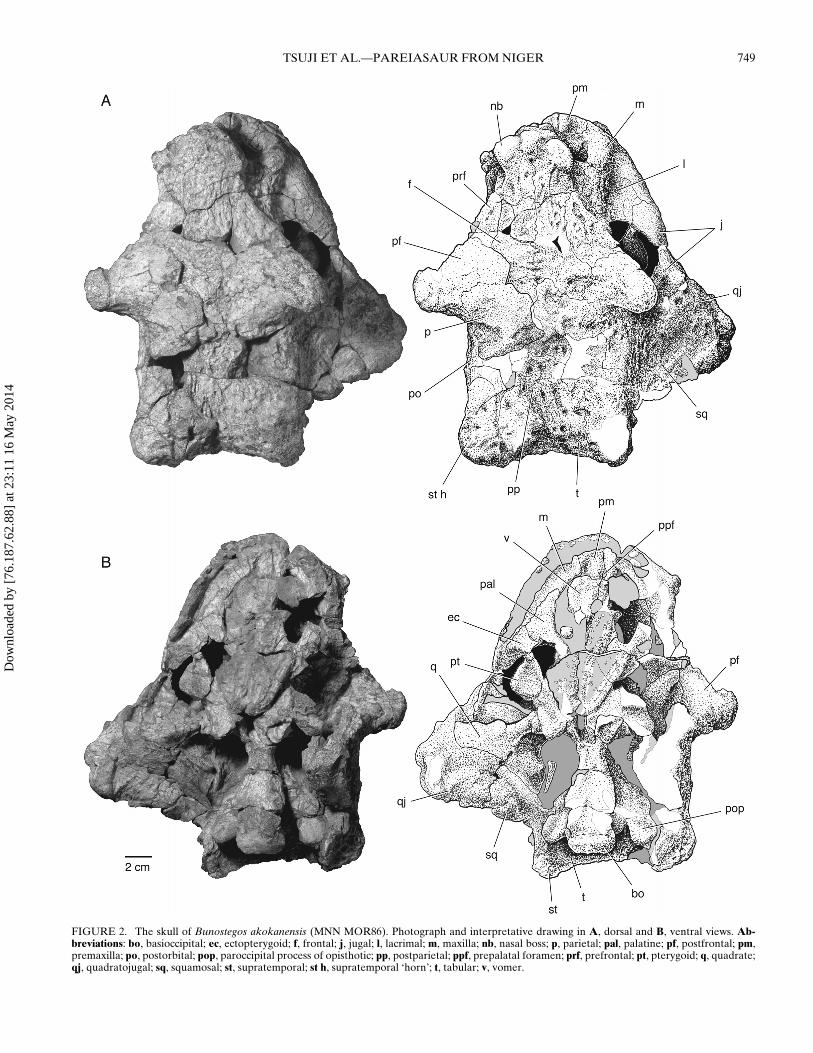
FIGURE 2. The skull of Bunostegos akokanensis (MNN MOR86). Photograph and interpretative drawing in A, dorsal and B, ventral views. Ab-breviations: bo, basioccipital; ec, ectopterygoid; f, frontal; j, jugal; l, lacrimal; m, maxilla; nb, nasal boss; p, parietal; pal, palatine; pf, postfrontal; pm,premaxilla; po, postorbital; pop, paroccipital process of opisthotic; pp, postparietal; ppf, prepalatal foramen; prf, prefrontal; pt, pterygoid; q, quadrate;qj, quadratojugal; sq, squamosal; st, supratemporal; st h, supratemporal ‘horn’; t, tabular; v, vomer.
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
750 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
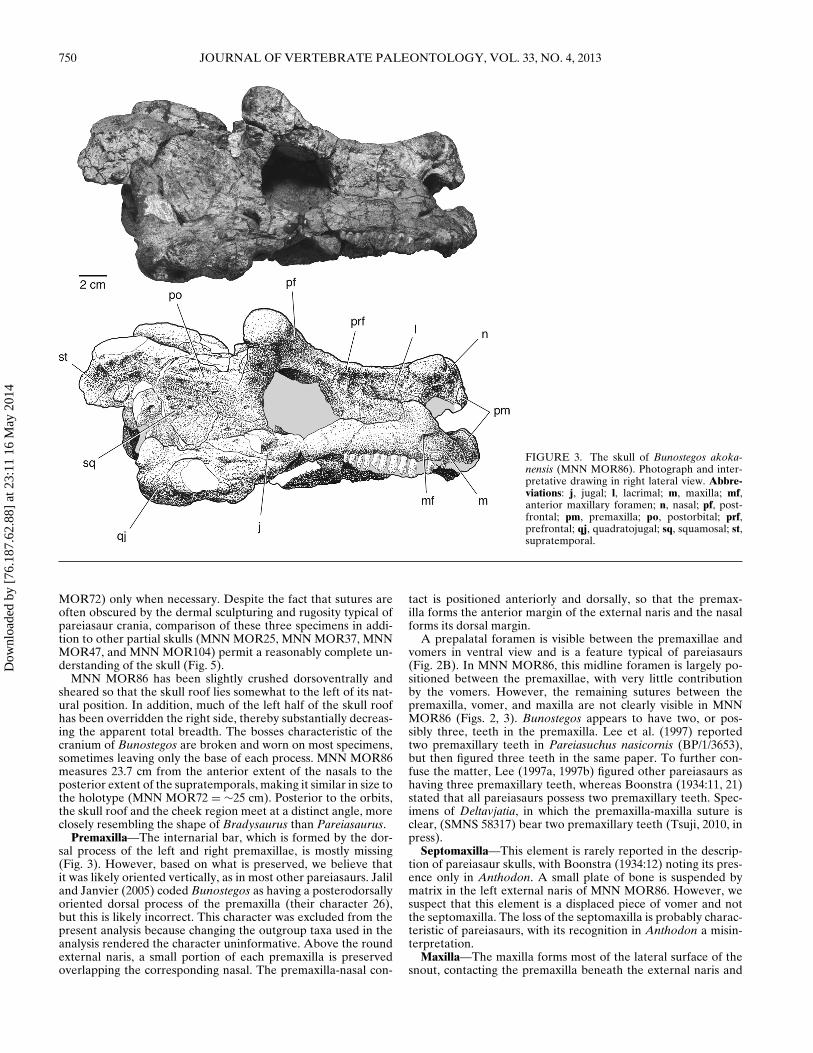
FIGURE 3. The skull of Bunostegos akoka-nensis (MNN MOR86). Photograph and inter-pretative drawing in right lateral view. Abbre-viations: j, jugal; l, lacrimal; m, maxilla; mf,anterior maxillary foramen; n, nasal; pf, post-frontal; pm, premaxilla; po, postorbital; prf,prefrontal; qj, quadratojugal; sq, squamosal; st,supratemporal.
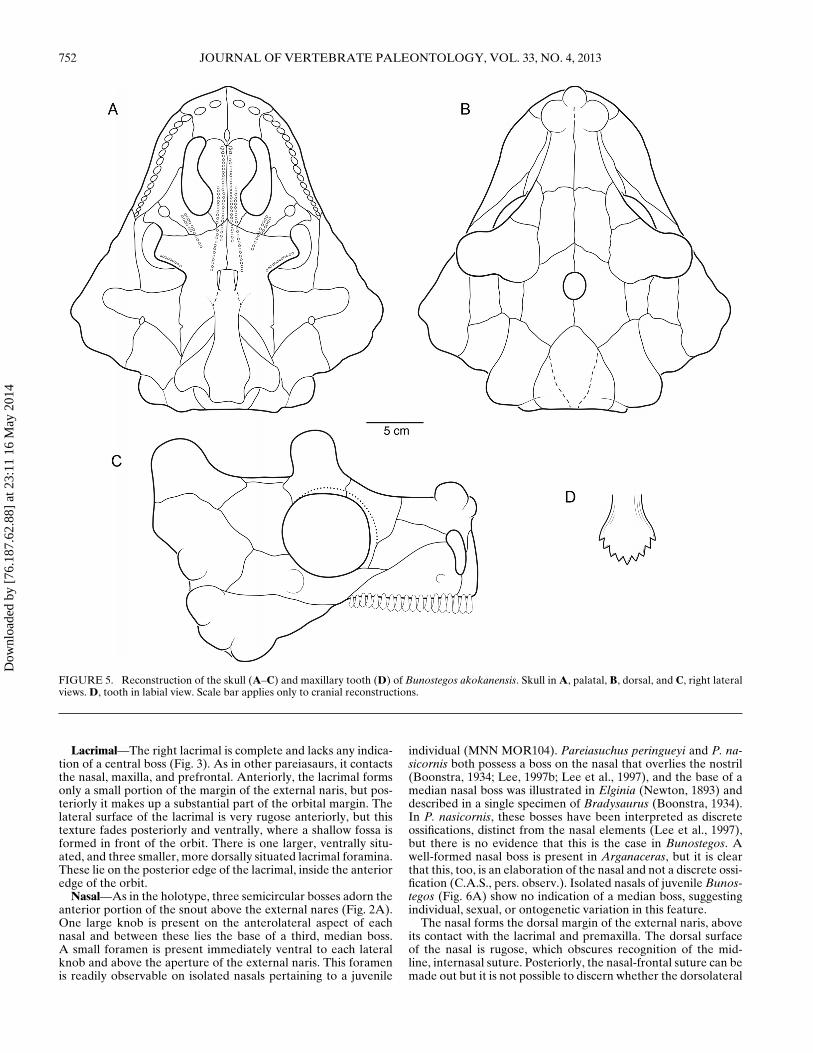
MOR72) only when necessary. Despite the fact that sutures areoften obscured by the dermal sculpturing and rugosity typical ofpareiasaur crania, comparison of these three specimens in addi-tion to other partial skulls (MNN MOR25, MNN MOR37, MNNMOR47, and MNN MOR104) permit a reasonably complete un-derstanding of the skull (Fig. 5).
MNN MOR86 has been slightly crushed dorsoventrally andsheared so that the skull roof lies somewhat to the left of its nat-ural position. In addition, much of the left half of the skull roofhas been overridden the right side, thereby substantially decreas-ing the apparent total breadth. The bosses characteristic of thecranium of Bunostegos are broken and worn on most specimens,sometimes leaving only the base of each process. MNN MOR86measures 23.7 cm from the anterior extent of the nasals to theposterior extent of the supratemporals, making it similar in size tothe holotype (MNN MOR72 = ∼25 cm). Posterior to the orbits,the skull roof and the cheek region meet at a distinct angle, moreclosely resembling the shape of Bradysaurus than Pareiasaurus.
Premaxilla—The internarial bar, which is formed by the dor-sal process of the left and right premaxillae, is mostly missing(Fig. 3). However, based on what is preserved, we believe thatit was likely oriented vertically, as in most other pareiasaurs. Jaliland Janvier (2005) coded Bunostegos as having a posterodorsallyoriented dorsal process of the premaxilla (their character 26),but this is likely incorrect. This character was excluded from thepresent analysis because changing the outgroup taxa used in theanalysis rendered the character uninformative. Above the roundexternal naris, a small portion of each premaxilla is preservedoverlapping the corresponding nasal. The premaxilla-nasal con-
tact is positioned anteriorly and dorsally, so that the premax-illa forms the anterior margin of the external naris and the nasalforms its dorsal margin.
A prepalatal foramen is visible between the premaxillae andvomers in ventral view and is a feature typical of pareiasaurs(Fig. 2B). In MNN MOR86, this midline foramen is largely po-sitioned between the premaxillae, with very little contributionby the vomers. However, the remaining sutures between thepremaxilla, vomer, and maxilla are not clearly visible in MNNMOR86 (Figs. 2, 3). Bunostegos appears to have two, or pos-sibly three, teeth in the premaxilla. Lee et al. (1997) reportedtwo premaxillary teeth in Pareiasuchus nasicornis (BP/1/3653),but then figured three teeth in the same paper. To further con-fuse the matter, Lee (1997a, 1997b) figured other pareiasaurs ashaving three premaxillary teeth, whereas Boonstra (1934:11, 21)stated that all pareiasaurs possess two premaxillary teeth. Spec-imens of Deltavjatia, in which the premaxilla-maxilla suture isclear, (SMNS 58317) bear two premaxillary teeth (Tsuji, 2010, inpress).
Septomaxilla—This element is rarely reported in the descrip-tion of pareiasaur skulls, with Boonstra (1934:12) noting its pres-ence only in Anthodon. A small plate of bone is suspended bymatrix in the left external naris of MNN MOR86. However, wesuspect that this element is a displaced piece of vomer and notthe septomaxilla. The loss of the septomaxilla is probably charac-teristic of pareiasaurs, with its recognition in Anthodon a misin-terpretation.
Maxilla—The maxilla forms most of the lateral surface of thesnout, contacting the premaxilla beneath the external naris and
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
TSUJI ET AL.—PAREIASAUR FROM NIGER 751
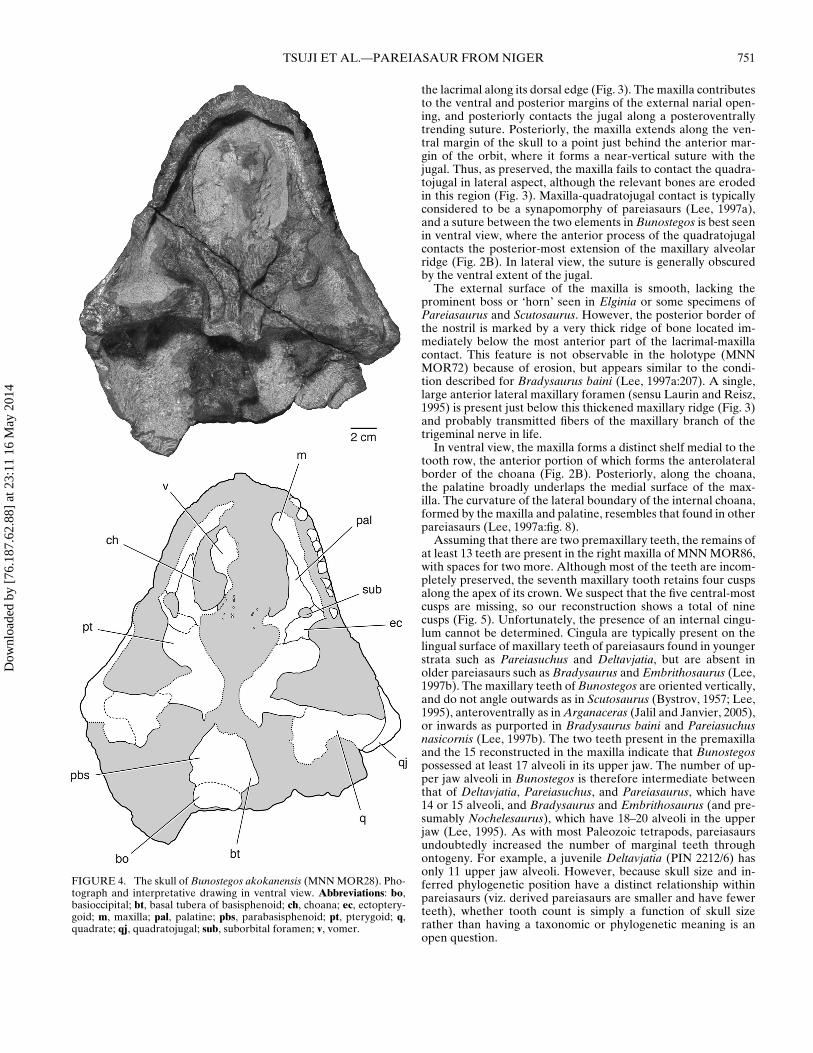
FIGURE 4. The skull of Bunostegos akokanensis (MNN MOR28). Pho-tograph and interpretative drawing in ventral view. Abbreviations: bo,basioccipital; bt, basal tubera of basisphenoid; ch, choana; ec, ectoptery-goid; m, maxilla; pal, palatine; pbs, parabasisphenoid; pt, pterygoid; q,quadrate; qj, quadratojugal; sub, suborbital foramen; v, vomer.
the lacrimal along its dorsal edge (Fig. 3). The maxilla contributesto the ventral and posterior margins of the external narial open-ing, and posteriorly contacts the jugal along a posteroventrallytrending suture. Posteriorly, the maxilla extends along the ven-tral margin of the skull to a point just behind the anterior mar-gin of the orbit, where it forms a near-vertical suture with thejugal. Thus, as preserved, the maxilla fails to contact the quadra-tojugal in lateral aspect, although the relevant bones are erodedin this region (Fig. 3). Maxilla-quadratojugal contact is typicallyconsidered to be a synapomorphy of pareiasaurs (Lee, 1997a),and a suture between the two elements in Bunostegos is best seenin ventral view, where the anterior process of the quadratojugalcontacts the posterior-most extension of the maxillary alveolarridge (Fig. 2B). In lateral view, the suture is generally obscuredby the ventral extent of the jugal.
The external surface of the maxilla is smooth, lacking theprominent boss or ‘horn’ seen in Elginia or some specimens ofPareiasaurus and Scutosaurus. However, the posterior border ofthe nostril is marked by a very thick ridge of bone located im-mediately below the most anterior part of the lacrimal-maxillacontact. This feature is not observable in the holotype (MNNMOR72) because of erosion, but appears similar to the condi-tion described for Bradysaurus baini (Lee, 1997a:207). A single,large anterior lateral maxillary foramen (sensu Laurin and Reisz,1995) is present just below this thickened maxillary ridge (Fig. 3)and probably transmitted fibers of the maxillary branch of thetrigeminal nerve in life.
In ventral view, the maxilla forms a distinct shelf medial to thetooth row, the anterior portion of which forms the anterolateralborder of the choana (Fig. 2B). Posteriorly, along the choana,the palatine broadly underlaps the medial surface of the max-illa. The curvature of the lateral boundary of the internal choana,formed by the maxilla and palatine, resembles that found in otherpareiasaurs (Lee, 1997a:fig. 8).
Assuming that there are two premaxillary teeth, the remains ofat least 13 teeth are present in the right maxilla of MNN MOR86,with spaces for two more. Although most of the teeth are incom-pletely preserved, the seventh maxillary tooth retains four cuspsalong the apex of its crown. We suspect that the five central-mostcusps are missing, so our reconstruction shows a total of ninecusps (Fig. 5). Unfortunately, the presence of an internal cingu-lum cannot be determined. Cingula are typically present on thelingual surface of maxillary teeth of pareiasaurs found in youngerstrata such as Pareiasuchus and Deltavjatia, but are absent inolder pareiasaurs such as Bradysaurus and Embrithosaurus (Lee,1997b). The maxillary teeth of Bunostegos are oriented vertically,and do not angle outwards as in Scutosaurus (Bystrov, 1957; Lee,1995), anteroventrally as in Arganaceras (Jalil and Janvier, 2005),or inwards as purported in Bradysaurus baini and Pareiasuchusnasicornis (Lee, 1997b). The two teeth present in the premaxillaand the 15 reconstructed in the maxilla indicate that Bunostegospossessed at least 17 alveoli in its upper jaw. The number of up-per jaw alveoli in Bunostegos is therefore intermediate betweenthat of Deltavjatia, Pareiasuchus, and Pareiasaurus, which have14 or 15 alveoli, and Bradysaurus and Embrithosaurus (and pre-sumably Nochelesaurus), which have 18–20 alveoli in the upperjaw (Lee, 1995). As with most Paleozoic tetrapods, pareiasaursundoubtedly increased the number of marginal teeth throughontogeny. For example, a juvenile Deltavjatia (PIN 2212/6) hasonly 11 upper jaw alveoli. However, because skull size and in-ferred phylogenetic position have a distinct relationship withinpareiasaurs (viz. derived pareiasaurs are smaller and have fewerteeth), whether tooth count is simply a function of skull sizerather than having a taxonomic or phylogenetic meaning is anopen question.
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
752 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
FIGURE 5. Reconstruction of the skull (A–C) and maxillary tooth (D) of Bunostegos akokanensis. Skull in A, palatal, B, dorsal, and C, right lateralviews. D, tooth in labial view. Scale bar applies only to cranial reconstructions.
Lacrimal—The right lacrimal is complete and lacks any indica-tion of a central boss (Fig. 3). As in other pareiasaurs, it contactsthe nasal, maxilla, and prefrontal. Anteriorly, the lacrimal formsonly a small portion of the margin of the external naris, but pos-teriorly it makes up a substantial part of the orbital margin. Thelateral surface of the lacrimal is very rugose anteriorly, but thistexture fades posteriorly and ventrally, where a shallow fossa isformed in front of the orbit. There is one larger, ventrally situ-ated, and three smaller, more dorsally situated lacrimal foramina.These lie on the posterior edge of the lacrimal, inside the anterioredge of the orbit.
Nasal—As in the holotype, three semicircular bosses adorn theanterior portion of the snout above the external nares (Fig. 2A).One large knob is present on the anterolateral aspect of eachnasal and between these lies the base of a third, median boss.A small foramen is present immediately ventral to each lateralknob and above the aperture of the external naris. This foramenis readily observable on isolated nasals pertaining to a juvenile
individual (MNN MOR104). Pareiasuchus peringueyi and P. na-sicornis both possess a boss on the nasal that overlies the nostril(Boonstra, 1934; Lee, 1997b; Lee et al., 1997), and the base of amedian nasal boss was illustrated in Elginia (Newton, 1893) anddescribed in a single specimen of Bradysaurus (Boonstra, 1934).In P. nasicornis, these bosses have been interpreted as discreteossifications, distinct from the nasal elements (Lee et al., 1997),but there is no evidence that this is the case in Bunostegos. Awell-formed nasal boss is present in Arganaceras, but it is clearthat this, too, is an elaboration of the nasal and not a discrete ossi-fication (C.A.S., pers. observ.). Isolated nasals of juvenile Bunos-tegos (Fig. 6A) show no indication of a median boss, suggestingindividual, sexual, or ontogenetic variation in this feature.
The nasal forms the dorsal margin of the external naris, aboveits contact with the lacrimal and premaxilla. The dorsal surfaceof the nasal is rugose, which obscures recognition of the mid-line, internasal suture. Posteriorly, the nasal-frontal suture can bemade out but it is not possible to discern whether the dorsolateral
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
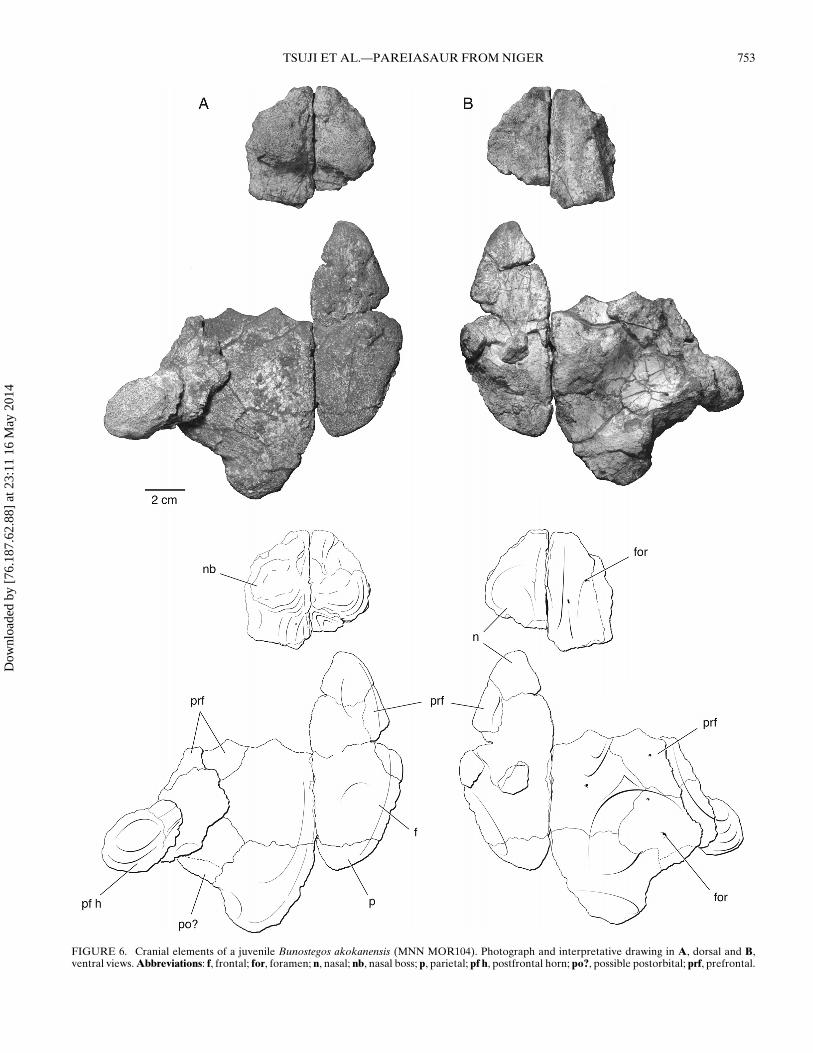
TSUJI ET AL.—PAREIASAUR FROM NIGER 753
FIGURE 6. Cranial elements of a juvenile Bunostegos akokanensis (MNN MOR104). Photograph and interpretative drawing in A, dorsal and B,ventral views. Abbreviations: f, frontal; for, foramen; n, nasal; nb, nasal boss; p, parietal; pf h, postfrontal horn; po?, possible postorbital; prf, prefrontal.
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

754 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
tab-like process of the posterior nasal, visible on the holotype, ispresent. The breadth of the nasals in Bunostegos gives the snoutan appearance that is broader than tall, similar to Pareiasuchusnasicornis (Lee et al., 1997), whereas in Arganaceras and Elginiathe snout appears taller than wide (Jalil and Janvier, 2005).
Prefrontal—The right prefrontal is more complete than theleft. It is clearly divided into dorsal and lateral faces by a thick-ened ridge of bone that extends anteriorly from the anterodorsalpart of the orbit to the lateral nasal boss. On its lateral surface,the prefrontal-lacrimal suture runs anteriorly and slightly dor-sally from the orbit to a position just posterior to the nasal boss(Fig. 3). Because a small part of the right orbital rim is missing,the frontal-prefrontal suture is visible in cross-section. It is diffi-cult to follow this suture dorsally on the skull roof, but from whatcan be made out, the prefrontal is restricted laterally (Fig. 2A).As in other pareiasaurs, the prefrontal contacts the postfrontal toexclude the frontal from contributing to the dorsal margin of theorbit. We suspect that Bunostegos either lacked a central bosson the prefrontal, which would be unusual among pareiasaurs,or, like Arganaceras, possessed a greatly reduced one. Unfortu-nately, the appropriate section of the orbital margin is missingon both sides of MNN MOR86 and is not available on any ofthe other referred skulls. The ventral process at the anterior edgeof the prefrontal forms a solid connection with the palate, run-ning just posterior to the lacrimal. This suture can best be seen inMNN MOR72, because this area of the palate is missing in MNNMOR86.
Frontal—The dorsal surface of the frontal is rugose, but lacksa central boss (Fig. 2A). In this character Bunostegos moreclosely resembles the condition seen in Arganaceras, Elginia, andPareiasaurus. It contacts the nasal, prefrontal, postfrontal, andparietal along sutures that correspond to the pattern seen in otherpareiasaurs. The element is relatively short in pareiasaurs com-pared with their outgroups, the nycteroleters, in which the frontalis a long, slim bone, more than four times as long as wide (Tsujiet al., 2012). The frontal-parietal suture is roughly perpendicularto the midline sutures of the frontals and parietals. This is slightlydifferent than the corresponding contact in the holotype (Sidoret al., 2003:fig. 2) in which the suture extends slightly anteriorlybefore proceeding directly lateral to the postfrontal. However,this difference may be due to the fact that the external surfaceof the holotype is severely weathered, such that the observed su-tures are all ‘deep’ to some degree.
Postfrontal—MNN MOR86, MOR101, and MOR104 all dis-play the diagnostic postfrontal morphology of Bunostegos; thiselement is very large and extends laterally to take the form ofa prominent, rounded ‘horn’ over the orbit. In contrast to otherpareiasaurs, the postfrontal in Bunostegos forms nearly the en-tire dorsal orbital border to obscure the orbit completely whenviewed from above. On all specimens where this area is acces-sible, a foramen can be seen to penetrate the ventral surfaceof the postfrontal (Fig. 6B). We suspect that this foramen car-ried an enriched blood supply to the supraorbital ‘horn,’ whichsuggests that it was covered by a keratinous sheath in life. Acorresponding, albeit smaller, foramen is present in Arganac-eras. Interestingly, although the skull of the Moroccan pareiasauris substantially larger than Bunostegos, its supraorbital horn issmaller, probably requiring a relatively smaller blood supply.Among pareiasaurs, an enlarged postfrontal ‘horn,’ or elongatepostfrontal boss, is also present in Elginia (Lee, 2000). A subadultDeltavjatia (SMNS 58317) has a correspondingly smaller boss,yet it is still clearly distinguishable from the surrounding dermalsculpturing.
Sutural contacts of the postfrontal are best seen on the rightside in MNN MOR86 (Figs. 2A, 3) and agree with those foundin the holotype (MNN MOR72) and MNN MOR104. In MNNMOR86, the right supraorbital ‘horn’ has been deformed up-wards, so that it does not extend as far laterally as in the holo-
type. Despite this deformation, this bone retains a shallow trans-versely oriented trough on its underside that continued later-ally under the main projection of the ‘horn.’ The left postfrontaland postorbital of MNN MOR86 have been translated medially.This movement is most apparent posteriorly, where the latter el-ements override the left frontal and parietal (Fig. 2A).
Postorbital—Considerably more information about the pos-torbital in Bunostegos can be added to the description of Sidoret al. (2003). On the right side, the postorbital can be seen to formthe dorsal half of posterior margin of the orbit (Fig. 3). Aboveit, the enlarged postfrontal forms the dorsal orbital margin. Thepostorbital-postfrontal contact can be seen along the posteriorbase of the right supraorbital ‘horn,’ until a triple junction withthe parietal is formed medially. The postorbital is sutured to thejugal ventrally, the latter forming the ventral half of the orbitalmargin. In frontal section, the postorbital is a curved plate form-ing a small horizontal contribution to the skull roof and a largervertical portion of the cheek. Posteriorly, the postorbital contactsthe squamosal and the supratemporal, and appears very long indorsal view (Fig. 2A). Bunostegos is unique among pareiasaurs inthat the postorbital extends farther posteriorly than the parietal.
Jugal—Aside from a small gap in the suborbital bar, nearlyall of the right jugal is present and well preserved. In lateralview, the jugal has a prominent anterior process that contactsthe maxilla and lacrimal at a level slightly in front of the orbit,and it has a well-developed ventral process projecting below thelevel of the tooth row (Fig. 3). Posteroventral to the orbit, thejugal forms a low-lying boss just dorsal of the anterior part ofthe jugal-quadratojugal suture. Contacts between the jugal andquadratojugal, the jugal and squamosal, and the jugal and pos-torbital are visible and correspond to the pattern commonly seenin pareiasaurs.
Squamosal—The squamosal is a large, rectangular elementforming the rear of the cheek above the quadratojugal in mostpareiasaurs. This element is mostly preserved on the right sidein MNN MOR86 (Fig. 3). The external surface of the squamosalbears small pits and relatively low furrows. The squamosal-jugalcontact is complete and angled dorsally and slightly anteriorlyto intersection of these two bones and the postorbital. Abovethis, the squamosal contacts the postorbital along a dorsally andslightly posteriorly oriented suture. The contact between thesquamosal and supratemporal begins along the posterior marginof the skull, just beneath the base of the supratemporal ‘horn.’This suture passes slightly upwards anteriorly, before arcing ven-trally along the postorbital. As in several other pareiasaur genera,a small boss is present on the squamosal along its posterior edge(e.g., Jalil and Janvier, 2005:fig. 9).
Quadratojugal—This element is entirely missing from theholotype and is incompletely preserved in MNN MOR86.Nonetheless, some details of its anatomy in Bunostegos are nowpossible to evaluate. As in other pareiasaurs, the quadratojugalis a large element that forms the ventral margin of the skull be-hind the orbit, with an anterior process that extends along theventral edge of the jugal to contact the maxilla. Anteriorly, theexternal surface of the quadratojugal is relatively unornamentedwhere it contacts the jugal. The remains of at least two bossesare present on the ventral and posterior margins of this element(Fig. 3). These appear to have been approximately equal in sizeto the nasal knobs and slightly larger than those of the jugal orsquamosal.
Parietal—The right parietal is relatively complete, although aportion of its posterior edge is broken. Deformation of the leftpostorbital-postfrontal complex has overrun the dorsal surface ofthe parietal and obscured most of the margin of the parietal fora-men. The position of the parietal foramen can be best seen inthe holotype (Sidor et al., 2003:fig. 2), where it is situated abouthalfway along the interparietal suture. In all other pareiasaurs,the foramen is closer to the frontoparietal suture; the atypical
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

TSUJI ET AL.—PAREIASAUR FROM NIGER 755
condition in Bunostegos is likely due to the large size and an-terior extent of the postparietal. As in the holotype (Sidor et al.,2003:fig. 2), the parietal contacts the frontal anteriorly, the post-frontal anterolaterally, and the postorbital laterally (Fig. 2A).The posterior contacts of the parietal are more difficult to makeout, but we suspect that the condition depicted by Sidor et al.(2003) is essentially correct: the parietal contacts the supratem-poral laterally, with the median postparietal interposed along theinterparietal suture behind the parietal foramen.
Postparietal—This element was depicted by Sidor et al.(2003:fig. 2) as a small, rectangular bone positioned entirely onthe skull table between the parietals and supratemporals. Thisarrangement mirrors that proposed by Lee (1997a; Spencer andLee, 2000) for Elginia, although Jalil and Janvier (2005) sug-gested that the conditions in Elginia and Bunostegos might not behomologous. We can tentatively identify the posterior border ofthe postparietal, where it presumably contacts the enlarged tabu-lar, but the lateral extent of the postparietal is unclear (Fig. 2A).Unfortunately, MNN MOR86 provides little new information onthe morphology of the postparietal. In ventral view, the contactbetween the tabular and the overlying supratemporal can be de-termined, but the suture between the postparietal and the formercannot be determined. Targeted preparation of other specimens(e.g., MNN MOR37) has also failed to shed light on this anatomy.
Supratemporal—We follow Lee (1995, 1997a, 1997b; Lee et al.,1997) in considering the large element of the posterolateral skullroof to be the supratemporal (contra Hartmann-Weinberg, 1933;Boonstra, 1934; Bystrov, 1957). Among pareiasaurs, Elginia hasthe best-developed projection on the supratemporal, where ittakes the form of a long, gently curved ‘horn’ (Newton, 1893;Spencer and Lee, 2000). A similar feature occurs in Arganaceras(Jalil and Janvier, 2005:fig. 9) and Bunostegos, although the ‘horn’in the latter ends in a rounded, globular swelling. In the holotypeof Bunostegos, these ‘horns’ are well developed and project lat-erally from the skull roof (Sidor et al., 2003:fig. 2). By contrast,in MNN MOR86, they are somewhat eroded but clearly did notproject as far laterally as those of the holotype (Figs. 2A). It istherefore possible that the holotypic skull is somewhat flatteneddorsoventrally. The region of the left supratemporal ‘horn’ inMNN MOR86 has been deformed so that the normally laterallyfacing part of the supratemporal faces ventrally.
Contacts between the supratemporal and the squamosal, pos-torbital, and parietal can be made out (Figs. 2A, 3). The suturebetween the supratemporal and tabular is oriented differently indorsal than in ventral view. Dorsally, the suture is broadly con-cave and is positioned approximately halfway between the lateraledge of the skull table and the midline. The nature of the contactbetween the supratemporal and the postparietal cannot be distin-guished in its entirety; it appears, however, that the large tabularsexclude contact between the postparietal and the supratemporal(Fig. 2A). On the ventral surface, the tabulars broadly underlapthe supratemporal, so the supratemporal appears much thinnerin ventral view (Fig. 2B).
Tabular—Lee (1997a) discussed the presence of a skull ele-ment interposed between the postparietal and supratemporal inpost-Embrithosaurus pareiasaurs, and commented on its variousinterpretation as a tabular (Wild, 1985), a cervical osteoderm in-corporated into the skull roof (Brink, 1955), or as a neomorphicossification. Jalil and Janvier (2005) supported the last proposalin Arganaceras, on the basis of its firm connection to the adjoiningbones. Most recently, Tsuji (2006, 2010) has provided evidencethat this element is homologous to the tabular in parareptilessuch as Macroleter and Nyctiphruretus. We adopt Tsuji’s (2006,2010) interpretation.
Sidor et al. (2003) found some evidence for the presence of anenlarged ‘supernumerary element’ (i.e., tabular) along the pos-terior margin of the skull in Bunostegos, although its anteriorsutural configuration was poorly understood. In MNN MOR86,
crushing of the left supratemporal region towards the midlinehas obscured much of the relevant area. However, as notedabove, a suture is visible between the more lateral, horn-bearingsupratemporal and the more medial element, here interpreted asthe tabular. In MNN MOR86, the area is well preserved but lacksobvious sutures on its dorsal surface. Preparation of the ventralsurface the posterior skull roof of MNN MOR86 has revealed anextensive underlap of the tabular under the supratemporal, withthe former element appearing to be quite extensive on the ventralsurface (Fig. 2B).
Palate and Braincase
The right half of the palate is mostly preserved, although a sub-stantial piece is missing in the region of the palatine and pterygoid(Fig. 2B). In addition, some portions of the palate remain un-prepared, such as the dentigerous parts of the vomer and ptery-goid. Nevertheless, much additional morphological detail is avail-able when compared with the severely eroded palate of the holo-type. The lateral exposure of the braincase in MNN MOR47 al-lows a rare lateral view of the pareiasaurian braincase (Fig. 7).As in most (adult) specimens, however, the braincase elementscan be fused together, making it difficult to delimit the individualelements.
Vomer—The morphology of the ventral surface of the vomeris mostly obscured by adhering matrix (Fig. 2B). However, theholotypic specimen shows that the vomer of Bunostegos is lat-erally expanded near its midpoint (the ‘alar flange’ of Damianiand Modesto, 2001), contributing to the ‘C’-shaped outline of thechoanae in ventral view (Sidor et al., 2003:fig. 3). Viewed throughthe incomplete left naris, the dorsal laminae of both vomers areexposed in cross-section. Damiani and Modesto (2001) noted thatthe dentition of the pareiasaurian vomer can be divided into ananterior row of larger teeth and paired posterior rows of smallerteeth. This pattern is present in MNN MOR86, with the roots ofat least four anterior teeth (∼2 mm diameter) visible on the leftside. Large teeth are also partially exposed in MNN MOR28, al-though they are still largely embedded in matrix (Fig. 4). Moreposteriorly, the roots of substantially smaller teeth (<1 mm di-ameter) form two subparallel rows. These rows are present onthe preserved left vomer and continue onto the correspondingpterygoid. Anteriorly, the vomer contacts the premaxilla, whereit forms a limited contribution to the prepalatal foramen, and pos-teriorly it contacts the pterygoid medially and the palatine later-ally. Although highly weathered, the basic outline of the posteriorcontacts of the vomer are exposed in MNN MOR25 (Fig. 8). Thevomer in MNN MOR25 also shows infillings that we interpret asthe remains of neurovascular canals that extended anteroposteri-orly within the vomer and pterygoid.
Palatine—Only the anterior portion of the right palatine is pre-served (Fig. 2B). It underlaps the palatal portion of the maxillaand forms the posterior half of the lateral margin of the rightchoana. Posteriorly, the palatine sutures to the ectopterygoid,with a comparatively small posterior palatal foramen in between.The posterior palatal (or suborbital) foramen is more clearly ex-posed in MNN MOR28, where it is larger and enclosed betweenthe palatine and ectopterygoid close to the triple junction of thesetwo elements with the maxilla (Fig. 4). Most of the body of thepalatine, where it contacts the pterygoid, is missing or obscuredby matrix, but the posterolaterally oriented palatine-pterygoidsuture can be seen in MNN MOR25 (Fig. 8). There are two paral-lel rows of denticles visible on the ventral surface of the palatinethat continue onto the pterygoid, although they are largely cov-ered with matrix (Fig. 2B).
Ectopterygoid—A small portion of the right ectopterygoid ispreserved (Fig. 2B). As in other pareiasaurs, it articulates withthe maxilla laterally and with the palatine anteriorly. In addi-tion, a portion of its posteromedial contact with the pterygoid is
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

756 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
FIGURE 7. The braincase of Bunostegosakokanensis (MNN MOR47). Photograph andinterpretative drawing in left lateral view. Ab-breviations: bo, basioccipital; cp, cultriformprocess; ex, exoccipital; f, frontal; fo, fora-men ovale; jf, jugular foramen; pop, paroccip-ital process; pro, prootic; pt, pterygoid; sph,sphenethmoid; II, III, IV, foramen for cranialnerves; XII, foramina for hypoglossal nerve.
preserved. The posterior edge of the ectopterygoid also under-laps the anterior process of the jugal.
Pterygoid—The body of the pterygoid is mostly preserved, butremains covered in a thin layer of red matrix. Nonetheless, fourrows of small teeth can be seen on each side (Fig. 2B). As in otherpareiasaurs, the medial two rows are parallel, oriented parasagi-tally, and pass anteriorly to continue on the corresponding vomer.The lateral two rows are oriented anterolaterally. The majority ofpareiasaurs, except perhaps Elginia, have a single row of promi-nent denticles on the posterior edge of the transverse flange of thepterygoid (Lee, 1997a). Unfortunately, no specimen of Bunoste-gos preserves the relevant section. Although the transverse flangeis not complete in any specimen, it is clear that it was reduced anddirected more anterolaterally than laterally and approached thecheek, as in the nycteroleters such as Macroleter (Tsuji, 2006). Itis also clear that the transverse flange in Bunostegos, as in mostpareiasaurs, did not extend as far ventrally as in their nycteroleterrelatives.
Posteromedially, the pterygoid forms an immobile suturalconnection with the basipterygoid process of the basisphenoid(Fig. 8), as occurs in all adequately known pareiasaurs (Lee,1997a). Sidor et al. (2003) indicated that the interpterygoid vacu-
ity of Bunostegos was present and ‘U’-shaped. However, this cod-ing resulted from a misinterpretation of Lee’s (1997a:character18) distinction between character states. Although the new mate-rial does not help address this issue, the morphology of the holo-type of Bunostegos remains most similar to that of Bradysaurusand Deltavjatia: the posterior margin of the pterygoids is orientedalmost transversely in front of the basisphenoid (Figs. 2B, 4, 7; seealso Newton, 1893:pl. 39). Although much of the ventral surfaceof the pterygoids is eroded, MNN MOR25 most clearly demon-strates this condition (Fig. 8). We have reworded this character(number 21) in our analysis.
Only the right quadrate ramus of the pterygoid is preserved.Its articulation with the quadrate is eroded, but as observed bySidor et al. (2003) in the holotype, the quadrate was positionedposteriorly when compared with most other pareiasaurs. In thisregard, Bunostegos is most similar to Pareiasuchus nasicornis(Lee et al., 1997). A conspicuous subhorizontal ridge also extendsalong the medial face of the quadrate ramus of the pterygoid(Fig. 8), attenuating in prominence posteriorly. Based on its loca-tion, we suspect that this ridge served to limit dorsal movementof the quadrate, but the distribution and significance of this fea-ture among pareiasaurs is unclear. The quadrate process of the
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
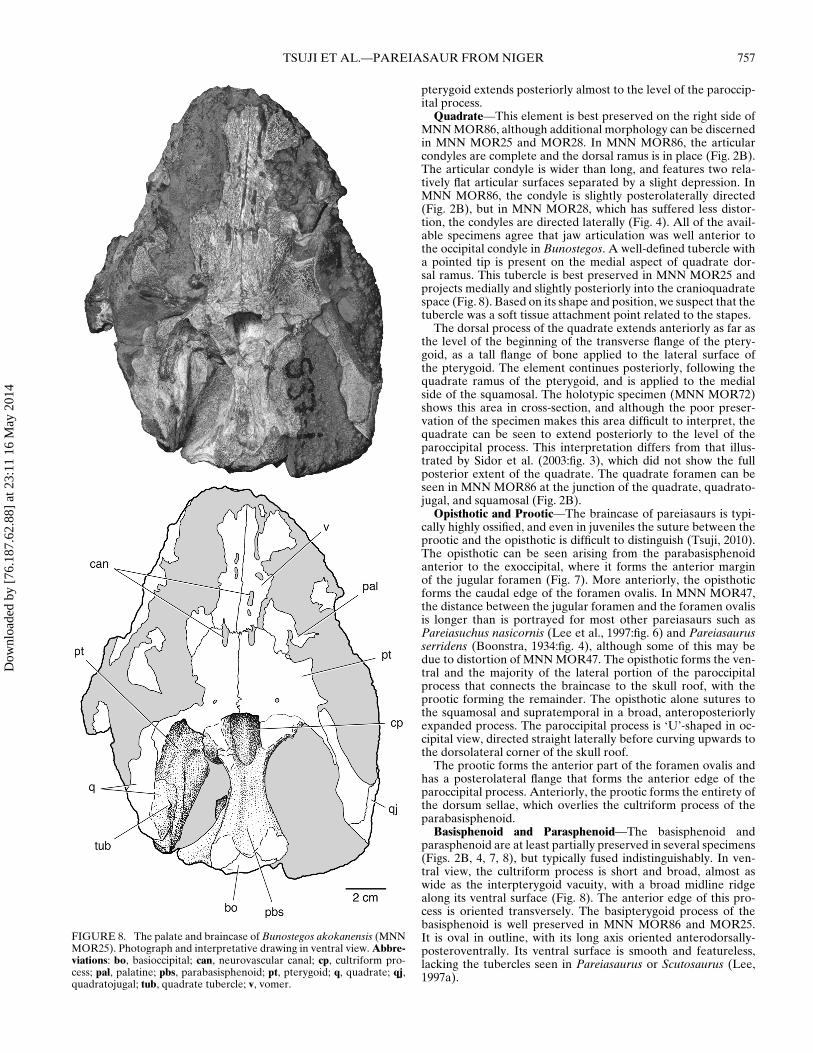
TSUJI ET AL.—PAREIASAUR FROM NIGER 757
FIGURE 8. The palate and braincase of Bunostegos akokanensis (MNNMOR25). Photograph and interpretative drawing in ventral view. Abbre-viations: bo, basioccipital; can, neurovascular canal; cp, cultriform pro-cess; pal, palatine; pbs, parabasisphenoid; pt, pterygoid; q, quadrate; qj,quadratojugal; tub, quadrate tubercle; v, vomer.
pterygoid extends posteriorly almost to the level of the paroccip-ital process.
Quadrate—This element is best preserved on the right side ofMNN MOR86, although additional morphology can be discernedin MNN MOR25 and MOR28. In MNN MOR86, the articularcondyles are complete and the dorsal ramus is in place (Fig. 2B).The articular condyle is wider than long, and features two rela-tively flat articular surfaces separated by a slight depression. InMNN MOR86, the condyle is slightly posterolaterally directed(Fig. 2B), but in MNN MOR28, which has suffered less distor-tion, the condyles are directed laterally (Fig. 4). All of the avail-able specimens agree that jaw articulation was well anterior tothe occipital condyle in Bunostegos. A well-defined tubercle witha pointed tip is present on the medial aspect of quadrate dor-sal ramus. This tubercle is best preserved in MNN MOR25 andprojects medially and slightly posteriorly into the cranioquadratespace (Fig. 8). Based on its shape and position, we suspect that thetubercle was a soft tissue attachment point related to the stapes.
The dorsal process of the quadrate extends anteriorly as far asthe level of the beginning of the transverse flange of the ptery-goid, as a tall flange of bone applied to the lateral surface ofthe pterygoid. The element continues posteriorly, following thequadrate ramus of the pterygoid, and is applied to the medialside of the squamosal. The holotypic specimen (MNN MOR72)shows this area in cross-section, and although the poor preser-vation of the specimen makes this area difficult to interpret, thequadrate can be seen to extend posteriorly to the level of theparoccipital process. This interpretation differs from that illus-trated by Sidor et al. (2003:fig. 3), which did not show the fullposterior extent of the quadrate. The quadrate foramen can beseen in MNN MOR86 at the junction of the quadrate, quadrato-jugal, and squamosal (Fig. 2B).
Opisthotic and Prootic—The braincase of pareiasaurs is typi-cally highly ossified, and even in juveniles the suture between theprootic and the opisthotic is difficult to distinguish (Tsuji, 2010).The opisthotic can be seen arising from the parabasisphenoidanterior to the exoccipital, where it forms the anterior marginof the jugular foramen (Fig. 7). More anteriorly, the opisthoticforms the caudal edge of the foramen ovalis. In MNN MOR47,the distance between the jugular foramen and the foramen ovalisis longer than is portrayed for most other pareiasaurs such asPareiasuchus nasicornis (Lee et al., 1997:fig. 6) and Pareiasaurusserridens (Boonstra, 1934:fig. 4), although some of this may bedue to distortion of MNN MOR47. The opisthotic forms the ven-tral and the majority of the lateral portion of the paroccipitalprocess that connects the braincase to the skull roof, with theprootic forming the remainder. The opisthotic alone sutures tothe squamosal and supratemporal in a broad, anteroposteriorlyexpanded process. The paroccipital process is ‘U’-shaped in oc-cipital view, directed straight laterally before curving upwards tothe dorsolateral corner of the skull roof.
The prootic forms the anterior part of the foramen ovalis andhas a posterolateral flange that forms the anterior edge of theparoccipital process. Anteriorly, the prootic forms the entirety ofthe dorsum sellae, which overlies the cultriform process of theparabasisphenoid.
Basisphenoid and Parasphenoid—The basisphenoid andparasphenoid are at least partially preserved in several specimens(Figs. 2B, 4, 7, 8), but typically fused indistinguishably. In ven-tral view, the cultriform process is short and broad, almost aswide as the interpterygoid vacuity, with a broad midline ridgealong its ventral surface (Fig. 8). The anterior edge of this pro-cess is oriented transversely. The basipterygoid process of thebasisphenoid is well preserved in MNN MOR86 and MOR25.It is oval in outline, with its long axis oriented anterodorsally-posteroventrally. Its ventral surface is smooth and featureless,lacking the tubercles seen in Pareiasaurus or Scutosaurus (Lee,1997a).
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

758 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
Despite the dorsal displacement experienced by the braincasein this specimen, MNN MOR28 best preserves the ventral surfaceof the basisphenoid and demonstrates the presence of broad, ex-panded basal tubera (Fig. 4). These were mostly eroded away inMNN MOR47 (Fig. 7) and slightly damaged in MNN MOR86(Fig. 2B). The basal tubera are positioned posteriorly, close tothe occipital condyle, a condition most similar to Bradysaurusand differing from taxa such as Anthodon where the tubera arelocated midway between the occipital condyle and the basiptery-goid processes (Lee 1997a). Lee (1997a) proposed a distinctionin the anatomy of the basisphenoid, where basal pareiasaurs suchas Bradysaurus and Nochelesaurus retain a primitive ‘waisting’ ofthe basisphenoid, and more-derived pareiasaurs show no signifi-cant transverse constriction. The anatomy of Bunostegos is diffi-cult to assign to either configuration, but in general it appears thatthe length and relative narrowness of the basisphenoid is mostsimilar to that depicted for Bradysaurus (Lee, 1997b:fig. 8a).
MNN MOR47 shows the cultriform process to cradle the baseof the sphenethmoid (Fig. 7). In lateral view, the prootic can beseen to suture along the dorsal margin of the basisphenoid, withthe anterior portion of the fenestra ovalis visible.
Sphenethmoid—The exposure of the braincase in lateral viewin MMN MOR47 affords a rare view of a pareiasaur spheneth-moid (Fig. 7), although portions of this element can also beseen in MNN MOR86. Lee (1995) interpreted this midline brain-case element as a pleurosphenoid, whereas deBraga and Riep-pel (1997) argued for its recognition as a sphenethmoid, whichwe adopt here. Boonstra (1934:18) provided a description ofthis element for Pareiasaurus serridens and Watson (1914:fig. 1)discussed it in a specimen that was subsequently attributed toEmbrithosaurus by Lee (1997a). An incomplete sphenethmoidof Pareiasuchus was figured by Haughton (1929:fig. 23). Thesphenethmoid is a trapezoidal element in lateral view that arisesfrom the cultriform process and contacts the underside of thefrontals. In sagittal section the element appears as a ‘V,’ withtwo plates of bone that diverge near their dorsal edge. As pro-posed by Boonstra (1934), a foramen near the posterior edge ofthe sphenethmoid likely transmitted cranial nerves II, III, and IVin life (Fig. 7).
Basioccipital—As noted by Sidor et al. (2003), the basioccip-ital forms most of the occipital condyle in Bunostegos, althoughthe limits of the adjacent exoccipitals are difficult to make out.The suture between the basisphenoid and basioccipital is visiblein the holotype, although a break along what appears to be thesuture confounds its interpretation. Among the available spec-imens, the eroded ventral surface of MNN MOR25 partiallyshows the path of this suture (Fig. 8). In contrast with the condi-tion depicted for Pareiasuchus nasicornis (Lee et al., 1997:fig. 6),the basisphenoid-basioccipital suture is concave anteriorly at itsmidpoint. The ventral surface of the basioccipital is smooth,lacking the median tubercle described for Arganaceras and Scu-tosaurus (Jalil and Janvier, 2005) as well as Pareiasuchus nasicor-nis (Lee et al., 1997).
Exoccipital—The lateral aspect of the exoccipital is well ex-posed in MNN MOR47 (Fig. 7), although sutures delimiting itfrom the surrounding elements are difficult to discern. The ele-ment extends dorsolaterally from the basioccipital, and the lateralprocess underlies the opisthotic where the latter begins to formthe paroccipital process. The exoccipital has a medial process, butnone of the available specimens makes it clear if it met its coun-terpart at the midline, thereby excluding the supraoccipital fromthe foramen magnum. A series of small foramina can be seen onthe ventrolateral edge of the exoccipital, close to where it fuseswith the basioccipital, for the hypoglossal nerve, similar to thecondition illustrated for P. nasicornis (Lee et al., 1997:fig. 6) andDeltavjatia (Tsuji, 2010:fig. 4.15). Dorsally, the exoccipital fusesindistinguishably with the supraoccipital. Lee (1995) consideredthe possession of a lateral flange on the exoccipital a diagnos-
tic character of pareiasaurs, but more recently this feature wasdemonstrated in Macroleter (Tsuji, 2006), and perhaps it may bea feature of all pareiasauromorphs.
DISCUSSION
Previous Analyses of Pareiasaur Phylogeny
The phylogenetic relationships of pareiasaurs have been thesubject of some cladistic work, but usually in the broader con-text of turtle origins or amniote phylogeny (Gauthier et al., 1988;Lee, 1993, 1995, 1997b; Laurin and Reisz, 1995; deBraga andRieppel, 1997; Hill, 2005; Tsuji and Muller, 2009; Lyson et al.,2010). Of these, Lee (1997b) has provided the only detailedanalysis of pareiasaur interrelationships, with 19 pareiasaur ter-minal taxa scored for 128 characters. This data matrix was re-vised and updated by Jalil and Janvier (2005), who included Ar-ganaceras vacanti as well as two types of unnamed pareiasaurpostcrania from the Ikakern Formation of Morocco. In one oftheir analyses, Jalil and Janvier (2005) also included Bunostegos,with codings based on the published description of Sidor et al.(2003). However, as with Lee (1997b), all of Jalil and Janvier’s(2005) analyses included turtles as an ingroup and a long list ofparareptile outgroups (viz. Millerettidae, Nycteroleteridae, Nyc-tiphruretidae, Owenetta, Barasaurus, Procolophonidae, Lantho-suchidae, and Sclerosaurus).
Tsuji and Muller (2008) rescored the German pareiasaurParasaurus and included it in a modified version of the Jaliland Janvier (2005) matrix. Tsuji and Muller (2008) eliminatedturtles from the ingroup and included three other parareptiles(Macroleter poezicus, Millerettidae, and Procolophonidae), usingMillerettidae and Procolophonidae as outgroups. In this analysis,Bunostegos remained relatively basally positioned as in Jalil andJanvier (2005). Tsuji (2010, in press) further modified the Jaliland Janvier (2005) and Tsuji and Muller (2008) matrices, also in-cluding all nycteroleter parareptiles in the analysis, but did notinclude a comprehensive reevaluation of Bunostegos.
Current Phylogenetic Analysis
We prefer to restrict our analysis to the interrelationships ofpareiasaurs and their closest relatives, because some of Lee’s(1997b) outgroups are of doubtful appropriateness (e.g., Scle-rosaurus has been identified as a derived procolophonid, not thesister taxon to Pareiasauria; see deBraga, 2003; Cisneros et al.,2005; Tsuji, 2006; Sues and Reisz, 2008) and turtles are nowconsidered to fall outside of Parareptilia (Zardoya and Meyer,1998; Rieppel and Reisz, 1999; Rieppel, 2000; Hill, 2005), al-though some debate remains (Lyson et al., 2010). The mono-phyly of traditional pareiasaurs has never been doubted seri-ously; Lee (1997b) suggested that the clade could be diagnosedby 37 synapomorphies, whereas deBraga and Rieppel (1997) sug-gested that it was diagnosed by 26 synapomorphies. Pareiasaursare clearly distinctive fossils, and probably would have beensimilarly distinctive in life (Fig. 9). More recent analyses haveshown that certain nycteroleter taxa form the closest outgroupsto pareiasaurs (Tsuji, 2006; Tsuji and Muller, 2009; Tsuji et al.,2012), but pareiasaur ingroup relationships remain little modifiedfrom the results of Lee’s (1997b) initial work.
Presented in Appendix 1 is a summary of the changes madeto the character list and character matrix presented in Tsuji (inpress), which combines selected characters from the study ofJalil and Janvier (2005) and Lee (1997a), as well as new char-acters developed in the course of our work on Bunostegos. Ap-pendix 2 includes our data matrix, with 127 characters codedfor 21 pareiasaur taxa, six nycteroleters, as well as ‘Owenetta’kitchingorum and Millerettidae. The matrix in Appendix 2 wassubjected to a traditional search with 5000 random addition se-quences and 1000 trees per replication using the Tree Bisection
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
TSUJI ET AL.—PAREIASAUR FROM NIGER 759
FIGURE 9. Restoration of a subadult Bunostegos akokanensis in life.Image © Marc Boulay.
Reconnection (TBR) algorithm in TNT (Goloboff et al., 2008).The default collapsing rule in TNT was used, which eliminatesbranches for which the minimum possible length is zero. All ofthe characters were left unordered and unweighted. Milleretti-dae was the sole outgroup. A bootstrap analysis was run using5000 replicates, and the Bremer decay indices were also deter-mined, both using TNT (Goloboff et al., 2008). Forty-five min-imum length trees of 213 steps were recovered, with a strictconsensus yielding a relatively well-resolved topology, althoughthe support values for most of the nodes are not particularlyhigh (Fig. 10; see online Supplementary Data). In all trees,Bunostegos is recovered as the sister taxon to a clade includ-ing all other non-Tapinocephalus and Pristerognathus assemblagezone pareiasaurs. Velosauria, a group of derived pareiasaurs de-fined by Lee (1994) as the ancestor of Therischia and Pumilio-pareiasauria and all its descendants, is recovered in this analysis.
In their initial description, Sidor et al. (2003) noted thatBunostegos possessed features of both primitive and derivedpareiasaurs. The present analysis suggests that, at least on ba-sis of its cranial anatomy, Bunostegos is best considered a rela-tively basal pareiasaur. This position can be attributed to severalspecific codings. In particular, the shape (character 54) and highnumber of marginal teeth (character 51), the shape of the ante-rior edge of the interpterygoid vacuity (character 21), and theshape and orientation of the paroccipital process (character 6).We were unable to determine with confidence the sutural con-tacts of the postparietal and supratemporal, which has been con-sidered a key feature linking Elginia, Arganaceras, and possiblyBunostegos (Jalil and Janvier, 2005). As such, character 33 wascoded as unknown (Appendix 2) for Bunostegos. However, whenthis character was rescored to the state present in Elginia and Ar-ganaceras and the analysis rerun, the resulting strict consensustree does not change (tree length 213 steps; consistency index =0.751; retention index = 0.877), which reinforces the interpreta-tion that this character is convergent.
Age of the Moradi Formation
The Moradi Formation has been considered Late Permianin age (Taquet, 1972; de Ricqles and Taquet, 1982; Sidor et al.,2005b; Smiley et al., 2008). This age assignment is based pri-marily on biostratigraphic correlations with other, better-knownPermian faunas, but such correlations are not without conflict.Taquet (1972) indicated a Late Permian age based on the
FIGURE 10. Cladistic relationships of Bunostegos akokanensis withinPareiasauria. A strict consensus of the 45 most parsimonious topologiesrecovered from a TNT analysis. Bremer decay values above one andbootstrap values >50% are listed above and below, respectively, eachwell-supported branch.
presence of a pareiasaur and a captorhinid (Moradisaurus) thatwas similar to the Russian taxon Gecatogomphius. However,Bunostegos is arguably more similar to middle Permian forms,and Gecatagomphius is now considered to be from the middlePermian of Russia. As outlined by Sidor et al. (2005b), thetemnospondyls of the Moradi Formation are members of cladesthat were primarily known from the Carboniferous. The onlyevidence supporting a Late Permian (as opposed to middlePermian) age for the fauna is the presence of a large, presumablyderived, gorgonopsid (Smiley et al., 2008). The phylogeneticposition of Bunostegos recovered here is equivocal, becausethis taxon fits between middle Permian and Late Permianpareiasaurs. The absence of biostratigraphically useful dicyn-odonts in the Moradi Formation makes it particularly difficult totie the Moradi vertebrate fauna to other Pangean assemblages.
Biogeographic Implications
Lee (1997b) suggested that pareiasaurs had their origin inGondwana, based on the early appearance of Bradysaurus,Embrithosaurus, and Nochelesaurus in the middle Permian ofSouth Africa. However, the remainder of his pareiasaur clado-gram was claimed to have little biogeographic structure (Lee,1997b:291–292). More recently, Tsuji (2006) found Macroleterpoezicus, a parareptile from Russia, to be the closest relativeof Pareiasauria within Parareptilia. Subsequent analyses have
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

760 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
supported the hypothesis that the nycteroleters are either themonophyletic sister taxon of, or grade into, pareiasaurs (Tsuji,2010; Tsuji et al., 2012). Most nycteroleters are from the mid-dle Permian of Russia, and therefore Laurasian, and althougha member of the group has been identified from the ChickashaFormation in Oklahoma (Reisz and Laurin, 2001), another spec-imen has been recovered from the Tapinocephalus AssemblageZone in South Africa (Cisneros and Tsuji, 2009). We suggest thatno biogeographic scenario for the origin of pareiasaurs can bedefinitively supported at present. However, given the current fos-sil record, we agree with Lee (1997b) that pareiasaurs probablyradiated in southern Pangea, as all of the middle Permian mem-bers of the group have been recovered from southern Africanrocks.
Sidor et al. (2005b) argued that the composition of the MoradiFormation tetrapod fauna was distinctive and tied its endemicnature to the arid conditions inferred for central Pangea (Gibbset al., 2002; Rees et al., 2002). More recent analyses of Moradi pa-leosols by Tabor et al. (2011) have confirmed that deposition ofthe Moradi Formation occurred under semiarid to hyperarid con-ditions. According to the climate hypothesis, Moradi tetrapodsowe some of their distinctive qualities (e.g., highly autapomor-phic morphology, long ghost lineages) to a long period of isola-tion from the remainder of tetrapod faunas on Pangea. For ex-ample, the temnospondyls Nigerpeton and Saharastega are mostclosely related to taxa known from the Permo-Carboniferous ofEuramerica. Although less pronounced, the phylogenetic posi-tion and anatomy of Bunostegos mirrors that of the Moradi tem-nospondyls; Bunostegos clusters with relatively early divergingmembers of Pareiasauria, but it is also highly autapomorphic, par-ticularly with regards to its cranial sculpture, which appears toshare convergent characters with more derived pareiasaurs suchas Elginia and Arganaceras.
Among middle or Upper Permian tetrapod assemblages, theMoradi Formation is closest in composition to that of the Ikak-ern Formation of Morocco. Both formations have at least onepareiasaur (Bunostegos, Arganaceras), a large captorhinid withmultiple toothrows (Moradisaurus, unnamed taxon; Jalil and Du-tuit, 1996), and amphibians belonging to groups that typically oc-cur in the Permo-Carboniferous of North America (Nigerpeton,Diplocaulus; Dutuit, 1988). Steyer and Jalil (2009) recently notedthe presence of the first temnospondyl from the Ikakern For-mation, noting that it likely belonged to a non-stereospondyl ornon-euskelian (sensu Yates and Warren, 2000). Given these sim-ilarities, we tentatively suggest that the tetrapod assemblages ofthe Moradi and Ikakern formations represent a faunal provincedistinct from that of southern Pangea (e.g., Karoo-type assem-blages).
Other Pareiasaurs from the Moradi Formation
Sidor et al. (2003) noted that a pareiasaur skull from theMoradi Formation was figured by Taquet (1976:pl. 1). This skull,which appears similar to Bunostegos in its possession of supraor-bital bosses, was never named or described, and is now missingfrom the MNHN (P. Taquet, pers. comm., 2008).
Additional pareiasaur material collected in 2003 and 2006is undergoing preparation. Fortunately, several specimens dis-cussed here include both cranial and postcranial elements, whichshould permit much of the postcranial skeleton of Bunostegos tobe identified. A preliminary examination of our Moradi collec-tion suggests that there is nothing definitive to establish greaterthan one pareiasaur taxon in the fauna. However, it is importantto recognize that pareiasaur genera are known to co-occur in thesame formation in South Africa and in China, leaving open thepossibility that Bunostegos coexisted with a second pareiasaur inthe Permian of Niger.
CONCLUSIONS
Fossils referable to Bunostegos akokanensis permit an im-proved understanding of the cranial morphology and phyloge-netic position of this taxon among pareiasaurs. Phylogenetically,Bunostegos sits between early diverging taxa such as Bradysaurusand Embrithosaurus and the derived velosaurs (e.g., Anthodon,Pareiasuchus, Scutosaurus). However, Bunostegos is also highlyautapomorphic, with at least eight unique features identified inthis work. The presence of Bunostegos at a low paleolatitude addssupport to the theory that the tetrapod fauna of central Pangeawas isolated by xeric conditions during the Late Permian (Sidoret al., 2005b; Tabor et al., 2011).
ACKNOWLEDGMENTS
We thank D. Blackburn, D. Chaney, A. Dindine, B. Gado, H.Larsson, C. Looy, T. Lyman, A. Maga, R. O’Keefe, D. Sindy, T.Smiley, B. Tahirou, and S. Thomas for assistance in the field, andthe personnel of the U.S. Embassy in Niamey for support. Addi-tionally, we acknowledge J. Alexander, B. Crowley, J. Groenke,V. Heisey, and A. Huttenlocker for their excellent preparationof the material reported in this paper. For the illustrations inFigures 2–7, we thank J. Swales. We are grateful to M. Boulayfor permission to feature his restoration. Field work in 2003 wassupported by National Geographic Society grant 7258–02 (toC.A.S.). Field work in 2006 and continuing research has beensupported by NSF EAR-0617718 (to C.A.S.) and EAR-0617250(to N.J.T.).
LITERATURE CITED
Araujo, D. C. F. 1985. Sobre Pareiasaurus americanus sp. nov., do Permi-ano Superior do Rio Grande do Sul, Brasil. I—diagnose especıfica.Anais da Academia Brasileira de Ciencias 57:63–66.
Boonstra, L. D. 1934. Pareiasaurian studies. Part IX.—the cranial osteol-ogy. Annals of the South African Museum 31:1–38.
Brink, A. S. 1955. On Nanoparia Broom. Palaeontologia africana 3:57–63.Bystrov, A. P. 1957. [The pareiasaur skull]. Trudy Paleontologischeskogo
Instituta AN SSSR 68:3–18. [Russian]Cisneros, J. C., and L. A. Tsuji. 2009. Nycteroleter affinities of a Permian
parareptile from the South African Karoo Basin. Acta Palaeonto-logica Polonica 54:165–169.
Cisneros, J. C., F. Abdala, and M. C. Malabarba. 2005. Pareiasaurids fromthe Rio do Rasto Formation, southern Brazil: biostratigraphic impli-cations for Permian faunas of the Parana Basin. Revista Brasileirade Paleontologia 8:13–24.
Damiani, R. J., and S. P. Modesto. 2001. The morphology of thepareiasaurian vomer. Neues Jahrbuch fur Geologie und Palaontolo-gie, Monatshefte 2000:423–434.
Damiani, R., C. A. Sidor, J. S. Steyer, R. M. H. Smith, H. C. E. Larsson, A.Maga, and O. Ide. 2006. The vertebrate fauna of the Upper Permianof Niger. V. The primitive temnospondyl Saharastega moradiensis.Journal of Vertebrate Paleontology 26:559–572.
deBraga, M. 2003. The postcranial skeleton, phylogenetic position, andprobable lifestyle of the Early Triassic reptile Procolophon trigoni-ceps. Canadian Journal of Earth Sciences 40:527–556.
deBraga, M., and O. Rieppel. 1997. Reptile phylogeny and the inter-relationships of turtles. Zoological Journal of the Linnean Society120:281–354.
Dutuit, J. M. 1988. Diplocaulus minimus n. sp. (Amphibia: Nectridea),lepospondyle de la formation d’Argana dans l’Atlas occidentalmarocain. Comptes Rendus de l’Academie des Sciences, Paris307:851–854.
Gauthier, J., A. Kluge, and T. Rowe. 1988. Amniote phylogeny and theimportance of fossils. Cladistics 4:105–209.
Gibbs, M. T., P. M. Rees, J. E. Kutzbach, A. M. Ziegler, P. J. Behling,and D. B. Rowley. 2002. Simulations of Permian climate andcomparisons with climate-sensitive sediments. Journal of Geology110:33–55.
Goloboff, P. A., J. S. Farris, and K. C. Nixon. 2008. TNT, a free programfor phylogenetic analysis. Cladistics 24:774–786.
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

TSUJI ET AL.—PAREIASAUR FROM NIGER 761
Hartmann-Weinberg, A. P. 1933. Die evolution der Pareiasauriden.Trudy Paleontologischeskogo Instituta AN SSSR 3:3–66.
Haughton, S. H. 1929. Pareiasaurian studies. Part II.—notes on somepareiasaurian brain-cases. Annals of the South African Museum28:88–96.
Hill, R. V. 2005. Integration of morphological data sets for phylogeneticanalysis of Amniota: the importance of integumentary charactersand increased taxonomic sampling. Systematic Biology 58:530–547.
Huene, F. von. 1944. Pareiasaurierreste aus dem Ruhuhu-Gebiet. Palaon-tologisches Zeitschrifte 23:386–410.
Jalil, N.-E., and J. M. Dutuit. 1996. Permian captorhinid reptiles from theArgana Formation, Morocco. Palaeontology 39:907–918.
Jalil, N.-E., and P. Janvier. 2005. Les pareiasaures (Amniota, Pararep-tilia) du Permien superieur du Bassin d’Argana, Maroc. Geodiver-sitas 27:35–132.
Laurin, M., and R. R. Reisz. 1995. A reevaluation of early amniote phy-logeny. Zoological Journal of the Linnean Society 113:165–223.
Lee, M. S. Y. 1993. The origin of the turtle body plan: bridging a famousmorphological gap. Science 261:1716–1720.
Lee, M. S. Y. 1994. Evolutionary morphology of pareiasaurs. Ph.D. Dis-sertation, University of Cambridge, Cambridge, U.K., 499 pp.
Lee, M. S. Y. 1995. Historical burden in systematics and the interrelation-ships of ‘parareptiles.’ Biological Reviews 70:459–547.
Lee, M. S. Y. 1997a. A taxonomic revision of pareiasaurian reptiles: im-plications for Permian terrestrial palaeoecology. Modern Geology21:231–298.
Lee, M. S. Y. 1997b. Pareiasaur phylogeny and origin of turtles. Zoologi-cal Journal of the Linnean Society 120:197–280.
Lee, M. S. Y. 2000. The Russian pareiasaurs; pp. 71–85 in M. J. Benton,M. A. Shishkin, D. M. Unwin, and E. N. Kurochkin (eds.), The Ageof Dinosaurs in Russia and Mongolia. Cambridge University Press,New York.
Lee, M. S. Y., C. E. Gow, and J. W. Kitching. 1997. Anatomy and rela-tionships of the pareiasaur Pareiasuchus nasicornis from the UpperPermian of Zambia. Palaeontology 40:307–335.
Lyson, T. R., G. S. Bever, B.-A. S. Bhullar, W. G. Joyce, and J. A. Gau-thier. 2010. Transitional fossils and the origin of turtles. Biology Let-ters 6:830–833.
Newton, E. T. 1893. On some new reptiles from the Elgin Sandstones.Philosophical Transactions of the Royal Society of London, SeriesB 184:431–503.
O’Keefe, F. R., C. A. Sidor, H. C. E. Larsson, A. Maga, and O. Ide.2005. The vertebrate fauna of the Upper Permian of Niger—III,morphology and ontogeny of the hindlimb of Moradisaurus gran-dis (Reptilia, Captorhinidae). Journal of Vertebrate Paleontology25:309–319.
O’Keefe, F. R., C. A. Sidor, H. C. E. Larsson, A. Maga, and O. Ide. 2006.Evolution and homology of the astragalus in early amniotes: newfossils, new perspectives. Journal of Morphology 267:415–425.
Rees, P. M., A. M. Ziegler, M. T. Gibbs, J. E. Kutzbach, P. J. Behling, andD. B. Rowley. 2002. Permian phytogeographic patterns and climatedata/model comparisons. Journal of Geology 110:1–31.
Reisz, R. R., and M. Laurin. 2001. The reptile Macroleter: first verte-brate evidence for correlation of Upper Permian continental strataof North America and Russia. Geological Society of America Bul-letin 113:1229–1233.
Ricqles, A. de, and P. Taquet. 1982. La faune de vertebres du PermienSuperieur du Niger. I. Le captorhinomorphe Moradisaurus grandis(Reptilia, Cotylosauria). Annales de Paleontologie (Vert.-Invert.)68:33–106.
Rieppel, O. 2000. Turtles as diapsid reptiles. Zoologica Scripta29:199–212.
Rieppel, O., and R. R. Reisz. 1999. The origin and early evolution of tur-tles. Annual Review of Ecology and Systematics 30:1–22.
Seeley, H. G. 1888. Researches on the structure, organisation, and clas-sification of the fossil Reptilia.—II. On Pareiasaurus bombidens(Owen), and the significance of its affinities to amphibians, reptiles,and mammals. Philosophical Transactions of the Royal Society ofLondon, Series B 179:59–109.
Sidor, C. A., D. C. Blackburn, and B. Gado. 2003. The verte-brate fauna of the Upper Permian of Niger—II, preliminarydescription of a new pareiasaur. Palaeontologia africana 39:45–52.
Sidor, C. A., H. C. E. Larsson, F. R. O’Keefe, D. C. Blackburn, and R.M. H. Smith. 2005a. Upper Permian captorhinid and pareiasaur rep-
tiles from Niger: new data and interpretation. Journal of VertebratePaleontology 25(3, Supplement):115A.
Sidor, C. A., K. D. Angielczyk, D. M. Weide, R. M. H. Smith, S. J. Nes-bitt, and L. A. Tsuji. 2010. Tetrapod fauna of the lowermost UsiliFormation (Songea Group, Ruhuhu Basin) of southern Tanzania,with a new burnetiid record. Journal of Vertebrate Paleontology30:696–703.
Sidor, C. A., F. R. O’Keefe, R. Damiani, J. S. Steyer, R. M. H. Smith,H. C. E. Larsson, P. C. Sereno, O. Ide, and A. Maga. 2005b. Per-mian tetrapods from the Sahara show climate-controlled endemismin Pangaea. Nature 343:886–889.
Smiley, T. M., C. A. Sidor, O. Ide, and A. Maga. 2008. Vertebrate faunaof the Upper Permian of Niger. VI. First evidence of a gorgonopsiantherapsid. Journal of Vertebrate Paleontology 28:543–547.
Spencer, P. S., and M. S. Y. Lee. 2000. A juvenile Elginia and early growthin pareiasaurs. Journal of Paleontology 74:1191–1195.
Steyer, J. S., and N.-E. Jalil. 2009. First evidence of a temnospondyl inthe Late Permian of the Argana Basin, Morocco. Special Papers inPalaeontology 81:155–160.
Steyer, J. S., R. Damiani, C. A. Sidor, F. R. O’Keefe, H. C. E. Larsson, A.Maga, and O. Ide. 2006. The vertebrate fauna of the Upper Permianof Niger. IV. Nigerpeton ricqlesi (Temnospondyli: Cochleosauri-dae), and the edopoid colonization of Gondwana. Journal of Ver-tebrate Paleontology 26:18–28.
Sues, H.-D., and R. R. Reisz. 2008. Anatomy and phylogenetic rela-tionships of Sclerosaurus armatus (Amniota: Parareptilia) from theBuntsandstein (Triassic) of Europe. Journal of Vertebrate Paleon-tology 28:1031–1042.
Tabor, N. J., R. M. H. Smith, J. S. Steyer, C. A. Sidor, and C. J. Poulsen.2011. The Permian Moradi Formation of northern Niger: pale-osol morphology, petrography and mineralogy. Palaeogeography,Palaeoclimatology, Palaeoecology 299:200–213.
Taquet, P. 1967. Decouvertes paleontologiques recentes dans le Norddu Niger; pp. 415–418 in Anonymous (ed.), Problemes actuels dePaleontologie—Evolution des Vertebres. Centre National de laRechererche Scientifique, Paris.
Taquet, P. 1972. Un exemple de datation et de correlation stratigraphiquebase sur les Captorhinomorphes (Reptiles cotylosauriens).Memoires du Bureau de Recherches Geologiques et Minieres77:407–409.
Taquet, P. 1976. Geologie et paleontologie du gisement de Gadoufaoua(Aptien du Niger). Cahiers de Paleontologie, Paris, 191 pp.
Taquet, P. 1978. Niger et Gondwana. Annales de la Societe Geologiquedu Nord 97:337–341.
Tsuji, L. A. 2006. Cranial anatomy and phylogenetic affinities of the Per-mian parareptile Macroleter poezicus. Journal of Vertebrate Pale-ontology 26:849–865.
Tsuji, L. A. 2010. Evolution, morphology and paleobiology of thePareiasauria and their relatives (Amniota: Parareptilia). Ph.D.dissertation, Humboldt Universitat zu Berlin, Berlin, Germany,220 pp.
Tsuji, L. A. In press. Anatomy, cranial ontogeny, and phylogenetic rela-tionships of the pareiasaur Deltavjatia rossicus from the Late Per-mian of central Russia. Earth and Environmental Science Transac-tions of the Royal Society of Edinburgh.
Tsuji, L. A., and J. Muller. 2008. A re-evaluation of Parasaurus geinitzi,the first named pareiasaur (Amniota, Parareptilia). Canadian Jour-nal of Earth Sciences 45:1111–1121.
Tsuji, L. A., and J. Muller. 2009. Assembling the history of the Pararep-tilia: phylogeny, diversification, and a new definition of the clade.Fossil Record 12:71–81.
Tsuji, L. A., J. Muller, and R. R. Reisz. 2012. Anatomy of Emeroleterlevis and the phylogeny of the nycteroleter parareptiles. Journal ofVertebrate Paleontology 32:45–67.
Watson, D. M. S. 1914. On the skull of a pareiasaurian reptile, and on therelationship of that type. Proceedings of the Zoological Society ofLondon 1914:155–180.
Wild, R. 1985. Ein schadelrest von Parasaurus geinitzi H. v. Meyer(Reptilia; Cotylosauria) aus dem Kupferschiefer (Permian) vonRichelsdorf (Hessen). Geologische Blatter fur Nordost-Bayern(Gedenkschrift B. v. Freyberg) 34/35:897–920.
Yates, A. M., and A. Warren. 2000. The phylogeny of the ‘higher’temnospondyls (Vertebrata: Choanata) and its implications for themonophyly and origins of the Stereospondyli. Zoological Journal ofthe Linnean Society 128:77–121.
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014

762 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 33, NO. 4, 2013
Zardoya, R., and A. Meyer. 1998. Complete mitochondrial genomesuggests diapsid affinities of turtles. Proceedings of the Na-tional Academy of Sciences of the United States of America95:14226–14231.
Submitted May 31, 2012; revisions received September 14, 2012;accepted October 8, 2012.Handling editor: Sean Modesto.
APPENDIX 1. Modifications to character list of Tsuji (in press).
Tsuji (in press) constructed the most recent cladistic data ma-trix for pareiasauromorphs, which included 125 characters (73cranial, 52 postcranial) in 29 taxa. This matrix was a revision of
the matrix of Jalil and Janvier (2005), which was itself a revisionof Lee (1997b). We redefined one character from Tsuji (in press),added two new characters, and substantially rescored one taxon,Bunostegos akokanensis.
Redefined Character
(21) Interpterygoid vacuity anterior shape: ‘V’-shaped (0); ‘U’-shaped (1); transversely oriented (2).
New Characters
(126) Postfrontal ‘horn’: absent (0); present (1).(127) Central boss on frontal: absent (0); present (1).
APPENDIX 2. Data matrix used in the cladistic analysis of pareiasauromorph interrelationships (Fig. 10). This matrix corresponds to that of Tsuji(in press), but incorporating the revisions noted in Appendix 1. ‘?’ denotes missing data. ‘A’ denotes polymorphism for 0, 1, and 2. ‘B’ denotespolymorphism for 0 and 1. Abbreviations: Ba., Bashkyroleter; Br., Bradysaurus; O., ‘Owenetta’; P., Pareiasuchus.
1 2 3 4 5 6 7Taxon 0 0 0 0 0 0 0Millerettidae 0??0000000 0000?00000 0000000000 0000000000 A000??0000 00?00001?0 00000?0000O. kitchingorum 0?0000??10 0001000000 0000001001 11?0000000 2000?000?0 00?00000?0 1?10000000Ba. bashkyricus 1????0??20 0000?110?1 000?1010?1 100??01010 0100?????0 00???000?? ??????????Ba. mesensis 1???????2? 0000?110?1 000?101011 000??01010 0100?????0 00?0?0000? ??????????Rhipaeosaurus ?????????? 0000?????? ?00??????? ?????????? ???????0?? ?00?1?0??0 ???????0?0Nycteroleter 1???????2? 0000?11001 000?101011 100000101? 0100?00??0 00000000?? ??????????Emeroleter 1??1????2? 0000?110?1 000?101011 000??01010 0100??0??0 0000000000 ?110???11?Macroleter 101110002? 0000?110?1 0000101011 1002101010 1100000000 00?0000000 011????000Br. baini 1112111111 0001021111 2111000010 ???2101210 0111012111 0101100111 ?020101110Br. seeleyi 11?2111111 000102?111 2111000010 1102101110 01110121?0 0101100111 0020101110Nochelesaurus 11??111111 000102?1?1 2111000010 1????01220 01110121?0 01011??11? ???????110Embrithosaurus 111?111111 000102?1?1 2111000010 1102101210 0111012110 0102200111 0020111110Deltavjatia 1??2101111 1001121111 1111000010 1202101221 0111012110 0113301102 ?020?11111Shansisaurus ?????????? ?????????? ?????????? ?????????? ????????1? ?11?410??? ???????121Shihtienfenia ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?03????121P. peringueyi 11?210??11 1001121111 1111000010 1202111221 0111012110 1113410102 ?021???121P. nasicornis 11?210??11 1001121111 1111000010 1202111221 0111012111 111341010? ??2????1??Pareiasaurus 11?210??11 11?11211?1 1111010110 1?02111221 0111012110 11134??10? 002????111Scutosaurus 1112101111 1111121111 1111010110 1202111221 0111012110 1113411102 ?0?1111111Sanchuansaurus ?????????? ?????????? ??1?01???? ?????????? ?????????0 11134?1??? ??????????Parasaurus ?????????? ?????????? 11???????0 1??2??12?0 01???????? ?113?01??? ?02????11?Elginia 11??10??1? ?????21?1? 111101?1?0 1212111231 01111????0 1113????0? ?0???1?1?1Provelosaurus ????10??1? ?????????? ???1000010 ???2111221 01110????0 1113?????? ?02??1?11?Anthodon 1???????11 10?1121111 0111000010 1??2111221 0111011110 12144?0102 ?03??1?111Pumiliopareia 1???????11 10?112?1?1 01110000?0 1??2111221 0111011??0 121440??0? ?0????????Nanoparia 1???????11 10?11211?1 0111000010 1202111??? 01110????0 11????010? ??????????Arganaceras ???2?0111? ?11112?1?1 ???10?0110 1212111231 01111???10 1113??1??? ??????????Obirkovia ?????????? ?????????? ?????????? ????????31 ?????????? ?????????? ??????????Bunostegos 111?11??11 1001021111 2111000010 02?2101221 01110????0 01?2???10? ???????110
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014
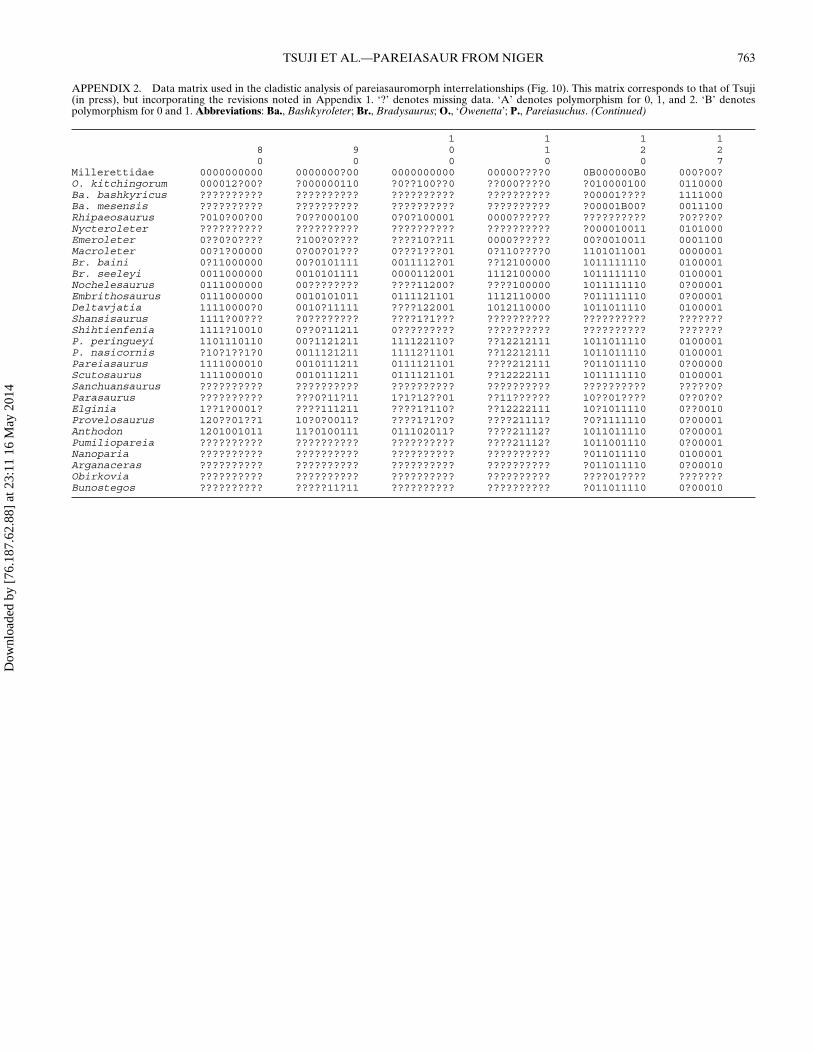
TSUJI ET AL.—PAREIASAUR FROM NIGER 763
APPENDIX 2. Data matrix used in the cladistic analysis of pareiasauromorph interrelationships (Fig. 10). This matrix corresponds to that of Tsuji(in press), but incorporating the revisions noted in Appendix 1. ‘?’ denotes missing data. ‘A’ denotes polymorphism for 0, 1, and 2. ‘B’ denotespolymorphism for 0 and 1. Abbreviations: Ba., Bashkyroleter; Br., Bradysaurus; O., ‘Owenetta’; P., Pareiasuchus. (Continued)
1 1 1 18 9 0 1 2 20 0 0 0 0 7
Millerettidae 0000000000 0000000?00 0000000000 00000????0 0B000000B0 000?00?O. kitchingorum 000012?00? ?000000110 ?0??100??0 ??000????0 ?010000100 0110000Ba. bashkyricus ?????????? ?????????? ?????????? ?????????? ?00001???? 1111000Ba. mesensis ?????????? ?????????? ?????????? ?????????? ?00001B00? 0011100Rhipaeosaurus ?010?00?00 ?0??000100 0?0?100001 0000?????? ?????????? ?0???0?Nycteroleter ?????????? ?????????? ?????????? ?????????? ?000010011 0101000Emeroleter 0??0?0???? ?100?0???? ????10??11 0000?????? 00?0010011 0001100Macroleter 00?1?00000 0?00?01??? 0???1???01 0?110????0 1101011001 0000001Br. baini 0?11000000 00?0101111 0011112?01 ??12100000 1011111110 0100001Br. seeleyi 0011000000 0010101111 0000112001 1112100000 1011111110 0100001Nochelesaurus 0111000000 00???????? ????11200? ????100000 1011111110 0?00001Embrithosaurus 0111000000 0010101011 0111121101 1112110000 ?011111110 0?00001Deltavjatia 11110000?0 0010?11111 ????122001 1012110000 1011011110 0100001Shansisaurus 1111?00??? ?0???????? ????1?1??? ?????????? ?????????? ???????Shihtienfenia 1111?10010 0??0?11211 0????????? ?????????? ?????????? ???????P. peringueyi 1101110110 00?1121211 111122110? ??12212111 1011011110 0100001P. nasicornis ?10?1??1?0 0011121211 11112?1101 ??12212111 1011011110 0100001Pareiasaurus 1111000010 0010111211 0111121101 ????212111 ?011011110 0?00000Scutosaurus 1111000010 0010111211 0111121101 ??12222111 1011111110 0100001Sanchuansaurus ?????????? ?????????? ?????????? ?????????? ?????????? ?????0?Parasaurus ?????????? ???0?11?11 1?1?12??01 ??11?????? 10??01???? 0??0?0?Elginia 1??1?0001? ????111211 ????1?110? ??12222111 10?1011110 0??0010Provelosaurus 120??01??1 10?0?0011? ????1?1?0? ????21111? ?0?1111110 0?00001Anthodon 1201001011 11?0100111 011102011? ????21112? 1011011110 0?00001Pumiliopareia ?????????? ?????????? ?????????? ????21112? 1011001110 0?00001Nanoparia ?????????? ?????????? ?????????? ?????????? ?011011110 0100001Arganaceras ?????????? ?????????? ?????????? ?????????? ?011011110 0?00010Obirkovia ?????????? ?????????? ?????????? ?????????? ????01???? ???????Bunostegos ?????????? ?????11?11 ?????????? ?????????? ?011011110 0?00010
Dow
nloa
ded
by [
76.1
87.6
2.88
] at
23:
11 1
6 M
ay 2
014