transport capabilities of eleven gram-positive bacteria: comparative genomic analyses
TRANSCRIPT

Transport Capabilities of Eleven Gram-positive Bacteria:Comparative Genomic Analyses
Graciela L. Lorca†,1, Ravi D. Barabote1, Vladimir Zlotopolski, Can Tran, Brit Winnen, RikkiN. Hvorup, Aaron J. Stonestrom, Elizabeth Nguyen, Li-Wen Huang, David S. Kim, and MiltonH. Saier Jr.*Division of Biological Sciences, University of California at San Diego, La Jolla, CA 92093-0116
AbstractThe genomes of eleven Gram-positive bacteria that are important for human health and the foodindustry, nine low G+C lactic acid bacteria and two high G+C Gram-positive organisms, wereanalyzed for their complement of genes encoding transport proteins. Thirteen to eighteen percent oftheir genes encode transport proteins, larger percentages than observed for most other bacteria. Allof these bacteria possess channel proteins, some of which probably function to relieve osmotic stress.Amino acid uptake systems predominate over sugar and peptide cation symporters, and of the sugaruptake porters, those specific for oligosaccharides and glycosides often outnumber those for freesugars. About 10% of the total transport proteins are constituents of putative multidrug efflux pumpswith Major Facilitator Superfamily (MFS)-type pumps (55%) being more prevalent than ATP-binding cassette (ABC)-type pumps (33%), which, however, usually greatly outnumber all othertypes. An exception to this generalization is Streptococcus thermophilus with 54% of its drug effluxpumps belonging to the ABC superfamily and 23% belonging each to the Multidrug/Oligosaccharide/Polysaccharide (MOP) superfamily and the MFS. These bacteria also display peptide efflux pumpsthat may function in intercellular signalling, and macromolecular efflux pumps, many of predictablespecificities. Most of the bacteria analyzed have no pmf-coupled or transmembrane flow electroncarriers. The one exception is Brevibacterium linens, which in addition to these carriers, also hastransporters of several families not represented in the other ten bacteria examined. Comparisons withthe genomes of organisms from other bacterial kingdoms revealed that lactic acid bacteria possessdistinctive proportions of recognized transporter types (e.g., more porters specific for glycosides thanreducing sugars). Some homologues of transporters identified had previously been identified onlyin Gram-negative bacteria or in eukaryotes. Our studies reveal unique characteristics of the lacticacid bacteria such as the universal presence of genes encoding mechanosensitive channels,competence systems and large numbers of sugar transporters of the phosphotransferase system. Theanalyses lead to important physiological predictions regarding the preferred signalling and metabolicactivities of these industrially important bacteria.
KeywordsLactic acid bacteria; transport proteins; genomic analyses; energetics
*Corresponding author: Phone: 858-534-4084, Fax: 858-534-7108, E-mail: [email protected].†Current address: Department of Microbiology and Cell Science, University of Florida, Museum Road 1052, Gainesville, FL 326111These two authors contributed equally to the work reported.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptBiochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
Published in final edited form as:Biochim Biophys Acta. 2007 June ; 1768(6): 1342–1366. doi:10.1016/j.bbamem.2007.02.007.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1. IntroductionMembrane transport systems catalyze the uptake of essential nutrients, ions and metabolites,as well as the expulsion of toxic compounds, cell envelope macromolecules, secondarymetabolites and the end products of metabolism. Transporters also participate in energygeneration and interconversion, and they allow communication between cells and theirenvironments. Transport is therefore essential to virtually every aspect of life.
Many Gram-positive bacteria are important in the food industry (see Table 1). These bacteriaare used for the fermentation of vegetables and fruits, for the conversion of dairy products intocheeses and yogurt, and for the preparation of beer and wine. A key feature of lactic acidbacteria (LABs) when grown in laboratory growth media that lack hematin and relatedcompounds is the inability to synthesize porphyrin (e.g., heme). Consequently, LABs grownunder these conditions are devoid of “true” catalase activity and cytochromes. They lackelectron transfer chains and rely on fermentation (i.e., substrate level phosphorylation) for thegeneration of energy. However if hematin is added to the growth medium, catalase andcytochromes may be formed, in some cases resulting in respiration [1–3].
Some LABs are both useful and harmful. For example, Lactobacillus brevis promotes both thepreparation and the spoilage of beer [4]. Other Gram-positive bacteria, such as Lactobacilluscasei, Lactibacillus gasseri, Lactobacillus acidophilus, and Bifidobacterium longum, inaddition to being useful in the food industry, are natural inhabitants of the humangastrointestinal (GI) tract where they synthesize probiotic-like substances. These compoundsstimulate the immune system, provide key nutrients and promote the development of a healthyorganism [5].
The Lactic Acid Bacterial Genomics Consortium has recently carried out genome sequencingof eleven Gram-positive bacterial species [6–9]. The initial shotgun sequencing was conductedat the Joint Genome Institute in Walnut Creek, CA, and genome closure was achievedsubsequently [9]. Nine of the sequenced bacteria are low G+C Gram-positive lactic acidbacteria (LABs) (Pediococcus pentosaceus, Lactobacillus brevis, L. casei, L. delbruekii, L.gasseri, Lactococcus lactis, subspecies cremoris, Streptococcus thermophilus, Leuconostocmesenteroides and Oenococcus oeni) while two of them (Bifidobacterium longum andBrevibacterium linens) are high G+C Gram-positive bacteria. These bacteria exhibitconsiderable diversity with respect to their ecological habitats, being found in milk, meat,plants, the GI tracts of humans and other animals, and elsewhere (see Table 1). They also differwith respect to their carbohydrate metabolic pathways (Table 2). Little is known about thetransport capabilities of these important organisms.
In this review we report comprehensive analyses of the transport capabilities encoded withinthe eleven Gram-positive bacterial genomes noted above. Identification of all homologues ofrecognized transport proteins included in the transporter classification database (TCDB) [6,7] and topological prediction were achieved using established methods [10]. The identifiedtransporters were grouped according to topology, class, and function. The most importantconclusions are summarized in this report.
2. Computer MethodsGenomes of the eleven Gram-positive bacteria were screened for homologues of all proteinscontained in TCDB, a membrane transport protein classification database(http://tcdb.ucsd.edu) [6,11]. FASTA-formatted protein sequences of the completed genomeswere used. Each putative open reading frame was used as a query in the BLAST software[12,13] to search for homologues of proteins in the TCDB sequence library. In addition, each
Lorca et al. Page 2
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ORF was scanned with HMMTOP [14] to predict the number of putative transmembranesegments as reported in Table 3.
All of the potential transport proteins thus identified were subsequently examined in greaterdetail. Each transport protein identified was classified on the basis of sequence similarity intofamilies and subfamilies of homologous transporters based on the classification systempresented in TCDB. Each family/subfamily was noted with its standard abbreviation, its TCnumber and its typical substrates. The substrate specificities of particular homologuesidentified in the sequenced genomes have been predicted based on homology to functionallycharacterized genes and from their genomic context (see Table 1). Assignment to a family orsubfamily within the Transporter Classification System usually allows specification ofsubstrate type with high confidence [6,7,11]. The genome sequencing projects are describedin Klaenhammer et al. [8] and Makarova et al. [9]. Phylogenetic trees (Neighbor joining) werebased on multiple alignments generated with the Clustal X program [15] and drawn using theTreeView program [16].
Some readers may be interested in greater detail than provided in this paper regarding theidentified transporters and the families/superfamilies to which they belong. These readers areadvised to read this paper with the Transporter Classification Database (TCDB) at hand. Thespecificities, polarities of transport, mechanisms of transport, and transport protein and familydescriptions are provided in detail therein. Novel substrates are continually being discoveredfor many transporters, especially drug exporters, some of which function physiologically inthe absence of drugs. Moreover, some transport systems, initially characterized as narrowspecificity systems, prove to exhibit broad specificities when examined in greater detail. Whensuch substrates are known, they can be identified by searching TCDB, a database which iscontinually being updated as new experimental data become available. Table 5 provides ahelpful guide as to where in TCDB to look for specific information. Because of the continualupdating of TCDB, the interested reader who uses TCDB in conjunction with this paper willbe up to date for years to come.
Thousands of published papers have described the functional characterization of transportsystems and their constituent proteins. Several thousand such references can be found in TCDB.It is not possible to include them all in a review such as this one. We apologize to any researcherwhose work we have mentioned without providing the reference(s) to the experimental work.
3. Genome StatisticsBetween 89 and 95% of all predicted transport proteins for the various genomes had significantmatches (e-values <10−3) in TCDB. Remaining predicted transmembrane transport proteinsproved to be more distantly related to proteins included in TCDB. Some of these distantlyrelated proteins could not be classified into the established TC families/subfamilies. They weretherefore listed under the “Not Assigned” (N/A) category (see Table 4). The percentage ofrecognized genes that are predicted to encode transport proteins varies according to themicroorganism analyzed between 12.8% (Lactococcus lactis spp. cremoris) and 18.0%(Oenococcus oeni). Percent transporters encoded within the various genomes showed nocorrelation with genome size (Table 1).
4. Transporter ClassificationFive distinct well-defined types of functionally characterized transport systems, based uponmode of transport and energy coupling mechanism, are recognized by the TC system [6,7,11]. These classes are: (1) channels, (2) secondary carriers, (3) primary active transporters, (4)group translocators of the sugar transporting phosphoenolpyruvate-dependentphosphotransferase system, and (5) transmembrane electron flow carriers. Of these primary
Lorca et al. Page 3
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

classes, classes 1, 3 and 5 are subdivided (see Table 4). In addition, class 8 is reserved foraccessory proteins involved in transport while class 9 specifies incompletely characterizedtransport systems. Class 9 is subdivided into classes 9A (recognized transporters of unknownmechanism) and 9B (putative transporters).
5. Organismal RepresentationTable 1 presents the eleven organisms included in this study. Two of these organisms,Bifidobacterium longum (Blo) and Brevibacterium linens (Bli) are high G+C Gram-positivebacteria and therefore in a bacterial phylum different from that of the other nine organisms,which are classified as low G+C Gram-positive lactic acid bacteria. As expected, thephylogenetic tree based on the 16S rRNAs of the eleven organisms (Fig. 1) shows Blo and Bliclustering loosely together, distant from all other organisms.
Two other organisms cluster together and separately from all others. These are Leuconostocmesenteroides (Lme) and Oenococcus oeni (Ooe) (Fig. 1). Moreover, situated in between thiscluster and the large cluster of remaining lactic acid bacteria is a cluster including Lactococcuslactic spp. cremoris (Lcr) and Streptococcus thermophilus (Sth). Of the five remaining closelyrelated organisms, Pediococcus pentosaceous (Ppe), Lactobacillus brevis (Lbr), andLactobacillus casei (Lca) cluster together as do L. delbruckii (Lde) and L. gasseri (Lga). Alltables included in this study will have the organisms presented according to their phylogeneticgroupings starting with Ppe and proceeding clockwise around the tree shown in Figure 1.
Table 1 presents the approximate genome sizes, G+C content, numbers of identified genes,and percent transport proteins (TP) available for analysis. As can be seen from Table 1, mostof the genomes sequenced are about 2 Mbp in size (1.7–2.8 Mbp), corresponding to 1788–2900 recognized genes, but that of Bli proved to be substantially larger (4.0 Mbp) with 4040genes. Between 12.8 and 18.0% of these genes encode recognizable transport proteins. Exceptfor the two high G+C Gram-positive bacteria, which have 60–62% G+C content, these bacteriahave G+C contents ranging from 35 to 49%.
Table 2 summarizes some of the key properties of these bacteria, particularly with respect tocarbohydrate utilization. This table summarizes the modes of sugar fermentation utilized andalso presents other key properties, particularly those of commercial significance.
6. Topological Distribution Among Transport Proteins (TPs)von Heijne and his associates have published several predictions and statistical analyses oftransmembrane protein topologies [17,18] in specific organisms such as Saccharomycescerevisiae [19], E. coli [20], and over 50,000 bacterial inner membrane proteins [21]. Theseand other studies have revealed that for certain classes and families of transporters, topologicalpredictions are surprisingly reliable although they are much less so for some other families oftransport proteins [22]. The topological results reported here for the 11 Gram-positive bacteriaexamined in this report are in general agreement with previous analyses for transport proteinsin other bacteria [6,23–30].
Table 3 and Figure 2 present the predicted topologies of the transport proteins (TPs) identifiedon the basis of their homology to entries in TCDB. The percentages of these proteins in generaldecrease as the numbers of TMSs increase from 0 to 3; then they increase as the numbers ofTMSs increase from 3 to 6; then they decrease again as the numbers of TMSs increase from 6to 9 before again increasing. There are consistently more proteins with even numbers of TMSsthan odd numbers of TMSs (Fig. 2). For example, there are more 10 TMS proteins than 9 or11 TMS proteins, far more 12 TMS proteins than 11 or 13 TMS proteins, and more 14 TMSproteins that 13 or 15 TMS proteins. These facts undoubtedly reflect the mechanisms by which
Lorca et al. Page 4
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

these proteins arose through evolutionary history, mechanisms that involved intragenicduplication [28]. Only Bli has proteins predicted to have homologues of 17 and 19–25 TMSs(see Table 3), while only Ppe and Lca have proteins with 18 putative TMSs (one such proteinper organism).
The greatest variation between the organisms with respect to the numbers of TP homologueswith a single predicted topology are the 5 and 12 TMS proteins. Bli has the most predicted 12TMS TPs (18.8%) and the least 5 TMS TPs (2.4%) while Lde and Sth have the reciprocalrelationship: 10.0% and 7.4% 5 TMS proteins but only 4.1% and 4.9% 12 TMS proteins,respectively. This correlates with the percentages of integral membrane proteins of MFS-typesecondary active transporters versus ABC-type primary active transporters (see below). ABCuptake systems most commonly have integral membrane constituents of 5 TMSs perpolypeptide chain while MFS permeases usually have 12. It is noteworthy that proportions of5 or 12 TMS proteins in Bli and Lde relative to the other organisms do not correlate with therelative proportions of 4 and 6 TMS proteins, or 11 and 13 TMS proteins, respectively. Thisfact suggests that the TMS predictions are remarkably accurate, at least for the members of theMFS and ABC superfamilies. Since Lde (with 4.1% of its TPs having 12 TMSs) clusters inFig. 1 with Lga (with 11.7% of its TPs having 12 TMSs), it is clear that TP topologicaldistribution does not correlate with organismal phylogeny. The percentage of 12 TMS TPs forLga is the fifth largest of the eleven organisms examined (Table 3). Similarly, although Lbrand Lca cluster together in Figure 1, they exhibit about a 2-fold difference in numbers of 12TMS proteins. Based on these analyses, we conclude that transporter topology (Table 3) andmode of energy metabolism (Table 2) do not correlate well with organismal phylogeny (seebelow).
Three additional observations are worthy of note: (1) Blo has 16.7% 6 TMS proteins, morethan any other organism examined; (2) Lde, which has the greatest percentage of 5 TMSproteins but the lowest percentage of 12 TMS proteins, also has the lowest percentage of 8TMS proteins (Fig. 2), and (3) only Bli has proteins of greater than 18 TMSs. These facts mustall be explained in terms of the types of transporters employed by these organisms.
7. Classes of TPs in the 11 organismsTable 4 summarizes the distribution of TPs in the seven TC categories, and these values fordefined classes 1–4 are shown in bar graphs in Figure 3. Predicted transport proteins that couldnot be classified with certainty into specific TC families are indicated in the Not Assigned (N/A) category (Table 4). All organisms have only a small percentage of their TPs as channelproteins (5–14 proteins per organism or 1.7–4.1%). Almost all of these channel proteins areα-helical type channels of subclass 1.A. No β-type channels of class 1.B were detected, andvery few channel-forming toxins (class 1.C) were identified; none of the latter was found forLbr, Lca, Lde, Sth, Ooe, Blo and Bli. Ppe and Lcr each have 1, Lga has 2 and Lme has 6. Thepresence of class 1.A α-type channel proteins in these Gram-positive bacteria may relate to theneed of these organisms to respond to stress such as osmotic shock [31]. Channel-formingproteins will be analyzed in greater detail below.
Secondary carriers (such as major facilitator superfamily (MFS) carriers; class 2) usuallyconsist of single polypeptide chains, but primary active transporters (such as ABC carriers;class 3) usually consist of heterooligomeric complexes. The numbers of TPs listed in class 2therefore probably accurately reflect the numbers of secondary carriers, but the numbers ofTPs listed in class 3 are greatly in excess of the numbers of transport systems representedbecause of their multicomponent nature, probably by a factor of 3 or more. These facts mustbe taken into account when evaluating the data presented in Table 4 and Figure 3. Bli has byfar the highest class 2/3 ratio while Sth and Lde have by far the lowest, in agreement with the
Lorca et al. Page 5
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

topological analyses discussed above. Excluding these three extreme organisms, this ratio forthe remaining eight organisms studied differs by less than a factor of two.
Almost all of the primary active transport proteins are ATP hydrolysis-driven systems of class3.A. Only one organism (Lca) has the transmembrane β-subunit of a decarboxylase-driventransporter, and consequently only Lca is predicted to have a decarboxylase-driven Na+ pumpof class 3.B. Only five organisms (Lbr, Lcr, Ooe, Blo and Bli) have subunits homologous tothose of some form of oxidoreductase-driven cation transporting systems (class 3.D). However,since most of these electron transfer complexes are multisubunit, it is possible that only Blipossesses complete proton pumping electron flow systems (see Table 5).
Class 4 proteins (PTS) show tremendous variation in representation among the elevenorganisms. Thus, Blo has only one recognized PTS permease, and this system is in the Glucose(Glc) Family (4.A.1). Lbr has just 7 recognized PTS protein homologues with four EnzymeIIC permeases, two of the Lac family, one of the Gat family (just the IIC constituent), and oneof the mannose (Man) family. Of these, only the Man family system is apparently a full EnzymeII complex with all requisite constituents. Lca has the most (63) PTS protein homologues withall 7 PTS families represented. This organism has between 1 and 8 intact PTS permeases perPTS family with 8 representatives in the Glc family and 1 in the Gut family. Lga and Ppe have36 and 37 PTS proteins, respectively. These two organisms possess complete glucose (Glc),fructose (Fru), lactose (Lac) and mannose (Man) family systems, and Lga also has a galactitol(Gat)-type system. Lcr and Ooe encode in their genomes 19 and 30 PTS proteins, respectively.Both have glucose, lactose, mannose and L-ascorbate (Asc)-type systems, and Lcr also has afructose-type system while Ooe additionally has a galactitol-type system. There is a reasonablecorrelation between prevalence of PTS permeases (Table 4) and organismal phylogeny (Fig.1).
Transmembrane electron carriers that transfer electrons from one side of the cytoplasmicmembrane to the other (TC class 5) are lacking in 10 of the 11 organisms. Bli is the onlyexception. It has 7 such proteins (1.3%) with representation in the DsbD and PMO families.
The eleven organisms have between 4 and 13 auxiliary transport proteins each (class 8). Allexcept Bli possess Enzyme I and HPr of the phosphotransferase system (PTS). Two organismseach possess a membrane fusion protein (MFP) family member, which in Gram-positivebacteria usually facilitates export of peptides via an ABC efflux pump [32]. Several organismshave homologues of the K+ transport/nucleotide binding regulatory protein (KTN) family, butthese homologues do not necessarily function to regulate transport and may serve an unrelatedfunction.
Of the poorly characterized class 9 transporters, the percentages are fairly constant in the 11organisms (3.6–7.4%). Within subclass 9A, we find putative transporters for heavy metals,including Fe3+ (OFeT and FeoB), Pb2+ (PbrT), Hg2+ (MerTP), and Mg2+ (MgtE). Threeorganisms also possess homologues of the nicotinamide mononucleotide (NMN) uptakepermease (PnuC) family.
Within subclass 9B, several potentially important families are represented. However, theevidence that many of these function in transport is marginal or nonexistent. For example, allor most members of the FAT family of acyl CoA synthetases may not play a role in fatty acidtransport [33]. The PerM, TEGT and YbbM families include 7 TMS protein members, and theYdjX–Z and YqiH families include 5 or 6 TMS members. The HCC family includes membersthat may be pore-forming hemolysins, and VGP family members may catalyze vectorialpolymerization of polysaccharides destined to be extracellular. Of greatest interest is thePutative Bacterial Murine Precursor Exporter (MPE) family (9.B.3) which is represented in all
Lorca et al. Page 6
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

11 organisms, usually with 2 or 3 paralogues per organism. These putative transporters maybe required for synthesis of the bacterial cell wall.
8. Channel-forming ProteinsAs summarized in Table 5, all of the organisms examined contain recognized α-type channelproteins of TC class 1.A. These vary in number from 5 for Lde and Bli to 11 for Lcr.
Three channel protein families (Voltage-gated Ion Channel (VIC), Large ConductanceMechanosensitive Channel (MscL) and Small Conductance Mechanosensitive Channel(MscS)) as well as some aquaporins of the MIP family (see below) may function primarily toprotect the bacteria against osmotic stress [31,34–41]. All of the 11 bacteria examined have atleast one such channel suggesting that relief from osmotic stress is of general importance tothese bacteria as has been demonstrated for Lactococcus lactis [31]. However, the numbersand types of these channels vary. For example, five organisms (Ppe, Lbr, Lga, Ooe and Blo)have at least one of each of these three channel types. Lca, Lde and Lcr have two such channelproteins, one of the MscL family and one of the MscS family. In L. lactis, only the mscL geneis apparently expressed although both the MscL and MscS channel proteins from this organismare functional [31]. Lme is the only organism with just one of these channel proteins (MscL).In fact, Lme is the only bacterium that lacks an MscS channel homologue. Surprisingly, noorganism has more than one member of either the VIC, MscL or MscS family. Multiple MscSfamily paralogues are found in many bacteria although few bacteria have more than one MscLchannel [39].
The eleven organisms have from one to three chloride channel proteins of the ClC family (Table5; ref. 7). Thus, Ppe, Lbr, Lca, Lde and Ooe have just one of these anion-selective channels;Lga, Lcr, Lme, Blo have 2, and Sth has three. In E. coli, these channels are known to promoteresistance to extreme acid stress [42].
All eleven bacteria contain at least one heavy metal ion channel of the MIT (CorA) family[43,44], and five organisms have between two and five members. These divalent metal ionchannels are important for the uptake of Mg2+, Co2+ and Ni2+ in other bacteria in response tothe membrane potential, negative inside [45]. It can be presumed that these channels providethe same function in the organisms examined here.
Glycerol facilitators and aquaporins of the MIP family allow transmembrane passage of severalsmall neutral molecules including glycerol and other polyols, dihydroxyacetone, water, CO2,urea and ammonia [46–48]. In prokaryotes, most MIP family members transport glycerol astheir primary function. However, synthesis of aquaporins has been shown to be induced aftertransfer of certain bacteria to hyperosmotic media [49,50]. This fact suggests a role in osmoticshock protection.
Members of the MIP family of aquaporins and glycerol facilitators are found in all 11 bacteriaexamined here. Four of the LABs have a single putative glycerol facilitator, and the rest havetwo or more of them. Lactic acid bacteria in general cannot utilize glycerol as a sole carbonsource [51], but they can often co-metabolize glycerol and glucose [52]. The universaloccurrence of glycerol facilitators in these organisms may therefore reflect their abilities to useglycerol in the presence of a second carbon source. Five of the LABs (Ppe, Lbr, Lme, Blo andBli) have putative aquaporins that may play a role in osmotic shock responses. A singlebacterium (Sth) has a urea/short chain amide (UAC) family member. This protein may allowuptake of various amides as sources of nitrogen [53,54] (Table 5).
Hsp70 chaperone proteins are listed in Table 5 under class 1.A. This is because some eukaryoticHsp70 proteins have been shown to insert into membranes to form channels [55,56]. All of the
Lorca et al. Page 7
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

organisms examined here have such a homologue. These proteins function primarily in proteinfolding as part of the general stress response system [57–60]. There is no evidence for channelformation by Hsp70 homologues in prokaryotes. However, merely because of thedocumentation of channel formation in eukaryotic homologues, we have included thesechaperones in this tabulation.
Several toxins of TC class 1.C were identified. In general, these are rare in the lactic acidbacteria examined. Six members of the large protein CCT toxin family were identified in Lme,but none was found in the other ten organisms analyzed. Members of this family are found inboth Gram-negative and Gram-positive bacteria [61,62]. Surprisingly, Lga has two colicinhomologues. Colicins are usually restricted to enteric bacteria. Finally, two of the organisms(Ppe and Lcr) encode peptide toxins (bacteriocins), Pediocin and Lactococcin A, respectively,which are exported to exert their toxic activities on other microorganisms [63,64].
Ppe, Lbr, Lcr and Sth each encode either one or two recognizable holin(s). These smalloligomeric proteins may promote cell death in response to stress stimuli by allowing export ofautolysins that digest the bacterial cell wall [65]. These proteins might therefore play a role inprogrammed cell death [66–68].
9. Secondary Carrier Superfamilies9.1. MFS (2.A.1)
There is tremendous variation in the numbers of secondary carriers in the different bacteriaexamined, and the same is true for the individual families of secondary carriers. The largestsuch family is the major facilitator superfamily (MFS) [69,70]. Sth has 5 such carriers whileBli has 70. All other bacteria have intermediate numbers (Table 5). Lde has 14, Ooe has 41and all other bacteria have between 21 and 35. Of equal interest are the probable substratespecificities of these transporters. In all but two of the bacteria analyzed, Lga and Bli, drugexporters comprise the majority of all MFS permeases. This is surprising since only 3 of the47 recognized MFS families are primarily concerned with export of hydrophobic andamphipathic substances. These three MFS families (called DHA1–3) are represented in alleleven bacteria with the exceptions of Ppe which lacks a DHA3 member. In Sth, 3 of the 5MFS permeases are putative drug exporters; in Lde, 8 of the 14 have this function, and in Lcr,20 of the 24 MFS homologues may export drugs. Of all the organisms analyzed, the high G+C Gram-positive bacterium, Bli, has the most non-drug-specific MFS transporters. Thus, inBli, 28 of the 70 MFS permeases probably act on drug-like substances, and more of thesesystems transport anionic compounds than drugs. It should be noted that DHA1–3 exporterscan act on a wide variety of compounds including hydrophilic metabolites (see TCDB).However, members of these three families always catalyze substrate efflux [69].
The sugar uptake systems of the MFS include members of the sugar porter (SP) family and theoligosaccharide:H+ symporter (OHS) family as well as the fucose:H+ symporter (FHS),sialate:H+ symporter (SHS) and polyol porter (PP) families. These families are poorlyrepresented in most of these Gram-positive bacteria. In fact, all but Lbr (5), Ooe (7), Blo (6)and Bli (5) have just 1–3 such systems.
Virtually all remaining MFS families represented (see Table 5) consist of permeases withspecificity for inorganic and organic anions. With the sole exception of Bli, these aniontransporting MFS permease families are poorly represented. The BST family is found only inLga which has 3 members and in Lca and Lde which have 1 member each. Multiple paraloguesof the OFA, ACS and AAHS families are found only in Bli, which has 2, 2 and 5 members ofthese three families, respectively, although Sth has three AAHS family paralogues. It is clear
Lorca et al. Page 8
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

that Bli exhibits a diversity of MFS permeases that far exceeds that of the other organismsexamined.
9.2. GPH (2.A.2)In view of the paucity of MFS uptake systems for sugars, it is surprising that putative reducingoligosaccharide and non-reducing glycoside uptake systems of the GPH family (2.A.2) arerepresented in all of the eleven organisms except Bli, the one organism with by far the mostMFS permeases. The other bacteria have between 1 and 10 GPH permeases. Of the lactic acidbacteria, only Lca, Lde, Lga and Ooe have more MFS sugar permeases than GPH permeases.All functionally characterized members of the GPH family take up oligosaccharidespreferentially (hence the name Glycoside-Pentoside-Hexuronide:Cation Symporter Family(TC #2.A.2)). Some reports suggest that reducing sugars may also be substrates, but thissuggestion has been disputed for some GPH members [71,72]. This suggests that most lacticacid bacteria preferentially take up oligosaccharides and non-reducing glycosides over simplesugars via secondary carriers since almost all members of the GPH family take up glycosidesand oligosaccharides rather than reducing monosaccharides [71,73–75]. This observation mayreflect the fact that the former compounds are common plant products. They are also generatedin animals by the degradation of glycolipids and glycoproteins [76,77]. In lactic acid bacteria,simple hexoses may be taken up primarily via the PTS (see below).
9.3. APC (2.A.3)The Amino Acid/Polyamine/Organocation superfamily, the largest superfamily of amino acidand amine uptake transporters [78], is represented in all 11 Gram-positive bacteria studied here.Seven to 18 members of this superfamily are encode in each of the various genomes. Of theeleven currently recognized families within the APC superfamily, the AAT family (2.A.3.1)is most prevalent with 1–8 representatives per genome. However, ten of the eleven APCfamilies are found in these Gram-positive bacteria, a most surprising observation sincemembers of three of these families had not been identified in bacteria when this superfamilywas last described [78]. These three families are the ACT (2.A.3.4), LAT (2.A.3.8) and YAT(2.A.3.10) families. The ACT family is represented only in Bli (2 members), and the YATfamily is found only in Ppe (1 member), Lde (2) and Sth (1). However, members of the LATfamily are found in six of the eleven organisms (1–2 members each).
The glutamate:γ-aminobutyrate (GABA) antiporter (GGA) family (2.A.3.7) is found with from1 to 5 members in 7 of the 11 bacteria. All bacteria except Lde have 1–3 members of the cationicamino acid transporter (CAT) family. Two organisms (Ppe and Bli) have 1 member each ofthe ethanolamine transporter (EAT) family. One to 4 members of the archaeal/bacterialtransporter (ABT) family are found in 6 of the 11 organisms, the exceptions being Lbr, Lca,Lde, Ooe and Blo. Four organisms (Ppe, Lbr, Lca and Lga) have 1–2 members of the aspartate/glutamate transporter (AGT) family, while four organisms (Ppe, Lbr, Lde and Lcr) have 2members of the basic amino acid/polyamine antiporter (APA) family. It is clear that theseorganisms can take up a wide range of amino acids and their derivatives. The characteristicsof these families and primary references describing their members can be found in TCDB[78].
9.4. RND (2.A.6)The RND superfamily of efflux pumps is represented in 7 of the 11 organisms studied (1–7members per organism) (Table 5). This superfamily includes eight currently recognizedfamilies. Four of these families (HME, HAE1, NFE and ORF4) predominate in Gram-negativebacteria; a fifth (SecDF) is found in both Gram-negative and Gram-positive bacteria; a sixth(HAE2) is found only in Gram-positive bacteria; a seventh (HAE3) is found primarily inarchaea; and the eighth (EST) is restricted largely to eukaryotes [79].
Lorca et al. Page 9
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The HAE1 family of drug and lipid exporters is represented only in Bli (2 member), as is theSecDF family of protein secretion factors (1 member). However, the HAE2 family of drug andlipid exporters is found in 7 of the 11 organisms with 1–3 members each. No other RND familyis represented. It is clear that the RND superfamily is underrepresented in these bacteria.
9.5. DMT (2.A.7)The Drug Metabolite Transporter (DMT) superfamily consists of 18 currently recognizedfamilies [80], seven of which are represented in the Gram-positive bacteria under study. Infact, all eleven organisms have DMT representation (2–7 members/organism) (Table 5).
Of the 18 families, by far the greatest representation is in the Drug/Metabolite Efflux (DME)family, the largest of the prokaryotic families found in the DMT superfamily. Ten of the elevenorganisms have DME pumps, and each organism may have from 1 to 4 members. Six organisms(Ppe, Lbr, Lca, Lcr, Lme and Bli) have small multidrug resistance (SMR) export systems, andthe 9 lactic acid bacteria have representation in the Glucose/Ribose Uptake Porter (GRP) family(1–2 members). These proteins probably take up glucose via a proton symport mechanism[81]. Only the two high G+C Gram-positive bacteria (Blo and Bli) lack GRP family members.
Members of other DMT families are sparsely represented. For example, Lca has 1 member ofthe choline uptake transporter (LicB–T) family, Sth, Lme and Bli have 1, 1 and 2 members,respectively, of the chloramphenicol-sensitivity protein (RarD) family, while Blo has a singlemember of the paraquat (methyl viologen) exporter (PE) family. Surprisingly, Sth has amember of the plant drug/metabolite exporter (P-DME) family, which, however, is closelyrelated to the prokaryotic DME family.
9.6. SSS (2.A.21)The Sodium:Solute Symporter (SSS) superfamily [82] was not represented in the low G+CGram-positive bacteria studied but was found in the high G+C Gram-positive bacteria. Bli hasnine members while Blo has one. The single SSS protein in Blo may be a proline uptake porter,and the SSS proteins in Bli most closely resemble sodium iodide (3), proline (2), sugar (2),monocarboxylate (1), and phenylacetate (1) uptake permeases of other organisms.
9.7. MOP (2.A.66)The multidrug/oligosaccharidyl-lipid/polysaccharide (MOP) superfamily [83] is present in alleleven bacteria under study with 2–7 members each. The vast majority of these belong to thepolysaccharide transporter (PST) family within the MOP superfamily. Most organisms have1–7 such proteins, although Lbr and Blo lack these proteins. These exporters catalyze secretionof a variety of polysaccharides and their precursors using a proton antiport mechanism. All but2 of the organisms have at least one MATE-type multidrug resistance pump with 1–3 suchsystems each. Three organisms, a single low G+C Gram-positive bacterium (Lcr) and the twohigh G+C Gram-positive bacteria (Blo and Bli) have representation in the bacterial MouseVirulence Factor (MVF) family. The transport substrates of members of this family are notknown.
10. Smaller Families of Secondary Carriers10.1. Divalent cations
Monovalent and divalent cation concentrations must be strictly controlled for optimal growth.Bacteria therefore possess channels, carriers and primary active pumps that regulate theseconcentrations. Moreover, among the carriers, some are strict cation:cation antiporters, othersare electrogenic uniporters, and still others are cation:cation symporters as documentedparticularly well in E. coli [84–87].
Lorca et al. Page 10
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Five families of carriers for divalent cations are represented in these Gram-positive bacteria.The Calcium:Cation Antiporter (CaCA) family (2.A.19) is found only in Bli. These systemscatalyze Ca2+ efflux. The Cation Diffusion Facilitator (CDF) family (2.A.4) is found in 10 ofthe 11 organisms. Only Ppe lacks representation. These are likely to export heavy metal divalentcations such as Zn2+, Cd2+ and Co2+. The Ni2+-Co2+ Transporter (NiCoT) family (2.A.52) isfound only in three organisms (Ppe, Lbr and Lme). All functionally characterized members ofthis family function in the uptake of Ni2+ and/or Co2+. The Nramp family (2.A.55) isrepresented in all but two (Lde and Lga) of the eleven organisms examined, and these 9organisms have between 1 and 3 members. Nramp carriers take up a variety of heavy metals(Fe2+, Mn2+, Zn2+, Cu2+, Cd2+, Ni2+, Co2+, Pb2+) using a cation symport mechanism. Thepoorly characterized CadC family (2.A.77) is also represented in Table 5. Five organisms eachhave 1–3 members of this family of putative Cd2+ efflux pumps.
10.2. Monovalent cationsNine families of monovalent cation transporters are represented in Table 5. None of these isfound in all of the eleven organisms. NhaA family (2.A.33) members are found only in thehigh G+C Gram-positive bacteria, Blo and Bli, each having one member. The NhaC family(2.A.35) is found in 7 of the 11 organisms with 1 or 2 members each. The CPA1 family (2.A.36) was found in all organisms except Lde while the distantly related CPA2 family (2.A.37)was lacking only in Sth and Blo. A maximum of 4 members of each of these cation:protonantiporters is encoded in any one genome.
The Trk family (2.A.38) of monovalent cation symporters is represented in 6 of the 11organisms, but one bacterium (Sth) has three of these systems while another (Bli) has four. Asingle protein of the NhaD family (2.A.62) of Na+/H+ antiporters is found only in Lca.
Subunits of the CPA3 family of K+:H+ and Na+:H+ antiporters (2.A.63) are found only in Bli.Normally these systems have 7 subunits, but 10 are reported for this organism. Possibly thesesubunits are constituents of two distinct systems. Finally, the KUP family (2.A.72) of K+ uptakesystems is found in 7 of the 11 bacteria, and the Amt family (2.A.49) of NH3 channel proteins[88] is found in nine. This last family was previously thought to transport NH4
+ by a carrier-type mechanism, but high-resolution x-ray structures of the E. coli AmtB protein clearlysuggest that at least this member of the family deprotonates NH4
+ at the external surface ofthe pore and transports NH3 via a simple hydrophobic channel-type mechanism [88,89].
10.3. AnionsSeventeen families of secondary carriers represented in Table 5 include members that probablytransport anionic compounds. Most of these are specific for organic anions that can be used assources of carbon (gluconate, ketodeoxygluconate, tri-, di- and monovalent acids such ascitrate, succinate and lactate, respectively, and bile salts), but several can transport inorganicanions as well (formate, nitrate, arsenite, sulfate, phosphate). Perusal of Table 5 reveals thatthe gluconate porter (GntP) family (2.A.8) is represented in seven organisms. Three additionalwell-represented families (CCS, DASS and DAACS) primarily transport organic anions whilethree other well-represented families (PiT, FNT and SulP) transport inorganic anions. Otheranion transporting families are represented to a lesser degree.
Interestingly, the poorly characterized auxin efflux carrier (AEC) family (2.A.69) is found in10 of the 11 organisms examined with between 1 and 5 members per organism. The fewcharacterized prokaryotic members of this family catalyze uptake of dicarboxylates such asmalate and malonate [90,91]. Many of these anion-transporting families are members of theIT superfamily [92].
Lorca et al. Page 11
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

10.4. Amino acids and peptidesIn addition to the APC superfamily, several other families represented in Table 5 includemembers that catalyze amino acid uptake [93]. The NSS family (2.A.22) is represented onlyin Bli. Glycine and alanine permeases of the AGCS family of semipolar amino acid uptakeporters (2.A.25) are represented in 9 of the eleven organisms.
Three families of amino acid efflux porters are represented in Table 5. The first of these is theLysE basic amino acid exporters (2.A.75), present only in Bli. The LysE family is a memberof the LysE superfamily, all members of which catalyze substrate efflux [94]. The secondamino acid efflux family represented is the RhtB semipolar amino acid exporters (2.A.76),present in Sth, Lme and Bli. This family is also a member of the LysE superfamily [94]. Finally,members of the LIV-E hydrophobic amino acid exporter family (2.A.78) are present in Ppe,Lbr, Lcr, Sth, Lme, Blo and Bli. The LIV-E family is unrelated to the LysE superfamily, andall characterized members of this family export hydrophobic amino acids [95,96].
Peptide uptake permeases represented include those of the POT family (2.A.17) found in 7organisms, the OPT family (2.A.67) found in 2 organisms, and the AbgT family (2.A.68) foundin 1 organism. The Tat family of pmf-driven protein secretion permeases (2.A.64) was foundin Sth and Bli. The Oxa1 family (2.A.9), involved in the insertion of integral membrane proteinsinto the plasma membrane bilayer, is represented in all organisms examined (Table 5).
10.5. Nucleobases and nucleosidesThree families found in Table 5 are concerned primarily with the uptake of nucleobases andnucleosides. These are the NCS1 family (2.A.39) (4 organisms represented), the NCS2 family(2.A.40), present in all 11 organisms, and the CNT family (2.A.41), present in 4 of the 11bacteria. Multiple such systems are present in all eleven bacteria.
11. Primary Active TransportersPrimary active transporters couple a primary source of energy to transport, and they areconsequently often multicomponent and always multidomain systems. They couple transportto processes such as ATP hydrolysis, organic acid decarboxylation and electron transfer. Aquick look at Table 5 reveals that each of the eleven organisms has multiple (66–175) ABCsystem proteins, a single F-type ATPase, always with 8 dissimilar subunits as expected, several(4–13) P-type ATPases, and a number of ATP-driven macromolecular transport systems. Oneorganism has an organic acid decarboxylation-driven Na+ exporter, but with the exception ofBli, none has a monovalent cation (H+ or Na+)-translocating electron carrier. The individualfamilies of primary active transporters and the members identified will be discussed below. Inthe following sections we will discuss homologues of known primary active transporters.
11.1. The ABC superfamily (3.A.1)ABC transporters segregate phylogenetically into uptake and efflux systems [97,98]. Theformer but not the latter act in conjunction with extracytoplasmic solute binding receptors (R)that feed the solute into the transmembrane channel (M) [99] which is energized by thecytoplasmic ATP hydrolyzing subunit/domain (C) (see TCDB). The receptors can be lipidanchored or attached to non-cleavable, N-terminal, leader sequences. In some cases they areeven fused to the transmembrane transporter component. Sometimes several receptors can feeddifferent, but structurally related solutes into a single ABC channel [100]. Transport isenergized by the hydrolysis of ATP, catalyzed by a membrane-associated ATP-binding cassette(ABC) protein or protein domain [101].
Lorca et al. Page 12
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Both the integral membrane channel forming protein (CFP; M) and the ABC protein (C) areusually present as dimers, each consisting either of a single gene product, yielding ahomodimer, or two gene products, yielding a heterodimer [102]. Sometimes, but seldom, thesethree protein types are fused to form multidomain proteins. ABC uptake permease systemstherefore normally consist of 3–5 distinct proteins although the number can be smaller (due togene fusions) or larger (due to the presence of multiple receptors) (see TCDB). The averagenumber of proteins per ABC uptake system is roughly 4.
By contrast, ABC efflux systems consist only of the ABC (C) and CFPs (M), and these twodomains are often fused. Consequently, there are either one or two proteins per ABC exporter;thus, about 1.5 proteins are present on an average per ABC exporter. The quantitation presentedin Table 5 is always in terms of numbers of proteins; consequently, these numbers should bedivided by about 4 and 1.5 for families of uptake and efflux systems, respectively, in order toestimate the numbers of systems represented.
As summarized in Table 5, ABC transporters occur in substantial numbers in all Gram-positivebacteria studied. The numbers of ABC proteins encoded within the various genomes vary from66 (for Ppe) to 175 (for Bli). The average number of ABC system proteins per organism is 127.
Two families within the ABC superfamily are concerned exclusively with carbohydrate uptake,the carbohydrate uptake transporter-1 and -2 (CUT1 and CUT2) families. While CUT1 systemsare largely concerned with oligosaccharide uptake, CUT2 systems usually exhibit specificityfor monosaccharides, particularly aldopentoses and hexoses. CUT1 homologues are found inall organisms except Ppe, Sth and Lme. Blo has 40, probably comprising about 10 systems. Itis interesting to note that Bli, with a larger genome, has only 5. Other bacteria probably have1–4 such transporters. Lcr and Ooe probably have four such systems. CUT2 family homologuesare found in all organisms studied, but the numbers of CUT2 family proteins never reach thelarge numbers sometimes observed for CUT1 family proteins. Note that the PepT family (3.A.1.5) includes members that like CUT1 family members can transport oligosaccharides (seebelow).
The two principal amino acid uptake families in the ABC superfamily (PAAT and HAAT) arespecific for polar and hydrophobic amino acids, respectively. Representation in the formerfamily (11–32 proteins) far exceeds that in the latter family (0–11 proteins). The PepT family,which includes members that transport peptides and oligosaccharides, is well represented (6–40 proteins per organism). The methionine uptake transporter (MUT) family (TC #3.A.1.24)is represented in all 11 organisms.
The next few uptake families listed in Table 5 are largely concerned with inorganic iontransport. Sulfate-transporting SulT family homologues are found in 5 of the 11 bacteria, butall 11 organisms have at least one phosphate transporter of the PhoT family, and five also havea phosphonate transporter of the PhnT family.
The POPT family includes a diverse group of transporters exhibiting specificity forpolyamines, opines and phosphonates. One or two systems are present in all bacteria exceptPpe and Blo. Similarly, at least one (and sometimes 2 or 3) QAT family systems for the uptakeof quaternary amine osmolytes and amino acid derivatives are present in all but threeorganisms. TauT family systems, specific for taurine, are found in a few bacteria, and thesesystems may, in addition to allowing utilization of taurine, provide defense against osmoticstress.
The next several families are concerned with heavy metal uptake. They include the FeCT andBIT iron (complex) transporters, the MZT, Mg2+, Mn2+, Zn2+, Fe2+ transporters, the CoTcobalt uptake systems and the NiCoT nickel/cobalt uptake transporters. Surprisingly, the MZT
Lorca et al. Page 13
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

systems are present in all eleven bacteria although the FeCT family is present in only 5 of them,and the Bit and CoT families are represented in only three. A single NitT nitrate/nitritetransporter is found in Lbr. The NiCoT family is represented in all eleven organisms.Homologues of the putative yersiniabactin Fe3+ uptake transporter (YbtPQ) family were foundin Lbr, Lca and Ooe. Members of all other ABC uptake families are absent from all elevenorganisms.
ABC efflux systems fall into two principal subdivisions, those found primarily in prokaryotes(the 3.A.1.100 series) and those found primarily in eukaryotes (the 3.A.1.200 series). Althoughthe capsular polysaccharide exporter (CPSE) family is lacking in the 11 organisms examined,the Lipooligosaccharide Exporter (LOSE) family, previously characterized only in Gram-negative bacteria, is represented in 7 of the 11 organisms. Members of the LipopolysaccharideExporter (LPSE) and Teichoic Acid Exporter (TAE) families are also represented. AlthoughGram-positive bacteria do not make LPS, they do incorporate teichoic acids into their cell walls[103].
Four of the six prokaryotic ABC drug efflux families (Drug E2, Drug RA1, Drug RA2 andMacB) are all nearly universally present in the eleven bacteria, and the two remaining drugexport families (Drug E1 and Drug E3) are also present in select organisms. It is interestingthat many of these organisms seem to have multiple MacB macrolide exporters. Three bacteriamay also have eukaryotic-type MDR pumps, and nine may have a member of the β-exotoxin1 Exporter (βETE) family.
Four of the five ABC peptide exporter families are represented in Table 5 (the Pep1, 2, 4 and5E families). While Pep3E pumps are lacking in all organisms, and a Pep1E family memberis present in each of four organisms, the other three families, Pep2E, Pep4E and Pep5E, arewell represented. Multiple such peptide exporters are found in many of the bacteria.Homologues of the protein exporter (Prot1E) family are found in four of the eleven organisms,while a single homologue of the Prot2E family is present in Bli.
A few other ABC exporter families are represented in Table 5. These include the poorlycharacterized DevE family of putative glycolipid exporters, the LPT family of lipoproteintranslocases, and the lipid exporter (LipidE) family. Two of these families (DevE and LPT)occur in 8 of the 11 organisms examined. Most surprisingly, the LipidE family is representedin all of the 11 organisms except Ppe with as many as 11 such proteins per organism.
11.2. P-type ATPases (3.A.3)The eleven organisms studied have between 4 and 13 P-type ATPases per organism. These canbe classified according to the substrates most likely transported, based on sequence similarityto functionally characterized systems (data not shown).
One organism (Lca) has a protein most closely resembling eukaryotic Na+, K+ ATPases of P-ATPase subfamily 1, a novel finding for a prokaryote. In eukaryotes, these enzymes catalyzeuptake of 3K+ coupled to efflux of 2Na+ in an electrogenic process [104]. All organisms exceptBli have multiple Ca2+/Mn2+ ATPases of subfamily 2. The numbers of these systems perorganism vary between 1 and 5.
In subfamily 3, which includes the characterized Mn2+/Cd2+ ATPase of Lactobacillusplantarum [105] as well as H+-translocating ATPases of fungi and protozoans, we find singlemember representation in two organisms, Lbr and Sth. Lca, Lde, Lga and Lcr each have twosuch members. These four bacteria also have one member of subfamily 4, similar to theMg2+/Ni2+ ATPase of Salmonella typhimurium.
Lorca et al. Page 14
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

All organisms have at least one, and as many as 3 Cu+, Ag+-ATPases of subfamily 5. InEnterococcus hirae, two such systems are characterized (TC #3.A.3.5.1 and 3.A.3.5.2) [106].One catalyzes copper uptake, the other copper efflux, presumably allowing proper maintenanceof copper homeostatic conditions in the cell [106].
Finally, all but two organisms (Sth and Blo) have 1–3 broad specificity heavy metal (Zn2+,Co2+, Cd2+, Hg2+, Ni2+, Cu2+, Pb2+ and Ag2+) efflux pumps of subfamily 6. Surprisingly, noK+ ATPases of subfamily 7, no phospholipid flippases of subfamily 8, and no monovalentalkali cation (Na+, K+) pumps of subfamily 9 were detected.
11.3. Macromolecular secretion systemsConstituents of three protein export systems as well as two DNA translocases were identified.The general secretory pathway (Sec) complex is ubiquitous, being found in every organismfor which it has been sought [107]. However, it can consist of as many as 8 proteins and onesmall (4.5 S) RNA, as is true for E. coli, or as few as 6 proteins plus a small RNA, as is truefor Mycoplasma genitalium. The SecA, SecY, SecE and SecG proteins as well as the Ffh andFtsY proteins (also required for membrane protein insertion) are always present. When proteinsare inserted into the membrane, YidC homologues of the Oxa1 family (2.A.9) often participate[108]. The SecDF complex (homologous to RND transporters), which serves an auxiliaryfunction, is present only in B. linens. Surprisingly, B. linens also has two SecA proteins. Finally,only Ppe has two SecYs, one for general secretion and one for specific secretion of large cellsurface adhesions [109–111]. The latter system uses different auxiliary proteins, the Asp1–4proteins. While Ppe has Asp2–4, Lme has Asp1–3 but lacks a second SecY. What these proteinsare doing in these two bacteria has yet to be determined (Table 5).
Gram-negative bacteria have multicomponent conjugal protein/DNA export systems. TheGram-positive bacterium, Staphylococcus aureus, also has such a system, the Trs system,which consists of 15 dissimilar proteins [112]. Ten of the eleven bacteria examined here havehomologues of this system (3.A.7), but the numbers of such proteins vary between 2 (for Lbr,Lga and Bli) and 8 (for Blo). It is therefore not clear that these bacteria have complete systems.
Constituents of the Bacterial Competence-related DNA Transformation Transporter (DNA-T)family (3.A.11) are found in all of the bacteria, but the numbers of such homologues varybetween 2 and 6. All but Lme, Blo and Bli have all three of the proteins required for competence[113]. Similarly, Septal DNA Translocators of the S-DNA-T family (3.A.12) are found in oneor two copies in all eleven organisms. These proteins may be required for translocation ofchromosomal or plasmid DNA across the septal membrane [114,115].
The Fimbrilin/Protein Exporter (FPE) family is represented in all nine low G+C Gram-positivebacteria under study, but it is not found in the two high G+C Gram-positive bacteria. Thesesystems consist of two proteins, an integral membrane channel protein, and an energizingATPase. Eight of the low G+C Gram-positive bacteria have both such constituents while Lcahas just the ATPase homologue.
11.4. Na+-translocating organocarboxylate decarboxylasesFour organisms, two low G+C and the two high G+C Gram-positive bacteria, have multiple(2–7) subunits of putative Na+-translocating decarboxylases (3.B.1) [116,117]. Of these, onlyone (Lca) has the decarboxylase subunit (α), the integral membrane transporter subunit (β) andthe small γ-subunit. Consequently, only Lca can be expected to export Na+ in response to thecytoplasmic decarboxylation of substrate carboxylates.
Lorca et al. Page 15
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

12. The Phosphotransferase System (PTS)PTS permeases consist of 3 or 4 domains, the IIA, IIB, IIC (present in all families), and IID(present only in the Mannose (Man) family). These domains can be fused together or detachedas separate polypeptide chains [22,118]. Variations in representation of PTS proteins in theeleven organisms were extensive. The two PTS energy-coupling proteins, Enzyme I and HPr,are present in all organisms except in the high G+C Gram-positive bacterium, Bli, which alsolacks a PTS permease. The other high G+C Gram-positive bacterium, Blo, has both EnzymeI and HPr and seems to have a single PTS permease, possibly specific for β-glucosides. Lbrhas far fewer PTS permeases than the other low G+C Gram-positive bacteria with only twosystems, a lactose-type (Lac) system and a Man system.
The representation of PTS permeases in the remaining low G+C lactic acid bacteria variestremendously. Lca has the most with 63 PTS permease proteins, and all seven known familiesof these permeases are encoded within the Lca genome. The first such family, the Glucose/Glucoside (Glc) family (4.A.1) is represented in all of the low G+C lactic acid bacteria exceptLbr, with up to 12 proteins belonging to this family per organism. Surprisingly, Sth has 8 ofits 15 PTS proteins within this family while Lca, with 4 times as many PTS proteins, has only12 in the Glc family.
Within the Glc family, putative glucoside transporting systems outnumber hexose-transportingsystems 4 to 1, emphasizing the dependency of these bacteria on plant glycosides. The Fructose/Mannitol (Fru) family (4.A.2) is found in seven organisms with 1–7 proteins per organism.The Lactose/Diacetylchitobiose/Lichenan Oligosaccharide (Lac) family (4.A.3) is wellrepresented with approximately four times as many proteins as for the Fru family. This is inagreement with the observation that many of these bacteria are dependent on plant glycosidesfor growth. It contrasts with the preponderance of Fru systems in γ-proteobacteria [22]. TheGlucitol (Gut) family (4.A.4) is represented in only two organisms (Ppe and Lca) while proteinsof the GAT family (4.A.5) are present in four (Lbr, Lca, Lga and Ooe). However, Lbr has onlythe IIC component, implying that the protein does not function by a PTS-type mechanism. Itmay be a secondary carrier as discussed previously for other organisms [119,120]. The L-ascorbate (Asc) family (4.A.7) is represented in four lactic acid bacteria (Lca, Lcr, Lme andOoe). All of the low G+C Gram-positive bacteria studied have at least one mannose/fructose/sorbose (Man)-type system (4.A.6), and at least three of them have 3–6 such systems.
13. Transmembrane Electron Transport SystemsWhen grown in the presence of heme, at least some lactic acid bacteria gain additionalmetabolic energy in the form of a proton motive force. The mechanisms are not known, andthese observations could be a result of indirect effects rather than the direct coupling of electronflow to proton efflux. In fact, none of the currently recognized bacterial H+- or Na+-translocating electron flow systems was identified (families of oxidoreduction-driven cationtransporters; TC section 3.D).
Transmembrane electron flow systems of TC classes 5.A and 5.B do not transport cationsacross the membrane, but electrons do flow from one side of it to the other. These systemstherefore contribute to or subtract from the membrane potential. Recognized transmembraneelectron flow systems are present in only one organism, the large genome high G+C Gram-positive bacterium, Bli. Bli has 4 members of the Disulfide Bond Oxidoreductase (DsbD)family (5.A.1) and three protein members of the Prokaryotic Molybdopterin-containingOxidoreductase (PMO) family (5.A.3). However, of these three proteins, two are α-subunitsand one is a β-subunit. Therefore, only one complete PMO family member may be present.Recognizable pmf-generating or -dissipating electron flow carriers are essentially absent in allother bacteria examined.
Lorca et al. Page 16
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

14. Auxiliary Transport ProteinsAuxiliary transport proteins are relegated to TC class 8. Gram-positive bacteria have“membrane fusion proteins” (MFPs) of family 8.A.1. These proteins were named followingtheir discovery in Gram-negative bacteria [121], but they have since been described in Gram-positive bacteria where they can be essential for the activity of particular ABC-type exporttransporters [122].
Only two of the bacteria studied here have such a protein (Sth and Lme). These probablyfunction in conjunction with peptide efflux pumps of the ABC superfamily [122]. In fact, bothSth and Lme have genes encoding ABC transporters of the Pep2E family (TC #3.A.1.112)adjacent to the genes coding for their MFPs. Two Gram-positive bacterial MFPs have beenshown to be essential for ABC transport function [123–125].
Seven organisms have MPA1 family members (8.A.3) that regulate complex capsularpolysaccharide export via PST-type transporters of the MOP superfamily (TC #2.A.66.2)[83]. The K+ Transport/Nucleotide Binding Regulatory Subunit (KTN) family (8.A.5), whichcontrols the activities of Trk family K+ porters, is found in 9 of the 11 organisms, and all ofthe low G+C lactic acid bacteria have homologues of the eukaryotic rBAT (1–5 copies perorganism). rBAT homologues stimulate amino acid transport via members of the APCsuperfamily, but they are homologous to various glycosidases [126–128]. There is no evidencethat these homologues regulate amino acid transport in bacteria.
15. Poorly Characterized TransportersSeveral poorly characterized transporter families of TC class 9 are sparsely represented in Table5. These include MerTP Hg2+ transporters (9.A.2) (1 organism), the PnuC family (9.A.4) ofnicotinamide mononucleotide (NMN) transporters (3 organisms) and the FeoB (9.A.8) andOFeT (9.A.10) families of iron transporters (4 and 2 organisms, respectively). A putative LeadUptake Porter (PbrT family; 9.A.17) is found in each of two organisms, Sth and Blo. MgtEMg2+ transporters (9.A.19) are found in single copy in Ppe, Lme and Bli.
Several families of unknown function (TC Class 9.B) are worthy of brief mention. The putativemurein precursor exporter (9.B.3), responsible for export of lipid-linked oligosaccharideprecursors of peptidoglycan, is found in all eleven organisms. These transporters may berequired for the assembly of the bacterial cell wall. The MgtC family of putative Mg2+
transporters (9.B.20) is represented in single copy in 7 organisms. The AI-2E family (TC #2.A.86), one member of which exports the form-containing autoinducer AI-2 signalling molecule[129–131] is found in all eleven organisms (1–5 copies per organism). As AI-2 is believed tobe required for interspecies communication, it is possible that this type of signalling isimportant for the lifestyles of LABs and other Gram-positive bacteria.
The Hly III family (9.B.30) of putative hemolysins is present in 10 of the 11 organisms. It willbe interesting to know the function of these homologues. Members of the Putative VectorialGlycosyl Polymerization (VGP) family (9.B.32) are found in 9 of the 11 organisms, and asmany as 6 paralogues may be present in a single organism. It can be presumed that these proteinsfunction in the assembly of exopolysaccharides and capsular polysaccharides and possiblyteichoic acids as well [132].
16. ConclusionsThe fully sequenced genomes of eleven Gram-positive bacteria were searched for homologuesin TCDB in order to analyze the transport capabilities of these bacteria. These eleven bacteriadevote at least 13 to 18% of their genes to transport. Of the total known transport proteins
Lorca et al. Page 17
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

encoded in these genomes, on average, about 48% constitute uptake systems, 32% constituteefflux systems, and 5% of the proteins encode systems that transport substrates bidirectionally(Table 6). Fifteen percent are of unknown polarity and mechanism of action. Most of thebidirectional transporters are channels that either have low specificity for their substrates orare nonspecific. The putative uptake systems in these lactic acid bacteria transport primarilyamino acids, sugars, cations, anions and peptides. The amino acid transporters are the mostabundant while peptide transporters are the least abundant of these uptake systems.
Lactic acid bacteria have many drug efflux systems, but they also have substantial numbers ofpeptide and macromolecular exporters. Comparison of the relative proportions of transportproteins in lactic acid bacteria with those in Escherichia coli, Bacillus subtilis andMycobacterium tuberculosis revealed interesting features of the lactic acid bacteria (Table 7and Fig. 4). Twenty-four percent of the transport proteins in lactic acid bacteria seem to beinvolved in transport of inorganic molecules, and 23% of the transport proteins may transportamino acids and their derivatives. E. coli, B. subtilis and M. tuberculosis devote about 18%,21% and 42% to transport of inorganic molecules and 29%, 24% and 14% to transport of aminoacids and their derivatives. About 17% of the transport proteins in lactic acid bacteria may beinvolved in the uptake of carbon sources while in E. coli, B. subtilis and M. tuberculosis thesesystems constitute 29%, 20% and 6% of the total transport systems, respectively. In all of theserespects the LABs most closely resemble B. subtilis. Transport proteins that may be involvedin efflux of drugs and toxic compounds constitute 10% of the total transport proteins in theLABs unlike B. subtilis (17%) and M. tuberculosis (18%) but similar to E. coli which has 9%of its transport proteins concerned with efflux of these molecules. Eleven percent of thetransport proteins in lactic acid bacteria may be involved in transport of macromolecules unlikeE. coli, B. subtilis and M. tuberculosis which have only 2%, 2% and 4% of their transportproteins devoted to this function, respectively.
Of the 11 bacteria examined, B. linens has the largest genome, and many transporter familiesare represented that are found in none of the others (see Table 5). Families of secondary carriersthat were only found in B. linens are the UMF2, YnfM, EntS and NNP families in the MFS,KDGT, CaCA, NSS, APC(ACT), CPA3, CHR, LysE and RND (Hae1 and SecDF) families.Three ABC-type families, Prot2E, DrugE3 and CT(ABCC), were also found only in B.linens. Moreover, only in this organism were electron carriers of the DsbD, PMO, QCR andCOX families identified. Finally, certain oversized proteins with 19–25 TMS were found onlyin B. linens, and these proteins belong to the FeCT (19 TMSs), UIT10 (19 TMSs) and the CPA3(24 and 25 TMSs) families.
Analysis of the MFS transporters in these bacteria revealed that the greatest proportion of thesesystems transport drugs (Table 8). Forty to eighty-three percent of MFS transporters in thesebacteria probably exhibit specificity for drugs and other amphipathic compounds. MFStransporters that translocate anions are the second most abundant of the MFS carriers followedby those that transport sugars. However, Bli has more MFS transporters that transport anionsthan drugs or other substrates. Surprisingly, Sth has no sugar transporters of the MFS-type atall. These systems are almost ubiquitous among free-living bacteria.
Analysis of ABC transporters in the eleven bacteria revealed that of these systems, theseorganisms generally have more uptake systems than efflux systems (Table 9). However, Lca,Lde and Lga have more ABC efflux systems than ABC uptake systems. Drug efflux pumpsconstitute the largest proportion of the ABC systems, and amino acid uptake systems are thesecond most abundant. However, the opposite is true for Sth and Lme which have more aminoacid uptake systems than drug efflux pumps. Interestingly, in Lde, most ABC systems effluxlipids and hydrophobic compounds while in Lga most ABC systems efflux peptides. In Blo,
Lorca et al. Page 18
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ABC-type sugar uptake systems dominate. A large number of drug exporters is a commonfeature of soil bacteria which presumably must cope with such toxic substances frequently.
Bacterial multidrug resistance efflux pumps fall into five superfamilies, MFS (TC #2.A.1),RND (TC #2.A.6), DMT (TC #2.A.7), MOP (TC #2.A.66 and ABC (TC #3.A.1). Several ofthese superfamilies include multiple families that export drugs and other toxic molecules[80,83,133,134]. We analyzed the distribution of these various types of pumps in the elevenGram-positive bacteria as well as several other well-characterized bacteria.
Analyses of the putative drug efflux pumps (Table 10) in the eleven bacteria reveals that overall,55% of these pumps belong to the MFS followed by ABC (33%), MOP (6%), RND (4%) andDMT (2%). Some organisms provided exceptions to this trend. Sth had more ABC familymembers (54%) than MFS (23%) or MOP (23%), but this was the only organism in whichMOP and MFS drug efflux pumps are present in equal numbers. No RND or DMT familymembers were found in Sth, while no representatives of the MOP or DMT families were foundin Lde and Lga. RND and DMT systems were absent in Ooe and Blo. Although the greatestproportions of drug efflux pumps in E. coli and B. subtilis are MFS-types, the relativeproportions of these and other drug efflux pumps in these bacteria differ. E. coli has 58% MFS,16% RND, 14% ABC, 7% DMT and 5% MOP-type systems while B. subtilis has 61% MFS,14% MOP, about 11% each of ABC and DMT and only 3% (one system) of RND-type systems.
Sugar transporters can be secondary carriers, ABC transporters or PTS group translocators.Half of the sugar uptake systems in the low G+C Gram-positive lactic acid bacteria are PTS-type (50%) while 34% are secondary carriers and only 16% are ABC transporters (Table 11).However, in the high G+C Gram-positive bacteria, Bli and Blo, most of the sugar transportersare either secondary carriers (48%) or ABC-type transporters (50%). Bli does not encode anyPTS permeases while just one sugar transport system in Blo is a PTS permease.
In three representative Gram-negative bacterial pathogens, Borrelia burgdorferi (Bbu),Treponema pallidum (Tpa) and Haemophilus influenzae (Hin), 70% of all sugar transportersare ABC-type while 22% are PTS and only 8% are secondary carriers (Table 11). However,in three representative Gram-positive pathogens, Listeria monocytogenes (Lmo),Streptococcus pyogenes (Spy) and Mycoplasma genitalium (Mge), about 75% of all sugartransporters are PTS permeases and 16% are ABC-type while only 9% are secondary carriers.These considerations demonstrate the unique features of the LABs and reveal which organismaltypes they most resemble.
The low G+C Gram-positive lactic acid bacteria seem to encode slightly more transportersspecific for glycosides (51%) as compared with those for free sugars (49%) (Table 12). Thetwo high G+C Gram-positive bacteria, Bli and Blo, show tremendous bias in the utilization ofglycosides versus sugars, but in opposite directions. Blo has more than twice as many uptakesystems for glycosides as for free sugars while Bli has more than ten times more transportersfor free sugars than for glycosides. In comparison, E. coli and B. subtilis transport more freesugars (65% and 55%, respectively) than glycosides (35% and 45%, respectively).Representative Gram-negative pathogens (Bbu, Tpa and Hin) have five times more uptakesystems for free sugars than for glycosides while Gram-positive bacterial pathogens (Lmo, Spyand Mge) have nearly two fold more sugar uptake proteins than glycoside uptake systems.These observations illustrate the unusual sugar utilization capabilities of the Gram-positivebacteria that live primarily in association with plants.
All eleven bacteria studied here have at least one peptide uptake system (Fig. 5), and all butBli have at least one peptide efflux system. Bli has disproportionately greater numbers ofpeptide uptake systems (13 systems) while Lga has many more peptide efflux systems (11systems) than the other bacteria. Lca, Lde, Lga, Sth and Lme have more peptide efflux systems
Lorca et al. Page 19
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

than peptide uptake systems, while Ppe, Ooe and Blo have more uptake systems for peptidesthan efflux systems. Lbr and Lcr have equal proportions of these two types of systems (Fig.5).
All low G+C lactic acid bacteria possess mechanosensitive channels for osmotic adaptation,suggesting that these organisms are exposed to extremes of osmotic conditions in their naturalhabitat. They also have chloride channels that may function in pH homeostasis. For the mostpart, they lack electron transfer, suggesting a predominantly fermentative mode of energygeneration. They have peptide uptake and export systems that function in nutrition, signalling,regulation and biological warfare. They have complements of transporters that distinguish themfrom other types of bacteria, and the different LABs also exhibit species-specificcharacteristics. All eleven organisms have competence-related and septal DNA translocationsystems although competence has not been demonstrated for any of these bacteria. They allpossess one or more putative murine precursor exporter(s), possibly required for extracellularpeptidoglycan assembly. Finally, they all possess the Sec/Oxa1 systems for secretion,membrane insertion and assembly of proteins that include all of the transporters discussed inthis article.
AcknowledgementsWe wish to acknowledge the enormous contribution of the Lactic Acid Bacterial Genome Sequencing Consortiumwithout the effort of which these analyses would not have been possible. We also thank Mary Beth Hiller for assistancein preparing this manuscript. This work was supported by NIH grant GM 55434.
References1. Gaudu P, Vido K, Cesselin B, Kulakauskas S, Tremblay J, Rezaiki L, Lamberret G, Sourice S, Duwat
P, Gruss A. Respiration capacity and consequences in Lactococcus lactis. Antonie Van Leeuwenhoek2002;82:263–269. [PubMed: 12369192]
2. Whittenbury R. Biochemical characteristics of Streptococcus species. Soc Appl Bacteriol Symp Ser1978;7:51–69. [PubMed: 364662]
3. Wolf G, Strahl A, Meisel J, Hammes WP. Heme-dependent catalase activity of lactobacilli. Int J FoodMicrobiol 1991;12:133–140. [PubMed: 1904757]
4. Sakamoto K, Konings WN. Beer spoilage bacteria and hop resistance. Int J Food Microbiol2003;89:105–124. [PubMed: 14623377]
5. Salminen, S.; Deighton, MA.; Benno, Y.; Gorbach, SL. Lactic acid bacteria in health and disease. In:Salminen, S.; von Wright, A., editors. Lactic Acid Bacteria Microbiology and Functional Aspects,Chapter 7. Marcel Dekker, Inc; New York: 1998. p. 211-253.
6. Busch W, Saier MH Jr. The Transporter Classification (TC) System, 2002. CRC Crit Rev BiochemMol Biol 2002;37:287–337. [PubMed: 12449427]
7. Saier MH Jr. A functional-phylogenetic classification system for transmembrane solute transporters.Microbiol Mol Biol Rev 2000a;64:354–411. [PubMed: 10839820]
8. Klaenhammer T, Altermann E, Arigoni F, Bolotin A, Breidt F, Broadbent J, Cano R, Chaillou S,Deutscher J, Gasson M, van de Guchte M, Guzzo J, Hartke A, Hawkins T, Hols P, Hutkins R,Kleerebezem M, Kok J, Kuipers O, Lubbers M, Maguin E, McKay L, Mills D, Nauta A, Overbeek R,Pel H, Pridmore D, Saier M, van Sinderen D, Sorokin A, Steele J, O’Sullivan D, de Vos W, WeimerB, Zagorec M, Siezen R. Discovering lactic acid bacteria by genomics. Antoine van Leeuwenhoek2002;82:29–58.
9. Makarova K, Slesarev A, Wolf Y, Sorokin A, Mirkin B, Koonin E, Pavlov A, Pavlova N, KaramychevV, Polouchine N, Shakhova V, Grigoriev I, Lou Y, Rohksar D, Lucas S, Huang K, Goodstein DM,Hawkins T, Plengvidhya V, Welker D, Hughes J, Goh Y, Benson A, Baldwin K, Lee JH, Diaz-MunizI, Dosti B, Smeianov V, Wechter W, Barabote R, Lorca G, Altermann E, Barrangou R, Ganesan B,Xie Y, Rawsthorne H, Tamir D, Parker C, Breidt F, Broadbent J, Hutkins R, O’Sullivan D, Steele J,Unlu G, Saier M, Klaenhammer T, Richardson P, Kozyavkin S, Weimer B, Mills D. Comparative
Lorca et al. Page 20
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

genomics of the lactic acid bacteria. Proc Natl Acad Sci USA 2006;103:15611–15616. [PubMed:17030793]
10. Khwaja M, Ma Q, Saier MH Jr. Topological analysis of integral membrane constituents of prokaryoticABC efflux systems. Res Microbiol 2005;156:270–277. [PubMed: 15748994]
11. Saier MH Jr, Tran CV, Barabote RD. TCDB: The transporter classification database for membranetransport protein analyses and information. Nucleic Acids Res 2006;34:D181–D186. [PubMed:16381841]Database issue
12. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J MolBiol 1990;215:403–410. [PubMed: 2231712]
13. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLASTand PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Re1997;25:3389–3402.
14. Tusnady GE, Simon I. Principles governing amino acid composition of integral membrane proteins:application to topology prediction. J Mol Biol 1998;283:489–506. [PubMed: 9769220]
15. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL X windowsinterface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. NucleicAcids Res 1997;25:4876–4882. [PubMed: 9396791]
16. Zhai Y, Tchieu J, Saier MH Jr. A web-based Tree View (TV) program for the visualization ofphylogenetic trees. J Mol Microbiol Biotechnol 2002;4:69–70. [PubMed: 11763971]
17. Amico M, Finelli M, Rossi I, Zauli A, Elofsson A, Viklund H, von Heijne G, Jones D, Krogh A,Fariselli P, Luigi Martelli P, Casadio R. PONGO: a web server for multiple predictions of all-alphatransmembrane proteins. Nucleic Acids Res 2006;34:W169–172. [PubMed: 16844984]Web Serverissue
18. von Heijne G. Membrane-protein topology. Nat Rev Mol Cell Biol 2006;7:909–918. [PubMed:17139331]
19. Kim H, Melen K, Osterberg M, von Heijne G. A global topology map of the Saccharomycescerevisiae membrane proteome. Proc Natl Acad Sci USA 2006;103:11142–11147. [PubMed:16847258]
20. Daley DO, Rapp M, Granseth E, Melen K, Drew D, von Heijne G. Global topology analysis of theEscherichia coli inner membrane proteome. Science 2005;308:1321–1323. [PubMed: 15919996]
21. Granseth E, Daley DO, Rapp M, Melen K, von Heijne G. Experimentally constrained topology modelsfor 51,208 bacterial inner membrane proteins. J Mol Biol 2005;352:489–494. [PubMed: 16120447]
22. Nguyen TX, Yen MR, Saier MH Jr. Topology predictions for integral membrane permeases of thephosphoenolpyruvate:sugar phosphotransferase system. J Mol Microbiol Biotechnol 2006;11:345–360. [PubMed: 17114898]
23. Barabote RD, Saier MH Jr. Comparative genomic analyses of the bacterial phosphotransferase system.Microbiol Mol Biol Rev 2005;69:608–634. [PubMed: 16339738]
24. Paulsen IT, Sliwinski MK, Saier MH Jr. Microbial genome analyses: global comparisons of transportcapabilities based on phylogenies, bioenergetics and substrate specificities. J Mol Biol1998;277:573–592. [PubMed: 9533881]
25. Paulsen IT, Nguyen L, Sliwinski MK, Rabus R, Saier MH Jr. Microbial genome analyses: comparativetransport capabilities in eighteen prokaryotes. J Mol Biol 2000;301:75–100. [PubMed: 10926494]
26. Saier, MH., Jr; Paulsen, IT. Whole genome analyses of transporters in spirochetes: Borreliaburgdorferi and Treponema pallidum (Ch. 8). In: Saier, MH.; Garcia-Lara, J., editors. TheSpirochetes: Molecular and Cellular Biology. Wymondham, Norfolk, UK: Horizon Scientific Press;2001. p. 67-73.
27. Saier MH Jr, Goldman SR, Maile RR, Moreno MS, Weyler W, Yang N, Paulsen IT. Transportcapabilities encoded within the Bacillus subtilis genome. J Mol Microbiol Biotechnol 2002;4:37–67.[PubMed: 11763970]
28. Saier MH Jr. Tracing pathways of transport protein evolution. Mol Microbiol 2003;48:1145–1156.[PubMed: 12787345]
29. von T, Nies DH, Saier MH Jr. Genomic analyses of transport proteins in Ralstonia metallidurans,Comparative Funct. Genomics 2005;6:17–56. [PubMed: 15715905]
Lorca et al. Page 21
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

30. Winnen, B.; Felce, J.; Saier, MH, Jr. Genomic analyses of transporter proteins in Corynebacteriumglutamicum and Corynebacterium efficiens. In: Eggeling, L.; Bott, M., editors. Handbook ofCorynebacterium glutamicum, Ch. 8. Boca Raton, FL: CRC Press; 2005. p. 149-186.
31. Folgering JHA, Moe PC, Schuurman-Wolters GK, Blount P, Poolman B. Lactococcus lactis usesMscL as its principal mechanosensitive channel. J Biol Chem 2005;280:8784–8792. [PubMed:15613476]
32. Harley KT, Saier MH Jr. A novel ubiquitous family of putative efflux transporters. J Mol MicrobiolBiotechnol 2000;2:195–198. [PubMed: 10939244]
33. Saier MH Jr, Kollman J. Is FatP a long chain fatty acid transporter? Mol Microbiol 1999;33:670–672. [PubMed: 10417658]
34. Bass RB, Strop P, Barclay M, Rees DC. Crystal structure of Escherichia coli MscS, a voltage-modulated and mechanosensitive channel. Science 2002;298:1582–1587. [PubMed: 12446901]
35. Biggin PC, Sansom MS. Mechanosensitive channels: stress relief. Curr Biol 2003;13:R183–R185.[PubMed: 12620208]
36. Koprowski P, Kubalski A. Bacterial ion channels and their eukaryotic homologues. BioEssays2001;23:1148–1158. [PubMed: 11746234]
37. Kuo MMC, Saimi Y, Kung C. Gain-of-function mutations indicate that Escherichia coli Kch formsa functional K+ conduit in vivo. EMBO J 2003;22:4049–4058. [PubMed: 12912904]
38. Perozo E, Cortes DM, Sompornpisut P, Kloda A, Martinac B. Open channel structure of MscL andthe gating mechanism of mechanosensitive channels. Nature 2002;418:942–948. [PubMed:12198539]
39. Pivetti CD, Yen MR, Miller S, Busch W, Tseng YH, Booth IR, Saier MH Jr. Two families ofprokaryotic mechanosensitive channel proteins. Microbiol Mol Biol Rev 2003;67:66–85. [PubMed:12626684]
40. Ruta V, Jiang Y, Lee A, Chen J, MacKinnon R. Functional analysis of an archaebacterial voltage-dependent K+ channel. Nature 2003;422:180–185. [PubMed: 12629550]
41. Stokes NR, Murray HD, Subramaniam C, Gourse RL, Louis P, Bartlett W, Miller S, Booth IR. A rolefor mechanosensitive channels in survival of stationary phase: regulation of channel expression byRpoS. Proc Natl Acad Sci USA 2003;100:15959–15964. [PubMed: 14671322]
42. Iyer R, Iverson TM, Accardi A, Miller C. A biological role for prokaryotic ClC chloride channels.Nature 2002;419:715–718. [PubMed: 12384697]
43. Eshaghi S, Niegowski D, Kohl A, Martinez Molina D, Lesley SA, Nordlund P. Crystal structure ofa divalent metal ion transporter CorA at 2.9 Å resolution. Science 2006;313:354–357. [PubMed:16857941]Erratum in: Science (2006) 313(5792):1389
44. Lunin VV, Dobrovetsky E, Khutoreskaya G, Zhang R, Joachimiak A, Doyle DA, Bochkarev A,Maguire ME, Edwards AM, Koth CM. Crystal structure of the CorA Mg2+ transporter. Nature2006;440:833–837. [PubMed: 16598263]
45. Kehres DG, Maguire ME. Structure, properties and regulation of magnesium transport proteins.BioMetals 2002;15:261–270. [PubMed: 12206392]
46. Carbrey JM, Gorelick-Feldman DA, Kozono D, Praetorius J, Nielsen S, Agre P. AquaglyceroporinAQP9: solute permeation and metabolic control of expression in liver. Proc Natl Acad Sci USA2003;100:2945–2950. [PubMed: 12594337]
47. Jensen MO, Park S, Tajkhorshid E, Schulten K. Energetics of glycerol conduction throughaquaglyceroporin GlpF. Proc Natl Acad Sci USA 2002;99:6731–6736. [PubMed: 11997475]
48. Uehlein N, Lovisolo C, Siefritz F, Kaldenhoff R. The tobacco aquaporins NtAQP1 is a membraneCO2 pore with physiological functions. Nature 2003;425:734–737. [PubMed: 14520414]
49. Hernández-Castro R, Rodríguez MC, Seoane A, Lobo JMG. The aquaporins gene aqpX of Brucellaabortus is induced in hyperosmotic conditions. Microbiology 2003;149:3185–3192. [PubMed:14600230]
50. Kozono D, Ding X, Iwasaki I, Meng X, Kamagata Y, Agre P, Kitagawa Y. Functional expressionand characterization of an archaeal aquaporins. AqpM from Methanothermobacter marburgensis. JBiol Chem 2003;278:10649–10656. [PubMed: 12519768]
Lorca et al. Page 22
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

51. Alvarez, Mde F.; Medina, R.; Pasteris, SE.; Strasser de Saad, AM.; Sesma, F. Glycerol metabolismof Lactobacillus rhamnosus ATCC 7469: cloning and expression of two glycerol kinase genes. J MolMicrobiol Biotechnol 2004;7:170–181. [PubMed: 15383715]
52. Luthi-Peng Q, Dileme FB, Puhan Z. Effect of glucose on glycerol bioconversion by Lactobacillusreuteri. Appl Microbiol Biotechnol 2002;59:289–296. [PubMed: 12111160]
53. Weeks DL, Eskandari S, Scott DR, Sachs GA. H+-gated urea channel: the link between Helicobacterpylori urease and gastric colonization. Science 2000;287:482–485. [PubMed: 10642549]
54. Weeks DL, Sachs G. Sites of pH regulation of the urea channel of Helicobacter pylori. Mol Microbiol2001;40:1249–1259. [PubMed: 11442825]
55. Arispe N, DeMaio A. ATP and ADP modulate a cation channel formed by Hsc70 in acidicphospholipid membranes. J Biol Chem 2000;275:30839–30843. [PubMed: 10899168]
56. Gross C, Koelch W, DeMaio A, Arispe N, Multhoff G. Cell surface-bound heat shock protein 70(Hsp70) mediates perforin-independent apoptosis by specific binding and uptake of granzyme B. JBiol Chem 2003;278:41173–41181. [PubMed: 12874291]
57. Agashe VR, Hartl FU. Roles of molecular chaperones in cytoplasmic protein folding. Cell Dev Biol2000;11:15–25.
58. Dougan DA, Mogk A, Bukau B. Protein folding and degradation in bacteria: to degrade or not todegrade? That is the question. Cell Mol Life Sci 2002;59:1607–1616. [PubMed: 12475170]
59. Houry WA. Chaperone-assisted protein folding in the cell cytoplasm. Curr Prot Pept Sci 2001;2:227–244.
60. Rassow J, von Ahsen O, Bömer U, Pfanner N. Molecular chaperones: towards a characterization ofthe heat-shock protein 70 family. Trends Cell Biol 1997;7:129–133. [PubMed: 17708923]
61. Barth H, Pfeifer G, Hofmann F, Maier E, Benz R, Aktories K. Low pH-induced formation of ionchannels by Clostridium difficile toxin B in target cells. J Biol Chem 2001;276:10670–10676.[PubMed: 11152463]
62. Belland RJ, Scidmore MA, Crane DD, Hogan DM, Whitmire W, McClarty G, Caldwell HD.Chlamydia trachomatis cytotoxicity associated with complete and partial cytotoxin genes. Proc NatlAcad Sci USA 2001;98:13984–13989. [PubMed: 11707582]
63. Moll GN, Konings WN, Driessen AJM. Bacteriocins: mechanism of membrane insertion and poreformation. Antonie van Leeuwenhoek 1999;76:185–198. [PubMed: 10532378]
64. Venema K, Venema G, Kok J. Lactococcal bacteriocins: mode of action and immunity. TrendsMicrobiol 1995;3:299–304. [PubMed: 8528613]
65. Young R. Bacteriophage holins: deadly diversity. J Mol Microbiol Biotechnol 2002;4:21–36.[PubMed: 11763969]
66. Engelberg-Kulka H, Glaser G. Addiction modules and programmed cell death and antideath inbacterial cultures. Annu Rev Microbiol 1999;53:43–70. [PubMed: 10547685]
67. Lewis K. Programmed death in bacteria. Microbiol Mol Biol Rev 2000;64:503–514. [PubMed:10974124]
68. Rice KC, Bayles KW. Death’s toolbox: examining the molecular components of bacterialprogrammed cell death. Mol Microbiol 2003;50:729–738. [PubMed: 14617136]
69. Pao SS, Paulsen IT, Saier MH Jr. The major facilitator superfamily. Microbiol Mol Biol Rev1998;62:1–32. [PubMed: 9529885]
70. Saier MH Jr, Beatty JT, Goffeau A, Harley KT, Heijne WHM, Huang SC, Jack DL, Jahn PS, LewK, Liu J, Pao SS, Paulsen IT, Tseng TT, Virk PS. The major facilitator superfamily. J Mol MicrobiolBiotechnol 1999;1:257–279. [PubMed: 10943556]
71. Chaillou S, Postma PW, Pouwels PH. Functional expression in Lactobacillus plantarum of xylPencoding the isoprimeverose transporter of Lactobacillus pentosus. J Bacteriol 1998;180:4011–4014.[PubMed: 9683504]
72. Heuberger EH, Smits E, Poolman B. Xyloside transport by XylP, a member of the galactoside-pentoside-hexuronide family. J Biol Chem 2001;276:34465–34472. [PubMed: 11408491]
73. Grossiord BP, Luesink EJ, Vaughan EE, Arnaud A, de Vos WM. Characterization, expression, andmutation of the Lactococcus lactis galPMKTE genes, involved in galactose utilization via the Leloirpathway. J Bacteriol 2003;185:870–878. [PubMed: 12533462]
Lorca et al. Page 23
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

74. Hugouvieux-Cotte-Pattat N, Reverchon S. Two transporters, TogT and TogMNAB, are responsiblefor oligogalacturonide uptake in Erwinia chrysanthemi 3937. Mol Microbiol 2001;41:1125–1132.[PubMed: 11555292]
75. Poolman B, Knol J, van der Does C, Henderson PJF, Liang WJ, Leblanc G, Pourcher T, Mus-VeteauI. Cation and sugar selectivity determinants in a novel family of transport proteins. Mol Microbiol1996;19:911–922. [PubMed: 8830272]
76. Holemann A, Seeberger PH. Carbohydrate diversity: synthesis of glycoconjugates and complexcarbohydrates. Curr Opin Biotechnol 2004;15:615–622. [PubMed: 15560990]
77. O’Donnell N. Intracellular glycosylation and development. Biochim Biophys Acta 2002;1573:336–345. [PubMed: 12417416]
78. Jack DL, Paulsen IT, Saier MH Jr. The amino acid/polyamine/organocation (APC) superfamily oftransporters specific for amino acids, polyamines and organocations. Microbiology 2000;146:1797–1814. [PubMed: 10931886]
79. Tseng T-T, Gratwick KS, Kollman J, Park D, Nies DH, Goffeau A, Saier MH Jr. The RND permeasesuperfamily: an ancient, ubiquitous and diverse family that includes human disease and developmentproteins. J Mol Microbiol Biotechnol 1999;1:107–125. [PubMed: 10941792]
80. Jack DL, Yang NM, Saier MH Jr. The drug/metabolite transporter superfamily. Eur J Biochem2001;268:3620–3639. [PubMed: 11432728]
81. Fiegler H, Bassias J, Jankovic I, Brückner R. Identification of a gene in Staphylococcus xylosusencoding a novel glucose uptake protein. J Bacteriol 1999;181:4929–4936. [PubMed: 10438764]
82. Jung H. The sodium/substrate symporter family: structural and functional features. FEBS Lett2002;529:73–77. [PubMed: 12354616]
83. Hvorup RN, Winnen B, Chang AB, Jiang Y, Zhou XF, Saier MH Jr. The multidrug/oligosaccharidyl-lipid/polysaccharide (MOP) exporter superfamily. Eur J Biochem 2003b;270:799–813. [PubMed:12603313]
84. Epstein W. The roles and regulation of potassium in bacteria. Prog Nucleic Acid Res Mol Biol2003;75:293–320. [PubMed: 14604015]
85. Hase CC, Fedorova ND, Galperin MY, Dibrov PA. Sodium ion cycle in bacterial pathogens: evidencefrom cross-genome comparisons. Microbiol Mol Biol Rev 2001;65:353–370. [PubMed: 11528000]
86. Maguire ME. The structure of CorA: a Mg2+-selective channel. Curr Opin Struct Biol 2006a;16:432–438. [PubMed: 16828282]
87. Maguire ME. Magnesium transporters: properties, regulation and structure. Front Biosci 2006b;11:3149–3163. [PubMed: 16720382]
88. Khademi S, O’Connell J III, Remis J, Robles-Colmenares Y, Miercke LJW, Stroud RM. Mechanismof ammonia transport by Amt/MEP/Rh: Structure of AmtB at 1.35 Å. Science 2004;305:1587–1574.[PubMed: 15361618]
89. Zheng L, Kostrewa D, Berneche S, Winkler FK, Li XD. The mechanism of ammonia transport basedon the crystal structure of AmtB of Escherichia coli. Proc Natl Acad Sci USA 2004;101:17090–17095. [PubMed: 15563598]
90. Hoenke S, Schmid M, Dimroth P. Sequence of a gene cluster from Klebsiella pneumoniae encodingmalonate decarboxylase and expression of the enzyme in Escherichia coli. Eur J Biochem1997;246:530–538. [PubMed: 9208947]
91. Labarre C, Divies C, Guzzo J. Genetic organization of the mle locus and identification of a mleR-likegene from Leuconostoc oenos. Appl Env Microbiol 1996;62:4493–4498. [PubMed: 8953720]
92. Prakash S, Cooper G, Singhi S, Saier MH Jr. The ion transporter superfamily. Biochim Biophys Acta2003;1618:79–92. [PubMed: 14643936]
93. Saier MH Jr. Families of transmembrane transporters selective for amino acids and their derivatives.Microbiology 2000b;146:1775–1795. [PubMed: 10931885]
94. Vrljic M, Garg J, Bellmann A, Wachi S, Freudl R, Malecki MJ, Sahm H, Kozina VJ, Eggeling L,Saier MH Jr. The LysE superfamily: topology of the lysine exporter LysE of Corynebacteriumglutamicum, a paradyme for a novel superfamily of transmembrane solute translocators. J MolMicrobiol Biotechnol 1999;1:327–336. [PubMed: 10943564]
Lorca et al. Page 24
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

95. Kennerknecht N, Sahm H, Yen MR, Patek M, Saier MH Jr, Eggeling L. Export of L-isoleucine fromCorynebacterium glutamicum: a two-gene-encoded member of a new translocator family. J Bacteriol2002;184:3947–3956. [PubMed: 12081967]
96. Trötschel C, Deutenberg D, Bathe B, Burkovski A, Kramer R. Characterization of methionine exportin Corynebacterium glutamicum. J Bacteriol 2005;187:3786–3794. [PubMed: 15901702]
97. Saurin W, Hofnung M, Dassa E. Getting in or out: Early segregation between importers and exportersin the evolution of ATP-binding cassette (ABC) transporters. J Mol Evol 1999;48:22–41. [PubMed:9873074]
98. Tomii K, Kanehisa M. A comparative analysis of ABC transporters in complete microbial genomes.Genome Res 1998;8:1048–1059. [PubMed: 9799792]
99. Tam R, Saier MH Jr. Structural, functional, and evolutionary relationships among extracellular solute-binding receptors of bacteria. Microbiol Rev 1993;57:320–346. [PubMed: 8336670]
100. Kang CH, Kim SH, Nikaido K, Gokcen S, Ames GF. Crystallization and preliminary X-ray studiesof HisJ and LAO periplasmic proteins from Salmonella typhimurium. J Mol Biol 1989;207:643–644. [PubMed: 2668538]
101. Higgins CF, Linton KJ. The ATP switch model for ABC transporters. Nat Struct Mol Biol2004;11:918–926. [PubMed: 15452563]
102. Higgins CF. ABC transporters: from microorganisms to man. Annu Rev Cell Biol 1992;8:67–113.[PubMed: 1282354]
103. Delcour J, Ferain T, Deghorain M, Palumbo E, Hols P. The biosynthesis and functionality of thecell-wall of lactic acid bacteria. Antoine Van Leeuwenhoek 1999;76:159–184.
104. Martin DW. Structure-function relationships in the Na+,K+-pump. Semin Nephrol 2005;25:282–291. [PubMed: 16139683]
105. Hao Z, Chen S, Wilson DB. Cloning, expression, and characterization of cadmium and manganeseuptake genes from Lactobacillus plantarum. Appl Environ Microbiol 1999;65:4746–4752.[PubMed: 10543781]
106. Solioz M, Stoyanov JV. Copper omeostasis in Enterococcus hirae. FEMS Microbiol Rev2003;27:183–195. [PubMed: 12829267]
107. Cao TB, Saier MH Jr. The general protein secretory pathway: Phylogenetic analyses leading toevolutionary conclusions. Biochim Biophys Acta 2003;1609:115–125. [PubMed: 12507766]
108. Yen MR, Harley KT, Tseng YH, Saier MH Jr. Phylogenetic and structural analyses of the Oxa1family of putative protein translocation constituents. FEMS Microbiol Lett 2001;204:223–231.[PubMed: 11731127]
109. Bensing BA, Sullam PM. An accessory sec locus of Streptococcus gordonii is required for exportof the surface protein GspB and for normal levels of binding to human platelets. Mol Microbiol2002;44:1081–1094. [PubMed: 12010500]
110. Bensing BA, Gibson BW, Sullam PM. The Streptococcus gordonii platelet binding protein GspBundergoes glycosylation independently of export. J Bacteriol 2004;186:638–645. [PubMed:14729688]
111. Takamatsu D, Bensing BA, Sullam PM. Genes in the accessory sec locus of Streptococcusgordonii have three functionally distinct effects on the expression of the platelet-binding proteinGspB. Mol Microbiol 2004;52:189–203. [PubMed: 15049820]
112. Grohmann E, Muth G, Espinosa M. Conjugative plasmid transfer in Gram-positive bacteria.Microbiol Mol Biol Rev 2003;67:277–301. [PubMed: 12794193]
113. Dubnau D. DNA uptake in bacteria. Annu Rev Microbiol 1999;53:217–244. [PubMed: 10547691]114. Pogliano J, Sharp MD, Pogliano K. Partitioning of chromosomal DNA during establishment of
cellular asymmetry in Bacillus subtilis. J Bacteriol 2002;184:1743–1749. [PubMed: 11872726]115. Sharpe ME, Errington J. Upheaval in the bacterial nucleoid: an active chromosome segregation
mechanism. Trends Genet 1999;15:70–74. [PubMed: 10098410]116. Buckel W. Sodium ion-translocating decarboxylases. Biochim Biophys Acta 2001;1505:15–27.
[PubMed: 11248185]117. Dimroth P. Primary sodium ion translocating enzymes. Biochim Biophys Acta 1997;1318:11–51.
[PubMed: 9030254]
Lorca et al. Page 25
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

118. Reizer J, Saier MH Jr. Modular multidomain phosphoryl transfer proteins of bacteria. Curr OpinStruc Biol 1997;7:407–415.
119. Hvorup RN, Chang AB, Saier MH Jr. Bioinformatic analyses of homologues of the bacterial L-ascorbate PTS permeases. J Mol Microbiol Biotechnol 2003a;6:191–205. [PubMed: 15153772]
120. Saier MH Jr, Hvorup RN, Barabote RD. Evolution of the bacterial phosphotransferase system: fromcarriers and enzymes to group translocators. Biochem Soc Trans 2005;33:220–224. [PubMed:15667312]
121. Dinh T, Paulsen IT, Saier MH Jr. A family of extracytoplasmic proteins that allow transport of largemolecules across the outer membranes of Gram-negative bacteria. J Bacteriol 1994;176:3825–3831.[PubMed: 8021163]
122. Harley KT, Djordjevic GM, Tseng TT, Saier MH Jr. Membrane-fusion protein homologues in Gram-positive bacteria. Mol Microbiol 2000;36:516–517. [PubMed: 10792737]
123. Axelsson L, Holck A. The genes involved in production of and immunity to sakacin A, a bacteriocinfrom Lactobacillus sake Lb706. J Bacteriol 1995;177:2125–2137. [PubMed: 7721704]
124. Quadri LE, Kleerebezem M, Kuipers OP, de Vos WM, Roy KL, Vederas JC, Stiles ME.Characterization of a locus from Carnobacterium piscicola LV17B involved in bacteriocinproduction and immunity: evidence for global inducer-mediated transcriptional regulation. JBacteriol 1997;179:6163–6171. [PubMed: 9324267]
125. Venema K, Dost MH, Beun PA, Haandrikman AJ, Venema G, Kok J. The genes for secretion andmaturation of lactococcins are located on the chromosome of Lactococcus lactis IL1403. ApplEnviron Microbiol 1996;62:1689–1692. [PubMed: 8633867]
126. Devés R, Boyd CAR. Surface Antigen CD98(4F2): not a single membrane protein, but a family ofproteins with multiple functions. J Membr Biol 2000;173:165–177. [PubMed: 10667913]
127. Malandro MS, Kilberg MS. Molecular biology of mammalian amino acid transporters. Annu RevBiochem 1996;65:305–336. [PubMed: 8811182]
128. Palacín M, Estévez R, Bertran J, Zorzano A. Molecular biology of mammalian plasma membraneamino acid transporters. Physiol Rev 1998;78:969–1054. [PubMed: 9790568]
129. Camilli A, Bassler BL. Bacterial small-molecule signaling pathways. Science 2006;311:1113–1116.[PubMed: 16497924]
130. Chen X, Schauder S, Potier N, Van Dorsselaer A, Pelczer I, Bassler BL, Hughson FM. Structuralidentification of a bacterial quorum-sensing signal containing boron. Nature 2002;415:488–489.[PubMed: 11823842]
131. Herzberg M, Kaye IK, Peti W, Wood TK. YdgG (TqsA) controls biofilm formation in Escherichiacoli K-12 through autoinducer 2 transport. J Bacteriol 2006;188:587–598. [PubMed: 16385049]
132. Morath S, von Aulock S, Hartung T. Structure/function relationships of lipoteichoic acids. JEndotoxin Res 2005;11:348–356. [PubMed: 16303090]
133. Paulsen IT, Chen J, Nelson KE, Saier MH Jr. Comparative genomics of microbial drug effluxsystems. J Mol Microbiol Biotechnol 2001;3:145–150. [PubMed: 11321566]
134. Saier MH Jr, Paulsen IT. Phylogeny of multidrug transporters. Semin Cell Dev Biol 2001;12:205–213. [PubMed: 11428913]
135. Kandler O. Carbohydrate metabolism in lactic acid bacteria. Antonie van Leeuwenhoek1983;49:209–224. [PubMed: 6354079]
136. Kandler O. Current taxonomy of lactobacilli. Dev Ind Microbiol 1984;25:109–123.137. Kandler, O.; Weiss, N. Regular, non-sporing Gram-positive rods. In: Sneath, PHA.; Mair, NS.;
Sharpe, ME.; Holt, JG., editors. Bergey’s Manual of Systematic Bacteriology. 2. Williams andWilkins; Baltimore: 1986. p. 1208-1234.
Lorca et al. Page 26
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
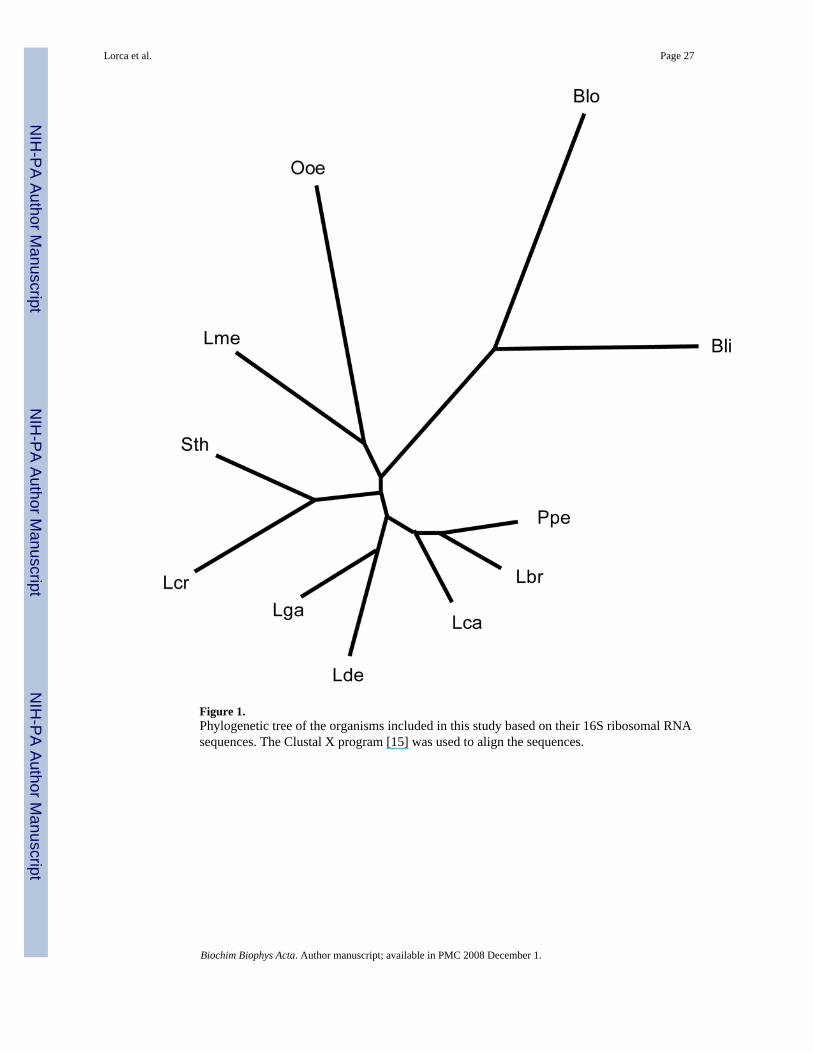
Figure 1.Phylogenetic tree of the organisms included in this study based on their 16S ribosomal RNAsequences. The Clustal X program [15] was used to align the sequences.
Lorca et al. Page 27
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
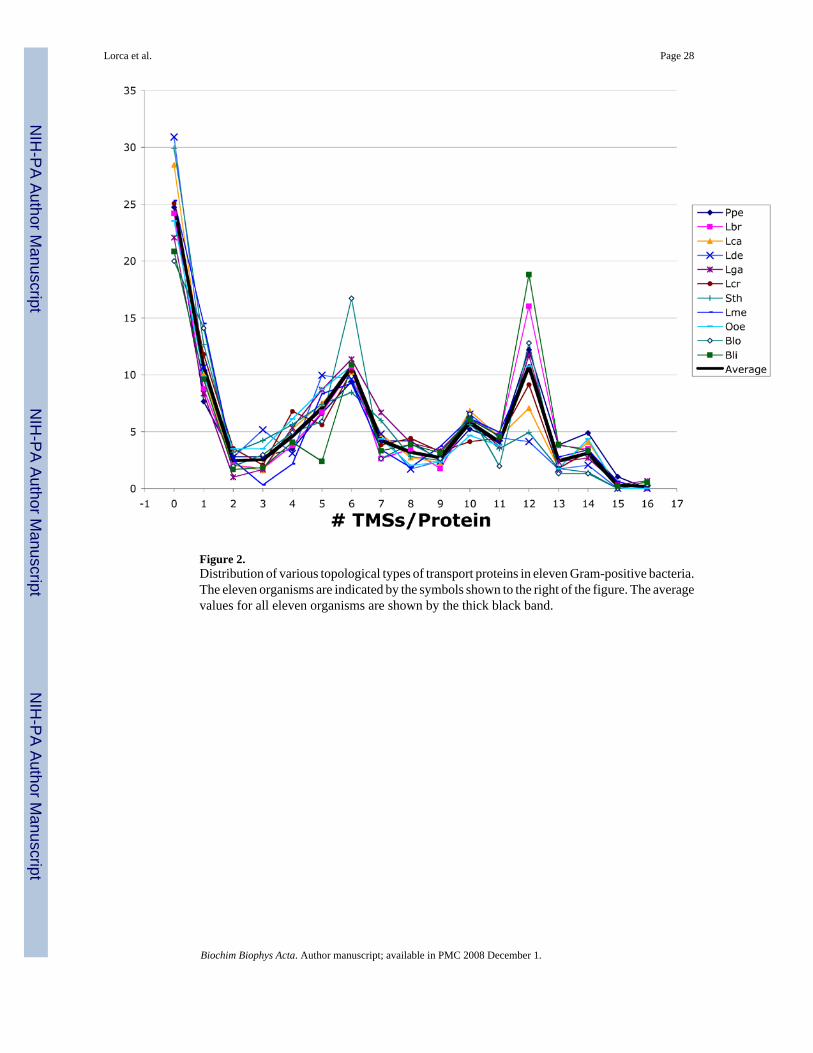
Figure 2.Distribution of various topological types of transport proteins in eleven Gram-positive bacteria.The eleven organisms are indicated by the symbols shown to the right of the figure. The averagevalues for all eleven organisms are shown by the thick black band.
Lorca et al. Page 28
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
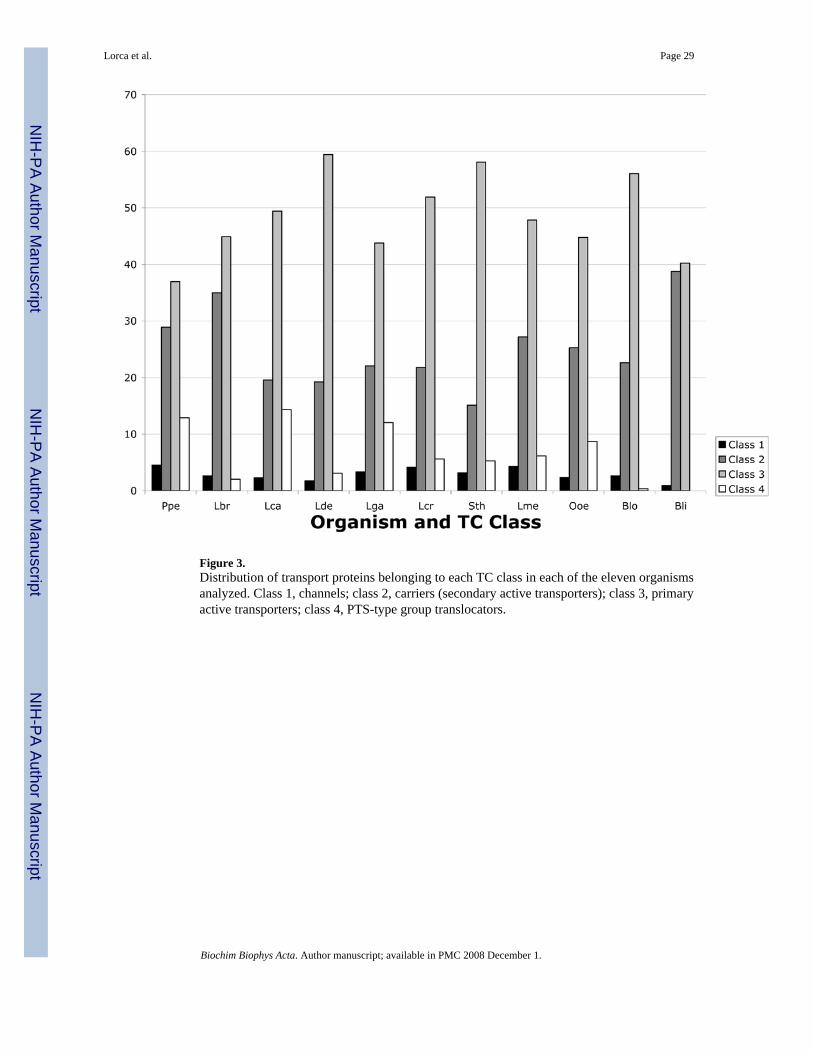
Figure 3.Distribution of transport proteins belonging to each TC class in each of the eleven organismsanalyzed. Class 1, channels; class 2, carriers (secondary active transporters); class 3, primaryactive transporters; class 4, PTS-type group translocators.
Lorca et al. Page 29
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
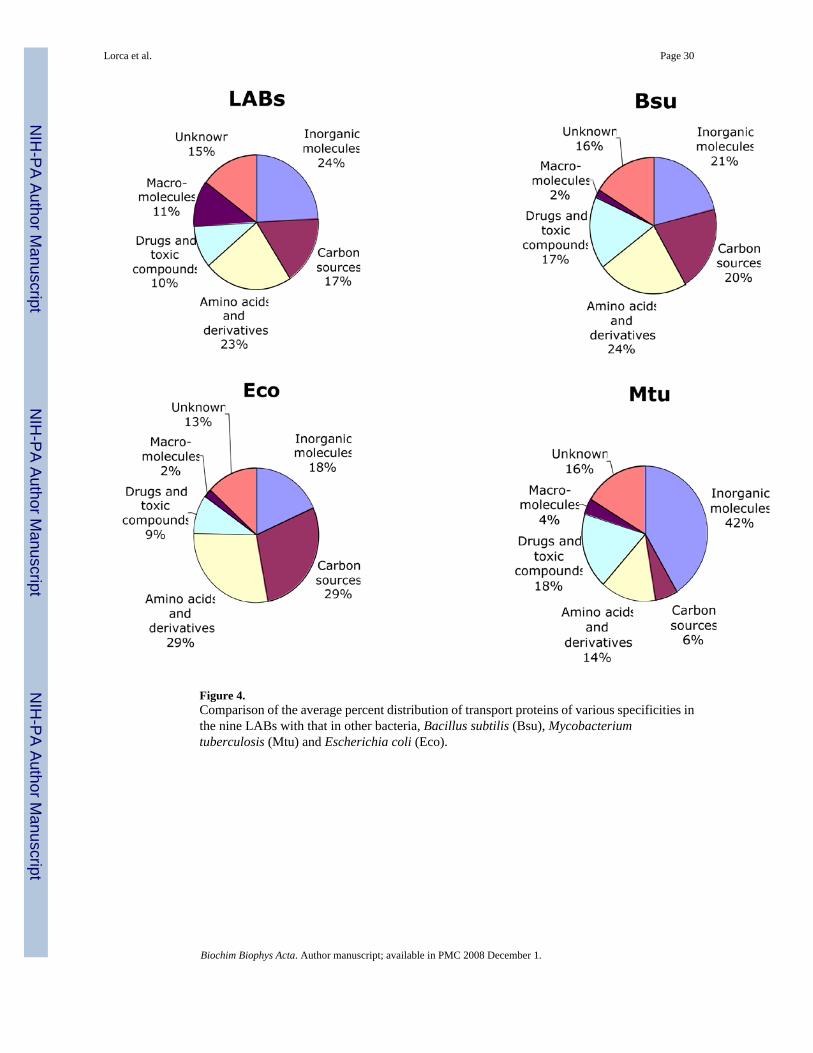
Figure 4.Comparison of the average percent distribution of transport proteins of various specificities inthe nine LABs with that in other bacteria, Bacillus subtilis (Bsu), Mycobacteriumtuberculosis (Mtu) and Escherichia coli (Eco).
Lorca et al. Page 30
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
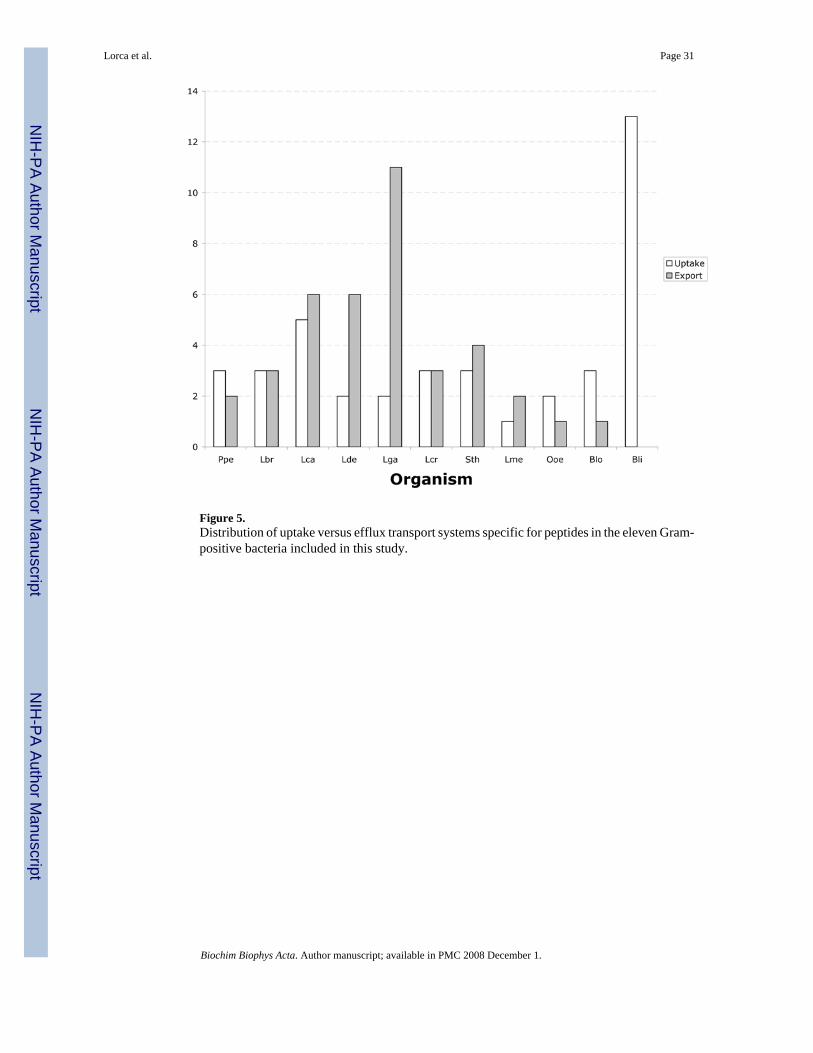
Figure 5.Distribution of uptake versus efflux transport systems specific for peptides in the eleven Gram-positive bacteria included in this study.
Lorca et al. Page 31
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
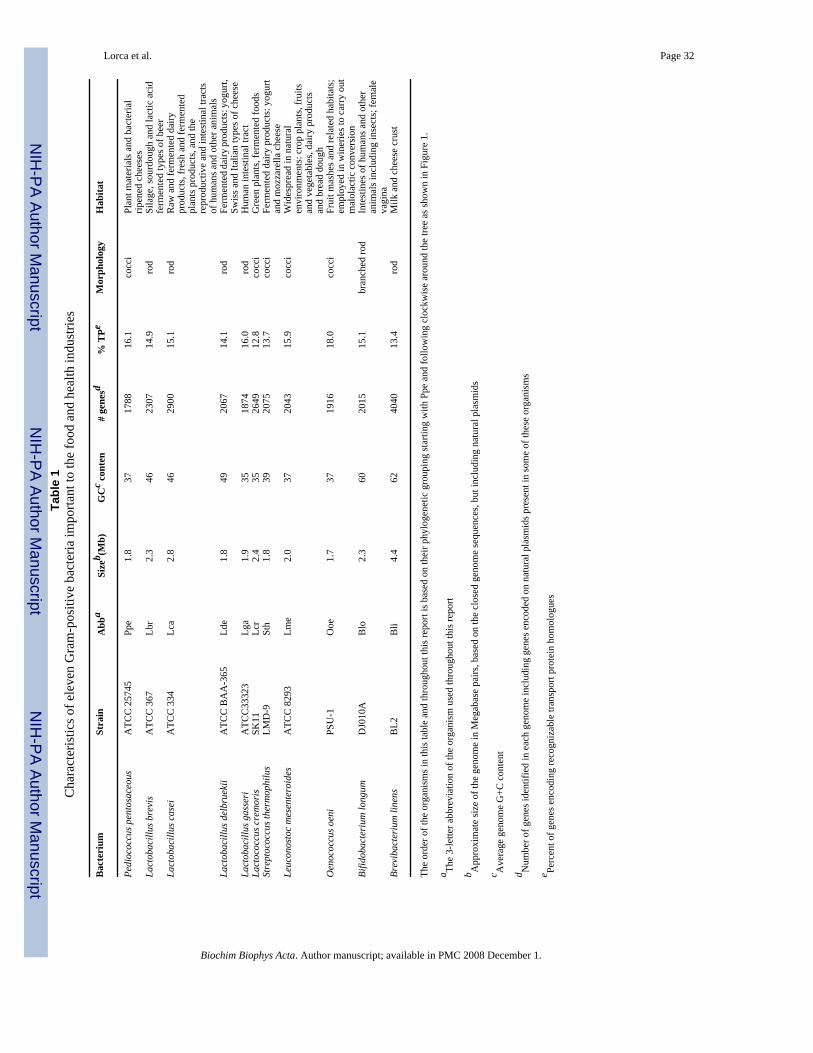
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 32Ta
ble
1C
hara
cter
istic
s of e
leve
n G
ram
-pos
itive
bac
teria
impo
rtant
to th
e fo
od a
nd h
ealth
indu
strie
s
Bac
teri
umSt
rain
Abb
aSi
zeb (M
b)G
Cc c
onte
n#
gene
sd%
TPe
Mor
phol
ogy
Hab
itat
Pedi
ococ
cus p
ento
sace
ous
ATC
C 2
5745
Ppe
1.8
3717
8816
.1co
cci
Plan
t mat
eria
ls a
nd b
acte
rial
ripen
ed c
hees
esLa
ctob
acill
us b
revi
sA
TCC
367
Lbr
2.3
4623
0714
.9ro
dSi
lage
, sou
rdou
gh a
nd la
ctic
aci
dfe
rmen
ted
type
s of b
eer
Lact
obac
illus
cas
eiA
TCC
334
Lca
2.8
4629
0015
.1ro
dR
aw a
nd fe
rmen
ted
dairy
prod
ucts
, fre
sh a
nd fe
rmen
ted
plan
ts p
rodu
cts,
and
the
repr
oduc
tive
and
inte
stin
al tr
acts
of h
uman
s and
oth
er a
nim
als
Lact
obac
illus
del
brue
kii
ATC
C B
AA
-365
Lde
1.8
4920
6714
.1ro
dFe
rmen
ted
dairy
pro
duct
s: y
ogur
t,Sw
iss a
nd It
alia
n ty
pes o
f che
ese
Lact
obac
illus
gas
seri
ATC
C33
323
Lga
1.9
3518
7416
.0ro
dH
uman
inte
stin
al tr
act
Lact
ococ
cus c
rem
oris
SK11
Lcr
2.4
3526
4912
.8co
cci
Gre
en p
lant
s, fe
rmen
ted
food
sSt
rept
ococ
cus t
herm
ophi
lus
LMD
-9St
h1.
839
2075
13.7
cocc
iFe
rmen
ted
dairy
pro
duct
s: y
ogur
tan
d m
ozza
rella
che
ese
Leuc
onos
toc
mes
ente
roid
esA
TCC
829
3Lm
e2.
037
2043
15.9
cocc
iW
ides
prea
d in
nat
ural
envi
ronm
ents
: cro
p pl
ants
, fru
itsan
d ve
geta
bles
, dai
ry p
rodu
cts
and
brea
d do
ugh
Oen
ococ
cus o
eni
PSU
-1O
oe1.
737
1916
18.0
cocc
iFr
uit m
ashe
s and
rela
ted
habi
tats
;em
ploy
ed in
win
erie
s to
carr
y ou
tm
alol
actic
con
vers
ion
Bifid
obac
teri
um lo
ngum
DJ0
10A
Blo
2.3
6020
1515
.1br
anch
ed ro
dIn
test
ines
of h
uman
s and
oth
eran
imal
s inc
ludi
ng in
sect
s; fe
mal
eva
gina
Brev
ibac
teri
um li
nens
BL2
Bli
4.4
6240
4013
.4ro
dM
ilk a
nd c
hees
e cr
ust
The
orde
r of t
he o
rgan
ism
s in
this
tabl
e an
d th
roug
hout
this
repo
rt is
bas
ed o
n th
eir p
hylo
gene
tic g
roup
ing
star
ting
with
Ppe
and
follo
win
g cl
ockw
ise
arou
nd th
e tre
e as
show
n in
Fig
ure
1.
a The
3-le
tter a
bbre
viat
ion
of th
e or
gani
sm u
sed
thro
ugho
ut th
is re
port
b App
roxi
mat
e si
ze o
f the
gen
ome
in M
egab
ase
pairs
, bas
ed o
n th
e cl
osed
gen
ome
sequ
ence
s, bu
t inc
ludi
ng n
atur
al p
lasm
ids
c Ave
rage
gen
ome
G+C
con
tent
d Num
ber o
f gen
es id
entif
ied
in e
ach
geno
me
incl
udin
g ge
nes e
ncod
ed o
n na
tura
l pla
smid
s pre
sent
in so
me
of th
ese
orga
nism
s
e Perc
ent o
f gen
es e
ncod
ing
reco
gniz
able
tran
spor
t pro
tein
hom
olog
ues
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 33Ta
ble
2C
arbo
hydr
ate
utili
zatio
n an
d ke
y pr
oper
ties o
f the
ele
ven
Gra
m-p
ositi
ve b
acte
ria in
clud
ed in
this
stud
y
Abb
.aM
ode
of F
erm
enta
tionb
Hex
ose
Ferm
enta
tion
Path
way
Oth
er In
form
atio
n
Ppe
Hom
ofer
men
tativ
eG
lyco
lysi
sPe
ntos
es fe
rmen
ted
thro
ugh
6-PG
/PK
. Hyd
roly
ze a
rgin
ine.
Lbr
Gro
up II
I (ob
ligat
ely
hete
rofe
rmen
tativ
e)6-
phos
phog
luco
nate
/pho
spho
keto
lase
pat
hway
(6-P
G/P
K)
Res
ista
nt to
hop
bitt
erin
g su
bsta
nces
like
isoh
umul
one.
Pro
duce
biog
enic
am
ines
.Lc
aG
roup
II (f
acul
tativ
ely
hete
rofe
rmen
tativ
e)G
lyco
lysi
sPe
ntos
es fe
rmen
ted
thro
ugh
the
6-PG
/PK
pat
hway
.
Lde
Gro
up I
(obl
igat
ely
hom
ofer
men
tativ
e)G
lyco
lysi
sD
o no
t fer
men
t pen
tose
s.
Lga
Gro
up I
(obl
igat
ely
hom
ofer
men
tativ
e)G
lyco
lysi
sPr
oduc
e ba
cter
ioci
ns; a
ssoc
iate
d w
ith p
robi
otic
act
iviti
es.
Lcr
Hom
ofer
men
tativ
eG
lyco
lysi
sPr
oduc
e un
ique
com
poun
ds v
ia a
min
o ac
id c
atab
olis
m.
Sth
Hom
ofer
men
tativ
eG
lyco
lysi
sPr
oduc
e ex
opol
ysac
char
ides
; util
ize
prot
eins
and
pep
tides
.Lm
eH
eter
ofer
men
tativ
e6-
PG/P
KFe
rmen
t citr
ate
to d
iace
tyl a
nd a
ceto
in; p
rodu
ce d
extra
ns a
ndle
vans
.O
oeH
eter
ofer
men
tativ
e6-
PG/P
KH
ighl
y re
sist
ant t
o et
hano
l.B
loH
eter
ofer
men
tativ
eFr
ucto
se-6
-pho
spha
te p
hosp
hoke
tola
se (F
6PPK
)R
esis
tant
to m
ethy
l qui
nolo
nes,
hete
rocy
clic
am
ines
,ni
trosa
min
es a
nd a
zom
etha
ne.
Bli
Stric
t aer
obe
Do
not f
erm
ent s
ugar
sPr
oduc
e ba
se, h
igh
leve
ls o
f sul
fur c
ompo
unds
, bac
terio
cins
and
caro
teno
ids;
deg
rade
pes
ticid
es (D
TT a
nd D
DE)
; met
abol
ize
amin
o ac
ids t
o pl
ant g
row
th h
orm
ones
.
a See
Tabl
e 1
for o
rgan
ism
al a
bbre
viat
ions
.
b The
phys
iolo
gica
l bas
is fo
r the
div
isio
n of
lact
obac
illi i
n th
ree
cate
gorie
s (G
roup
s I–I
II) i
s the
pre
senc
e or
abs
ence
of t
he k
ey e
nzym
es o
f hom
o- a
nd h
eter
ofer
men
tativ
e su
gar m
etab
olis
m, f
ruct
ose-
1,6-
bisp
hosp
hate
ald
olas
e an
d ph
osph
oket
olas
e, re
spec
tivel
y [1
35–1
37].
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 34Ta
ble
3N
umbe
rs o
f put
ativ
e tra
nspo
rt pr
otei
ns w
ith v
aryi
ng p
redi
cted
topo
logi
es
TM
SPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
071
8312
590
6685
8582
8161
113
122
3044
3125
4036
4734
4352
28
79
83
129
812
69
38
67
155
712
112
910
410
1323
916
2316
721
1522
519
2333
2926
1921
2730
1813
627
3645
2834
3524
3037
5159
712
921
1420
1317
1115
818
812
1212
512
158
67
1221
97
610
710
117
128
817
1015
2130
1918
1416
2016
2033
1112
1620
1311
1510
1613
625
1235
5531
1235
3114
3537
3910
213
1113
85
76
59
64
2114
1412
186
811
411
154
1815
31
11
12
116
12
11
317
118
11
192
241
251
Tota
l28
734
343
929
129
933
928
432
434
430
554
2
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 35Ta
ble
4To
tal n
umbe
rs (u
pper
) and
tota
l per
cent
(low
er) o
f tra
nspo
rt pr
otei
ns th
at b
elon
g to
eac
h TC
cla
ss a
nd su
bcla
ss
Cla
ssSu
bcla
ssPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
1. C
hann
els
139
105
1014
914
88
51.
A. α
-Hel
ical
pro
tein
cha
nnel
s10
810
58
118
88
85
1.C
. Tox
in c
hann
els
10
00
21
06
00
01.
E. H
olin
s2
10
00
21
00
00
2. E
lect
roch
emic
al P
oten
tial-d
riven
2.A
. Por
ters
8312
086
5666
7443
8887
6921
03.
Prim
ary
Act
ive
Tran
spor
ters
106
154
217
173
131
176
165
155
154
171
218
3.A
. P-P
-bon
d hy
drol
ysis
-driv
ensy
stem
s10
315
121
317
113
116
916
315
314
816
520
4
3.B
. Dec
arbo
xyla
tion-
driv
en sy
stem
s3
24
20
52
23
26
3.D
. Oxi
dore
duct
ion-
driv
en sy
stem
s0
10
00
20
03
48
4. G
roup
Tra
nslo
cato
rs4.
A. P
hosp
hotra
nsfe
r-dr
iven
syst
ems
377
639
3619
1520
301
05.
Tra
nsm
embr
ane E
lect
ron
Car
riers
5.A
. Tw
o-el
ectro
n tra
nsfe
r car
riers
00
00
00
00
00
78.
Acc
esso
ry F
acto
rs In
volv
ed in
Tran
spor
t8.
A A
uxili
ary
trans
port
prot
eins
811
125
96
913
74
6
9. In
com
plet
ely
Cha
ract
eriz
edTr
ansp
ort S
yste
ms
2124
1617
1725
1918
1920
38
9.A
Rec
ogni
zed
trans
porte
rs o
fun
know
n bi
oche
mic
al m
echa
nism
22
11
03
23
02
1
9.B
Put
ativ
e un
char
acte
rized
tran
spor
tpr
otei
ns19
2215
1617
2217
1519
1837
N/A
119
1835
2630
2524
1639
3258
Tota
l28
734
343
929
129
933
928
432
434
430
554
2
Cla
ssSu
bcla
ssPp
eLb
rLc
aLd
eLg
aLc
rSt
hLm
eO
oeB
loB
li
1. C
hann
els
4.5
2.6
2.3
1.7
3.3
4.1
3.2
4.3
2.3
2.6
0.9
1.A
. α-H
elic
al p
rote
in c
hann
els
3.5
2.3
2.3
1.7
2.7
3.2
2.8
2.5
2.3
2.6
0.9
1.C
. Tox
in c
hann
els
0.3
0.0
0.0
0.0
0.7
0.3
0.0
1.9
0.0
0.0
0.0
1.E.
Hol
ins
0.7
0.3
0.0
0.0
0.0
0.6
0.4
0.0
0.0
0.0
0.0
2. E
lect
roch
emic
al P
oten
tial-d
riven
2.A
. Por
ters
28.9
35.0
19.6
19.2
22.1
21.8
15.1
27.2
25.3
22.6
38.7
3. P
rimar
y A
ctiv
e Tr
ansp
orte
rs36
.944
.949
.459
.543
.851
.958
.147
.844
.856
.140
.23.
A. P
-P-b
ond
hydr
olys
is-d
riven
syst
ems
35.9
44.0
48.5
58.8
43.8
49.9
57.4
47.2
43.0
54.1
37.6
3.B
. Dec
arbo
xyla
tion-
driv
en sy
stem
s1.
00.
60.
90.
70.
01.
50.
70.
60.
90.
71.
13.
D. O
xido
redu
ctio
n-dr
iven
syst
ems
0.0
0.3
0.0
0.0
0.0
0.6
0.0
0.0
0.9
1.3
1.5
4. G
roup
Tra
nslo
cato
rs4.
A. P
hosp
hotra
nsfe
r-dr
iven
syst
ems
12.9
2.0
14.4
3.1
12.0
5.6
5.3
6.2
8.7
0.3
0.0
5. T
rans
mem
bran
e Ele
ctro
n C
arrie
rs5.
A. T
wo-
elec
tron
trans
fer c
arrie
rs0.
00.
00.
00.
00.
00.
00.
00.
00.
00.
01.
38.
Acc
esso
ry F
acto
rs In
volv
ed in
Tran
spor
t8.
A A
uxili
ary
trans
port
prot
eins
2.8
3.2
2.7
1.7
3.0
1.8
3.2
4.0
2.0
1.3
1.1
9. In
com
plet
ely
Cha
ract
eriz
edTr
ansp
ort S
yste
ms
7.3
7.0
3.6
5.8
5.7
7.4
6.7
5.6
5.5
6.6
7.0
9.A
Rec
ogni
zed
trans
porte
rs o
fun
know
n bi
oche
mic
al m
echa
nism
0.7
0.6
0.2
0.3
0.0
0.9
0.7
0.9
0.0
0.7
0.2
9.B
Put
ativ
e un
char
acte
rized
tran
spor
tpr
otei
ns6.
66.
43.
45.
55.
76.
56.
04.
65.
55.
96.
8
N/A
a6.
65.
28.
08.
910
.07.
48.
54.
911
.310
.510
.7To
tal
100
100
100
100
100
100
100
100
100
100
100
a Not
ass
igne
d
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
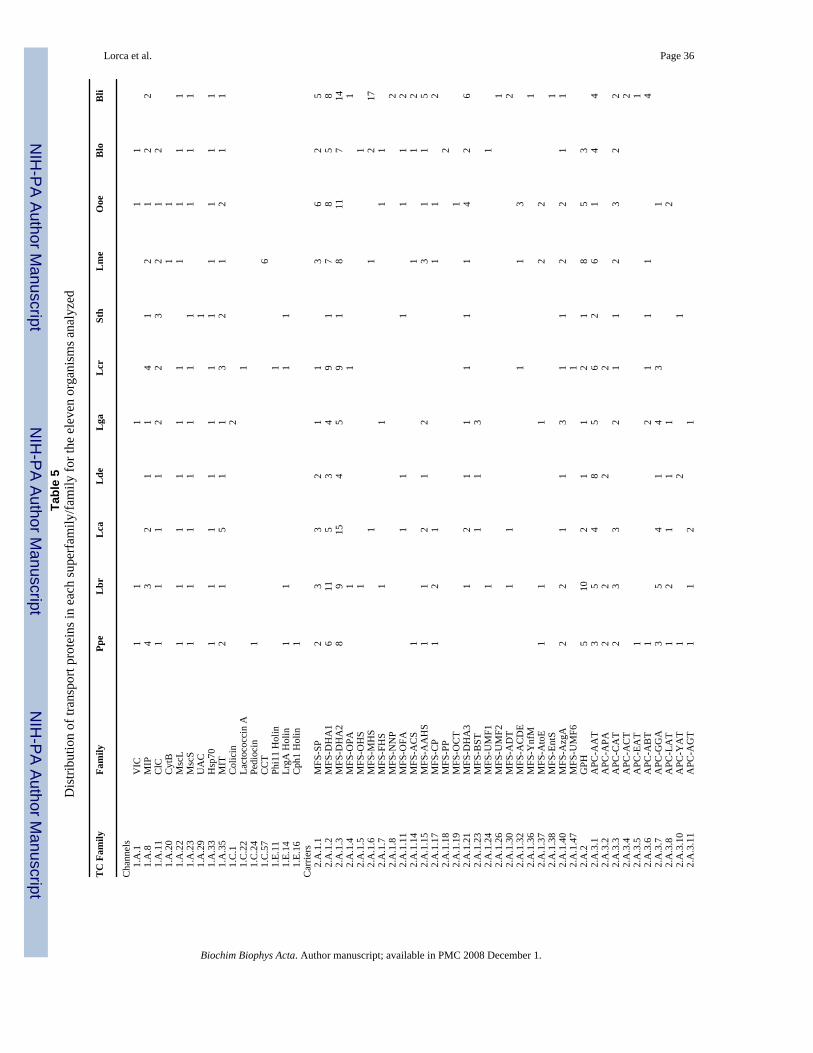
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 36Ta
ble
5D
istri
butio
n of
tran
spor
t pro
tein
s in
each
supe
rfam
ily/fa
mily
for t
he e
leve
n or
gani
sms a
naly
zed
TC
Fam
ilyFa
mily
Ppe
Lbr
Lca
Lde
Lga
Lcr
Sth
Lm
eO
oeB
loB
li
Cha
nnel
s
1.A
.1V
IC1
11
11
1.
A.8
MIP
43
21
14
12
12
2
1.A
.11
ClC
11
11
22
32
12
1.
A.2
0C
ytB
11
1.
A.2
2M
scL
11
11
11
11
11
1.
A.2
3M
scS
11
11
11
11
11
1.
A.2
9U
AC
1
1.A
.33
Hsp
701
11
11
11
11
11
1.
A.3
5M
IT2
15
11
32
12
11
1.
C.1
Col
icin
2
1.C
.22
Lact
ococ
cin
A1
1.
C.2
4Pe
dioc
in1
1.
C.5
7C
CT
6
1.E.
11Ph
i11
Hol
in1
1.
E.14
LrgA
Hol
in1
11
1
1.E.
16C
ph1
Hol
in1
Car
riers
2.
A.1
.1M
FS-S
P2
33
21
13
62
5
2.A
.1.2
MFS
-DH
A1
611
53
49
17
85
8
2.A
.1.3
MFS
-DH
A2
89
154
59
18
117
14
2.A
.1.4
MFS
-OPA
11
1
2.A
.1.5
MFS
-OH
S1
1
2.A
.1.6
MFS
-MH
S1
12
17
2.A
.1.7
MFS
-FH
S1
11
1
2.A
.1.8
MFS
-NN
P2
2.
A.1
.11
MFS
-OFA
11
11
12
2.
A.1
.14
MFS
-AC
S1
11
2
2.A
.1.1
5M
FS-A
AH
S1
12
12
31
15
2.
A.1
.17
MFS
-CP
12
11
12
2.
A.1
.18
MFS
-PP
2
2.A
.1.1
9M
FS-O
CT
1
2.A
.1.2
1M
FS-D
HA
31
21
11
11
42
6
2.A
.1.2
3M
FS-B
ST1
13
2.
A.1
.24
MFS
-UM
F11
1
2.A
.1.2
6M
FS-U
MF2
1
2.A
.1.3
0M
FS-A
DT
11
2
2.A
.1.3
2M
FS-A
CD
E1
13
2.
A.1
.36
MFS
-Ynf
M1
2.
A.1
.37
MFS
-Ato
E1
11
22
2.
A.1
.38
MFS
-Ent
S1
2.
A.1
.40
MFS
-Azg
A2
21
13
11
22
11
2.
A.1
.47
MFS
-UM
F61
2.
A.2
GPH
510
21
12
18
53
2.
A.3
.1A
PC-A
AT
35
48
56
26
14
4
2.A
.3.2
APC
-APA
22
22
2.
A.3
.3A
PC-C
AT
23
32
11
23
22
2.
A.3
.4A
PC-A
CT
2
2.A
.3.5
APC
-EA
T1
1
2.A
.3.6
APC
-AB
T1
21
11
4
2.A
.3.7
APC
-GG
A3
54
14
31
2.
A.3
.8A
PC-L
AT
12
11
12
2.
A.3
.10
APC
-YA
T1
21
2.
A.3
.11
APC
-AG
T1
12
1
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
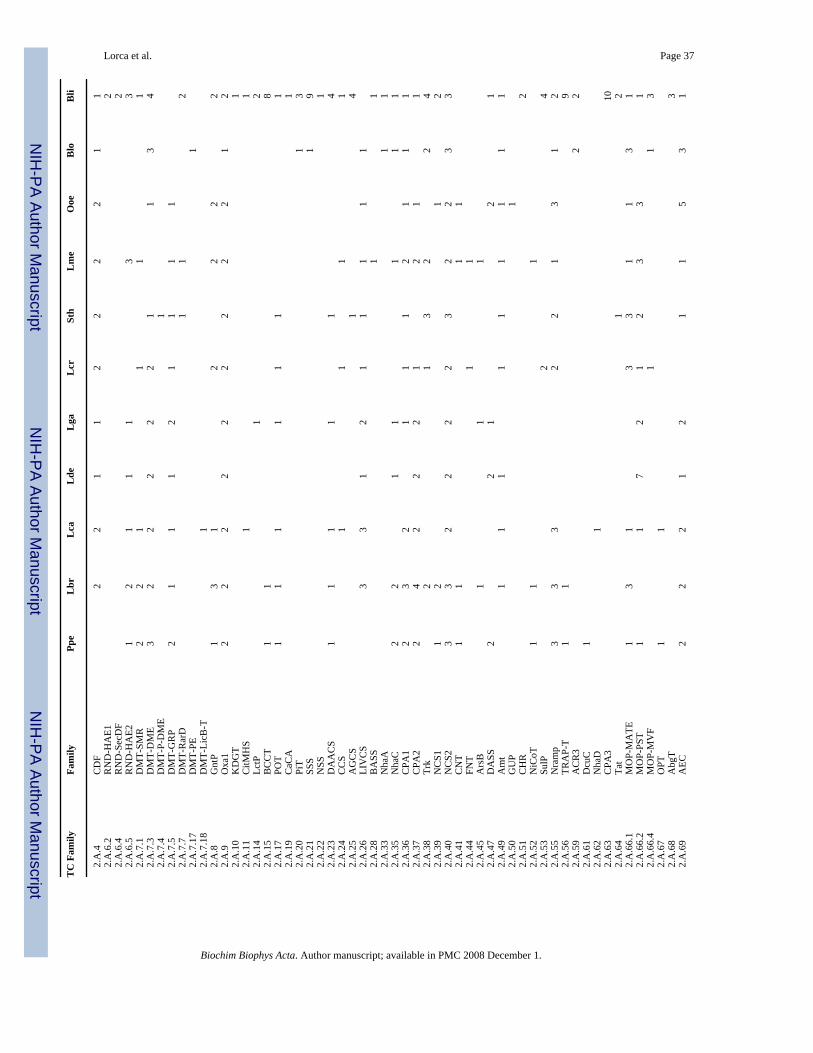
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 37T
C F
amily
Fam
ilyPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
2.
A.4
CD
F2
21
12
22
21
1
2.A
.6.2
RN
D-H
AE1
2
2.A
.6.4
RN
D-S
ecD
F2
2.
A.6
.5R
ND
-HA
E21
21
11
33
2.
A.7
.1D
MT-
SMR
22
11
11
2.
A.7
.3D
MT-
DM
E3
22
22
21
13
4
2.A
.7.4
DM
T-P-
DM
E1
2.
A.7
.5D
MT-
GR
P2
11
12
11
11
2.
A.7
.7D
MT-
Rar
D1
12
2.
A.7
.17
DM
T-PE
1
2.A
.7.1
8D
MT-
LicB
-T1
2.
A.8
Gnt
P1
31
22
22
2.
A.9
Oxa
12
22
22
22
22
12
2.
A.1
0K
DG
T1
2.
A.1
1C
itMH
S1
1
2.A
.14
LctP
12
2.
A.1
5B
CC
T1
18
2.
A.1
7PO
T1
11
11
11
2.
A.1
9C
aCA
1
2.A
.20
PiT
13
2.
A.2
1SS
S1
9
2.A
.22
NSS
1
2.A
.23
DA
AC
S1
11
11
4
2.A
.24
CC
S1
11
1
2.A
.25
AG
CS
14
2.
A.2
6LI
VC
S3
31
21
11
11
2.
A.2
8B
ASS
11
2.
A.3
3N
haA
11
2.
A.3
5N
haC
22
11
11
1
2.A
.36
CPA
12
32
11
12
11
1
2.A
.37
CPA
22
42
22
12
11
2.
A.3
8Tr
k2
13
22
4
2.A
.39
NC
S11
21
2
2.A
.40
NC
S23
32
22
23
22
33
2.
A.4
1C
NT
11
11
2.
A.4
4FN
T1
1
2.A
.45
Ars
B1
11
2.
A.4
7D
ASS
22
12
1
2.A
.49
Am
t1
11
11
11
11
2.
A.5
0G
UP
1
2.A
.51
CH
R2
2.
A.5
2N
iCoT
11
1
2.A
.53
SulP
24
2.
A.5
5N
ram
p3
33
22
13
12
2.
A.5
6TR
AP-
T1
19
2.
A.5
9A
CR
32
2
2.A
.61
Dcu
C1
2.
A.6
2N
haD
1
2.A
.63
CPA
310
2.
A.6
4Ta
t1
2
2.A
.66.
1M
OP-
MA
TE1
31
33
11
31
2.
A.6
6.2
MO
P-PS
T1
17
21
23
31
2.
A.6
6.4
MO
P-M
VF
11
3
2.A
.67
OPT
11
2.
A.6
8A
bgT
3
2.A
.69
AEC
22
21
21
15
31
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
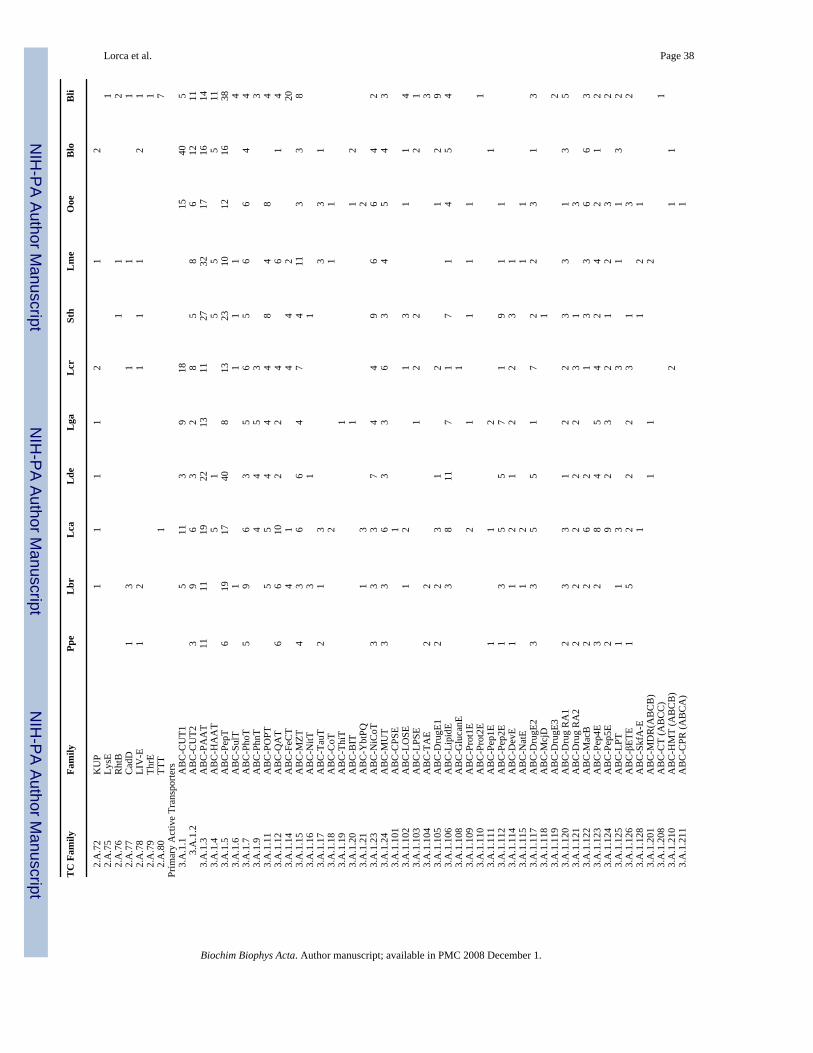
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 38T
C F
amily
Fam
ilyPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
2.
A.7
2K
UP
11
11
21
2
2.A
.75
LysE
1
2.A
.76
Rht
B1
12
2.
A.7
7C
adD
13
11
1
2.A
.78
LIV
-E1
21
11
21
2.
A.7
9Th
rE1
2.
A.8
0TT
T1
7Pr
imar
y A
ctiv
e Tr
ansp
orte
rs
3.A
.1.1
AB
C-C
UT1
511
39
1815
405
3.A
.1.2
AB
C-C
UT2
39
63
28
58
612
11
3.A
.1.3
AB
C-P
AA
T11
1119
2213
1127
3217
1614
3.
A.1
.4A
BC
-HA
AT
51
55
511
3.
A.1
.5A
BC
-Pep
T6
1917
408
1323
1012
1638
3.
A.1
.6A
BC
-Sul
T1
11
14
3.
A.1
.7A
BC
-Pho
T5
96
35
65
66
44
3.
A.1
.9A
BC
-Phn
T4
45
33
3.
A.1
.11
AB
C-P
OPT
55
44
48
48
4
3.A
.1.1
2A
BC
-QA
T6
610
22
46
14
3.
A.1
.14
AB
C-F
eCT
41
44
220
3.
A.1
.15
AB
C-M
ZT4
36
64
74
113
38
3.
A.1
.16
AB
C-N
itT3
11
3.
A.1
.17
AB
C-T
auT
21
33
31
3.
A.1
.18
AB
C-C
oT2
11
3.
A.1
.19
AB
C-T
hiT
1
3.A
.1.2
0A
BC
-BIT
11
2
3.A
.1.2
1A
BC
-Ybt
PQ1
32
3.
A.1
.23
AB
C-N
iCoT
33
37
44
96
64
2
3.A
.1.2
4A
BC
-MU
T3
36
33
63
45
43
3.
A.1
.101
AB
C-C
PSE
1
3.A
.1.1
02A
BC
-LO
SE1
21
31
14
3.
A.1
.103
AB
C-L
PSE
12
22
1
3.A
.1.1
04A
BC
-TA
E2
23
3.
A.1
.105
AB
C-D
rugE
12
23
12
12
9
3.A
.1.1
06A
BC
-Lip
idE
38
117
17
14
54
3.
A.1
.108
AB
C-G
luca
nE1
3.
A.1
.109
AB
C-P
rot1
E2
11
11
3.
A.1
.110
AB
C-P
rot2
E1
3.
A.1
.111
AB
C-P
ep1E
11
21
3.
A.1
.112
AB
C-P
ep2E
13
55
71
91
1
3.A
.1.1
14A
BC
-Dev
E1
12
12
23
1
3.A
.1.1
15A
BC
-Nat
E1
21
1
3.A
.1.1
17A
BC
-Dru
gE2
33
55
17
22
31
3
3.A
.1.1
18A
BC
-Mcj
D1
3.
A.1
.119
AB
C-D
rugE
32
3.
A.1
.120
AB
C-D
rug
RA
12
33
12
23
31
35
3.
A.1
.121
AB
C-D
rug
RA
22
22
22
31
3
3.A
.1.1
22A
BC
-Mac
B2
26
21
33
66
3
3.A
.1.1
23A
BC
-Pep
4E3
28
45
42
42
12
3.
A.1
.124
AB
C-P
ep5E
29
23
21
23
2
3.A
.1.1
25A
BC
-LPT
11
33
11
32
3.
A.1
.126
AB
C-β
ETE
15
22
23
13
2
3.A
.1.1
28A
BC
-Skf
A-E
11
21
3.
A.1
.201
AB
C-M
DR
(AB
CB
)1
12
3.
A.1
.208
AB
C-C
T (A
BC
C)
1
3.A
.1.2
10A
BC
-HM
T (A
BC
B)
21
1
3.A
.1.2
11A
BC
-CPR
(AB
CA
)1
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.
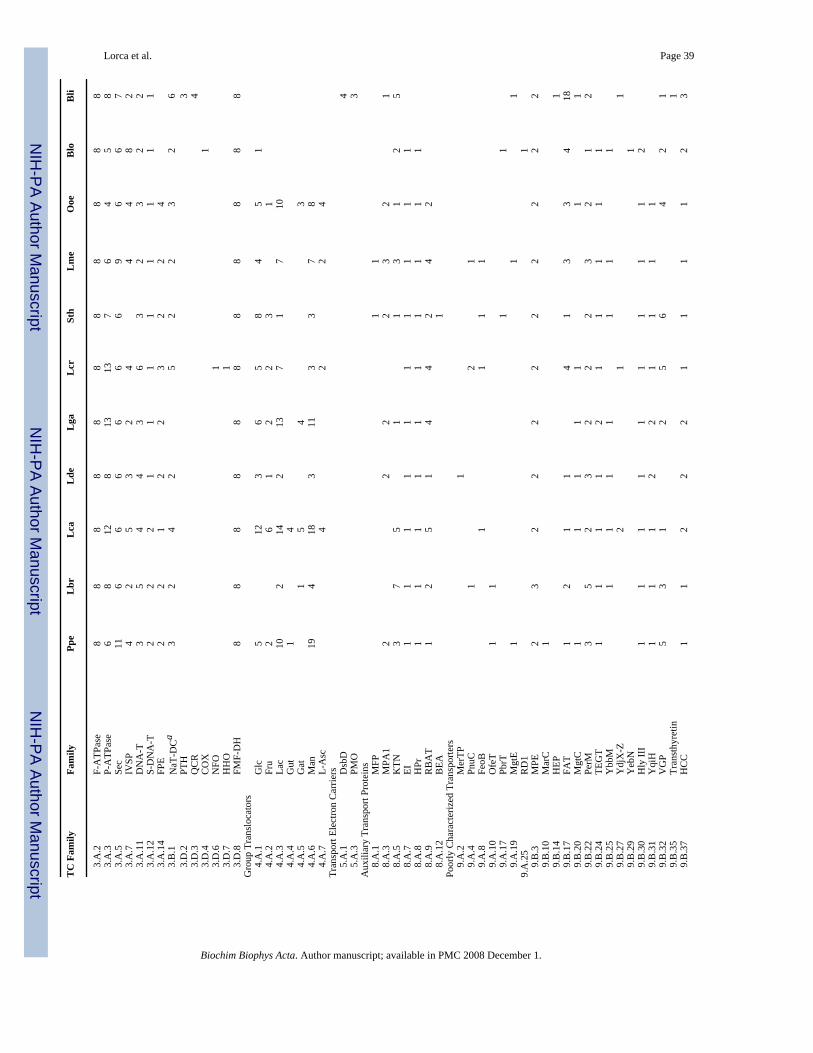
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 39T
C F
amily
Fam
ilyPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
3.
A.2
F-A
TPas
e8
88
88
88
88
88
3.
A.3
P-A
TPas
e6
812
813
137
64
58
3.
A.5
Sec
116
66
66
69
66
7
3.A
.7IV
SP4
25
32
44
48
2
3.A
.11
DN
A-T
35
44
36
32
32
2
3.A
.12
S-D
NA
-T2
22
11
11
11
11
3.
A.1
4FP
E2
21
22
32
24
3.
B.1
NaT
-DC
a3
24
25
22
32
6
3.D
.2PT
H3
3.
D.3
QC
R4
3.
D.4
CO
X1
3.
D.6
NFO
1
3.D
.7H
HO
1
3.D
.8FM
F-D
H8
88
88
88
88
88
Gro
up T
rans
loca
tors
4.
A.1
Glc
512
36
58
45
1
4.A
.2Fr
u2
61
22
31
4.
A.3
Lac
102
142
137
17
10
4.A
.4G
ut1
4
4.A
.5G
at1
54
3
4.A
.6M
an19
418
311
33
78
4.
A.7
L-A
sc4
22
4Tr
ansp
ort E
lect
ron
Car
riers
5.
A.1
Dsb
D4
5.
A.3
PMO
3A
uxili
ary
Tran
spor
t Pro
tein
s
8.A
.1M
FP1
1
8.A
.3M
PA1
22
22
32
1
8.A
.5K
TN3
75
11
31
25
8.
A.7
EI1
11
11
11
11
1
8.A
.8H
Pr1
11
11
11
11
1
8.A
.9R
BA
T1
25
14
42
42
8.
A.1
2B
EA1
Poor
ly C
hara
cter
ized
Tra
nspo
rters
9.
A.2
Mer
TP1
9.
A.4
PnuC
12
1
9.A
.8Fe
oB1
11
1
9.A
.10
Ofe
T1
1
9.A
.17
PbrT
11
9.
A.1
9M
gtE
11
19.
A.2
5 R
D1
1
9.B
.3M
PE2
32
22
22
22
22
9.
B.1
0M
arC
1
9.B
.14
HEP
1
9.B
.17
FAT
12
11
41
33
418
9.
B.2
0M
gtC
11
11
11
1
9.B
.22
PerM
35
23
22
23
21
2
9.B
.24
TEG
T1
11
12
11
11
1
9.B
.25
Ybb
M1
11
11
11
9.
B.2
7Y
djX
-Z2
11
9.
B.2
9Y
ebN
1
9.B
.30
Hly
III
11
11
11
11
12
9.
B.3
1Y
qiH
11
12
21
11
1
9.B
.32
VG
P5
31
25
64
21
9.
B.3
5Tr
anst
hyre
tin1
9.
B.3
7H
CC
11
22
21
11
12
3
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 40T
C F
amily
Fam
ilyPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
9.
B.4
4Y
iaA
B1
9.
B.4
8U
IT1
2
9.B
.50
UIT
31
21
11
9.
B.5
3U
IT6
1
9.B
.55
UIT
81
12
9.
B.5
7U
IT10
1
9.B
.59
Cst
A1
11
9.
B.6
39-
PME
11
11
11
11
11
Unc
lass
ified
tran
spor
ters
N
/AN
/A19
1835
2630
2524
1639
3258
a In th
e N
aT-D
C fa
mily
(3.B
.1),
the
pres
ence
of a
hom
olog
ue o
f the
dec
arbo
xyla
se is
not
indi
cativ
e of
the
pres
ence
of a
tran
spor
ter a
s the
se su
buni
ts c
an fu
nctio
n in
cyt
opla
smic
dec
arbo
xyla
tion
inde
pend
ently
of a
β-s
ubun
it w
hich
tran
spor
ts N
a+.
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 41Ta
ble
6N
umbe
rs o
f put
ativ
e tra
nspo
rters
pre
dict
ed to
exh
ibit
pola
r cha
ract
eris
tics:
cha
nnel
-type
bid
irect
iona
l tra
nspo
rt, u
ptak
e, o
r exp
ort
Dir
ectio
nPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
Bid
irect
iona
l7
75
35
44
73
56
Upt
ake
4748
5148
4645
4854
4849
49Ex
port
3233
3133
3135
3229
3029
30U
nkno
wna
1513
1316
1715
1710
1917
16
a The
pola
ritie
s of t
hese
tran
spor
ters
cou
ld n
ot b
e pr
edic
ted.
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 42Ta
ble
7Pe
rcen
t dis
tribu
tion
of tr
ansp
ort p
rote
ins a
ccor
ding
to su
bstra
te ty
pe
Cat
egor
yPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
Am
ino
acid
s & d
eriv
ativ
es19
2222
3322
1732
2516
1923
Car
bon
sour
ces
2315
239
2018
1015
2324
10D
rugs
and
toxi
c co
mpo
unds
1013
108
612
711
1211
12In
orga
nic
mol
ecul
es23
2823
2023
2524
3020
2030
Mac
rom
olec
ules
1110
1015
1112
139
1010
7U
nkno
wn
1413
1315
1715
1510
1817
18
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 43Ta
ble
8Pe
rcen
t dis
tribu
tion
of M
FS tr
ansp
orte
rs a
ccor
ding
to su
bstra
te ty
pe
Subs
trat
eaPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
Dru
gs64
6067
5748
8360
5763
5240
Ani
ons
1814
1821
294
2027
1219
44Su
gars
914
914
104
010
1722
7O
ther
s9
116
714
820
77
79
a All
drug
pum
ps c
atal
yze
efflu
x w
hile
ani
on a
nd su
gar t
rans
porte
rs c
atal
yze
upta
ke a
nd/o
r sub
stra
te e
xcha
nge.
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 44Ta
ble
9Pe
rcen
t dis
tribu
tion
of A
BC
eff
lux
and
upta
ke sy
stem
s acc
ordi
ng to
subs
trate
type
Cat
egor
yPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
A. E
FFLU
X
Dru
gs38
3022
1812
2918
2322
12
Lipi
ds0
516
2120
216
38
12
Pept
ides
88
1016
277
116
32
C
ompl
ex c
arbo
hydr
ates
43
00
25
30
02
Pr
otei
ns0
05
02
20
33
2
Inor
gani
c ca
tions
00
20
02
00
32
B. U
PTA
KE
A
min
o ac
ids
1914
1618
1510
2431
1916
Su
gars
48
55
717
86
1733
In
orga
nic
catio
ns8
119
52
108
148
7
Pept
ides
45
55
25
53
67
O
rgan
ic c
atio
ns8
85
55
55
66
0
Inor
gani
c an
ions
48
35
55
33
32
O
rgan
ic a
nion
s4
02
00
00
33
2
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 45Ta
ble
10Pe
rcen
t of p
utat
ive
mul
tidru
g re
sist
ance
(MD
R) e
fflu
x pu
mps
acc
ordi
ng to
supe
rfam
ily
Supe
rfam
ilya
Ppe
Lbr
Lca
Lde
Lga
Lcr
Sth
Lm
eO
oeB
loB
li
MFS
5055
5850
6354
2357
7264
57A
BC
3629
3444
3134
5429
2523
29M
OP
48
30
09
234
314
2R
ND
45
36
60
07
00
10D
MT
73
30
03
04
00
2
a The a
bbre
viat
ions
are:
MFS
, Maj
or F
acili
tato
r Sup
erfa
mily
; AB
C, A
TP-b
indi
ng C
asse
tte S
uper
fam
ily; M
OP,
Mul
tidru
g/O
ligos
acch
arid
yl-li
pid/
Poly
sacc
harid
e Flip
pase
Sup
erfa
mily
; RN
D, R
esis
tanc
e-N
odul
atio
n-D
ivis
ion
Supe
rfam
ily; D
MT,
Dru
g/M
etab
olite
Tra
nspo
rter S
uper
fam
ily
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 46Ta
ble
11Pe
rcen
t sug
ar u
ptak
e sy
stem
s fal
ling
into
the
thre
e ca
tego
ries o
f tra
nspo
rters
Fam
ilyPp
eL
brL
caL
deL
gaL
crSt
hL
me
Ooe
Blo
Bli
Eco
Bsu
Gra
m-
nega
tive
Path
ogen
s(B
bu, T
pa,
Hin
)
Gra
m-
posi
tive
Path
ogen
s(L
mo,
Spy
,M
ge)
2° c
arrie
r32
7016
4016
1818
5246
3858
2834
89
AB
C4
138
2010
3227
921
5842
2326
7016
PTS
6417
7640
7450
5539
324
049
4022
75
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lorca et al. Page 47Ta
ble
12Pe
rcen
t dis
tribu
tion
of fr
ee su
gar v
ersu
s olig
osac
char
ide/
glyc
osid
e tra
nspo
rt sy
stem
s
Det
ails
Ppe
Lbr
Lca
Lde
Lga
Lcr
Sth
Lm
eO
oeB
loB
liE
coB
suG
ram
-ne
gativ
ePa
thog
ens
(Bbu
, Tpa
,hi
n)
Gra
m-
posi
tive
Path
ogen
s(L
mo,
Spy
,M
ge)
Free
suga
rs46
3959
6045
3664
3950
3392
6555
8363
Olig
osac
char
ides
5461
4140
5564
3661
5067
835
4517
37
Biochim Biophys Acta. Author manuscript; available in PMC 2008 December 1.