transgene silencing in grapevines transformed with gflv resistance genes: analysis of variable...
TRANSCRIPT

ORIGINAL PAPER
Transgene silencing in grapevines transformed with GFLVresistance genes: analysis of variable expressionof transgene, siRNAs production and cytosine methylation
Giorgio Gambino Æ Irene Perrone Æ Andrea Carra ÆWalter Chitarra Æ Paolo Boccacci Æ Daniela Torello Marinoni ÆMarco Barberis Æ Fatemeh Maghuly Æ Margit Laimer ÆIvana Gribaudo
Received: 30 October 2008 / Accepted: 26 May 2009
� Springer Science+Business Media B.V. 2009
Abstract Eight transgenic grapevine lines trans-
formed with the coat protein gene of Grapevine fanleaf
virus (GFLV-CP) were analyzed for a correlation
between transgene expression, siRNAs production and
DNA methylation. Bisulphite genome sequencing was
used for a comprehensive analysis of DNA methylation.
Methylated cytosine residues of CpG and CpNpG sites
were detected in the GFLV-CP transgene, in the T7
terminator and in the 35S promoter of three grapevines
without transgene expression, but no detectable level of
siRNAs was recorded in these lines. The detailed
analysis of 8 lines revealed the complex arrangements
of T-DNA and integrated binary vector sequences as
crucial factors that influence transgene expression.
After inoculation with GFLV, no change in the levels
of cytosine methylation was observed, but transgenic
and untransformed plants produced short siRNAs (21–
22 nt) indicating that the grapevine plants responded to
GFLV infection by activating a post-transcriptional
gene silencing mechanism.
Keywords Bisulphite genomic sequencing �Coat protein � DNA methylation �Transgene expression stability � Vitis
Introduction
RNA silencing, a process leading to the degradation
of homologous mRNAs, has been widely observed in
animals, plants, and fungi (Hannon 2002). A key
feature of RNA silencing is the presence of small
RNAs such as micro RNAs (miRNAs) and small
interfering RNAs (siRNAs) that are processed from
double-stranded RNA (dsRNA) by a family of the
RNase III-like enzyme known as DICER (Vaucheret
2006). RNA silencing is an important mechanism
used by plants to defend themselves against viral
infection. In this process the dsRNA are originated
during viral replication and/or from internal pairing
Electronic supplementary material The online version ofthis article (doi:10.1007/s11248-009-9289-5) containssupplementary material, which is available to authorized users.
G. Gambino (&) � W. Chitarra � P. Boccacci �I. Gribaudo
Plant Virology Institute CNR (IVV), Grugliasco Unit. Via
L. da Vinci 44, 10095 Grugliasco-TO, Italy
e-mail: [email protected]
I. Perrone � A. Carra � W. Chitarra � D. Torello Marinoni
Department of Arboriculture and Pomology, University of
Torino, Via L. da Vinci 44, 10095 Grugliasco-TO, Italy
F. Maghuly � M. Laimer
Plant Biotechnology Unit, Institute of Applied
Microbiology BOKU, Nussdorfer Lande 11, 1190 Vienna,
Austria
M. Barberis
S.C.D.U. Medical Genetics, A.O.U. San Giovanni
Battista, Corso Bramante 88/90, 10126 Turin, Italy
123
Transgenic Res
DOI 10.1007/s11248-009-9289-5

of long RNA molecules (Vaucheret 2006). In addi-
tion, it has been suggested that host-encoded RNA-
dependent RNA polymerases also assist in battling
viral infection by using the viral genome as a
template for the production of dsRNA, which are
then diced to siRNAs (Xie et al. 2001). Virus- and
transgene-derived siRNAs accumulate in two distinct
size classes of 21 and 24 nt in plants (Hamilton et al.
2002). The 21 nt siRNAs are sufficient to guide the
cleavage of target RNA mediated by RNA-inducd
silencing complex (RISC), while the 24 nt species are
probably involved in chromatin modifications (Ham-
ilton et al. 2002; Fusaro et al. 2006). RNA silencing
also involves RNA-directed DNA methylation
(RdDM), in which DNA homologous to a dsRNA
that triggers gene silencing is methylated de novo
(Mette et al. 2000). RdDM has been documented in
many plant systems, and requires the enzyme know
as Domains Rearranged Methylase which could be
guided by siRNAs to the target sequences (Cao et al.
2003). In mammals, DNA methylation occurs almost
exclusively on cytosines in the symmetric dinucleo-
tides CpG. In plants, cytosine methylation occurs at
both symmetric sites (CpG and CpNpG, where N is
A, T or C) and at asymmetric sites (CpNpN). In
plants, several distinct RNA silencing pathways
operate to repress gene expression at the transcrip-
tional or post-transcriptional level (Baulcombe 2004).
In both cases silencing is generally associated with
siRNAs and/or DNA methylation: siRNAs homolo-
gous to the promoter region of a target gene induces
transcriptional gene silencing (TGS), which is asso-
ciated with promoter methylation. siRNAs homolo-
gous to the coding region of the target gene induces
post-transcriptional gene silencing (PTGS), which
involves mRNA specific degradation in the cyto-
plasm and, in some case, methylation of the coding
sequences (Baulcombe 2004).
Grapevine fanleaf virus (GFLV) is the main causal
agent of grapevine fanleaf disease, one of the most
damaging and widespread viral diseases affecting
grapevine (Vitis spp.). GFLV is spread through
propagating material and the nematode vector Xiph-
inema index. Control of viral vectors in the vineyards
either remains inefficient or is being restricted
because of the detrimental effects of the pesticides
on the environment (Andret-Link et al. 2004).
Therefore the approaches to engineer resistance in
grapevine would be desirable. Resistance to GFLV in
a few lines of transgenic rootstocks expressing the
coat protein gene has been reported after a three-year
trial in a naturally infected vineyard in France (Vigne
et al. 2004).
In grapevines transformed with the coat protein of
GFLV (GFLV-CP) obtained previously (Gambino
et al. 2005) we did not observe a strict correlation
between number of T-DNA insertion and mRNA
accumulation levels. Moreover, in these lines multi-
copy transgene insertions with repeated sequences
and integrated binary vector sequences in complex
arrangements were observed (Gambino et al. 2009).
In these transgenic grapevines, bisulphite genome
sequencing was employed for a comprehensive
analysis of DNA methylation on lines grown under
in vivo and in vitro conditions. Besides, the correla-
tions among transgene expression, siRNA produc-
tion and DNA methylation were analyzed in three
healthy transgenic grapevines and in the same
inoculated with GFLV through grafting onto infected
rootstocks.
Materials and methods
Plant material
Transgenic plants of grapevine (Vitis spp.) were
regenerated from embryogenic calli after co-cultivation
with Agrobacterium tumefaciens LBA4404 carrying a
binary Ti vector pGA643 (An et al. 1988) with
sequences of the GFLV-CP gene in sense and antisense
orientation (Golles et al. 1998) as described previously
(Gambino et al. 2005). Transgenic lines were obtained
from V. vinifera cv Nebbiolo (NE) and Blaufrankisch
(BF) and from V. berlandieri 9 V. rupestris 110 Richter
(110R). Eight lines were chosen for the analyses as
follow: NE-A and NE-C, a transgene multicopy and a
single-copy line respectively, without vector backbone
sequences integrated; NE-E having a single-copy of
T-DNA truncated; NE-B and NE-D, two multicopy
lines with vector backbone sequences integrated; NE-F,
a multicopy line with cytosine methylation at GFLV-CP
sequence; BF-A and 110R, two multicopy lines trans-
formed with GFLV-CP gene in antisense orientation
and with vector backbone sequences integrated (Gam-
bino et al. 2005, 2009).
Transgenic lines were micropropagated in vitro for
6 years (2002–2007), by repeatedly subculturing
Transgenic Res
123

apical cuttings (3–4 cm long) on a modified Murash-
ige and Skoog (1962) medium with half strength
mineral salts and 20 g l-1 sucrose without plant
growth regulators.
In 2004 some plants of each transgenic line were
acclimatized and transferred to a contained green-
house under natural daylight conditions; scions from
these plants were wedge-grafted in 2006 onto non-
transgenic rootstocks Kober 5BB (V. berlandi-
eri 9 V. riparia) infected by GFLV. For each line
10 scions were grafted onto the infected root-
stocks and grown in the greenhouse. Untransformed
V. vinifera cv Nebbiolo plants were used as controls.
Serological assays
The reaction of the plants to GFLV infection was
investigated by visually scoring symptom develop-
ment and by serological assays. Samples of young
apical leaves from transgenic and non transgenic
scions were collected from each tested plant in June
and September for 2 years after grafting. Samples of
phloem were scraped from mature canes collected
during winter from the same tested plants. Double
antibody sandwich enzyme-linked immunosorbent
assay (DAS-ELISA) was carried out using polyclonal
antisera to GFLV purchased from Agritest (Valenz-
ano, Italy). Samples were regarded as positive if their
OD405 value was at least three times the negative
control value.
Northern blot analysis
Total RNA was extracted from micropropagated
grapevine plantlets and from young apical leaves
collected from plants grown under greenhouse con-
ditions following the protocol of Gambino et al.
(2008). Total RNA (10 lg) was separated by elec-
trophoresis on a 1.2% denaturing formaldehyde-
agarose gel and capillary-blotted in 209 SSC (3 M
NaCl, 300 mM sodium citrate) onto a nylon mem-
brane positively charged (Roche). The GFLV-CP
gene, used as probe, was amplified by PCR using the
primers indicated by Gambino et al. (2005) and
digoxigenin labeled with a PCR Dig Probe Synthe-
sis Kit (Roche) according to the manufacturer’s
instructions. Labeled probe (20 ng/ml) was added
to the DIG easy Hyb (Roche) and the hybridization
was carried out overnight at 50�C. Following
hybridization, the membranes were incubated in the
CSPD chemiluminescent substrate following the
Roche protocol and exposed to Kodak� BioMaxTM
light film (Sigma). After autoradiographs the blots
were stripped (according to the Roche protocol) and
reprobed with neomycin phosphotransferase II (nptII)
(Supplemental Table 1) and 18S rDNA probes
(Gambino et al. 2008).
Bisulphite genomic sequencing
DNA was extracted using the method of Thomas et al.
(1993) from all grapevine lines grown in vitro, and
from young apical leaves collected from healthy
plants of lines NE-B, NE-C, NE-D, NE-F and 110R,
and from GFLV-infected transgenic plants of lines
NE-C, NE-F and 110R grown under greenhouse
conditions. To investigate the methylation status of
transgene, 35S promoter and T7 terminator sequences,
the bisulphite genomic sequencing technique was
used. Bisulphite treatment allows the conversion of all
cytosines to uracils, except those that are methylated
at the carbon-5 position. Since after the bisulphite
analyses the two strands of DNA are no longer
complementary, single strands can be amplified and
sequenced. The modified cytosines appear as thymi-
dines, whereas, methylated cytosines remain uncon-
verted. Bisulphite modification and desulfonation of
approximately 2 lg genomic DNA were performed
using the EpiTect Bisulfite kit (Qiagen) according to
the manufacturer’s instructions. In addition, to con-
firm that bisulphite mutagenesis was complete a
fragment of the GFLV movement protein (GFLV-
MP) was amplified by RT-PCR (Gambino et al. 2008)
with the primers reported in Supplemental Table 2
and cloned into the pGEM-T Easy vector (Promega)
according to the manufacturer’s instructions. In all
samples 50 pg of plasmid digested with SacII
(Promega) were mixed with grapevine genomic
DNA and subjected to bisulphite modification.
For bisulphite-PCR analysis of methylation differ-
ent primer pairs were used to amplify three fragments
of the sense strand of 35S promoter, GFLV-CP and
T7 terminator sequence (Fig. 1 ; Supplemental
Table 2). A mixture of primers with different nucle-
otides [either cytosine or thymine (Y) for the forward
primers, and adenine or guanine (R) for the reverse
primers], at positions corresponding to cytosine
residues in the target sequence, were used to cover
Transgenic Res
123

the possible states of methylation of cytosines,
including those bases that had not been converted
to uracils through the bisulphite reaction. Primers
designed using the software Primer 3 (Rozen and
Skaletsky 2000; http://frodo.wi.mit.edu/cgi-bin/primer3/
primer3_www.cgi) are listed in Supplemental Table 2.
The PCR reaction mix (20 ll) contained 200 ng of
bisulphite-treated DNA, 0.2 mM dNTPs, 2 lM each
primer, 2 mM MgSO4 and 0.5 unit Taq polymerase
(AccuPrimeTM Taq DNA Polymerase High Fidelity,
Invitrogen Life Technologies).
Cycling conditions for all primer pairs consisted of
initial denaturation at 94�C for 30 s, followed by 35
cycles at 94�C for 30 s, 50�C for 30 s and 68�C for
2 min. Reaction products were analyzed by electro-
phoresis on 1% agarose gels buffered in TBE
(45 mM Tris–borate, 1 mM EDTA) and visualised
by UV-light after staining with ethidium bromide.
Amplified PCR products were gel purified by Wiz-
ard� SV Gel and PCR Clean-Up System (Promega)
and cloned into the pGEM-T Easy vector (Promega).
The plasmid DNA was isolated by Wizard Plus SV
Minipreps DNA Purification Systems (Promega)
following the Promega protocol and sequenced using
M13 forward and reverse primers.
Five independent clones from each PCR product
were sequenced using a Big-Dye Terminator v1.1
Cycle Sequencing kit (Applied Biosystems) follow-
ing the manufacturer’s instructions. PCR products
were purified using an AutoSeq G-50 Dye Terminator
Removal kit (GE Healthcare) and analysed using a
3130 Genetic Analyzer capillary sequencer (Applied
Biosystems).
The sequences of individual clones were aligned
with the original references using the ClustalW
program 1.83 (Chenna et al. 2003; http://www.ebi.ac.uk/
c110Rstalw/). These alignment data were processed
by CyMATE program (Cytosine Methylation Analy-
sis Tool for Everyone; Hetzl et al. 2007; http://www.
gmi.oeaw.ac.at/CyMATE) for in silico analysis of
DNA sequences after bisulphite conversion of plant
DNA. The program allows to distinguish methylation
at CpG, CpNpG and CpNpN sites.
siRNAs detection
Small RNAs were extracted from young apical leaves
collected from healthy and GFLV-infected transgenic
plants (NE-C, NE-F and 110R) grown under green-
house conditions, following the protocol of Carra
Fig. 1 Schematic
representation of GFLV-CP
gene in a sense (GFLV-CP)
and b antisense (GFLV-AS)
orientation. Arrows indicate
direction of primers used
for bisulphite-PCR analysis
of methylation. The three
primer pairs were used to
amplify in three fragments
the filament sense of
transgene. Primers
sequences and locations
within the plasmid pGA643
and the GFLV-CP gene are
reported in Supplemental
Table 2. P 35S: CaMV 35S
promoter; T7:
Agrobacterium gene 7
transcription termination
region
Transgenic Res
123

et al. (2007). Low molecular weight RNA (3 lg per
lane) was separated on a 15% (v/w) polyacrylamide
gel containing 8 M urea. Gels were stained with
ethidium bromide and the RNA was transferred to
nylon membranes (Roche) with a Trans-blot semi-dry
transfer cell (Bio-Rad) at 2.5 mA/cm2 for 30 min.
RNA was cross-linked by exposing each side of the
membranes for 2.5 min to UV on a transilluminator,
followed by baking at 120�C for 30 min. Membranes
were analyzed both with 32P-a-UTP labelled probes
following the protocol of Szittya et al. (2002) and
with digoxigenin-labelled (DIG-dUTP) probes. For
DIG hybridization the membranes were prehybrid-
ized with PerfectHybTM Plus (Sigma) and hybridized
with DNA probe at 50�C overnight. Filters were
washed twice with 29 SSC-0.2% SDS and twice with
19 SSC-0.1% SDS at 50�C. The membranes were
then incubated in the CSPD chemiluminescent sub-
strate following the Roche protocol and exposed to
Kodak� BioMaxTM light film (Sigma). After autora-
diographs the blots were stripped (according to the
Roche protocol) and reprobed. Two regions of the
GFLV-CP gene were used as probes for this study: 50
region (50CP) and 30 region (30CP), amplified with the
primers reported in Supplemental Table 1.
Results
Transgene expression
The expression levels of the GFLV-CP gene were
studied in transgenic lines by Northern blot on in
vitro and in vivo plant material. The Northern assays
carried out on 6-year-old in vitro cultures showed a
stable GFLV-CP expression in 3/8 analyzed trans-
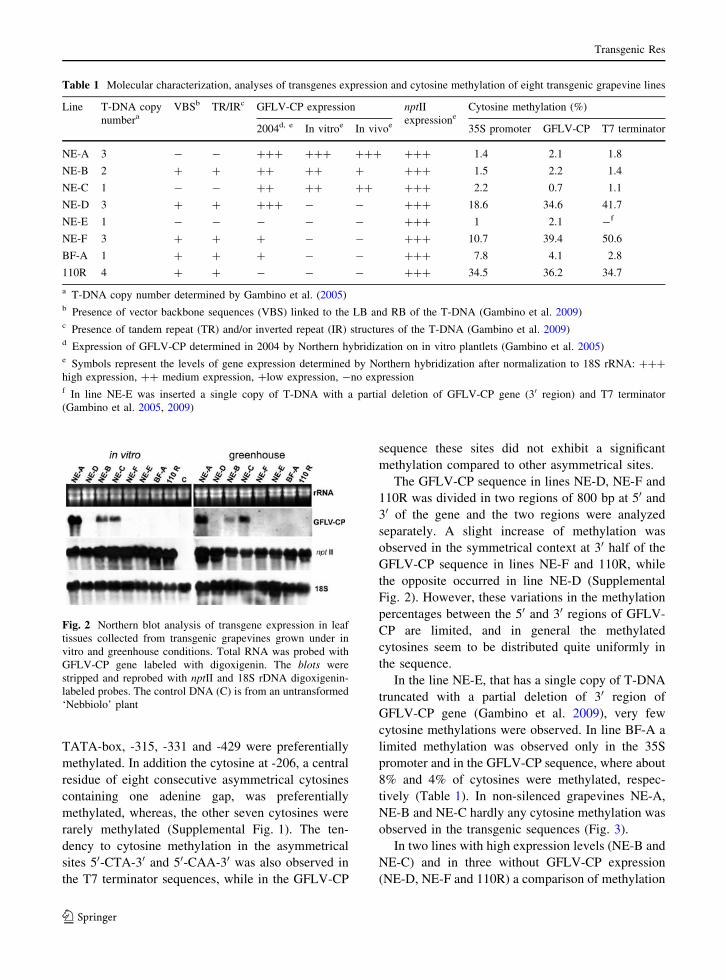
genic grapevines (Table 1; Fig. 2): high in NE-A,
slightly lower in NE-C and NE-B, while the mRNA
transcript was not detected in NE-E and 110R as well
as in the untransformed control plant. In three lines
(NE-D, NE-F and BF-A) a reduction of transgene
expression after the long-term in vitro culture was
observed, as transcripts of the GFLV-CP gene were
not detected by Northern hybridization. Previously a
low signal in Northern blot was detected for lines
NE-F and BF-A, while the GFLV-CP expression in
line NE-D was high with a hybridization signal
similar to that observed in NE-A (Table 1; Fig. 2)
(Gambino et al. 2005). In grapevines transferred to
the greenhouse the expression analyses were carried
out on young apical leaves (1-4 from the top)
collected in spring. The transgene expression in
plants grown in vivo conditions (Table 1; Fig. 2) is
not substantially different from that observed in the
same lines cultivated in vitro, except for NE-B, where
a slight reduction of GFLV-CP expression was
observed.
The gene for nptII, that was transferred with
GFLV-CP gene for selection of transgenic plants,
showed a high and stable expression in all grape-
vines, both in vitro and in vivo, including the lines
NE-D, NE-E, NE-F, BF-A and 110R that showed
GFLV-CP silencing (Table 1; Fig. 2).
Methylation status of the transgene
in grapevine plants
The methylation status of transgene, 35S promoter
and T7 terminator sequences were analyzed by
bisulphite genomic sequencing technique. In the
GFLV-MP gene used as unmethylated control of
the reaction, a high conversion rate of cytosines to
thymidines was obtained: the percentage of methyl-
ation varied from 0 to 1.1% for all samples. The
correlation between transgene expression and meth-
ylation was analyzed in all the 8 lines cultivated in
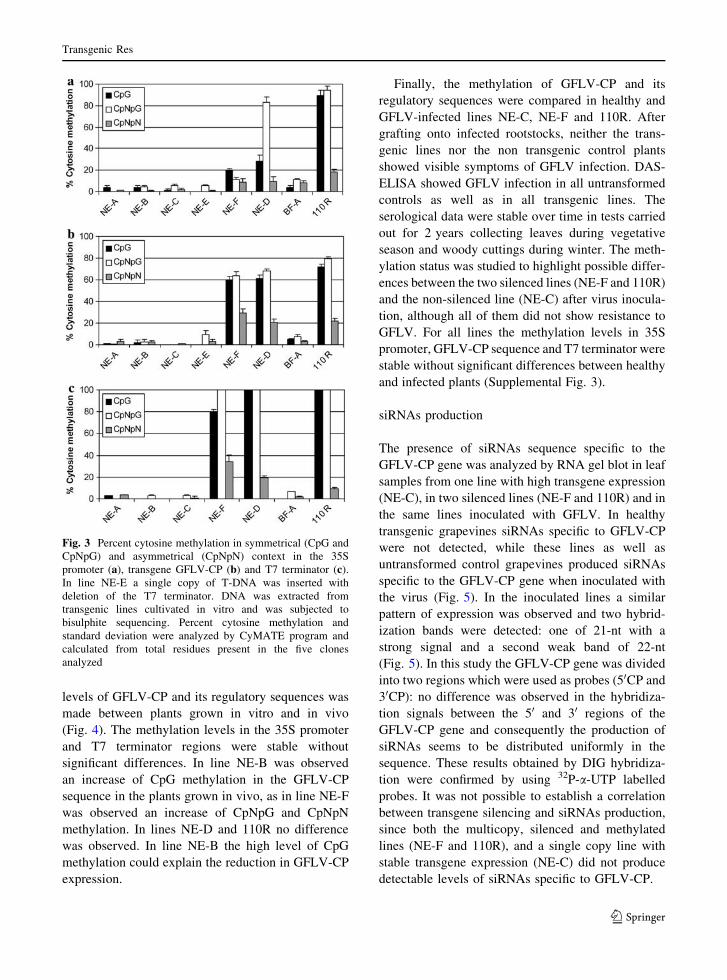
vitro (Table 1; Fig. 3). Comparisons of the methyl-
ation status in the non-methylated line NE-C and in
the highly methylated lines 110R for the 35S
promoter are reported in Supplemental Fig. 1. The
transgenic grapevine lines in which GFLV-CP tran-
script was not detected (NE-D, NE-F and 110R)
consistently exhibited cytosine methylation at the
CpG and CpNpG sites (Fig. 3) which were distrib-
uted in the GFLV-CP gene and in the T7 terminator
sequence. Cytosine methylation in asymmetrical
sequences (CpNpN) was also observed throughout
these regions, but at a lower frequency.
Furthermore, in lines NE-D and 110R high
frequencies of methylation were observed in sym-
metrical sites within the 35S promoter sequence
(Fig. 3), while in the asymmetrical cytosines the
methylation was observed at some residues of both
lines (Supplemental Fig. 1). 50-CTA-30 and 50-CAA-
30 cytosine sites had a much more prominent
tendency for methylation than other types of asym-
metrical cytosine sequence: cytosines at the positions
-76 (from the transcription start site) adjacent to
Transgenic Res
123
TATA-box, -315, -331 and -429 were preferentially
methylated. In addition the cytosine at -206, a central
residue of eight consecutive asymmetrical cytosines
containing one adenine gap, was preferentially
methylated, whereas, the other seven cytosines were
rarely methylated (Supplemental Fig. 1). The ten-
dency to cytosine methylation in the asymmetrical
sites 50-CTA-30 and 50-CAA-30 was also observed in
the T7 terminator sequences, while in the GFLV-CP
sequence these sites did not exhibit a significant
methylation compared to other asymmetrical sites.
The GFLV-CP sequence in lines NE-D, NE-F and
110R was divided in two regions of 800 bp at 50 and
30 of the gene and the two regions were analyzed
separately. A slight increase of methylation was
observed in the symmetrical context at 30 half of the
GFLV-CP sequence in lines NE-F and 110R, while
the opposite occurred in line NE-D (Supplemental
Fig. 2). However, these variations in the methylation
percentages between the 50 and 30 regions of GFLV-
CP are limited, and in general the methylated
cytosines seem to be distributed quite uniformly in
the sequence.
In the line NE-E, that has a single copy of T-DNA
truncated with a partial deletion of 30 region of
GFLV-CP gene (Gambino et al. 2009), very few
cytosine methylations were observed. In line BF-A a
limited methylation was observed only in the 35S
promoter and in the GFLV-CP sequence, where about
8% and 4% of cytosines were methylated, respec-
tively (Table 1). In non-silenced grapevines NE-A,
NE-B and NE-C hardly any cytosine methylation was
observed in the transgenic sequences (Fig. 3).
In two lines with high expression levels (NE-B and
NE-C) and in three without GFLV-CP expression
(NE-D, NE-F and 110R) a comparison of methylation
Table 1 Molecular characterization, analyses of transgenes expression and cytosine methylation of eight transgenic grapevine lines
Line T-DNA copy
numberaVBSb TR/IRc GFLV-CP expression nptII
expressioneCytosine methylation (%)
2004d, e In vitroe In vivoe 35S promoter GFLV-CP T7 terminator
NE-A 3 - - ??? ??? ??? ??? 1.4 2.1 1.8
NE-B 2 ? ? ?? ?? ? ??? 1.5 2.2 1.4
NE-C 1 - - ?? ?? ?? ??? 2.2 0.7 1.1
NE-D 3 ? ? ??? - - ??? 18.6 34.6 41.7
NE-E 1 - - - - - ??? 1 2.1 -f
NE-F 3 ? ? ? - - ??? 10.7 39.4 50.6
BF-A 1 ? ? ? - - ??? 7.8 4.1 2.8
110R 4 ? ? - - - ??? 34.5 36.2 34.7
a T-DNA copy number determined by Gambino et al. (2005)b Presence of vector backbone sequences (VBS) linked to the LB and RB of the T-DNA (Gambino et al. 2009)c Presence of tandem repeat (TR) and/or inverted repeat (IR) structures of the T-DNA (Gambino et al. 2009)d Expression of GFLV-CP determined in 2004 by Northern hybridization on in vitro plantlets (Gambino et al. 2005)e Symbols represent the levels of gene expression determined by Northern hybridization after normalization to 18S rRNA: ???
high expression, ?? medium expression, ?low expression, -no expressionf In line NE-E was inserted a single copy of T-DNA with a partial deletion of GFLV-CP gene (30 region) and T7 terminator
(Gambino et al. 2005, 2009)
Fig. 2 Northern blot analysis of transgene expression in leaf
tissues collected from transgenic grapevines grown under in
vitro and greenhouse conditions. Total RNA was probed with
GFLV-CP gene labeled with digoxigenin. The blots were
stripped and reprobed with nptII and 18S rDNA digoxigenin-
labeled probes. The control DNA (C) is from an untransformed
‘Nebbiolo’ plant
Transgenic Res
123
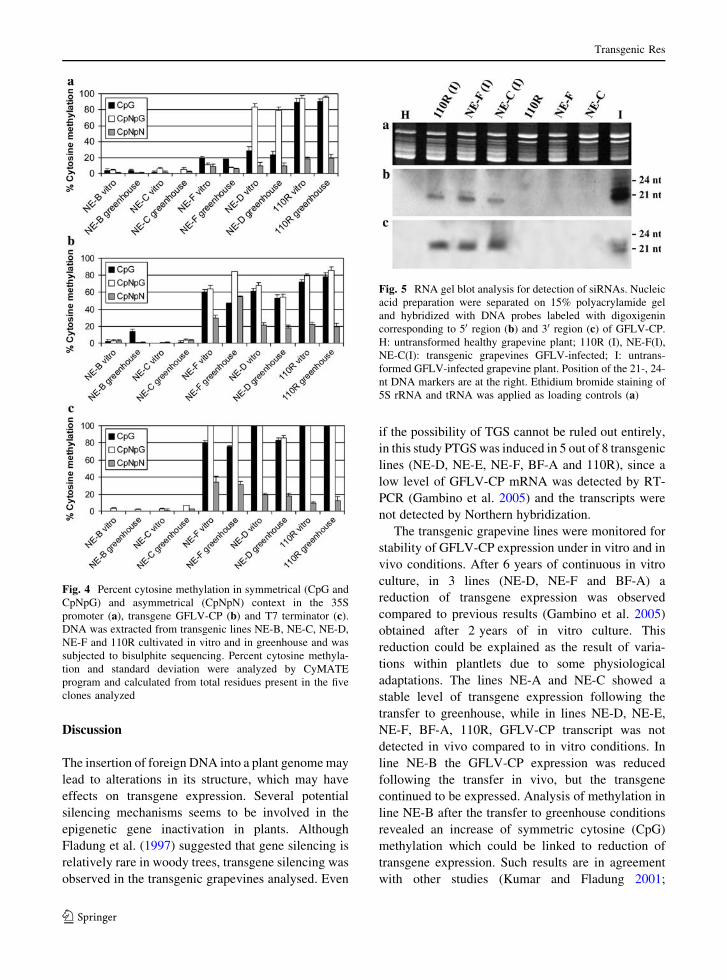
levels of GFLV-CP and its regulatory sequences was
made between plants grown in vitro and in vivo
(Fig. 4). The methylation levels in the 35S promoter
and T7 terminator regions were stable without
significant differences. In line NE-B was observed
an increase of CpG methylation in the GFLV-CP
sequence in the plants grown in vivo, as in line NE-F
was observed an increase of CpNpG and CpNpN
methylation. In lines NE-D and 110R no difference
was observed. In line NE-B the high level of CpG
methylation could explain the reduction in GFLV-CP
expression.
Finally, the methylation of GFLV-CP and its
regulatory sequences were compared in healthy and
GFLV-infected lines NE-C, NE-F and 110R. After
grafting onto infected rootstocks, neither the trans-
genic lines nor the non transgenic control plants
showed visible symptoms of GFLV infection. DAS-
ELISA showed GFLV infection in all untransformed
controls as well as in all transgenic lines. The
serological data were stable over time in tests carried
out for 2 years collecting leaves during vegetative
season and woody cuttings during winter. The meth-
ylation status was studied to highlight possible differ-
ences between the two silenced lines (NE-F and 110R)
and the non-silenced line (NE-C) after virus inocula-
tion, although all of them did not show resistance to
GFLV. For all lines the methylation levels in 35S
promoter, GFLV-CP sequence and T7 terminator were
stable without significant differences between healthy
and infected plants (Supplemental Fig. 3).
siRNAs production
The presence of siRNAs sequence specific to the
GFLV-CP gene was analyzed by RNA gel blot in leaf
samples from one line with high transgene expression
(NE-C), in two silenced lines (NE-F and 110R) and in
the same lines inoculated with GFLV. In healthy
transgenic grapevines siRNAs specific to GFLV-CP
were not detected, while these lines as well as
untransformed control grapevines produced siRNAs
specific to the GFLV-CP gene when inoculated with
the virus (Fig. 5). In the inoculated lines a similar
pattern of expression was observed and two hybrid-
ization bands were detected: one of 21-nt with a
strong signal and a second weak band of 22-nt
(Fig. 5). In this study the GFLV-CP gene was divided
into two regions which were used as probes (50CP and
30CP): no difference was observed in the hybridiza-
tion signals between the 50 and 30 regions of the
GFLV-CP gene and consequently the production of
siRNAs seems to be distributed uniformly in the
sequence. These results obtained by DIG hybridiza-
tion were confirmed by using 32P-a-UTP labelled
probes. It was not possible to establish a correlation
between transgene silencing and siRNAs production,
since both the multicopy, silenced and methylated
lines (NE-F and 110R), and a single copy line with
stable transgene expression (NE-C) did not produce
detectable levels of siRNAs specific to GFLV-CP.
Fig. 3 Percent cytosine methylation in symmetrical (CpG and
CpNpG) and asymmetrical (CpNpN) context in the 35S
promoter (a), transgene GFLV-CP (b) and T7 terminator (c).
In line NE-E a single copy of T-DNA was inserted with
deletion of the T7 terminator. DNA was extracted from
transgenic lines cultivated in vitro and was subjected to
bisulphite sequencing. Percent cytosine methylation and
standard deviation were analyzed by CyMATE program and
calculated from total residues present in the five clones
analyzed
Transgenic Res
123
Discussion
The insertion of foreign DNA into a plant genome may
lead to alterations in its structure, which may have
effects on transgene expression. Several potential
silencing mechanisms seems to be involved in the
epigenetic gene inactivation in plants. Although
Fladung et al. (1997) suggested that gene silencing is
relatively rare in woody trees, transgene silencing was
observed in the transgenic grapevines analysed. Even
if the possibility of TGS cannot be ruled out entirely,
in this study PTGS was induced in 5 out of 8 transgenic
lines (NE-D, NE-E, NE-F, BF-A and 110R), since a
low level of GFLV-CP mRNA was detected by RT-
PCR (Gambino et al. 2005) and the transcripts were
not detected by Northern hybridization.
The transgenic grapevine lines were monitored for
stability of GFLV-CP expression under in vitro and in
vivo conditions. After 6 years of continuous in vitro
culture, in 3 lines (NE-D, NE-F and BF-A) a
reduction of transgene expression was observed
compared to previous results (Gambino et al. 2005)
obtained after 2 years of in vitro culture. This
reduction could be explained as the result of varia-
tions within plantlets due to some physiological
adaptations. The lines NE-A and NE-C showed a
stable level of transgene expression following the
transfer to greenhouse, while in lines NE-D, NE-E,
NE-F, BF-A, 110R, GFLV-CP transcript was not
detected in vivo compared to in vitro conditions. In
line NE-B the GFLV-CP expression was reduced
following the transfer in vivo, but the transgene
continued to be expressed. Analysis of methylation in
line NE-B after the transfer to greenhouse conditions
revealed an increase of symmetric cytosine (CpG)
methylation which could be linked to reduction of
transgene expression. Such results are in agreement
with other studies (Kumar and Fladung 2001;
Fig. 4 Percent cytosine methylation in symmetrical (CpG and
CpNpG) and asymmetrical (CpNpN) context in the 35S
promoter (a), transgene GFLV-CP (b) and T7 terminator (c).
DNA was extracted from transgenic lines NE-B, NE-C, NE-D,
NE-F and 110R cultivated in vitro and in greenhouse and was
subjected to bisulphite sequencing. Percent cytosine methyla-
tion and standard deviation were analyzed by CyMATE
program and calculated from total residues present in the five
clones analyzed
Fig. 5 RNA gel blot analysis for detection of siRNAs. Nucleic
acid preparation were separated on 15% polyacrylamide gel
and hybridized with DNA probes labeled with digoxigenin
corresponding to 50 region (b) and 30 region (c) of GFLV-CP.
H: untransformed healthy grapevine plant; 110R (I), NE-F(I),
NE-C(I): transgenic grapevines GFLV-infected; I: untrans-
formed GFLV-infected grapevine plant. Position of the 21-, 24-
nt DNA markers are at the right. Ethidium bromide staining of
5S rRNA and tRNA was applied as loading controls (a)
Transgenic Res
123

Maghuly et al. 2007), which reported that stress and
other changes in environmental conditions as well as
the developmental stage of the plant could affect the
expression level of transgenes. The explants for
expression analyses were sampled from young leaves
in spring, according to previous works that reported
that highest transgene activity was usually observed
in young rather than in mature leaves (Maghuly et al.
2007). These seasonal variations are most probably
correlated with the physiological activity of the plant
material under study. The expression of the nptII
gene inserted in the T-DNA next to GFLV-CP was
stable both in vitro and in vivo, in all lines. The
kanamycin resistance gene is regulated by the nos
promoter, while the GFLV-CP gene is driven by the
35S, a strong promoter that appears more susceptible
to inactivation (McCabe et al. 1999).
The methylation status of cytosines present in the
transgene coding sequence was previously analyzed
by Southern hybridization using methylation-sensi-
tive restriction enzymes: cytosine methylation was
detected only in the GFLV-CP region in the trans-
genic line NE-F (Gambino et al. 2005). Those
methylation-sensitive restriction enzymes however,
allowed the investigation of the methylation status of
a limited number of cytosine residues because only a
limited number of recognition sites are available to
assess DNA methylation. Therefore in this study
bisulphite genomic sequencing was employed which
allows the conversion of all cytosines to uracils,
except those that are methylated at the carbon-5
position of C. The methylation status of GFLV-MP
gene cloned into the pGEM-T Easy vector was
determined to ascertain the complete chemical con-
version of the DNA. The low percentages (0–1.1%)
of methylation observed in this gene might indicate a
random base exchanges during PCR steps or an
incomplete conversion of non-methylated cytosines
after treatment with bisulphite. These technical
problems are not uncommon in the bisulphite geno-
mic sequencing approach. However, the high vari-
ability observed in the methylation levels of the
grapevines transgene sequences can be considered
reliable.
In three silenced lines (NE-D, NE-F and 110R)
transgene methylation was observed in both symmet-
rical and asymmetrical (at lower levels) contexts, and
a very low level of DNA methylation was observed in
non-silenced lines. In the 35S promoter sequence, the
50-CTA-30 and 50-CAA-30 cytosine sites have shown a
much more prominent tendency to methylation than
other types of asymmetrical cytosine. These results
coincide with the methylation observed in the SUP
locus in METI antisense transformants of Arabidopsis
(Kishimoto et al. 2001) and in the 35S promoter of
transgenic gentian (Mishiba et al. 2005). Inverted
repeats have been considered to trigger transgene
silencing and to be susceptible to de novo methylation
(Selker 1999). In this study the lines NE-F and 110R,
which contain inverted T-DNAs (Gambino et al.
2009), were silenced and had high methylation levels
in the 35S promoter, GFLV-CP and T7 terminator
sequences. However, the correlation between DNA
methylation and multiple copies of the integrated
transgene was not in agreement with cytosine meth-
ylation patterns observed in all analyzed lines. In
grapevine NE-B, that carries two T-DNA copies
linked to each other in tandem configuration with two
vector backbone sequences that are connected to the
left (LB) and right border (RB; Gambino et al. 2009),
no silencing and no methylation was observed. In line
BF-A, which carries one T-DNA copy linked by
vector backbone sequences to a second copy of
truncated T-DNA (Gambino et al. 2009), the trans-
gene silencing was associated with very low percent-
ages of cytosines methylation in the 35S promoter and
in the GFLV-CP transgene. The levels of transgene
cytosine methylation in silenced and non-silenced
grapevines indicated that transgene silencing was not
always associated with DNA methylation.
Methylation in CpG, CpNpG and CpNpN sites is a
notable feature of RNA-dependent DNA methylation
(RdDM) in plants, as reported by several authors in
different species (Aufsatz et al. 2002; Haque et al.
2007). Hamilton et al. (2002) proposed that siRNAs of
the longer class (24-25 nt) could be the trigger
molecule for RdDM. The short class of siRNAs (21
nt) is part of natural plant defense mechanism against
viruses (Voinnet 2005) and was proposed to guide the
RISC in the degrading specific mRNAs in herbaceous
plants (Hamilton et al. 2002). The accumulation of
different siRNAs size classes could be the result of
different inducers, virus infection or transgenes. In
plants, transgene- and virus-induced RNA silencing
pathways are overlapping but not identical (Voinnet
2005). The virus seems to act both as inducer and
target in this process, as observed in grapevine (this
study), in Prunus (Hily et al. 2005), in peach
Transgenic Res
123

inoculated with Peach latent mosaic viroid and in
chrysanthemum inoculated with Chrysanthemum clo-
rotic mottle viroid (Martinez de Alba et al. 2002). Hily
et al. (2005) demonstrated the accumulation of two
classes of siRNAs in transgenic Prunus resistant to
Plum pox virus and showed that those siRNAs are
constitutively present only in the resistant clone. The
resistance mounted by the shorter siRNAs is appar-
ently overcome and the longer siRNAs are responsible
for high levels of resistance against Plum pox virus. In
our grapevine lines the correlation between accumu-
lation of longer class of siRNAs, transgene methyl-
ation and RNA silencing could not be confirmed. In
the GFLV-CP transcript no small RNAs could be
detected both in methylated and non methylated lines,
while short siRNAs were detected only in transgenic
and non transgenic grapevines following GFLV
infection. These small RNAs originate from virus
replication and not from the GFLV-CP transgene.
However, it is possible that RNA signalling molecules
were responsible for the DNA methylation patterns
observed in these lines, even if siRNAs were below
the detection level. Similar results were observed
in Arabidospis, in which gene PA1 was methylated
at both symmetrical and asymmetrical contexts, but
no PA1 siRNAs cold be detected (Melquist and
Bender 2004); likewise, no siRNAs could be detected
in methylated CP gene of SPFMV in transgenic
N. benthamiana (Haque et al. 2007).
In this study eight transgenic grapevine lines
expressing GFLV-CP gene were tested for virus
resistance in greenhouse by grafting onto infected
rootstocks. All inoculated transgenic plants were
positive in ELISA, even when not showing GFLV
symptoms. The severity of symptoms following a
GFLV infection may be linked to environment and to
plant physiological conditions. The growth in pots
and in greenhouse could cause the lack of symp-
toms—at least for the first 2 years after inoculation—
both in the transgenic and in the control infected
grapevines; however, the serological results indicated
that the transgenic grapevines analyzed were suscep-
tible to GFLV transmitted by grafting onto infected
rootstocks. On the contrary Nicotiana benthamiana,
transformed with the same constructs and expressing
the full-length GFLV-CP gene, showed protection
against virus inoculation (Golles et al. 2000). Vigne
et al. (2004) showed that 3/18 transgenic grapevine
lines were resistant in the vineyard to natural GFLV
transmission by nematode vectors. Therefore, the
susceptibility to GFLV observed in the analyzed
transgenic grapevines could be due to high viral
inoculum from the rootstock applied to relatively
young and small plants, since Sonoda et al. (1999)
suggested a dose-dependent resistance. Probably
under these conditions the transgenic plants are
unable to suppress GFLV replication. Future tests
might include the use of natural vectors of the virus to
keep the viral pressure comparable to field condi-
tions. Further analyses will be necessary to investi-
gate thoroughly the complex relationships between
GFLV and silencing process in order to increase our
knowledge of the molecular mechanisms related to
transgene silencing and virus resistance in grapevine
as well as other woody species.
Acknowledgments We thank Federico Ghilino for excellent
support in the grafting, Danila Cuozzo and Tiziano Strano for
their technical assistance in micropropagation of transgenic
grapevines and in greenhouse management.
References
An G, Ebert PR, Mitra A, Ha SB (1988) Binary vectors. In:
Gelvin SB, Schilperoort RA (eds) Plant molecular biology
manual. Kluwer, Dortrecht p A3/1-19
Andret-Link P, Laporte C, Valat L et al (2004) Grapevine
fanleaf virus: still a major threat to the grapevine industry.
J Plant Pathol 86:183–195
Aufsatz W, Mette MF, van der Winden J et al (2002) RNA-
directed DNA methylation in Arabidopsis. Proc Natl Acad
Sci USA 99:16499–16506. doi:10.1073/pnas.162371499
Baulcombe D (2004) RNA silencing in plants. Nature
431:356–363. doi:10.1038/nature02874
Cao XF, Aufsatz W, Zilberman D et al (2003) Role of the
DRM and CMT3 Methyltransferases in RNA-directed
DNA methylation. Curr Biol 13:2212–2217. doi:10.1016/
j.cub.2003.11.052
Carra A, Gambino G, Schubert A (2007) A Cetyltrimethyl
ammonium bromide -based method to extract low molec-
ular weight RNA from polysaccharide-rich plant tissues.
Anal Biochem 360:318–320. doi:10.1016/j.ab.2006.09.022
Chenna R, Sugawara H, Koike T et al (2003) Multiple sequence
alignment with the Clustal series of programs. Nucleic
Acids Res 31:3497–3500. doi:10.1093/nar/gkg500
Fladung M, Kumar S, Ahuja R (1997) Genetic transformation
of Populus genotypes with different chimaeric gene con-
structs: transformation efficiency and molecular analysis.
Transgenic Res 6:111–121. doi:10.1023/A:101842162
0040
Fusaro AF, Matthew L, Smith NA et al (2006) RNA interfer-
ence-inducing hairpin RNAs in plants act through the
viral defence pathway. EMBO Rep 7:1168–1175. doi:
10.1038/sj.embor.7400837
Transgenic Res
123

Gambino G, Gribaudo I, Leopold S et al (2005) Molecular
characterization of grapevine plants transformed with
GFLV resistance genes: I. Plant Cell Rep 24:655–662.
doi:10.1007/s00299-005-0006-4
Gambino G, Perrone I, Gribaudo I (2008) A rapid and effective
method for RNA extraction from different tissues of
grapevine and other woody plants. Phytochem Anal
19:520–525. doi:10.1002/pca.1078
Gambino G, Chitarra W, Maghuly F, et al (2009) Character-
ization of T-DNA insertions in transgenic grapevines
obtained by Agrobacterium-mediated transformation. Mol
Breed. doi:10.1007/s11032-009-9293-8
Golles R, da Camara Machado A, Tsolova V et al (1998)
Transformation of somatic embryos of Vitis sp. with dif-
ferent constructs containing nucleotide sequences from
nepovirus coat protein genes. Acta Hortic 447:265–270
Golles R, Moser M, Puhringer H et al (2000) Transgenic
grapevines expressing coat protein gene sequences of
Grapevine fanleaf virus, Arabis mosaic virus, Grape-vine virus A and Grapevine virus B. Acta Hortic
528:305–311
Hamilton A, Voinnet O, Chappell L, Baulcombe D (2002) Two
classes of short interfering RNA in RNA silencing.
EMBO J 21:4671–4679. doi:10.1093/emboj/cdf464
Hannon GJ (2002) RNA interference. Nature 418:244–251.
doi:10.1038/418244a
Haque AKMN, Yamaoka N, Nishiguchi M (2007) Cytosine
methylation is associated with RNA silencing in silenced
plants but not with systemic and transitive RNA silencing
through grafting. Gene 396:321–331. doi:10.1016/j.gene.
2007.04.003
Hetzl J, Foerster AM, Raidl G, Scheid OM (2007) CyMATE: a
new tool for methylation analysis of plant genornic DNA
after bisulphite sequencing. Plant J 51:526–536. doi:
10.1111/j.1365-313X.2007.03152.x
Hily JM, Scorza R, Webb K, Ravelonandro M (2005) Accu-
mulation of the long class of siRNA is associated with
resistance to Plum pox virus in a transgenic woody
perennial plum tree. Mol Plant Microbe Interact 18:794–
799. doi:10.1094/MPMI-18-0794
Kishimoto N, Sakai H, Jackson J et al (2001) Site specificity of
the Arabidopsis METI DNA methyltransferase demon-
strated through hypermethylation of the superman locus.
Plant Mol Biol 46:171–183. doi:10.1023/A:10106362
22327
Kumar S, Fladung M (2001) Gene stability in transgenic aspen
(Populus). II. Molecular characterization of variable
expression of transgene in wild and hybrid aspen. Planta
213:731–740. doi:10.1007/s004250100535
Maghuly F, Machado A, Leopold S et al (2007) Long-term
stability of marker gene expression in Prunus subhirtella:
a model fruit tree species. J Biotechnol 127:310–321. doi:
10.1016/j.jbiotec.2006.06.016
Martinez de Alba AE, Flores R, Hernandez C (2002) Two
chloroplastic viroids induce the accumulation of small
RNAs associated with posttranscriptional gene silencing. J
Virol 76:13094–13096. doi:10.1128/JVI.76.24.13094-130
96.2002
McCabe MS, Schepers F, van der Arend A et al (1999)
Increased stable inheritance of herbicide resistance in
transgenic lettuce carrying a petE promoter-bar gene
compared with a CaMV 35S-bar gene. Theor Appl Genet
99:587–592. doi:10.1007/s001220051272
Melquist S, Bender J (2004) An internal rearrangement in an
Arabidopsis inverted repeat locus impairs DNA methyl-
ation triggered by the locus. Genetics 16:437–448. doi:
10.1534/genetics.166.1.437
Mette MF, Aufsatz W, van der Winden J et al (2000) Tran-
scriptional silencing and promoter methylation triggered
by double-stranded RNA. EMBO J 19:5194–5201. doi:
10.1093/emboj/19.19.5194
Mishiba K, Nishihara M, Nakatsuka T et al (2005) Consistent
transcriptional silencing of 35S-driven transgenes in
gentian. Plant J 44:541–556. doi:10.1111/j.1365-313X.
2005.02556.x
Murashige T, Skoog F (1962) A revised medium for rapid growth
and bio assays with tobacco tissue cultures. Physiol Plant
15:473–497. doi:10.1111/j.1399-3054.1962.tb08052.x
Rozen S, Skaletsky HJ (2000) Primer3 on the WWW for
general users and for biologist programmers. In: Krawetz
S, Misener S (eds) Bioinformatics methods and protocols:
methods in molecular biology. Humana Press, Totowa,
pp 365–386
Selker EU (1999) Gene silencing: repeats that count. Cell
97:157–160. doi:10.1016/S0092-8674(00)80725-4
Sonoda S, Mori M, Nishiguchi M (1999) Homology-dependent
virus resistance in transgenic plants with the coat protein
gene of Sweet potato feathery mottle potyvirus: target
specificity and transgene methylation. Phytopathology
89:385–391. doi:10.1094/PHYTO.1999.89.5.385
Szittya G, Molnar A, Silhavy D et al (2002) Short defective
interfering RNAs of tombusviruses are not targeted but
trigger post-transcriptional gene silencing against their
helper virus. Plant Cell 14:359–372. doi:10.1105/tpc.
010366
Thomas MR, Matsumoto S, Cain P, Scott NS (1993) Repetitive
DNA of grapevine: classes present and sequences suitable
for cultivar identification. Theor Appl Genet 86:173–180
Vaucheret H (2006) Post-transcriptional small RNA pathways
in plants: mechanisms and regulations. Genes Dev
20:759–771. doi:10.1101/gad.1410506
Vigne E, Komar V, Fuchs M (2004) Field safety assessment of
recombination in transgenic grapevines expressing the coat
protein gene of Grapevine fanleaf virus. Transgenic Res
13:165–179. doi:10.1023/B:TRAG.0000026075.79097.c9
Voinnet O (2005) Induction and suppression of RNA silencing:
insights from viral infections. Nat Rev Genet 6:206–220.
doi:10.1038/nrg1555
Xie Z, Fan B, Chen C, Chen Z (2001) An important role of an
inducible RNA-dependent RNA polymerase in plant
antiviral defense. Proc Natl Acad Sci USA 98:6516. doi:
10.1073/pnas.111440998
Transgenic Res
123