transcriptional and secretory responses of entamoeba histolytica to mucins, epithelial cells and...
TRANSCRIPT

www.elsevier.com/locate/ijpara
International Journal for Parasitology 37 (2007) 897–906
Transcriptional and secretory responses of Entamoeba histolyticato mucins, epithelial cells and bacteria
Anjan Debnath, Jessica S. Tashker 1, Mohammed Sajid, James H. McKerrow *
Sandler Center for Basic Research in Parasitic Diseases, University of California, San Francisco, San Francisco, CA 94158, USA
Received 20 October 2006; received in revised form 17 December 2006; accepted 9 January 2007
Abstract
Invasive intestinal amebiasis, caused by Entamoeba histolytica, is initiated with attachment of trophozoites to the colonic mucouslayer, mucous disruption and/or depletion, and adherence to and cytolysis of host epithelial and inflammatory cells. A current workingmodel of intestinal amebiasis suggests that the microenvironment of the host intestine, particularly intestinal mucins and the bacterialbiofilm, may influence the behavior of pathogenic amebae. The invasive phenotype is dependent on expression of a number of virulencefactors of which cysteine proteases provide the most convenient experimental probe because their activity is readily monitored. In thepresent study, we examined the interaction of E. histolytica with GalNAc, mucin, different epithelial cell lines and bacteria both by bio-chemical assays of protease release and transcriptional profiling using a previously validated genomic microarray. A significant down-regulation of released cysteine protease activity was observed when amebic trophozoites were grown with GalNAc, specific colonic celllines and bacteria. Transcriptional profiling during GalNAc interaction revealed enhanced expression of the 170-kDa Gal/GalNAc lec-tin. Decreased protease activity during GalNAc interaction and enhanced expression of the Gal/GalNAc lectin gene are consistent with aprogram of commensal infection and mucus coat colonization mediated by the lectin. The down-regulation of cysteine protease activityfollowing interaction with a colonic epithelial cell line parallels the presence of secretory mucin having a complex carbohydrate structurerich in Gal and GalNAc. In contrast, interaction of E. histolytica trophozoites with stomach porcine mucin enhanced cysteine protease(EhCP1 and EhCP2) secretion 3-fold. This suggests the specific composition of mucins may affect the Entamoeba phenotype. Transcrip-tional profiling revealed interaction of Entamoeba with intestinal bacteria induced protein kinase, ABC transporter, Rab family GTPaseand hsp 90 gene expression. The enhanced expression of this gene cluster is consistent with enhanced phagocytosis of E. histolytica duringinteraction with bacteria.� 2007 Published by Elsevier Ltd on behalf of Australian Society for Parasitology Inc.
Keywords: Entamoeba histolytica; Microarray; Protease; GalNAc; Mucin; Epithelial cells; Bacteria; Transcription
1. Introduction
Entamoeba histolytica, a protozoan intestinal parasite, isthe causative agent of human amebiasis. Infection with thisparasite is the fourth leading cause of death and the thirdleading cause of morbidity due to protozoan infectionsworldwide and is responsible for about 70,000 deaths annu-ally (World Health Organization, 1998). Infection can lead
0020-7519/$30.00 � 2007 Published by Elsevier Ltd on behalf of Australian S
doi:10.1016/j.ijpara.2007.01.016
* Corresponding author. Tel.: +1 415 476 2940; fax: +1 415 502 8193.E-mail address: [email protected] (J.H. McKerrow).
1 Present address: Institute for OneWorld Health, San Francisco, CA94111, USA.
to amebic colitis, colonic ulceration and, less frequently,dissemination to the liver, resulting in amebic liver abscess.
The pathogenesis of intestinal amebiasis has beenrelated to at least three separate and distinct phases: (i)attachment of trophozoites to the colonic mucous layerby the parasite surface galactose and N-acetylgalactose-amine-inhibitable lectin, (ii) mucous disruption and/ordepletion, and (iii) adherence to and cytolysis of host epi-thelial and inflammatory cells by the amoebapore and abattery of cysteine proteases (CPs) released by the parasiteinto its microenvironment.
Mucus is the first layer that protects the colonic epithe-lium from potentially pathogenic viruses, bacteria and
ociety for Parasitology Inc.

898 A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906
parasites (Belley et al., 1999). Colonic mucus also functionsas a lubricant and supports the growth of normal bacterialflora as a biofilm (Allen et al., 1982). Crude colonic mucuscontains large molecular weight mucin glycoproteins andnon-mucin components. Amebic invasiveness is controlledby the mucin glycoprotein layer on the intestinal epithe-lium. As amebae enter the mucin coat, their ability toinvade may be prevented by mucins binding to and inhib-iting the activity of the lectin and by the physical barrierbetween the parasite and host intestinal epithelium (Petriet al., 2002). This protective effect of mucin is preventedby free galactose or by treatment of the mucin withenzymes that remove galactose (Chadee et al., 1987). Theprotective effect is not necessarily a permanent one as thepresence of E. histolytica in the colon can alter the locallevel and composition of mucin in several ways (Eichinger,2001a). Amebae can degrade mucin (Connaris and Green-well, 1997) or bind and internalize it, resulting in the localproduction of mucopenic depressions (Prathap and Gil-man, 1970). Amebae also have a heat-stable mucus secreta-gogue activity, which results in goblet cell mucus secretionfrom both glandular and interglandular regions, the com-position of which may differ from the pre-existing mucinof the mucus layer (Chadee et al., 1991).
Another ‘‘host’’ factor influencing amebic virulence maybe the bacterial flora of the intestine. Amebic trophozoitesin the large intestine reside among bacteria in the mucinblanket that coats the intestinal epithelium. Bacteria influ-ence the balance between production of trophozoites andcysts, with only certain mixtures of bacteria capable of sup-porting encystation of trophozoites in vitro (Eichinger,2001b). Because of the inability of the cyst to invade, con-ditions that promote encystment should inhibit invasive-ness. Certain strains of bacteria have also been shown toalter the adherence and cytotoxicity of the trophozoitein vitro (Padilla-Vaca et al., 1999).
Entamoeba histolytica is known to release high levels of CPactivity into its environment and a direct correlation betweenCP activity and amebic virulence and invasiveness has beenconfirmed (Reed et al., 1989). The contribution of CPs in thepathogenesis of amebiasis includes facilitation of degradationof basement membrane and extracellular matrix componentsincluding fibronectin and collagen as well as immunoglobulinsand complement (Keene et al., 1986; Schulte and Scholze,1989; Tran et al., 1998). Nevertheless, the level of CP activityreleased by E. histolytica in culture fluctuates and this may rep-resent an ‘‘in vitro’’ correlate to how trophozoites respond totheir environment in the host and therefore the balancebetween commensal and invasive phenotypes. In order todetermine the role played by specific host factors, we examinedthe interaction of E. histolytica with GalNAc, mucin, differentepithelial cell lines and bacteria both by biochemical assays ofprotease release and transcriptional profiling using a previ-ously validated genomic microarray (Debnath et al., 2004).A significant down-regulation of released CP activity wasobserved when amebic trophozoites were grown with Gal-NAc, specific colonic cell lines and bacteria. The transcrip-
tional program identified during GalNAc, mucin andbacteria interaction provides new clues to how Entamoeba
orchestrates responses to the host environment.
2. Materials and methods
2.1. Cell cultures
The human colon adenocarcinoma cell line DLD1, Chi-nese hamster ovary (CHO) cell line and mutant CHO cellline Lec1 were obtained from the American Type CultureCollection (Rockville, MD). They were cultured in RPMI1640 with 25 mM N-2-Hydroxyethylpiperazine-N 0-2-ethanesulfonic acid (HEPES) medium (UCSF Cell CultureFacility) supplemented with 10% heat inactivated FBS,100 lg of streptomycin sulfate per ml and 100 U of penicil-lin per ml. Cultures were grown in plastic tissue cultureflasks and maintained in a humidified 5% CO2 atmosphereat 37 �C. Once the cells were confluent, they were harvestedby 0.25% trypsin digestion for 5 min.
2.2. Maintenance of Entamoeba cultures
Entamoeba histolytica trophozoites of the HM1:IMSSstrain were maintained in TYI-S-33 (Trypticase-YeastExtract-Iron-Serum) medium supplemented with penicillin(100 U/ml), streptomycin (100 lg/ml) and 10% bovineserum, according to the methods of Diamond (1984). Allexperiments were performed using trophozoites harvestedduring the logarithmic phase of growth.
2.3. Collection of ameba secretory products after GalNAc
incubation
A 0.5 M master solution of GalNAc (Sigma) was made upin PBS and sterilized by syringe filtration. Following harvest,trophozoites were washed with serum-free TYI medium andincubated in 2 ml of PBS (1 · 106 ameba/ml) in the presenceof different concentrations of GalNAc for 4 h at 37 �C in aGasPak� EZ Anaerobe Gas Generating Pouch System (Bec-ton, Dickinson and Company). In another experiment, thesame number of trophozoites was incubated in the absenceof GalNAc for 4 h at 37 �C. After incubation, the cells werechilled (4 �C) and collected by centrifugation (230g for5 min). The PBS conditioned by trophozoites was collectedand protein concentrations were determined by the methodof Bradford, using BSA as a standard (Bradford, 1976). Thesecretory products were stored at �80 �C until use.
2.4. Collection of ameba secretory products after mucin
interaction
A 1 mM master solution of porcine stomach mucin(Sigma) was prepared in sterile PBS. Trophozoites(1 · 106 ameba/ml) were incubated in 2 ml of PBS in pres-ence of different concentrations (ranging from 1 to 100 lM)of porcine stomach mucin for 4 h at 37 �C in a GasPak�

A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906 899
EZ Anaerobe Gas Generating Pouch System. In anotherexperiment, the same number of trophozoites was incu-bated in the absence of mucin for 4 h at 37 �C. The secre-tory products were stored at �80 �C until use.
2.5. Collection of ameba secretory products after interaction
with different cell lines
CHO, Lec1, and DLD1 cells were trypsinized andcounted using a hemocytometer, and variable number ofcells (5 · 104, 10 · 104, 25 · 104, 50 · 104 and 100 · 104)was plated in each well of a 6-well plate. Media withoutcells was used as a control. Cells were left to grow over-night at 37 �C. Mammalian cells were treated with 10 lME64d (Peptides International) and incubated for 1 h.E64d-treated cells were washed with warm PBS. Two mil-lilitres of trophozoites (25 · 104/ml) in serum-free M199medium (GIBCO) were placed in each well of the 6-wellplate. Plates were placed in a GasPak� EZ AnaerobeGas Generating Pouch System and incubated at 37 �C for4 h. The conditioned supernatant was collected and filteredthrough a 0.45 lM cellulose acetate syringe filter. Proteinconcentrations were measured using Bradford reagent.
2.6. Collection of ameba secretory products after bacteria
interaction
Trophozoites (2 · 105 ameba/ml) and Escherichia coli
K12 strain MG1655 (at a ratio of 1:1000) were co-culturedfor 1 h in 2 ml of serum-free TYI medium at 37 �C. Thesame number of trophozoites was also assayed in theabsence of bacteria. Secretory products were collected bothfrom control and bacteria-interacted trophozoites.
2.7. Enzymatic assays
All assays were performed in 100 mM Tris–HCl, 5 mMEDTA and 2 mM dithiothreitol (DTT) (pH 7.4). The abso-lute level of CP activity present in the supernatant wasassayed by measuring the activity of amebic secretions ona relatively specific peptide substrate, benzyloxycarbonyl-arginyl-4-amino-7-methylcoumarin (ZRR-AMC) (EnzymeSystem Products) as previously described (Keene et al.,1986). This peptide substrate was used because E. histolyti-
ca CP degrades peptides with arginine at P-1 and P-2(Keene et al., 1986). The rate of substrate hydrolysis atambient temperature was determined from the rate ofincrease of fluorescence, monitored on a continuouslyrecording spectrofluorometer and measured as RFU/min/lg protein. In one control for the GalNAc experiment,before measuring the CP activity the supernatant wassupplemented with the respective amount of GalNAc.
2.8. 125I-DCG-04 labeling
DCG-04 was iodinated using iodogen-coated beads(Pierce) as described by Greenbaum et al. (2002). Equal
amounts of protein (as determined by the protein standardcurve, data not shown) present in the supernatant releasedby the CHO, Lec1 and bacteria-treated trophozoites anduntreated trophozoites were labeled with 4 ll of [125I]-DCG-04 after adding 5 mM DTT. After 1 h incubationat room temperature the supernatants were separated ona 15% polyacrylamide gel and labeled polypeptides werevisualized with a PhosphorImager (Molecular Dynamics).Densitometric quantification of band intensities wasperformed using ImageQuant TL software (AmershamBiosciences). Values were obtained after subtracting thebackground value from each lane.
2.9. RNA isolation, labeled cDNA preparation and
microarray hybridization
Total RNA was isolated from untreated HM1, 4 h50 mM GalNAc-treated HM1, 3 h 50 lM porcine stomachmucin-treated HM1, 4 h 100 · 104 CHO-treated HM1 and1 h E. coli MG1655-treated HM1 using TRIZOL (Invitro-gen). cDNA synthesis, microarray hybridization andmicroarray scanning were performed as described (Deb-nath et al., 2004). Briefly, cDNA was synthesized fromtotal RNA extracted from untreated and treated HM1,coupled to CyScribe Cy3 or Cy5 (Amersham), and hybrid-ized to the shotgun DNA microarray constructed by use of6144 random inserts from an E. histolytica genomic DNAlibrary (Debnath et al., 2004). The hybridization was per-formed at 63 �C for 16 h in a humidified slide chamber con-taining the labeled probe, 3· sodium chloride–sodiumcitrate (SSC) and 0.2% SDS. After hybridization, thehybridization chamber was removed from the 63 �C waterbath, washed with 0.6· SSC-0.03% SDS, and then 0.06·SSC. Eleven microarray hybridizations were performed:two hybridizations with HM1 trophozoite RNA labeledwith Cy3 and GalNAc-treated trophozoite RNA labeledwith Cy5 (EhCS1p1-079 and EhCS1p1-080); two hybrid-izations with HM1 trophozoite RNA labeled with Cy3and mucin-treated trophozoite RNA labeled with Cy5(EhCS1p1-121 and EhCS1p1-122); three hybridizationswith HM1 trophozoite RNA labeled with Cy3 and CHO-treated trophozoite RNA labeled with Cy5 (EhCS1p1-125, EhCS1p1-126 and EhCS1p1-127); four hybridizationswith HM1 trophozoite RNA labeled with Cy3 and E. coli-treated trophozoite RNA labeled with Cy5 (EhCS1p1-110,EhCS1p1-111, EhCS1p1-112 and EhCS1p1-113). Scanningand data analysis were performed as previously described(Debnath et al., 2004). Only spots in which almost morethan 70% of the pixels had a signal of at least twice theSD of the local background were considered in subsequentdata analysis. The data were stored and normalized by useof the NOMAD microarray database (http://nomad2.ucs-f.edu/NOMAD/). The Cy5/Cy3 fluorescence ratios andlog2-transformed ratios were calculated from the normal-ized values. Data from related clones in different hybridiza-tion experiments were analyzed using CLUSTER analysisand were visualized using TREEVIEW. Clones whose

Fig. 1. Percentage of protease activity secreted by trophozoites in thepresence of GalNAc relative to protease secreted by trophozoites in theabsence of GalNAc. HM1:IMSS trophozoites (2 · 106) were incubated inthe absence and presence of different concentrations of GalNAc for 4 h at37 �C. Cysteine protease activity was determined as described in Section 2and measured as RFU/min/lg protein. The data represent the means andSD of four independent experiments.
900 A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906
expression increased or decreased at least 2.5-fold intreated HM1 relative to untreated HM1 were selected forfurther sequencing analysis.
2.10. Real-time PCR
Total RNA was extracted from trophozoites using TRI-ZOL (Invitrogen) and cDNA was synthesized with oligo(dT) (Invitrogen). PCR reactions were performed in dupli-cate using SYBR Green PCR Master Mix (Applied Biosys-tems). The PCR product was continually monitored in aMx3005P� QPCR System with MxPro� QPCR software(Stratagene), which analyzes the amplification curvesderived from the incorporation of SYBR Green into thedouble-stranded DNA product. Quantitative values forreal-time PCR were obtained from the threshold cyclenumber at which the increase in the signal associated withan exponential growth of PCR products was first detected.Final results, expressed as N-fold differences in target geneexpression relative to the reference gene E. histolytica actin(GenBank Accession No. M16339), termed ‘Ntarget’, weredetermined as follows:
Ntarget ¼ 2ðdCt sample�dCt reference geneÞ
where dCt values of the sample and reference were deter-mined by subtracting the average Ct value of the test genefrom the average Ct value of the actin gene.
The specificity of the PCR amplification was verified bymelting curves of the final amplicons. Changes in mRNAexpression were considered significant if the degree of expres-sion differed by P1.5-fold between two groups (Shazandet al., 2004; Kaushik et al., 2005). The primer sequences usedto amplify the clones were as follows: for the AEC 2837 clone,50-TGCACATGTCCAATGTGTTG-30 and 50-CCATGCATTTCACTTTTGGTT-30; for the AEC 3223 clone, 50-TGGACCATTTGCTGCTATGA-30 and 50-ATGACCCACAATTCCCTTGA-30; for the AEC 1278 clone, 50-GGACCTGTTGCTGTCGGTAT-30 and 50-ATGCTGTTCCCCATGAGTTT-30; for the AEC 5872 clone, 50-CCAATTTGGACAAGAAATCCA-30 and 50-TCAAATGGTGCTCTTTGTGG-30. The primer sequences for E. histolytica actinwere 50-CTTGATCTTGCAGGACGTGA-30 and 50-CTGCTTGATGCAGCTTTTTG-30.
2.11. Statistical analysis
The protease activity data represent the means and SEMof four independent experiments.
3. Results
3.1. GalNAc decreases protease release by Entamoeba
trophozoites
In order to determine whether GalNAc affects proteasesecretion into the media by E. histolytica, trophozoiteswere treated with concentrations of GalNAc ranging from
10 lM to 50 mM. A 4 h incubation was selected becauseduring this time period E. histolytica release significant lev-els of CP activity into the media without lysis of trophozo-ites as determined by alcohol dehydrogenase activity.GalNAc concentrations as low as 50 lM inhibited proteaserelease and at 50 mM GalNAc, protease secretion into themedia was inhibited by almost 65% (Fig. 1). In one control,sugar did not inhibit CP activity after supplementing thesupernatant with the respective amount of GalNAc.
3.2. Porcine gastric mucin enhances release of protease
activity by Entamoeba trophozoites
To determine how non-colonic mucin modulates CPsecretion into the media, trophozoites were treated withdifferent concentrations of porcine stomach mucin rangingfrom 1 to 100 lM. As shown in Fig. 2a, 4 h interaction of50 lM porcine stomach mucin with E. histolytica tropho-zoites enhanced CP secretion into the media almost 3.5-fold. The dose-dependent effect of mucin was observedbetween 0 and 5 lM. As the concentration of mucinincreased in 0–5 lM range protease secretion by thetrophozoites increased (Fig. 2b).
3.3. Release of protease activity varies by interaction of
trophozoites with different epithelial cells
We compared the released CP activity of trophozitesduring their interaction with different mammalian cell lines.Entamoeba histolytica trophozoites are able to disruptmonolayers of cultured mammalian cells. Compared withthe trophozoites incubated without cells, HM1 cells inter-acting with 100 · 104 CHO released 2-fold greater proteaseinto the media. Secretion of protease correlated with thenumber of interacting CHO cells. In contrast, trophozoitesincubated with 100 · 104 DLD1 cells released 50% less CP.The protease secretion decreased as the number of interact-
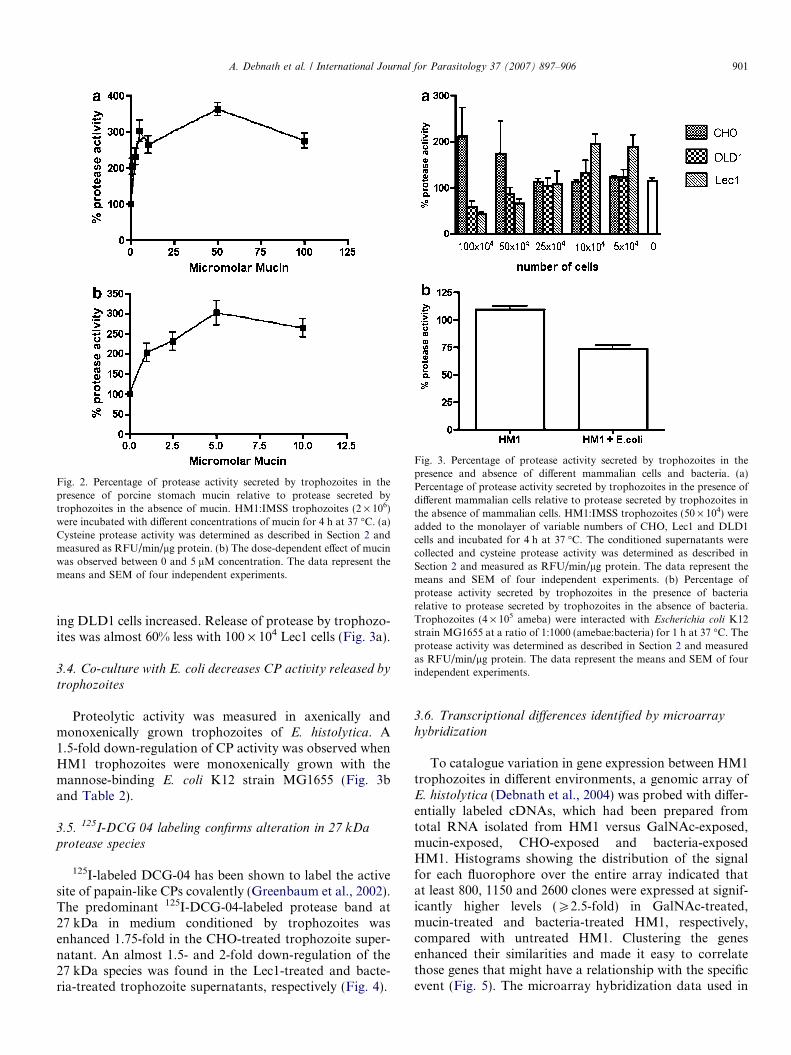
Fig. 2. Percentage of protease activity secreted by trophozoites in thepresence of porcine stomach mucin relative to protease secreted bytrophozoites in the absence of mucin. HM1:IMSS trophozoites (2 · 106)were incubated with different concentrations of mucin for 4 h at 37 �C. (a)Cysteine protease activity was determined as described in Section 2 andmeasured as RFU/min/lg protein. (b) The dose-dependent effect of mucinwas observed between 0 and 5 lM concentration. The data represent themeans and SEM of four independent experiments.
Fig. 3. Percentage of protease activity secreted by trophozoites in thepresence and absence of different mammalian cells and bacteria. (a)Percentage of protease activity secreted by trophozoites in the presence ofdifferent mammalian cells relative to protease secreted by trophozoites inthe absence of mammalian cells. HM1:IMSS trophozoites (50 · 104) wereadded to the monolayer of variable numbers of CHO, Lec1 and DLD1cells and incubated for 4 h at 37 �C. The conditioned supernatants werecollected and cysteine protease activity was determined as described inSection 2 and measured as RFU/min/lg protein. The data represent themeans and SEM of four independent experiments. (b) Percentage ofprotease activity secreted by trophozoites in the presence of bacteriarelative to protease secreted by trophozoites in the absence of bacteria.Trophozoites (4 · 105 ameba) were interacted with Escherichia coli K12strain MG1655 at a ratio of 1:1000 (amebae:bacteria) for 1 h at 37 �C. Theprotease activity was determined as described in Section 2 and measuredas RFU/min/lg protein. The data represent the means and SEM of fourindependent experiments.
A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906 901
ing DLD1 cells increased. Release of protease by trophozo-ites was almost 60% less with 100 · 104 Lec1 cells (Fig. 3a).
3.4. Co-culture with E. coli decreases CP activity released by
trophozoites
Proteolytic activity was measured in axenically andmonoxenically grown trophozoites of E. histolytica. A1.5-fold down-regulation of CP activity was observed whenHM1 trophozoites were monoxenically grown with themannose-binding E. coli K12 strain MG1655 (Fig. 3band Table 2).
3.5. 125I-DCG 04 labeling confirms alteration in 27 kDa
protease species
125I-labeled DCG-04 has been shown to label the activesite of papain-like CPs covalently (Greenbaum et al., 2002).The predominant 125I-DCG-04-labeled protease band at27 kDa in medium conditioned by trophozoites wasenhanced 1.75-fold in the CHO-treated trophozoite super-natant. An almost 1.5- and 2-fold down-regulation of the27 kDa species was found in the Lec1-treated and bacte-ria-treated trophozoite supernatants, respectively (Fig. 4).
3.6. Transcriptional differences identified by microarray
hybridization
To catalogue variation in gene expression between HM1trophozoites in different environments, a genomic array ofE. histolytica (Debnath et al., 2004) was probed with differ-entially labeled cDNAs, which had been prepared fromtotal RNA isolated from HM1 versus GalNAc-exposed,mucin-exposed, CHO-exposed and bacteria-exposedHM1. Histograms showing the distribution of the signalfor each fluorophore over the entire array indicated thatat least 800, 1150 and 2600 clones were expressed at signif-icantly higher levels (P2.5-fold) in GalNAc-treated,mucin-treated and bacteria-treated HM1, respectively,compared with untreated HM1. Clustering the genesenhanced their similarities and made it easy to correlatethose genes that might have a relationship with the specificevent (Fig. 5). The microarray hybridization data used in

Table 1Transcriptional changes in Entamoeba histolytica trophozoites duringinteraction with GalNAc, mucin, mammalian cell lines and bacteria
Treatment Fold change of the transcripts
Eh + GalNAc 1.5-fold up-regulation of 170-kDa Gal/GalNAc letin3-fold up-regulation of peroxiredoxin
Eh + Mucin 3-fold up-regulation of EhCP1 and EhCP2Eh + CHO 3-fold down-regulation of CP3Eh + Escherichia coli
MG16553-fold up-regulation of protein kinase
3.5-fold up-regulation of ABC transporter3-fold up-regulation of Rab family GTPase3-fold up-regulation of hsp 90
CHO, Chinese hamster ovary.
Table 2Changes in protease activity in Entamoeba histolytica trophozoites duringinteraction with GalNAc, mucin, mammalian cell lines and bacteria
Treatment Fold change in protease activity released bytreated E. histolytica compared with untreatedtrophozoites
Eh + GalNAc 3-fold down-regulationEh + Mucin 3.5-fold up-regulationEh + CHO 2-fold up-regulationEh + Lec1 CHO 2.5-fold down-regulationEh + DLD1 2-fold down-regulationEh + Escherichia coli
MG16551.5-fold down-regulation
CHO, Chinese hamster ovary.
Fig. 4. 125I-DCG-04 labeling of secreted protease from untreatedEntamoeba histolytica and CHO-, Lec1-, and bacteria-treated Entamoeba.Equal amounts of protein (as determined by protein standard curve, datanot shown) released by the treated and untreated trophozoites werelabeled with 125I-DCG-04 and separated on a 15% polyacrylamide gel.Lane 1, protease released by trophozoites incubated without epithelialcells; lane 2, protease released by trophozoites incubated with 100 · 104
CHO cells; lane 3, protease released by trophozoites incubated with100 · 104 Lec1 cells; lane 4, protease released by trophozoites incubatedwithout bacteria; lane 5, protease released by trophozoites incubated withbacteria. Densitometric quantification of band intensities was performedusing ImageQuant TL software (Amersham Biosciences) and the valuesare indicated below each lane on the gel. Values were obtained aftersubtracting the background value from each lane.
902 A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906
this report are available at http://www.ucsf.edu/mckerrow/array2.html. The sequencing data from clones that weresequenced because they were transcriptionally regulatedare available as supplementary data (Supplmentary Data).
The most abundant gene transcript found up-regulatedwith GalNAc interaction was the 170-kDa Gal/GalNAclectin (clone AEC 2837). Peroxiredoxin or thiol-specificantioxidant transcripts (clone AEC 2482) were expressed3-fold higher during GalNAc interaction.
The proteases, EhCP1 and EhCP2 (clone AEC 3223)were found to be up-regulated 3-fold during E. histolytica
interaction with porcine stomach mucin interaction whereasCP3 (clone AEC 1278) was found to be down-regulated 3-fold during its interaction with 100 · 104 CHO (Table 1).
Bacteria induced higher transcript levels for proteinkinase (clone AEC 5059), ABC transporter (AEC 1729),Rab family GTPase (clone AEC 5255) and hsp 90 (cloneAEC 5872) gene expression in E. histolytica. Protein kinaseexpression was almost 3-fold enhanced by exposure of tro-phozoites to bacteria. The ABC transporter, Rab familyGTPase and hsp 90 were increased 3.5-, 3-, and 3-fold,respectively.
3.7. Validation of microarray data by real-time PCR
Selected clones that were shown by the DNA microarrayto be differentially expressed were validated by real-time
PCR, affirming the reliability of the microarray (Fig. 6).Clone AEC 2837 was found to be up-regulated 1.5-foldin GalNAc treated trophozoites. Clone AEC 3223 encod-ing EhCP1 and EhCP2 was over expressed almost 1.6-foldin real-time PCR during E. histolytica interaction with por-cine stomach mucin whereas the same transcript remainedunchanged during bacteria interaction. Clone AEC 1278encoding CP3 was found to be down-regulated 3-foldduring CHO interaction and clone AEC 5872 encodingE. histolytica hsp 90 was enhanced 9-fold by exposure oftrophozoites to bacteria.
4. Discussion
In developing countries, colonization with E. histolyticahas been observed in 5% or more of children. Less than10% of colonized individuals develop colitis, with the restclearing the infection within months. It is in the intestinethat the determinants of invasive versus commensal infec-tion must come into play. One model proposes that deple-tion of mucins allows the parasite to interact with theepithelial cell surface and ingest exudated erythrocytes,initiating an invasion pathway (Petri et al., 2002).
Entamoeba histolytica trophozoites express numerouscell surface and secreted molecules that assist with feeding

Fig. 5. Clustered display of data from a portion of the entire microarray,indicating bacteria- and GalNAc-induced gene expression from fourdifferent experiments (EhCS1p1-110, EhCS1p1-111, EhCS1p1-112,EhCS1p1-113; EhCS1p1-079, EhCS1p1-080, EhCS1p1-081 andEhCS1p1-082). Data from related genes in four separate hybridizationsfrom four independent cDNA preparations, prepared from both bacteria-treated and GalNAc-treated trophozoites, were analyzed by use of suchdata mining tools as CLUSTER analysis and were visualized usingTREEVIEW. Increasingly positive log ratios due to bacteria and GalNAcactivation are colored red.
A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906 903
and retention in the intestine, and which also contribute tothe virulence of the parasite. Adhesion of trophozoites tocolonic mucins, host cells and bacteria occurs mainlythrough a galactose binding lectin (Frederick and Petri,2005).
CPs of E. histolytica are involved in several steps ofinvasion and tissue damage. The role of CPs in the patho-genesis of invasive amebiasis is not yet fully understoodand the factors responsible for inducing amebic CP arenot clear. Previous reports suggested that CP activity of
E. histolytica was dependent on Gal/GalNAc-mediatedcontact with target cells (Singh et al., 2004). Our resultsindicate that release of CP by trophozoites is inhibited byfree GalNAc. MUC2 is the major glycoprotein componentof the colonic mucous gel layer and GalNAc residues ofMUC2 mucin protect the colonic epithelium from amebicinvasion (Petri et al., 2002). Chadee et al. (1987) alsoshowed that free galactose abrogated binding of inhibitoryrat colonic mucins to the amebae. Decreased proteaseactivity during GalNAc interaction and over expressionof the Gal/GalNAc lectin gene are consistent with a pro-gram of commensal infection and mucus coat colonizationmediated by the Gal lectin.
In microarray hybridization, E. histolytica peroxiredox-in or thiol-specific antioxidant (TSA) was over-expressedalong with GalNAc lectin during GalNAc interaction. Thisis consistent with an earlier report of co-localization ofTSA and GalNAc lectin at sites of ameba:host cell contact(Hughes et al., 2003). Gilchrist et al. (2006) also found thelevels of thioredoxin transcripts were increased duringintestinal colonization and invasion by E. histolytica. Theinteraction of trophozoites with GalNAc rich mucin maystimulate the lectin–TSA interaction, and thus E. histolyti-
ca trophozoites protect themselves during tissue adherenceand invasion from oxidative attack by activated hostinflammatory cells.
While GalNAc, a component of MUC2, down-regu-lated protease release, interaction of porcine stomachmucin with E. histolytica trophozoites enhanced cysteineprotease secretion into the media. Moncada et al. (2003)reported the ability of EhCPs to degrade colonic mucinbut could not identify a specific CP involved in mucin deg-radation. In the present study, transcripts for EhCP1 andEhCP2 were up-regulated during E. histolytica and porcinestomach mucin interaction. The enhanced secretion of CPinto the media during interaction with porcine stomachmucin suggests that the specific composition of mucinsmay be important in influencing the Entamoeba phenotypeas was proposed by Chadee et al. (1991). In a recent study,Lidell et al. (2006) also indicated the role of EhCP1 andEhCP2 for the cleavage of MUC2.
Adherence of ameba to target cells is presumed to be anecessary first step in production of the cytopathic effect(Martinez-Palomo et al., 1985). As contact with target cellsis achieved, the concentration of secreted CP in the regionof the target cell would be expected to increase substan-tially. Furthermore, the surface of E. histolytica trophozo-ites has many thiol groups, which could provide a suitablereducing environment for the proteinase, especially as thespace between ameba and the target cell becomes negligible(Keene et al., 1990). Indeed, trophozoites interacting withCHO cells released more CP activity compared with non-interacting trophozoites. However, as was the case withmucins, different epithelial cell types elicited differentameba phenotypes. Ameba released less CP when the samenumber of Lec1 CHO mutants was used. This mutant lacksGlcNAc glycosyl transferase so that N-linked

Fig. 6. Confirmation of the microarray gene expression seen in GalNAc-, mucin-, CHO- and bacteria-exposed HM1 by real-time PCR analysis.Quantitative values for real-time PCR were obtained from the threshold cycle number at which the increase in the signal associated with an exponentialgrowth of PCR products was first detected. Final results were expressed as N-fold differences in target gene expression relative to the reference geneEntamoeba histolytica actin. The clones selected for confirmation by real-time PCR analysis are AEC 2837, AEC 3223, AEC 1278 and AEC 5872.
904 A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906
carbohydrates are blocked at the intermediate Man5-Glc-NAc2-Asn. Ravdin et al. (1989) found reduced lectin-med-iated amebic adherence when the Lec1 mutant was used.Therefore, the CHO versus Lec1 results are consistent witha model of lectin-mediated adherence leading to an invasivephenotype. We recognized the marked increase in proteasesecretion at a lower number of Lec1 cells as a paradox. Thisresult raises the possibility that there are two competing sig-nals during this interaction. While a significant decrease inprotease activity in the presence of high numbers of Lec1cells is clear from the data, at lower cell density proteaseactivity was induced. The exact mechanism of this latterphenomenon is not clear. MUC2, the major secreted mucinin human intestine, is strongly expressed in the mucinouscarcinomas but not in CHO cells. Ookawa et al. (2002)showed that MUC2 expression is transcriptionally regu-lated by p53 protein in DLD1. The down-regulation ofCP activity during DLD1 interaction might therefore bedue to the presence of secretory mucin, having a complexcarbohydrate structure rich in Gal and GalNAc.
The bacterial flora of the intestine has been proposed toplay a role in determining the virulence of E. histolytica
(Bracha and Mirelman, 1983; Mirelman, 1987). Strains ofE. coli that utilize mucus as a source of carbon and nitro-gen can also bind mucus proteins and are more effectivein colonizing the intestine (Cohen et al., 1983). Entamoeba
histolytica trophozoites attach and ingest bacteria withtheir mannose-containing cell surface components servingas receptors for the mannose-binding lectins of bacteria(Mirelman, 1988). A comparison between the CP activityof axenically grown trophozoites of strain HM1:IMSSand of the same strain grown with the mannose-bindingE. coli K12 strain MG1655 showed less CP activity releasedby the bacteria-associated amebae. Transcriptional profil-ing revealed bacteria induced protein kinase, ABC trans-porter, Rab family GTPase and hsp 90 gene expression inE. histolytica. Protein kinases play a crucial role in signaltransduction for a variety of cellular responses includingcell proliferation, differentiation and phagocytosis (Nish-izuka, 1986). Rab family GTPases have been shown to playa role in polarized vesicle trafficking, regulation of the actincytoskeleton and adhesion (Zahraoui et al., 1994; Imamuraet al., 1998). Vega and De Maio (2005) recently showedthat the Hsp90 family of proteins plays a role in modula-tion of the phagocytic process. The over-expression ofgenes encoding these proteins suggests enhanced phagocy-tosis of E. histolytica during interaction with bacteria with-out leakage of CP activity into the environment. Theincreased expressions of hsp 90, protein kinase and Rabfamily GTPase are consistent with previous findings byGilchrist et al. (2006) where they showed adaptation tothe intestinal environment was accompanied by over-

Mucincoat
Epithelialcells
PhagocytosedbacteriaCysteine
protease
Protein KinaseABC transporterRab GTPasehsp90
Transcripts for:
Gal/GalNAc lectinThiol-specific antioxidantEhCP1 and EhCP2
Transcripts for:
CHOCell lineDLDCell lineLecCell line
CP
Bacteria
with
CP with
CP with
(CP)
CP
Fig. 7. A diagrammatic summary of the key transcriptional changes andinduction of protease secretion in response to interaction of Entamoeba
histolytica trophozoites with components of the intestinalmicroenvironment.
A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906 905
expression of stress-related transcripts and also increases ina subset of cell signaling genes including kinases and smallGTPases.
Finally, the discordance of CP activity directly measuredby a fluorescent substrate and transcriptional levels of CPsis consistent with a model whereby trophozoites containprestored CPs in vesicles that can be released rapidly uponinteraction of amebae with appropriate host inducers. Insome, but not all, cases this release of CP activity is paral-leled by CP transcript. In the remaining cases post-tran-scriptional regulation is likely either at the level oftranslation or by release of protease from pre-existingpools. Fig. 7 summarizes the results of interaction of E. his-
tolytica with mucin, epithelial cells and bacteria as well asthe roles played by lectins and CP.
Acknowledgements
This work was presented in part at the XV Seminar onAmebiasis, Oaxaca, Mexico, January 31 to February 3,
2006. This work was supported by the Sandler FamilySupporting Foundation and NIAID TDRU AI-35707.We thank Maha Abdulla for assisting in real-time PCRdata analysis; Christopher Franklin for the photographicwork.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.ijpara.2007.01.016.
References
Allen, A., Bell, A., Mantel, M., Pearson, J.P., 1982. The structureand physiology of gastrointestinal mucus. Adv. Exp. Med. Biol. 144,115–133.
Belley, A., Keller, K., Goettke, M., Chadee, K., 1999. Intestinal mucins incolonization and host defense against pathogens. Am. J. Trop. Med.Hyg. 60, 10–15.
Bracha, R., Mirelman, D., 1983. Adherence and ingestion of Escherichia
coli serotype 055 by trophozoites of Entamoeba histolytica. Infect.Immun. 40, 882–887.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254.
Chadee, K., Petri, W.A. Jr., Innes, D.J., Ravdin, J.I., 1987. Rat andhuman colonic mucins bind to and inhibit adherence lectin ofEntamoeba histolytica. J. Clin. Invest. 80, 1245–1254.
Chadee, K., Keller, K., Forstner, J., Innes, D.J., Ravdin, J.I., 1991. Mucinand nonmucin secretagogue activity of Entamoeba histolytica andcholera toxin in rat colon. Gastroenterology 100, 986–997.
Cohen, P.S., Rossoll, R., Cabelli, V.J., Yang, S.L., Lanx, D.C., 1983.Relationship between the mouse colonizing ability of a human fecalEscherichia coli strain and the ability to bind a specific mouse colonicmucous gel protein. Infect. Immun. 40, 62–69.
Connaris, S., Greenwell, P., 1997. Glycosidases in mucin-dwellingprotozoans. Glycoconj. J. 14, 879–882.
Debnath, A., Das, P., Sajid, M., McKerrow, J.H., 2004. Identification ofgenomic responses to collagen binding by trophozoites of Entamoeba
histolytica. J. Infect. Dis. 190, 448–457.Diamond, L.S., 1984. Lumen dwelling protozoa: Entamoeba, Trichomo-
nads, and Giardia. In: Jensen, J.B. (Ed.), In vitro Cultivation ofProtozoan Parasites. CRC Press, Florida, pp. 65–109.
Eichinger, D., 2001a. A role for a galactose lectin and its ligands duringencystment of Entamoeba. J. Eukayot. Microbiol. 48, 17–21.
Eichinger, D., 2001b. Encystation in parasitic protozoa. Curr. Opin.Microbiol. 4, 421–426.
Frederick, J.R., Petri, W.A. Jr., 2005. Roles for the galactose/N-acetylgalatosamine binding lectin of Entamoeba in parasite virulenceand differentiation. Glycobiology 15, 53R–59R.
Gilchrist, C.A., Houpt, E., Trapaidze, N., Fei, Z., Crasta, O., Asgharpour,A., Evans, C., Martino-Catt, S., Baba, D.J., Stroup, S., Hamano, S.,Ehrenkaufer, G., Okada, M., Singh, U., Nozaki, T., Mann, B.J., PetriJr., W.A., 2006. Impact of intestinal colonization and invasion on theEntamoeba histolytica transcriptome. Mol. Biochem. Parasitol. 147,163–176.
Greenbaum, D.C., Baruch, A., Grainger, M., Bozdech, Z., Med-zihradszky, K.F., Engel, J., DeRisi, J., Holder, A.A., Bogyo, M.,2002. A role for the protease falcipain 1 in host cell invasion by thehuman malaria parasite. Science 298, 2002–2006.
Hughes, M.A., Lee, C.W., Holm, C.F., Ghosh, S., Mills, A., Lockhart,L.A., Reed, S.L., Mann, B.J., 2003. Identification of Entamoeba
histolytica thiol-specific antioxidant as a GalNAc lectin-associatedprotein. Mol. Biochem. Parasitol. 127, 113–120.

906 A. Debnath et al. / International Journal for Parasitology 37 (2007) 897–906
Imamura, H., Takaishi, K., Nakano, K., Kodama, A., Oishi, H., Shiozaki,H., Monden, M., Sasaki, T., Takai, Y., 1998. Rho and Rab small Gproteins coordinately reorganize stress fibers and focal adhesions inMDCK cells. Mol. Biol. Cell 9, 2561–2575.
Kaushik, N., Fear, D., Richards, S.C.M., McDermott, C.R., Nuwaysir,E.F., Kellam, P., Harrison, T.J., Wilkinson, R.J., Tyrrell, D.A.J.,Holgate, S.T., Kerr, J.R., 2005. Gene expression in peripheral bloodmononuclear cells from patients with chronic fatigue syndrome. J.Clin. Pathol. 58, 826–832.
Keene, W.E., Pettit, M.G., Allen, S., McKerrow, J.H., 1986. Themajor neutral proteinase of Entamoeba histolytica. J. Exp. Med. 163,536–549.
Keene, W.E., Hidalgo, M.E., Orozco, E., McKerrow, J.H., 1990.Entamoeba histolytica: correlation of the cytopathic effect of virulenttrophozoites with secretion of a cysteine proteinase. Exp. Parasitol. 71,199–206.
Lidell, M.E., Moncada, D.M., Chadee, K., Hansson, G.C., 2006.Entamoeba histolytica cysteine proteases cleave the MUC2 mucin inits C-terminal domain and dissolve the protective colonic mucus gel.Proc. Natl. Acad. Sci. USA 103, 9298–9303.
Martinez-Palomo, A., Gonzalez-Robles, A., Chavez, B., Orozco,E., Fernandez-Castelo, S., Cervantes, A., 1985. Structural basesof the cytolytic mechanisms of Entamoeba histolytica. J.Protozool. 32, 166–175.
Mirelman, D., 1987. Ameba-bacterium relationship in amebiasis. Micro-biol. Rev. 51, 272–284.
Mirelman, D., 1988. Ameba-bacterial relationship in amebiasis. In:Ravdin, J.I. (Ed.), Amebiasis. Human Infection by Entamoeba
histolytica. Wiley, New York, pp. 351–369.Moncada, D., Keller, K., Chadee, K., 2003. Entamoeba histolytica cysteine
proteinases disrupt the polymeric structure of colonic mucin and alterits protective function. Infect. Immun. 71, 838–844.
Nishizuka, Y., 1986. Studies and perspectives of protein kinase C. Science233, 305–312.
Ookawa, K., Kudo, T., Aizawa, S., Saito, H., Tsuchida, S., 2002.Transcriptional activation of the MUC2 gene by p53. J. Biol. Chem.277, 48270–48275.
Padilla-Vaca, F., Ankri, S., Bracha, R., Koole, L.A., Mirelman, D., 1999.Down regulation of Entamoeba histolytica virulence by monoxenic
cultivation with Escherichia coli O55 is related to a decrease inexpression of the light (35-Kilodalton) subunit of the Gal/GalNAclectin. Infect. Immun. 67, 2096–2102.
Petri Jr., W.A., Haque, R., Mann, B.J., 2002. The bittersweet interface ofparasite and host:lectin–carbohydrate interactions during humaninvasion by the parasite Entamoeba histolytica. Annu. Rev. Microbiol.56, 39–64.
Prathap, K., Gilman, R., 1970. The histopathology of acute intestinalamebiasis. A rectal biopsy study. Am. J. Pathol. 60, 229–245.
Ravdin, J.I., Stanley, P., Murphy, C.F., Petri Jr., W.A., 1989. Charac-terization of cell surface carbohydrate receptors for Entamoeba
histolytica adherence lectin. Infect. Immun. 57, 2179–2186.Reed, S.L., Keene, W.E., McKerrow, J.H., 1989. Thiol proteinase
expression and pathogenicity of Entamoeba histolytica. J. Clin.Microbiol. 27, 2772–2777.
Schulte, W., Scholze, H., 1989. Action of the major protease fromEntamoeba histolytica on proteins of the extracellular matrix. J.Protozool. 36, 538–543.
Shazand, K., Baban, S., Prive, C., Malette, B., Croteau, P., Lagace, M.,Racine, J.-B., Hugo, P., 2004. FOXO1 and c-jun transcriptionfactors mRNA are modulated in endometrosis. Mol. Hum. Reprod.10, 871–877.
Singh, D., Naik, S.R., Naik, S., 2004. Contact of Entamoeba histolytica
with baby hamster kidney-21 (BHK-21) cell line on cysteine proteinaseactivity. Indian J. Med. Res. 119, 157–161.
Tran, V.Q., Herdman, D.S., Torian, B.E., Reed, S.L., 1998. The neutralcysteine proteinase of Entamoeba histolytica degrades IgG andprevents its binding. J. Infect. Dis. 177, 508–511.
Vega, V.L., De Maio, A., 2005. Increase in phagocytosis after geldana-mycin treatment or heat shock: role of heat shock proteins. J.Immunol. 175, 5280–5287.
World Health Organization, 1998. The World Health Report 1998: Life inthe 21st Century: A Vision for All. World Health Organization,Geneva.
Zahraoui, A., Joberty, G., Arpin, M., Fontaine, J.J., Hellio, R., Tavitian,A., Louvard, D., 1994. A small rab GTPase is distributed incytoplasmic vesicles in nonpolarized cells but colocalizes with thetight junction marker ZO-1 in polarized epithelial cells. J. Cell Biol.124, 101–115.