towards antigen-specific apheresis of pathogenic autoantibodies as a further step in the treatment...
TRANSCRIPT

Journal of Neuroimmunology 201–202 (2008) 95–103www.elsevier.com/locate/jneuroim
Review article
Towards antigen-specific apheresis of pathogenic autoantibodies as a furtherstep in the treatment of myasthenia gravis by plasmapheresis
Paraskevi Zisimopoulou a, George Lagoumintzis b, Kalliopi Kostelidou a, Kalliopi Bitzopoulou a,b,Gregory Kordas b, Nikolaos Trakas a, Konstantinos Poulas b, Socrates J. Tzartos a,b,⁎
a Department of Biochemistry, Hellenic Pasteur Institute, 127, V. Sofias Ave., GR11521, Athens, Greeceb Department of Pharmacy, University of Patras, Rio, GR26500, Patras, Greece
Received 1 April 2008; received in revised form 6 June 2008; accepted 6 June 2008
Abstract
Myasthenia gravis (MG), a prototypic antibody-mediated autoimmune disease, presents an excellent target for scientific research aimed at abetter understanding of the disease itself and the source that triggers an autoimmune reaction in an organism. MG is a neuromuscular diseasecaused mainly by an autoimmune response against the nicotinic acetylcholine receptor (AChR) which interferes with neuromuscular transmission.This review focuses on our studies on the extracellular domains of human muscle AChR subunits in an effort to develop an approach for thespecific therapeutic apheresis of autoantibodies from patients' sera using the immobilized subunits as immunoadsorbents. The ability of the anti-AChR antibodies isolated by this technique, but not of the depleted sera, to induce disease is also described.
This review is dedicated to the late Prof. John Newsom-Davis, who was the first to introduce the use of plasmapheresis for MG.© 2008 Elsevier B.V. All rights reserved.
Keywords: Acetylcholine receptor autoantibodies; Extracellular domains; Immunoadsorption; Myasthenia gravis; Therapy; Plasmapheresis
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 952. Expression of muscle-type human AChR ECDs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 963. Use of recombinant ECDs as immunoadsorbents for the clearance of MG sera . . . . . . . . . . . . . . . . . . . . . . . . . . . . 984. Characterization of the ECD immunoadsorbents . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 985. Activity of antibody-depleted sera and of purified anti-subunit autoantibodies in vitro and in vivo . . . . . . . . . . . . . . . . . 1006. Conclusions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102
Abbreviations: ACh, acetylcholine; AChR, nicotinic acetylcholine receptor;MG, myasthenia gravis; ECD, extracellular domain; RIA, radioimmunoassay;IVIg, intravenous immunoglobulin; EAMG, experimental autoimmunemyasthenia gravis.⁎ Corresponding author. Department of Biochemistry, Hellenic Pasteur Institute,
127, V. Sofias Ave., GR11521, Athens, Greece. Tel.: +30 2106478844; fax: +302106478842.
E-mail addresses: [email protected], [email protected] (S.J. Tzartos).
0165-5728/$ - see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.jneuroim.2008.06.020
1. Introduction
One of the important contributions of the late Prof. JohnNewsom-Davis to the treatment of myasthenia gravis (MG) wasthe introduction of the use of plasmapheresis (Pinching et al.,1976). In this paper, dedicated to JND, we review some of ourrecent efforts towards the development of an advanced apheresis

96 P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
approach, namely antigen-specific autoantibody apheresis.This method involves the use of immunoadsorption columnsof recombinant extracellular domains (ECDs) of all the sub-units of the human muscle nicotinic acetylcholine receptor(AChR) for the selective removal of anti-AChR antibodies frompatients' sera.
The neuromuscular junction (NMJ) is a specialized synapsedesigned to transmit nerve impulses to the muscle via achemical transmitter, acetylcholine (ACh), which is stored insynaptic vesicles at the nerve terminal. Processes that target theNMJ can result in clinical syndromes characterized by muscleweakness. Among these disorders, the most common and beststudied is autoimmune MG, an acquired disease in which theAChR is targeted in the majority of affected patients.
The AChR is an acetylcholine-gated cation channel with akey role in communication between motor nerves and skeletalmuscles, and in the peripheral and central nervous system (Berget al., 2000; Lindstrom, 2000). At the normal NMJ, the AChR ishighly concentrated on the terminal expansions of thepostsynaptic junctional folds. Its principal function is to permitthe flow of current through its ion channel after binding ACh. Inthe central nervous system, AChRs also act presynaptically andextrasynaptically to modulate transmission by facilitating therelease of many transmitters (Kaiser et al., 2000; Zoli, 2000).Additionally, they are present in the skin (Grando and Horton,1997), bronchial and vascular epithelia (Macklin et al., 1998;Maus et al., 1998), and other non-neuronal tissues (Sekhon et al.,1999; Wang et al., 2003; Skok et al., 2005), where they are alsoinvolved in intercellular communication.
The AChR and other proteins located on the cell membrane ofthe motor nerve or muscle endplate with an ECD accessible tocirculating substances are targets for autoimmune attack.Following attack, the disruption of communication in the NMJresults in muscle weakness, the best characterized conditionsbeing the autoimmune myasthenic disorders. Molecules in theNMJ identified as targets include calcium and potassium channelsin the motor nerve terminal, AChR and muscle specific kinase(MuSK) postsynaptically, and rapsyn, titin, and the ryanodinereceptor intracellularly in the muscle cell (Agius et al., 2003).
Antibodies to AChRs, which result in the clinical picture ofacquired MG, were the first demonstrated to be pathogenic. Thisrole ofAChRantibodies has been established by severalmeasures.These include the demonstration of the presence of circulatingAChR antibodies in nearly 90% of patients with MG (Lindstromet al., 1976), passive transfer of several features of the disease fromhuman to mouse with IgG (Lang et al., 1983; Mossman et al.,1988; Shillito et al., 1995), localization of immune complexes(IgG and complement) on the postsynaptic membrane (Engelet al., 1982), the beneficial effects of plasmapheresis (Pinchinget al., 1976; Dau et al., 1977), and the induction of experimentalMG by immunization with AChR (Patrick and Lindstrom, 1973)or by injection of anti-AChR antibodies (Tzartos et al., 1987). Inaddition, a more direct proof of pathogenicity of MG patients'antibodies comes from the recent observation that experimentalMG is readily induced in animals, injected i.p. with isolated anti-AChR antibodies fromMG patients (Tzartos et al., 2008; Kordas,Sideris, Poulas and Tzartos, unpublished data).
Initial treatment for MG includes acetylcholinesterase inhibi-tors, which retard the hydrolysis of acetylcholine at the NMJ, andcorticosteroids, which affect the immune system, leading to generalimmunosuppression (Conti-Fine and Kaminski, 2001; Kaminski,2001; Dalakas, 2002). Other commonly used immunomodulatingdrugs primarily interfere with T and B cell proliferation and/orfunction. Some of the major adverse effects linked to immuno-suppression are osteoporosis, hypertension, gastrointestinal dis-comfort, and psychiatric changes (Kaminski, 2003; Fostieri et al.,2006).
A different therapeutic approach, plasma exchange (plasma-pheresis), is used mostly in severe MG symptoms or inmyasthenic crisis and also in cases where muscle function mustbe optimized, such as before surgery (including thymectomy). Itproduces a rapid clinical improvement regardless of whether thepatients are seronegative or seropositive for anti-AChR antibodies(Chiu et al., 2000). This suggests that the removal of circulatingpathogenic antibodies and possibly other pathogenic agents leadsto the clinical improvement. Administration of intravenous Ig(IVIg) is also used in similar circumstances to plasma exchange(Gajdos et al., 1987; Ferrero et al., 1993). IVIg administration hasbeen commonly linked to adverse effects, which, however, areonly minor (Brannagan et al., 1996; Kazatchkine and Kaveri,2001; Dalakas, 2002). IVIg seems to impact on the autoimmuneprocess by several mechanisms including interfering with thebinding of antibodies to Fc receptors on macrophages (Samuels-son et al., 2001). Nevertheless, as with the other therapies forMG,neither IVIg nor plasma exchange are specific for MG.
An attractive MG-specific therapeutic approach proposedhere is the selective antigen-specific depletion of autoantibodies(immunoadsorption) and the return of the “cleared” plasma tothe patient. This might be achieved through the use ofimmunoadsorbent columns carrying immobilized humanAChR (whole or part), which would necessitate the productionand use of large amounts of recombinant AChR or AChRdomains. The immobilization of such recombinant proteins onSepharose beads and the incubation of these matrices with avariety of seropositive MG sera results in a significant reductionin the concentration of autoantibodies in these sera (Psaridi-Linardaki et al., 2005; Kostelidou et al., 2007). We thereforepropose the therapeutic selective depletion of autoantibodiesusing this technique.
2. Expression of muscle-type human AChR ECDs
In contrast to the many subtypes of neuronal AChRs, there areonly two subtypes of muscle AChRs, a fetal subtype with an (α1)2β1γδ stoichiometry and an adult subtype with an (α1)2β1εδstoichiometry. Each AChR subunit consists of an N-terminalECD (containing the characteristic 13-residue long Cys-loop of aligand-gated ion channel), four membrane-spanning α-helices, alarge cytoplasmic loop, and a small extracellular C-terminal tail(Karlin, 2002).
The development of a specific immunoadsorption approachfor the selective depletion of pathogenic autoantibodies fromMGsera requires large amounts of recombinant AChR, a challengingtask, as the AChR is a membrane protein. However, the use of the

Fig. 1. Schematic representation of A) the consensus AChR subunit, B) the constructs used for the expression of the ECDs and C) a representative mutated ECD. SP:signal peptide; M1–4: transmembrane domains; αFactor: signal peptide from yeast; flag, myc, 6xHis: the tags expressed with the polypeptides.
Fig. 2. Schematic representation of the work on the AChR ECDs in ourlaboratory and summarized in this review.
97P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
ECD of each of the AChR subunits should overcome thisproblem, especially as these ECDs carry the full range of epitopesrecognized by the pathogenic autoantibodies.
We therefore expressed and purified the ECD of each humanmuscle AChR subunit (Fig. 1). The ECDs of the human α1(amino acids 1–210),β1 (amino acids 1–221), γ (amino acids 1–218), δ (amino acids 1–219), and ε (amino acids 1–219) AChRsubunits, carrying a His-tag at their C-terminus, were expressed inP. pastoris, a yeast expression system, and purified under non-denaturing conditions (Psaridi-Linardaki et al., 2002; Kostelidouet al., 2006). These ECDs were then used as immunoadsorbentsfor the specific depletion of antibodies from seropositiveMG sera(see next chapter) and for structural studies (Fig. 2).
When the size of each protein in solution was estimated by gelfiltration analysis, the α1 ECD was found to be mainly in themonomeric form, the β1 ECD had an apparent molecular weightof 69 kDa corresponding to a dimeric form, the γ and ε ECDswere mainly oligomers, while the δ ECD eluted in the form ofmicroaggregates. The α1 ECD bound conformation-dependentmonoclonal antibodies, the competitive antagonist α-bungarotoxin with a satisfactory Kd value (∼5.1 nM); andwith low affinity, other cholinergic ligands, such as gallamineand d-tubocurarine (Psaridi-Linardaki et al., 2002), implyingcorrect protein folding. Both the β1 and γ ECDs also appearedto fold in a near-native conformation, as they were able torecognize and bind conformation-dependent, subunit-specificmonoclonal antibodies (Kostelidou et al., 2006).
In order to improve the solubility, and possibly the yield, ofthe expressed proteins, we replaced the Cys-loop in all wild-type ECDs with the cognate, more hydrophilic loop region ofthe Lymnea stagnalis acetylcholine binding protein (AChBP).This resulted in increased yield and solubility, although, in thecase of the δ ECD, a high proportion of microaggregates wasstill seen. Furthermore, mutation of Cys106 and Cys115 to Serin the Cys-loop mutant γ ECD resulted in a further improve-ment in solubility and expression yield compared to either the
wild-type γ ECD or the Cys-loop γ mutant alone (Bitzopoulouet al., submitted for publication).
The purified α1,β1, γ, and ε ECDs have been used in far- andnear-UV CD studies in an attempt to further probe their structure(Zouridakis et al., 2007). The results demonstrated that the majorstructural feature of the humanAChRECDswas a high content ofβ-sheet structure, with a small α-helix-content. These results arein very good agreement with the solved structures for thehomologous AChBPs from various mollusk species (Brejc et al.,

98 P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
2001) and for the ECDof the AChR from the electric ray, Torpedo(Unwin, 2005). Taken together, these findings strongly suggestthat the expressed human AChR ECDs may well be anappropriate starting material for more detailed structural studies.
Due to the relatively moderate ECD yield in the yeastexpression system (usually around 1 mg/l), we explored thepossibility of usingE. coli-expressed ECDs as immunoadsorbents(Zisimopoulou et al., in press). The proteins were expressed ininclusion bodies and purified under denaturing conditions. Asexpected, the yield for all ECDs was significantly higher than thatof the corresponding yeast product, namely around 12 mg/l.
3. Use of recombinant ECDs as immunoadsorbents for theclearance of MG sera
A number of techniques have been devised as alternatives toplasmapheresis. These include (a) adsorption of immunoglobu-lins using a tryptophan-linked polyvinyl alcohol gel (TR-350) thatadsorbs most large proteins, including most anti-AChR anti-bodies, by hydrophobic interactions (Yamazaki et al., 1982), (b)columns carrying staphylococcal protein A, which avidly bindsmost human immunoglobulins (Matic et al., 2001), and (c)columns carrying sheep anti-human IgG antibodies (Ig-Adsopak)to remove human IgGmolecules (Ptak, 2004); themain drawbackof these approaches is their non-specificity, namely theindiscriminate removal of all, or most, immunoglobulins,including potentially useful antibodies. To this end, threealternative approaches to the development of an antigen-specificapproach for the selective depletion of anti-AChR autoanti-bodies from MG patients' sera have been described. The firsttwo consisted of the use of columns bearing (a) a peptidecorresponding to amino acids 183–200 of the Torpedo AChR αsubunit (Takamori and Maruta, 2001) or (b) the ECD of theTorpedoα-subunit expressed inE. coli as a fusion protein with themaltose binding protein immobilized on amylase-resin; thisimmunoadsorbent was successfully tested on only two MG sera(Guo et al., 2005). However, the majority of anti-AChRantibodies are directed against several epitopes other than thatused by Takamori and Maruta, whereas, in the vast majority ofMG patients, few autoantibodies bind to Torpedo AChR(Lindstrom et al., 1978; Loutrari et al., 1992). These factsnecessitated the development of a more effective approach forselective serum clearance. The third approach, successfullyapplied by our own group, was the depletion of autoantibodiesfrom anti-AChR antibody-positive MG sera using recombinantECDs of human muscle AChR subunits immobilized on CNBr-activated Sepharose beads as immunoadsorbents (Psaridi-Linardaki et al., 2003, 2005; Kostelidou et al., 2007).
The average degree of immunoadsorption obtained for 41randomly chosen MG sera using E. coli-produced ECDs was24.1% for anti-α1 antibodies, 12.9% for anti-β1 antibodies,15.8% for anti-γ antibodies, 7.8% for anti-δ antibodies, and13.5% for anti-ε antibodies, whereas the corresponding resultsfor the yeast P. pastoris ECDs “matrix” were 31%, 13.9%, 7%,5.1%, and 12.7% (Zisimopoulou et al., in press). A different setof 50 randomly chosen MG sera tested only with yeast-ECDs(Fig. 3) showed higher levels of immunoadsorption, namely
35% for anti-α1 antibodies, 22% for anti-β1 antibodies, 20%for anti-γ antibodies, and 15.5% for anti-ε antibodies (Psaridi-Linardaki et al., 2005; Kostelidou et al., 2007). The unexpectedability of the denatured E. coli-expressed ECDs to efficientlyimmunoadsorb MG autoantibodies (often with a similarefficiency to that of the yeast-expressed folded ECDs) suggeststhat the initially unfolded E. coli-expressed ECDs acquire thecorrect conformation after immobilization on the resin andremoval of the denaturing reagent.
Use of the δ ECD did not result in considerable immunoad-sorption of autoantibodies even when a set of 100 additional MGsera were tested. Two possible reasons for the lack of autoanti-body depletion are that anti-δ autoantibodies are strictlyconformation-dependent and were not be removed due to animperfect conformation of the recombinant δ ECD (E. coli- oryeast-expressed) or that anti-δ antibodies are rare in MG.
The availability of the ECDs of all AChR subunits hasallowed the construction of immunoadsorption columns withimproved efficiency for removing MG autoantibodies. Our datashow that the combination of more than one ECD on a singleimmunoadsorbent leads to increased depletion of anti-AChRantibodies and, in some cases, to near-complete clearance(Fig. 4). In addition, further improvement was observed when theabove-mentioned γ ECD mutants were used for immunoadsorp-tion. Both the γCysLoop mutant and the γCysLoop-C106S-C115S mutant acted as more efficient immunoadsorbents thanthe wild-type γ1-218, while the γC61S and γC106S-C115Smutants lacking the transferred Cys-loop from the AChBP didnot display significantly improved adsorption (Bitzopoulouet al., submitted for publication). Overall it was found that allthe γ ECD mutants retain the characteristics of the wild-typeγ ECD, but those containing the AChBP loop have animproved expression yield, solubility, and antigenicity, andcould thus serve as more suitable immunoadsorbents for MGtreatment.
4. Characterization of the ECD immunoadsorbents
The potential use of the immobilized ECDs as immunoad-sorbents would require them to have a set of additionalcharacteristics, such as (a) a considerable capacity, (b) stabilityto long-term incubation with human serum and (c) specificityfor the anti-AChR antibodies. These characteristics aredescribed below, supporting the suitability of the immobilizedECDs as immunoadsorbents for MG therapy.
The maximal amount of anti-AChR antibodies by α1 ECD-Sepharose removed was about 1.5 pmol of antibody permicrogram of immobilized protein (Psaridi-Linardaki et al.,2005), while the β1 ECD-Sepharose removed 5 pmol (Koste-lidou et al., 2007). These results suggest that the plasma from anadult patient (approximately 3 l) with an average anti-AChRtiter of 5–10 nM would be cleared of anti-β1 antibodies by asmall column bearing 3–6 mg of yeast-expressed β1 ECD.Although such amounts are currently easily produced the futureuse of fermentors and the establishment of optimal cultureconditions could multiply the ECD yield and allow thetreatment of patients with almost any anti-AChR titer.
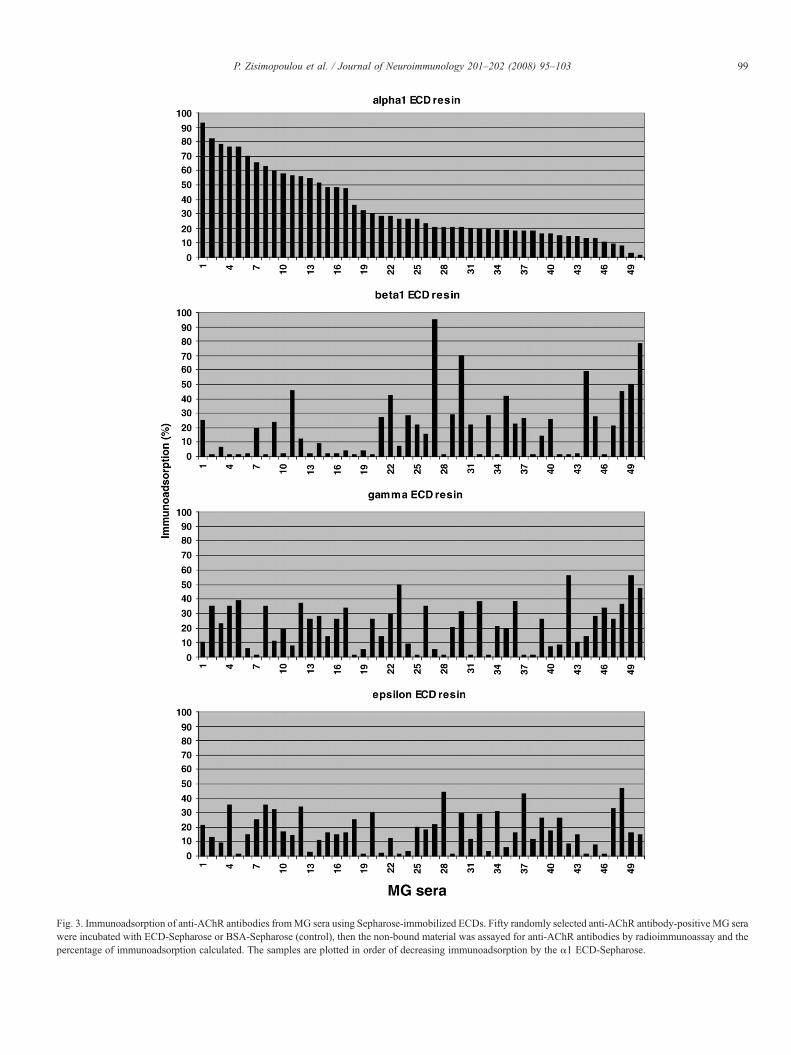
Fig. 3. Immunoadsorption of anti-AChR antibodies fromMG sera using Sepharose-immobilized ECDs. Fifty randomly selected anti-AChR antibody-positive MG serawere incubated with ECD-Sepharose or BSA-Sepharose (control), then the non-bound material was assayed for anti-AChR antibodies by radioimmunoassay and thepercentage of immunoadsorption calculated. The samples are plotted in order of decreasing immunoadsorption by the α1 ECD-Sepharose.
99P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103

Fig. 4. Immunoadsorption of anti-AChR antibodies from MG sera using columns containing more than one ECD expressed in yeast. Combined use of the α1, β1, γ,and ε ECDs. Ten selected anti-AChR antibody-positive MG sera were incubated with α1 ECD-Sepharose (gray bars), β1 ECD-Sepharose (chequered bars), γ ECD-Sepharose (white bars), ε ECD-Sepharose (dashed bars), and Sepharose bearing all four ECDs (black bars), with BSA/Sepharose as the control.
100 P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
Stability of the column is a prerequisite, since any polypeptidereleased from the column during immunoadsorption might act asan immunogen, despite the fact that the ECDs are of human originand so would be only weakly immunogenic. However, toeliminate this possibility, we showed that no detectable amountsof ECD (yeast- or E. Coli-expressed) were removed from theresin even after 24 h incubation with human serum, thus ensuringthat no potentially antigenic ECDmolecules would be introducedinto the patient during the brief therapeutic apheresis procedure(Psaridi-Linardaki et al., 2005; Zisimopoulou et al., in press). Tofurther verify the safety of the immunoadsorption process, wehave performed preliminary experiments in experimental rabbits.Two rabbits were used, one not immunized and one immunizedtwice with the β1 ECD. The immunized rabbit developed anti-ECD and anti-AChR antibodies, although no disease symptomsappeared. Antibody apheresis was performed one month after thesecond injection when the antibody titer had decreased. About20 ml of blood were collected from each rabbit and the plasmaisolated by centrifugation, passed through a 10 ml Sepharoseimmunoadsorbent containing 1 mg/ml of immobilized β1 ECD,and the treated plasma reintroduced i.v. into the correspondingrabbit. After 8 days, blood samples were collected and assayed foranti-β1 ECD and anti-AChR antibodies. The procedure wasrepeated after another 8 days. After this process, the non-immunized rabbit did not develop any anti-AChR antibodies andthe antibody titer in the immunized rabbit did not increase. Moreimportantly, the rabbits remained healthy for a long period(Tzartos et al., 2008). This preliminary experiment suggests thatthe ECD immunoadsorption procedure may be safe for use onpatients.
The specificity of the ECD resin was tested using known anti-MuSK antibody-positive sera. These experiments showed that noanti-MuSK antibodywas removed after extended incubation withthe ECD resin (Zisimopoulou et al., in press). Confirmation of the
specificity of the binding of the anti-AChR antibodies to the ECDimmunoadsorbents was provided by the finding that theantibodies did not bind to the control BSA immunoadsorbent.In addition, measurement of the total protein content in a testserum before and after immunoadsorption did not reveal anymeasurable reduction in protein after treatment, further verifyingthe specificity of immunoadsorption.
Additionally, it was found that immunoadsorption takes place invery short time (∼5 min), whereas the immunoadsorbents may beused repeatedly for 4–6 times, characteristics useful for the clinicalapplication of the technique. Finally, in preliminary studies, whereabout 4ml-columnswith ECD-Sepharose were used, the estimatedtime necessary for every 8 ml of serum with moderate titer (10–50 nM) was found less than 1 min, which means that a 50 ml ECDcolumn might be able to clear much more than 3 l serum per hour.
5. Activity of antibody-depleted sera and of purifiedanti-subunit autoantibodies in vitro and in vivo
The effectiveness of either plasmapheresis or Ig apheresisdoes not exclude the possibility that non-anti-AChR antibodyfactors in the plasma also contribute to the development of MGsymptoms. If this were true, the effectiveness of selective anti-AChR antibody depletion could be moderate. To examine this,we investigated the ability of the autoantibody-depleted seraand the purified subunit-specific autoantibodies to causereceptor loss in vitro and experimental MG (EAMG) in vivo.More specifically, using α1 and β1 ECD immunoadsorbents,we initially depleted MG sera of the corresponding antibodies,then isolated the bound antibodies from the immunoadsorbents.
The ability of the “depleted” MG sera and the purified ECDbound antibodies to cause AChR antigenic modulation (AChRloss) was first tested in vitro in TE671 cell cultures (Sideris et al.,2007). Sera partially depleted of anti-AChR antibodies still

Fig. 5. Proposed therapeutic immunoadsorption using a combination of twocolumns in series. The first one is made from immobilized E. coli-expressedECDs followed by the much smaller column with the immobilized yeast-expressed ECDs in order to adsorb antibodies which are not depleted from the E.coli column. Orange, green and black antibodies denote antibodies binding to E.coli-ECDs, yeast-ECDs or to neither, respectively. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web versionof this article.)
101P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
caused AChR antigenic modulation, but at a considerably lowerlevel, whereas completely depleted sera did not. The auto-antibodies isolated from the α1 or β1 ECD column were able tocause dose-dependent AChR loss, with the anti-α1 autoanti-bodies being approximately 4 times more potent than the anti-β1antibodies in triggering AChR modulation. This observationsupports the notion that the α1 subunit is the main target of thepathogenic autoantibodies.
Our next aimwas to determine the ability of the untreated sera,depleted sera, and isolated autoantibodies to passively transferEAMG to Lewis rats. TwoMG sera with high titers of antibodiesto both human and rat AChR and rich in anti-α ECD antibodieswere used in preliminary in vivo experiments. Four- to five-week-old Lewis rats were injected once i.p. with the different samplesand disease severity was assessed on the basis of clinicalsymptoms. The untreated sera and isolated anti-α1 subunit-specific antibodies were equally capable of inducing severe MGsymptoms and death. Interestingly, the depleted sera, which didnot contain antibodies reactive with rat AChR, did not induce anyMG-like symptoms (Tzartos et al., 2008; Kordas, Sideris, Poulasand Tzartos, unpublished).
Even though the latter result remains to be confirmed usingadditional sera, the fact that the antibody-depleted fractions ofthe MG sera did not induce MG-like symptoms suggests that theanti-AChR autoantibodies are the sole pathogenic factor inthese sera. Therefore, the proposed therapeutic approach aimedat the depletion of anti-AChR autoantibodies fromMG patients'plasma should be able to remove all or most of the pathogenicfactors.
6. Conclusions
The use of columns carrying immobilized ECDs of humanAChR subunits has a number of advantages. First, it would bespecific for anti-AChR antibodies, thus avoiding the removal ofuseful antibodies and other molecules from the patient andeliminating the need for replacement fluids. Second, it would bereasonably affordable, since the production of substantialamounts of recombinant AChR subunit ECDs, especiallyusing the prokaryotic expression system, is now a standardlaboratory procedure. Third, the immunoadsorbents are re-usable, since we showed that they can be used at least 4 times.Fourth, the titration of the immunoadsorbents showed asatisfactory efficiency in terms of the amount of recombinantECD needed for complete clearance of serum.
The described results suggest that a mixture of either yeast- orE. coli-expressed human AChR ECDs immobilized on aninsoluble material, such as CNBr-activated Sepharose, may beused in clinical trials for antigen-specificMG therapy as efficientimmunoadsorbents of anti-AChR autoantibodies from manyMG patients. E. coli-expressed ECD columns (which are muchmore economical) may be sufficient for some MG patients,whereas a combination of two columns in series (E. coli-expressed followed by yeast-expressed ECDs) may be neededfor others (Fig. 5). We envisage two possible procedures,depending on the urgency of each case: a. the use of bothcomplete columns (E. coli and yeast) each with the five subunit
ECDs, especially in the emergency case of a new patient, and b.an individualized approach, where a screening with the differentcolumns must be performed prior to the decision, which columnis suitable for the specific patient. This screening lasts only 1–2 days, a time in most cases affordable for the patient.
Importantly, anti-AChR autoantibody depletion was found tobe sufficient for the elimination of the pathogenic factor(s) fromtwo tested MG sera and the procedure seemed to be safe in twoexperimental rabbits. Finally, scaling up of the procedure isunderway as a necessary step prior to proceeding to clinicaltrials. Additional improvements in immunoadsorption effi-ciency might be achieved by the coexpression of all AChRECDs (free, or covalently linked with flexible peptides), whichmight result in their assembly into a pentameric AChR ECD.Such a pentamer might also contain inter-subunit epitopes, thusbinding more, or even all, anti-AChR antibodies. We arecurrently working towards this aim.
Acknowledgements
This work was supported by grants from the EuropeanCommission (Contract LSHM-CT-2006-037833), the GreekMinistry of Development-GSRT, the Muscular Dystrophy

102 P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
Association of the USA (MDA USA) and the AssociationFrancaise contre les Myopathies (AFM).
References
Agius, M.A., Richman, D.P., Vincent, A., 2003. Specific antibodies in thediagnosis and management of autoimmune disorders of neuromusculartransmission and related diseases. In: Kaminsky, H.J. (Ed.), Current ClinicalNeurology: Myasthenia Gravis and Related Disorders, pp. 177–196.
Berg, D., Shoop, R., Chang, K., et al., 2000. Nicotinic acetylcholine receptors inganglionic transmission. Experimental Pharmacology, vol. 144. Springer,New York, pp. 247–270.
Bitzopoulou, K., Kostelidou, K., Poulas, K., Tzartos, S.J., submitted forpublication. Mutant forms of the extracellular domain of the humanacetylcholine receptor-subunit with improved solubility and enhancedantigenicity. The importance of the Cys-loop. BBA.
Brannagan 3rd, T.H., Nagle, K.J., Lange, D.J., Rowland, L.P., 1996.Complications of intravenous immune globulin treatment in neurologicdisease. Neurology 47, 674–677.
Brejc, K., van Dijk, W.J., Klaassen, R.V., Schuurmans, M., van Der Oost, J., Smit,A.B., Sixma, T.K., 2001. Crystal structure of an ACh-binding protein revealsthe ligand-binding domain of nicotinic receptors. Nature 411, 269–276.
Chiu, H.C., Chen, W.H., Yeh, J.H., 2000. The six year experience ofplasmapheresis in patients withmyasthenia gravis. Therap. Apher. 4, 291–295.
Conti-Fine, B.M., Kaminski, H.J., 2001. Autoimmune neuromuscular transmis-sion disorders: myasthenia gravis and Lambert-Eaton myasthenic syndrome.Continuum 7.
Dalakas, M.C., 2002. Immunotherapies in the treatment of neuromusculardisorders. Neuromuscular Disorders in Clinical Practice. Butterworth Heine-mann, Boston, pp. 364–383.
Dau, P.C., Lindstrom, J.M., Cassel, C.K., Denys, E.H., Shev, E.E., Spitler, L.E.,1977. Plasmapheresis and immunosuppressive drug therapy in myastheniagravis. N. Engl. J. Med. 297, 1134–1140.
Engel, A.G., Lambert, E.H., Mulder, D.M., Torres, C.F., Sahashi, K., Ber, torini,T.E., Whitaker, J.N., 1982. A newly recognized congenital myasthenicsyndrome attributed to a prolonged open time of the acetylcholine-inducedion channel. Ann. Neurol. 11, 553–569.
Ferrero, B., Durelli, L., Cavallo, R., Dutto, A., Aimo, G., Pecchio, F.,Bergamasco, B., 1993. Therapies for exacerbation of myasthenia gravis. Themechanism of action of intravenous high-dose immunoglobulin G. Ann. N.Y. Acad. Sci. 681, 563–566.
Fostieri, E., Kostelidou, K., Poulas, K., Tzartos, S.J., 2006. Recent advances in theunderstanding and therapy of myasthenia gravis. Future Neurol. 1, 799–801.
Gajdos, P., Outin, H.D., Morel, E., Raphael, J.C., Goulon, M., 1987. High-doseintravenous gamma globulin for myasthenia gravis: an alternative to plasmaexchange. Ann. N.Y. Acad. Sci. 505, 842–844.
Grando, S., Horton, R., 1997. The keratinocyte cholinergic system with acetylcho-line as an epidermal cytotransmitter. Curr. Opin. Dermatol. 4, 262–268.
Guo, C.Y., Li, Z.Y., Xu, M.Q., Yuan, J.M., 2005. Preparation of animmunoadsorbent coupled with a recombinant antigen to remove anti-acetylcholine receptor antibodies in abnormal serum. J. Immunol. Methods303, 142–147.
Kaiser, S., Soliokov, L., Wonnacott, S., 2000. Presynaptic neuronal nicotinicreceptors: pharmacology, heterogeneity and cellular mechanisms. Experi-mental Pharmacology, vol. 144. Springer, New York, pp. 193–212.
Kaminski, H.J., 2001. Myasthenia gravis. Neuromuscular Disorders in ClinicalPractice. Butterworth Heinemann, MA, pp. 916–930.
Kaminski, H.J., 2003. Treatment of myasthenia gravis. Current ClinicalNeurology: Myasthenia Gravis and Related Disorders, pp. 197–221.
Karlin, A., 2002. Emerging structure of the nicotinic acetylcholine receptors.Nat. Rev. Neurosci. 3, 102–114.
Kazatchkine, M.D., Kaveri, S.V., 2001. Immunomodulation of autoimmune andinflammatory diseases with intravenous immune globulin. N. Engl. J. Med.345, 747–755.
Kostelidou, K., Trakas, N., Zouridakis, M., Bitzopoulou, K., Sotiriadis, A.,Gavra, I., Tzartos, S.J., 2006. Expression and characterization of soluble
forms of the extracellular domains of the beta, gamma and epsilon subunitsof the human muscle acetylcholine receptor. FEBS J. 273, 3557–3568.
Kostelidou, K., Trakas, N., Tzartos, S.J., 2007. Extracellular domains of thebeta, gamma and epsilon subunits of the human acetylcholine receptor asimmunoadsorbents for myasthenic autoantibodies: a combination ofimmunoadsorbents results in increased efficiency. J. Neuroimmunol.
Lang, B., Newsom-Davis, J., Prior, C., Wray, D., 1983. Antibodies to motornerve terminals: an electrophysiological study of a human myasthenicsyndrome transferred to mouse. J. Physiol. 344, 335–345.
Lindstrom, J., 2000. The structures of neuronal nicotinic receptors. Experi-mental Pharmacology, vol. 144. Springer, New York, pp. 101–162.
Lindstrom, J.M., Seybold, M.E., Lennon, V.A., Whittingham, S., Duane, D.D.,1976. Antibody to acetylcholine receptor in myasthenia gravis. Prevalence,clinical correlates, and diagnostic value. Neurology 26, 1054–1059.
Lindstrom, J., Campbell, M., Nave, B., 1978. Specificities of antibodies toacetylcholine receptors. Muscle Nerve 1, 140–145.
Loutrari, H., Tzartos, S.J., Claudio, T., 1992. Use of Torpedo-mouse hybridacetylcholine receptors reveals immunodominance of the alpha subunit inmyasthenia gravis antisera. Eur. J. Immunol. 22, 2949–2956.
Macklin, K.D., Maus, A.D., Pereira, E.F., Albuquerque, E.X., Conti-Fine, B.M.,1998. Human vascular endothelial cells express functional nicotinicacetylcholine receptors. J. Pharmacol. Exp. Ther. 287, 435–439.
Mat ic, G., Winkler, R.E., Tiess, M., Ramlow, W., 2001. Selective apheresis—time for a change. Int. J. Artif. Organs 24, 4–7.
Maus, A.D., Pereira, E.F., Karachunski, P.I., Horton, R.M., Navaneetham, D.,Macklin, K., Cortes, W.S., Albuquerque, E.X., Conti-Fine, B.M., 1998.Human and rodent bronchial epithelial cells express functional nicotinicacetylcholine receptors. Mol. Pharmacol. 54, 779–788.
Mossman, S., Vincent, A., Newsom-Davis, J., 1988. Passive transfer ofmyasthenia gravis by immunoglobulins: lack of correlation between AChRwith antibody bound, acetylcholine receptor loss and transmission defect.J. Neurol. Sci. 84, 15–28.
Patrick, J., Lindstrom, J., 1973. Autoimmune response to acetylcholine receptor.Science 180, 871–872.
Pinching, A.J., Peters, D.K., Newso,m-Davis, J., 1976. Remission of myastheniagravis following plasma-exchange. Lancet 2, 1373–1376.
Psaridi-Linardaki, L., Mamalaki, A., Remoundos, M., Tzartos, S.J., 2002.Expression of soluble ligand- and antibody-binding extracellular domain ofhuman muscle acetylcholine receptor alpha subunit in yeast Pichia pastoris.Role of glycosylation in alpha-bungarotoxin binding. J. Biol. Chem. 277,26980–26986.
Psaridi-Linardaki, L., Mamalaki, A., Tzartos, S.J., 2003. Future therapeuticstrategies in autoimmune myasthenia gravis. Ann. N. Y. Acad. Sci. 998,539–548.
Psaridi-Linardaki, L., Trakas, N., Mamalaki, A., Tzartos, S.J., 2005. Specificimmunoadsorption of the autoantibodies from myasthenic patients using theextracellular domain of the human muscle acetylcholine receptor alpha-subunit. Development of an antigen-specific therapeutic strategy. J. Neuroim-munol. 159, 183–191.
Ptak, J., 2004. Changes of plasma proteins after immunoadsorption using Ig-Adsopak columns in patients with myasthenia gravis. Transfus. Apher. Sci.30, 125–129.
Samuelsson, A., Towers, T.L., Ravetch, J.V., 2001. Anti-inflammatory activityof IVIG mediated through the inhibitory Fc receptor. Science 291, 484–486.
Sekhon, H.S., Jia, Y., Raab, R., Kuryatov, A., Pankow, J.F., Whitsett, J.A.,Lindstrom, J., S, pindel, E.R., 1999. Prenatal nicotine increases pulmonaryalpha7 nicotinic receptor expression and alters fetal lung development inmonkeys. J. Clin. Invest. 103, 637–647.
Shillito, P., Molenaar, P.C., Vincent, A., Leys, K., Zheng, W., van den Berg, R.J.,Plo, mp, J.J., van Kempen, G.T., Chauplannaz, G., Wintzen, A.R., et al.,1995. Acquired neuromyotonia: evidence for autoantibodies directed againstK+ channels of peripheral nerves. Ann. Neurol. 38, 714–722.
Sideris, S., Lagoumintzis, G., Kordas, G., Kostelidou, K., Sotiriadis, A., Poulas,K., Tzartos, S.J., 2007. Isolation and functional characterization of anti-acetylcholine receptor subunit-specific autoantibodies from myasthenicpatients: receptor loss in cell culture. J. Neuroimmunol. 189, 111–117.
Skok, M., Gr, ailhe, R., Changeux, J.P., 2005. Nicotinic receptors regulate Blymphocyte activation and immune response. Eur. J. Pharmacol. 517, 246–251.

103P. Zisimopoulou et al. / Journal of Neuroimmunology 201–202 (2008) 95–103
Takamori, M., Maruta, T., 2001. Immunoadsorption in myasthenia gravis basedon specific ligands mimicking the immunogenic sites of the acetylcholinereceptor. Therap. Apher. 5, 340–350.
Tzartos, S., Hochschwender, S., Vasquez, P., Lindstrom, J., 1987. Passivetransfer of experimental autoimmune myasthenia gravis by monoclonalantibodies to the main immunogenic region of the acetylcholine receptor. J.Neuroimmunol. 15, 185–194.
Tzartos, S.J., Bitzopoulou, K., Gavra, I., Kordas, G., Jacobson, L., Kostelidou,K., Lagoumintzis, G., Lazos, O., Poulas, K., Sideris, S., Sotiriadis, A.,Trakas, N., Zisimopoulou, P., 2008. Antigen-specific apheresis ofpathogenic autoantibodies from myasthenia gravis sera. Antigen-specificapheresis of pathogenic autoantibodies from myasthenia gravis sera.Myasthenia Gravis and Related Disorders: Ann N Y Acad Sci XIthInternational Conference, vol. 1131. est. pub date May 2008.
Unwin, N., 2005. Refined structure of the nicotinic acetylcholine receptor at 4Aresolution. J. Mol. Biol. 346, 967–989.
Wang, H., Yu, M., Ochani, M., Amella, C.A., Tanovic, M., Susarla, S., Li, J.H.,Yang, H., Ulloa, L., Al-Abed, Y., Czura, C.J., Tracey, K.J., 2003. Nicotinic
acetylcholine receptor alpha7 subunit is an essential regulator of inflamma-tion. Nature 421, 384–388.
Yamazaki, Z., Fujimori, Y., Takahama, T., Inoue, N., Wada, T., Kazama, M.,Morioka, M., Abe, T., Yamawaki, N., Inagaki, K., 1982. Efficiency andbiocompatibility of a new immunosorbent. Trans Am Soc Artif. Intern.Organs 28, 318–323.
Zisimopoulou, P., Lagoumintzis, G., Poulas, K., Tzartos, S., in press. Antigen-specific apheresis of human anti-acetylcholine receptor autoantibodies frommyasthenia gravis patients' sera using Escherichia coli-expressed receptordomains. J Neuroimmunol.
Zoli, M., 2000. Distribution of cholinergic neurons in the mammalian brain withspecial reference to their relationship with neuronal nicotinic receptors.Experimental Pharmacology, vol. 144. Springer, New York, pp. 13–30.
Zouridakis, M., Kostelidou, K., Sotiriadis, A., Stergiou, C., Eliopoulos, E.,Poulas, K., Tzartos, S.J., 2007. Circular dichroism studies of extracellulardomains of human nicotinic acetylcholine receptors provide an insight intotheir structure. Int. J. Biol. Macromol. 41, 423–429.