tissue-level biomarkers and biological effect of mercury on sentinel slugs, arion ater
TRANSCRIPT

Arch. Environ. Contam. Toxicol. 31, 54-62 (1996) A R C H I V E S O F
Environmental C o n t a m i n a t i o n a n d T o x i c o l o g y © 1996 Springer-Verlag N~w York Inc.
Tissue-Level Biomarkers and Biological Effect of Mercury on Sentinel Slugs, Arion ater
I. Marig6mez, M. Soto, M. Kortabitarte
Biologia Zelularra eta Zientzia Morfologikoen Saila, Zientzi Fakultatea, Euskal Herriko Unibertsitatea, 644 RK., E-48080 Bilbo, Basque Country, Spain
Received: 27 July 1995/Revised: 11 December 1995
Abstract. The present investigation was to determine the extent to which mercury (Hg) provokes measurable effects on the structure of the digestive gland of slugs as well as to relate the extent of these effects to the cell and tissue distribution of Hg. For this purpose, slugs (Arion ater) received various dietary concentrations of Hg (from 0 to 1,000 Ixg Hg/g food) as chloride for 30 days and were histologically examined every third day. Autometallography was used to demonstrate Hg as black silver deposits (BSD) in paraffin sections. The lysosomes and residual bodies of digestive cells resulted to be the major accumulation sites. In addition, Hg was also evidenced in lipofuscine granules of vacuoles in excretory cells but, however, it was rarely ob- served within calcium cells. Generally, the extent of BSD in- creased with dietary Hg concentration and exposure time but, however, it became significant lowered after exposure to 1,000 Ixg Hg/g food for 30 when the digestive epithelium appeared almost devoid of digestive cells. On the other hand, significant changes were recorded in the quantitative structure of digestive tubules. Mean Epithelial Thickness (MET), Mean Luminal Ra- dius (MLR) and Mean Diverticular Radius (MDR) were re- corded as measures of the sublethal biological effect of Hg. MET, MLR/MET and MET/MDR were affected by Hg concen- tration (C), exposure time (T) and CxT interaction, changes in MET, MLR, MLR/MET and MET/MDR being explained by regression models after logarithmic transformation of the data. In order to explain the nature of the changes in the quantitative structure of the digestive tubules this investigation was comple- mented with qualitative histological observations. According to them, the excretory activity in digestive cells was initially enhanced. Afterwards, the relative numbers of digestive cells declined until the extreme cases of exposure to 1,000 Ixg Hg/g for 27 to 30 days in which the digestive epithelium was mostly comprised of calcium and excretory cells. Concomi- tantly, some changes took place in blood vessels where Leydig cells became disrupted and the connective tissue layers thick- ened. Finally, it is suggested to use slugs in soil quality assess-
Correspondence to: I. Marig6mez
ment as sentinel organisms ("Slug Watch") in which biomarkers of exposure to metallic pollutants and of biological effect are re- corded.
One of the problems in soil quality assessment is the fact that exposure to pollutants generally occurs at sublethal doses for long periods. This is a problem common to all the environment but in aquatic systems well-developed monitoring programs are currently being performed (Bayne 1989). These programs are based on the use of sentinel species, mainly molluscs, able to detect changes in pollutant bioavailability and their biologi- cal effect.
Slugs and snails have been previously proposed as sentinels for soil quality assessment due to their ability to accumulate environmental toxicants as well as to their sensitive responses to environmental stress (Popham and D'Auria 1980; Berger and Dallinger 1993).
Previous reports have shown the effects of dietary copper (Cu) and zinc (Zn) on the digestive gland of slugs, Arion ater
(Marigdmez et al. 1986a, 1986b; Recio et al. 1988a, 1988b). The main results can be summarized in three points. First, metals are localized in different cell types but their distribution also changes according to the exposure-time and dietary concen- tration of the metal (Marigdmez et aI. 1986a; Recio et al.
1988a). Second, the epithelium of the digestive gland is reduced in thickness at increasing dietary intake of Cu and Zn in excess (Marigrmez et al. 1986b; Recio et al. 1988b). Third, the detoxi- fication of metals is done by excretion from the digestive gland to the faeces and is closely related to calcium metabolism (Marig6mez et al. 1986a; Recio et al. 1988a; Ireland and Mari- grmez 1992).
The first purpose of the present investigation was to determine whether toxic metals without known biological role, such as mercury (Hg), provoke similar effects on the quantitative struc- ture of the digestive gland of slugs. Marigrmez et al. (1986c) observed that both feeding activity and growth were more strongly affected by Hg than by either Cu or Zn under similar

Biomarkers of Hg Pollution in Sentinel Slugs 55
experimental conditions. Nevertheless, Hg did not affect mortal- ity and for this reason we thought it would be possible to use biomarkers of environmental stress to assess sublethal toxicity of Hg. A series of studies have demonstrated that the digestive gland tubules of molluscs are reduced in thickness in response to a variety of environmental stressors (Lowe et al. 1981; Couch 1984; Tripp et al. 1984; Marig6mez et al. 1986b; 1993; Minnitti 1987; Axiak et al. 1988; Recio et al. 1988b; Lowe and Clarke 1989; Vega et aI. 1989; Cajaraville et al. 1992; Ireland and Marigrmez 1992) and have proposed the measure of such reduc- tion as a biomarker of the biological effect of sublethal expo- sures to pollutants.
The second purpose was to relate quantitative changes in the morphology of the digestive epithelium to the cellular and subcellular distribution of Hg in exposed slugs as determined by histochemistry. A similar approach was made by Marigdmez et al. (1986a), Recio et al. (1988a), Marigdmez et al. (1990a), and Ireland and Marigdmez (1992) to determine whether metal is accumulated in exposed organisms. It is simple, quicker and cheaper than chemical analysis and may offer an alternative for routine work able to help in discarding chemical analyses whenever possible. Histochemistry is a specially valuable tool in the particular case of Hg because this metal presents serious problems to be quantified by analytical chemistry.
In addition, it was planned to describe the changes in the cell type composition and the morphology of the cell types comprising the digestive epithelium since these histo- logical observations might he useful to understand the nature of the effects of Hg on the digestive epithelium morphol- ogy (Marig6mez et al. 1990b), as well as to explain the dynamics of Hg accumulation in the digestive gland tissue (Soto 1995).
Materials and Methods
Experiment and Histological Processing o f Samples
Two simultaneous sets of seven groups of slugs (Arion ater) were treated with different Hg concentrations in diet (0, 10, 25, 50, 100, 300, and 1,000 ~xg Hg/g food) for a month. For this purpose, 198 slugs measuring 4-5 cm in length (5-6 g) were collected from a field near the University Campus the llth of July and taken to the laboratory to be distributed in 14 plastic boxes (20 × 18 × 8 cm3). Animals were starved for 6 days prior to Hg treatment in order to minimize physiologi- cal differences and provide acclimation to laboratory conditions. Mer- cury treatment was started with 12 slugs per box. Mean temperature during the assay was 20°C and relative humidity was maintained at saturation levels. "Natural diet" (equiproportional triturate of lettuce, apple, carrot and pumpkin with an 1.5 agar aqueous solution) was supplied to slugs mixed with mercury chloride in the proper concentra- tion for each group of treatment. Exposure to Hg did not cause a significant mortality: three, one, one, two and five slugs died in the 0, 25, 50, 300, and 1,000 Ixg Hg/g food groups, respectively. Feeding activity and growth rate decreased at increasing Hg concentrations in diet. A more detailed description of the experimental conditions are given by Marigdmez et al. (1986c).
Slugs were always removed between 5:00 and 7:00 p.m. for histolog- ical examination after 3, 6, 9, 12, 18, 21, 24, 27, and 30 days of treatment. The digestive gland of 2 individuals per treatment was excised, fixed in 4% formaldehyde at 4°C overnight and embedded in paraffin.
Quantitative Morphology o f Digestive Tubules
Paraffin-sections (8 txm) were stained with haematoxylin-eosin. Draw- ings of digestive tubule sections were realized at a final × 650 magnifi- cation with the aid of a drawing-tube attachment onto a Nikon "Opti- phot" microscope. According to previous statistical analyses, five tubule sections in five histological sections, with at least 50/xm distance between each other, were drawn and measured by the planimetric procedure called Geometrical Transformation (Recio et al. 1988b; Soto et al. 1990) after introduction of tubule section profiles by a scanner (Scan jet IIp, Hewlett Packard) attached to a 486 PC computer (Eduskan- 93 Software Package, Sfinchez 1993). Five parameters were calculated (Vega et al. 1989): MET, Mean Epithelial Thickness; MDR, Mean Diverticular Radius; MLR, Mean Luminal Radius; MLR/MET and MET/MDR. Data analysis was carried out by means of two-way ANOVA and multiple regression analyses with the aid of the SPSS/ PC + statistical package (SPSS Inc, Microsoft Co). Three-dimensional surface responses attained after multiple regression analysis were drawn by using Mathcad 4.0 (Madsoft).
Mercury Histochemistry: AutometaIlography
Another set of paraffin sections was stained by autometallography, a procedure to demonstrate metals in tissue sections as black silver deposits (BSD), which is based on the principles of photography (Danscher 1984; Soto 1995). Paraffin sections were dewaxed in xylene, hydrated in ethanol-water mixtures and left in an oven at 40°C until sections were completely dried. Then, tissues sections were covered with a photographic emulsion (Ilford Nuclear Emulsion L4) under safety light conditions. After drying under completely dark conditions (above 30 min) sections were rinsed in developer bath (1:5, Ultrathin Tetenal SF) for 15 min, rinsed in stop bath (1% acetic acid) for 1 min, and finally rinsed in a fixative bath (1:9, Agefix) for 10 min. Sections were mounted in Kaiser's gelatin and sealed with nails protector (Soto 1995).
Some histochemical controls were performed in parallel on several series of tissue sections stained with either (a) the Cambar's method to demonstrate Hg (Pearse 1980), (b) alizarin red for calcium (Ca, Martoja and Martoja-Pierson 1970; Marig6mez et al. 1986b), or (c) the Stoeltzner's technique with cobalt nitrate for calcium (Ca, Martoja and Martoja-Pierson 1970; Marigrmez et al. 1986b).
Results
Mercury was taken up by the slugs and accumulated in their tissues as evidenced by the increasing BSD extent recorded in the digestive gland of slugs exposed to increasing dietary levels of the metal (Figures la-e). The histochemical method of Cambar for Hg, however, did not indicate the presence of the metal in the studied tissues. This technique was very corrosive and the structural integrity of the tissue sections was too poor to enable feasible observations at the light microscope. Con- versely, autometallography proved to be an excellent technique to visualize Hg in slug tissue sections (Figure i). Thus, the digestive gland exhibited a great ability to accumulate Hg, the lysosomal system (mainly heterolysosomes--or green gran- u l e s - a n d residual bodies--or yellow granules) of the digestive cells being the primary accumulation site (Figures lb-e). After threshold exposure, Hg was also accumulated in the lipofuscine granules of the excretory cells (Figures 1 d and e) but, however, BSD were rarely observed in calcium cells (Figures If and g). In addition, BSD were observed forming layers at the basal
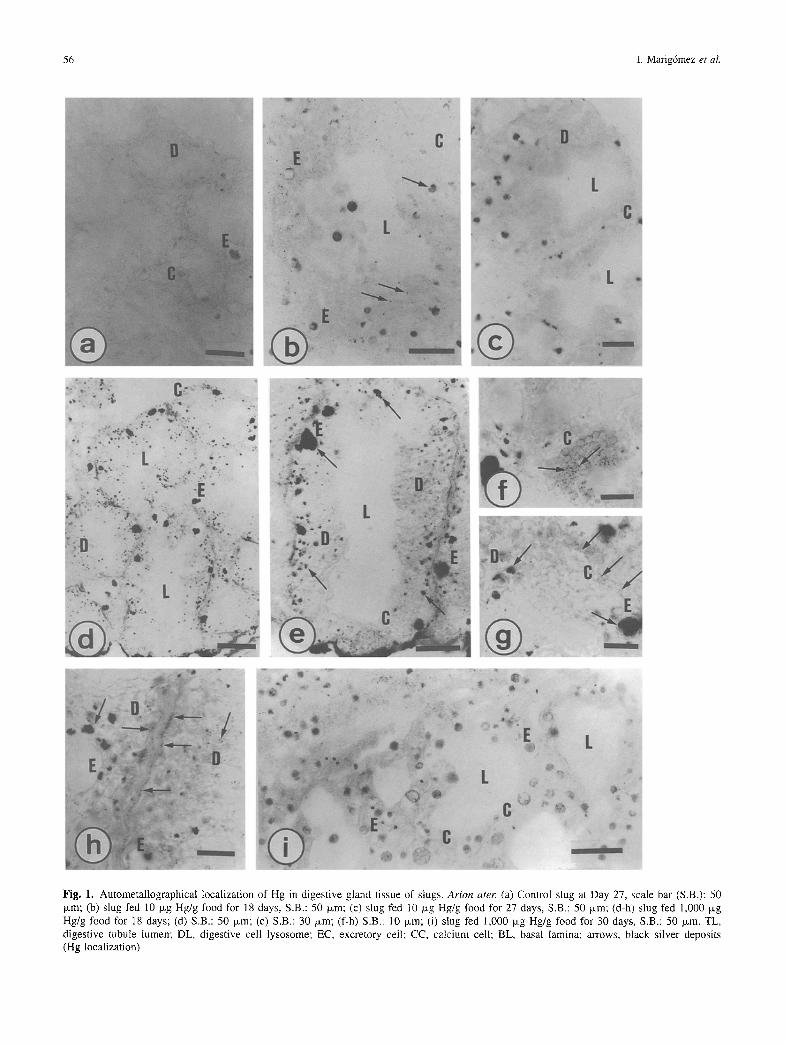
56 I. Marig6mez et al.
Fig. 1. Autometallographical localization of Hg in digestive gland tissue of slugs, Ar ion a ter (a) Control slug at Day 27, scale bar (S.B.): 50 p,m; (b) slug fed 10 o,g Hg/g food for 18 days, S.B.: 50 p,m; (c) slug fed 10 ~g Hg/g food for 27 days, S.B.: 50 ixm; (d-h) slug fed 1,000 Ixg Hg/g food for 18 days; (d) S.B.: 50 ixm; (e) S.B.: 30 txm; (f-h) S.B.: l0 ~m; (i) slug fed 1,000 #,g Hg/g food for 30 days, S.B.: 50 ~m. TL, digestive tubule lumen; DL, digestive cell lysosome; EC, excretotN cell; CC, calcium cell; BL, basal lamina; arrows, black silver deposits (Hg localization)
Biomarkers of Hg Pollution in Sentinel Slugs 57
@ pc @ ~-~ ~ EC CC
DC ~ ~ ~2)~ 4 "~
~'~ 3
(3) = 2
=
Dietary Hg
1000 ~tg/g 300 ~g/g
100 ~tg/g ::,: 50 ~g/g
: 10 ~lg/g
~l"l~["~ .................................... Control
I I I I I 0 0 6 12 18 24 30
Exposure Time (days)
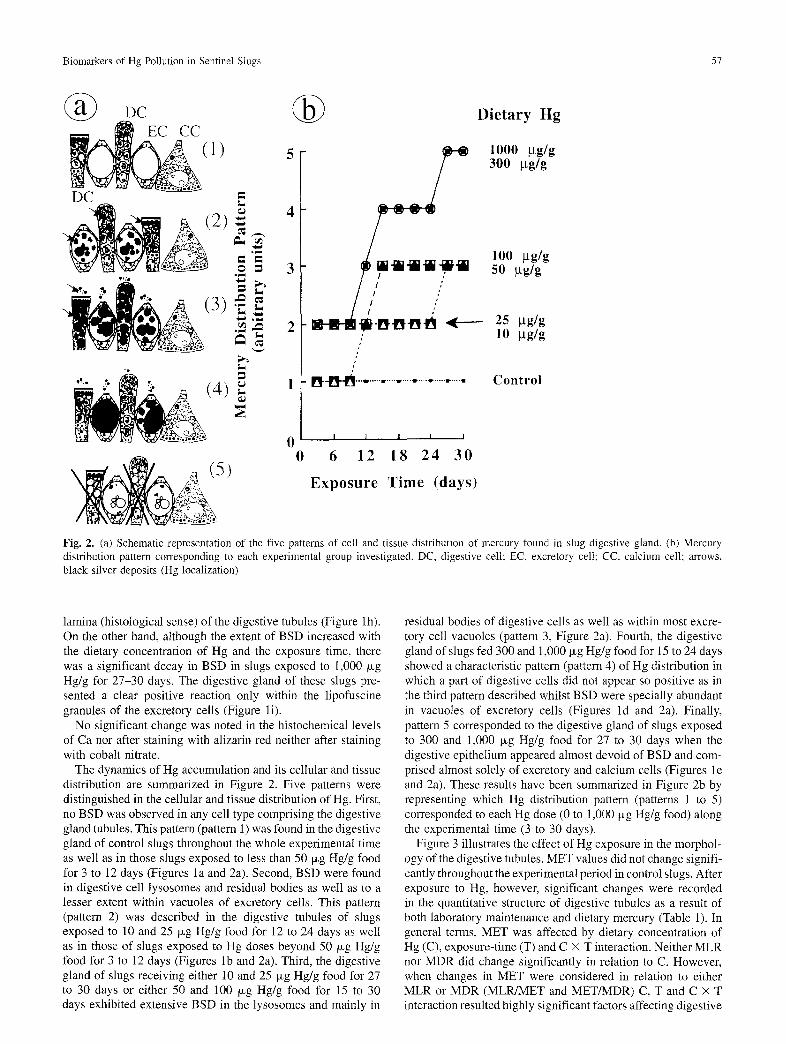
Fig. 2. (a) Schematic representation of the five patterns of cell and tissue distribution of mercury found in slug digestive gland. (b) Mercury distribution pattern corresponding to each experimental group investigated. DC, digestive cell; EC, excretory cell; CC, calcium cell; arrows, black silver deposits (Hg localization)
lamina (histological sense) of the digestive tubules (Figure lh). On the other hand, although the extent of BSD increased with the dietary concentration of Hg and the exposure time, there was a significant decay in BSD in slugs exposed to 1,000 ~g Hg/g for 27-30 days. The digestive gland of these slugs pre- sented a clear positive reaction only within the lipofuscine granules of the excretory cells (Figure li).
No significant change was noted in the histochemical levels of Ca nor after staining with alizarin red neither after staining with cobalt nitrate.
The dynamics of Hg accumulation and its cellular and tissue distribution are summarized in Figure 2. Five patterns were distinguished in the cellular and tissue distribution of Hg. First, no BSD was observed in any cell type comprising the digestive gland tubules. This pattern (pattern 1) was found in the digestive gland of control slugs throughout the whole experimental time as well as in those slugs exposed to less than 50 txg Hg/g food for 3 to 12 days (Figures la and 2a). Second, BSD were found in digestive cell lysosomes and residual bodies as well as to a lesser extent within vacuoles of excretory cells. This pattern (pattern 2) was described in the digestive tubules of slugs exposed to 10 and 25 txg Hg/g food for 12 to 24 days as well as in those of slugs exposed to Hg doses beyond 50 txg Hg/g food for 3 to 12 days (Figures lb and 2a). Third, the digestive gland of slugs receiving either 10 and 25 txg Hg/g food for 27 to 30 days or either 50 and 100 txg Hg/g food for 15 to 30 days exhibited extensive BSD in the lysosomes and mainly in
residual bodies of digestive cells as well as within most excre- tory cell vacuoles (pattern 3, Figure 2a). Fourth, the digestive gland of slugs fed 300 and 1,000 p~g Hg/g food for 15 to 24 days showed a characteristic pattern (pattern 4) of Hg distribution in which a part of digestive cells did not appear so positive as in the third pattern described whilst BSD were specially abundant in vacuoles of excretory cells (Figures ld and 2a). Finally, pattern 5 corresponded to the digestive gland of slugs exposed to 300 and 1,000 txg Hg/g food for 27 to 30 days when the digestive epithelium appeared almost devoid of BSD and com- prised almost solely of excretory and calcium cells (Figures le and 2a). These results have been summarized in Figure 2b by representing which Hg distribution pattern (patterns 1 to 5) corresponded to each Hg dose (0 to 1,000 txg Hg/g food) along the experimental time (3 to 30 days).
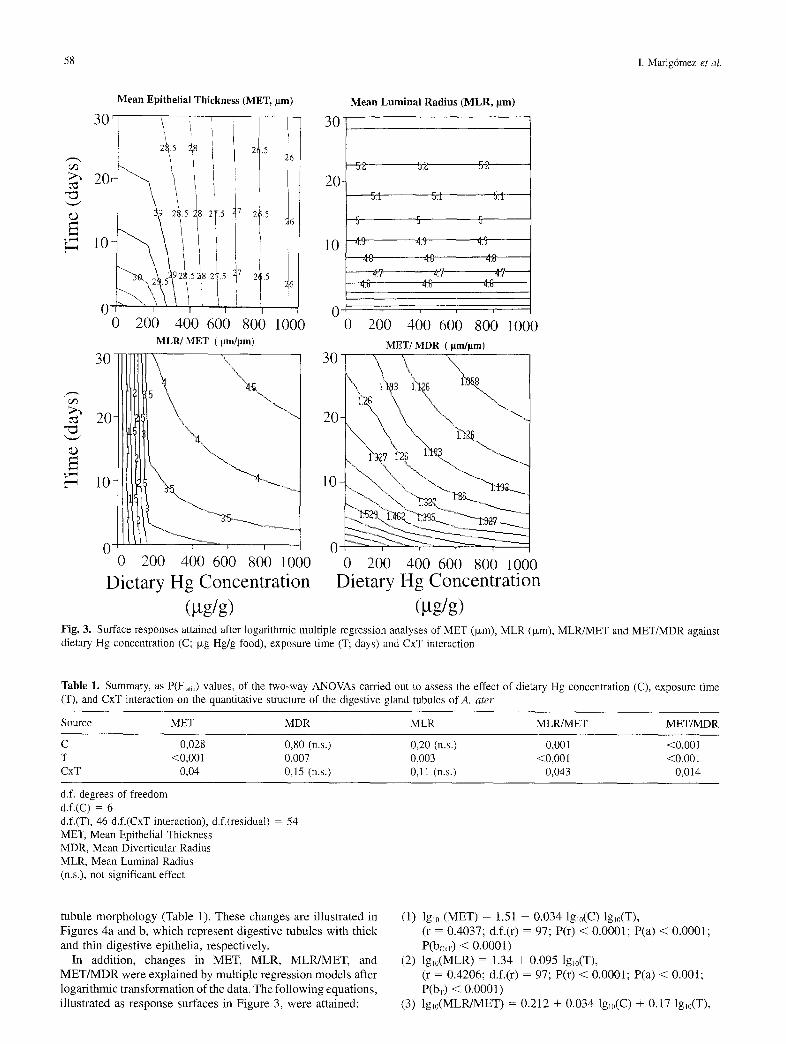
Figure 3 illustrates the effect of Hg exposure in the morphol- ogy of the digestive tubules. MET values did not change signifi- cantly throughout the experimental period in control slugs. After exposure to Hg, however, significant changes were recorded in the quantitative structure of digestive tubules as a result of both laboratory maintenance and dietary mercury (Table 1). In general terms, MET was affected by dietary concentration of Hg (C), exposure-time (T) and C × T interaction. Neither MLR nor MDR did change significantly in relation to C. However, when changes in MET were considered in relation to either MLR or MDR (MLR/MET and MET/MDR) C, T and C × T interaction resulted highly significant factors affecting digestive
58 I. Marig6mez et al.
° ,m,,~
[ -
Mean Epithelial Thickness (MET, ~tm)
MLR/MET (pro/pro)
30-
20-
I0
n [ l I
0 200 400 600 800 1000
Dietary Hg Concentration (gg/g)
30
20
10 ̧
Mean Luminal Radius (MLR, turn)
---~2 ~ ~ ....
5.1 5.1 5.1
---5 5 5
42 4 9 -
42 4:8 4~ 4~ 4.7
0 t 0 200 400 600 800 1000
MET/MDR (~m/lam) 30 r ',~ ~ , ~
1. 3 1~. G
1 i I I
0 200 400 600 800 1000 Dietary Hg Concentration
(gg/g) Fig. 3. Surface responses attained after logarithmic multiple regression analyses of MET (ixm), MLR (Ixm), MLR/MET and MET/MDR against dietary Hg concentration (C; Ixg Hg/g food), exposure time (T; days) and CxT interaction
Table 1. Summary, as P(Fr~,,) values, of the two-way ANOVAs carried out to assess the effect of dietary Hg concentration (C), exposure time (T), and CxT interaction on the quantitative structure of the digestive gland tubules of A. a ter
Source MET MDR MLR MLR/MET MET/MDR
C 0,028 0,80 (n.s.) 0,20 (n.s.) 0,001 <0,001 T <0,001 0,007 0,003 <0,00l <0,001 CxT 0,04 0,15 (n.s.) 0,11 (n.s.) 0,043 0,014
d.f. degrees of freedom d.f.(C) = 6 d.f.(T), 46 d.f.(CxT interaction), d.f.(residual) = 54 MET, Mean Epithelial Thickness MDR, Mean Diverticular Radius MLR, Mean Luminal Radius (n.s.), not significant effect
tubule morphology (Table 1). These changes are illustrated in Figures 4a and b, which represent digestive tubules with thick and thin digestive epithelia, respectively.
In addition, changes in MET, MLR, MLPUMET, and MET/MDR were explained by multiple regression models after logarithmic transformation of the data. The following equations, illustrated as response surfaces in Figure 3, were attained:
(1) lgl0 (MET) = 1.51 - 0.034 lglo(C) lgl0(T), (r = 0.4037; d.f.(r) = 97; P(r) < 0.0001; P(a) < 0.0001; P(bcxv) < 0.0001)
(2) lg~0(MLR) = 1.34 + 0.095 lglo(T), (r = 0.4206; d.f.(r) = 97; P(r) < 0.0001; P(a) < 0.001; P(bv) < 0.0001)
(3) lg~0(MLRfMET) = 0.212 + 0,034 lg~0(C) + 0.17 lgl0(T),
Biomarkers of Hg Pollution in Sentinel Slugs 59
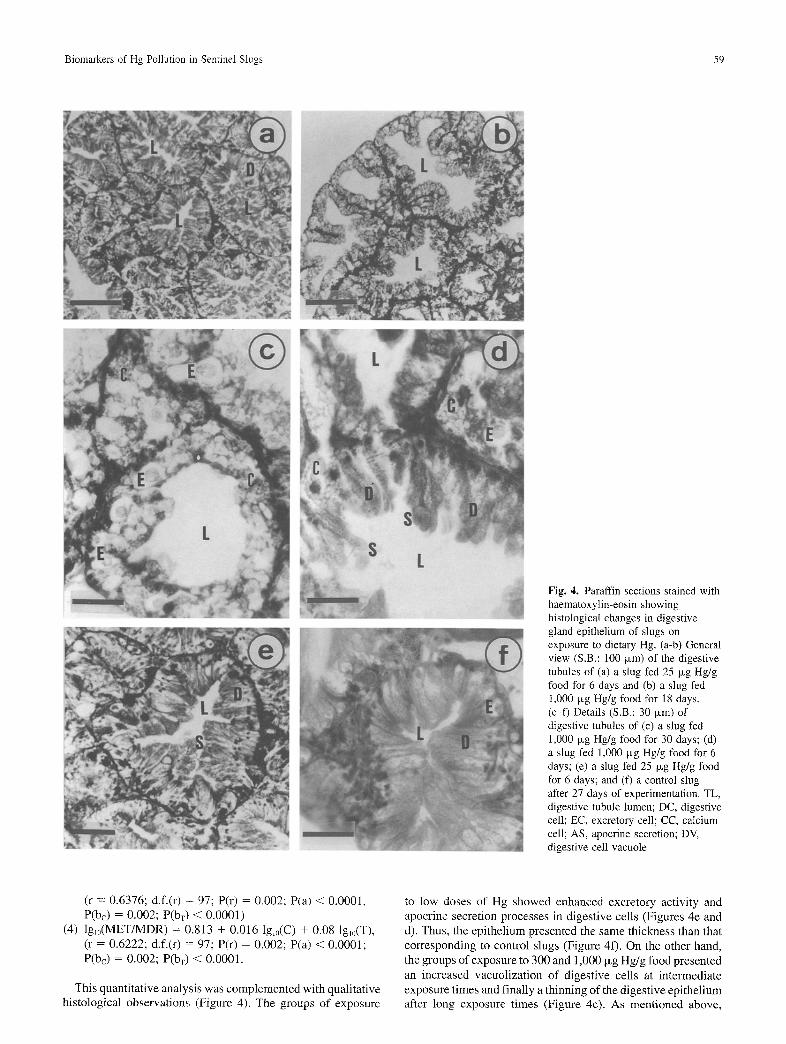
~!iiiii;i Fig. 4. Paraffin sections stained with haematoxylin-eosin showing histological changes in digestive gland epithelium of slugs on exposure to dietary Hg. (a-b) General view (S.B.: 100 Ixm) of the digestive tubules of (a) a slug fed 25 Ixg Hg/g food for 6 days and (b) a slug fed 1,000 txg Hg/g food for 18 days. (c-f) Details (S.B.: 30 Ixm) of digestive tubules of (c) a slug fed 1,000 Ixg Hg/g food for 30 days; (d) a slug fed 1,000 Ixg Hg/g food for 6 days; (e) a slug fed 25 ~g Hg/g food for 6 days; and (f) a control slug after 27 days of experimentation. TL, digestive tubule lumen; DC, digestive cell; EC, excretory cell; CC, calcium cell; AS, apocrine secretion; DV, digestive cell vacuole
(r = 0.6376; d.f.(r) = 97; P(r) = 0.002; P(a) < 0.0001, P(bc) = 0.002; P(br) < 0.0001)
(4) lgI0(MET/MDR) = 0.813 + 0.016 lgl0(C) + 0.08 lgl0(T), (r = 0.6222; d.f.(r) = 97; P(r) = 0.002; P(a) < 0.0001; P(bc) = 0.002; P(bT) < 0.0001.
This quantitative analysis was complemented with qualitative histological observations (Figure 4). The groups of exposure
to low doses of Hg showed enhanced excretory activity and apocrine secretion processes in digestive cells (Figures 4e and d). Thus, the epithelium presented the same thickness than that corresponding to control slugs (Figure 4f). On the other hand, the groups of exposure to 300 and 1,000 Ixg Hg/g food presented an increased vacuolization of digestive cells at intermediate exposure times and finally a thinning of the digestive epithelium after long exposure times (Figure 4c). As mentioned above,
60 I. Marig6mez et al.
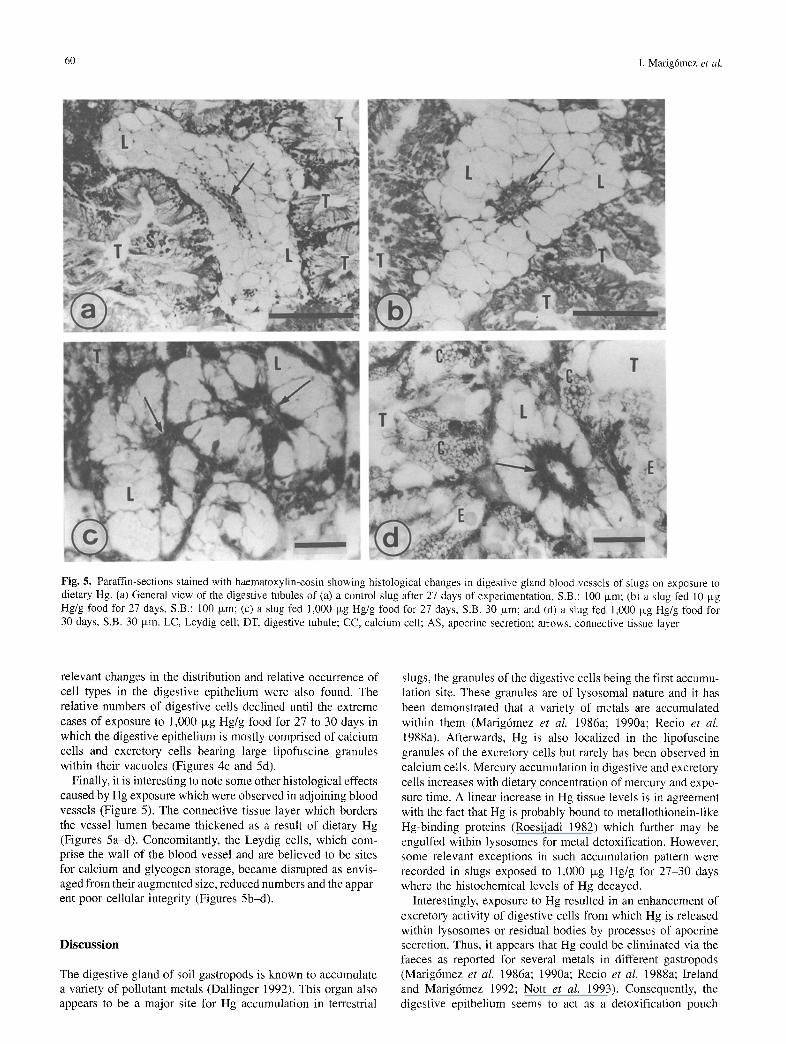
Fig. 5. Paraffin-sections stained with haematoxylin-eosin showing histological changes in digestive gland blood vessels of slugs on exposure to dietary Hg. (a) General view of the digestive tubules of (a) a control slug after 27 days of experimentation, S.B.: 100 Ixm; (b) a slug fed 10 p~g Hg/g food for 27 days, S.B.: 100 ~xm; (c) a slug fed 1,000 Ixg Hg/g food for 27 days, S.B. 30 p~m; and (d) a slug fed 1,000 Ixg Hg/g food for 30 days, S.B. 30 ixm. LC, Leydig cell; DT, digestive tubule; CC, calcium cell; AS, apocrine secretion; arrows, connective tissue layer
relevant changes in the distribution and relative occurrence of cell types in the digestive epithelium were also found. The relative numbers of digestive cells declined until the extreme cases of exposure to 1,000 Ixg Hg/g food for 27 to 30 days in which the digestive epithelium is mostly comprised of calcium cells and excretory cells bearing large lipofuscine granules within their vacuoles (Figures 4c and 5d).
Finally, it is interesting to note some other histological effects caused by Hg exposure which were observed in adjoining blood vessels (Figure 5). The connective tissue layer which borders the vessel lumen became thickened as a result of dietary Hg (Figures 5a~t). Concomitantly, the Leydig cells, which com- prise the wall of the blood vessel and are believed to be sites for calcium and glycogen storage, became disrupted as envis- aged from their augmented size, reduced numbers and the appar- ent poor cellular integrity (Figures 5b-d).
Discussion
The digestive gland of soil gastropods is known to accumulate a variety of pollutant metals (Dallinger 1992). This organ also appears to be a major site for Hg accumulation in terrestrial
slugs, the granules of the digestive cells being the first accumu- lation site. These granules are of lysosomal nature and it has been demonstrated that a variety of metals are accumulated within them (Marig6mez et al. 1986a; 1990a; Recio e t aI.
1988a). Afterwards, Hg is also localized in the lipofuscine granules of the excretory cells but rarely has been observed in calcium cells. Mercury accumulation in digestive and excretory cells increases with dietary concentration of mercury and expo- sure time. A linear increase in Hg tissue levels is in agreement with the fact that Hg is probably bound to metallothionein-like Hg-binding proteins (Roesijadi 1982) which further may be engulfed within lysosomes for metal detoxification. However, some relevant exceptions in such accumulation pattern were recorded in slugs exposed to 1,000 Ixg Hg/g for 27-30 days where the histochemical levels of Hg decayed.
Interestingly, exposure to Hg resulted in an enhancement of excretory activity of digestive cells from which Hg is released within lysosomes or residual bodies by processes of apocrine secretion. Thus, it appears that Hg could be eliminated via the faeces as reported for several metals in different gastropods (Marigdmez e t al. 1986a; 1990a; Recio e t al. 1988a; Ireland and Marigdmez 1992; Nott et al. 1993). Consequently, the digestive epithelium seems to act as a detoxification pouch

Biomarkers of Hg Pollution in Sentinel Slugs 61
rather than an accumulating site. The present morphological observations also support this hypothesis.
MET values remained constant throughout the experimental period in control slugs. Previously, Marigdmez et al. (1986a) and Recio et al. (1988a) performed identical experiments and found a higher variability in MET values of control slugs and a decay after 2-3 weeks of maintenance under laboratory condi- tions. Since these former experiments were carried out between August to September when slugs are entering the female phase of their life cycle (South 1992), it is likely that during this critical period they were more susceptible to the stress due to laboratory maintenance than during July (male phase) when the present experiment was carried out.
Significant changes were recorded, however, in the quantita- tive structure of digestive tubules on exposure to dietary Hg. The reduction in MET as a result of exposure to Hg agrees with those observations reported in molluscs exposed to sublethal concentrations of metals (Marigdmez et al. 1986b; Vega et al.
1989; Ireland and Marig6mez 1992) and to other sources of environmental stress (Lowe et aI. 1981; Axiak et al. 1988; Cajaraville et al. 1992) and suggests that MET could be used as a tissue-level biomarker of environmental stress. Vega et al.
(1989), Soto et al. (1990) and Marigdmez et al. (1992) have reported, however, natural variations in MET that could produce inconsistent conclusions when MET is used as an index of the biological effect of pollutants. For this reason, Vega et al. (1989) proposed that MET should be used simultaneously with other measures of the tubule morphology. Thus, MDR and MLR values, as well as the ratios MLR/MET and MDRfMET have been used together with MET as complementary measures of digestive tubule morphology.
Reductions in MDR have been described in extreme cases of environmental stress in both gastropod and bivalve molluscs (Vega et al. 1989; Cajaraville et al. 1992; Marigdmez et al.
1993). In addition, some stages of inflammation associated with haemocyte proliferation have been related to high MDR values (Marig6mez et al. 1992; 1993) in marine gastropods. In the present work, MDR values did not change in relation to the Hg concentration in diet. Since this is the first time this measure has been made in terrestrial molluscs, we do not know whether it was not affected under the present experimental conditions or whether MDR is not sensitive in terrestrial gastropods due to the particular morphology of their digestive gland. Similarly, although increased MLR values have been found in environ- mentally stressed molluscs (Vega et al. 1989; Cajaraville et
al, 1992; Marig6mez et al. 1993), presently MLR values also remained unchanged with the concentration of dietary Hg. On the other hand, when changes in MET were related to either diverticular or luminal sizes, the attained MLR/MET and MET/MDR ratios were significantly affected by Hg concentra- tion in diet, exposure time and CxT interaction. The high sensi- tivity of these relative parameters has been previously reported by Vega et al. (1989), Cajaraville et al. (1992), Soto et al.
(1992), and Marigdmez et al. (1992; 1993). Changes in MET, MLR, MLR/MET, and MET/MDR can be
explained by regression models after logarithmic transformation of the data. Similar conclusions were obtained by Vega et al.
(1989) in marine gastropods exposed to cadmium. Thus, the existence of a significant regression model provides a good basis to further develop reliable biomarkers since we can establish the degree of association between the conditions of exposure and the changes measured in the biological system.
Finally, some changes were observed in the distribution and relative occurrence of cell types in the digestive epithelium. In a first step, excretory activity increased in digestive cells as evidenced by an enhanced apocrine secretion. Similar results were obtained in Cu-exposed slugs by Marigdmez et al.
(1986b). Afterwards, the relative numbers of digestive cells declined until the extreme cases of exposure to 1,000 Ixg Hg/g for 27 to 30 days in which the digestive epithelium is mostly comprised of calcium and excretory cells bearing large lipofuscine granules within their vacuoles. A similar process has been previously described in marine gastropods exposed to sublethal levels of cadmium (Marig6mez et al. 1990b). The raise in the relative numbers of calcium cell-like cells (the so- called basophilic cells) in stressed marine molluscs is a well- known phenomenon (Widdows et al. 1984; Lowe and Clarke 1989; Cajaraville et al. 1990a; 1990b). Whether this change in the cellular composition of the digestive epithelium is an adap- tive or pathological response is a question which deserves fur- ther research efforts. From a practical point of view, the changes in the cell type composition is an important issue since cell type replacement provokes a change in the accumulative capacity of the digestive gland as envisaged from the decay in the histo- chemical levels of Hg. On the other hand, autometallography of digestive gland sections can be useful for the screening of bioavailable metals but routine records on cell type composition and extent of secretory activity must be done to attain feasible conclusions as recommended by Soto (1995). The observation of some other changes in the histological organization of the digestive gland tissue may be also useful to elucidate how sublethal levels of dietary Hg (Marig6mez et al. 1986c) provoke a nonspecific stress response in terrestrial slugs. The thickening of blood vessel wall and the disruption of Leydig cells can be attributed to glycogen reserve consumption and obliteration of blood vessels. Similar effects have been previously described in terrestrial (Recio et al. 1988b; Marig6mez and Ireland 1992) and marine (Cajaraville et al. 1990; Marig6mez et al. 1990b; 1993) gastropods on exposure to a variety of environmental stressors. Therefore, it appears that Hg acts as an environmental stressor under the present experimental conditions instead of exerting direct toxic effects on cell systems of the slug diges- tive gland.
For all these reasons, it is suggested that slugs can be used in soil quality assessment in the manner that other molluscs are used in monitoring programs to assess water quality ("Mussel- Watch"; Bayne 1989). Therefore, slugs (Popham and D'Auria 1980) or equivalent terrestrial gastropods (Russell et al. 1981; Berger and Dallinger 1993) could be used as sentinels ("Slug- Watch"), which are examined to record biomarkers of exposure to pollutants and biomarkers of biological effect at various levels of biological organization including, of course, the cell and tissue levels (Widdows 1982; Cajaraville et al. 1993).
References
Axiak V, George JJ, Moore MN (t988) Petroleum hydrocarbons in the marine bivalve Venus verrucosa: accumulation and cellular responses. Mar Biol 97:225-230
Bayne BL (1989) Measuring the biological effects of pollution: the Mussel Watch approach. Wat Sci Technol 21:1089-1100
Berger B, Dallinger R (1993) Terrestrial snails as quantitative indicators of environmental metal pollution. Environ Monitor Assess 25:65-84

62 I. Marig6mez et al.
Cajaraville MR Diez G, Marig6mez I, Angulo E (1990a) Responses of basophilic cells of the digestive gland of mussels to petroleum hydrocarbon exposure. Dis Aquat Org 9:221-228
Cajaraville MR Marigdmez I, Angulo E (1993) Correlation between cellular and organismic responses to oil-induced environmental stress in mussels. Sci Tot Environ Supph1353-1371
Cajaraville MR Marig6mez I, Dfez G, Angulo E (1992) Comparative effects of the water accommodated fractions (WAF) of three oils on mussels. 2.--Quantitative alterations in the structure of the digestive tubules. Comp Biochem Physiol 102C: 1 1 3 - 1 2 3
Cajaraville MR Marigdmez JA, Angulo E (1990b) Short-term toxic effects of 1-naphthol on the digestive gland-gonad complex of the marine prosobranch Littorina littorea (L.): a light microscopic study. Arch Environ Contam Toxicol 19:17-24
Couch JA (1984) Atrophy of diverticular epithelium as an indicator of environmental irritants in the oyster, Crassostrea virginica. Mar Environ Res 14:525-526
Dallinger R (1992) Strategies of metal detoxification in terrestrial invertebrates. In: Dallinger R, Rainbow PS (eds) Ecoxoticology of metals in invertebrates. Lewis Publ., Boca Raton, FL, pp 245-289
Danscher G (1984) Autometallography. A new technique for light and electron microscopic visualization of metals in biological tissues (gold, silver, metal sulphides and metal selenides). Histochemis- try 81:331-335
Ireland MR Marig6mez I (1992) The influence of dietary calcium on the tissue distribution of Cu, Zn, Mn and P and histological changes in the digestive cells in the snail Achatinafulica. Bowdich. J Moll Stud 58:157-168
Lowe DM, Clarke KR (1989) Contaminant-induced changes in the structure of the digestive epithelium of Mytilus edulis. Aquat Tox- icol 15:345-358
Lowe DM, Moore MN, Clarke KR (1981) Effects of oil in the digestive ceils in mussels: quantitative alterations in cellular and lysosomal structure. Aquat Toxicol 1:213-226
Marig6mez I, Soto M, Angulo E (1992) Seasonal variability in the quantitative structure of the digestive tubules of Littorina littorea. Aquat Living Resour 2:299-305
Marig6mez I, Soto M, Etxeberria M, Angulo E (1993) Effects of size, sex, reproduction, and trematode infestation on the quantitative structure of digestive tubules in stressed winkles. Zool Jb Anat 123:319-336
Marig6mez JA, Angulo E, Moya J (1986a) Copper treatment of the digestive gland of the slug Arion ater L. 1. Bioassay conduction and histochemical analysis. Bull Environ Contain Toxicol 36:600-607
, , (1986b) Copper treatment of the digestive gland of the slug Arion ater L. 2. Morphometrics and histophysiol- ogy. Bull Environ Contain Toxicol 36:608-615
Marig6mez JA, Angulo E, Saez V (1986c) Feeding and growth re- sponses to copper, zinc, mercury and lead in the terrestrial gastro- pod Arion ater (Linn6). J Moll Stud 52:68-78
Marig6mez JA, Cajaraville MP, Angulo E (1990a) Cellular cadmium distribution in the common winkle, Littorina littorea (L.) deter- mined by X-ray microprobe analysis and histochemistry. Histo- chemistry 94:191-199
, , - - (1990b) Histopathology of the digestive gland- gonad complex of the marine prosobranch Littorina littorea ex- posed to cadmium. Dis Aquat Org 9:229-238
Martoja R, Martoja-Pierson M (1970) T6cnicas de histologfa animal. Toray-Mason, Barcelona, 350 pp
Minniti F (1987). Effects of copper pollution on the hepatopancreas of Cyclope neritea L. (Mollusca: Gastropoda). Zool Anz 219:141-146
Nott JA, Bebbiano MJ, Langston WJ, Ryan KP (1993) Cadmium in the gastropod Littorina littorea. J Mar Biol Assoc UK 73:655-665
Pearse AGE (1980) Histochemistry, theoretical and applied. Churchill Livingstone, Edinburgh, 439 pp
Popham JD, D'Auria (1980) Arion ater (Mollusca: Pulmonata) as an indicator of terrestrial environmental pollution. Water Air Soil Pollut 14:115-124
Recio A, Marig6mez JA, Angulo E, Moya J (1988a) Zinc treatment of the digestive gland of the slug Arion ater L. 1. Cellular distribu- tion of zinc and calcium. Bull Environ Contam Toxicol 41:858-864
, , , (1988b) Zinc treatment of the digestive gland of the slug Arion ater L. 2. Sublethal effects at the histological level. Bull Environ Contam Toxicol 41:865-871
Roesijadi G (1982) Uptake and incorporation of mercury into mercury- binding proteins of gills of Mytilus edulis as a function of time. Mar Biol 66:151-157
Russell LK, DeHaven JI, Botts RP (1981) Toxic effects of cadmium on the garden snail (Helix aspersa). Bull Environ Contain Tox- icoI 30:245-251
Sfinchez E (1993) Desarrollo de un sistema de amilisis plauim6trico automatizado con toma de datos en un lector de barrido digital: bases te6ricas y desarrollo tecnol6gico. Licenciature Thesis, Uni- versity of the Basque Country, Leioa (Basque Country), 50 pp.
Soto M (1995) Simultaneous quantification of bioavailable metals in molluscs by means of cellular and tissue analysis. Implications for monitoring metal pollution in water quality assessment. Euro- pean PhD Thesis, University of the Basque Country, Leioa (Basque Country), 330 pp
Soto M, Agirregoikoa MG, P6rez MA, Marigdmez I (1990) A planimet- ric study of morphological variability in the digestive diverticula of Littorina linorea (Linnaeus) and Mytilus edulis Linnaeus. J Moll Stud 56:339-344
South A (1992) Terrestrial slugs. Biology, ecology and control. Chap- man and Hall, London, 428 pp
Tripp MR, Fries CR, Craven MA, Grier CE (1984) Histopathology of Mercenaria mercenaria as an indicator of pollutant stress. Mar Environ Res 14:521-524
Vega MM, Marig6mez JA, Angulo E (1989) Quantitative alterations in the structure of the digestive cell of Littorina littorea on exposure to cadmium. Mar Biol 103:547-553
Widdows J (1982) Field measurements of the biological impacts of pollutants. In: Assimilative capacity of the oceans for man's wastes. SCOPE/ICSU Academia Sinica, Taipei, Pop. Rep. China, pp 111-129
Widdows J, Bakke T, Bayne BL, Donkin R Livingstone DR, Lowe DM, Moore MN, Evans SV, Moore SL (1984) Responses of Mytilus edulis on exposure to the WAF of North Sea oil. Mar Bio167:15-31