thymic hiv-2 infection uncovers posttranscriptional control of viral replication in human thymocytes
TRANSCRIPT

1
Thymic HIV-2 infection uncovers post-transcriptional control of viral replication in 1
human thymocytes 2
3
Helena Nunes-Cabaço#, Paula Matoso, Russell B. Foxall, Rita Tendeiro*, Ana R. Pires, Tânia 4
Carvalho, Ana I. Pinheiro, Rui S. Soares and Ana E. Sousa# 5
6
Instituto de Medicina Molecular, Faculdade de Medicina, Universidade de Lisboa, Lisbon, 7
Portugal. 8
9
Running Head: HIV-2 Infection of the Human Thymus 10
11
Word count (Abstract/Importance): 190/131 12
Word count (Text): 3566 13
14
#Address correspondence to: 15
Helena Nunes-Cabaço ([email protected]) 16
Ana E. Sousa ([email protected]) 17
Instituto de Medicina Molecular, Faculdade de Medicina da Universidade de Lisboa 18
Av. Prof. Egas Moniz, 1649-028 Lisboa, Portugal 19
Phone +351 217 999 525; Fax +351 217 999 527 20
21
22
*Present address: Rita Tendeiro, Division of Infection and Immunity, UCL Institute of Immunity 23
and Transplantation, Rowland Hill Street, London NW3 2PF, UK 24
25
JVI Accepts, published online ahead of print on 3 December 2014J. Virol. doi:10.1128/JVI.03047-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

2
ABSTRACT 26
A unique HIV-host equilibrium exists in untreated HIV-2-infected individuals, characterized by 27
low-to-undetectable viremia throughout the disease course despite the establishment of 28
disseminated HIV-2 reservoirs at levels comparable to those observed in untreated HIV-1 29
infection. Although the clinical spectrum is similar in the two infections, HIV-2 is associated 30
with a much slower rate of CD4 T-cell decline and limited impact on the mortality of infected 31
adults. Here we investigated HIV-2 infection of the human thymus, the primary organ for T-32
cell production. Human thymic tissue and suspensions of total or purified CD4 single-positive 33
thymocytes were infected with HIV-2 or HIV-1 primary isolates using either CCR5 or CXCR4 34
co-receptors. We found that HIV-2 infected both thymic organ cultures and thymocyte 35
suspensions, as attested by total HIV DNA and cell-associated viral mRNA levels. 36
Nevertheless, thymocytes featured reduced levels of intracellular Gag viral protein, 37
irrespective of HIV-2 co-receptor tropism and cell differentiation stage, in agreement with the 38
low viral load in culture supernatants. Our data show that HIV-2 is able to infect the human 39
thymus but leads to reduced viral production by thymocytes, providing a new model to identify 40
therapeutic targets for viral replication control. 41
42
IMPORTANCE 43
HIV-1 infects the thymus, leading to a decrease in CD4 T-cell production that contributes to 44
the characteristic CD4 T-cell loss. HIV-2 is associated with a very slow rate of progression to 45
AIDS, and is therefore considered a unique naturally-occurring model of attenuated HIV 46
disease. HIV-2-infected individuals feature low-to-undetectable plasma viral load, in spite of 47
the numbers of circulating infected T-cells being similar to those found in patients infected 48
with HIV-1. We assessed, for the first time, the direct impact of HIV-2 infection on the human 49
thymus. We showed that HIV-2 is able to infect the thymus, but that the HIV-2 replication 50
cycle in thymocytes is impaired. We propose that this system will be important to devise 51
immunotherapies that target viral production, aiding the design of future therapeutic strategies 52
for HIV control. 53
54
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

3
INTRODUCTION 55
The thymus is the primary organ for T-cell production and, despite the age-associated 56
decline, thymic function is maintained until late in life (1, 2). Thymic activity is vital in clinical 57
settings requiring de novo T-cell generation, such as HIV infection (1, 3, 4). Accordingly, 58
impairment of thymic output impacts on the rate of HIV-1 disease progression, whilst the 59
degree of immunological reconstitution achieved after antiretroviral therapy has been shown 60
to rely on thymus recovery (1, 3, 4). Moreover, a “functional” cure for HIV infection is thought 61
to entail a diverse T-cell repertoire, which can only be generated by the thymus. 62
HIV-1 targets the thymus in both children and adults, resulting in severe disruption of the 63
thymic microenvironment, as demonstrated by the morphological changes and thymocyte 64
depletion reported in thymuses of HIV-1-infected individuals (5, 6). Several studies, either 65
based on HIV-1 infection of the human thymus in vitro (7-9), or in vivo using the SCID/hu 66
mouse model (10, 11), have indicated that both direct infection of thymic cells and indirect 67
viral effects upon the microenvironment play a role in HIV-1-associated thymic pathology. 68
Furthermore, viral entry, replication kinetics and cytopathicity of HIV-1 in human thymocytes 69
have been shown to be highly dependent on viral tropism, due to the predominance of 70
CXCR4 (X4) vs. CCR5 (R5) expression in the human thymus (9, 12, 13). Thymic disruption 71
has also been described in non-human primate models of SIV infection (14). 72
Here we addressed, for the first time, the direct impact of HIV-2 infection on the human 73
thymus. This is particularly relevant because HIV-2-infected individuals feature slow rates of 74
CD4 T-cell decline and disease progression (15-17). Moreover, they typically have low-to-75
undetectable viremia, this being observed even in AIDS patients with <200 CD4 T cells/µl 76
(18, 19). The low levels of circulating virus account for the reduced horizontal and vertical 77
transmission observed in HIV-2 infection (20, 21), as well as for its geographical confinement 78
to West Africa and connected countries, such as Portugal. Despite the high prevalence of 79
HIV-2 in several regions of West Africa, such as in Guinea Bissau (8% in adults and up to 80
20% in people over 40 years of age) (22), there is no significant impact on the mortality of 81
infected adults. HIV-2 thus constitutes a unique naturally-occurring model of attenuated HIV 82
disease, valuable for the study of HIV pathogenesis. 83
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

4
In spite of the low-to-undetectable HIV-2 plasma viral load, HIV-2 and HIV-1-infected patients 84
at equivalent stages of CD4 T-cell depletion feature comparable levels of cell-associated viral 85
burden (18, 23, 24), indicating the establishment of disseminated HIV-2 reservoirs. They also 86
feature similar levels of T-cell activation (19), suggesting distinct control of viral replication in 87
the presence of cell activation in HIV-2 and HIV-1 infections. 88
Our previous data support preservation of thymic function in HIV-2 infection, as estimated by 89
signal-joint (sj)/TREC quantification (3) in circulating T-cells of HIV-2 infected patients (25). 90
However, there are no studies on the direct impact of HIV-2 infection on the human thymus, 91
either in vivo, due to the difficulty of obtaining thymic tissue from HIV-2-infected patients, or in 92
vitro. We show here that HIV-2 was able to infect the human thymus, but that this was 93
associated with limited viral replication, irrespectively of viral co-receptor tropism and 94
thymocyte differentiation stage. HIV-2 infection of the human thymus thus offers a novel 95
approach to investigate the mechanisms underlying the establishment of HIV reservoirs and 96
the control of viral replication. 97
98
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

5
MATERIALS AND METHODS 99
100
Ethical Statement 101
Thymic specimens (from newborns to 4-year-old children) were obtained from routine 102
thymectomy performed during pediatric corrective cardiac surgery at Hospital de Santa Cruz, 103
Carnaxide, Portugal, after parent’s written informed consent. Buffy coats from healthy donors 104
were provided by Instituto Português do Sangue e da Transplantação, after written informed 105
consent. The study was approved by the Ethical Boards of Faculty of Medicine of the 106
University of Lisbon and of Hospital de Santa Cruz, Portugal. 107
108
HIV stocks 109
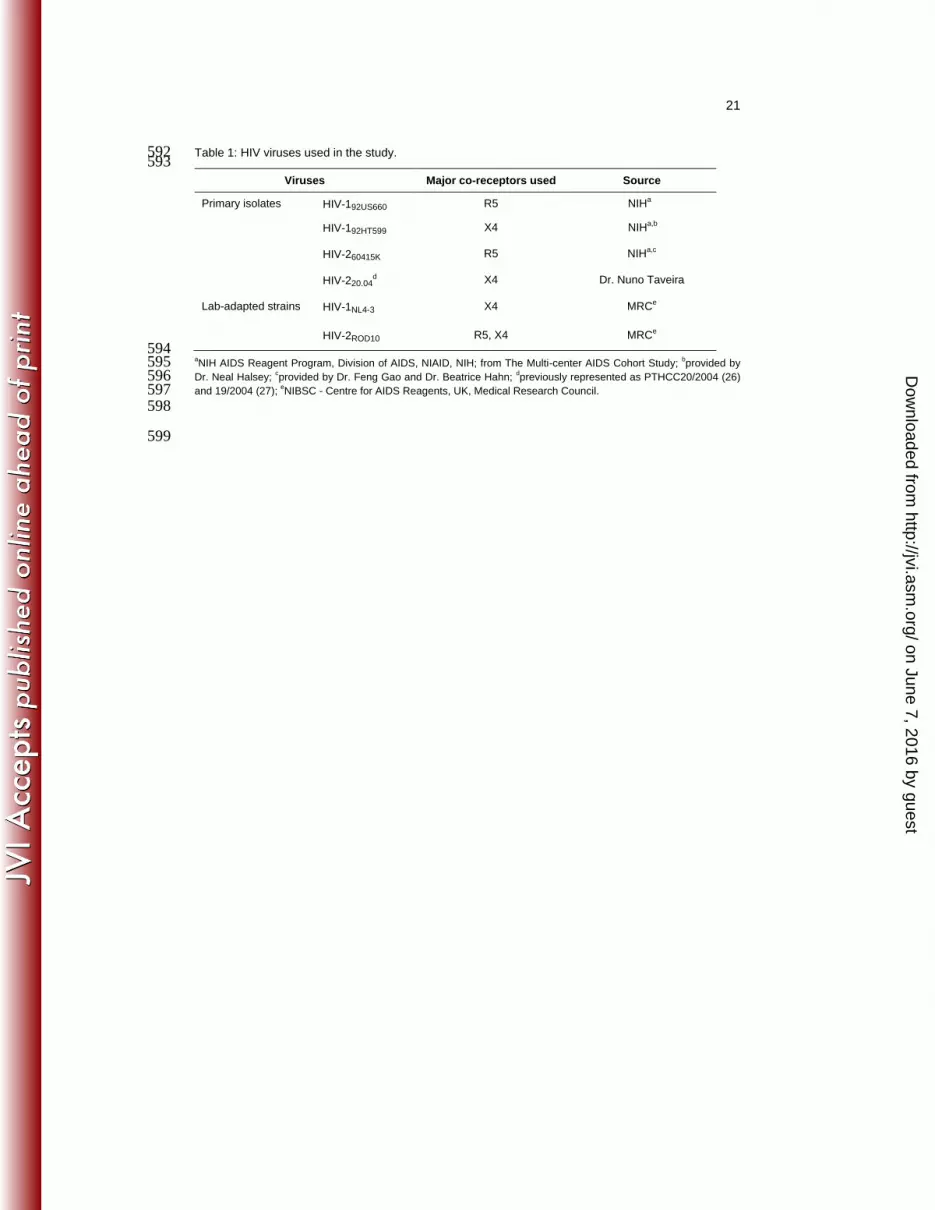
Viruses used are described in Table 1. Viral stocks were propagated in pools of isolated 110
peripheral blood mononuclear cells (PBMC) stimulated for 3 days with phytohemagglutinin 111
(PHA; 5μg/ml, Sigma) and maintained with human recombinant IL-2 (10U/ml; Dr. Maurice 112
Gately, Hoffmann – La Roche Inc, NIH AIDS Reagent Program), as described (28). Virus in 113
cell-free culture supernatants was quantified by measuring reverse transcriptase (RT) activity 114
using Lenti-RT Activity Kit (Cavidi). 115
116
HIV infection of thymocyte suspensions 117
Total thymocytes were recovered through tissue dispersion and separation on Ficoll-Paque 118
PLUS (GE Healthcare) density gradient. The CD4 single-positive (SP) population was sorted 119
from total thymocytes as CD3high
CD8neg
cells (purity>98%), using a FACSAria High-Speed 120
Cell Sorter (BD Biosciences). Viral stocks were ultra-centrifuged for 30min at 50,000g and 121
4ºC (Beckmann L8 Ultracentrifuge), resuspended in complete medium (RPMI1640 with 10% 122
fetal bovine serum (FBS), 2mM L-glutamine, 50U/ml penicillin/streptomycin and 50g/ml 123
gentamycin (all from GIBCO/Invitrogen) plus 3g/ml polybrene (Sigma)) and added to the 124
cells at 0.3ng RT/106 thymocytes. Total and CD4SP thymocytes were cultured for 3-4h at 10
8 125
and 3.5x107 cells/ml, respectively, in absence or presence of virus. After infection, cells were 126
washed and cultured at 107 cells/ml in complete medium supplemented with IL-2 (20U/ml), IL-127
4 (20ng/ml; R&D) and IL-7 (10ng/ml; R&D) at 37ºC. A quarter of the media was replaced 128
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

6
every 3-4 days. On day 10 thymocyte number per well was determined using 10M latex 129
beads (Coulter), and cells were analyzed by flow-cytometry or stored as pellets at -80ºC. Viral 130
production in supernatants was quantified at day 10 post-infection of 131
CD11cneg
CD14neg
CD123neg
TCRneg
thymocytes by measuring RT activity using SYBR 132
Green–product enhanced RT (SG-PERT), as described (29, 30). 133
134
HIV infection of Thymic Organ Cultures (TOC) 135
Thymic tissue blocks (1-2mm diameter) were placed on Millicell organotypic inserts (Millipore) 136
in a 6-well plate containing 1ml of TOC medium (complete medium with 15% FBS, 10mM 137
HEPES, 1mM sodium pyruvate and 1% MEM non-essential amino-acids, all from 138
GIBCO/Invitrogen). TOC were placed at 37ºC, 5%CO2 overnight and half of the medium 139
replaced prior to infection. HIV infection was performed by placing a 5l drop containing 3ng 140
RT of virus on top of each TOC. A third of the media was replaced every 2-3 days. On day 141
10/11 post-infection TOC were placed in 4% formaldehyde or mashed. Thymocytes were 142
analyzed by flow-cytometry or stored as cell pellets at -80ºC. The role of Env proteins was 143
assessed by culturing TOC for 7 days on Isopore membranes (Millipore) placed in TOC 144
medium containing 1g/ml of purified recombinant glycoproteins: gp105ROD (MRC EVA621), 145
gp120BaL (NIH Cat. No.4961) and gp120IIIB (MRC EVA607); or anti-CD4 mAb (BD 146
Biosciences) as control. 147
148
Flow-cytometry 149
Surface staining was performed for 20min at room temperature and always included Fixable 150
Viability Dye (eBioscience) for dead cell exclusion. Thymocytes were fixed, permeabilized and 151
stained using an intracellular staining kit (eBioscience), as described (31). Anti-human 152
monoclonal antibodies (mAb) used were (clones in brackets): CD3 (UCHT1), CD4 (RPA-T4), 153
CD8 (RPA-T8), CD11c (3.9), CD14 (61D3), CD16 (eBioCB16), CD19 (HIB19), CD123 (6H6) 154
and TCR(B1.1) from eBioscience. The following anti-Gag mAb were used: KC57 (Beckman 155
Coulter), anti-p24 (Kal-1; Dako), anti-p24 (AG3.0 or 183-H12-5C, NIH); anti-p27 156
(ARP396/397; Dr.Szawlowski, NIBSC-Centre for AIDS Reagents, Medical Research Council). 157
AlexaFluor®488 goat anti-Mouse IgG(H+L) antibody (Molecular Probes) was used for 158
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

7
secondary detection. Cells were acquired using LSRFortessa (BD Biosciences) and data was 159
analyzed in FlowJo (TreeStar). 160
161
Immunohistochemistry 162
Fixed TOC were embedded in paraffin and cut into 3μm sections (Minot Microtome Leica 163
RM2145). Epitope-retrieval was performed at pH9 (Leica Biosystems buffer) for 15min using 164
a microwave (800W). Samples were stained with the appropriate anti-Gag primary antibodies, 165
incubated with a peroxidase/DAB detection system (EnVision, Dako) and counterstained with 166
Harris’ hematoxylin (BioOptica). Images were acquired using a Leica DM2500 microscope. 167
168
Quantification of total HIV DNA and gag mRNA by real-time PCR 169
Total HIV DNA was quantified in cell lysates prepared by treating cell pellets with 100g/ml 170
proteinase K in 10mM Tris-HCl for 1h at 56ºC, followed by 10min enzyme inactivation at 171
95ºC. For gag mRNA quantification, 200ng of total RNA, purified using mirVana (Ambion), 172
were used to synthesize cDNA using oligo(dT)20 and SuperscriptIII (Invitrogen), according to 173
manufacturer’s instructions. Real-time PCR was performed using Platinum Taq plus ROX or 174
Taqman Gene Expression Master Mix (Applied Biosystems), with primers and probes 175
described in Supplementary Table 1. Standard curves were generated from serial dilutions of 176
cDNA prepared from mRNA of 3 different thymuses (for GAPDH quantification) or of plasmids 177
containing the amplicons of HIV-1 Gag and CD3 (kind gift from Rémi Cheynier) (32) or HIV-2 178
Gag. HIV-2 Gag plasmid was generated by inserting a sequence including LTR/Gag from 179
HIV-2ROD into the pGEM®-TEasy vector (Promega). Quantification was performed using an 180
Applied Biosystems 7500 Fast Real-Time PCR System. 181
182
Statistical Analysis 183
Statistical analysis was performed using GraphPad Prism v5.01 (GraphPad Software Inc.). 184
Two-sample data were compared using Wilcoxon-matched pairs test. Data from more than 185
two samples were compared using Friedman test or Kruskal-Wallis test with Dunn’s multiple 186
comparison post-test. P values <0.05 were considered significant. 187
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

8
RESULTS 188
189
HIV-2 infects the human thymus 190
We investigated HIV-2 infection of the human thymus in vitro using both TOC, which preserve 191
the thymic microenvironment and have been shown to be permissive to HIV-1 infection (7, 8), 192
and thymocyte suspensions. TOC and thymocyte suspensions were infected with HIV-2 or 193
HIV-1 primary isolates with selective R5 or X4 co-receptor specificity and cultured for 10 days. 194
RT activity was used to normalize viral input, as HIV-1 and HIV-2 RT enzymes were shown to 195
possess similar specific activities (33). 196
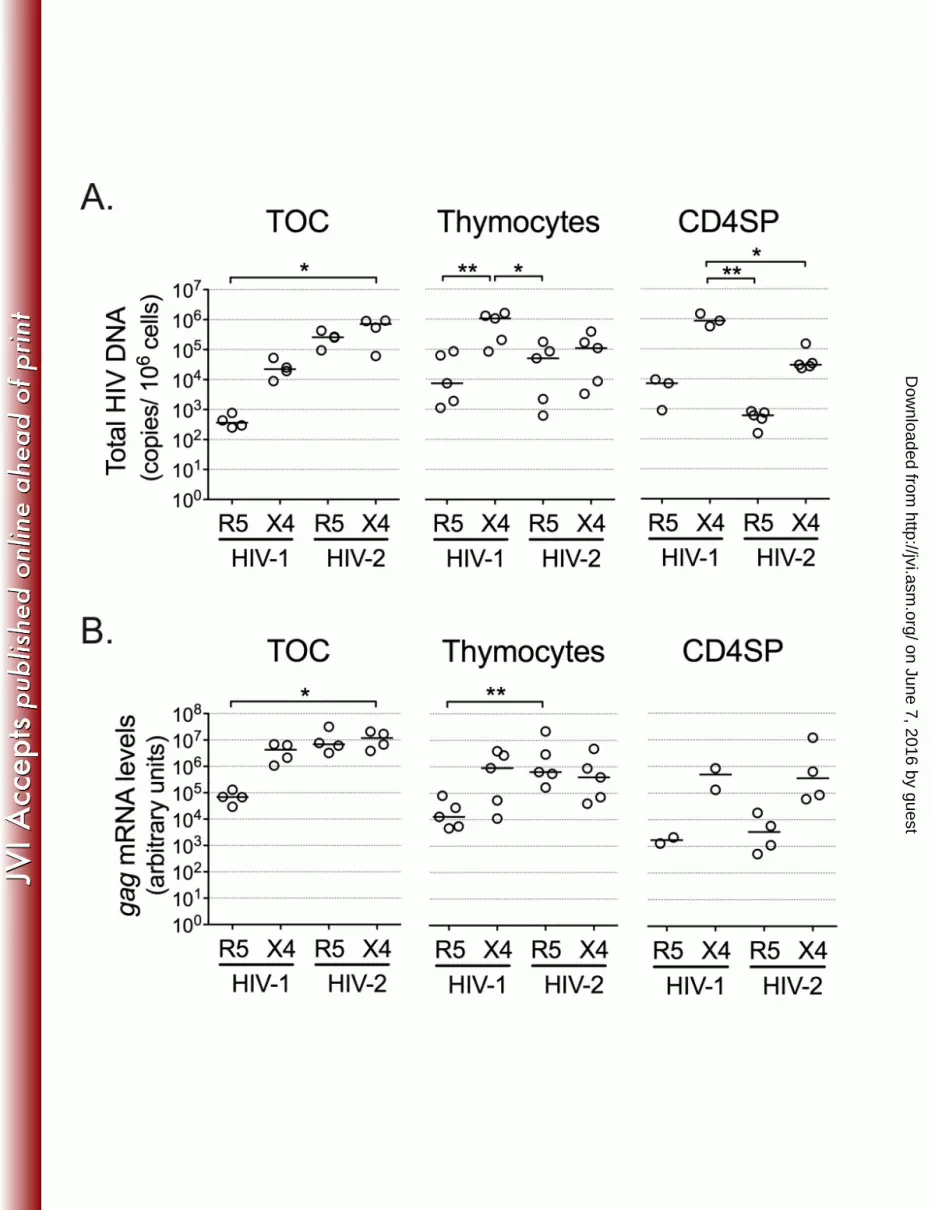
We found that HIV-2 was able to infect both thymic tissue and total thymocyte suspensions, 197
as indicated by the levels of total HIV DNA, which includes unintegrated and integrated 198
proviral DNA, in HIV-2-infected samples (Figure 1A). Furthermore, we observed that HIV-2 199
infection of both thymic tissue and total thymocyte suspensions was less co-receptor 200
dependent than HIV-1 infection. 201
Next we sort-purified mature CD4SP thymocytes, which constitute a target of HIV-1 (11), and 202
cultured them for 10 days after infection with the above-mentioned viruses. In contrast to TOC 203
and thymocyte suspensions, HIV-2 infection of CD4SP thymocytes resulted in co-receptor-204
dependent levels of total HIV DNA (Figure 1A). The limited ability of HIV-2 R5 to infect the 205
CD4SP subset was in contrast to our observations in TOC and total thymocyte suspensions 206
(Figure 1A), suggesting that HIV-2 R5 targeted other human thymocyte populations. 207
We next assessed the levels of ongoing viral transcription in HIV-2-infected TOC and 208
thymocytes by measuring viral gag mRNA. Both HIV-2 R5 and X4 isolates generated gag 209
mRNA levels comparable to those found upon infection with the HIV-1 X4 isolate (Figure 1B), 210
indicating efficient integration and transcription of HIV-2 in TOC and thymocyte cultures. Of 211
note, HIV-2 gag mRNA levels showed co-receptor dependency in the case of purified CD4SP 212
thymocytes, but not in TOC or thymocyte suspensions (Figure 1B), in line with the total HIV 213
DNA levels that were observed in the same samples. 214
In summary, our results indicated that HIV-2 was able to successfully enter, reverse 215
transcribe and integrate in human thymocytes in TOC and in suspension, and that viral 216
transcription was ongoing in these systems. 217
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

9
218
Limited viral production in HIV-2 infection of human thymocytes 219
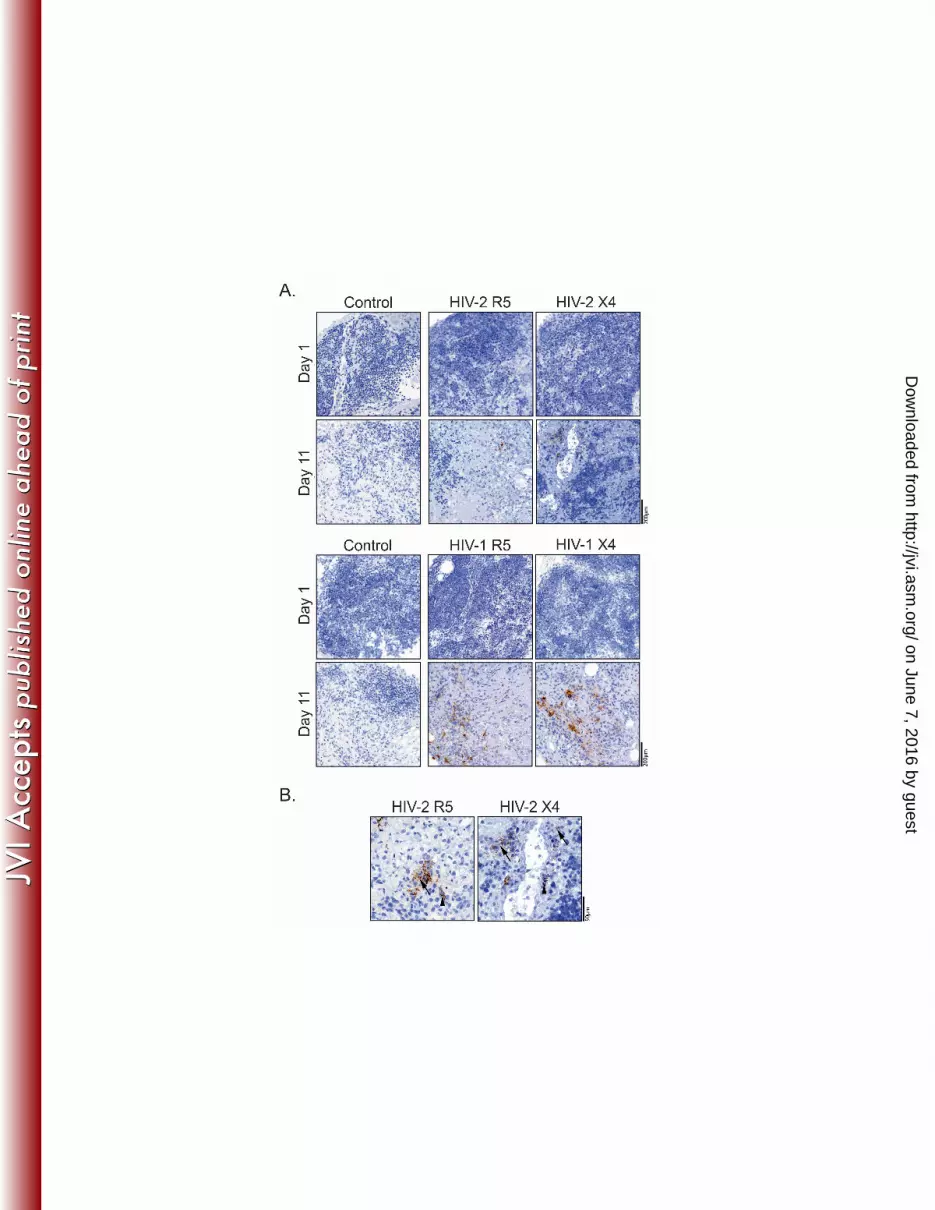
Next we assessed viral production in HIV-2-infected thymic tissue using 220
immunohistochemistry. Gag protein was detected in HIV-2 R5- and X4-infected TOC after 221
culture (Figure 2A), using validated antibodies (Supplementary Figure 1) (34). Gag protein 222
expression was both cell-associated and extracellular (Figure 2B), with the latter likely 223
corresponding to virus produced by HIV-infected cells during TOC culture, as no Gag staining 224
was observed on day 1 (Figure 2A). Of note, Gag protein levels were often indistinguishable 225
in X4 and R5 HIV-1 TOC infections (Figure 2A), despite the differences in the levels of cell-226
associated viral burden (Figure 1), indicating lack of correlation between the amount of virus 227
detected by immunohistochemistry and the levels of thymocyte viral production. Therefore, 228
although our immunohistochemistry data confirmed the ability of HIV-2 to productively infect 229
the human thymus, it was not possible to infer the specific contribution of thymocytes vs. 230
thymic stroma to the viral protein detected in the histological analysis. 231
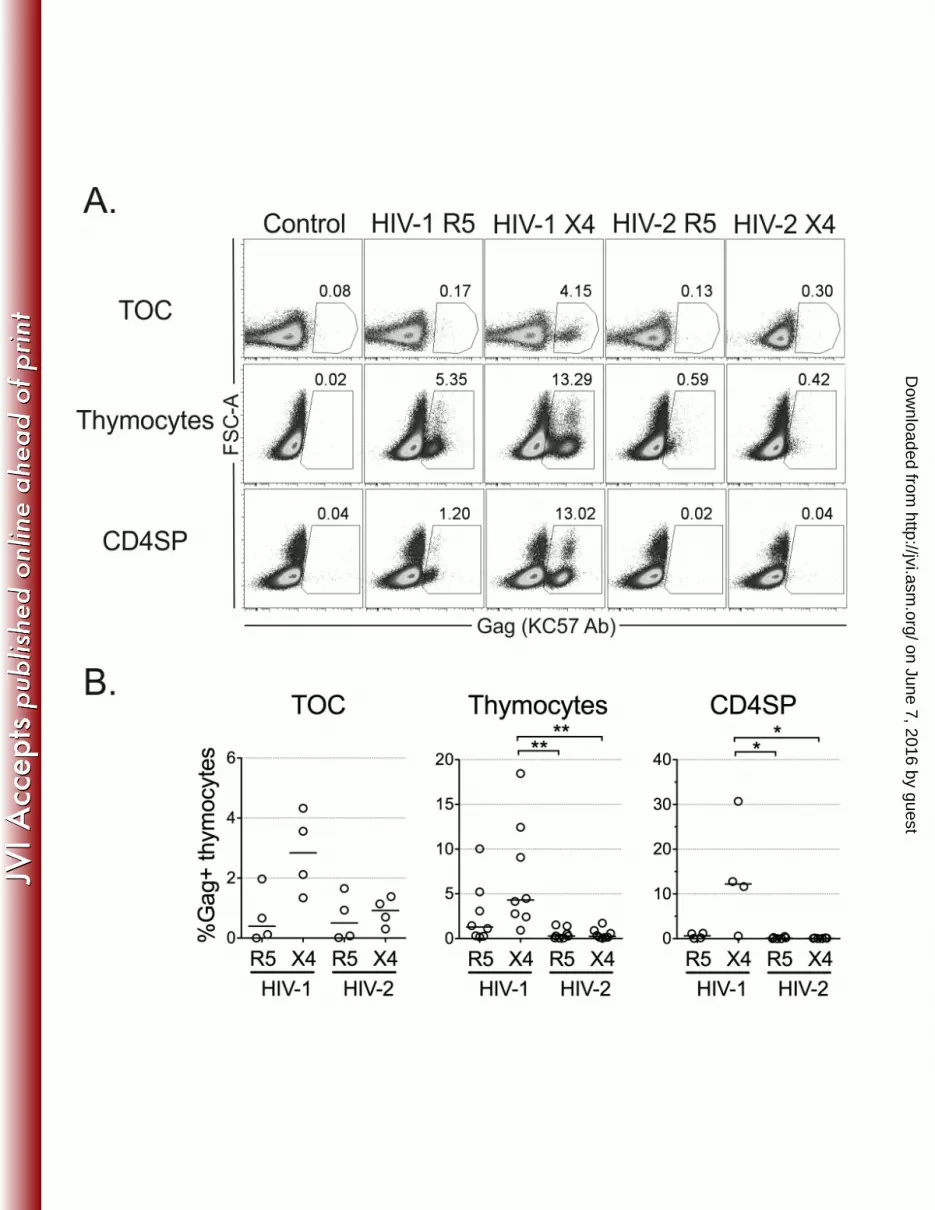
We thus evaluated viral production at the single-cell level by flow-cytometry. Notably, we 232
found very low levels of intracellular Gag protein in cells isolated from HIV-2-infected TOC or 233
in thymocyte suspensions, irrespective of the virus used (Figure 3) and time-point analyzed 234
(Supplementary Figure 2). In contrast, Gag protein expression was clearly detected in cells 235
infected with HIV-1, particularly with the X4 isolate (Figure 3), excluding the possibility that the 236
low Gag detection in HIV-2 infections was due to sample processing. Of note, intracellular 237
Gag protein was also barely detected upon thymocyte infection with the lab-adapted strain 238
HIV-2ROD (Supplementary Figure 2), despite the high levels of total HIV DNA detected 239
(181,499copies/106 cells in the example presented). Low Gag levels upon HIV-2 infection 240
were particularly evident in CD4SP thymocytes, which featured extremely low levels of 241
intracellular Gag after infection with HIV-2 primary isolates or HIV-2ROD (Figure 3 and 242
Supplementary Figure 2). 243
The antibody used (KC57) has been previously shown to be specific for both HIV-2 and HIV-1 244
Gag (35). We further confirmed the ability of the antibody to efficiently detect HIV-2 Gag by 245
flow-cytometry in activated PBMCs infected with the HIV-2 strains used in the current study 246
(Supplementary Figure 3). Moreover, KC57 staining was comparable to that obtained with 247
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

10
other available anti-Gag antibodies in HIV-2-infected PBMCs or thymocytes (Supplementary 248
Figures 2 and 3), confirming low viral production in HIV-2-infected thymocytes. 249
The low levels of intracellular Gag protein translated into reduced viral production in HIV-2 250
infections, as indicated by the lower levels of virus in the supernatant of HIV-2-infected 251
thymocyte cultures than in their HIV-1-infected counterparts (RT activity: HIV-1 R5: 252
5.97±3.23pg/ml; HIV-1 X4: 1318±583.5pg/ml; HIV-2 R5: 0.31±0.15pg/ml; HIV-2 X4: 253
8.54±3.40pg/ml; n=3). 254
Overall, we found that HIV-2 infection of human thymocytes occurred without significant viral 255
production per cell, despite evidence of cell-associated viral burden and active ongoing viral 256
transcription. 257
258
Impact of HIV-2 infection on thymocyte populations 259
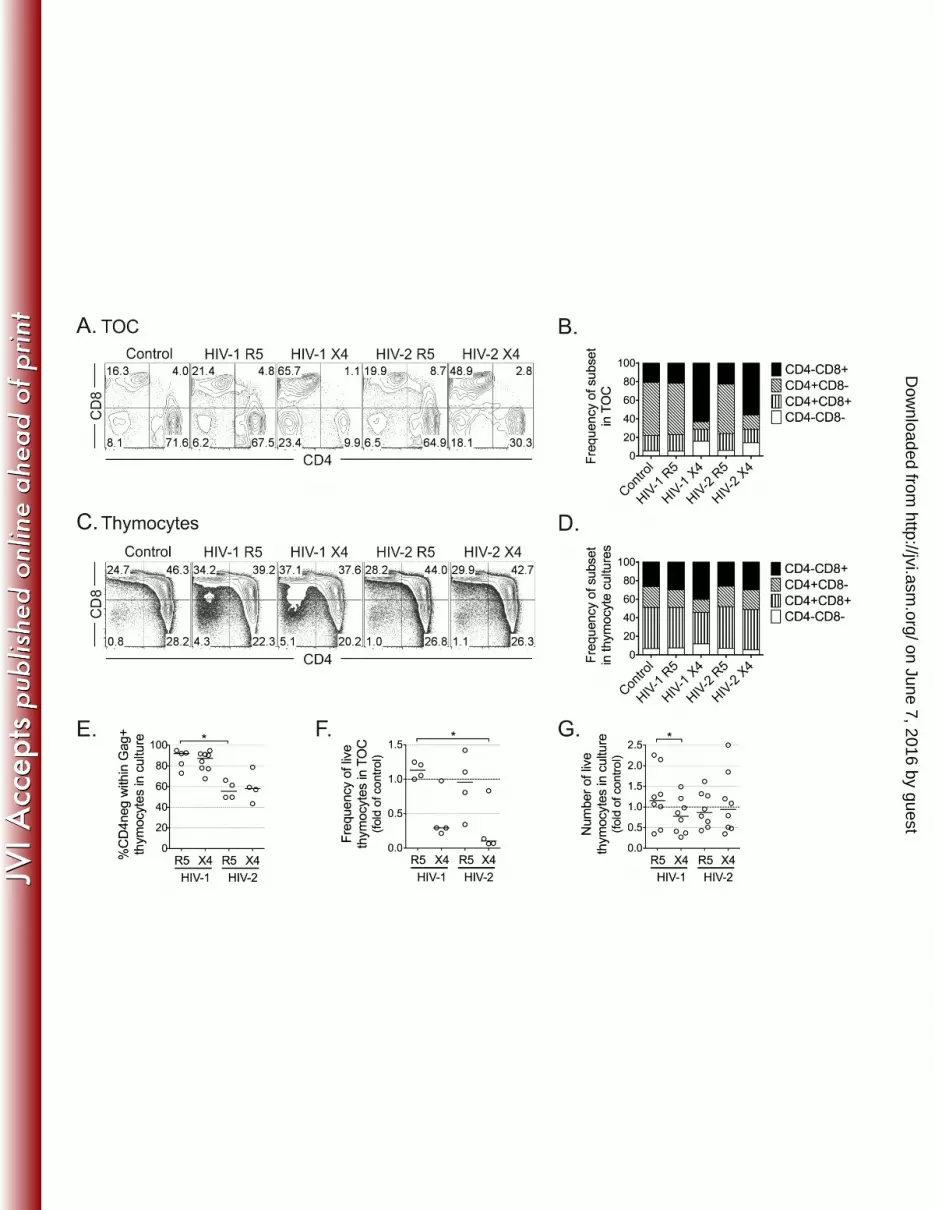
Finally, we assessed the impact of HIV-2 on the distribution of thymocyte populations in 260
infected TOC and thymocyte suspensions. TOC infected with X4-using viruses, whether HIV-261
2 or HIV-1, featured a dramatic decrease in CD4+ thymocytes as compared to R5 viruses 262
(Figures 4A and 4B). This was also observed in thymocyte suspensions infected with HIV-1 263
X4, but not with HIV-2, which did not result in significant alterations of the thymocyte 264
phenotype in culture (Figures 4C and 4D). One possible explanation for these results is that 265
down-regulation of the CD4 co-receptor is distinctly induced by HIV-2 and HIV-1. Accordingly, 266
our analysis of productively infected cells indicated that HIV-2 was less efficient than HIV-1 at 267
down-regulating CD4 (Figure 4E). 268
The effects on subset distribution were mirrored by the cytopathic effect of the respective 269
virus: both HIV-2 and HIV-1 X4 but not R5 viruses were cytopathic in TOC (Figure 4F), whilst 270
HIV-1 X4 had a small effect on cell viability in thymocyte suspensions (Figure 4G). The low 271
cell recovery in HIV-2 X4-infected TOC, together with the imbalance in cell subsets and the 272
apparent low impact of HIV-2 infection on CD4 down-regulation, suggested that HIV-2 X4 273
infection of TOC led, directly or indirectly, to the death of CD4+ thymocytes. This occurred 274
despite the lack of impact of HIV-2 X4 infection on total thymic cells suspensions, supporting 275
the contribution of other cell populations present in the whole tissue. 276
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

11
Given our previous data supporting immunosuppressive properties of the HIV-2 envelope 277
proteins in peripheral blood lymphocytes mediated, at least in part, by monocytes (36-38), we 278
also evaluated whether the alterations observed in HIV-2 X4-infected TOC were induced by 279
direct action of viral envelope proteins. We cultured TOC in the presence of recombinant HIV-280
2 and HIV-1 Env proteins, either binding to X4 (HIV-2 gp105ROD and HIV-1 gp120IIIB) or R5 281
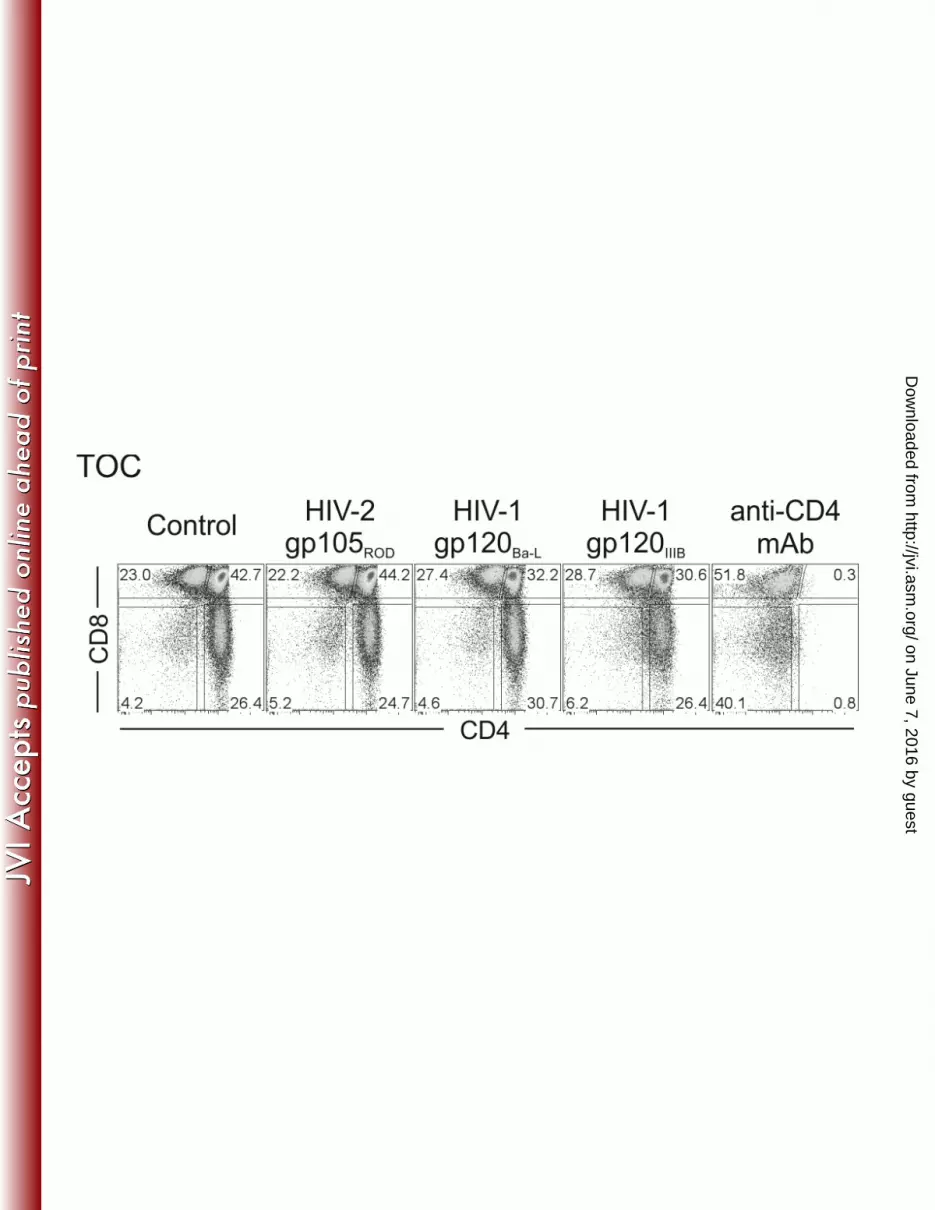
(HIV-1 gp120Ba-L). In our system, Env proteins did not induce CD4+ thymocyte depletion or 282
subset imbalances (Figure 5), indicating that the effects observed may require viral entry 283
and/or post-entry steps. 284
Altogether, our data support a cytopathic effect of HIV-2 X4 on the human thymus that was 285
not observed in thymocyte suspensions, suggesting a dependency on the stromal 286
compartment. Importantly, HIV-2 R5 had no significant cytopathic effect on either TOC or 287
thymocyte suspensions. 288
289
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

12
DISCUSSION 290
We addressed here, for the first time, the ability of HIV-2 to infect the human thymus. We 291
showed that HIV-2 is able to infect thymic tissue and thymocyte suspensions in the absence 292
of exogenous stimulation, although with a very low level of viral production per thymocyte, 293
supporting the existence of post-transcriptional control in viral replication. 294
HIV-2 infection of the human thymus was confirmed by the levels of cell-associated HIV DNA 295
and gag mRNA measured following HIV-2 infection of either TOC or thymocyte suspensions. 296
The high levels of total HIV DNA observed were consistent with previous reports where 297
unintegrated plus integrated proviral DNA were quantified (39, 40). HIV-1 infection of human 298
thymocytes was shown to be co-receptor-dependent (9, 12), and our data recapitulated these 299
results. Conversely, we showed that HIV-2 infection of TOC and thymocyte suspensions was 300
much less dependent than HIV-1 on co-receptor tropism, both in terms of cell-associated HIV 301
DNA and viral RNA, which could be related to the broader co-receptor usage reported for 302
HIV-2 as compared to HIV-1 (41). 303
On the other hand, HIV-2 cytopathicity in human thymic tissue was co-receptor-dependent, 304
with X4 virus inducing significant T-cell death in TOC, which was not observed for HIV-2 R5 305
virus. This was in line with a previous study of HIV-2 infection of lymphoid tissue, where 306
lymphocyte depletion also occurred with X4 but not R5-tropic isolates (42). Importantly, we 307
showed that HIV-2 X4 cytopathicity in TOC was not due to the direct action of HIV-2 Env, 308
indicating a requirement for viral entry and/or post-entry steps for CD4+ thymocyte depletion 309
in the human thymus. 310
The observed cytopathicity caused by HIV-2 X4 in tissue but not in thymocyte suspensions 311
could be related to the infection of components of the thymic stroma that are present in TOC. 312
Thymic stromal cells, including DCs, macrophages, and even thymic epithelial cells, have 313
been reported to be permissive to HIV-1 infection (43-45). It is not known whether HIV-2 314
infects the thymic stroma and, due to alterations in tissue morphology that occurred during the 315
culture process, we were not able to directly infer on the type of infected cells from our 316
immunohistochemistry data. However, in contrast to HIV-1, HIV-2 encodes the lentiviral 317
accessory protein Vpx, which targets the restriction factor sterile alpha motif and 318
histidine/aspartic acid domain–containing protein 1 (SAMHD1) for degradation, a factor that 319
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

13
has been shown to limit the productive infection of HIV-1 in myeloid cells (46). Moreover, 320
additional cell targets may be considered, given the broader co-receptor usage reported for 321
HIV-2 (41). We are currently addressing the possibility that infection of thymic stroma 322
components might have distinct consequences in HIV-1 and HIV-2 infections. 323
We had previously reported that thymic function, estimated via sj/βTREC measurement, was 324
better preserved in HIV-2 as compared to HIV-1-infected patients (25). It is likely that most of 325
these individuals were infected with R5-tropic HIV-2 (25). Importantly, we showed here that 326
HIV-2 R5 infection did not significantly impact on TOC or thymocyte suspensions, in terms of 327
cytopathicity or subset distribution, or induce CD4 down-regulation in the latter. Nevertheless, 328
the elevated levels of total HIV DNA and viral mRNA support the potential establishment of 329
HIV-2 viral reservoirs in human thymocytes. It would thus be important to investigate whether, 330
as reported for HIV-1 (32), CD4+ recent thymic emigrant T-cells from HIV-2-infected patients 331
may constitute viral reservoirs. 332
Our data suggest that, in our system, distinct regulation of the replicative cycle of HIV-2 and 333
HIV-1 occurs at the post-transcriptional/translational level. The discrepant levels of Gag 334
protein in the two infections markedly contrasted with the similarly high levels of total HIV 335
DNA and viral mRNA documented. HIV-2 and HIV-1 have been reported to differ significantly 336
in their mechanisms of translation initiation (47). For instance, HIV-2 was described to have 337
lower translational efficiency than HIV-1 due to differences in the 5’-untranslated region (5’-338
UTR) of viral genomic RNA (48, 49). In agreement with our observations, lower Gag protein 339
production in the presence of equivalent T-cell-associated viral gag mRNA levels were 340
reported in HIV-2 as compared to HIV-1 infection of T-cell lines and macrophages (49). 341
Importantly, and in relation to our data, this was not due to lower stability of the Gag protein, 342
nor to higher rate of viral particle release (49). Interestingly, the authors raised the possibility 343
that host factors may be differentially regulating the initiation of Gag translation in HIV-2 and 344
HIV-1 (49). HIV-2 infection of the human thymus provides, to our knowledge, the first model of 345
HIV-2 post-transcriptional regulation in ex vivo T-cells using primary isolates, providing a 346
unique opportunity to study the molecular factors and mechanisms involved in the regulation 347
of HIV translation. 348
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

14
Human thymocytes represent an ideal system for the study of HIV latency due to their ability 349
to become infected in the absence of external activation (50). Models using peripheral CD4+ 350
T-cells require cellular activation followed by quiescence induction (51), thus inducing 351
molecular modifications that may impact the processes under study. The system described 352
here, utilizing HIV-2 infection of the human thymus in the absence of exogenous stimulation, 353
thus provides an important cellular model for the study of latency and reservoir generation in 354
HIV pathogenesis. 355
Our results regarding HIV-2 infection also highlight the potential importance of post-356
transcriptional control of viral production, possibly through regulation of translation, in viral 357
pathogenesis. Further studies using this model will enable the discovery of potential 358
molecular targets to be used as the basis for new immunotherapies aimed at achieving a 359
“functional” cure for HIV. 360
361
362
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

15
ACKNOWLEDGEMENTS 363
This work was supported by “Fundação para a Ciência e a Tecnologia” (FCT) and by 364
“Programa Operacional Ciência e Inovação 2010” (POCI2010), grant PTDC/SAU-365
MII/66248/2006 to AES. HNC, RBF, RT and RSS received scholarships from FCT co-366
financed by POCI2010. 367
The authors thank Dr. Miguel Abecasis and Dr. Rui Anjos for human thymus sample 368
collection; Dr. Rémi Cheynier, Dr. Andreia Amaral and Dr. Íris Caramalho for discussion; Dr. 369
Francisca Matos and nurses and staff at Hospital de Santa Cruz for technical assistance; Dr. 370
Nuno Taveira, Dr. Rémi Cheynier and Dr. Nicolas Manel for reagents; patients and families. 371
372
373
CONFLICT OF INTERESTS 374
The authors declare no conflicting financial interests. 375
376
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

16
REFERENCES 377
1. Douek DC, McFarland RD, Keiser PH, Gage EA, Massey JM, Haynes BF, Polis 378 MA, Haase AT, Feinberg MB, Sullivan JL, Jamieson BD, Zack JA, Picker LJ, 379 Koup RA. 1998. Changes in thymic function with age and during the treatment of HIV 380 infection. Nature 396:690–695. 381
2. Nunes-Cabaço H, Sousa AE. 2013. Repairing thymic function. Curr. Opin. Organ 382 Transplant. 18:363–368. 383
3. Dion ML, Poulin JF, Bordi R, Sylvestre M, Corsini R, Kettaf N, Dalloul A, 384 Boulassel MR, Debre P, Routy JP, Grossman Z, Sekaly RP, Cheynier R. 2004. 385 HIV infection rapidly induces and maintains a substantial suppression of thymocyte 386 proliferation. Immunity 21:757–768. 387
4. Li T, Wu N, Dai Y, Qiu Z, Han Y, Xie J, Zhu T, Li Y. 2011. Reduced Thymic Output Is 388 a Major Mechanism of Immune Reconstitution Failure in HIV-Infected Patients After 389 Long-term Antiretroviral Therapy. Clin. Infect. Dis. 53:944–951. 390
5. Haynes BF, Hale LP, Weinhold KJ, Patel DD, Liao H-X, Bressler PB, Jones DM, 391 Demarest JF, Gebhard-Mitchell K, Haase AT, Bartlett JA. 1999. Analysis of the 392 adult thymus in reconstitution of T lymphocytes in HIV-1 infection. J. Clin. Invest. 393 103:453–460. 394
6. Joshi VV, Oleske JM, Saad S, Gadol C, Connor E, Bobila R, Minnefor, AB. 2012. 395 Thymus Biopsy in Children with Acquired-Immunodeficiency-Syndrome. Arch. Pathol. 396 Lab. Med. 110:837–842. 397
7. Bonyhadi ML, Su L, Auten J, McCune JM, Kaneshima H. 1995. Development of a 398 human thymic organ culture model for the study of HIV pathogenesis. AIDS Res. Hum. 399 Retroviruses 11:1073–1080. 400
8. Choudhary SK, Choudhary NR, Kimbrell KC, Colasanti J, Ziogas A, Kwa D, 401 Schuitemaker H, Camerini D. 2004. R5 Human Immunodeficiency Virus Type 1 402 Infection of Fetal Thymic Organ Culture Induces Cytokine and CCR5 Expression. J. 403 Virol. 79:458–471. 404
9. Gurney KB, Uittenbogaart CH. 2006. Human Immunodeficiency Virus Persistence 405 and Production in T-Cell Development. Clin. Vaccine Immunol. 13:1237–1245. 406
10. Bonyhadi ML, Rabin L, Salimi S, Brown DA, Kosek J, McCune JM, Kaneshima H. 407 1993. HIV induces thymus depletion in vivo. Nature 363:728–732. 408
11. Stanley SK, McCune JM, Kaneshima H, Justement JS, Sullivan M, Boone E, 409 Baseler M, Adelsberger J, Bonyhadi M, Orenstein J. 1993. Human 410 immunodeficiency virus infection of the human thymus and disruption of the thymic 411 microenvironment in the SCID-hu mouse. J. Exp. Med. 178:1151–1163. 412
12. Pedroza-Martins L, Gurney KB, Torbett BE, Uittenbogaart CH. 1998. Differential 413 tropism and replication kinetics of human immunodeficiency virus type 1 isolates in 414 thymocytes: coreceptor expression allows viral entry, but productive infection of 415 distinct subsets is determined at the postentry level. J. Virol. 72:9441–9452. 416
13. Berkowitz RD, Alexander S, Bare C, Linquist-Stepps V, Bogan M, Moreno ME, 417 Gibson L, Wieder ED, Kosek J, Stoddart CA, McCune JM. 1998. CCR5- and 418 CXCR4-utilizing strains of human immunodeficiency virus type 1 exhibit differential 419 tropism and pathogenesis in vivo. J. Virol. 72:10108–10117. 420
14. Dutrieux J, Fabre-Mersseman V, Charmeteau-De Muylder B, Rancez M, Ponte R, 421 Rozlan S, Figueiredo-Morgado S, Bernard A, Beq S, Couëdel-Courteille A, 422 Cheynier R. 2014. Modified interferon-α subtypes production and chemokine 423 networks in the thymus during acute simian immunodeficiency virus infection, impact 424 on thymopoiesis. AIDS 28:1101–1113. 425
15. Marlink R, Kanki P, Thior I, Travers K, Eisen G, Siby T, Traore I, Hsieh CC, Dia 426 MC, Gueye EH. 1994. Reduced rate of disease development after HIV-2 infection as 427 compared to HIV-1. Science 265:1587–1590. 428
16. Poulsen AG, Aaby P, Larsen O, Jensen H, Nauclér A, Lisse IM, Christiansen CB, 429 Dias F, Melbye M. 1997. 9-year HIV-2-associated mortality in an urban community in 430 Bissau, west Africa. The Lancet 349:911–914. 431
17. Clavel F, Mansinho K, Chamaret S, Guetard D, Favier V, Nina J, Santos-Ferreira 432 MO, Champalimaud JL, Montagnier L. 1987. Human immunodeficiency virus type 2 433 infection associated with AIDS in West Africa. N. Engl. J. Med. 316:1180–1185. 434
18. Popper SJ, Sarr AD, Gueye-NDiaye A, Mboup S, Essex ME, Kanki PJ. 2000. Low 435
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

17
plasma human immunodeficiency virus type 2 viral load is independent of proviral 436 load: low virus production in vivo. J. Virol. 74:1554–1557. 437
19. Sousa AE, Carneiro J, Meier-Schellersheim M, Grossman Z, Victorino R. 2002. 438 CD4 T cell depletion is linked directly to immune activation in the pathogenesis of HIV-439 1 and HIV-2 but only indirectly to the viral load. J. Immunol. 169:3400–3406. 440
20. Kanki PJ, Travers KU, Mboup S, Hsieh CC, Marlink RG, Gueye-NDiaye A, Siby T, 441 Thior I, Hernandez-Avila M, Sankalé JL. 1994. Slower heterosexual spread of HIV-2 442 than HIV-1. Lancet 343:943–946. 443
21. O'Donovan D, Ariyoshi K, Milligan P, Ota M, Yamuah L, Sarge-Njie R, Whittle H. 444 2000. Maternal plasma viral RNA levels determine marked differences in mother-to-445 child transmission rates of HIV-1 and HIV-2 in The Gambia. MRC/Gambia 446 Government/University College London Medical School working group on mother-child 447 transmission of HIV. AIDS 14:441–448. 448
22. Poulsen AG, Aaby P, Gottschau A, Kvinesdal BB, Dias F, Mølbak K, Lauritzen E. 449 1993. HIV-2 infection in Bissau, West Africa, 1987-1989: incidence, prevalences, and 450 routes of transmission. J. Acquir. Immune Defic. Syndr. 6:941–948. 451
23. Soares R, Foxall R, Albuquerque A, Cortesao C, Garcia M, Victorino RMM, Sousa 452 AE. 2006. Increased Frequency of Circulating CCR5+ CD4+ T Cells in Human 453 Immunodeficiency Virus Type 2 Infection. J. Virol. 80:12425–12429. 454
24. Soares RS, Tendeiro R, Foxall RB, Baptista AP, Cavaleiro R, Gomes P, Camacho 455 R, Valadas E, Doroana M, Lucas M, Antunes F, Victorino RMM, Sousa AE. 2011. 456 Cell-Associated Viral Burden Provides Evidence of Ongoing Viral Replication in 457 Aviremic HIV-2-Infected Patients. J. Virol. 85:2429–2438. 458
25. Gautier D, Beq S, Cortesao CS, Sousa AE, Cheynier R. 2007. Efficient 459 Thymopoiesis Contributes to the Maintenance of Peripheral CD4 T Cells during 460 Chronic Human Immunodeficiency Virus Type 2 Infection. J. Virol. 81:12685–12688. 461
26. Borrego P, Marcelino J, Rocha C, Doroana M, Antunes F, Maltez F, Gomes P, 462 Novo C, Barroso H, Taveira N. 2008. The role of the humoral immune response in 463 the molecular evolution of the envelope C2, V3 and C3 regions in chronically HIV-2 464 infected patients. Retrovirology 5:78. 465
27. Marcelino JM, Borrego P, Nilsson C, Família C, Barroso H, Maltez F, Doroana M, 466 Antunes F, Quintas A, Taveira N. 2012. Resistance to antibody neutralization in HIV-467 2 infection occurs in late stage disease and is associated with X4 tropism. AIDS 468 26:2275–2284. 469
28. Soares RS, Matoso P, Calado M, Sousa AE. 2011. Strategies to quantify unspliced 470 and multiply spliced mRNA expression in HIV-2 infection. J. Virol. Methods 175:38–45. 471
29. Pizzato M, Erlwein O, Bonsall D, Kaye S, Muir D, McClure MO. 2009. A one-step 472 SYBR Green I-based product-enhanced reverse transcriptase assay for the 473 quantitation of retroviruses in cell culture supernatants. J. Virol. Methods 156:1–7. 474
30. Vermeire J, Naessens E, Vanderstraeten H, Landi A, Iannucci V, Van Nuffel A, 475 Taghon T, Pizzato M, Verhasselt B. 2012. Quantification of Reverse Transcriptase 476 Activity by Real-Time PCR as a Fast and Accurate Method for Titration of HIV, Lenti- 477 and Retroviral Vectors. PLoS ONE 7:e50859. 478
31. Nunes-Cabaço H, Caramalho Í, Sepúlveda N, Sousa AE. 2011. Differentiation of 479 human thymic regulatory T cells at the double positive stage. Eur. J. Immunol. 480 41:3604–3614. 481
32. Fabre-Mersseman V, Dutrieux J, Louise A, Rozlan S, Lamine A, Parker R, Rancez 482 M, Nunes-Cabaço H, Sousa AE, Lambotte O, Cheynier R. 2011. CD4+ recent 483 thymic emigrants are infected by HIV in vivo, implication for pathogenesis. AIDS 484 25:1153–1162. 485
33. Hizi A, Tal R, Shaharabany M, Loya S. 1991. Catalytic properties of the reverse 486 transcriptases of human immunodeficiency viruses type 1 and type 2. J. Biol. Chem. 487 266:6230–6239. 488
34. Fernandes SM, Pires AR, Ferreira C, Tendeiro R, Correia L, Paulo SE, Victorino 489 RMM, Sousa AE. 2014. Gut disruption in HIV-2 infection despite reduced viremia. 490 AIDS 28:290–292. 491
35. Duvall MG, Lore K, Blaak H, Ambrozak DA, Adams WC, Santos K, Geldmacher C, 492 Mascola JR, McMichael AJ, Jaye A, Whittle HC, Rowland-Jones SL, Koup RA. 493 2007. Dendritic Cells Are Less Susceptible to Human Immunodeficiency Virus Type 2 494 (HIV-2) Infection than to HIV-1 Infection. J. Virol. 81:13486–13498. 495
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

18
36. Cavaleiro R, Sousa AE, Loureiro A, Victorino RMM. 2000. Marked 496 immunosuppressive effects of the HIV-2 envelope protein in spite of the lower HIV-2 497 pathogenicity. AIDS 14:2679–2686. 498
37. Cavaleiro R, Brunn GJ, Albuquerque AS, Victorino RMM, Platt JL, Sousa AE. 499 2007. Monocyte-mediated T cell suppression by HIV-2 envelope proteins. Eur. J. 500 Immunol. 37:3435–3444. 501
38. Cavaleiro R, Baptista AP, Foxall RB, Victorino RMM, Sousa AE. 2009. Dendritic 502 Cell Differentiation and Maturation in the Presence of HIV Type 2 Envelope. AIDS 503 Res. Hum. Retroviruses 25:425–431. 504
39. MacNeil A, Sarr AD, Sankalé JL, Meloni ST, Mboup S, Kanki P. 2007. Direct 505 Evidence of Lower Viral Replication Rates In Vivo in Human Immunodeficiency Virus 506 Type 2 (HIV-2) Infection than in HIV-1 Infection. Journal of Virology 81:5325–5330. 507
40. Suspene R, Meyerhans A. 2012. Quantification of Unintegrated HIV-1 DNA at the 508 Single Cell Level In Vivo. PLoS ONE 7. 509
41. Mörner A, Björndal Å, Albert J, KewalRamani VN, Littman DR, Inoue R, 510 Thorstensson R, Fenyö EM, Björling E. 1999. Primary human immunodeficiency 511 virus type 2 (HIV-2) isolates, like HIV-1 isolates, frequently use CCR5 but show 512 promiscuity in coreceptor usage. J. Virol. 73:2343–2349. 513
42. Schramm B, Penn ML, Palacios EH, Grant RM, Kirchhoff F, Goldsmith MA. 2000. 514 Cytopathicity of human immunodeficiency virus type 2 (HIV-2) in human lymphoid 515 tissue is coreceptor dependent and comparable to that of HIV-1. J. Virol. 74:9594–516 9600. 517
43. Schmitt N, Nugeyre M-T, Scott-Algara D, Cumont M-C, Barré-Sinoussi F, Pancino 518 G, Israël N. 2006. Differential susceptibility of human thymic dendritic cell subsets to 519 X4 and R5 HIV-1 infection. AIDS 20:533–542. 520
44. Rozmyslowicz T, Murphy SL, Conover DO, Gaulton GN. 2010. HIV-1 infection 521 inhibits cytokine production in human thymic macrophages. Exp. Hematol. 38:1157–522 1166. 523
45. Braun J, Valentin H, Nugeyre M-T, Ohayon H, Gounon P, Barré-Sinoussi F. 1996. 524 Productive and Persistent Infection of Human Thymic Epithelial Cells in Vitro with HIV-525 1. Virology 225:413–418. 526
46. Laguette N, Sobhian B, Casartelli N, Ringeard M, Chable-Bessia C, Ségéral E, 527 Yatim A, Emiliani S, Schwartz O, Benkirane M. 2011. SAMHD1 is the dendritic- and 528 myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 474:654–529 657. 530
47. Ricci EP, Soto-Rifo R, Herbreteau CH, Decimo D, Ohlmann T. 2008. Lentiviral 531 RNAs can use different mechanisms for translation initiation. Biochem. Soc. Trans 532 36:690. 533
48. Strong CL, Lanchy J-M, Dieng Sarr A, Kanki PJ, Lodmell JS. 2009. A 5′UTR-534 Spliced mRNA Isoform Is Specialized for Enhanced HIV-2 gag Translation. J. Mol. 535 Biol. 391:426–437. 536
49. Soto-Rifo R, Limousin T, Rubilar PS, Ricci EP, Decimo D, Moncorge O, Trabaud 537 MA, Andre P, Cimarelli A, Ohlmann T. 2012. Different effects of the TAR structure 538 on HIV-1 and HIV-2 genomic RNA translation. Nucleic Acids Res. 40:2653–2667. 539
50. Brooks DG, Kitchen SG, Kitchen C, Scripture-Adams DD, Zack JA. 2001. 540 Generation of HIV latency during thymopoiesis. Nat. Med. 7:459–464. 541
51. Marini A, Harper JM, Romerio F. 2008. An In Vitro System to Model the 542 Establishment and Reactivation of HIV-1 Latency. J. Immunol. 181:7713–7720. 543
544 545
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

19
FIGURE LEGENDS 546
547
Figure 1: HIV-2 infects human thymic tissue and isolated thymocytes. Thymic organ 548
cultures (TOC) were infected with R5- or X4-using HIV-2 or HIV-1 primary isolates and 549
cultured for 10/11 days on Millipore inserts. Single cell suspensions of total thymocytes or 550
CD4SP thymocytes, sorted as CD3high
CD8neg
cells, were also infected with the above-551
mentioned viruses and cultured for 10 days. Graphs show total HIV DNA (A), as assessed by 552
quantitative real-time PCR, or viral gag mRNA (B), as assessed by real time RT-PCR 553
(normalized to GAPDH expression). Each dot represents a single thymus. Lines indicate 554
median values. *: P<0.05; **: P<0.01. 555
556
Figure 2: Viral production in HIV-2-infected TOC. TOC infected with R5- or X4-using HIV-2 557
or HIV-1 primary isolates were analyzed for viral production by immunohistochemistry, using 558
anti-Gag antibodies (brown) and hematoxylin counterstain (blue). (A) Viral production at days 559
1 and 11 after infection with HIV-2 (top panels) or HIV-1 (bottom panels), as assessed using 560
anti-p27 (ARP396/397 from MRC) or anti-p24 (Kal-1 from DAKO) mAb, respectively. (B) 561
Extracellular (arrow) and cytoplasmic (arrowhead) Gag expression in R5 or X4 HIV-2-infected 562
TOC at day 11 (ARP396/397 anti-p27 antibody). 563
564
Figure 3: Limited replication of HIV-2 in human thymocytes. Viral production at the single-565
cell level was determined by flow-cytometry using the anti-Gag antibody KC57 10 days post-566
infection of TOC, total thymocytes or CD4SP thymocytes with R5- or X4-tropic HIV-1 and 567
HIV-2 primary isolates. Representative dot plots of intracellular Gag expression (A) and 568
frequency of productively infected KC57+ thymocytes (B) in each condition are shown. Flow 569
cytometric analysis was performed after exclusion of dead cells and aggregates. Each dot 570
represents a single thymus. Lines indicate median values. *: P<0.05; **: P<0.01. 571
572
Figure 4: Distinct cytopathic impact of HIV-2 infection on TOC and thymocytes. HIV-573
infected TOC (A and B) or thymocyte suspensions (C and D) were cultured for 10 days. 574
Representative dot plots of CD4 and CD8 expression in cells in TOC (A) and in total 575
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from

20
thymocyte suspensions (C), with graphs showing the mean frequency of thymocyte subsets in 576
all thymuses analyzed (B, n=4, and D, n=8; respectively). TOC analysis was performed in 577
CD14neg
CD16neg
CD19neg
CD123neg
cells. Dead cells and aggregates were excluded from the 578
flow-cytometric analysis. (E) Frequency of CD4neg
cells within Gag+ (KC57
+) thymocytes in 579
HIV-infected thymocyte suspensions. (F) Fold-change in the frequency of live cells in TOC 580
relative to uninfected control, as assessed by flow-cytometry. (G) Fold-change in the number 581
of live thymocytes in culture relative to uninfected control. Lines indicate median values. *: 582
P<0.05. 583
584
Figure 5: HIV-2 X4 envelope per se does not impact on thymocyte distribution in TOC. 585
TOC were cultured for 7 days with medium only (control) or in presence of the recombinant 586
envelope proteins HIV-2 gp105ROD, HIV-1 gp120Ba-L or gp120IIIB (all at 1g/ml) or anti-CD4 587
mAb. Dot plots show a representative example (one of 3 independent experiments with 588
different thymuses) of the frequency of subsets in TOC, as determined by CD4 and CD8 589
expression. 590
591
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from
21
Table 1: HIV viruses used in the study. 592 593
Viruses Major co-receptors used Source
Primary isolates HIV-192US660 R5 NIHa
HIV-192HT599 X4 NIHa,b
HIV-260415K R5 NIHa,c
HIV-220.04d X4 Dr. Nuno Taveira
Lab-adapted strains HIV-1NL4-3 X4 MRCe
HIV-2ROD10 R5, X4 MRCe
594 aNIH AIDS Reagent Program, Division of AIDS, NIAID, NIH; from The Multi-center AIDS Cohort Study;
bprovided by 595
Dr. Neal Halsey; cprovided by Dr. Feng Gao and Dr. Beatrice Hahn;
dpreviously represented as PTHCC20/2004 (26) 596
and 19/2004 (27); eNIBSC - Centre for AIDS Reagents, UK, Medical Research Council. 597
598
599
on June 7, 2016 by guesthttp://jvi.asm
.org/D
ownloaded from