thoracopterus bronn (osteichthyes: actinopterygii): a gliding fish from the upper triassic of europe
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/254313549
ThoracopterusBronn(Osteichthyes:Actinopterygii):AglidingfishfromtheUpperTriassicofEurope
ARTICLEinJOURNALOFVERTEBRATEPALEONTOLOGY·SEPTEMBER1992
ImpactFactor:1.98·DOI:10.1080/02724634.1992.10011459
CITATIONS
28
READS
61
2AUTHORS:
AndreaTintori
UniversityofMilan
105PUBLICATIONS1,056CITATIONS
SEEPROFILE
DavideSassi
UniversitàdegliStudidiMIlano
26PUBLICATIONS92CITATIONS
SEEPROFILE
Availablefrom:DavideSassi
Retrievedon:04February2016

! Joumal of Vertebrate Paleontolo gy | 2(3):26 5-283, September l 992@ 1992 by the Society ofvertebrate Paleontology
INTRODUCTION
In the present-day seas, a group offishes belongingto the Exocoetidae has achieved the capability of glid-ing in the air; they are often called "flying fishes" inthe scientific literature (Bertin, 1958; Bruun, 1935;Hubbs, 1932;Lauder and Liem, 1983). Though wide-spread in most temperate and tropical seas, their tax-onomic diversity is very low. The Exocoetidae are firstknown from the Eocene (Mùller, 1985). No other groupofliving fishes seem to have a similar behavior (Hubbs,1932:577).
From the studies by Hubbs (1932) and Bruun (1935)on exocoetids it is clear that two main characters de-termine whether a fish is a glider or not: a high ratioof fin length to standard length (up to 60 or 70olo) andan associated hypobatic caudal fin with an upper lobelength equivalent to 58-750/o the length of the lowerone.
Among Actinopterygii, only the Late Triassic Thora-copteridae shows the same morphological characters.This family is known with certainty from Austria(Reingrabener Schiefer, early Carnian, Grifrth Í19771)and Italy (Calcare del Predil, early Carnian, Bronn[858]; Dolomie di Forni, early Norian, this paper;Dolomie di Giffoni, Bassani [1895]; Calcare diZorzinoand Argilliti di Riva di Solto, late Norian, Tintori andSassi [987]) .
A complete taxonomic discussion, regarding boththe species and family relationships, is also possible inthe light of better anatomical knowledge based on newwell-preserved material from the late Norian ofNorth-ern Italy. We consider the four thoracopterid speciesas belonging to the same genus, Thoracopterus. Twospecies, T. niederristi and T. telleri (the latter previ-ously referred to Gigantopterus) are of early Carnian
T:HORACOPT:ERUS BRONN (OSTEICHTHYES: ACTINOPTERYGII):A GLIDING FISH FROM THE UPPER TRIASSIC OF EUROPE
ANDREA TINTORI and DAVIDE SASSIDipartimento di Scienze della Terra, Universita degli Studi di Milano,
Via Mangiagalll 34,20133 Milano, Italy
ABSTRACT-The Late Triassic gliding fishes of the gents Thoracopterus (Actinopterygii, Peltopleu-riformes) are reviewed, and Thoracopterus martinisi, sp. nov. is described. We consider the familyThoracopteridae a valid taxon, but place it closer to Peltopleurus, rather than to Perleidus or Luganoia,especially on the basis ofthe pattern ofskull roofing bones, shape ofthe preopercular, scale similarities,and caudal fin structure. Morphological characters ofthree species of Thoracopterzs, such as length ofpaired fins, proportion of caudal fin lobes, and position of dorsal and anal fins, are compared withcorresponding characters in extant monoplane and biplane gliding fishes. On the basis of this com-parison, Thoracopteru.s is viewed as a biplane gliding fish, so far the only one known outside the extant"flying fishes" of the family Exocoetidae.
age; the other two are from the early Norian (7. mar-tinisi, sp. nov.) and late Norian (7. magnificus).
Thoracopleru.r was considered as "flying fish" in itsoriginal description (Bronn, 1858) and compared toexocoetids. In this paper we propose that thoracop-terids, based on a detailed morphometric comparison,were biplane gliders, thus similar to the most derivedextant Exocoetidae (Cypse lurus).
SYSTEMATIC PALEONTOLOGY
Family Tnonacop"rERrDAr Griffith, 1977
Emended Diagnosis - Small to medium-sized fi shes.Skull roof with broad paired frontals; large dermop-terotics sometimes fused together; no distinct parietals.Nasals separated by large rostral. Supraorbital singleor few in number. Opercular series with opercular,subopercular, several branchiostegal rays, lateral andmedian gulars. Vertical, narrow preopercular abuttingthe rear edge of the maxilla. One suborbital. Maxillaextending beyond posterior border of orbit and withlarge postero-ventral plate. Mandible with low coro-noid process. Spiracular bones present. Large otoliths.Sensory canal system reduced. Body scaled or naked.Tail diphycercal with more than ten epaxial rays. Cau-dal fin deeply forked with lower lobe larger than upper.Pectoral fins very large. Pelvic fins well developed.Anal and dorsal fin lepidotrichia reduced in length.
Remarks-So far only four species of fossil glidingfishes, other than Exocoetidae, are known: Thoracop-terus niederristi, T. magnifirus, T- martinisi, sp. nov.,and the closely related, but poorly known, Gigantop-terus telleri (Bronn, 1858; Kner, 1866, Abel, 1906;Griffith, 1977;I-nhman,1979:' Tintori and Sassi, 1987).Wade (1941, 1942) recorded ?Thoracopterus frorn the
265

I
266 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 12, NO. 3. 1992
Middle Triassic ofNew South Wales, but, in both cases,he only referred to isolated large pectoral fins. Theattribution was therefore very doubtful, and Wade(1942) noted ". . . they are referred to Thoracopterusfor convenience of record only." Actually, the branch-ing of the rays is not dichotomic, as it is in all speciesof Thoracopterus. Moreover, the posterior outline ofthe figured specimen (Wade, 1942) is straight ratherthan rounded, the anterior lepidotrichium being thelongest. We thus believe that this fin cannot be ascribedto Thoracopterus or to any other flying fish.
Genus TnonacorrERus Bronn, 1858
Diagnosis-As for family; only known genus.Remarks-The abundance and the better preser-
vation of the Norian specimens from the Calcare diZorzino allow a more accurate anatomical restorationofboth the skull and the body ofthis genus. Therefore,a few characteristics, such as the loss ofscale coveringand the large range in size, appear as mere intragenericvariations. In our view, Gigantopterus Abel 1906 mustbe considered a junior synonym of Thoracopterus, fheonly differences being a size larger than Thoracopterusniederristi, the loss of scales, and, possibly, the size ofthe subopercular. Griffith (1977) also found that thedermal skull bones of the two genera are very similar.Because large size and naked body are present also inthe Norian species T. magnificus and T. martinisi, sp.nov., it seems that size of the subopercular alone is notsuftcient to support generic distinction.
A few alleged differences between the Carnian andthe Norian thoracopterids are due to misinterpretationof the poorly preserved Carnian specimens (Grimth,1977; Llnhman, 1979). For example, the shape andposition of the rostral (in our opinion separating thenasals in both Thoracopterus niederristi and T. telleri)and the shape of the maxilla very much resemble thoseof the Norian species (pers. obs.).
One specimen of Thoracopterus from the Norian ofGiffoni (Salerno, Italy) was described and figured byCosta (1862:pl. 5, fig. 2) as belonging to Urocomuspicenus; it was, in fact, mistaken for a fragment of acoelacanth described by Costa himself, type of whichis the specimen figured in the same paper (Costa, 1862:pl. 6, fig. l). Later, Bassani realized the misinterpre-tation and referred the specimen figured by Costa (1862:pl. 5, fig. 2) to Pterygopterus (Bassani, 1892) and laterto Thoracopterus (Bassani, 1895). We now believe thatit can be referred fo T. magnificus.
Four species of the gews Thoracopterus are consid-ered here: T. niederristi and T. telleri from the Carnianand Z. martinisi, sp. nov. and Z. magnfficus from theNorian. Only the first species is completely covered byscales; the others have a naked body. Z. telleri has avery reduced subopercular. Furthermore, the length ofits pectoral fins is up to 70olo of standard length, whichis considerably longer than in the Norian species, whereit reaches 35olo in T. martinisi, sp. nov. and up to 600/oin T. magnificrzs (Figs. 17,l8). In our opinion, the dif-
ferent relative lengths ofthe pectoral fins alone is suf-ficient to justify the separation of these three latterspecies.
Abbreviations-GIUf , Geologisches Institut derUniversitàt Innsbruck; MCSNIO, Museo Civico diScienze Naturali di Induno Olona; MCSNM, MuseoCivico di Storia Naturale di Milano; MFSNU, MuseoFriulano di Storia Naturale di Udine; MPUN, MuseoPaleontologico dell'IJniversita' di Napoli; SM, For-schungsinstitut Senckenberg, Frankfurt am Main.
af, anal fin; ant, antorbital; ap, anterior pit line; ar,anal fin radial; br, branchiostegal rays; cl, cleithrum;cs, caudal scute; df, dorsal fin; dpt, dermopterotic; dr,dorsal fin radial; dsp, dermosphenotic; ethco, eth-moidal commissure; exsc, extrascapular; fns, first neu-ral spine; fr, frontal; frdpt, frontal-dermopterotic; gu,gular plate; ha, haemal arch; hs, haemal spine; hypl-2, hypurals; inf, infraorbital; infc, infraorbital sensorycanal; isl, inner scaly lobe; lhy, last hypural; mand,mandible; mc, mandibular sensory canal; mmc, maincephalic sensory canal; mns, modified neural spine;mx, maxilla; na, nasal; nal, anterior naris; na2, pos-terior naris; nar, neural arch; ns, neural spine; op, oper-cular; pa, parietal; pcf, pectoral fin; phyp, parhypural(haemal spine of first pre-ural centrum); pmx, pre-maxilla; pop, preopercular; pp, posterior pit-line; pscl,presupracleithrum; pul*ul*u2, centrum formed byfusion offirst pre-ural and ural centra; pu2-7, pre-uralcentra; pvf, pelvic fin; r, rib; rad, radial; ro, rostral;sbo, suborbital; scco, scapulocoracoid; scl, supracleith-rum; sd, supradorsal; sn, supraneural; soc, supraorbitalsensory canal; sop, subopercular; sp, spiracular; unl-2, uroneurals.
Genus THonacoyrERus Bronn, 1858
TnonecoprERus MAGMFrcusTintori and Sassi. 1987
(Figs. 2-8, 10, 12B)
1862 Urocomus picenus p.p., Costa:32 and pl. 5,fig. 2.1892 Pterygopterus? sp., Bassani: I 0.1895 Thoracopterus(?) sp., Bassani:2O8 and pl. 4, frg.
4, pl. 7, fig. 64.1987 Thoracopterus magnfficus Tintori and Sassi:338
and figs. l-3.
Diagnosis -Small Thoracopterus ; oînamentationpresent, particularly prominent on the skull roof; su-praorbital sensory canal ending on the anterior thirdof the frontal and followed by two parallel, curved andtransverse pit lines; posterior lobe ofthe pectoral finscovered by scales; naked body; pectoral fin length about5o-600/o of standard length; pelvic fins about 25o/o ofstandard length.
Type Locality - Enden na, Zogno (Bergamo, Italy).Type Horizon-Calcare di Zorzino (Zorzino Lime-
stone); age: middle-late Norian (Late Triassic).Holotype-Museo Civico di StoriaNaturale "E. Caf-
fi," Bergamo, no. 7820.
:
II
I
I
lrcscct(

TINTORI AND SASSI_TRIASSIC GLIDING FISII 267
Aust ra I osorrus PeI toDI eurus Habroi chthys Thoragopterus Cte i th ro teo i s Per Ieidus LUganol a
2 , 3 ó , 8 3 2
FIGURE l. Cladogram showing our interpretation of the nodes S-U sensu Gardiner and Schaeffer (1989: flg. l2). Characters:1, presence oflarge epaxial rays;2, deep flank scale-row(s); 3, infraorbital process ofthe preopercular;4, dorsal and anal finrays equal supports; 5, preopercular narrow in dorsal region; 6, hypobate caudal fin; 7, epaxial rays less than l0; 8, coronoidprocess; 9, free posterior end of the maxilla.
2 , 3
J
n
ddIS
rtlf
e-
Lf-
Material-70 specimens (20 almost complete); 66specimens (Tintori and Sassi, 1987) in the Museo Civi-co di Scienze Naturali "Caffi," Bergamo; MCSNMVll9l from Ponte Giurino, SM P5468 from Cene,MPUN Ml45 from Giffoni (Salerno), GIUI P9030from Seefeld, and MCSNIO P264 from Endenna-Zo-gno.
Description
The rostral (Figs. 4, 5) is large, abutting to the an-terior edge of the frontals and thus separating the twonasals. Anterior to the rostral, there is a median den-tigerous premaxilla (Fig. aA); its dorsal part is coveredby the rostral itself. The latter has a transverse ridgeon its inner surface against which the premaxilla abuts.This ridge corresponds in position to the traces of theethmoidal commissure on the external surface. Theteeth of the premaxilla are slightly smaller than thosecarried by the maxilla and number up to l3; the mediantooth is larger than the others. Only one tooth-row isvisible, but it seems reasonable to assume that morethan one row was originally present.
The presence of a median premaxilla is rather un-usual (e.g., Canobius ramsayi Traquair; Moy-Thomasand Dyne, 1938); a median "rostropremaxilla" hasbeen described in Birgeria groenlandica and perhapsin some parasemionotids (Gardiner, 1963:314). Morerecently, Gardiner (1984) wrote that the so called"postrostral" is considered a true rostral, which is amedian bone; thus the rostropremaxilla is simply thepremaxilla. We confirmed this on a specimen of Bir-geria acuminata from Cene, on which we have beenable to observe the "rostropremaxilla" sensu Nielsen(1949). This bone is much lower than in Birgeria groen-landica and there are no traces of the ethmoidal com-missure; it is a premaxilla. Thus the posterior largemedian bone must be the rostral (postrostral; Nielsen,1949). Furthermore, one of us (A.T.), while preparingnew specimens of Perleidus altolepis and of a new spe-cies of the perleidid Dipteronotu^s (Tintori, 1990a), ob-served that the median shield-like bone, which wascalled "postrostral" in the Early Triassic Perleidus spe-cies (Stensió,l92l; Lehman 1953) or "nasal" in Dip-teronotus aculeatus (Gall et al., 1974), bears the eth-
moidal commissure. The paired "rostrals" of Lehman(1953) or "antorbitals" of Stensió (1921) and Gall etal. (1974) are nasals because they show the nostrilnotches and the supraorbital sensory canal. As a con-sequence, the dentigerous bone(s) in front ofthe rostralmust be considered true premaxilla(e), unpaired in Bir-geria(at least in B. acuminata), Canobius ramsayi, andThoracopterus and paired in all other actinopterygians.
As P ho lidop horus, I c ht hyoke nte ma, and primitiveteleosts (Gardiner, 1963) have a median dentigerousbone in the upper jaw (rostral), the presence of a me-dian premaxilla may be considered a structural con-vergence.
The nasals (Figs. 4, 5) are slender bones, with thetwo narine notches in the anterior third. Sparse, talltubercles on the nasals as well as on the rostral contrastwith the denser, but lower, ornamentation on the skullroofing bones.
The frontals (Figs. 4, 5) are roughly rectangular,sometimes with a slight constriction at the level of theorbits. The median suture is straight, but, in a fewspecimens (as in the case of the holotype), the twobones are fused together.
Posterior to the frontals, there is usually only a pairof bones, which are called dermopterotics (Figs. 4, 5)because they were related to the sensory canal. Thesuture between the frontals and the dermopteroticsgently slopes forward from the midline.
Ornamentation is homogeneous on the whole skullroof; it comprises densely arranged low tubercles andridges.
Posterior to the dermopterotics, there are three pairsof small plates (Figs. 4, 5), usually in contact with theposterior edge of the dermopterotics themselves. Thelateral pairs are presupracleithra (sensu Nybelin, 1976;postspiraculars ofauthors) because they do not showany traces of sensory canals. The two median pairs arethe extrascapulars, bearing pit lines that replace theoccipital commissure. This bone occupies the sameposition as in Perleidus piveteaui (Lehman, 1952).
Posterior to the presupracleithrum and the lateralextrascapular, there is only alarge supracleithrum. Thispattern may be related to the unusual median positionof the sensory organs in the dermopterotics.
The maxilla (Figs. 4, 5) is elongated, very low an-
L

268 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 12, NO. 3. 1gg2
l+.+I
//:'í;í,
1l
'ì;
. l î t
B .FIGURE 2. Thoracopterus magnificus. A, Holotype, MCSNBG 7820. B, MCSNBG 3300. Scale bars equal l0 mm.
teriorly but ventrally expanded in the posterior part,where it is in contact with the preopercular. The den-tition is powerful, with the teeth being arranged in atleast three rows on a rather large horizontal platform
of the anterior region. The largest teeth, at least 25,make up the innermost row; three to four teeth arealso visible where the maxilla expands to overlap themandible.
FIGURE 3. Thoracopterus magni,fìcus. Body restoration. Scale bar equals l0 mm.
. . 1 ' - . , , .
r ' , 1 j 1 ' - " . ; " ' '

TIN'TORI AND SASSI_TRIASSIC GLIDING FISH
FIGURE4.Thoracopterusmagni f icus.skul l restorat ion 'A,dorsalv iew;B' la tera lv iew'scalebarsequal5mm'
269
The lower jaw (Fig. 4) is equally well supplied with
t..ìrr. Tlt" o.àl -utgitt is straight and bears teeth only
aiÀng lts anterior two-third. The articular notch is on
itr. ótt..oOorsal corner behind the low coronoid pro-
c.ss, *ttictt occupies the posterior third of the man-
;ìbi;. ih" anteribr five oi six teeth are the largest in-
ttre Ào,rttt, the others being similar in size to those of
the maxilla. Three tooth-rows are found also in the
io*"iju*, but the largest teeth here lie externally' All
itt. i.étrt úave pointeé ".o*ns with an acrodine tip'"--ltr" posltion of the preopercular (Fig' 4).is vertical;
it shows an anteriorly directed process' which encom-
futret ttt" posterodorsal edge of the-maxilla' The pre-
àperc.rtat is about three-and-one-half times as deep as
long, including the infraorbital process'- ii" op"t"uiar bones (Fig. a) are usually very poorly
pr"r"rrrù. We can only nóte that the subopercular is
much smaller than the opercular' The boundary be-
t*";; them is steeply anteroventrally directed' so that
the anteroventral corner ofthe opercular is near to the
"""tJ éOg" ofthe subopercular' Eleven branchiostegal
;;t;;;" p"receoeo bv làteral and median gulars' The
;;;;";"" -oitttt""
grriutt is interpreted as a very prim-
iii't " f"ut"t" (Gardiner, 1984). The branchiostegal rays
are well ornamented with large ganoine tubercles'
It"r"u, the opercular and subopercular are nearly
smooth.- The basioccipital and the posterior end ofthe para-
rpft""oiJ are ràrely visible. The latter bone is of the;ittt"t-"aiute type;' (sensu Lehman, 1966); it extends
b;t;"d the otió^fissure but it does not overlap the
."óipit"f i"gion. This pattern re-sembles very much that
in Perleidui (Patterson, 1975:fig' I 16)'- The sensory canal system is peculiar and reduced'

270 JOURNAL OF VERTEBRATE PALEONT-OLOGY, VOL. 12. NO. 3. 1gg2
cFIGURE 5. Thoracopterus magniJicus. Drawings of (A)MCSNBG 7817, (B) MCSNBG 7816, (C) MCSNBG 7820(holotype). Scale bars equal 5 mm.
Traces of the ethmoidal commissure are present on therostral as a transverse furrow, which does not reachthe lateral margins; perhaps a pit-line replaced thecommissure. The supraorbital canal extended from the
anterior region ofthe nasal back to the anterior thirdofthe frontal. Several pores are present on both sidesofthe canal itself. Posterior to the end ofthe canal inthe frontal are two transverse pit-lines; the anteriorone had two neuromasts and the posterior five to sev_en. The dermopterotic shows a curved canal endinsblindly in the posterior third. This curious shape iitypical of Thoracopterus and was already noticeà bvGriffith (1977) and Lehman (1979) in i. niederristí.The pores are smaller than those on the frontal. Tracesof the temporal commissure are seen on the extrascap_ulars as a pit-line on both sides of the skull. Numeroùspores are also visible on the mandible and on the ven_tral region ofthe preopercular.
The cleithrum is curved and its posterior margin isslightly notched; its posteroventral corner is widè androunded.
The axial skeleton (Figs. 3, l0) consists of about 40vertebrae. At least 26-28 long, slender paired neuralarches and spines are present in the region between theskull and the posterior margin of the dorsal fin. A bonyrod is associated with each paired spine in its distalthird and is considered a supradorsal, marking the po_sition ofthe longitudinal ligament. Supraneurals ocèuras well as thin ribs, the latter being rarely visible. TheI I to l3 most posterior neural arches give rise to un_paired spines. These spines are peculiarly shaped, be_coming more and more expanded posteriorly, and havedeeply incised distal ends. The last three or four spinesare longer than the preceding ones and decrease grad_ually in size. They are more transverse and horizontal,supporting the epaxial rays ofthe caudal fin.
Ventral to the unconstricted notochord, in the an_terior trunk region, there are keeled parapophyses bear_ing thin ventral ribs. In the posterior trunk region, fiveor six larger parapophyses are present, possibly withoutany associated ribs. In the caudal region, haemal arch_es, each with a large unpaired spine, are present. Theposterior seven spines are more elongated and supportthe lower lobe of the caudal fin. The last elemenì hasa, short, fan-shaped spine with a vertical posterior edge,though the body axis is only slightly upturned. Thelateral surfaces of these spines are more or less deeplyfurrowed.
Apart from their large size, the pectoral fins (Figs.3, 8). show other peculiar characters, including'all,o-metric growth from about 4Oo/o of ttre standard length
m m c
,\r -.,
\dpl
----------- B.
FIGURE 6. Thoracopterus magnfficus. otolith of specimen MCSNBG 7825.

TINTORI AND SASSI_TRIASSIC GLIDING FISHior thirdrth sidescanal inanterior) to sev_ending
Ihape isiced bylerytsi.Tracesrascap-nerousre ven_
rgn isle and
rut 40reural,n thebonylistal3 po-rccurTheun-be-
|aveinesad-tal,
27r
tn-ar-veruth-tertts
e
FIGURE 7.region ofthe
Thoracopterw *"g"rfily:-_!:p€,crorar girdre and fins of MCSNBG 7815. Scale bar equalsverrebral column of MCSNBGiSf Z. S"iì"Gr equats 5 mm.l0 mm. B, caudal
in s.ma_ll specimens (up to 40 mm in s.l.) îo about 6Ooloin the longest. In rhe lossils, the fi;;-úk"*rt "r ry__metrical -when spread, the longest tepiOotrictrit bÉ_gthe fourrh and the fifth of the ú ".ii il;ily makingup the fin. The first lepidotrichium is saber_tike andmuch larger than the following o.r"s. fu"À has a lon!
proximal segment and branches three times. The prox_imal segments each bear an anterior win!_6." process,which supported the preceding orr" *t "-r, the fin wascompletely spread. Small denli"t"s arJì.rbercles arefould at least on the lar-gest .p""iÀ""r;*they cover thewhole anterior region ofltre "i"trui-rr"i"iiàpidotrichia

272
FIGURE 8. Thoracopterus magnificus. Drawing of the pec-toral girdle and fins of MCSNBG 7815 (same as Fig. 7A).Scale bar equals 5 mm.
and the posterior region of the dorsal ones. As a con-sequence, adjacent lepidotrichia were held tightly to-gether when the fins were closed.
The proximal end of each lepidotrichium is bentbackward, most sharply in the posterior elements, inorder to provide stronger support to each other. Lep-idotrichia articulate through three or four strong ra-dials on the ventral side of the scapulocoracoid. Fur-thermore, we observed the presence of an inner scalylobe ("sensenÍÌirmig," sensu Abel, 1906; "voile falci-forme" sensu Lehman, 1979) that filled the space be-tween the last ray and the body, providing better lift.
The pelvic fins have only seven or eight lepidotrich-ia, which are very similar in shape to those of thepectorals. The maximum length of the fins, almostindependent of the specimen dimension, is 20 to 23o/oof standard length. Consequently, the ratio betweenpelvic and pectoral length varies from 37o/o in the larg-est fishes to 58o/o in the small ones. The pelvics arerather posterior, being inserted at about 670lo of stan-dard length.
The dorsal fin comprises eight to ten lepidotrichia.They are placed rather far apart from each other, sothat the fin insertion is fairly long. The dorsal usuallybegins at about 75olo of standard length, anterior to theanal, which begins at 87o/o. There are more than 15anal fin lepidotrichia; they are very thin, short, unseg-mented, and unbranched. Anterior to the anal, thereare a couple ofnarrow, elongated scales. We only foundthe endoskeletal support of the dorsal fin, in whicheach large radial supports one lepidotrichium.
The caudal fin (Figs. 78, 8) shows peculiarities clear-ly linked to the flight function. It has a very wide sur-face with an hypobatic shape, the upper lobe beingonly 74o/o of the lower lobe length. The dermal skeletonconsists of 30 lepidotrichia; the central elements branchup to five times. The number of lepidotrichia in thetwo lobes is the same, and in the lower lobe they arelarger and longer than in the upper. Segmentation startsafter a first long segment, like in the other fins. Thismay represent a further improvement ofthis character.
JOURNALOF VERTEBRATE PALEONTOLOGY, VOL. 12, NO.3, 1992
In Thoracopterus niederristi, the oldest known species,the first segment is approximately as long as the fol-lowing ones. Basal fulcra are more numerous on theupper than on the lower edge.
TnonacoprERus MARTINTST, sp. nov.(Figs. 12-14, l5A)
Diagnosis - Medium- sized Thoracopterus; skullroofing bones often fused together; ornamentation con-sisting only of tubercles; supraorbital sensory canalending in the posterior half of the frontal; pectoral finlength not exceeding35o/o ofthe standard length; nakedbody.
Holotype-MFSNU 1885.Etymology-Named for Mr. E. Martinis, from Am-
pezzo (Udine), who first found fossil fishes in the Preonearea.
Type Locality-Rio Seazza Valley, near Preone(Udine, Italy).
Type Horizon-Dolomia di Forni, middle parq age:early Norian (Late Triassic).
Other Material-20 specimens, all from the Preonearea (MFSNU l87l -1,1881-4, 1886-90, l90 l -4) .
Description
The rostral (Figs. 13, 15) is large, rapidly taperingforwards. Only its posterior region is usually visible,but sometimes the rostral itselfis preserved uncrushed,showing the original shape with the ventral part lessdeveloped than the dorsal one. Owing to the preser-vation, no traces of the ethmoidal furrow are visible.
The nasals (Figs. 13, I 5) are shorter and more squar-ish than in Thoracopterus magnificus, tt'e boundarywith the rostral being shorter as well.
The frontals (Figs. 13, l5) may be fused to eachother. In MFSNU 1785, the dermopterotics are alsofused to the frontals so that the main portion of theskull roof is occupied by a single, very large bone. Theembayment for the orbit is somewhat deeper than inThoracopterus magnificzs and the region anterior to
FIGURE 9. Cypselurus heterurus (Raflnesque, 1810). Prox-imal end of the anterior lepidotrichia of the pectoral fin. A,dorsal view; B, ventral view. Scale bar equals l0 mm.
A
I

pecies,he fol-on the
skullcon-:analtl fintked
|.m-one
)ne
ge:
ne
*::,"",
AND SASSI_TRIASSrc GLIDING FISH273
I
I
sFIGURE I0. Thoraconîcruî tunob;A-..^ ^
1a 1q1 reeiot " ; ;;; ;:í:í:6 #if#{" f ,#élffir"B. MCSNBG 7817 (same as eig. zÉt.'S-."1" i"i "q*r
bitals borderins. ilÀ; ;;; i;: ix!ÈlLi ;: "ffiL:,i I ii : xil,l"::X:mosphenodc and *re. posieri;;;;;' infraorbiral arel":-:i,! .larse ruuoruitat'uuJir' o'i,t " anrerodorsatmargln of the preopercular.
The operculàr .
opercurar Gú; i f ffl *T";:",T..?11::"#".if;same shape as in Thora}op,irir-^ààr,firur.
r,,lh#îlr'f:j"t'" comprises "u3"t 40 verrebrae.deil iT,;, ;;;#r 1"É: .:lÉ1 ft ì: j :J: ffiji#::#ral spin e s are m odi fi e a nui- i"'Fnî i oi à o, ", u, m a g n iJi _
,rJ**ffil?.î:*Ì:'*r" in rength rrom 20 6 3so/o orra.,,iir,iì-it"idill"f, ff #:H::?xTi,..1.,;ili;TJ;only in rheir distal portion. The À.riìipiaotrichium issa ber-li ke and larser ^trr"n it . ilr ià*ìn'r'on".. A n reriorwingJike pro"".ré, are present in ÀE first proximalsegments, as in Thoraîo_pJ9lus -ognirt"rr. only a cou_ple orsrrons radiars.(MFSNú fii;í;lve been iden_trhed in the endoskelàr"n of in.
';;., , '"
The pelvic fins are.well dev;lo;;d, reaching l9 ro2oo/o of rhe standard r""girrl É""Iffà ,is
maoe up of:?tt f3:Í.Xilchia,
whicí ";";ilir;;i; ,nuo" t. fro;;- About 12 lepidotrichia are found in the dorsal fin;the endoskeletal rays are large ""0 UL"t backwards.They show a one_lo_one ratió wittr th-e tepidotrichiarhemselves. Three ro four u*àr rurc*,i;"" tt " fin andare supported bv tl
other. _ tree rays, partially fused to "u"À
Nothing can be r1i9 glrlr" anal fin, which is alwavscovered by the lenidorrichi" "i'tr,.'pàrli" n"..The caudal fin ìs rrn T h o r a c o p, ", u, -)oÎi!!-u
t . l " t t a s v m m e1 ri cal th a n
roue.ueini"íùo;;;i'if ::{rr::;Jff :.îjff IrT:ifjiilelidotrichia are preced:g grl ilil;t futcra bothln the dorsal and ventral IobL.
TAXONOMIC REMARKSThe taxonomic positio n of Thoracopterus is dis_cussed here taking into account tfre cfrarícìers used bv
FIGURE ^l t. Cypselurus,heterurus,^ufin.rou.,ììl
í#]j ff ,:l te vertebrar cor u m n i; ;.';;il;i region. Scaíe
of the7 8 1 8 ;
5 mm.
the embayment is n"f rh " ;;; ;;#"J:iilfii'H iff T:,H:J_""Tffi:the dermopreror ics,. when ì;;;;à"l"of rhe fron ral s,are very simitar ro those ;f T.;;;;;À;r.Rounded tubercles are sparse orrìnó.ortrul and vervdense on the lateralas we, ", ";;;;;1XÍ,o;ì::ffi : J:'óîì; ii:r*:i3jportion of the frontals, ú;;;;;"i,ij.ao.uitar sen_sory canals, is almost smooth.Behind the dermopterotic, three pairs of bones arepresenr (Figs. 13. t 5.y: the U.à. pr"ríp*it"ith..r_ urrd
:!: ;-; "7;,î:;::urars are a"àn geo ì s
- in r n o i o ii i
A few spiracular boSes (Figs. 13, l5) lie posterior tothe dermosphenoric, u.ti""l ilrl,rtfi'ioor and thedorsal edges of the suborbital ""O"oià.o"rcular; rhearea is poorly preserved.
,#th'ì:il'j:. ##:.:jaw (Figs. 13, 15) are very
toothcounÀ";;"';;:::i:í::,"ítr".magntfi cus,andthéThe position of the. preopercular (Figs. 13, 15) isverticat, with a rarher. ú.c" i#;;;ir!ì iìo""r..
,"ll:_'-1r.,"9,?,jul (Figs. -r3, r 5t;;;; lonrirr, or urarge anrorbiral followed by one ó rrv.;;"w infraor_

274 JOURNAL OF VERTEBRATE PALEONT-OLOGY, VOL. T2, NO. 3, ]gg2
pPc l
f r dpr
-.{FIGURE l2' Thoracopterus martinisi, sp' nov. Skull restoration. A, dorsal view; B, lateral view. scale bars equal l0 mm.
A
=l-:li:::H7-:7-:2:-::
fr dpt
,ii.,.Xi:..i.,.iÌ..i,.iÌ.:i:
Gardiner and Schaeffer (19g9) as well as other synapo_morphies.
Following Lehman (1919), we think thaf Thoracon_terus cannot be included in the Luganoiform es (Lu_ganoia group ofGardiner and Schaeffer, l9g9) as pro_pos.eg by Griffith (197i).In Thoracopterus, the -uiiltuis tightly fixed to the preopercular ìhrough the infra_orbital process (sensu Gardiner, lggg); ín Luganoia,however, the maxilla is free posteriorlyiGardiner andSchaef fer ,1989).
Thoracopterus should be placed close to Node T of1f: glu9og.am published by Gardiner and Schaeffer(1989) because it has as many dorsal and anal fin raysas radials, and the upp-er caudal fin rays are elongatédbeyond termination of body axis. Thé propositión of
Lauder and I iem (19g3) that in the perleidiformes finrays ofdorsar and anal fins do not equar their internalsupports in number cannot be maintained (see alsoGardiner and Schaeffer, l9g9).
The Perleidiformes are charact eized by only few(ten or less) epaxial rays, which are elongated béyondtermination of body axis. Furthermore,"the pr"óp*cular is not_vertical, with an expanded dorsai ."gìor;the supraorbital canal extends Uack into ttre pari"etal;there are stout grinding teeth; the opercular is smallerthan. the subopercular; a dermofryat is present; themaxilla is dorsally well expanded ín the posterior re_gron; the flank scales are not dorsoventralfu elongated.None of these features are observe a h iioracopterus.
Neither does Thoracopterus show resemblances to

TINTORI AND SASST_TRIASSIC GLIDING FISH
,î?3ftH"l;;T: by a single canal_bearing bone. HowLehman (rs7s)"::^?:li"Y"d.is difficutt ó "ti"oììri.
r*ii::ì#m#'"T:"i"":1'"Ti'ff !::ffj:: jiib.o út ; ;;;;; ; :' 1: T"'v can al traj ecto'v "o ùr a r'ì,,"Iik; ;;i;.prffitfr1ilil,X1?fiili#e sman, tube-the sensory "unurìrr-rrin rhe parier", ,to"t#tl:ltj#as yet to give a de
,g.i:: ùilhili,t Ji :xli?T:?1.&i"Tî: B::ixajrr bears rraces of the sensory ;;i."i; any case, rhepattern of the sensory canals;"rù; iànsidered as anautapomorphy.The presence ofsuborbital bones related to a narrow
,l*:31'ff : r:Hn: r,'.-T b o i the' o'à.o J..",1u. i,
- "".ytr," pu.u;"?il. ** iîi*ff ì,il',ftTffi ;f ru-'man, 1952; parters on,,!97 j;óìr;;,ì.ró; in the latrer,
:Uì*":?'il:Tff J:oo'ri;c;;;;'i""1î,r*""ì".i.iaé s z) ;t;#;i."11ular (Lehman, I s s2). i;ù;;p",t"iaiiat,;;;;ufi1:,:'ii,:'lf, àiìn:'..r?",Hiiorbital in perteidus fàrtrlouen'ji;;'ffg.il in perteidusatrolepis; pers. obs.).cuu."o"rt i. ;;;il. to be over_Iooked by mosr uuí19r.. excepr scrraeHi ( t 973). Fur_rherm ore, rh e suborbi r ̂ t i"- ií i iià,irì, li,,, associaredIi lL lJr?#:'il; jl,:'::yr"T' y;t ;xiianded i n i t sorrh;-;;r;;fi iit:in:1,;'*,1î:,?:,'1",3:';Íf; :::t.1'J#Îff [Îl#i' i g t n À,í iià.iJj o *'i d a e, r h e:îy^i ; ;; ft*,-,:#t?Jff3;1"fJt*:;x x:srtron, though maintaining ttre contaci-iuirh ,h" ,.u.edge of rhe maxilta. lr| irr:"";;";"r;;:;;;, group, rhe
275
A
FIGU_RE 13. Thoracopterustype. MFSNU tgg5. B, skul loars equal l0 mm.
maninisi, sp. nov. A, Holo_root o t .MFSNU 1902. Sca le
the Cleithroleois eroup sensu Gardiner and Schaeffer( I e 8 e : a cco rd i n g t."If'.1;; i iùó; i."i",,'u, ", o, or Lts d oe snot belong here) in rb"dr ;l,ff"; ;;; ;iliii:iT l:ff :Tn: Ii3:,fJ *ifence ofa short bodv_r"b";;t;;;il::i* ten epaxials(Gardiner, 1988), ana trre aùi;;;;fi:"ronoid pro_Some characters in Thoracopte,zs resemble those of'::":,::,ií':I:nS;..""(c"'d#;;;;s"iu"n",,re8e):
r"l ;6i ;; il;'rt;,ì,lt"oo"rcular' n arro wi ns i " i tt ""í-
fl fI,*",,:TiÈfl1'ii"i{,i:}#'.f :"ilT,,1?#.,?Trt'î" r"; ;;;; d;JTh::: :{l;:,?;:T:#il lffi:ejther individually or in coupl"r, i, othì. groups. Intne Australosomus sroup. the ilank ;;"i;;." deep, andseveral epaxial ravi
11e,or9,sent, trre aìirlt uooy tou"oemg absenr; however, in the aórsat àiJurrur fins theradials do nor corresoond i" "r;;;;ìJti"i"ptAot.i"rr_ra. In rhe Luganoia group, rhe nu;k"r;;ì", are deep;the.preopercurar is "..ti.aì, uur;il;#ill it, u"nr.ulregron though free frg_ Jt-" postérior"margin of themax'la. The sim'aritv or'reie cÀa;;;#: is here in_terpreted as convergent.
- r ne dermopterotic is reduced to a narrow, lube_likebone in both perteidus and,-pdb;t;iili.'t, Habro_tchthys and Thoracopterus. rh" ;ùi;;;iri"riol. ro the
FIGURE I4. Thoracontcr,rî hnvt;-:^: ^-
"n"l,Jii,lidí3ffi[í','íí'i,?{{,liJìiì,8Ì3lr"l:X,,."?;l

276
FIGURE 15. A, Thoracopterus martiniil, sp. nov., caudalportion of the vertebral column of MFSN 1904. B, Thora-copterus magniftcus, skull of MCSNBG 7830, showing oto-liths. Scale bars equal l0 mm.
preopercular narrowing thus coincides with that of theParasemionotidae, but this stage was achieved main-taining a wide contact with the maxilla, whereas thepreopercular soon separated from the maxilla itself inthe Parasemionotidae (Lehman, 1952; Olsen, 1984).
Apparently, upper caudal fin rays occur indepen-dently in both lower actinopterygians and neopteryg-ians (Gardiner and Schaeffer, 1989). This character,however, is attained in different ways in one group ofgenera (Saurichthys, Australosomus, Pholidopleurus,P e lt o p I eurus, H ab r oic ht hy s, and T h o r ac opt e rus), thenin a second group (Cleithrolepis, Perleidus, and Lu-ganoia) and, finally, in the neopterygians. In the firstgroup, in fact, the notochord is straight (or almoststraight) as far as the end of the body axis and thedorsal body lobe is not present; therefore, the tail isexternally symmetrical, with the whole dorsal lobe ofthe caudal fin consisting ofelongated epaxial rays. Inthe second group, because the dorsal body lobe is veryshort, there are only ten or fewer epaxials. In this lattercase, the vertebral column is probably slightly up-turned in the ural region. There are few (ifany) epaxialsin neopterygians where the vertebral column is moreremarkably upturned (homocercal tail).
Thus, in Node T of Gardiner and Schaeffer (1989),
JOURNAL OF VERTEBRATE PALEONT'OLOGY, VOL. 12, NO. 3, T992
tlne Peltopleurus grouît (including Ilabroichthys) can bedistinguished from the Perleidus plus Cleithrolepisgroups by the number of epaxials.
If Gardiner and Schaeffer's (1989) phylogeny is ac-cepted, the large epaxials have appeared at least twice,in Saurichthys and then in the Australosomus group.
A deep flank scale row is common in taxa aboveNode O (Gardiner and Schaeffer, 1989), and it seemsto be related to an externally symmetric (or nearlysymmetric) tall (S auric ht hys cost as Etamosus, Austra-Iosomus, Pholidop leurus, Cephaloxenus, Peltopleurus,Habroichthys, Thoracopterus niederristi-whose hy-pobate caudal fin is probably related to glíding-Lu-ganoia, and a few neopterygians). Thus, we think thatthis feature may have repeatedly and independentlyoccurred; it is possible that it was related to the highswimming speed inferred from the elongated andstreamlined body shape of most these genera.
In Node T ofthe cladogram by Gardinerand Schaef-fer (1989), this character appears only in the Pelto-pleurus group, which includes also Habroichthys.Therefore, it can be considered as a synapomorphy,distinguishing the Peltopleurus group from the Perlei-dus and Cleithrolepis groups.
Perleidus altolepis shows a well developed coronoidprocess (pers. obs. on specimens from Ca' del Frate,uppermost Ladinian). Thoracopterus also has a coro-noid process, though it is less developed than in Per-leidus. We think this process has been achieved in-dependently in the two genera (see cladogram below).
A few other characters, derived or not, are sharedby Thoracopterus and Habroichthys. They include:preopercular narrow in the dorsal region and with awell-developed infraorbital process; supraorbital sen-sory canal very reduced, ending in the frontal; skullroofbones often fused; very large otoliths; absence ofparietals; and sensory canal on the dermopterotic run-ning in the middle of the bone.
The presence of unpaired neural spines in Thora-copterus is interesting because it is considered a hale-costome character (Patterson, 1973). Apparently, out-side this group, this feature has been found only inAustralosomzs and Birgeria (Nielsen, 1949), Moytho-masia (Gardiner, I 984) and, Pholidopleurus typus (pers.obs.). Both Australosomus and Pholidopleurzs belongto Pholidopleuridae, with peculiar ring-shaped centra,while Birgeria and Moythomasia have no vertebralcentra. However, we do not believe Thoracopterus tobe closely related to any of the aforementioned generaor to the halecostomes; possibly this feature is morecommon in chondrostean fishes than known so far.Furthermore , in Thoracopterus, unpaired neural spinesmay well represent a specialization to provide alargerinsertion surface for the powerful muscles involved inthe run-offbefore flight.
Unfortunately, owing to the compression ofthe spec-imens of Thoracopterus, it is impossible to determinethe presence and the evolution ofother characters suchas the posttemporal fossa noted in Perleidus by Pat-terson (1975).

:an beolepis
is ac-wice,)up.bove,ems:arlyttra-rus,hy-Lu-lhatrtlyLightnd
Lef-to-ys.tY,ei-
id
t-r-ì-).d
a
II
TINTORI AND SASSI_TRIASSIC GLIDING FISH 27In the light of the new data from our specimens,Thoracopteru.î cannot ue inctuaeà
-iri?" perleidi_
formes (perleidus eroup of Gardinei- a.rA Schaeffer,J99gl, as_proposediy Lehman (1966,1979) and Tin_tori and Sassi ( l9g7)._.We consider Habroichthys the closest relative ofThoracopterrs, borh r,a"i"gîp.e;o;;;;l;; that is nar_row in the dorsal reeion, but still ti"A-àìi" posteriorend of rhe maxilla. it " ."rt oi in" i"iirii"urus group(sensu Gardiner and -Schaeffer,-i 9àò; ;í';heir sisrer_group, with deep flank scale_ro.frl uíOu infraorbitalprocess of the preooercul.ar.as ryù6;;;hies. FigureI illytlatel our inierpretarion tf N.a"-iif GardinerandSchaeffer (1989), with th; _"ì;;;i rhoràcip_
DISCUSSIONExtant flying fishes are common in arì the tropicarand temperate seas,- both near urJ ofrfro.", and in afew tropical rivers. This p"""ri- _ùJàirif" t achievedin differenr_wavs.
llr^eig "i" _.;ùjJn" uno biplanegliding and flapping ..flying" flrh;;.1il;" fossil record,"flying" fishes are- rare, apparently limited to a shortrime (carnian and lroiigi,i. ò;ì;,;;'senus, Thora_copterus, has been found in the fuestern Tethys. Wedo not think this is due to l""o_pf"t"nlss of the fossilrecord itself; all over the world,^dozeris of Mesozoicand Cenozoic localities fra"e yieiOed lù" numbers offi sh genera from di ffere", """í.o"i Jii.' r,,ran"rmore,it is curious that flight *u. u"frìà""J fi pri_itive ac_tinopterygians when ttrev were-ar.*oí declining andulT::, complerety replaéed Oy n"àptÈrvgtun..
I ne answer 1o why fishes glide haj not yet beenfound: aerial feeding-can be "*itua"al"cause controrofthe direction is sliÀht a".i"g tÀ" ntgiilìruuus, tg32).Besides, the tooth-row pattern on both jaws of Thor_acopterus magnificus leads us to supposé that this fish:^.*
,9 catch prey while swim-i"i'up*urO. This di_rectron allowed in f-c,t the widest-polsible Cap;, ;;_pecially if the mandible coutd ;;; 6" j;;"red much.This predation partern is also ;cÀ"d L"r,rr. presenceotlhe hypobaric caudat fin *h;; À;Jriln rvu, ro pro_
yi.de an upward push during th";;;ing (Beiles_Isles. 1987) orheithan propèlling the nsfr Auring tnetaxi. Feeding during the ìaxi .""_-, ;;-*"1 impossible:the fish was angled upward *friG _""i"liorizontally,and its clear gape was natrow. On thé other hand,during both taxi and gliding, tn" _"rrf, had to beclosed in order to reduòe arie.
--
Flight could be used to "róup" from predators. Infact, one of the main predaro.. éigriJi"gofishes is Cor_yphaena, the dolphin fish, which if.o Ép, out of thewater in pursuit of its prey (Rayner, f qdOl. Further_more, gliding fishes ofren siart their óigúi:"rt in fronrofboats andships, escaprng fiom a perceived predator..P:edators_largerlhan"Th"oracàpt"-r^i",urelyfoundin Carnian faunas (criffiú, l9?t; pe;.;ts.). Norianf_au1as show large (up.to 165 cm) sàiriiiìnv, and Bir_geria, which were able to pr"y or, _olf of tfre smatt
fishes collected in the same localities; in any case, foorremains found in rheir gut. u.";;riiy identifiable aPholidophoridae and y".y v.""g -ia'ratepiaotus
ornalut -(p."!.obs.). Ac_cording ío trrÉÀó.pnology of thei:body (Tintori, 1990b), ""ítnirliiiiíùhys nor Birgericappear as fast cruising swimmers simiia, to the moderrCoryphaena, and thèy *".. prluuiliv no, capable orpursuing Thoracopterus fo1 al,""c;irà""". w" "urrrroìexclude, however, that other n.fr"r-pr"V" a on fhora,-copterus.
Alternative n"r^o_,n"r1to escape can be proposed,at least for the triassic gliding nr_ii"r. Éro"oetidae sea_sorrally migrate to warm wateis (Bruun, 1935). Severalother,species lay gggs-9n seagrasses, near shore, eventhoush they usually live in;;;;"; (Bruun, 1935;P".,il, 195g). We can tryp"tt"iirà a-Jimitar behaviorfor T h o r ac op t er u s ̂ry a g i ífic; ;;J;;;ed specie s; rhepaleogeography of ttre siies ,""-À, io i,rppoa this hy_pothesis. In fact, small .p""i_"rx u.à fir.rrrA in theCalcare diZorzino,which *", a"p..it".ì in rather smallbasins inside a huge carb.onare ú;;À;;; larger speci_mens come from theArgilliti diRiv" Oi.S.fó a"-p"r_i1e{.in a large gutf, pos-sibtt,;.;;à;; took place inshallow waters llaqoons i"riA" tÀ" "uìU"'nate platform),where vouns tivà.untii ,h"r";;;;ffi fur maturiry.Then rhey migrated to more'of;;;;ì".r. Thoracop_terus martinzsi is generally larger than- f. *ognifuiír;in facr, it is found'in the seJiir-e";;; a large anoxicbasin. We can assume that shaiiow,ffienateO settingswere rather far away, since no allochif,onous benthicorganisms have been F_""d trrere.-ii ifitows rhat our
:fl:;::*r can be considered _"_rv "à"lts livin! ofl
We think that eliding fliglrr-may be retated to longdistance migratiol. n"i*r"lìetió'oóì"ì"o out rhat insome marine mammals of interméiiui".ir", a kind of^,Tqlr"
is presenr. ro--o"" r,àìi;;;;. and half inarr seems to require a lower energy than simple .*ir*mrng; the same pattem appears in-tuna and other fast_swimming fi shes, including C riii n" ii " "]:rrr"r" advan_tages are apparently referred tó'ai.iv ,t.rct size con_straints. Gliding fishes are smaller than the above citedanimals, so that the energy saving could be almostnegligible if they swam hÈé a t.rnal i-o�r"u"r, an ad-vantage comparable to that.obtained by tu.g". lrùì,a small one, such as-a gliding nrfr,-_,i.r reduce thenumber. of complele leaps. reíral"irg-ìn tfr" air for alonger t ime. This al lows the reducl ion-of i t , involve_::ll
with viscosiry and surface t""rio", which aremaxlmum when the fish goes out or returns into thewater.
_,Y_oOu of.Gìiding_Extant gliding fishes can be mono_plane or biplane eliders 6tubbs, jSli; B".fin, l95g).The former mode of flight i. *;;É. ;íì ."quirine urun on the water surface before tak_À ili 6axiy. zxo-coetus is a rypicat monoplane glid;;.é;ìhe conrrary,we will consider Cvoselurus drep;;;,ing the most:"91y:-d .ryav of gtiàìng, the biptane one, for which ataxi (Hubbs, te32) anà the u; ;i;;i;it fins is .,ec_essary.

278 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 12, NO. 3. ]gg2
-Thoracopterus do not share any characteristics withGasteropelecus, a flapping ..flying', fish, which showsa peculiar modification of the endoskeleton of the pec_toral fin to accommodate^the very expanded ..flyi'ng,,
lyryles (Hubbs, 1932). On rhe othei hand, faveÀe(1974) shows a peculiar linking structure between thepectoral fin lepidotrichia of pantodon buchholzi, ri-_sembling that of Thoracopterzs. Taverne regards thisfeature as a stiffening of ihe fin itself in order to im-prove the flapping fligbt. However, because Thoracop_terus scapular region does not show any modificatiónwe can only compare it with monoplane and biplanemodern gliders.
We will not know with certainty whether or notThoracopteruJ was actually a gliding fish, but we cancompare as many characters as possible with those ofthe modern Exocoetidae, to achieve plausible conclu_sions.
We think that two main features have to be foundtogether to determine whether a fossil fish was a glideror not: high ratio of fin length to standard tengtl (upto 6O-70o/o) and an associated hypobatic caudal fin with31 rp-_p_el lobe length 58-75o/o the length of the lowerlobe (Hubbs, 1932). A third character"is the pattern ofthe.sensory canal system; in modern Exocoetidae, thecephalic sensory system is reduced and the lateral lineof the fish runs very low along the flank (Tretiakov,1938). The lateral line position-has not been observedi2 Tloygo4terus (G.iffith, 1977; Lehman, 1979; pers.obs.). S^chrJltze (1966 and pers.'"o--.j found .p""i_mens of Thoracopterus niederrisli show.ing two laterallines:.one dorsal, on the third scale_row below the dor_sal midline, the other in the middle ofthe deepest scale_row. The reduction of the head sensory canals, how_ever, is visible in all of the three wel_Lnown speciesof Thoracopterus.
Other supposed..flyirg" fishes are simply long_finnedspecies, such as lcarealcyon malagasium'(Beltai,l 9g4).Regarding lcarealcyonwe note tliat the dorsal and analnns are very long (Beltan, l9g4:p1.3), in contrast to theLloÍ"9fr of unpaired fins in both extant and fossil"flying" fishes. Furthermore, the posterior outline ofthe caudal fin is not preserved (sèe below). We thusthink that these specimens may well be related to oneof the common parasemionotid genera in ttrat fauna,perhaps to Watsonulus. Chirothrú libanicuspictet andHumbert from the Upper Cretaceous oilebanon hasbeen shown to be a pelagic fish (Abel, 1906) on thebasis of its peculiar pectoial fin structuie;fuftúermorethey are smaller than the pelvic ones. Cóncerning theC^oenozoic, specimens referred to Dactylopteriformes(Gayet and Barbin, l9g5) and to Exócoetus are re_corded from the Eocene (Mùller, l9S5). The formerauthors interpreted the dactylopteriform prevolitansfaedoensis Gayet and Barbin, l9S5 as;;flying,, fish,,!9-"gtt nothing in the specimen could be OescrlhA aia "flying" structure. On the other hand Hubbs, as earlyas in 1932, believed Da ctylopterus *u, ,roià flying fislqmostly because of the perfect symmetry of ifs càudaífin. Many living long-Ànned fishes, inÉrràirrg Dacty-
topterus, use those fins for purposes other than flying(Lo Bianco, 1909).Most of our work has been carried out on Thora_
copte-rus magnfficus, the best known species. The sameconclusion will then be applied in part to the otherthree species.Axial Skeleton-The modifications of the neural
spines of the caudal region give rise to a wide area formuscle insertion. The shape of these enlarged spines(Figs. 7R_, 10, I l) contrasts with that ofalmoit ail òtherfishes,_where they are elongated but thin. This may bèrelated to the different function of the caudal fin infish^es that glide and those that do not. Gtidirrg nst "s,in fact, need an asymmetrical stroke of the taiía"ringthe taxi, _the upper lobe being ."i.iìn" water. Theenlarged haemal arches_and thé hypurals have groovedsurfaces providing a stiffer support for ttre tepia-otricfrlaof the lower lobe.
Paired Fins-A few characters are interpreted as re_lated to .gliding capability: large size, shape of the firstlepidotrichium, presenceof thé anterior wing_like pio_cess on the proximal segment of each lepid-otrichìum(Figs.
]{, 8, 9), position of the ornamentation, pres_9nc9 o^f the scaly inner lobe, and position of the iaàiats(only for the pectorals).
Saber-like first lepidotrichia in both the pectorat andpelvic fins point to a biplane flight, as shown by Bruun(1935). The wing-like process ofthe repidotrichia couldhelp-in sliffening the fins when spreadl through it, eachlepidotrichium gave support to the previoùs orr". Asimilar structure is known-in nantodoi lfaverne, 1974)and was interpreted by that author aì rehted to thésupposed flapping flight of pantodon. The fact thatboth pectoral and pelvic lepidotrichia show this pro_cess clearly shows they were both involved in the flyingprocess. Possibly the peculiar ornamentation patieri,on the lepidotrichia also played a role. ft may travebeen useful either in the àt li" which case only theproximal parts of the lepidotrichia remaìn in contactwith each other) or underwater, when the fish speedup to reach the surface and begin the taxi. In the làtter9a9e, in fact, the long fins weie probably closed andheld tightly against the body to reduce A.àg. A similaiornamentation may have served as a helpful lock. Thetnner scaty tobe provided an even better lift to the openfins, perhaps also reducing the ui, t";b"i;"ce betweenthe body and the fin itself.
Because the radials and the very proximal end of thelepidotrichia are ventral to-the scàputoco.acoid, up_ward vertical movements of the pecìoral fins are'tó_ited. This is a furtherelement to "i"t.rO" lapping flight.As we have seen, all the structures ;ith" paired finsseem to be related to gliding. The fins couid be heldvery stiff when spread, and their upward movement
was. strongly limited above the horiiontal plane._ . Size and position of the paired nns furttier supportbiplane gliding flight in Thoracopterr" lÉigr. tiqfl.The-relative length of pectoral fins in the monoptaneand. biplane gliders is similar, being slightly more than7Oo/o of the standard length; this latier is"lérre.atty more
L

han flying
n Tltora_fhe same.he other
e neuralarea forJ spinesill othermay bel f i n i n! fishes,during:r. Theroovedtrichia
as re_Le firste pro-rhiumpres_rdials
I andruunouldeach3 . 4)74)theihatrro-ing3rnlveherctederrdlrIC
nn
TINTORI AND SASSI_TRIASSIC GLIDING FISH
:?1îJ i!,#ft#:re sjgnincanr is rhe rerarive rengrhg",nirl';.
'r; ;;;|:;):n Iî1,?,, ri1y
or be ,,"o-in
;;n",;5:.1xfl i"#*, o;i ffi ;àt:i:i l!;": {:A:i*:íiini:$ii,\î{#:i#í;^::"ri".1ixili:.'x1',:r",cy;;d;;;""??Jx.'JllJ"ti::"f n::l:X;i"r}!:(,"! :{ {;i!7 ;;", #3J: Ti t i v e À o i Jpi ài., *i t r, a pe r _to the 'h.;;;:' ;i"ili1':'."r33%' rhis is simplv iueor .turrau.à'i;;;;.
tne pectoral fi n s t h emser "es i3l "7,
^.u:^:,!: :;,":#;"" {: !' r' u s, :v e n o red a po s i r i vethe fact tnl rfr"."i.the
pectorat fins. This ir-;;;,i;square of the lengrh.l-91,'h:
"wings" grows with rhe
its cube' Ti;;ri""",j*If:':'ih" fish weignt grows withbody d oes ili,;;Í il?TJi.i5J:y"?ff1T,Hlli
FIGURE 16. Thoracowi th respecr r"' à'ri"*'J' iirz-s' useful gape wi r hu pwa rd .",,, i, i i'i i,ì ; [::J T#jì%i*F;T'f tT' il llll
:?t:r:' {: "KI?f': ff 1î"p: beca u se or th e smalee"tuuírvl'i"n';#1iid the larger tir" ortrt" .pJ"iì
tr#:ffi ,TlÉjlil,j,:"jf,isjt+,;1,",.#:iJlmaximum l"nnrl
tertainlv the tepidotri"hi;-;;
fi :.:f glT:tr".Ti:$i:l;$jdT:,ff l+*"7more effecrirr" tlll
a greater musóular -ur., ir'�rarge flying 6r6.1u11
tl9er supporting area inontogenetic data ,
t'or Thoracoplerus niederyisti,
Í"rqlùil.ì;;,"* available' but. al an averase snn s i s a bo u t o j "2,'ffi-H,i:t}: i_"àr;';f rÀ;;;
o9..). tr is possibte^,_hu: il, rurd"i p"".ral fins vriflffiii.offi ill':::: :tu h "u;; ;;i; io ve.i n g, whf f;:r:1tiil#!ii!,,rf=iii'.;;:i;x;,:f;r;capability. rr," o"l-?r
the fins' without.loss;î;il;,;;;;;;;,ì;;::l:;:T:îff#::.$iil;k;:Í^ "TI" rff:: ;'" :;; ;: "T ! :: : c ot ! e r u s m a g n ffi c u s, as wlîî.g.ry;;;í;;'Ji#:irr;i:91i",.'1,.#il jl*#;;'!.'f, :'ff 'J|f,":n
: fl ît' p r' " ó. r i à iij Ti' " ro.,, ",, pt.8olo. Furtheìri;;::^t:t 2l.o/o^and the tater of abJratty strortei ;nJ;;::
pelvicfns become proporrioralso said to ii"rn" Ì"*
as fish size increases. This r.t nus. the propoftioc-ase^of cypselurus (B.ruu. I93j.length ""d'fi ;;;;]l -?"tt""n- -pel vic uno p".toruii,for our N..i";";:::"{::' we observe a trigt .aiim asn trt i u i ài,l' rí"ir'ill'
wtth 4 1 o/o ror rn o ro íipìàiiabove rhe ";i;.";;,j:i
I marrinisÌ, respectiveti. wei. Another r.;;#'#:l!rn binl-an es @ [o/os.is the posiri;;-;1",í:'::1"9 t9 a biplane tvpe orglidingcopterus, rh;;;n;';:"1:i" fins' ln specielgi 7i;';;:srandardr"";;..;;;i;:i:Tffil#ou.î,:"qJ,r,í,,r,1,ff
7 a
a a
E A
r
H 4 a
:ri: '
o e ò
P - E o a@ o 3 : f l 3 9 P
magnincus'aorcr-nìiti;ffi .H.rJ.*f i:J,L'ff irJÍ.iiiiihfi ;5ii?r1.rl:.*Í,xff :

280 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 12, NO. 3, 1992
oo
. 3 4
-o
t i a a
9 9
a a
7 0
j 6 a
F
9 4 sI
3 A
2 A
t a
r"t ax xr x l r a a
a o o o o
x _ a! a
l r t a
^ x a at a
o 6 t s o o o $ o o o o o a o o o- N O î n @ N O O o - r u O f 0
J . L .
s o o o 6 r o o o o o o o 6 , 6 t o 6 l- N O r n 6 N 6 0 6 l i N m r n
c l
a
FUn 7 A
, 3 4I
tf)
à a a
x
A 2
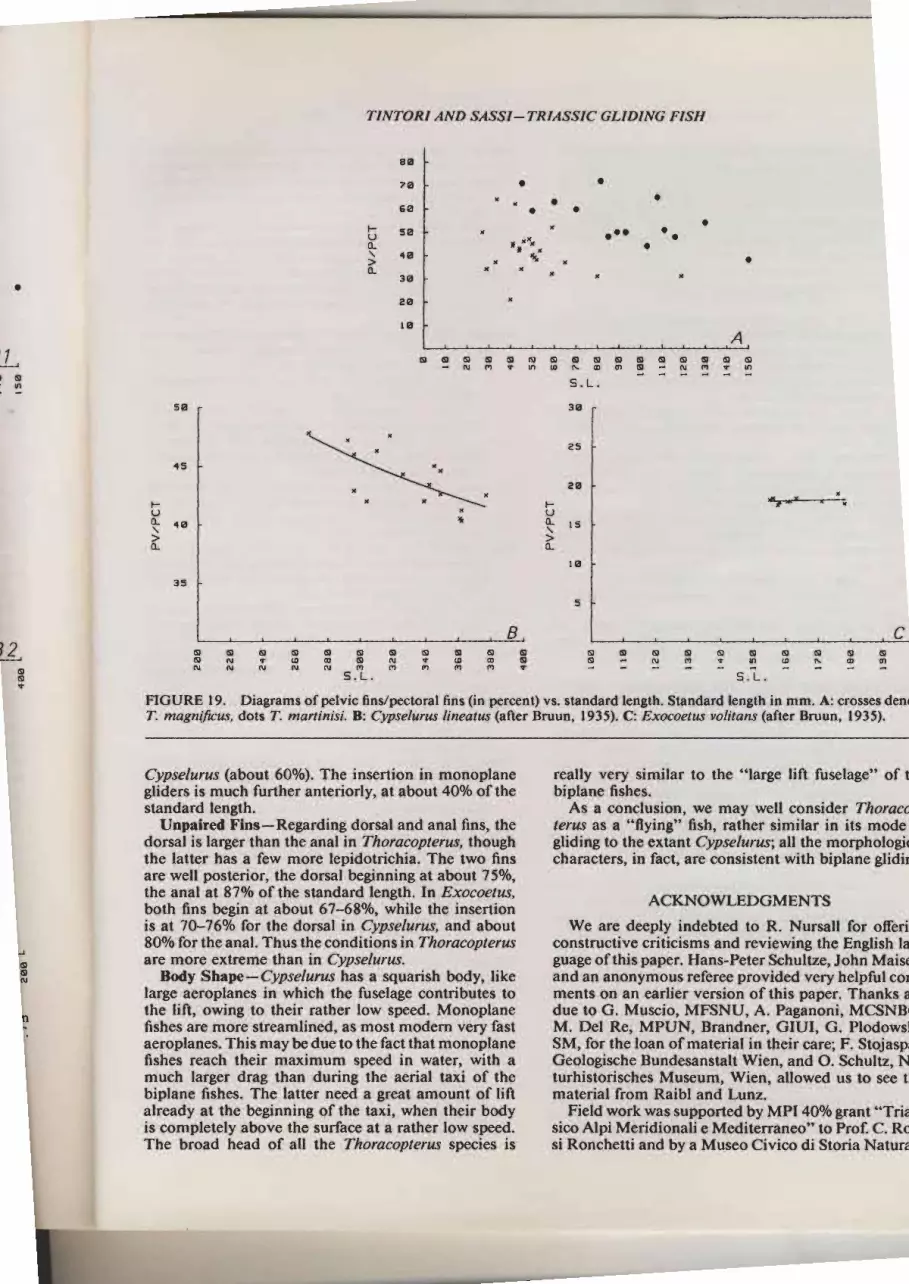
FIGURE 18. A, diagram of pectoral fln length/standard length (in percent) vs. standard length. B, diagrams of pelvic finlength/standard length (in percent) vs. standard length. Standard length in mm. Al, B1: crosses denote T. magniftcus, dots ?nmartinisi A2,B.2: Cypselurus lineatus (after Bruun, 1935). A3, E.3: Exocoetus volitans (after Bruun, 1935).
6)N
o o o a o a s a o 6 | sO N r @ o $ N q @ @ AN N N N a
- q O O O o ÉJ . L .
s o 6 t 6 t E $ o N 6 t or u î @ o O N î @ @ OT N N N m O O O O 9
5 . L .
2 A
)óF
ù
j
ó r s
0_t a
o o s o o( ! N o o O
N
o o o o sO i N o c
q o s a o a o o o o aS d N o î n @ N 6 o A
- ^ : N5 . L .
TINTORI AND SASST_TRIASSIC GLIDING FISH
a a
n
0_
R 7@ OÎ o
E : ! i i iS A6 o
f-U
t 4 aa_
FQ
t r s0_
t q
O O
f l P T }- ; r ' ' O
$ f i : f ;t 2
os A O
@ @ ;r . r O t - : '
, . 1 .t. magniJtcus, dots T. n-
ryantntsl B: Cvpselurus rtneatrs (after Bruun ,lÍÍ{3 t:Tjn. Standard length
7*iàr'ìl;r, o:t:?T: "f.pglylg fins/pectorar
fns (in percent) vs. standa*r rpn^*L o._ .
- s'i '
- : : In a n t n t s i. B : C y p s e t u r u s t i ;; à;;," ;;à;i ilil:ij""Ig ppjn. S ran dard r t,nearus (arter Bruun, u,;. ó,.";;":;Zi;:;:;i:::1,:_g. A: crosses denorJ/. C: Exocoetus volitans tufr". nruun,",r"rxì
oaN
n
Cypselu.rus (abour g.OU"l. The insertion .glrders is much furtr,", í-r^*l^:i:Ì"'î"1 tn monoplanestandard i;;;r;."tn"r
anteriorlv' at about 4o/,;i;;;
..y':ilJì:l#î;::t:ting dorsar and anar nns, rhet he latre; ;;;;' dr:
t:"^ 1"?l i n .T h o r ac o p t e rrzs, rh ough
are well p.r;;;;: :' m ore l?idotrichia. Th; ;;;ffi i
rhe anal ;iA;t"ùiî. dorsal beginning at about 75o/o,
uotrr nnsìelir"uì',ln" standard lenethl h È;;;;";;::ts^at 7o-760/o 15, ,t'l",lt
67-68%, ritrite ttre inserrion8 oo/o ro1 h ; ;;;;: i'; :' :-?':" 1 in' C vp s e t u r u-s' an d a bo u tare more exreme ,ilt-'ft
condidons i" Ti";;;";;""r;;. Body sh;;;;;lin,1n c)tPseturus-targ" a"rofiJni "-ilo.!ÍJ!?t.hul u squarish body, likertrJ rirr, oii;;;'
',,n::'ll,tht fuselage con tri buies iJfishes are -;;: r"#::Ti-Jatler
low speed' M.";;l;;;u"roptun "r.
.f1,ì r"il1Î1 13o' .as m ost m odern very fasr
fishes reach d;;'H:-: clue to the fact thar monoptane
t#, J mi JTf; "Tr"T{frE ti ;"�;"f lì :it:already ar;;; b;;i.l-1::' 199d u sÍeat amounl of tifr
f*^.r_r*'",' "ùil'iil:'.:ill: :1"j',Lî:x,tili#r ne broad head of al the Th;;";;;;;;:;, species is
;fi1J"$?tsimilar to the "large lift fuserage,, of thAs a conclusion
Éi'íjl*{;i#{l':;8;t},#":ffi 'li'fftr:i
ch araclers, i n ract, are co n si srerì ; ffifri#j, !iLf, fl.ACKNOWLEDGMENTS
".ffJ#*":*il"ls:t,:q ro R. Nursau for offerine:ffi T".fi :f ff diir*lIril!:"1,i,?#th',ilil:}:ments o' u;';;;;;:lei:: ero^vi'dga "erv t'gnr;il;#
$*f**gl*l[l$Éfi !+Jt:àitr{i#;Geot ogi sche' ffi ;; jl.llÎÎÎl jl th eir care; I: i;;;uilí;rurhistórisci;-ili;r."".ralr Wien, and O. Schulrz, Na_-X,,:îfl *;.-"ili;ì?,il'#l?' arrowed us to see the. . ,:ru wOrI( WaS SUnlrrA,qiM;;;;#fi ?:i":i,ii#IJ"1e,'f F::?:;k::sr Koncherti and by a Museo Civicoii iì.ri" Narurale

282 JOURNAL OF VERTEBRATE PALEONT-OLOGY, VOL. 12, NO. 3, ]gg2di Bergamo fund. The Museo Friulano di Scienze Na_turali operated with- grants rrom cNn à c. vtuscio...Severat people, orhér than "",h;;;;iìecred the fos_sil s.peci,mens and presented them to _,i.".r_.. C. Cor_selti and D. Basso orovided ,;;";;;; r Cypseturus.To all of them our.i.r".." thanis. pfràìo. Ov G. Chiodi,final drawings by D. Cortu urrà-S. ii"""?r..
LITERATURE CITEDAbel, O. 1906. Fossile Flugfische. Jahrbuch der Geologisch__ en Bundesanstalt Wien 56:l_gi.-
-" '^'* '
Bassani, F. 1g92. Sui fossiliL suìl,"eia,degli scisti bituminosidi Monte petrine presso Giffoni Vuff""iìu"u in provinciadi Saterno (Dotomia priryro"lei fri"_"i" S""ieta ltalia_na Scienze dei XL, s.Z, S:i_Zí.
1895. La ittiofauna della Dolomia principale diGiffoni (prov. di Salerno)- p;ì""-.;ì;;;;phia rtalica t:169_21o.Belles-Isles, M. 19g7. La.nage etl,hydrodynamique de deuxagnathes du paléozoique: Araspis màcrotubercurata efpteraspis rostrata. N""è, ran.ul"i niióJér"ei" und pa_^ . ìàontologie, Abhandl""g"" rir:?):ià.Beltan, L. 1984. A pro_pos d,un poisson vltant biplan def 'Eotrias
du NW à" ry"g"ry."'".
";;;;;í;;", matagas_tum Beltan. Annales de la-Société é;Gíq"" du Nord53:7 5-82.
Bertin, L. 1958. Modifications des nageoir es; pp. 74g_7g2t1P; P Grassé (ed.), Traité a" Z.ri8g*, ".1. I 3. Massonet Cie., paris.Bronn, H. G. lg5g.
.Beitràge zvî triassischen Fauna undFlora der bituminósen Sóni"r", "à" ii"fur. Neues Jahr_l;:n
*. Mineralogie. Georogìe ""a iiiàor,rologie l:l_Bruun, A. F. 1935. Flying fishes (Exocoetidae) of the At_Iantic. Sysrematic and biol.ci.;l;t;d;;. Dana Reporr6: I -l 06. Carlsberg pounaati"on, è;;;h"r"".costa, o. c. r 862. sr;di s;;;;i;n"ìiíài,ii",ui der Regnodi Napoli. Atti Regia a"*d"Àiu i-"f,i)"'Nupor i,44 pp.Gall, J. C., L. Grauvogel, and l. F. i"rr_"íl 1974. Faunedu Buntsandstein- V. I-es poisson-s^'ilisìi", de la collec_tior Grauvoget_Gall. an"ur"r-a"-puìéà"iàrogi" 6O: | 29_147 .
Gardiner, B. G. 1963. Certain palaeoniscoid fìshes and theevolution of the snout,l" "'"ti""pi".ygiàs. Bulletin ofthe British Museum (Natural ffistory.l] óeology g:255_
1984. The relationships of the palaeoniscid fishes,a review based on _n-ew specime r" if ùi*ia and Moy_t h o m a s i a fro m the upp".-o""oiiu"" o i''#" *"* A u s tra -lia. Buletin of ttre eritistì M";; N;tìrrat History),Geology 37:17342g. -----^ \r !s!!
1988. A new Cleithrolepl.s from the Triassic of cen_tral Cyrenaica, northeast Lrbt", !|.'líg_zas in A. Et_ftffiff,;i1eds.), suusu.a"à'iarf,iortrutig.upnvìr
and B. Schieffer. l9g9- Interrelationships of loweracrinoprerygian fi shes. zo otogicit i i".ài "rrrr" r_inneanSociery 97:135_1g7.Gayet, M., and V. Barbin.. 19g5. Cephalacanthidae fossiledu priabonien des environ, d. p;;;;;;frì"rr"l. Bullerinde Muséum nar ional d. H ir,;;; ;;;;;il.ì' ó o, ru r_rr r.Griffith. J. 1e77. The
lnoerT.i":;;;';r;; Èom potzbergbei Lunz, Ausrria. zòàrogi""iJo"_Jàiìn" LinneanSociety 60:l-93.
Hubbs, C. L. 1932. Observations on the flights of fishes,wirh a sratistt"l:1"Or.f ttÉ filgtt oitne Cvpselurinaeand remarks ",1 ,hî ""of"tio" ? ir'e flight of fishes.papers of Michigan e"uo"Àv ori"i;;;"., Arts and Let-t e r s l 7 :575_611 .Kner, R. 1g66. Die Frl9he der, bituminósen Schiefer vonRaibl in Kàrnthen.
_Sitr""grU"iJit"'jer Akademie der_ Wissenschafren in Wien sii i ;;ì i ;;
- '
Lauder, G. V., and K. I .Lie;.
-íifÉ. The evolurion andinterrelationships of rhe u"ti"àpióg*"n fishes. Bulletinof the Museum
versitv r s o' s s- lé zl om parati ve zo ol ogv' Harvard uni-
Lehman, J. p. 1952.- Etude complémentaire des poissonsde I'Eorrias de Madagasc;;. ri;;;lt;;svenska veren_skaps A kadem i en s Ha*ndlirg" r-à, i1)"ór1953. Erude d,un perteiàisO" frr". A" fufadagascar.Annales de paléontologie :9:H g.^'^-"1966. Actinoîr"ryÀlt pl. t_iqz inJ. piveteau (ed.),Traité de paléontoloeie] vóli+ò1. ir;#;" et cie., paris.1979. Nore. suitei poirràí, àJîìiu, de Lunz. r.Thoracopterus Bro11. Annalen Oes Naturfristorischen
- _Museums in Wien g2:53_66t.Lo Bianco. S. 1909.
, Notizie biologiche riguardanti spe_cialmenre i1 periodo_dj _","ri a?lrri,ur" o"gt animalidel golfo di Napoti. Mirpift_À*"àur'i"r roorogischenSrar ion zu Neaiet r9:sr3_zài l
' r gqo u!moy- r homas, J. A., and M. B. Dyne. 193g. The actinopte_rygian fishes from the ro*". óuìUoíi"dro.,. ofGlencarr,Eskdate, Dumfriesshire. T;;"-r;;;."J,n" Royal So_- -...:i"ry of Edinburgh SS:qyJ8l.Mùller, A. H. 19g5.*-Lrchrbuch j"', putaoroologie. Band 3,Teil l. 2. Auflage. vps Custav Fis.i"iîJ.,"r, Jena, 655pp.Nielsen, E. 1949. Studies on Triassic fishes from EastGreen I an d. rr. A u s t r a I o s o ;;, ;;;" ; ;, r'r, *. Meddet et serNybelin, O. 1976. On the so_called postspiracular bones incrossopterygian s, brachiopterygiàns a-na actinopterygi_ans. Acra Resiae Societaris ;";.;i;r;; et tiuerarumGorhoburgensls t 0: l_3 l.L.,fsen. P. E. lgg4. The.skull and pecloral girdle of theparasemionotid fi sh Wat s onu I us àiài'itriao from theEarly Triassic Sakamena prouo ó? ruruJàgur"ar, withcomments on the relationships àf rn" ùfà.tean frshes.Jo u rn at of Ve rr ebra te pur "o"t-J Jev" i", o' ài ) g s .Patterson, C. 1973. I233_30i i"t' n df#i,lil::ì|:"J ii,::]."'i:T,":,;terson (eds-), Interrelario"rr,ipr'o"i ;ffi;. AcademicPress, London.
I
1975. The O:"ll_:,":" of photidophorid and lepro_lepid fishes, with a review of the acdfop-terygian brain_case. philosophicat Transa.ti"", .iiirl"iàyd Society of_ London 269:275_579.Kayner, J. M. V. 19g6. pleuston: animals which move in^ - wnter and air. Endeavour l0:5g_6a.Schaeffer,B. 1973. Inr207 _226 i, ". ". 3[!11,ffiI'r $lH:,i.ff:lt,l:l:rerson (eds.,), Inrerrelatio"stipì' ;i ;H"r. AcademicPress, London-Schultze, H.-p. 1966. Morphologische und histologischef.Inrersuchpnsen an S"h;p;;;;;#J"r,", Acrinop_terygier (Ubereanq von Gànoid_-r" nì"or"huppen).Neues Jahrbu"t, È, Ceotogie-unOiuù'Jrrrorogie, Ab_^ handlungen 126:232_314.
- '--- ' s'qv'� i
slensió. E. A. 192 l. Triassic _Fishes from Spitzbergen, Vol.l . A. Hotzhausen. Wien. 30i ' ; ; " " " ' "v" '

TINTORI AND SASSI_z'RIASSIC GLIDING FISH 283
Taveme, L. 1974. Sur une adaptation au vol des lépido-
triches pectoraux de Pantodon Peters (Pisces, Osteo-gtàssiformes). Revue de zoologie et botanique africaine
88:221-223.Tintori, A. 1990a. Dipteronotus olgiatii n' sp' (Actinopte-
rygii, Perleidiformes) from the Kalkschieferzone of Ca'
dti Frate (N. Iîaly). Preliminary note' Atti Ticinensi di
Scienze della Terra 33:.19l-197.1990b. The vertebral column of the Triassic fish
Saurichthys (Actinopterygii) and its stratigraphical si-g-
nificance. Rivista Italiana di Paleontologia e Stratigrafia96:93-102.
and D. Sassi. 1987. Pesci volanti del genere Thora-
copterus nelNorico lombardo. Nota preliminare' RivistaItaliana di Paleontologia e Stratigrafi a 93:337 -345'
Tretiakov, D. K. 1938. Seismosensory canals in the flying
fishes. Comptes Rendus de I'Académie des Sciences de
|URSS l8:487-490.Wade, R. T. 1941 . The Triassic fishes of St' Peter's' Journal
and Proceedings ofthe Royal Society ofNew SouthWales74:377-397.
1942. Triassic fishes of New South Wales' Journaland Proceedingsofthe Royal Society ofNew SouthWales74:377-397.
Received 30 January 1990; accepted 2 October I99l'
,"-- ;Lúi- :- .a-*;L.,.-,a&.-.- _