thermotolerant yeast strains producers of galactooligosaccharides
TRANSCRIPT

1612 Biotechnol. & Biotechnol. eq. 24/2010/1
Article DOi: 10.2478/v10133-010-0014-6 FB
Keywords: Kluyveromyces yeasts, β-galactosidase, galacto-oligosaccharides (GOS), whey, transgalactosylation
Biotechnol. & Biotechnol. Eq. 2010, 24(1), 1612-1619
IntroductionIn recent years, some of the most significant developments in functional food science have related to the development of dietary supplements that beneficiary affect microbial composition of gastrointestinal tract. Such compounds with well documented prebiotic action are oligosaccharides and their derivatives (5, 20, 28). Galacto-oligosaccharides (GOS) are also known as ‘Bifidus growth factors’ for their ability to stimulate the growth of Bifidobacteria in the human body (29, 31). For this reason, ingested GOS are now wildly used as a foodstuff valuable additives, as they have numerous dietetic benefits for intestinal lactic acid bacteria and Bifidobacteria: 1) maintenance of normal intestinal balance of microflora; 2) improvement of lactose tolerance and digestibility of milk products; 3) antitumorigenic activity; 4) reduction of serum cholesterol levels; 5) impr oved synthesis of B-complex vitamins and 6) enhanced absorption of dietary calcium (12).
Enzymatic methods for synthesis of galacto-oligosaccharides and preparation of whole cells biocatalysts are promising alternatives to chemical synthesis (20, 24) since the last one is usually complicated by the formation of unwanted anantiomers. For avoiding these undesirable products long protection - deprotection steps for selective control are required, as well as low final yields are achieved (11). In the last years GOS has been produced through employing the transgalactosylation activity of various β-galactosydases (lactase, EC.3.2.1.23) (3, 19). β-Galactosydases can catalyze the transgalactosyl reaction as well as the hydrolysis of lactose. The proportion of transgalactosylation to hydrolysis reaction varies, depending on the different sources of the enzyme. β-Galactosydases from E. coli or Aspergillus niger
exerts strong hydrolytic activity, whereas β-galactosydase from Aspergillus oryzae and Bacillus circulans possess strong transgalactosylation activity (17). Moreover, amongst other microorganisms, yeasts have emerged as an important source of β-galactosydase, since the yeast enzyme has an optimum pH suitable for lactose hydrolysis in milk and cheese-whey (13, 26). Furthermore, yeasts belonging to genus Kluyveromyces, offer several great advantages such as good growth yield, acceptability as safe microorganisms for human (GRAS status), thermotolerancy, utilization of cheese-whey and higher β-galacytosydase activity than other yeasts.
In this respect the aim of this work is to study the capacity of different yeast strains, isolated from various dairy products (white cow cheese, cow yogurt, cow milk, yellow cheese and melted cheese), to produce different galacto-oligosaccharides and to select a strain with enhanced transgalactosylation activity.
Materials and MethodsMicroorganisms and growth conditionsThe yeasts used in this investigation were isolated in the Laboratory of Applied Microbiology (Biological Faculty, Sofia University) from different dairy products: white cow cheese, cow yogurt, cow milk, yellow cheese and melted cheese (Table 1) and were described elsewhere (9). The purified wild isolates were grown at 400C on a reciprocal shaker (204 rpm) in a liquid Rider medium with the following composition: 20 g l-1 lactose; 3 g l-1 (NH4)2SO4; 7 g l-1 MgSO4 . 7 H2O; 0.5 g l-1 NaCl; 1 g l-1 Kh2Po4; 0.1 g l-1 K2hPo4; 1 g l-1 yeast extract; 0.1 g l-1 CaCl2; 0.05 g l-1 FeSO4 . 7H2O; 0.01 g l-1 MnSO4 . 5H2O; 0.07 g l-1 ZnSO4 . 7H2O; 0.01 g l-1 CuSO4 . 5H2o.
Biochemical analysesThe specific β-galactosidase [EC: 3.2.1.23] activity was measured using chromogenic substrate o-nitrophenyl-β-D-
THERMOTOLERANT YEAST STRAINS PRODUCERS OF GALACTO-OLIGOSACCHARIDES
V.Y. Petrova and A.V. KujumdzievaSofia University “St. Kliment Ohridski”, Faculty of Biology, Soifia, BulgariaCorrespondence to: Anna V. KujumdzievaE-mail: [email protected]
ABSTRACTTwenty five thermotolerant yeasts isolated from different dairy products were tested for their ability to grow on lactose containing media and to produce β-galactosidase enzyme with enhanced transgalactosylation activity. It was confirmed that the most effective strains are those belonging to the genus Kluyveromyces. Two K. marxianus strains - 904 and t3 with relatively high specific activity of β-galactosidase (480 and 500 U/mg protein, respectively) were selected. It was shown that these strains possess a high yield of lactose transformation (hydrolysis and transgalactosylation) (98%) and high tri- and tetra- galacto-oligosaccharides (GOS) yield. More than 50% conversion rate of lactose to GOS was achieved when K. marxianus 904 and t3 were used as whole cell biocatalysts. The composition of the synthesized GOS was determined to be exclusively consisted of galactosyl residues.
1613Biotechnol. & Biotechnol. eq. 24/2010/1
galactoside (ONPG) as described by Ausbel et al. (1989) (1). One unit of β-galactosidase activity is defined as the amount of enzyme that hydrolyzes 1 µmol of ONPG per minute at 300c and pH 7.0.
Analysis of lactose concentrationLactose was measured by the DNSA method, calibrated with pure lactose (Sigma Co.).
Determination of dry weight and kinetic parametersCell dry weight was determined gravimetrically after drying washed cells to constant weight at 1050C. Specific growth rate (m) and cell yield coefficient (Ys) were calculated according to Pirt (1975) (27).
Cell-free extract preparationCells from 18th - 22nd hour of cultivation were harvested by centrifugation at 4500 rpm for 10 min and washed twice with distilled H2O. Cell wall disruption was carried out mechanically according to the procedure of Nedeva et al. (2002) (23). The cell debris was removed by centrifugation at 5000 rpm and 40C for 10 min and the obtained supernatants were used for enzymatic analysis.
Protein determinationProtein content was determined by the method of Lowry et al. (1951) (16). Bovine serum albumin (Sigma Co.) was used as a standard.
Biotransformation of lactoseThe yeast cells were precultured on lactose at the above mentioned conditions, harvested after centrifugation at 4500 rpm for 10 min and washed twice with distilled water. Next, 250 mg wet biomass were resuspended in 2 ml potassium-phosphate buffer pH 7.0 and 0.5 ml toluene was added. The samples were vortexed vigorously and another 3 ml potassium-phosphate buffer pH 7.0, containing 0.4 g ml-1 lactose was added. Yeast cells were incubated at 280C for 24 h and samples were withdrawn at 4th and 24th hour. The enzymatic reaction was stopped through sample treatment for 10 min in boiling water bath. Then centrifugation was carried out for 10 min at 5000 rpm and the obtained supernatant was used for further chromatographic analysis.
TLC analysisTLC analysis was performed on DC Keisel Gel 60 (sheets 25x25 cm) using a mobile phase - acetone:butanol:H2o (4:4:1,v/v/v). After completion of the procedure, plates were dried and carbohydrates were visualized with mixture of solution A (0.2 M 4-aminobenzoic acid, 8 M acetic acid, 0.5 M phosphorous acid) and solution B (acetone) (A:B = 2:3, v/v), prepared ex tempore. Finally, the plates were dried in a vacuum oven (1000C) for 5 minutes. Internal standards of lactose, glucose and galactose at a known concentration were spotted on each TLC plate.
TABLE 1Yeast strains isolated from different dairy products and subjected on screening procedure for production of β-galactosidase enzyme with enhanced transgalactosylation activity
No. Strain taxonomic status
Strain denotation Source
1. Kluyveromyces marxianus t1 Cow yogurt
2. Kluyveromyces marxianus t3 Cow yogurt
3. Kluyveromyces marxianus 901 Cow white cheese
4. Kluyveromyces marxianus 902 Cow white cheese
5. Kluyveromyces marxianus 903 Cow white cheese
6. Kluyveromyces marxianus 904 Cow white cheese
7. Kluyveromyces marxianus 905 Cow white cheese
8. Kluyveromyces marxianus 906 Cow white cheese
9. Kluyveromyces aestuarii 907 Cow yogurt
10. Kluyveromyces marxianus 908 Cow yogurt
11. Kluyveromyces marxianus 909 Cow yogurt
12. Kluyveromyces marxianus 9010 Yellow cheese
13. Candida bombicola 9011 Melted cheese
14. Kluyveromyces marxianus 9012 Melted cheese
15. Blastobotritis proliferans 9013 Cow white cheese
16. Pichia xylosa 9014 Cow white cheese
17. Candida blankii 9015 Cow milk
18. Kluyveromyces fragilis 9016 Yellow cheese
19. Kluyveromyces fragilis 9017 Yellow cheese
20. Dekkera anomala 9018 Cow milk
21. Dekkera anomala 9019 Cow milk
22. Dekkera anomala 9020 Cow milk
23. Dekkera anomala 9021 Cow milk
24. Dekkera anomala 9022 Cow milk
25. Shizosaccharomyces servazzii 9023 Cow milk

1614 Biotechnol. & Biotechnol. eq. 24/2010/1
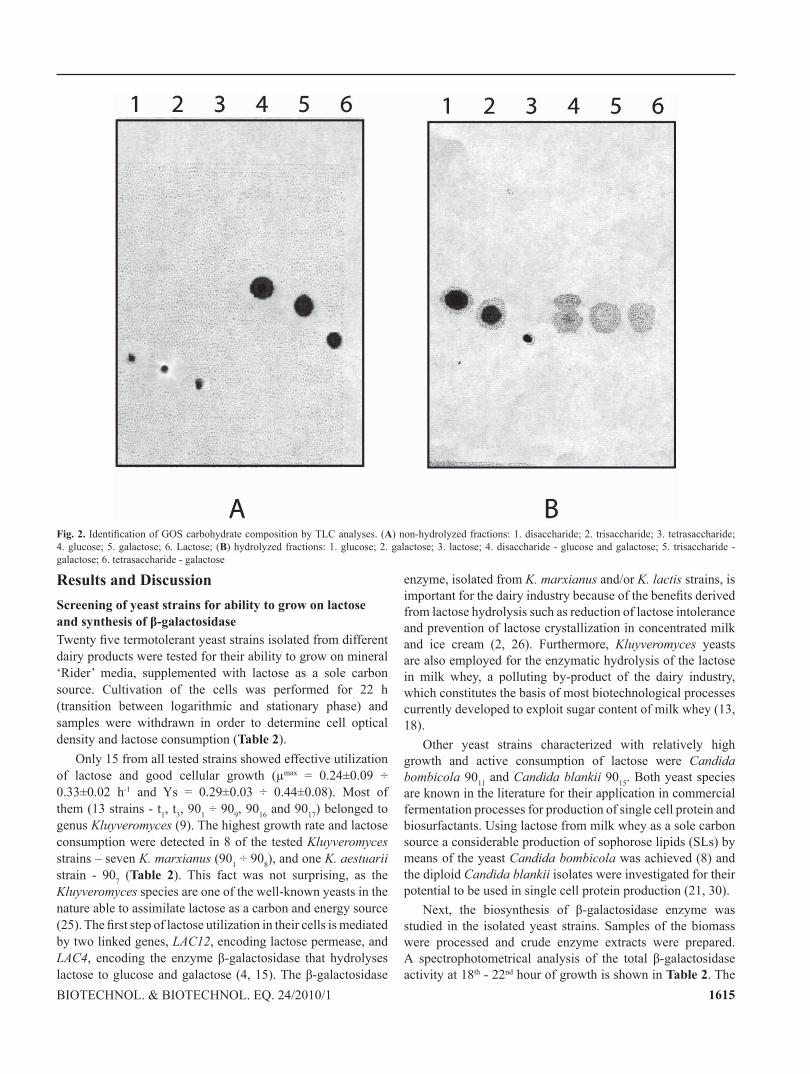
Fig. 1. TLC analyses for detection of GOS after transformation of lactose for 4 h (a) and 24 h (b). (A) yeast strains belonging to Group 1; (B) yeast strains belonging to Group 2; (C) yeast strains belonging to Group 3; (D) yeast strains belonging to Group 4
HPLC analysisQuantitative analyses of GOS were performed on Waters Millipore HPLC apparatus. Oligosaccharides were separated on Hibar LiChrosorb NH2 coulmn (250 x 4 mm, 5 µm), with elution by acetonitrile/water (4:1) at 0.8 ml min-1. Before injecting, the samples were 10 fold diluted with deionized water and centrifuged for 10 min at 13 000 rpm.
Yields of GOS were presented as a weight percentage of the appropriate carbohydrate based on total saccharides in the reaction medium. They were calculated as g l-1, and GOS conversion rate was expressed by converting their percentage according to the initial lactose (substrate) concentration.
1615Biotechnol. & Biotechnol. eq. 24/2010/1
Results and DiscussionScreening of yeast strains for ability to grow on lactose and synthesis of β-galactosidaseTwenty five termotolerant yeast strains isolated from different dairy products were tested for their ability to grow on mineral ‘Rider’ media, supplemented with lactose as a sole carbon source. Cultivation of the cells was performed for 22 h (transition between logarithmic and stationary phase) and samples were withdrawn in order to determine cell optical density and lactose consumption (Table 2).
Only 15 from all tested strains showed effective utilization of lactose and good cellular growth (µmax = 0.24±0.09 ÷ 0.33±0.02 h-1 and Ys = 0.29±0.03 ÷ 0.44±0.08). Most of them (13 strains - t1, t3, 901 ÷ 909, 9016 and 9017) belonged to genus Kluyveromyces (9). The highest growth rate and lactose consumption were detected in 8 of the tested Kluyveromyces strains – seven K. marxianus (901 ÷ 908), and one K. aestuarii strain - 907 (Table 2). This fact was not surprising, as the Kluyveromyces species are one of the well-known yeasts in the nature able to assimilate lactose as a carbon and energy source (25). The first step of lactose utilization in their cells is mediated by two linked genes, LAC12, encoding lactose permease, and LAC4, encoding the enzyme β-galactosidase that hydrolyses lactose to glucose and galactose (4, 15). The β-galactosidase
enzyme, isolated from K. marxianus and/or K. lactis strains, is important for the dairy industry because of the benefits derived from lactose hydrolysis such as reduction of lactose intolerance and prevention of lactose crystallization in concentrated milk and ice cream (2, 26). Furthermore, Kluyveromyces yeasts are also employed for the enzymatic hydrolysis of the lactose in milk whey, a polluting by-product of the dairy industry, which constitutes the basis of most biotechnological processes currently developed to exploit sugar content of milk whey (13, 18).
Other yeast strains characterized with relatively high growth and active consumption of lactose were Candida bombicola 9011 and Candida blankii 9015. Both yeast species are known in the literature for their application in commercial fermentation processes for production of single cell protein and biosurfactants. Using lactose from milk whey as a sole carbon source a considerable production of sophorose lipids (SLs) by means of the yeast Candida bombicola was achieved (8) and the diploid Candida blankii isolates were investigated for their potential to be used in single cell protein production (21, 30).
Next, the biosynthesis of β-galactosidase enzyme was studied in the isolated yeast strains. Samples of the biomass were processed and crude enzyme extracts were prepared. А spectrophotometrical analysis of the total β-galactosidase activity at 18th - 22nd hour of growth is shown in Table 2. The
Fig. 2. Identification of GOS carbohydrate composition by TLC analyses. (A) non-hydrolyzed fractions: 1. disaccharide; 2. trisaccharide; 3. tetrasaccharide; 4. glucose; 5. galactose; 6. Lactose; (B) hydrolyzed fractions: 1. glucose; 2. galactose; 3. lactose; 4. disaccharide - glucose and galactose; 5. trisaccharide - galactose; 6. tetrasaccharide - galactose
1616 Biotechnol. & Biotechnol. eq. 24/2010/1
TABLE 2Cell growth parameters on ‘Rider’ media, supplemented with 20 g L-1 lactose, and induction of β-galactosidase activity
No. Strain denotation µmax [h-1] YS qS [h-1] Residual lactose [g L-1] β-galactosidase [U mg-1]
1. t1 0.26 ± 0.09 0.29 ± 0.03 0.14 ± 0.01 12.11 ± 1.40 280.3 ± 23.42. t3 0.29 ± 0.03 0.38 ± 0.05 0.25 ± 0.03 0.35 ± 1.82 480.0 ± 11.33. 901 0.28 ± 0.03 0.36 ± 0.01 0.28 ± 0.01 0.17 ± 0.95 412.4 ± 25.84. 902 0.30 ± 0.04 0.38 ± 0.08 0.24 ± 0.02 0.07 ± 1.21 435.1 ± 10.45. 903 0.30 ± 0.06 0.38 ± 0.09 0.25 ± 0.02 0.21 ± 1.41 420.0 ± 15.86. 904 0.33 ± 0.02 0.44 ± 0.08 0.32 ± 0.04 0.70 ± 1.40 500.0 ± 22.47. 905 0.31 ± 0.01 0.40 ± 0.02 0.32 ± 0.03 0.24 ± 0.90 450.5 ± 22.48. 906 0.30 ± 0.01 0.42 ± 0.02 0.31 ± 0.03 1.87 ± 1.35 450.4 ± 18.89. 907 0.30 ± 0.08 0.40 ± 0.03 0.32 ± 0.01 1.29 ± 1.87 420.5 ± 16.210. 908 0.30 ± 0.07 0.38 ± 0.04 0.25 ± 0.02 0.22 ± 2.10 395.5 ± 13.511. 909 0.24 ± 0.09 0.31 ± 0.05 0.24 ± 0.03 0.37 ± 1.46 320.0 ± 11.412. 9010 0.13 ± 0.02 0.15 ± 0.05 0.20 ± 0.02 16.52 ± 1.55 120.2 ± 11.613. 9011 0.23 ± 0.01 0.28 ± 0.07 0.11 ± 0.02 6.51 ± 1.80 410.4 ± 10.214. 9012 0.10 ± 0.02 0.15 ± 0.04 0.19 ± 0.01 16.04 ± 2.10 210.1 ± 9.815. 9013 - - - 21.12 ± 2.34 10.0 ± 3.216. 9014 - - - 20.42 ± 2.10 10.2 ± 2.417. 9015 0.26 ± 0.02 0.33 ± 0.08 0.27 ± 0.04 0.42 ± 1.10 385.1 ± 21.518. 9016 0.26 ± 0.05 0.33 ± 0.08 0.28 ± 0.01 0.21 ± 0.80 492.0 ± 23.419. 9017 0.24 ± 0.08 0.28 ± 0.09 0.25 ± 0.02 2.69 ± 1.40 300.5 ± 18.620. 9018 0.13 ± 0.01 0.16 ± 0.03 0.10 ± 0.02 14.87 ± 1.40 212.0 ± 18.121. 9019 - - - 18.00 ± 2.00 50.2 ± 5.422. 9020 - - - 18.40 ± 1.35 62.8 ± 3.223. 9021 - - - 20.00 ± 2.25 62.4 ± 4.424. 9022 0.09 ± 0.02 0.12 ± 0.01 0.05 ± 0.01 17.70 ± 1.41 70.1 ± 4.425. 9023 - - - 19.20 ± 1.85 75.9 ± 3.6
detected values vary between 10 and 500 U mg-1 protein in different strains. The maximal enzyme activity was measured in two K. marxianus strains - t3 and 904 (480 and 500 U mg-1 protein, respectively) and in the K. fragilis 9016 strain (492 U mg-1 protein). On the other hand, strains 9010, 9012, 9013, 9014, 9018, 9019, 9020, 9021, 9022, and 9023 showed very poor growth on lactose, as well as low β-galactosidase activity. All these results are in agreement with the already known fact that Kluyveromyces yeasts are the major commercial source of β-galactosidase, as their natural habitat is the dairy environment (14). The obtained data also correspond to the earlier studies performed with K. lactis strains, which have shown that maximum β-galactosidase activity is associated with particular growth phase, namely late exponential – early stationary phase (or approximately 20th hour of cultivation) (10, 25).
Transgalactosylation activity and GOS productionAll 25 yeast strains tested were analyzed for their ability to transform lactose to the corresponding GOS. To reduce the permeability barrier and prepare whole cell biocatalysts with high activities, the harvested wet yeast biomass was treated
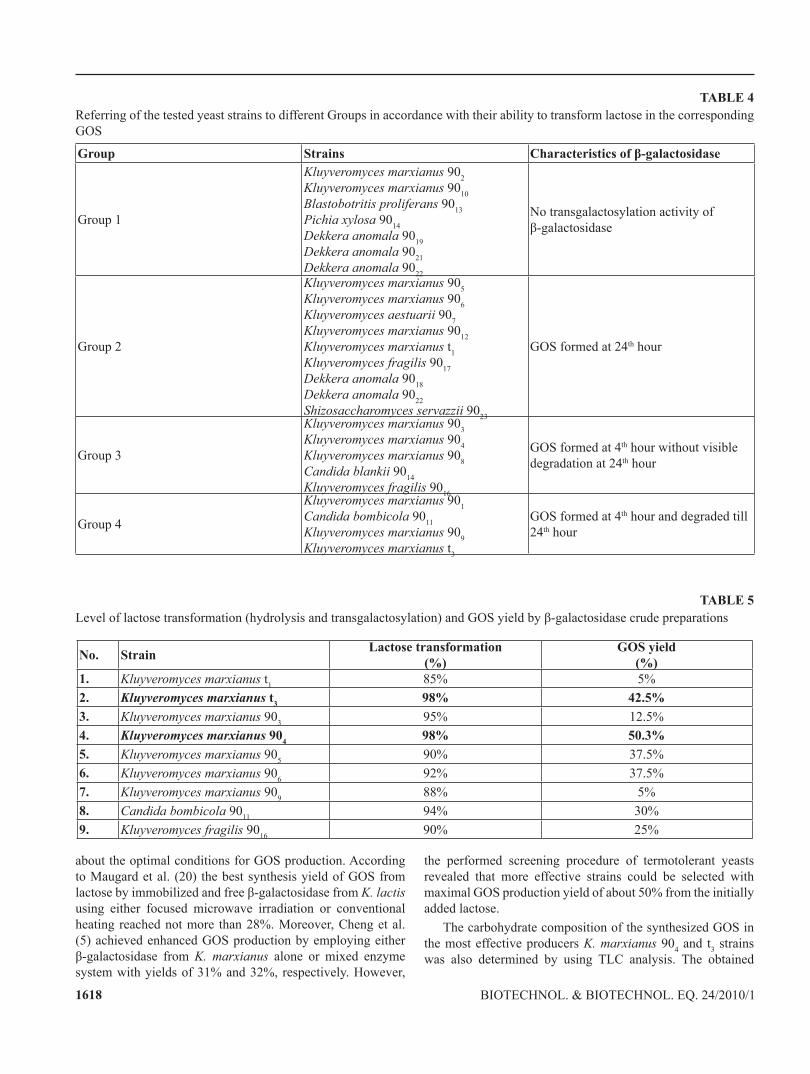
with toluene (22). Next, lactose was added in concentration 0.4 g ml-1 and the transgalactosylation reaction was followed up after 4 and 24 hours of incubation. The obtained results are presented in Table 3. It is evident that in correlation with the data presented for the cellular growth and β-galactosidase activity, the transferring activity was also observed in the yeast strains belonging to genus Kluyveromyces with positive transgalactosylation detected in strains 901, 903, 904, 905, 9016 and 9017. An interesting phenomenon presents the strain K. marxianus 9012, which although lacking significant growth on lactose, showed relatively high β-galactosidase activity (210.1±9.8 U mg-1) with transglactosylation properties. In four of the tested strains transgalactosylation activity was not detected at all (9010, 9013, 9014 and 9019). In accordance with the obtained results, the studied yeast strains could be referred in 4 corresponding groups: Group 1 - strains without transgalactosylation activity; Group 2 - strains forming GOS at 24th hour; Group 3 - strains forming GOS at 4th hour of incubation and not degrading them or degrading them in minimal quantities till 24th hour; and Group 4 - strains forming GOS at 4th hour and degrading them until 24th hour (Table 4).
1617Biotechnol. & Biotechnol. eq. 24/2010/1
TABLE 3Screening of the investigated yeast strains for presence of transgalactosylation activity. The yeast cells were incubated in media with high lactose concentration (0.4 g ml-1) and samples were withdrawn at 4th and 24th hour of incubation. Transgalactosylation efficacy of β-galacosidase enzyme was expressed in relative units in accordance with the relative quantity of product detected after performance of TLC analysis
No. Strain denotationTransgalactosylation activity of β-galactosidase (relative units)
4th hour 24th hour1. t1 - +2. t3 + -3. 901 + -4. 902 - -5. 903 + +6. 904 + +7. 905 - +8. 906 - +9. 907 - +10. 908 + +11. 909 + -12. 9010 - -13. 9011 + -14. 9012 - +15. 9013 - -16. 9014 - -17. 9015 + +18. 9016 + +19. 9017 - +20. 9018 - +21. 9019 - -22. 9020 - -23. 9021 - -24. 9022 - +25. 9023 - +
‘+’ - GOS synthesis was detected‘-‘ - no GOS synthesis was detected
Further, to visualize and characterize the types of GOS formed during the transgalactosylation process, the crude enzyme preparations were analyzed through TLC (Fig. 1A, B, C and D). The obtained chromatograms revealed that in most of the crude preparations of the tested strains a trisaccharide is produced. In addition, K. marxianus 904 and t3 strains besides the trisaccharides also formed a tetrasaccharide. This result is supported by other authors, reporting that GOS synthesized through β-galactosidase from Kluyveromyces spp., containing mainly trisaccharides. Only a small amount of tetrasaccharides, and GOS with higher degree of polymerization could be observed (3, 6). In all tested strains both galactose and glucose were still present in the mixture while lactose in most cases was almost completely hydrolyzed (about 98% lactose transformation was achieved). The best yield of tri-
and tertasacchrarides was observed in K. marxianus 904 and t3 strains.
Quantitative comparative analyses of the produced GOS were performed by applying high-performance liquid chromatography (HPLC). Calibration curves of glucose, galactose and lactose were constructed and used to convert peak areas of each component to concentration. GOS were assumed to give the same detector response as lactose and therefore, a lactose calibration curve was used to calculate their concentrations. Data are presented in Table 5. Maximum conversion rate is observed in K. marxianus 904 and t3 strains, which is 42.5% and 50.3%, respectively. In the other tested yeast strains the average conversion of lactose to GOS is with lower efficacy, varying from 5% to 37.5%. The obtained values correspond to those already reported in the literature
1618 Biotechnol. & Biotechnol. eq. 24/2010/1
TABLE 4Referring of the tested yeast strains to different Groups in accordance with their ability to transform lactose in the corresponding GOS
Group Strains Characteristics of β-galactosidase
Group 1
Kluyveromyces marxianus 902Kluyveromyces marxianus 9010Blastobotritis proliferans 9013Pichia xylosa 9014Dekkera anomala 9019Dekkera anomala 9021Dekkera anomala 9022
No transgalactosylation activity of β-galactosidase
Group 2
Kluyveromyces marxianus 905Kluyveromyces marxianus 906Kluyveromyces aestuarii 907Kluyveromyces marxianus 9012Kluyveromyces marxianus t1Kluyveromyces fragilis 9017Dekkera anomala 9018Dekkera anomala 9022Shizosaccharomyces servazzii 9023
GOS formed at 24th hour
Group 3
Kluyveromyces marxianus 903Kluyveromyces marxianus 904Kluyveromyces marxianus 908Candida blankii 9014Kluyveromyces fragilis 9016
GOS formed at 4th hour without visible degradation at 24th hour
Group 4
Kluyveromyces marxianus 901Candida bombicola 9011Kluyveromyces marxianus 909Kluyveromyces marxianus t3
GOS formed at 4th hour and degraded till 24th hour
TABLE 5Level of lactose transformation (hydrolysis and transgalactosylation) and GOS yield by β-galactosidase crude preparations
No. Strain Lactose transformation(%)
GOS yield(%)
1. Kluyveromyces marxianus t1 85% 5%2. Kluyveromyces marxianus t3 98% 42.5%3. Kluyveromyces marxianus 903 95% 12.5%4. Kluyveromyces marxianus 904 98% 50.3%5. Kluyveromyces marxianus 905 90% 37.5%6. Kluyveromyces marxianus 906 92% 37.5%7. Kluyveromyces marxianus 909 88% 5%8. Candida bombicola 9011 94% 30%9. Kluyveromyces fragilis 9016 90% 25%
about the optimal conditions for GOS production. According to Maugard et al. (20) the best synthesis yield of GOS from lactose by immobilized and free β-galactosidase from K. lactis using either focused microwave irradiation or conventional heating reached not more than 28%. Moreover, Cheng et al. (5) achieved enhanced GOS production by employing either β-galactosidase from K. marxianus alone or mixed enzyme system with yields of 31% and 32%, respectively. However,
the performed screening procedure of termotolerant yeasts revealed that more effective strains could be selected with maximal GOS production yield of about 50% from the initially added lactose.
The carbohydrate composition of the synthesized GOS in the most effective producers K. marxianus 904 and t3 strains was also determined by using TLC analysis. The obtained

1619Biotechnol. & Biotechnol. eq. 24/2010/1
samples were hydrolyzed with 1N HCl at 700C according to Coughlin and Nickerson (7) and were then analyzed on Silica gel aluminum plates (Fig. 2). The obtained results showed that both tri- and tetrasaccharides were composed mainly from galactose. Correspondingly, Chockchaisawasdsee et al. (6) revealed that depending of lactose concentration subjected to conversion by β-galactosidase from K. lactis, the composition of tri- and tetrasaccharides is mainly consisted either only from glucose or from galactose residues.
ConclusionsDisposal of lactose in whey and whey permeates is one of the significant problems with regards to economics and environmental impact faced by the dairy industry. The enzymatic hydrolysis of whey lactose to glucose and galactose by β-galactosidase constitutes the basis of most biotechnological processes that are currently developed to exploit the sugar content of the whey (2, 13, 18, 25, 26). Moreover, employment of transgalactosylation activity of β-galactosidase enzymes and production of prebiotic GOS will further enhance the value of the process.
Here, it was demonstrated that two new termotolerant strains of Kluyveromyces marxianus yeasts have been isolated with enhanced transgalagtosylation activity and high degree of lactose conversion to important tri- and tetragalacto-oligosaccharides. Thus, this work can contribute to the further development of effective wasteless technologies for production of high-content of valuable galacto-oligosaccharides by lactose assimilation and enzyme catalysis with whole cell biocatalysist from Kluyveromyces marxianus strains.
AcknowledgementsThis work was supported by a grant from the Sofia University “St. Kliment Ohridski”, Thematic Project № 148/09.
REFERENCES1. Ausubel F.M., Brent R., Kingston R.E., Moore D.D.,
Smith J.A. (1989) Current protocols in molecular biology, Vol. 2., New York, John Wiley & Sons, Inc.
2. Bonekamp F.J. and Oosteron J. (1994) Appl. Microbiol. Biotechnol., 41, 1-3.
3. Boon M.A., Janssen A.E.M., Riet K. (2000) Enz. Microb. Techn., 26, 271-281.
4. Chang Y.D. and Dickson R.C. (1988) J. Biol. Chem., 263, 16696-16703.
5. Cheng C.-C., Yu M.-C., Cheng T.-Z., Sheu D.-C., Duan K.-J., Tai W.-L. (2006) Biotechnol. Lett., 28, 793-797.
6. Chockchaisawasdee S., Vasileios I., Athanasopoulos K.N., Rastall R.A. (2004) Biotechnol. Bioeng., 89, 434-443.
7. Coughlin J.R. and Nickerson T.A. (1974) J. Dairy Sci., 58, 169-174.
8. Daverey A., Pakshirajan K., Sangeetha P. (2009) Int. J. Environ. Sci. Eng., 1, 173-175.
9. Deneva D., Zamfirova D., Kujumdzieva A. (2003) Biotechnol. & Biotechnol. Eq., 17, 85-90.
10. Dickson R.C. and Martin J.S. (1980) J. Bacteriol., 142, 777-785.
11. Flowers H.M. (1978) Meth. Enzymol., 50, 93-121.12. Hughes D.B. and Hoover D.G. (1995) J. Dairy Sci., 78,
268-276.13. Kauer G., Panesar P.S., Bera M.B., Kumar H. (2009)
Bioprocess Biosyst. Eng., 32, 63-67.14. Lee Y.-J., Kim C.S., Oh D.-K. (2004) Appl. Microbiol.
Biotechnol., 64, 787-793.15. Lodi T. and Donnini C. (2005) FEMS Yeast Res., 5, 727-
734.16. Lowry O.H., Rosenbrough N.J., Farr O.L., Randle R.J.
(1951) J. Biol. Chem., 193, 265-275.17. Mahoney R.R. (1985) In: Developments in Dairy
Chemistry 3 (P.F. Fox, Ed.), Amsterdam, Elsevier Applied Science, 69-108.
18. Martins D.B.G., de Souza C.G., Simoes D.A., Morais M.A. (2002) Curr. Microbiol., 44, 379-382.
19. Martinez-Villaluenga C., Cardelle-Cobas A., Olano A., Corzo N., Villamiel M., Jimeno M.J. (2008) J. Agric. Food Chem., 56, 557-563.
20. Maugard T., Gaunt D., Legoy M.D., Besson T. (2003) Biotechnol. Lett., 25, 623-629.
21. Meyer P.S., du Preez J.C., Wingfield B.D., Kilian S.G. (1993) J. Biotechnol., 29, 267-275.
22. Murakami K., Nagura H., Yoshino M. (1980) Anal. Biochem., 105, 407-413.
23. Nedeva T, Petrova V, Hristozova T, Kujumdzieva A. (2002) Z. Naturforsch, 57c, 960-961.
24. Ohrlein R. (1999) Top. Curr. Chem., 200, 227-254.25. Ornelas A.P., Silveira W.B., Sampaio F.C., Passos
F.M.L. (2007) J. Appl. MIcrobiol., 104, 1008-1013.26. Panesar P.S., Panesar R., Singh R.S., Kennedy J.F., and
Kumar H. (2006) J. Chem. Toxicol. Biotechnol., 81, 530-543.
27. Pirt S.H. (1975) In: Principles of Microbe and Cell Cultivation, Blackwell Scientific Publications, Oxford.
28. Sako T., Matsumoto K., Tanaka R. (1999) Int. Dairy J., 9, 69-80.Sharp R., Fishbain S., Macfarlane G.T. (2001) J. Med. Microbiol., 50, 152-160.
29. Tzvetkova B., Hristozova Ts., Angeov A., Paskaleva D. (2004) World J. Microbiol. Biotechnol., 20, 219-223.
30. Vandebplas Y. (2002) Br. J. Nutr., 87, 293-296.