thermoregulation in a large bird, the emu (dromaius novaehollandiae)
TRANSCRIPT

J Comp Physiol B (1994) 164:464-472 Journal of Comparative Biochemical, Systemic.
and Environ'-
Physiology B " - ' Physiology
�9 Sioringer-Verlag 1994
Thermoregulation in a large bird, the e m . (Dromaius novaehollandiae) S.K. Maloney*, T.J. Dawson
School of Biological Science, University of New South Wales, PO Box 1, Kensington, 2033, Australia
Accepted: 10 August 1994
Abstract. The emu is a large, flightless bird native to Aus- tralia. Its habitats range f rom the high snow coun t ry to the arid interior of the continent. Our experiments show that the emu mainta ins a cons tant b o d y tempera ture within the ambient tempera ture range - 5 to 45 ~ C. The males regulate their b o d y tempera ture abou t 0.5 ~ C low- er than the females. Wi th falling ambient tempera ture the emu regulates its b o d y temperature initially by reducing conduc tance and then by increasing heat product ion . At - 5 ~ C the cost of mainta in ing thermal balance is 2.6 times basal metabol ic rate. By sitting down and reducing heat loss f rom the legs the cost of h o m e o t h e r m y at - 5 ~ C is reduced to 1.5 times basal metabol ic rate. At high am- bient temperatures the emu utilises cu taneous evapora- tive water loss in addi t ion to panting. At 45 ~ C evapora- t ion is equal to 160% of heat product ion . Pant ing ac- counts for 70% of total evapora t ion at 45 ~ C. The cost of utilising cutaneous evapora t ion for the o ther 30% ap- pears to be an increase in dry conductance .
Key words: Cutaneous evapora t ion - Pant ing - Ratite - Thermoregu la t ion - Emu, Dromaius novaehollandiae
Introduction
The emu is a large flightless bird that inhabits mos t of the Austral ian continent. Its habitats range f rom the high snow count ry of the Grea t Dividing Range to the hot, arid interior (Grice et al. 1985). The emu's large size limits the extent to which it can exploit micro-cl imates and so its ability to exist in such thermally diverse habitats must largely reflect successful physiological thermoregulat ion. Some of this success would undoub ted ly be at t r ibutable to the degree of thermal inertia that large b o d y size con- fers. However , long- term main tenance of a temperature difference between an animal and its envi ronment re- quires regulat ion of heat p roduc t ion and loss.
The emu belongs to the suborder Ratiti which consists of several species of large flightless birds (Cracraft 1974). There have been reports on the response to high temper- ature in the ostrich Struthio camelus (Schmidt-Nielsen et al. 1969; Crawford and Schmidt-Nielsen 1967) and the response to the heat generated dur ing exercise in the rhea Rhea americana (Taylor et al. 1971). However, an overall picture of ratite physiological responses to T a changes remains unknown.
Present address: * Brain Function Research Unit, University of the Witwatersrand, 7 York Road, Parktown, 2193, South Africa Abbreviations: At, Effective radiating surface area; BMR, basal metabolic rate; Cdry, dry conductance; CEWL, cutaneous evapora- tive water loss; EHL, evaporative heat loss; EWL, evaporative wa- ter loss; FECO2, fractional concentration of CO2 in excurrent air; FEH2o, water content of chamber excurrent air; FEO2, fractional concentration of 02 in chamber excurrent air; FICO2, fractional concentration of CO2 in incurrent air; FIO2, fractional concentra- tion of 02 in chamber incurrent air; MHP, metabolic heat produc- tion; MR, metabolic rate; REWL, respiratory evaporative water loss; RH, relative humidity; RQ, respiratory quotient (12CO2/1202); SA, surface area; SEM, standard error of the mean; SNK, Student- Newman-Keuls multiple range test; STPD, standard temperature and pressure, dry; T a, ambient temperature(s); Tb, body tempera- ture(s); Te, surface temperature(s); ~ flow rate of air into the cham- ber; 17CO2, carbon dioxide production; 1202, oxygen consumption; VPHzo, vapour pressure of water
Materials and methods
Experimental animals. The ten emus used (five female and five male) were raised in captivity at the University of New South Wales Cow- an Field Station where the birds lived in a wooded enclosure (200 x 100 m). Sex was determined by cloacal examination. Experi- ments were carried out at the University of New South Wales in winter (July-September) and summer (November-February). The emus were familiarized with the experimental procedure for at least 2 weeks prior to data collection. After this time they would stand or sit quietly in the experimental chamber. None of the birds were laying or incubating during the experimental period. At the Univer- sity the emus were housed in pens (4.3 • 1.2 • 2.5 m) with food ('kangaroo' pellets, fruit, lettuce and bread) and water ad libitum. Food and water were witheld for 24 h prior to experimentation. The birds maintained body mass during the approximately 6 weeks they were used for the measurements (winter females 39.7 + 0.5 kg, males 37.0_+ 1.2 kg; summer females 45.4___2.8 kg, males 40.7_+3.4 kg).

S.K Maloney, T.J. Dawson: Emu thermoregulation 465
Metabolic measurements. Experiments were carried out during the quiet phase of the emu's diurnal cycle (between 1800 and 0330 hours), except for one set of measurements at Ta = 25 ~ C performed during the day. Animals were placed in a 1 . 8 x l . 0 x 0 . 6 m metabolism chamber which had a mesh floor above a bath of veg- etable oil to cover any excreta. The inside walls of the chamber were painted flat black to reduce radiation reflection in the chamber (Porter 1969). The chamber was placed in an environmental room in which Ta was controlled to within +_0.5 ~ C of a set point. Measure- ments were made at five temperatures in summer (5, 15, 25, 35 and 45 ~ C) and at eight in winter (-5, 5, 15, 20, 25, 30, 35 and 45 ~ C). T~ was measured (+_0.1 ~ C) with a Jenco electronic thermometer, the probe being situated in the excurrent port of the chamber. This thermometer was calibrated against a mercury in glass thermome- ter certified by the National Association of Testing Authorities, Australia.
Flow rate of dry air into the chamber was adjusted so that V P m o in the chamber remained below that which would give a relative humidity of 20% at the T b of emus (from 120 l-rain -1 at - 5 ~ C to 250 l-min -1 at 45 ~ C). Flow rate was measured upstream of the chamber with a Hastings Mass Flowmeter (model HFM-201). Approximately 125 ml-min -~ of chamber excurrent air was drawn through a capacitance type humidity sensor (CHK-Engineering, dgt-rh/t; _+0.1% RH). This sensor was calibrated at regular inter- vals with saturated solutions of LiC13 and NaC1 (Winston and Bates 1960).
The sample air stream was then dried and divided in two. One stream passed through an Anarad AR-50 CO 2 analyzer calibrated daily with ambient air and a gas of known higher CO2 concentra- tion (Commonwealth Industrial Gases, Sydney). The other stream was scrubbed of CO 2 with Ascarite, redried and passed through an Applied Electrochemistry Model S3A-II oxygen analyzer.
Outputs from the four sensors (mass flow, relative humidity, CO2 and 02) were logged every 5 s on a computer via a 16-bit analog/digital converter (Sable Systems, USA). Approximately 120 readings were averaged for each recorded value. Maximum resolu- tion for this combination of sensors and A/D converter was 0.002% for 02 and 0.01% for CO2. The system was calibrated several times each season by the iron-burn method of Young et al. (1984). Cali- bration factors varied between 0.97 and 1.02.
Experimental procedure. Before each experiment an emu was weighed to the nearest 0.1 kg on a platform balance (Wedderburn Scales, Sydney). At temperatures below 20~ it was desired to obtain data from animals in both sitting and standing positions. Animals could be forced to stand by placing a large plastic box in the chamber (this left insufficient room for sitting). Animals could be coaxed to sit but were not always willing to remain sitting for long periods. We allowed 1-2 h of equilibration at all T a before data collection was initiated. Data collection periods lasted from 1.5-4 h. In most cases total exposure to each T, was more than 3 h. At the end of an experiment plumage surface temperatures of seven repre- sentative areas (Fig. 1) were measured with an Everest infra-red thermometer. This thermometer was calibrated by reading the tem- perature of a plastic block painted flat black. Temperature of the block was measured with a Jenco electronic thermometer and al- tered by placing it in a temperature-controlled water bath. Emmis- sivity of the block was assumed to be 0.99. Distance of the ther- mometer from the emu was adjusted so that measurements from large areas (back, shoulder, belly, hip) were measurements of ap- proximately 200 cm 2, while the thermometer was held close to the neck, leg, and toe for measurements from an area of approximately 1 cm 2.
T b was then measured 10 cm into the cloaca (which would place the sensor in the rectum of the emu) with a calibrated Jenco elec- tronic thermometer. T b measurement was completed within 5 min of removing the bird from the chamber. Animals were sometimes measured at two T~ during one night. At least 48 h elapsed between experimental sessions for any one animal.
~ Neck
Back
Leg
T o e
Fig. 1. Diagram illustrating the areas of the emu where the radiation thermometer was aimed for surface temperature measurements
Measurement of cutaneous evaporative water loss. Five emus that were exposed to T~ of 25 and 45 ~ C in summer were fitted with two Perspex capsules containing Drierite. The capsules consisted of a round sleeve (5.68 cm diameter) and tight-fitting caps which con- tained the Drierite below wire mesh. Two sleeves were glued (Super- glue) to areas where the feathers had been trimmed (one to the back and one to the belly). After the bird had been exposed to the rele- vant T~ for at least 30 min a pre-weighed cap was fitted to each sleeve. At Ta = 25 ~ C the caps were left on the birds for 60 rain and at Ta=45 ~ C for 30 min. After removal, the caps were immediately weighed. CEWL was calculated from the weight gain of the cap.
Although this type of method is the most commonly used means of measuring CEWL from animals the results should be viewed within the limitations of the method. The use of a Drierite capsule creates an artificially dry environment (VPmo close to zero) and this will have a bearing on the results. Although chamber VPH2 o was maintained low (<0.5 kPa at T a up to 30 ~ C; <1.3 kPa at 45 ~ C) it was always higher than in the capsules. This factor, in addition to the removal of any resistance to water vapour move- ment normally offered by the feathers, will almost certainly have led to overestimates of CEWL.
Data analysis. IkO 2 was calculated as: 1202 = IkSTPD.(FIO2-FEO2)/(1- FEO2), where IkST•D = flOW rate of dry air into the chamber (STPD), and F I O 2 and F E Q were the fractional concentrations o f O 2 in air entering and leaving the chamber, respectively (Hill 1972). IkCO 2 was calculated as: 12CO2= ~Tt, D-(FECO2-FICO2), where FICO2 and FECO2 were the fractional concentrations of CO 2 in air enter- ing and leaving the chamber, respectively. EWL was calculated as: EWL=FEH20.(Ik+IkCO2-1)O2)/(1-FEH20), where FEH20 was the water content of the excurrent air [calculated from humidity using a predictive equation based on vapour pressure data (Weast and Astle 1983] and lkwas the flow rate at ambient temperature and pressure.
M R and EWL at each T a were defined as the lowest continuous 5 rain of lkO 2 and EWL, respectively, in any collection period. Due to condensation of water in the chamber and excurrent lines at T a < 0 ~ C, EWL was not calculated for - 5 ~ C. For calculation of conductance EWL at - 5 ~ C was assumed to be the same as at 5 ~ C. MHP and EHL were calculated assuming 20.08 kJ.1 02 -1 (based on RQ measurements, see Results) and 2.42 kJ.g H20 L Dry conduc- tance 0N -~ C -J) was calculated as:
MHP-EHL (Jdry - - Tb_~
Surface area specific estimates of dry conductance (W.m-2.~ 1) were obtained by dividing the above estimates by total plumage surface area.

466 S.K Maloney, T.J. Dawson: Emu thermoregulation
Surface area. The plumage surface area of each area in Fig. l was measured in four emus. The neck, legs, and toes were assumed to be cylinders with a surface area equal to the average circumference times the length. All other areas (back, shoulder, hip, belly) were traced with tissue paper and the area calculated by trigonometry. Total SA of the four emus was used to calculate a Meeh formula:
SA (cm2)=7.5 (+0.16) Mass (g)0.667 (1)
This is a smaller mass coefficient than the value of 8.11 obtained by Walsberg and King (1978) as a general formula for external plumage surface of birds. This may reflect the relatively simple geometry of the emu's body and its lack of wings. However, the emu's SA in- cludes the legs - which other SA analyses have not - because they form a large proportion (15%) of the standing emu's SA. Equation 1 was used to estimate the SA of each emu used in this study.
The average contribution of each surface in Fig. 1 to total SA was used to estimate the area of each region for each emu. These estimates and the T~ of each region were used to calculate radiative heat exchange for each region at each T~, assuming chamber wall temperature was equal to T~ (this was confirmed several times) using the formula:
~=~1 �9 ~z'd'(T~ -- T,)" S A 4 4 (2)
where: (b = radiant heat exchange (W) as = animal surface infrared emmissivity (assumed 0.98) e 2 = chamber wall infrared emmissivity (assumed 0.98) (~ = Stefan-Boltzmann constant (5.67 x 10 -8 W.m4.K 4) T~ = relevant surface temperature (Kelvin) T~ = chamber wall temperature (Kelvin) SA = relevant surface area
Total radiant heat exchange for an animal at any T a was the sum of the radiant heat exchange from all regions.
Investigators interested in the radiative exchange of heat from animals have reduced total SA to an effective radiating surface area (At) since some areas of the body will be exchanging radiation with other surfaces on the animal and not the chamber walls. Taking the shape of the emu into consideration (i.e. estimating the proportion of SA shielded from radiant exchange with chamber walls by other parts of the body, such as the inside of legs and the belly) and comparing it with published values of As for other animals (Folkow and Mercer 1986), a value of 0.9.SA was used as A r for the emu. Radiative heat exchange calculated from Eq. 2 was multiplied by 0.9 to obtain a final estimate of radiative heat exchange for each ani- mal.
Statistical analysis. Data were initially analysed using a two-way repeated measures ANOVA for season and the five common tem- peratures for each sex. Season did not prove significant for any
variable. Since pooling of seasons for a single analysis is statistically invalid (Hurlbert 1984) the majority of the further analysis was restricted to the winter results. Winter results were analysed using a Two-Way Repeated Measures ANOVA for sex and temperature. A Student-Newman-Keuls (SNK) multiple range test was applied when significant differences were indicated by the ANOVA (using Statistica/Mac software). If sex proved significant the sexes are pre- sented separately, otherwise data for both sexes are grouped. Twelve out of the sixty individual T~/T~ sets were not recorded due to problems with animals. Rather than reduce the data set to those individuals on which a full data set was obtained, thus allowing a paired analysis, sex and temperature differences were tested for with a factorial ANOVA and individual means compared with Fishers PLSD test. All graphs and tables show mean _+ SEM. Means are considered significantly different if P < 0.05.
Results
Body temperature
At the comple t ion of metabo l i sm experiments (3-5 h ex- posure) T b had not changed from the rmoneu t ra l levels at any T~, except at T~=45 ~ C, where there was a slight increase in Tb (Fig. 2). Males tended to have lower Tb than females, though not at higher T~.
Metabolism
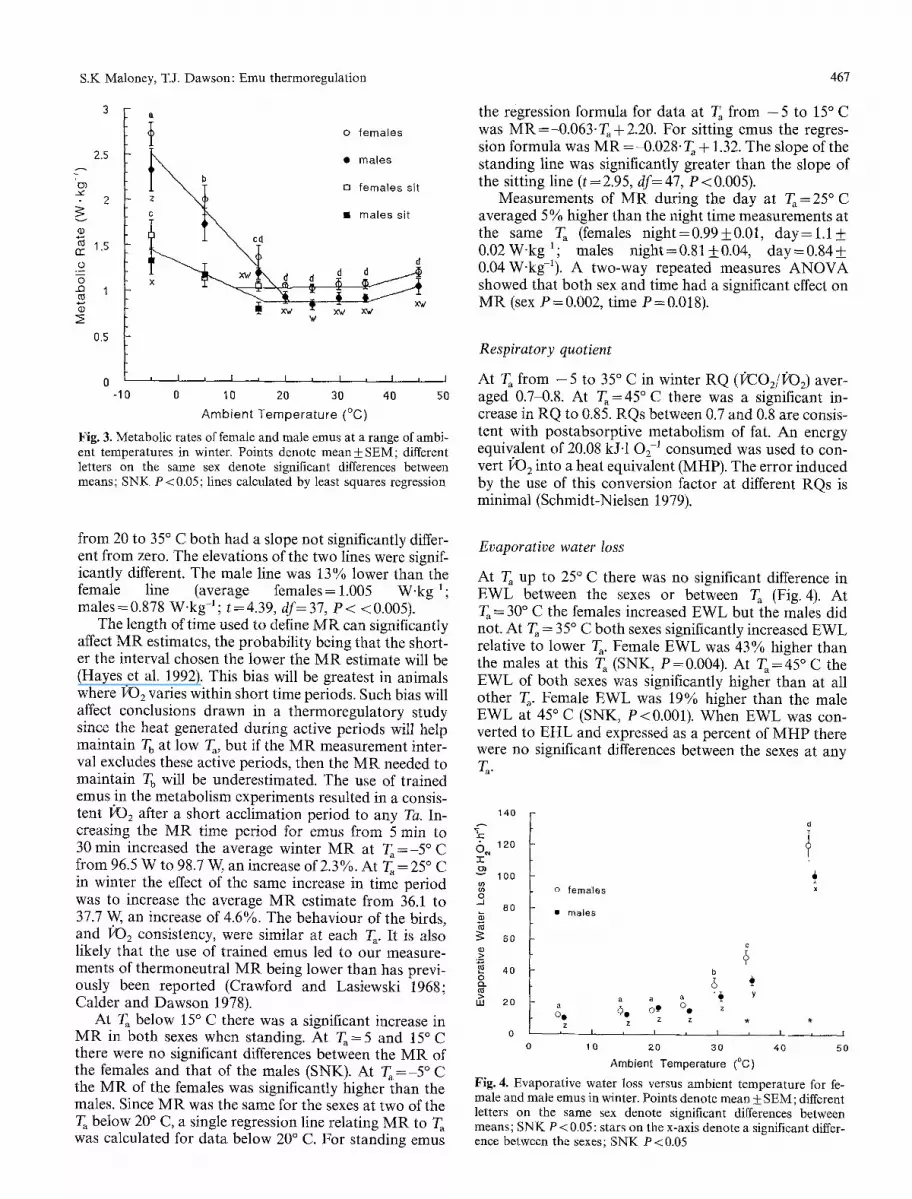
At T~s below 2 0 ~ data were ob ta ined from all emus while they were s tand ing in the chamber. At - 5 ~ C da ta were also ob ta ined from all emus sitt ing in the chamber , bu t at 5 and 15 ~ C not all the emus would remain sitt ing long enough to ob ta in stable measurements of MR. At 5 ~ sit t ing da ta were ob ta ined from two females and four males and at 15 ~ C from two females and three males. At 25 ~ C there was no difference between the M R of six emus for which bo th s tanding and sitt ing da ta were ob ta ined (s tanding 0.946 W ' k g -1, sitting 0.915 W.kg-1; n = 6 , paired t=1 .63 , P = 0 . 1 6 ) a nd so da ta at T~ above 20 ~ C were collected from either s i tuat ion.
Cons t an t M R for the two sexes at T, between 20 and 35 ~ C, bu t at different levels (Fig. 3), is further evidence that B M R differs between the sexes (Maloney and Daw- son 1993). The regression lines for female and male da ta
39 39
U o 38.5
4~
C~ 3 8 E
I---
o 37.5 nm
37
Winter
ab
Z z
b
Y
, i , l ~ I I I
0 0 I 0 20 30 40
Ambient Temperature (~
38.5
38
37.5
I 37
50
Slimmer b
{
Y
z 0 f e m a l e s
z �9 males
i I , I L I ~ t , I
I0 20 30 40 50
AmNenL TemperaLure (~
Fig. 2. Body temperatures of female and male emus taken at the completion of metabolism experiments in winter and summer. Points denote mean_+SEM; dif- ferent letters on the same sex denote sig- nificant differences between means; SNK P < 0.05: stars on the x-axis denote a sig- nificant difference between the sexes; SNK P < 0.05
S.K Maloney, T.J. Dawson: Emu thermoregulation 467
w-
2
C~ 1.5
O
-6 -Q 1
3
2,5
0.5
0 -10
x " Z ~ ~ \ - - - .4 .
o f e m a l e s
�9 m a l e s
r~ f e m a l e s s i t
�9 m a l e s s i t
d d d
I t I I I I I ~ I A I
0 10 20 30 40 50
Ambient Temperature (~
Fig. 3. Metabolic rates of female and male emus at a range of ambi- ent temperatures in winter. Points denote mean +_ SEM; different letters on the same sex denote significant differences between means; SNK P < 0.05; lines calculated by least squares regression
from 20 to 35 ~ C both had a slope not significantly differ- ent from zero. The elevations of the two lines were signif- icantly different. The male line was 13% lower than the female line (average females = 1.005 W . k g l ; males =0.878 W.kgq ; t=4.39, d f= 37, P < <0.005).
The length of time used to define M R can significantly affect M R estimates, the probabil i ty being that the short- er the interval chosen the lower the M R estimate will be (Hayes et al. 1992). This bias will be greatest in animals where ]20 2 varies within short time periods. Such bias will affect conclusions drawn in a thermoregulatory study since the heat generated during active periods will help maintain T b at low T,, but if the M R measurement inter- val excludes these active periods, then the M R needed to maintain T b will be underestimated. The use of trained emus in the metabolism experiments resulted in a consis- tent 1)O 2 after a short acclimation period to any Ta. In- creasing the M R time period for emus from 5 rain to 30 min increased the average winter M R at T~=-5~ C from 96.5 W to 98.7 W, an increase of 2.3%. At T ,=25 ~ C in winter the effect of the same increase in time period was to increase the average M R estimate from 36.1 to 37.7 W, an increase of 4.6%. The behaviour of the birds, and 1)O 2 consistency, were similar at each T~. It is also likely that the use of trained emus led to our measure- ments of thermoneutral M R being lower than has previ- ously been reported (Crawford and Lasiewski 1968; Calder and Dawson 1978).
At T, below 15 ~ C there was a significant increase in M R in both sexes when standing. At T~=5 and 15~ there were no significant differences between the M R of the females and that of the males (SNK). At T, = - 5 ~ the M R of the females was significantly higher than the males. Since M R was the same for the sexes at two of the T~ below 20 ~ C, a single regression line relating M R to T, was calculated for data below 20 ~ C. For standing emus
the regression formula for data at T~ from - 5 to 15 ~ C was MR=-0.063"Ta+2.20. For sitting emus the regres- sion formula was M R = ~).028. T~ + 1.32. The slope of the standing line was significantly greater than the slope of the sitting line (t = 2.95, df= 47, P < 0.005).
Measurements of M R during the day at T ,=25~ C averaged 5% higher than the night time measurements at the same T~ (females night=0.99+_0.01, d a y = l . l + 0.02 W - k g 1; males night = 0.81 + 0.04, day = 0.84-t- 0.04 W-kg-1). A two-way repeated measures A N O V A showed that both sex and time had a significant effect on M R (sex P=0.002, time P=0.018).
Respiratory quotient
At T~ from - 5 to 35 ~ C in winter RQ (fYCO2/l~2) aver- aged 0.7~0.8. At T,=45~ C there was a significant in- crease in RQ to 0.85. RQs between 0.7 and 0.8 are consis- tent with postabsorptive metabolism of fat. An energy equivalent of 20.08 kJ.1 02 J consumed was used to con- vert 1)O 2 into a heat equivalent (MHP). The error induced by the use of this conversion factor at different RQs is minimal (Schmidt-Nielsen 1979).
Evaporative water loss
At T, up to 25 ~ C there was no significant difference in EWL between the sexes or between T a (Fig. 4). At Ta = 30 ~ C the females increased EWL but the males did not. At T~ = 35 ~ C both sexes significantly increased EWL relative to lower T,. Female EWL was 43% higher than the males at this T a (SNK, P = 0.004). At T a = 45 ~ C the EWL of both sexes was significantly higher than at all other T,. Female EWL was 19% higher than the male EWL at 45 ~ C (SNK, P<0.001). When EWL was con- verted to E H L and expressed as a percent of M H P there were no significant differences between the sexes at any ra .
1 4 0
6 ~ 1 2 0
I
100 o .-1
8 0
60
4 0 x
I.U 2 0
o f e m a l e s
�9 m a l e s
x
b
a a a Y
a 0 0 ~ e o~ z o o z z . . z z
I I I I I
10 2 0 3 0 4 0 5 0
A m b i e n t T e m p e r a t u r e ( ~
Fig. 4. Evaporative water loss versus ambient temperature for fe- male and male emus in winter. Points denote mean __ SEM; different letters on the same sex denote significant differences between means; SNK P<0.05: stars on the x-axis denote a significant differ- ence between the sexes; SNK P<0.05

468 S.K Maloney, T.J, Dawson: Emu thermoregulation
Cutaneous evaporative water loss
Average CEWL measured using drierite capsules at T ,= 25 ~ C was 14.6 g H20-m-2-h -~, There was no differ- ence between the rates measured from the back and the belly�9 At T~ = 45 ~ C data could not be obtained from two emus. Average CEWL for the other three emus was 21.1 g H20.m-2"h -1. Rates of CEWL estimated from the differ- ence between EWL and calculated rates of respiratory EWL [calculated from ventilatory data in Maloney and Dawson (1994)] were 7.2 g HzO'm-2"h -~ at T~=25 ~ C and 28�9 g H20.m~-h ~ at T~=45 ~ C in summer. These esti- mates were made using the formula for skin surface area of birds obtained by Walsberg and King (1978)�9 The value obtained using the capsules at Ta=45~ C is within the 95% confidence intervals of the summer data. The value obtained at 25 ~ C is above the 95% confidence interval of the summer data (upper 95% interval = 9.1). The low hu- midity in the capsules compared to the chamber cannot alone account for this difference which may also be due to removal of the feathers�9
Conductance
Females and males did not differ in Cd~y at any T~, Cdry was minimal at 20 ~ C. Above 20 ~ C Ca~y increased to a maximum at 45 ~ C (Fig. 5A,B). Figure 5A also shows the
S0 [ A Neck f r B Shoulder [ '~ 3 0
~ . O T~ = T~
? ? , . . . . . . . '~176 . ,~
e ~ 40 /
'~ 30 d
09 20
-10 , t -10 0 10 20 30 40 50-10
[ C Back r
E Belly d ~ rx [ F Leg e e
T r i i i J E _ i i r i i i 10 20 30 40 50 - I0 O 10 20 30 40 50
Ambient Temperature (~
Fig. 6. Surface temperatures of various areas of emus in relation to ambient temperature. Points denote mean 4-SEM; different letters denote significant differences between means; Fisher's PLSD P<0 .05
Cd~y of sitting emus at T~ = - 5 ~ C, calculated using total SA (open circle) and exposed SA (open square, see Dis- cussion). A repeated measures ANOVA and SNK was used to compare these three measures of Cdry a t - - 5 ~ C with the values at 20 ~ C. All four means were significantly different from each other, in contrast to the analysis of the full range of standing data which indicated that at - 5 and 20 ~ C the values were not different.
7
cu
E
v
q9 o E t ~
O = 2
O 1
A 0
a a
o =
a b
b o
v
�9 o
o
7 2 E O o
B5
ab T
T a , a -r
T i I l l a
6 1'o 2o ~ 4b T e m p e r a t u r e (~
Fig. 5. A Emu dry conductance in winter expressed as W-m-2"~ -t. Open square and circle show the conductance of sitting emus calcu- lated using exposed and total surface area respectively. B Emu dry conductance in winter expressed as W.~ 1. Open square shows sitting emus. Points denote mean_+SEM; different letters denote significant differences between means; SNK P<0 .05
Surface temperature
All Te measurements were from standing emus. At any one Ta there were no differences between male and female T~ of any region. T~ of six of the seven areas identified in Fig. 1 are shown in Fig. 6A-F. Temperatures of all sur- faces decreased at lower T~. The temperature difference between emu surface and T~ was lower for the well insu- lated areas of the body (shoulder, back, hip) than other areas not as well insulated.
Heat balance
Heat loss from an animal's body occurs via four path- ways: radiation, evaporation, conduction, and convec- tion. We calculated the contribution of conduction/con- vection as the difference between total heat loss (which is M H P since Tb was constant) and EH L plus net radiative heat loss. The contribution of each pathway to total heat loss in winter and summer is shown in Fig. 7A,B. There were no significant seasonal differences in any of the parameters. The heat lost by conduction/convection was similar to that lost by radiation at each T~. The magni- tude of conduction/convection and radiation decreased as T~ rose to 35 ~ C, and CEWL and REWL increased across this temperature range. EH L (CEWL and REWL) at 45~ rose substantially in order to dispose of metabolic heat in addition to the heat being gained by conduction/convection and radiation from the warmer surroundings.

S.K Maloney, T.J. Dawson: Emu thermoregulation
200
150
O "J 100 I# "1- "5 50
0
n -50
- 1 0 0
Winter
200 Summer [Z] Cond + Cony ~ CEWL
150 . . . . . . . . . . . . .
O " 100 q~ "I-
"6 50
0
0_ -50
-100 ....... _; ; ;5 is A Ambient Temperature (~
Fig. 7. Partitioning of heat loss by evaporative and non-evaporative means in winter and summer
Discussion
Body temperature
The T b of emus is low in comparison with other bird species. This may be a feature of the ratite group (Withers 1983; Maloney and Dawson 1993). There was little change in the T b of the emus following 3-5 h exposure to T a from - 5 to 45 ~ C (Fig. 2).
Conductance and heat balance
A change in conductance with a change in temperature regime (e.g. with changes in season) is a principal means
469
of animal acclimation to environmental conditions (Briick 1986). In winter a low conductance is advanta- geous, while in summer increased conductance would seem advantageous. The emu did not show such a pattern. How- ever, the emu is active in summer on days when environ- mental temperatures are higher than Tb (Dawson et al. 1984). In these conditions a lower conductance (higher insulation) will reduce the environmental heat load.
The conductance of the emu is higher than predicted by the allometric relationships of Herreid and Kessel (1967), Aschoff (1981), and Drent and Stonehouse (1971; their Fig. 8). However, these studies used a generalised Meeh formula for skin surface area, whereas Walsberg and King (1978) have shown that the commonly quoted Meeh formula overestimates SA of the plumage by some 23%. Bech (1980) pointed out that studies using this Meeh factor would have underestimated true plumage surface area specific conductance. Posture changes may also affect exposed surface area (for example, if an emu sits down exposed SA is reduced by 15% by the covering of the legs; exposed SA may be further reduced by curling of the neck onto the shoulder). The conductance esti- mates of the sitting emus, calculated using exposed SA (Fig. 5A, open square), were significantly higher than esti- mates made using total SA (open circle). In species where postural adjustments contribute to temperature regula- tion in the cold, estimates of conductance made using total SA will also be underestimates. Bech (1980) has argued also that as body size increases insulation may reach a maximum and so the relationships cited above cannot be applied to large birds [the largest bird in the analysis of Herreid and Kessel (1967) was 2.7 kg, in the study by Aschoff (1981) it was 2.4 kg, and in the data set of Drent and Stonehouse (1971) it was 5 kg]. These fac- tors in addition to the implication that conductance is equivalent over all surfaces of the body (whereas this is definitely not the case) restrict the utility of such a mea- sure. Conductance expressed in whole animal terms (as W .~ C ~, Fig. 5B) is a useful estimate of the amount of resistance to heat flow between an animal and the envi- ronment.
Below the lower critical temperature of the emu, heat production was increased to balance the greater heat loss associated with larger gradients (Fig. 3) and conductance
40
v 30 ID3 e-
2o 0 x
LU 10
ID I
0 >
if5 -10 CE
-20 O
i--
A - 3 0
8
b
n=10
d
-5 5 15 25 35 45
Ambient Temperature (~
40 0") c
Z X
cu 30
I
~ 20
r r
"5
0 ~-B
b
b
j iiiiiiiiiiiii I (t 6
Illiii
-5 5 15 25
Ambient Temperature (~
45
Fig. 8. A Total radiative heat ex- change between emu and chamber walls in watts. B Leg radiative heat exchange as a percent of total ra- diative heat exchange (n for each temperature given in Fig. 8A; points denote mean 4- SEM; different let- ters denote significant differences between means; Fishers PLSD P < 0.05)

470 S.K Maloney, T.J. Dawson: Emu thermoregulation
was minimal (Fig. 5B), which resulted in low T e (Fig, 6). With decreasing T, the T e of the legs fell considerably (Fig. 6F). Because the legs are poorly insulated, such a low T~ suggests that the interior of the legs was not main- tained at Tb. Hyrtl (1863) has shown that the arteries and veins at the top of the emu's legs are arranged in a fashion that allows counter-current heat exchange [simple rete, sensu Midtggtrd (1981)]. If the T e of the legs was equal to T b at T~ = - 5 ~ C, radiant heat loss from the legs would be 29 W instead of the observed 9 W. Even with the rete 25-30% of total radiant heat loss from the emu occurred from the legs at low T~ (Fig. 8B). An indication of the importance of the legs as sources of heat loss can be gained by comparing the heat production of sitting and standing emus at low T~ (Fig. 3). MR of emus sitting and standing at thermoneutral T~ suggested that the metabol- ic cost of standing per se was not very large. Much of the reduction in MR in sitting emus would be due to reduced heat loss from the legs.
At T~ up to 30 ~ C radiation, conduction and convec- tion accounted for most of the heat exchange (Fig. 7). At T~=25~ total radiative heat loss was reduced com- pared to the lower T~, but the proportion of that ex- change that occurred from the legs increased (Fig. 8B). This indicates that the legs may act as 'heat windows'. Poorly insulated areas of animals can be important areas for heat dissipation, increasing dry heat loss at moderate T a. Examples of such heat windows are elephants' ears (Phillips and Heath 1992), bare skin areas under the wings of ostriches (Louw et al. 1969), and the legs and feet of birds (Steen and Steen 1965; Midtggtrd 1980; Arad et al. 1989). At T, = 35 ~ C, leg Te in emus was close to T~. In some individuals it was below T~, an indication that wa- ter was evaporating from the legs. CEWL from the legs may be important to emus exposed to high T~ and a radiant heat load. During periods of high solar radiation (i.e. around midday) the legs will be shaded by the body and with CEWL could act as efficient heat dissipators. This could also explain why emns are observed crouched, rather than sitting flat, in creek beds on very hot days in summer in the arid zone [T~>42~ Dawson et al. (1984)]. Crouching would expose the evaporating sur- faces of the belly and legs to any ambient convection and increase evaporation.
Evaporative water loss
Loss of water when it is not essential for temperature regulation is disadvantageous to animals inhabiting arid areas. The break even point for water loss/metabolic wa- ter production is 0.67mg H20.ml O2-1consumed (Schmidt-Nielsen and Schmidt-Nielsen 1952). Some of the standing emus in this study were gaining more metabolic water than they were losing via EWL. The average ratios of milligrams water lost per ml 0 2 corl- sumed were 0.67 at --5 ~ C, 0.92 at 5 ~ C, 1.6 at 15 ~ C, and 2.6 at 25 ~ C. None of the sitting emus were making a gain at low temperatures, although the ratios were low (1.06 at - 5 ~ C, 1.23 at 5 ~ C, and 1.99 at 15 ~ C). These calcula- tions do not include water lost in urine and feces. The
ability to minimize water loss will be important during times of limited water intake; for example, during the male's long incubation period which involves continuous nest attendance for 56 days. A factor contributing to the emus, low ventilatory water loss was the ability to exhale air cooler than T b (Maloney and Dawson 1994).
At T~ above 30 ~ C the emus relied on increased EWL to maintain thermal balance (Figs. 4, 7). At the higher T a used in this study (35 and 45 ~ C) the EWL of females was significantly higher than that of the males (Fig. 4). This was probably related to the higher metabolism of the females (Fig. 3). The slightly higher Tb of the females gave them a larger temperature gradient for dry heat loss at Ta = 35 ~ C, but this only allowed for an increase in dry heat loss of 2 W out of the 12 W higher heat production. The other 10 W were dissipated by evaporation. At Ta=45~ C the 25% higher metabolism of the females again placed a larger burden on their EWL mechanisms relative to those of the males, resulting in the females having a 20% higher rate of EWL at this T~.
At T~ slightly lower than Tb, a high conductance will allow dissipation of heat by non-evaporative means and thus a saving of water. The emu's conductance did in- crease at T a = 35 ~ C relative to lower Ta (Fig. 5B). Howev- er, when T~ exceeds Tb it is advantageous for the animal to decrease Cary. This means that less heat flows into the body, and the EWL needed to maintain T b is lower than if Cdry is high (Dawson and Schmidt-Nielsen 1966). This pattern of increasing Cdry as T~ approaches Tb from be- low, but then decreasing Cdry when T~ exceeds Tb has been observed in many species, including pyrrhuloxias and Arizona cardinals (Hinds and Calder 1973), starlings (Dmi'el and Tel-Tzur 1985), chukar and sand partridges (Frumkin et al. 1986) and jack rabbits (Dawson and Schmidt-Nielsen 1966). The emu did not conform to this pattern. Its Cdry at T~=45~ was greater than at any other Ta. At T~=45~ C there was 2.4 (females) and 2.7 (males) times as much heat flowing into the body, result- ing in EWL rates 24% (females) and 28% (males) higher than if Cary were the same as at T a = 25 ~ C. This begs the question why was the Cdry SO high at T a=45 ~ C? This may be the inevitable result of the contribution of CEWL to evaporative cooling. At T~ = 45 ~ C approximately 30% of total EWL was CEWL in the emu. In other bird spe- cies CEWL is dependent on high levels of skin blood flow (Marder and Raber 1989). The maintenance of high levels of skin blood flow will inevitably lead to high thermal conductance, further increasing heat flow from the envi- ronment into the body.
There would appear to be a trade off involved in using CEWL. The bird could vasoconstrict the periphery, ab- sorb less heat, and so need a lower EWL, but the majority of that EWL would have to be achieved by respiratory evaporation (Hinds and Calder 1973; Dmi'el and Tel- Tzur 1985). Alternatively it could rely more on CEWL but consequently need a higher EWL to dissipate the extra environmental heat load (Arad et al. 1987; Withers and Williams 1990). Given that most bird species are threatened by pH imbalance when panting (Calder and Schmidt-Nielsen 1968; Marder and Arad 1989), the trade offmay be one of water economy versus pH balance. This

S.K Maloney, T.J. Dawson: Emu thermoregulation 471
trade off is affected by acclimatization. Pigeons acclimat- ed to high T~ but with cons tant access to water do not pant at high T, but dissipate all the metabol ic and the external heat load by C E W L (Marder and Gavrieli-Levin 1987; Marder et al. 1989). Non-acc l imated pigeons pant and develop b lood alkalosis (Calder and Schmidt- Nielsen 1968; Bernstein and Samiengo 1981).
The emu presents an interesting case: it can pant with- out developing a severe pH imbalance (Jones et al. 1983), so why should it suffer the disadvantages associated with C E W L ? Perhaps diurnal foraging would be compro- mised by the need to pant cont inuously on hot days. Alternatively, the C E W L mechanisms m a y have evolved in response to the high heat loads experienced during exercise in this cursorial species. We have observed emus mainta in ing 25-30 km.h -1 for several kilometres. Evapo- rat ion into the moving airstream from the body surface would be an efficient means of dissipating the heat of exercise, while pant ing would be restricted by the need to adequate ly ventilate the lungs at high MR.
Conclusion
We have shown that the emu is capable of mainta ining thermal balance across a wide range of 7]. However, the l abora to ry condit ions dur ing this s tudy would not al- ways hold for an emu in the wild. In particular, the radia- tive envi ronment in the field will differ greatly f rom the control led radiative envi ronment of the metabol ism chamber. The greatest influence will be direct solar radia- tion, while objects absorbing solar radiat ion will tend to be hot ter than Ta and vegetat ion will tend to be cooler than T~ due to t ranspirat ion, in contras t to the homoge- nous radiant envi ronment of the chamber. In addition, the convective envi ronment will influence rates of heat exchange. While these factors make any field thermoreg- u la tory s tudy more complex, this s tudy will serve as a baseline for future field studies.
Acknowledgements. We thank Adam McLean, Geoff Vaughan and Ray Williams for help with animal capture and handling, and Prof. Mark Chappell for his constant technical advice and the loan of much hardware. SKM was supported by an Australian Post-Grad- uate Research Award. This research was funded in part by an Aus- tralian Research Council grant to TJD. Animals were held under a license from the New South Wales National Parks and Wildlife Service. This research was conducted with the approval of the Uni- versity of New South Wales Animal Care and Ethics Committee.
References
Arad Z, Gavrieli-Levin I, Eylath U, Marder J (1987) Effect of dehy- dration on cutaneous water evaporation in heat-exposed pi- geons (Columba Iivia). Physiot Zool 60:623-630
Arad Z, Midtgfird U, Bernstein MH (1989) Thermoregulation in turkey vultures: vascular anatomy, arteriovenous heat ex- change, and behaviour. Condor 91:505-514
Aschoff J (1981) Thermal conductance in mammals and birds: its dependence on body size and circadian phase. Comp Biochem Physiol 69A:611619
Bech C (1980) Body temperature, metabolic rate, and insulation in winter and summer acclimatized mute swans (Cygnus olor). J Comp Physiol 136:61-66
Bernstein MH, Samiengo FC (1981) Ventilation and acid-base status during thermal panting in pigeons (Columba livia). Physi- ol Zoot 54:303 315
Brfick K (1986) Basic mechanisms in thermal long-term and short- term adaptation. J Therm Biol 11:73-77
Calder WA, Schmidt-Nielsen K (1968) Panting and blood carbon dioxide in birds. Am J Physiol 215:477-482
Calder WA, Dawson TJ (1978) Resting metabolic rates of ratite birds: the kiwis and the emu. Comp Biochem Physiol 60A :479- 481
Cracraft J (1974) Phylogeny and evolution of the ratite birds. Ibis 116:494-521
Crawford EC, Schmidt-Nielsen K (1967) Temperature regulation and evaporative cooling in the ostrich. Am J Physiol 212:347- 353
Crawford EC, Lasiewski RC (1968) Oxygen consumption and res- piratory evaporation of the emu and rhea. Condor 70:333-339
Dawson TJ, Schmidt-Nielsen K (1966) Effect of thermal conduc- tance on water economy in the antelope jack rabbit, Lepus al- leni. J Cell Physiol 67:463-472
Dawson TJ, Read D, Russell EM, Herd RM (1984) Seasonal varia- tion in daily activity patterns, water relations and diet of emus. Emu 84:93-102
Dmi'el R, Tel-Tzur D (1985) Heat balance of two starling species (Sturnus vulgaris and Onychognathus tristrami) from temperate and desert habitats. J Comp Physiol B 155:395-402
Drent RH, Stonehouse B (1971) Thermoregulatory responses of the Peruvian penguin, Spheniscus humboldti. Comp Biochem Physi- ol 40A:689-710
Folkow LP, Mercer JB (1986) Partition of heat loss in resting and exercising winter- and summer-insulated reindeer. Am J Physiol 251 :R32-R40
Frumkin R, Pinshow B, Weinstein Y (1986) Metabolic heat produc- tion and evaporative heat loss in desert phasianids: chukar and sand partridge. Physiol Zool 59:592-605
Grice D, Caughley G, Short J (1985) Density and distribution of emus. Aust Wildl Res 12:69-73
Hayes JP, Speakman JR, Racey PA (1992) Sampling bias in respirometry. Physiol Zool 65:604-619
Herreid CF, Kessel B (1967) Thermal conductance in birds and mammals. Comp Biochem Physiol 21:405-414
Hill RW (1972) Determination of oxygen consumption by use of the paramagnetic oxygen analyzer. J Appl Physiol 33:261-263
Hinds DS, Calder WA (1973) Temperature regulation of the pyrrhuloxia and the Arizona cardinal. Physiol Zool 46:55-71
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:187-211
Hyrtl J (1863) Neue Wundernetze und Geflechte bei V6geln und S/iugetieren. Denkschr Mathem Naturw Classe 22:113-152
Jones JH, Grubb B, Schmidt-Nielsen K (1983) Panting in the emu causes arterial hypoxemia. Respir Physiol 54:189-195
Louw GN, Belonje PC, Coetzee HJ (1969) Renal function, respira- tion, heart rate, and thermoregulation in the ostrich (Struthio camelus). Sci Pap Namib Desert Res Stn 42:43-54
Maloney SK, Dawson TJ (1993) Sexual dimorphism in basal metabolism and body temperature of a large bird, the emu. Condor 95:1034-1037
Maloney SK, Dawson TJ (1994) Ventilatory accommodation of oxygen demand and respiratory water loss in a large bird, the emu (Dromaius novaehollandiae), and a re-examination of venti- latory allometry for birds. J Comp Physiol B (in Press)
Marder J, Gavrieli-Levin I (1987) The heat-acclimated pigeon: an ideal physiological model for a desert bird. J Appl Physiol 62:952-958
Marder J, Arad Z (1989) Panting and acid-base regulation in heat stressed birds. Comp Biochem Physiol 94A:395-400

472 S.K Maloney, T.J. Dawson: Emu thermoregulation
Marder J, Raber P (1989) Beta-adrenergic control of trans-cuta- neous evaporative cooling mechanisms in birds. J Comp Physiol B 159:97-103
Marder J, Arieli Y, Ben-Ashcr J (1989) Defense strategies against environmental heat stress in birds. Isr J Zool 36:61-75
Midtggtrd U (1980) Heat loss from the feet of mallards Anas platyrhynchos and arterio-venous heat exchange in the rete tibiotarsale. Ibis 122:354-359
Midtgfird U (1981) The fete tibiotarsale and arteriovenous associa- tion in the hind limb of birds: a comparative morphological study on counter-current heat exchange systems. Acta Zool (Stockholm) 62:67-87
Phillips PK, Heath JE (1992) Heat exchange by the pinna of the African elephant (Loxodonta africana). Comp Biochem Physiol 101A:693-699
Porter WP (1969) Thermal radiation in metabolic chambers. Sci- ence 166:115-117
Schmidt-Nielsen K (1979) Animal Physiology: adaptation and envi- ronment, 2nd edn. Cambridge University Press, Cambridge, UK
Schmidt-Nielsen K, Schmidt-Nielsen B (1952) Water metabolism of desert mammals. Physiol Rev 32:135-166
Schmidt-Nielsen K, Kanwisher J, Lasiewski RC, Cohn JE, Bretz WL (1969) Temperature regulation and respiration in the os- trich. Condor 71:341 352
Steen I, Steen JB (1965) The importance of the legs in the ther- moregulation of wading birds. Acta Physiol Scand 63:285-291
Taylor CR, Dmi'el R, Fedak M, Schmidt-Nielsen K (1971) Energet- ic cost of running and heat balance in a large bird, the rhea. Am J Physiol 221 : 596-601
Walsberg GA, King JR (1978) The relationship between external surface area of birds to skin surface area and body mass. J Exp Biol 76:185-189
Weast RC, Astle MJ (1983) CRC handbook of chemistry and physics. CRC Press, Boca Raton, Florida
Winston PW, Bates DH (1960) Saturated solutions for the control of humidity in biological research. Ecology 41:232-237
Withers PC (1983) Energy, water and solute balance of the ostrich, Struthio camelus. Physiol Zool 56:568-579
Withers PC, Williams JB (1990) Metabolic and respiratory physiol- ogy of an arid-adapted Australian bird, the spinifex pigeon. Condor 92:961-969
Young BA, Fenton TW, McLean JA (1984) Calibration methods in respiratory calorimetry. J Appl Physiol 56:1120-1125