thermodynamics and structure of a salmon cold active goose-type lysozyme
TRANSCRIPT

Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part B
j ourna l homepage: www.e lsev ie r.com/ locate /cbpb
Thermodynamics and structure of a salmon cold active goose-type lysozyme
Peter Kyomuhendo a,b,1, Bjørnar Myrnes a,1, Bjørn-Olav Brandsdal b,c, Arne O. Smalås b,Inge W. Nilsen c,1, Ronny Helland b,⁎a Nofima Marine, P.O.Box 6122, N-9291 Tromsø, Norwayb The Norwegian Structural Biology Centre, Department of Chemistry, University of Tromsø, 9037 Tromsø, Norwayc The Centre for Theoretical and Computational Chemistry, University of Tromsø, 9037 Tromsø, Norway
Abbreviations: SalG, Atlantic salmon goose-type lyslysozyme; GEWL, Goose egg-white lysozyme; OEL, OstrAtlantic cod g-type lysozyme; DSC, Differential scannin⁎ Corresponding author. The Norwegian Structural B
Chemistry, University of Tromsø, 9037 Tromsø, Norway.77644765.
E-mail address: [email protected] (R. Helland).1 Fax: +47 77629100.
1096-4959/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.cbpb.2010.04.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 1 February 2010Received in revised form 6 April 2010Accepted 6 April 2010Available online 14 April 2010
Keywords:SalmonGoose-typeLysozymeDifferential scanning calorimetryProteinRefoldingThermal toleranceX-ray crystallographyInnate immunity
Atlantic salmon goose-type lysozyme (SalG) was previously shown to display features of cold-adaptation aswell as renaturation following heat treatment. In this study differential scanning calorimetry (DSC) wascarried out to investigate unfolding and potential refolding, while X-ray crystallography was used to studystructural factors contributing to the temperature-related characteristics. The recombinant SalG has amelting temperature (Tm) of 36.8 °C under thermal denaturation conditions and regains activity afterreturning to permissive (low) temperature. Furthermore, refolding is dramatically reduced in solutions withhigh SalG concentrations, coupled with significant protein precipitation. The structural features of SalGclosely resemble those of other g-type lysozymes. However, the N-terminal region of SalG is less anchored tothe rest of the molecule due to the absence of disulphide bonds, thus, contributing significantly to the low Tmof SalG. The absence of disulphide bonds and the distribution of salt bridges may at the same time easerefolding leading to renaturation.
ozyme; HEWL, Hen egg-whiteich egg-white lysozyme; CLYS,g calorimetry.iology Centre, Department ofTel.:+47 77646474; fax: +47
l rights reserved.
© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Lysozymes (EC 3.2.1.17) hydrolyse the β-1,4-glycosidic bondbetween N-acetylmuramic acid (NAM) and N-acetyl-D-glucosamine(NAG) residues in peptidoglycans surrounding and protecting thebacterial cells. There are three types of lysozyme prevalent in theanimal kingdom; chicken c-type lysozyme (ex. hen egg-whitelysozyme, HEWL), the goose g-type lysozyme (ex. goose egg-whitelysozyme, GEWL) and the invertebrate i-type lysozyme (ex. Tapesjaponica lysozyme, TJL) (Jollès and Jollès, 1984; Jollès, 1996; Ito et al.,1999). In addition, lysozyme is found in phages (ex. bacteriophage T4lysozyme, T4L) (Weaver and Matthews, 1987), bacteria, and plants.Until a few years ago the g-type lysozymes were believed to belong inbirds only, but an increasing number of publications have since 2001documented the presence of g-type lysozymes in many other groupsof vertebrates (Hikima et al., 2001; Irwin and Gong, 2003) and even
invertebrates (Nilsen et al., 2003; Zou et al., 2005) where it is the soletype of lysozyme in some animals (Nilsen et al., 2003).
The majority of g-type lysozymes have been characterised only atthe gene level. Mammals and zebrafish carry two and someurochordates carry three g-type lysozyme genes arising from earlylineage-specific duplication (Irwin and Gong, 2003; Nilsen et al.,2003). There is a strong indication of alternative splicing in the signalpeptide coding region and an alternative transcription start in salmonand cod g-type lysozyme genes, respectively. This creates two variantsof each of these proteins, where the only difference is that one variantlacks a signal peptide (Kyomuhendo et al., 2007; Larsen et al., 2009).The thermodynamic parameters for a g-type lysozyme have so faronly been determined for ostrich egg-white lysozyme (OEL) revealinga Tm of 60 °C (Kawamura et al., 2006; Kawamura et al., 2008). Theoptimal temperature of lytic activity for some avian (Thammasiriraket al., 2001; Pooart et al., 2005) and marine (Hikima et al., 2001;Kyomuhendo et al., 2007) g-type lysozymes was reported at about30–40 °C and 22–30 °C respectively.
Although the lysozyme types differ in their molecular weights,amino acid compositions and enzymatic properties, their threedimensional (3D) folds display structural similarities and suggest acommon evolutionary origin (Grütter et al., 1983; Weaver et al.,1984). The crystal structures for all three classes of animal lysozymeand that of phage lysozyme have been solved (Weaver andMatthews,1987; Maenaka et al., 1995; Weaver et al., 1995; Goto et al., 2007).

255P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
Before the recent study on Atlantic cod g-type lysozyme (CLYS)(Helland et al., 2009), structural studies of g-type lysozymes had beenlimited to avian species (Isaacs et al., 1985; Weaver et al., 1995). Inaddition to non-covalent forces, the structures of some native proteinsare stabilised covalently by intramolecular disulphide bonds occur-ring between the sulphurs of two cysteine side chains. The stabilisingforce of disulphide bonds is mainly attributable to the loss ofconformational entropy of the unfolded polypeptide chain by cross-linking (Pace et al., 1988;Wetzel et al., 1988; Cooper et al., 1992; Betz,1993; Siddiqui and Cavicchioli, 2005; Kawamura et al., 2008; Matsuoet al., 2009). The g-type lysozymes show greater variation in cysteinecontent than c- and i-type subclasses. In fish g-type lysozymes, thisranges from zero in flounder and grouper (Hikima et al., 2001; Yinet al., 2003), through one is salmon and carp (Savan et al., 2003;Kyomuhendo et al., 2007), to two (non-bonded) in zebrafish. In otherwords, the fish g-type lysozymes studied so far do not possessdisulphide bonds. Avian g-type lysozymes have four cysteine residuesinvolved in two disulphide bonds. Orthologous mammalian g-typelysozyme genes code either for four or seven cysteines. The threeadditional cysteines in some orthologs have potential to form onemore disulphide bond. G-type lysozymes from invertebrate specieshave six to thirteen cysteines (Nilsen et al., 2003; Zou et al., 2005;Zhao et al., 2007) but there is as yet no report of howmany disulphidebonds are actually formed in their 3D structures.
Cold-adapted enzymes, often referred to as psychrophilic, havebeen studied for many years in order to understand what differ-entiates them from their mesophilic and thermophilic counterparts.These enzymes are often catalytically more efficient at moderate tolow temperatures, more unstable at higher temperatures and have alowered temperature optimum (Feller et al., 1996). No single factorcan explain cold-adaptation, but some common structural featureswhich in general distinguish them from their mesophilic counterpartshave been comprehensively reviewed (Smalås et al., 2000; Feller andGerday, 2003; Georlette et al., 2004; Siddiqui and Cavicchioli, 2006;Marx et al., 2007).
In a preceding study, SalG showed both cold-activity andreversible heat inactivation (Kyomuhendo et al., 2007). Here,differential scanning calorimetry (DSC) was used to determine thethermodynamic parameters of SalG andwhether it actually refolds. Itscrystal structure was solved in an attempt to explain its cold-activityand ease of renaturation. This is the first report of a calorimetric studyof a g-type lysozyme from an aquatic organism and the structure isonly the second of this lysozyme subclass from a non-avian species.Moreover, correlating biochemical properties to structure of g-typelysozymes of aquatic origin has previously not received any attention.
2. Materials and methods
2.1. Differential scanning calorimetry (DSC) of recombinant SalG andactivity measurements
Overexpression and purification to homogeneity were carried outas previously described (Kyomuhendo et al., 2007). DSC measure-ments were performed in an ultra-sensitive, fast scanning, model CSC6300 Nano-DSC III differential scanning calorimeter (CalorimetrySciences Corporation (CSC), Linden, UT). Samples were dialysed toequilibrium against the reference buffer (50 mM NaCl and 50 mMNaAc pH 5.2 buffer of ionic strength 0.1 mol kg−1). Protein concen-trations were determined by measuring the absorbance at 280 nmusing a UV-1700 spectrophotometer (Shimadzu). All buffers andprotein solutions were degassed for 15 min under a vacuum aspirator.Purified recombinant SalG at a concentration of 0.75 and 1.6 mg/mLwas heated in duplicate in the temperature range 0–60 °C usingheating rates of 1 and 2 °C/min under 3 atmospheres pressure. Thedenatured protein was further analysed by immediately cooling to4 °C, followed by an additional round of heating to 60 °C, after which
enzyme activity was measured. Data were analysed with CpCalc 2.1(Applied Thermodynamics).
Lysozyme activity was measured by recording spectrophotomet-rically at 450 nm the decrease in absorbance during lysis of asuspension of lyophilised Micrococcus lysodeikticus cells (Sigma) at aconcentration of 0.2 mg/mL in 50 mM sodium acetate buffer (pH 5.2)as previously described (Kyomuhendo et al., 2007). The activity wasdetermined from the first 2 min, and one unit of activity is defined asthe amount of enzyme that catalyses a reduction in absorbance of0.001 min−1.
2.2. Crystallisation, data collection, structure determinationand refinement
Lyophilised SalG was dissolved in 150 mM NaCl and 50 mM HEPESpH 7.25 to a final concentration of 8 mg/mL. Initial conditions forcrystallisation were screened using commercially available screen kitsfrom BioGenova and Hampton Research. Small salmon lysozymecrystalswere obtained using the hangingdropvapourdiffusionmethod,where 1 µL drops of SalGweremixedwith an equal volume of reservoirsolution. Crystals suitable for X-ray diffraction studies appeared fromreservoir solutions containing 43–45% ammonium sulphate, 0.01 Mcobalt chloride and 0.1 M MES buffer with pH 6.25–6.5 at 22 °C.Diffraction data collection was carried out at beamline BL-14.1 at BESSY(Berlin, Germany) to a resolution of 1.75 Å. The data were processedusing MOSFLM and SCALA and TRUNCATE of the CCP4 suite (Leslie,1992; Collaborative Computational Project, 1994; Evans, 1997). Thestructure of goose lysozyme (PDB entry 153L (Weaver et al., 1995))wasused as a starting model for molecular replacement using MOLREP(Vagin, 1997). Automatic main chain tracing and side-chain dockingwas carried outwith the ARP/wARP software (Perrakis et al., 1999). Themodel was further improved bymanual rebuilding using O (Jones et al.,1991) based on sigmaA-weighted 2mFo–DFc and mFo–DFc electrondensity maps and refinement using REFMAC5 in the CCP4 suite(Collaborative Computational Project, 1994; Murshudov et al., 1997).The structure has been deposited with the Protein Data Bank, entry3MGW.
2.3. Homology modelling
A homology model of ostrich lysozyme (OEL) was generated inSwiss Model (http://swissmodel.expasy.org/ (Arnold et al., 2006))using GEWL as template. Two models were generated where residues85–86 were in the orientations of GEWL and SalG, respectively. Bothmodels were subjected to 1000 steps of energy minimization usingthe MacroModel (version 9.5) option of the Schrödinger software(Schrödinger, LLC, New York, NY, 2008, http://www.schrodinger.com).
2.4. Structure analysis and comparison
Programs for structural comparison and analysis were taken fromthe CCP4 suite. Ion pair interactions were identified with WHAT IFprogram suite (Vriend, 1990) using the criterion that the distancebetween two oppositely charged atoms is less than or equal to 4 Å.Hydrogen bonds were calculated using HBPLUS (McDonald andThornton, 1994) with a 3.4 Å distance criteria and default donor–acceptor angles of 90°. Structural alignments were performed usingSTRAP (Gille and Frommel, 2001) and displayed with ESPript (Gouetet al., 1999). Images of molecules were prepared with PyMOL(DeLano, 2002).
3. Results
Shift in temperature affects the stability of molecular associations,structures and consequently functions of molecules. To furtherinvestigate previous observed effects of temperature on SalG, we

Table 1Differential scanning calorimetry study of SalG.
Calorimetric van't Hoffa
SalG in cell(mg/mL)
Scan rate(°C/min)
Enthalpy (ΔHcal)(kcal mol−1)
Transitiontemperature(Tm) (°C)
Enthalpy (ΔHvH)(kcal mol−1)
0.75 1 73.4 36.8 73.40.75 2 81.7 38.6 73.51.6 1 74.0 36.7 76.71.6 2 72.1 38.4 74.3
a Two-state unfolding model.
256 P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
employed thermodynamic and kinetic analyses of the protein insolution together with 3D structural studies on crystallised protein.
3.1. Temperature, unfolding and activity
DSCmeasures the change in energy in a sample as the temperatureis raised or lowered, and thus, can determine absolute thermody-namic data for thermally-induced transitions. The melting tempera-ture Tm (unfolding transition peak) is defined as the temperature atwhich 50% of the proteinmolecules are unfolded. SalG at two differentconcentrations (0.75 and 1.6 mg/mL) was analysed by DSC andshowed mean Tm of 36.8 °C and 38.5 °C at two different scan rates (1and 2 °C/min, respectively), as shown in Fig. 1. No exothermic peakwas observed in the cooling scans. The temperature of the samplecells was therefore rapidly adjusted to 4 °C and held overnight afterthe first heating scan to allow the protein to refold to the native state.The subsequent DSC analysis revealed denaturation peaks on heatingthe protein samples a second timewith a Tm of 33 °C for protein at lowconcentration. Thermodynamic parameters for the initial scans arelisted in Table 1. Denaturation enthalpy (ΔH) of the protein at Tm afterinitial denaturation and overnight incubation at 4 °C was substantiallyreduced by 40 kcal/mol, or 60%, relative to the initial denaturationthermogram. In order to examine a two-state transition thecalorimetric (ΔHcal) and the van't Hoff (ΔHvH) enthalpy changes arecompared. Two-state behaviour normally implies that the transientintermediate states are not populated at the transition temperature(Zhou et al., 1999). Table 1 shows the ΔHcal and ΔHvH obtained for thedifferent scan rates and protein concentrations. The ΔHcal/ΔHvH isclose to 1 which indicates a two-state transition with no population ofunfolding intermediates. The transition temperature of thermaldenaturation of SalG was dependent on scan rates since doublingthe scan rate resulted in a 2 °C increase in transition temperature.
Based on previous measurements showing SalG activity afterprolonged heating (Kyomuhendo et al., 2007), the proportion ofprotein molecules refolded after DSC studies was unexpectedly low.We therefore reproduced previous heat pre-incubation studies toinclude the high protein concentration used in the DSC analyses.Before enzyme activity assays were performed on the cooled proteinsolutions, additional aliquots were removed from the high concen-tration heated protein to allow cooling in diluted form. The results aresummarised in Table 2 and show that heating (60 and 90 °C) andcooling at high SalG concentrations have significant effects on enzymeactivity, while the protein at low concentration regains most or all of
Fig. 1. DSC thermograms of SalG. Primary scan 1 ismeasured at a scan rate of 1 °C min−1
for 1.6 mg/mL protein; primary scan 2 is measured at 2 °C min−1 for 1.6 mg/mLprotein; rescan 1 is the sample from primary scan 1, but after overnight incubation at4 °C.
its activity after the heating. Approximately 30% activity is recoveredwhen the concentrated protein is cooled from 60 °C in a 30-folddiluted concentration, and only 8% activity remains in diluted samplesafter cooling from 90 °C. Thus a rapid and irreversible inactivationtakes place during heating at high enzyme concentration. Inaccordance, significant aggregation or precipitation of proteins wasobserved when highly concentrated protein solutions were subjectedto heating and cooling. Protein concentration measurements revealedthat the loss of soluble SalG after heating (60 and 90 °C) wasproportional to the loss of enzyme activity in the samples.
3.2. Crystallisation, diffraction data collection and refinement
Salmon lysozyme crystals grew up to 0.2×0.2×0.05 mm withinthree weeks in an ammonium sulphate and MES buffer. The crystalsdiffracted to 1.75 Å and belonged to space group H3 (space group 146)where A-axis=B-axis=103 Å and C-axis=43 Å. The structure wassolved by molecular replacement using goose lysozyme as the startingmodel, and refined to a crystallographic R-factor of 17.63% and Rfree of21.52%. The data collection and refinement statistics are listed in Table 3.The structure is composed of 181 residues where residues Asp7 toTyr185 constitute the mature salmon lysozyme aligning withcorresponding residues (7–185) in GEWL. The first two His residues inthe deposited structure (PDB entry 3MGW) are an artefact from theexpression system. The SalG polypeptide is generally well defined inelectron density (Fig. 2), with the exception of the loop comprisingresidues 22 to 24, which is poorly defined in electron density.
3.3. Overall structure
The secondary structural elements shown in Fig. 3A were assignedto SalG on the basis of the DSSP program (Kabsch and Sander, 1983).The framework of the molecule closely resembles the well character-ised g-type lysozyme fold as in a two-lobed structure comprisedmainly of α-helices, an irregular β-sheet and a long substrate bindinggroove or active site cleft. The structure consists predominantly ofsix α-helices α1–α6 (residues 30–46, 49–59, 63–74, 110–130, 136–149, and 169–182) with three small strands of irregular antiparallelβ-sheet structure β1–β3 (residues 84–85, 89–90 and 96–97). Short
Table 2Effect of temperature on SalG activity at high and low protein concentrations.
SalG Heating Cooling Relative activity(mg/mL) 12 h on ice (%)
1.60 – – 1001.60 DSCa, 20→60 °C + 331.60 60 °C, 15 min + 3
+b 301.60 90 °C, 15 min + 3
+b 80.05 – – 1000.05 90 °C, 15 min + 91
a Scan rate 2 °C/min.b 30 fold dilution of heated SalG sample before cooling.

Table 3Data collection and refinement statistics. Values in parentheses are for the highestresolution shell (1.84–1.75 Å).
Data collectionNo. of crystals 1Wavelength (Å) 0.91841Resolution limit (Å) 1.75Unit cell parametersa (Å) 103.15b (Å) 103.15c (Å) 43.67
Space group ‘H3’ (number 146)Total no. of reflections 69820 (10097)Total no. of unique reflections 19410 (2840)Redundancy 3.6 (3.6)Completeness (%) 99.64 (99.9)I/σ(I) 4.5 (1.5)Rsym (%) 8.8 (49.0)Overall B factor from Wilson plot 24.557
RefinementTotal no. of reflections 19410R-factor (%) 17.63Rfree (%) 21.52Average B factor of all atoms (Å2) 24.96No. of amino acid residues 181No. of protein atoms 1444No. of solvent molecules 150Ramachandran plot, core (%) 90.7Ramachandran plot, allowed (%) 8.6Rmsd. bond lengths, refined atoms (Å) 0.014Rmsd. bond angles, refined atoms (°) 1.298
257P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
segments of residues from 8–10, 157–159 and 165–167 are identifiedas a 310 helices.
The structure of SalG was compared to GEWL (PDB entry 153L)CLYS (PDB entry 3GXK) and a homology model of OEL as shown inFig. 3. The sequence identity between SalG and GEWL, CLYS and theOEL is 59, 60 and 63%, respectively. Residues 7–185 of SalG super-imposes on the protein structures of goose and cod lysozyme withaverage xyz displacement (xyzd) values for main chain atoms of 0.76(GEWL) and 0.78–1.07 Å (CLYS molecules D and A, respectively). Theregions with the largest xyzd values include the N-terminal region,residues 22–24, the loop region from residues 82 to 88 ( Fig. 3B and C).Larger xyzd values are primarily attributed to differences in aminoacid composition and packing interactions, except for the loopcomprising residue D86, previously posited to be involved in thecatalytic mechanism of CLYS (Helland et al., 2009), and after residue
Fig. 2. Stereo plot displaying the electron density of selected residues within the subtrate bcontoured at 1 σ.
82, where CLYS has an insert of four residues. The conformation ofD86 in SalG deviates significantly from that in GEWL, and instead, theloop carrying D86 in SalG is folded towards the active site andcorresponds well with what is observed in the CLYS structure(Helland et al., 2009) (Fig. 3B). The conformation of the catalyticE73 is well conserved in all three structures, as well as D97, alsoimplicated in the catalytic mechanism of g-type lysozymes (Hirakawaet al., 2008; Helland et al., 2009).
SalG is already known from its primary structure to not containany disulphide bonds unlike GEWL. On the other hand, it has twomore single salt bridges (only 2 residues involved) than GEWL and 6–7 more than CLYS (different in CLYS molecules A and D), as shown inFig. 4. All three enzymes have three salt bridge networks. One suchnetwork in SalG and CLYS incorporates eight residues via (co-ordinated by) a unique salt bridge (D41Oδ2–R176Nη1) but is dividedinto two networks of four residues each in GEWL. SalG has a slightlyhigher number (29) of residues involved in ionic interactions thanGEWL (25) and CLYS (15–17). The homology model of OEL suggeststhat the number of ionic interactions is comparable to SalG, and thenumber of amino acids involved in such interactions (27) is betweenSalG and GEWL.
Besides the lack of disulphide bonds in SalG and CLYS, and differ-ences in ion pairs, other structural features associated with cold-activity (for reviews see Smalås et al., 2000; Feller and Gerday, 2003;Georlette et al., 2004; D'Amico et al., 2006; Siddiqui and Cavicchioli,2006; Marx et al., 2007) were thoroughly examined for in the twoavian and two fish proteins studied. These included hydrogen bondsand aromatic interactions which tend to be lowered in cold-adaptedproteins. Cold-adapted proteins often have an increased number ofglycine and polar residues, a lower arginine/lysine ratio, a lower prolinecontent, less hydrophobic residues, and surface loops may be longer.Noneof these features showed any significant trendswhich could singleout SalG as cold active relative to the other proteins studied.
4. Discussion
The present study reports the structure and thermal denaturationof a g-type lysozyme from an aquatic vertebrate species. Atlanticsalmon is a benthopelagic, anadromous cold-water fish that thrivesin water temperatures between 2 and 9 °C and having a bodytemperature corresponding to its surroundings. Geese, on the otherhand have a body temperature between 40 and 42 °C and preferenvironmental temperatures ranging from 20 to 28 °C.
inding site of SalG. The 2fo–fc electron density map (blue chicken-wire mesh) of SalG

Fig. 3. A: structure based sequence alignment of SalG and three other g-type lysozymes. Secondary structure elements of SalG are shown above the alignment. α-helices are markedas blue coils and β-strands as green arrows symbolise active site residues. Accessibility of SalG residues to solvent is rendered by the bar below: white is buried (b0.1), cyan isintermediate (0.1–0.4) and blue is accessible (N 0.4). B: stereo plot displaying the superposition of SalG ( ), CLYS ( ), GEWL ( ) and a homology model of OEL ( ).Residues involved in catalysis (E73, D86 and D97) are illustrated as ball-and-stick models. NAG units identified in the CLYS structure are illustrated as yellow transparent ball-and-stick model to display the location of the substrate binding cleft. C: graph showing the average xyz displacement of superposition of the crystal structures of SalG and CLYS ( ),GEWL ( ) and the homology model of OEL ( ). The average xyz displacement on superposition of CLYS and GEWL is illustrated in gray.
258 P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
4.1. Folding and stability
There are several factors responsible for the folding and stability ofnative proteins, including hydrophobic interactions, hydrogen bond-ing and conformational entropy (Dill, 1990; Pace et al., 1996). The
relationship between activity, flexibility and stability represents acentral issue in the adaptation of SalG to its environment. SalG is coldactive as shown by its 70% relative lytic activity at 4 °C (Kyomuhendoet al., 2007), and with a Tm of 36.8 °C (this work) it is not thermallystable when compared to its counterpart OEL having a Tm of 60 °C

Fig. 4. Stereo plot illustrating the salt bridge distribution in SalG, CLYS, GEWL and OEL. Only selected residues are labelled.
259P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263

260 P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
(Kawamura et al., 2008). In addition to salmon lysozyme, a fewlysozymes have been characterised as cold active, among them alysozyme from Icelandic scallops. This invertebrate-type lysozyme isvery active at low temperatures (4–35 °C) and it regains activity afterheating for 15min at 70 °C (Nilsen et al., 1999). By visual inspection ofthe initial calorimetric profiles of SalG (Fig. 1), we observe that theyconsist of single endothermic peaks. The thermal denaturationproduces narrow, relatively symmetric peaks suggesting the transi-tions are likely two-state, reversible and highly cooperative which isfurther supported by the ratio ΔHcal/ΔHvH of close to 1. Subsequentsize exclusion gel filtration demonstrated that SalG exists as amonomer (data not shown). High cooperativity of the first unfoldingreaction indicates that SalG existed initially as a compact and well-folded structure. However, the melting profile after allowing 12 h in-situ incubation and renaturation at 4 °C is a gradual, non-cooperativemelting reaction. This indicates that the refolded protein exists as aflexible, partially unfolded protein or as a heterogeneous populationof folded structures.
Tm is an indicator of thermal stability and higher Tm yields morethermodynamically stable proteins. Even though SalG showed a Tm of36.8 °C suggestive of a thermolabile protein, the rescanned protein iseven more heat labile as seen by the lowering of Tm to 33 °C (Fig. 1).This, however, demonstrates that the initial heat-induced unfoldingwas reversible. Tm at the highest and lowest protein concentrationsused was consistent, indicating that accurate thermodynamic char-acterisation of the protein is possible at both concentrations. Thiscould also mean that intermolecular cooperativity (protein–proteininteractions) might not be important to stability. Transition temper-ature was dependent on scan rate and doubling the scan rateincreased the melting temperature by 2 °C probably due to lessenergy absorbed per unit interval.
The observed aggregation of portions of the protein samplesfollowing scanning indicates irreversible processes associated withthe thermal denaturation of the protein. These are generally underkinetic control and are sensitive to scan rate and protein concentra-tion. Data obtained at high scan rate could therefore not be fitted to atwo-state unfolding model. ΔHvH, which depends on the shape of theendotherm and assumes a true two-state reversible equilibrium,could not be determined reliably from this high scan rate data.Following calorimetry, residual lytic activity (presumably fromrefolded protein) was found to be lower for the more concentratedsample relative to that of lower concentration. This together withthermal lability of salmon lysozyme at high concentration can beexplained by the observed considerable aggregation of the protein athigher concentrations. Activity measurements agreed with the loss ofsoluble protein concentration. The significantly increased moleculardensity probably creates misfolded populations of protein moleculesthat promote aggregation and hence loss of activity.
The DSC data is consistent with observations from our earlier workwhere the protein began to lose function at about 30 °C but withstoodprolonged heating at 90 °C which we attributed to refolding(Kyomuhendo et al., 2007). In the present study, the requiredconcentrations for use in DSC were too high for spectrophotometricactivity measurements and vice versa. The implication of this is two-fold: first, low concentration of SalG tolerates heat as earlier observed(Kyomuhendo et al., 2007) and secondly, SalG at higher concentra-tions loses this paradoxical ability and becomes thermolabile. InitialDSC of SalG, where the protein was scanned up and down in thetemperature range of 0–55 °C, showed no exothermic transition onthe cooling scan to suggest thermodynamic refolding (data notshown). However, this agrees with an earlier study in which activitiesof SalG and HEWLwere followed as the reaction temperature droppedfrom an initial 60 °C revealing a gradual recovery and loss of activity,respectively (Kyomuhendo et al., 2007). This combined with therefolding observed after overnight incubation probably indicates aslow, kinetically driven refolding process.
4.2. Cold-activity
Prior to SalG, only three 3D structures of g-type lysozymes wereknown (Isaacs et al., 1985; Weaver et al., 1995; Helland et al., 2009)and we find that fish and avian lysozymes superimpose well on eachother (Fig. 3). The structural features distinguishing cold-adaptedproteins and their mesophilic counterparts have previously beenthoroughly reviewed (Smalås et al., 2000; Feller and Gerday, 2003;Georlette et al., 2004; Siddiqui and Cavicchioli, 2006; Marx et al.,2007). The cold active SalG was here compared to the mesophilicGEWL regarding acknowledged characteristics of cold-activity. In-creased structural flexibility of cold-adapted enzymes is recognised asan adaptive strategy at low temperature (Siddiqui and Cavicchioli,2006). Increased intramolecular flexibility is achieved throughweakening or loss of interactions that stabilise the native proteinmolecules thereby producing an enzyme close to the lowest stabilitywithout abolishing activity (D'Amico et al., 2001). These changes inflexibility can be local including at active sites or of the overall proteinstructure (Fields and Somero, 1998; D'Amico et al., 2001; Georletteet al., 2003). The goose and salmon lysozymes were indistinguishableor marginally different for a range of cold-adaptation features.Arguably the most distinct distinguishing feature between the salmonand goose enzymes in terms of stabilising factors with significance forcold-activity is the absence of disulphide bonds in the salmonlysozyme.
4.3. Disulphide bonds and salt bridge networks
When compared to their mesophilic and thermophilic counter-parts, cold-adapted proteins have been shown to exhibit increasedflexibility at the expense of stability. Salt bridges often serveimportant roles in catalysis, ligand binding and stability. Fourconserved charged residues involved in salt bridge networks (R72,D168, D172 and R176) in SalG, GEWL and CLYS, are characterised byvery low solvent accessible surface area. This suggests that thenetworks are essential for ensuring structural integrity.
Six substrate binding subsites, designated B–G, have beenproposed for GEWL (Honda and Fukamizo, 1998). An earlier studyshowed substrate bound to subsites B, C and D in GEWL (Weaver et al.,1995), and recently, the crystal studies of cod CLYS demonstratedadditional binding of NAG in subsites E–G (Helland et al., 2009).Whilemost of the salt bridges in GEWL and the homology model of OEL arelocated near the E–G subsites and at the N-terminal end, salt bridgesin SalG are more spread out through the molecule (Figs. 4 and 5),including both sides of the cleft forming the B–D subsites. There arefive salt bridges (R1–D3, R10–D137, D12–K141, K19–D167 and E24–H75) in the N-terminal region of GEWL compared to four salt bridgesin the corresponding region of SalG (D7–K181, D12–H138, E18–R22and D24–R75/H40) although one might be non-existent or transient(because of poor electron density observed around SalG R22).Correspondingly, four salt bridges (D7–R1/K181, D12–K141 andE24–R40 (goose and salmon templates, respectively), R10–E137 andK19–D167) are predicted in the homology model of OEL, but only onesalt bridge is found in CLYS (D24–R75).
Loss of cysteine-mediated bridges also increases flexibility(Kawamura et al., 2008). The two disulphide bridges in GEWL andthe homology model of OEL (C4–C60 and C18–C29) may stabilise themolecule by reducing conformational entropy of the unfolded stateand increase rigidity. With reference to the high sequence identity toOEL, they do this after the protein has reached its final folded form inthe absence of both disulphide bonds (Kawamura et al., 2008).Dianoux and Jollès (Dianoux and Jolles, 1967) attributed relative heatstability of HEWL vis-à-vis GEWL, to the number of disulphide bonds,four in the former and two in the latter. Similarly, a compact structureand disulphide bonds have been credited for the thermal stability ofHEWL (Masschalck and Michiels, 2003) and T4L (Wetzel et al., 1988).
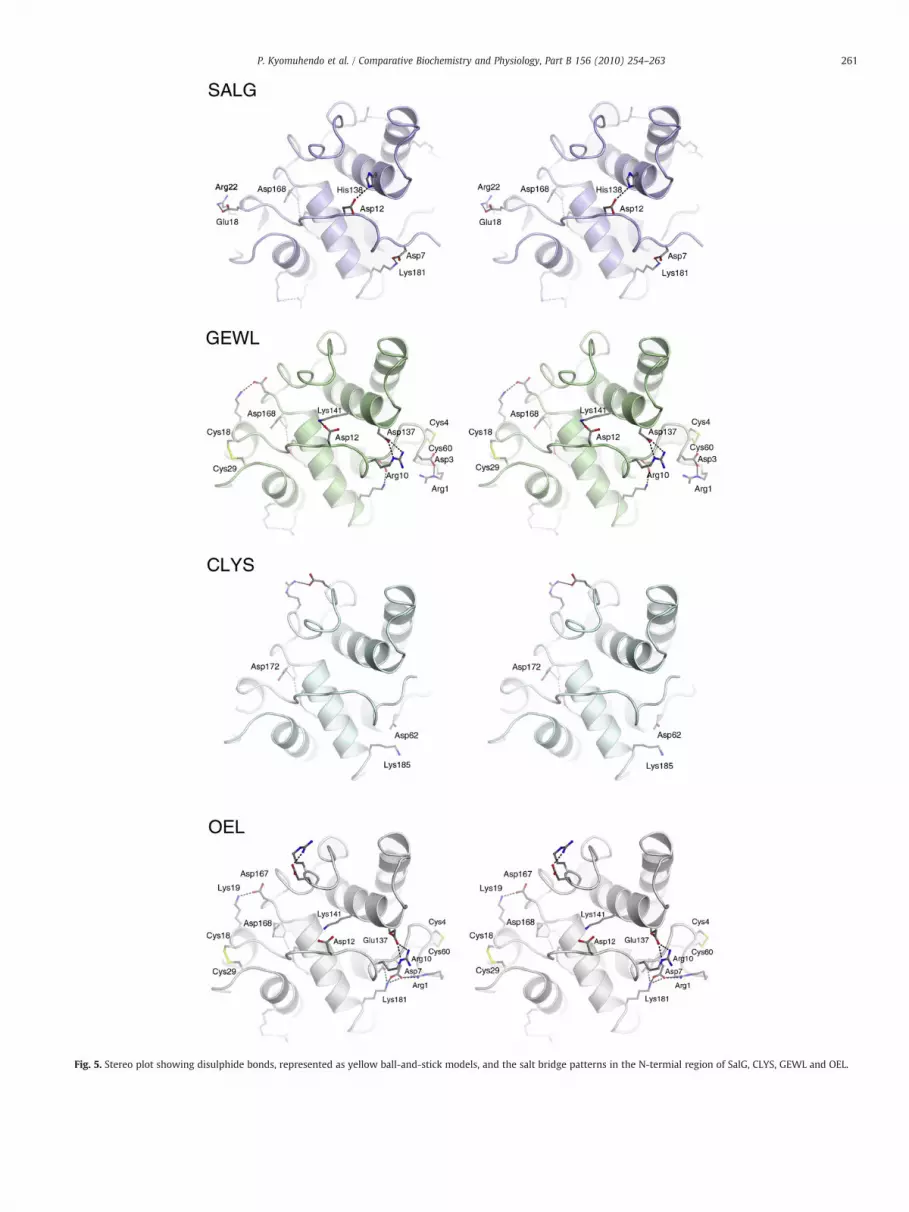
Fig. 5. Stereo plot showing disulphide bonds, represented as yellow ball-and-stick models, and the salt bridge patterns in the N-termial region of SalG, CLYS, GEWL and OEL.
261P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263

262 P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
In the latter, disulphide bondswere shown to interfere with reversiblerefolding presumably because they are chemically labile at hightemperatures. However, disulphide bonds alone cannot account forhigh heat stability as is evident from the lysozyme domain of the T7-like lytic phage φKMV (gp36C) (Lavigne et al., 2004), which lacksthese bonds and yet is highly thermostable. Conversely, the Icelandicscallop i-type lysozyme shows a low temperature optimumdespite itspredicted seven disulphide bonds (Nilsen et al., 1999). The role ofdisulphide bonds for catalytic activity and structural stability in OELhas recently been studied (Kawamura et al., 2008). The authorsreported that though crucial for the conformational stability, thesebonds were neither initiation sites for refolding nor did their absenceprevent folding into a catalytically active enzyme. Furthermore, theCys-free mutant of OEL showed similar enzyme reactivation as SalGafter heat treatment. Thammasirirak (Thammasirirak et al., 2002)found cassowary g-type lysozyme more heat-stable than GEWLdespite 83% sequence similarity and a predicted equal number ofdisulphide bridges. The authors concluded that in addition todisulphide bonds, differences in amino acid content, particularlyhydrophobic residues, led to changes in conformational stability ofsecondary and tertiary structure and hence affected thermal stability.
Our assumption is that the SalG polypeptide, due to the lack ofdisulphide bonds, would be relatively easier to unravel, especiallyfrom the N-terminus (Fig. 5), under thermal denaturation conditions.OEL containing two disulphide bonds and four salt bridges within the29 first residues, in contrast to the four salt bridges in SalG, has a Tm of60 °C (Kawamura et al., 2008) while that of SalG is 36–38 °C. SalG hasa slightly higher number and a more global distribution of salt bridgesthan GEWL, and this may improve the rigidity of the folded moleculein the absence of disulphide cross-linkages. However, caution shouldbe prescribed regarding the level of compensation offered. Moreoverso in light of their non-covalent nature as well as the observed low Tmcoupled with loss of function between 30 and 40 °C. There is someevidence that residual structure in the denatured state is stabilised bycharge–charge interactions and can influence folding (Trefethen et al.,2005), but the difference in total number of salt bridges is marginal inthe proteins albeit differing in distribution. It can thus be assumedthat these have a limited ability to prevent or delay completeunfolding of SalG where a slightly higher number was observed.However, they may influence folding considering that they may helpstabilise any residual structure in the denatured state thus forming agood starting point for refolding to take place. The increased flexibilitydue to the absence of disulphide bonds may help SalG to be active atthe low temperatures prevalent in the habitat occupied by Atlanticsalmon.
5. Conclusions
The recombinant g-type lysozyme from Atlantic salmon waspreviously characterised as a cold active enzyme with remarkablereversion to activity after prolonged heating. In this study it has beensubjected to thermal denaturation and structure analysis. It is clearfrom this and earlier studies that low concentrations of SalG toleratethermal-induced denaturation as opposed to high concentrations ofthe protein such as those used for calorimetry. The salmon lysozyme3D structure bears close resemblance to that of goose and codlysozyme. We have correlated its biochemical, biophysical, andstructural properties and concluded that the lack of disulphidebonds plays a great role in the thermal behaviour observed for SalG.In addition, the location of two disulphide bonds within the first 29residues at the N-terminus of GEWL and OEL, in combination withfour to five salt bridges effectively prevents the terminal from beingan initiation site for denaturation. The opposite is likely in SalG withno disulphide bonds and with only four salt bridges in thecorresponding region, thus probably contributing to the lower Tm.
The slightly higher number of salt bridges and their wider distributionmight be a factor in assisting SalG to refold.
Therefore, the results from the present work provide explanationat the atomic level for important biological properties of salmonlysozyme, and the information obtained here contributes to the generalknowledge of the structure–function relationship in lysozymes.
Acknowledgements
We thank the Research Council of Norway, the National FunctionalGenomics Program (FUGE) and the Nofima Marine for funding thisproject. Provision of beamtime at the Protein Structure Factorybeamlines of Berliner Elektronenspeicherring (BESSY) and theSwiss–Norwegian Beamlines (SNBL) at European Synchrotron Radi-ation Facility (ESRF) is gratefully acknowledged.
References
Arnold, K., Bordoli, L., Kopp, J., Schwede, T., 2006. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics 22,195–201.
Betz, S.F., 1993. Disulfide bonds and the stability of globular proteins. Protein Sci. 2,1551–1558.
Collaborative Computational Project, N., 1994. The CCP4 Suite: programs for proteincrystallography. Acta Crystallogr., Sect. D: Biol Crystallogr. 50, 760–763.
Cooper, A., Eyles, S.J., Radford, S.E., Dobson, C.M., 1992. Thermodynamic consequencesof the removal of a disulfide bridge from hen lysozyme. J. Mol. Biol. 225, 939–943.
D'Amico, S., Gerday, C., Feller, G., 2001. Structural determinants of cold adaptation andstability in a large protein. J. Biol. Chem. 276, 25791–25796.
D'Amico, S., Collins, T., Marx, J.C., Feller, G., Gerday, C., 2006. Psychrophilicmicroorganisms: challenges for life. EMBO Rep. 7, 385–389.
DeLano, W.L., 2002. The PyMOL Molecular Graphics System. DeLano Scientific, PaloAlto, CA, USA.
Dianoux, A.C., Jolles, P., 1967. Study of a lysozyme poor in cystine and tryptophan: thelysozyme of goose egg-white. Biochim. Biophys. Acta 133, 472–479.
Dill, K.A., 1990. Dominant forces in protein folding. Biochemistry-US 29, 7133–7155.Evans, P.R., 1997. SCALA. Proceedings of the CCP4 Study Weekend. Recent Advances in
Phasing. In: Wilson, K.S., Davies, G., Ashton, A.W., Bailey, S. (Eds.), DaresburyLaboratory, Warrington, pp. 97–102.
Feller, G., Gerday, C., 2003. Psychrophilic enzymes: hot topics in cold adaptation. Nat.Rev. Microbiol. 1, 200–208.
Feller, G., Narinx, E., Arpigny, J.L., Aittaleb, M., Baise, E., Genicot, S., Gerday, C., 1996.Enzymes from psychrophilic organisms. FEMS Microbiol. Rev. 18, 189–202.
Fields, P.A., Somero, G.N., 1998. Hot spots in cold adaptation: localized increases inconformational flexibility in lactate dehydrogenase A(4) orthologs of Antarcticnotothenioid fishes. Proc. Natl. Acad. Sci. U. S. A. 95, 11476–11481.
Georlette, D., Damien, B., Blaise, V., Depiereux, E., Uversky, V.N., Gerday, C., Feller, G.,2003. Structural and functional adaptations to extreme temperatures in psychro-philic, mesophilic, and thermophilic DNA ligases. J. Biol. Chem. 278, 37015–37023.
Georlette, D., Blaise, V., Collins, T., D'Amico, S., Gratia, E., Hoyoux, A., Marx, J.C., Sonan, G.,Feller, G., Gerday, C., 2004. Some like it cold: biocatalysis at low temperatures. FEMSMicrobiol. Rev. 28, 25–42.
Gille, C., Frommel, C., 2001. STRAP: editor for Structural Alignments of Proteins.Bioinformatics 17, 377–378.
Goto, T., Abe, Y., Kakuta, Y., Takeshita, K., Imoto, T., Ueda, T., 2007. Crystal structure ofTapes japonica Lysozyme with substrate analogue: structural basis of the catalyticmechanism and manifestation of its chitinase activity accompanied by quaternarystructural change. J. Biol. Chem. 282, 27459–27467.
Gouet, P., Courcelle, E., Stuart, D.I., Metoz, F., 1999. ESPript: analysis of multiplesequence alignments in PostScript. Bioinformatics 15, 305–308.
Grütter, M.G., Weaver, L.H., Matthews, B.W., 1983. Goose lysozyme structure: anevolutionary link between hen and bacteriophage lysozymes? Nature 303,828–831.
Helland, R., Larsen, L.R., Finstad, S., Kyomuhendo, P., Larsen, A.N., 2009. The crystalstructure of g-type lysozyme from Atlantic cod (Gadus morhua L.) in complex withNAG oligomers sheds new light on substrate binding and the catalytic mechanism.Cell. Mol. Life Sci. 66, 2585–2598.
Hikima, J., Minagawa, S., Hirono, I., Aoki, T., 2001. Molecular cloning, expression andevolution of the Japanese flounder goose-type lysozyme gene, and the lytic activityof its recombinant protein. Biochim. Biophys. Acta 1520, 35–44.
Hirakawa, H., Ochi, A., Kawahara, Y., Kawamura, S., Torikata, T., Kuhara, S., 2008.Catalytic reaction mechanism of goose egg-white lysozyme by molecularmodelling of enzyme substrate complex. J. Biochem. 144, 753–761.
Honda, Y., Fukamizo, T., 1998. Substrate binding subsites of chitinase from barley seedsand lysozyme from goose egg-white. Biochim. Biophys. Acta. Prot. Struct. Mol. Enz.1388, 53–65.
Irwin, D.M., Gong, Z., 2003. Molecular evolution of vertebrate goose-type lysozymegenes. J. Mol. Evol. 56, 234–242.
Isaacs, N.W., Machin, K.J., Masakuni, M., 1985. 3-Dimensional structure of goose-typelysozyme from the egg-white of the Australian Black Swan, Cygnus-Atratus. Aust. J.Biol. Sci. 38, 13–22.

263P. Kyomuhendo et al. / Comparative Biochemistry and Physiology, Part B 156 (2010) 254–263
Ito, Y., Yoshikawa, A., Hotani, T., Fukuda, S., Sugimura, K., Imoto, T., 1999. Amino acidsequences of lysozymes newly purified from invertebrates imply wide distributionof a novel class in the lysozyme family. Eur. J. Biochem. 259, 456–461.
Jollès, P., 1996. Lysozymes: Model Enzymes in Biochemistry and Biology. BirkhäuserVerlag, Basel.
Jollès, P., Jollès, J., 1984. What's new in lysozyme research? Always a model system,today as yesterday. Mol. Cell. Biochem. 63, 165–189.
Jones, T.A., Zou, J.Y., Cowan, S.W., Kjeldgaard, M., 1991. Improved methods for buildingprotein models in electron-density maps and the location of errors in these models.Acta Crystallogr., Sect. A: Found Crystallogr. 47, 110–119.
Kabsch, W., Sander, C., 1983. Dictionary of protein secondary structure — pattern-recognition of hydrogen-bonded and geometrical features. Biopolymers 22,2577–2637.
Kawamura, S., Ohno, K., Ohkuma, M., Chijiiwa, Y., Torikata, T., 2006. Experimentalverification of the crucial roles of Glu(73) in the catalytic activity and structuralstability of goose type lysozyme. J. Biochem. 140, 75–85.
Kawamura, S., Ohkuma, M., Chijiiwa, Y., Kohno, D., Nakagawa, H., Hirakawa, H., Kuhara,S., Torikata, T., 2008. Role of disulfide bonds in goose-type lysozyme. FEBS J. 275,2818–2830.
Kyomuhendo, P., Myrnes, B., Nilsen, I.W., 2007. A cold active salmon goose-typelysozyme with high heat tolerance. Cell. Moll. Life. Sci. 64, 2841–2847.
Larsen, A.N., Solstad, T., Svineng, G., Seppola, M., Jorgensen, T.O., 2009. Molecularcharacterisation of a goose-type lysozyme gene in Atlantic cod (Gadus morhua L.).Fish Shellfish Immunol 26, 122–132.
Lavigne, R., Briers, Y., Hertveldt, K., Robben, J., Volckaert, G., 2004. Identification andcharacterization of a highly thermostable bacteriophage lysozyme. Cell. Moll. Life.Sci. 61, 2753–2759.
Leslie, A.G.W., 1992. Recent changes to the MOSFLM package for processing film andimage plate data. In Joint CCP4 and ESF-EACBM Newsletters on ProteinCrystallography. Daresbury Laboratory, Warrington, UK, p. 26.
Maenaka, K., Matsushima, M., Song, H., Sunada, F., Watanabe, K., Kumagai, I., 1995.Dissection of protein–carbohydrate interactions in mutant hen egg-whitelysozyme complexes and their hydrolytic activity. J. Mol. Biol. 247, 281–293.
Marx, J.C., Collins, T., D'Amico, S., Feller, G., Gerday, C., 2007. Cold-adapted enzymesfrom marine Antarctic microorganisms. Mar. Biotechnol. (NY) 9, 293–304.
Masschalck, B., Michiels, C.W., 2003. Antimicrobial properties of lysozyme in relation tofoodborne vegetative bacteria. Crit. Rev. Microbiol. 29, 191–214.
Matsuo, K., Watanabe, H., Tate, S.I., Tachibana, H., Gekko, K., 2009. Comprehensivesecondary-structure analysis of disulfide variants of lysozyme by synchrotron-radiation vacuum-ultraviolet circular dichroism. Proteins 77, 191–201.
McDonald, I.K., Thornton, J.M., 1994. Satisfying hydrogen-bonding potential in proteins.J. Mol. Biol. 238, 777–793.
Murshudov, G.N., Vagin, A.A., Dodson, E.J., 1997. Refinement of macromolecularstructures by the maximum-likelihood method. Acta Crystallogr., Sect. D: Biol.Crystallogr. 53, 240–255.
Nilsen, I.W., Overbo, K., Sandsdalen, E., Sandaker, E., Sletten, K., Myrnes, B., 1999.Protein purification and gene isolation of chlamysin, a cold-active lysozyme-likeenzyme with antibacterial activity. FEBS Lett. 464, 153–158.
Nilsen, I.W., Myrnes, B., Edvardsen, R.B., Chourrout, D., 2003. Urochordates carrymultiple genes for goose-type lysozyme and no genes for chicken- or invertebrate-type lysozymes. Cell. Moll. Life. Sci. 60, 2210–2218.
Pace, C.N., Grimsley, G.R., Thomson, J.A., Barnett, B.J., 1988. Conformational stability andactivity of ribonuclease-T1 with zero, one, and two intact disulfide bonds. J. Biol.Chem. 263, 11820–11825.
Pace, C.N., Shirley, B.A., McNutt, M., Gajiwala, K., 1996. Forces contributing to theconformational stability of proteins. FASEB J. 10, 75–83.
Perrakis, A., Morris, R., Lamzin, V.S., 1999. Automated protein model building combinedwith iterative structure refinement. Nat. Struct. Biol. 6, 458–463.
Pooart, J., Torikata, T., Araki, T., 2005. Enzymatic properties of rhea lysozyme. Biosci.,Biotechnol. Biochem. 69, 103–112.
Savan, R., Aman, A., Sakai, M., 2003. Molecular cloning of G type lysozyme cDNA incommon carp (Cyprinus carpio L.). Fish Shellfish Immunol 15, 263–268.
Siddiqui, K.S., Cavicchioli, R., 2005. Improved thermal stability and activity in the cold-adapted lipase B from Candida antarctica following chemical modification withoxidized polysaccharides. Extremophiles 9, 471–476.
Siddiqui, K.S., Cavicchioli, R., 2006. Cold-adapted enzymes. Annu. Rev. Biochem. 75,403–433.
Smalås, A.O., Leiros, H.H., Os, V., Willassen, N.P., 2000. Cold adapted enzymes.Biotechnol. Annu. Rev. 6, 1–57.
Thammasirirak, S., Torikata, T., Takami, K., Murata, K., Araki, T., 2001. Purification andcharacterization of goose type lysozyme from cassowary (Casuarius casuarius) egg-white. Biosci., Biotechnol. Biochem. 65, 584–592.
Thammasirirak, S., Torikata, T., Takami, K., Murata, K., Araki, T., 2002. Study of thephysicochemical stability of cassowary (Casuarius casuarius) goose-type lysozyme.Proceedings of Faculty of Agriculture Kyushu Tokai University 21, 9–15.
Trefethen, J.M., Pace, C.N., Scholtz, J.M., Brems, D.N., 2005. Charge–charge interactionsin the denatured state influence the folding kinetics of ribonuclease Sa. Protein Sci.14, 1934–1938.
Vagin, A.A.T.A., 1997. MOLREP: an automated program for molecular replacement.J. Appl. Crystallogr. 30, 1022–1025.
Vriend, G., 1990. What If — a molecular modeling and dug design program. J. Mol.Graph. 8, 52–56.
Weaver, L.H., Matthews, B.W., 1987. Structure of bacteriophage-T4 lysozyme refined at1.7 a resolution. J. Mol. Biol. 193, 189–199.
Weaver, L.H., Grutter, M.G., Remington, S.J., Gray, T.M., Isaacs, N.W., Matthews, B.W.,1984. Comparison of goose-type, chicken-type, and phage-type lysozymesillustrates the changes that occur in both amino acid sequence and three-dimensional structure during evolution. J. Mol. Evol. 21, 97–111.
Weaver, L.H., Grutter, M.G., Matthews, B.W., 1995. The refined structures of gooselysozyme and its complex with a bound trisaccharide show that the “goose-type”lysozymes lack a catalytic aspartate residue. J. Mol. Biol. 245, 54–68.
Wetzel, R., Perry, L.J., Baase, W.A., Becktel, W.J., 1988. Disulfide bonds and thermal-stability in T4 lysozyme. Proc. Natl. Acad. Sci. U. S. A. 85, 401–405.
Yin, Z.X., He, J.G., Deng, W.X., Chan, S.M., 2003. Molecular cloning, expression of orange-spotted grouper goose-type lysozyme cDNA, and lytic activity of its recombinantprotein. Dis. Aquatic Org. 55, 117–123.
Zhao, J., Song, L., Li, C., Zou, H., Ni, D., Wang, W., Xu, W., 2007. Molecular cloning of aninvertebrate goose-type lysozyme gene from Chlamys farreri, and lytic activity ofthe recombinant protein. Mol. Immunol. 44, 1198–1208.
Zhou, Y.Q., Hall, C.K., Karplus, M., 1999. The calorimetric criterion for a two-stateprocess revisited. Protein Sci. 8, 1064–1074.
Zou, H., Song, L., Xu, W., Yang, G., 2005. Molecular cloning and characterisation analysisof cDNA encoding g-type lysozyme from scallop (Argopectens irradians). High Tech.Lett. 15, 101–106.