the signature of seeds in resurrection plants: a molecular and physiological comparison of...
TRANSCRIPT

771
INTEGR. COMP. BIOL., 45:771–787 (2005)
The Signature of Seeds in Resurrection Plants: A Molecular and Physiological Comparison ofDesiccation Tolerance in Seeds and Vegetative Tissues1
NICOLA ILLING, KATHERINE J. DENBY, HELEN COLLETT, ARTHUR SHEN, AND JILL M. FARRANT2
Department of Molecular and Cell Biology, University of Cape Town, Private Bag, Rondebosch, 7701, South Africa.
SYNOPSIS. Desiccation-tolerance in vegetative tissues of angiosperms has a polyphyletic origin and couldbe due to 1) appropriation of the seed-specific program of gene expression that protects orthodox seedsagainst desiccation, and/or 2) a sustainable version of the abiotic stress response. We tested these hypothesesby comparing molecular and physiological data from the development of orthodox seeds, the response ofdesiccation-sensitive plants to abiotic stress, and the response of desiccation-tolerant plants to extreme waterloss. Analysis of publicly-available gene expression data of 35 LEA proteins and 68 anti-oxidant enzymes inthe desiccation-sensitive Arabidopsis thaliana identified 13 LEAs and 4 anti-oxidants exclusively expressedin seeds. Two (a LEA6 and 1-cys-peroxiredoxin) are not expressed in vegetative tissues in A. thaliana, buthave orthologues that are specifically activated in desiccating leaves of Xerophyta humilis. A comparison ofantioxidant enzyme activity in two desiccation-sensitive species of Eragrostis with the desiccation-tolerant E.nindensis showed equivalent responses upon initial dehydration, but activity was retained at low watercontent in E. nindensis only. We propose that these antioxidants are housekeeping enzymes and that theyare protected from damage in the desiccation-tolerant species. Sucrose is considered an important protectantagainst desiccation in orthodox seeds, and we show that sucrose accumulates in drying leaves of E. nindensis,but not in the desiccation-sensitive Eragrostis species. The activation of ‘‘seed-specific’’ desiccation protectionmechanisms (sucrose accumulation and expression of LEA6 and 1-cys-peroxiredoxin genes) in the vegetativetissues of desiccation-tolerant plants points towards acquisition of desiccation tolerance from seeds.
INTRODUCTION
Desiccation tolerance (DT) is the ability of an or-ganism to survive the loss of most (.95%) of its cel-lular water. It is considered to be a complex trait thatis present in reproductive structures (pollen and seeds)of vascular plants. Desiccation tolerance in vegetativetissues is also relatively common in less complexplants such as bryophytes (Proctor, 1990) and lichens(Kappen and Valadares, 1999) but rare in pterido-phytes and angiosperms (Gaff, 1977, 1987; Alpert andOliver, 2002; Porembski and Barthlott, 2000) and ab-sent in gymnosperms (Gaff, 1980). Oliver et al. (2000)have speculated on the evolution of DT in vegetativetissue and argue that DT is the ancestral state for earlyland plants (e.g., mosses), which was lost early in theevolution of tracheophytes. The subsequent successfulradiation of vascular plants on land was probably aconsequence of the evolution of DT in seeds, in par-allel to the evolution of structural and morphologicalmodifications in vegetative tissue which allowed great-er control of water status. Oliver et al. (2000) speculatethat the emergence of DT in seeds was a modificationof vegetative DT in early ancestors. They suggest fur-thermore that vegetative DT in angiosperms subse-quently re-evolved independently at least eight timesas an adaptation of seed DT.
DT in seeds and vegetative tissues of angiospermsis different from that in extant lower orders. In the
1 From the Symposium Drying Without Dying: The ComparativeMechanisms and Evolution of Desiccation Tolerance in Animals,Microbes, and Plants presented at the Annual Meeting of the Societyfor Integrative and Comparative Biology, 4–8 January 2005, at SanDiego, California.
2 E-mail: [email protected]
former, it is based on induction of several relativelycomplex protection mechanisms during drying, withminimal reliance on repair of desiccation-induceddamage during rehydration (Gaff, 1989; Vertucci andFarrant, 1995; Oliver, 1996; Oliver et al., 2000). In thelower order plants, DT is constitutive, less reliant oncomplex cellular protection and more on repair duringrehydration (Bewley and Oliver, 1992; Oliver andBewley, 1997; Oliver et al., 2000). Thus, although DTin seeds appears to include some mechanisms presentin DT vegetative tissues of less-complex plants (re-viewed in Oliver, 1996; Dickie and Pritchard, 2002),additional, more sophisticated mechanisms of protec-tion must have evolved. We examine the possibilitythat DT in the vegetative tissues of angiosperms is asubsequent adaptation of these seed developmentally-regulated mechanisms.
Whereas DT is developmentally regulated in seedswith the putative mechanisms of tolerance being ac-cumulated only at precise times after fertilisation,drought is stochastic and DT vegetative tissues mustrespond to environmental signals to activate protectivemechanisms for the whole plant. Conceivably, envi-ronmental signals for extreme water loss activate anexisting repertoire of protective seed-specific genes invegetative tissue.
An alternative hypothesis is that DT evolved fromthe response of desiccation-sensitive plants to abioticstresses such as cold, salt and drought. Desiccation-sensitive plants use an interconnected signaling net-work to activate a common repertoire of responses toabiotic stress (Knight and Knight, 2001). These re-sponses appear to overlap with those described for DTplants during extreme water loss as they include the

772 N. ILLING ET AL.
accumulation of compatible osmolytes, and the up-reg-ulation of anti-oxidants and anti-oxidant enzymes(Knight and Knight, 2001). Under this scenario, wewould predict that there should be significant overlapbetween genes that are induced in response to abioticstresses and DT.
The stresses associated with extreme water loss in-clude the mechanical stress associated with turgor loss,oxidative stress from free radical-mediated processesand the destabilization or loss of macromolecular in-tegrity (reviewed by Vertucci and Farrant, 1995; Oli-ver and Bewley, 1997; Walters et al., 2002). Protectionmechanisms associated with prevention of damagefrom these stresses have been extensively reviewedand the maintenance of subcellular integrity has beenwidely attributed to the accumulation of stress-asso-ciated proteins, non-reducing sugars and free radicalscavenging systems (for example Farrant, 2000; Scott,2000; Oliver et al., 2000; Vicre et al., 2004).
Late Embryo Abundant (LEA) proteins accumulateduring the onset of DT in orthodox seeds and havebeen shown to occur in response to the drying of DTvegetative tissues (Bray, 1997; Ingram and Bartels,1996). Expression of LEAs has also been reported tobe up-regulated during abiotic stress such as cold,drought and osmotic stress (Wise and Tunnacliffe,2004). Several LEA mRNA transcripts and proteinshave been identified as being up-regulated in leaves ofthe resurrection plants Craterostigma plantagineum(4) and Xerophyta viscosa (2) during a cycle of des-iccation (Piatkowski et al., 1990; Mundree and Far-rant, 2000; Ndima et al., 2001). The largest set ofLEAs (16) from a single resurrection plant was re-cently identified in a mini-microarray screen of 400cDNAs from Xerophyta humilis (Collett et al., 2004).LEAs are low complexity proteins which have beenclassified into several unrelated groups on the basis ofconservation of peptide motifs (or Pfam domains)(Close, 1997; Wise, 2003). It has not been possible toassign structures to LEAs, or experimentally determinetheir exact cellular role because they are unfolded inthe hydrated state. Based on evidence of their abun-dant expression, and rich hydrophilic amino acid con-tent, it has been proposed that LEAs maintain subcel-lular integrity by protecting cellular structures from theeffects of water loss by either acting as a hydrationbuffer, by sequestering ions, by direct protection ofother proteins or membranes, or by renaturing unfold-ed proteins (Bray, 1991). Recently, it was shown thattwo LEAs can prevent protein aggregation during wa-ter stress (Goyal et al., 2005) and the ability of plantLEAs to confer increased tolerance to water deficitstress on yeast and other plants (for example, Swire-Clark and Marcotte [1999], Xu et al. [1996] and Siv-amani et al. [2000]) also suggests LEAs play an im-portant role in protecting tissues from the effects ofwater loss.
Sucrose is the only sugar commonly accumulated inDT tissues of seeds and resurrection plants (Scott,2000; Ghasempour et al., 1998). Like LEAs, it is com-
monly acknowledged to play an important and variedrole in DT, although the exact nature of this in vivohas yet to be demonstrated. The general protectiveroles ascribed to sucrose are as water replacement mol-ecules and a facilitator, together with proteins, of glassformation (Leopold et al., 1994; Vertucci and Farrant,1995; Crowe et al., 1998). It has also been suggestedthat the formation of sucrose reduces the monosaccha-ride pool, which in turn reduces the chances of dam-aging Maillard-type reactions occurring and puts a sta-sis on respiratory metabolism, both of which reducefree radical formation (Vertucci and Farrant, 1995).
Free radical scavenging systems are ubiquitous inplants and include well known antioxidants such asascorbate, glutathione and tocopherol, and enzymessuch as the peroxidases (ascorbate peroxidase [AP],glutathione peroxidase, thioredoxin peroxidase, cata-lase), glutathione reductase (GR) and superoxide dis-mutase (SOD) inter alia. Other anti-oxidant enzymessuch as 1-cys peroxiredoxin have been identified in theseeds of desiccation-sensitive angiosperms (Aalen,1999) and recently in the vegetative tissues of the res-urrection plant X. viscosa (Mowla et al., 2002).
We have focused on the above-mentioned mecha-nisms of DT to test the hypothesis that DT in vege-tative tissue in resurrection plants is an adaptation ofDT in seeds. We have compared some of the putativemechanisms of tolerance in the seeds of desiccation-sensitive angiosperms with those found to occur dur-ing desiccation of resurrection plants (viz. the accu-mulation of LEA proteins, anti-oxidants and sucrose)using a combination of molecular and physiologicalstudies. In this study we aim to distinguish betweenseed-specific and abiotic stress-specific LEA and anti-oxidant enzyme gene expression in Arabidopsis witha view to showing that some of the ‘‘seed-specific’’genes are expressed in the vegetative tissue of resur-rection plants such as X. humilis during desiccation.We also compare antioxidant activity and sucrose ac-cumulation in the seeds and vegetative tissue of mis-cellaneous desiccation-sensitive and desiccation-toler-ant plants to test our hypotheses.
MATERIALS AND METHODS
Plant material
Resurrection plants were collected and maintainedin a glasshouse at the University of Cape Town aspreviously reported (Sherwin and Farrant, 1996; Daceet al., 1998; Vander Willigen et al., 2001). Plants ofEragrostis curvula and E. teff were grown from seedssown in seedling flats (1:1 potting soil:river sand mix)and maintained in the glasshouse for three months. Pri-or to experimentation, fully hydrated plants were trans-ferred to a controlled environment chamber where theywere acclimated to conditions of 50–65% relative hu-midity, 14 hr light (500 mmol photons·m22·s21; 258C):10 hr dark (188C). After two weeks plants were usedfor dehydration/rehydration experiments. Plants werenaturally dried by withholding water until the plants
773SIGNATURE OF SEEDS IN RESURRECTION PLANTS
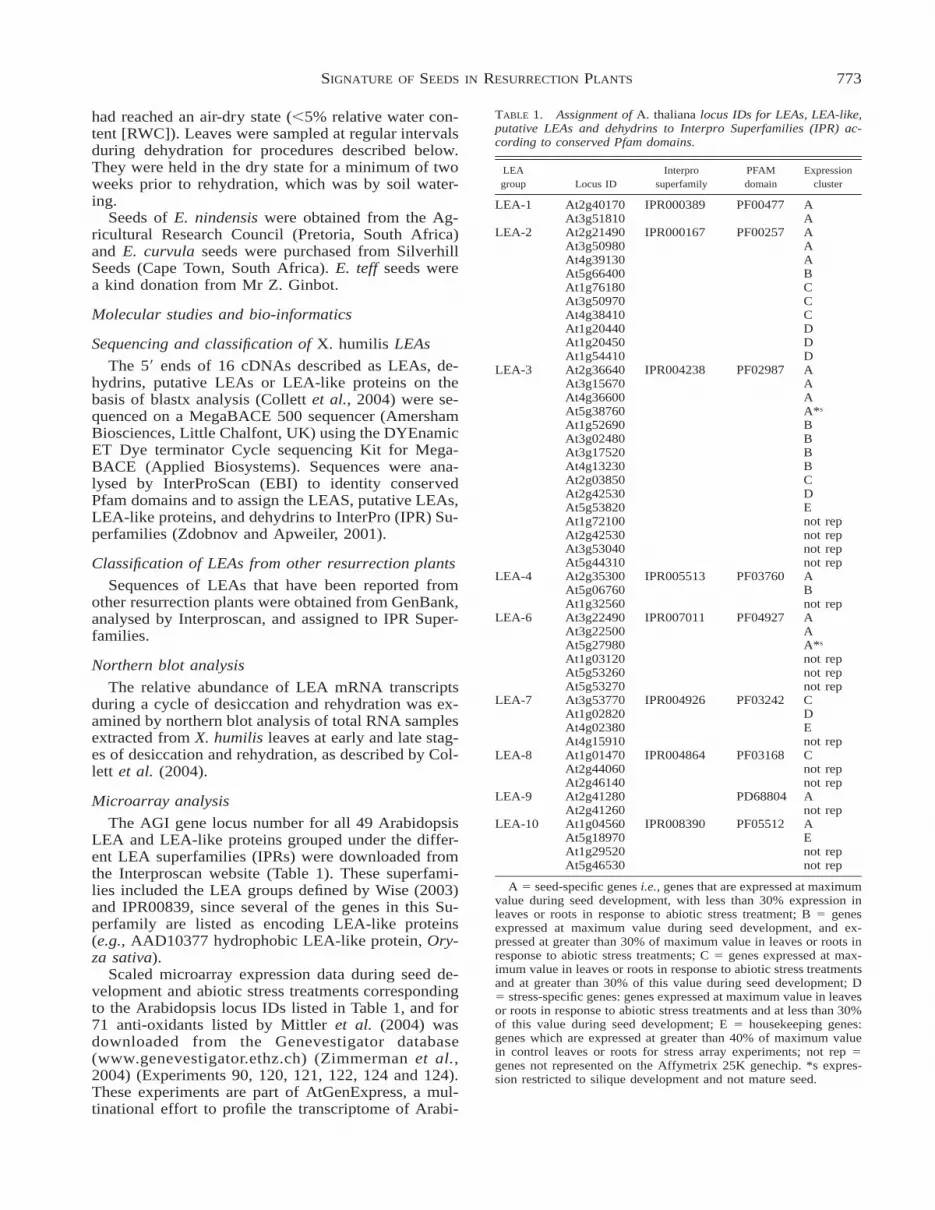
TABLE 1. Assignment of A. thaliana locus IDs for LEAs, LEA-like,putative LEAs and dehydrins to Interpro Superfamilies (IPR) ac-cording to conserved Pfam domains.
LEAgroup Locus ID
Interprosuperfamily
PFAMdomain
Expressioncluster
LEA-1 At2g40170 IPR000389 PF00477 AAt3g51810 A
LEA-2 At2g21490 IPR000167 PF00257 AAt3g50980 AAt4g39130 AAt5g66400 BAt1g76180 CAt3g50970 CAt4g38410 CAt1g20440 DAt1g20450 DAt1g54410 D
LEA-3 At2g36640 IPR004238 PF02987 AAt3g15670 AAt4g36600 AAt5g38760 A*s
At1g52690 BAt3g02480 BAt3g17520 BAt4g13230 BAt2g03850 CAt2g42530 DAt5g53820 EAt1g72100 not repAt2g42530 not repAt3g53040 not repAt5g44310 not rep
LEA-4 At2g35300 IPR005513 PF03760 AAt5g06760 BAt1g32560 not rep
LEA-6 At3g22490 IPR007011 PF04927 AAt3g22500 AAt5g27980 A*s
At1g03120 not repAt5g53260 not repAt5g53270 not rep
LEA-7 At3g53770 IPR004926 PF03242 CAt1g02820 DAt4g02380 EAt4g15910 not rep
LEA-8 At1g01470 IPR004864 PF03168 CAt2g44060 not repAt2g46140 not rep
LEA-9 At2g41280 PD68804 AAt2g41260 not rep
LEA-10 At1g04560 IPR008390 PF05512 AAt5g18970 EAt1g29520 not repAt5g46530 not rep
A 5 seed-specific genes i.e., genes that are expressed at maximumvalue during seed development, with less than 30% expression inleaves or roots in response to abiotic stress treatment; B 5 genesexpressed at maximum value during seed development, and ex-pressed at greater than 30% of maximum value in leaves or roots inresponse to abiotic stress treatments; C 5 genes expressed at max-imum value in leaves or roots in response to abiotic stress treatmentsand at greater than 30% of this value during seed development; D5 stress-specific genes: genes expressed at maximum value in leavesor roots in response to abiotic stress treatments and at less than 30%of this value during seed development; E 5 housekeeping genes:genes which are expressed at greater than 40% of maximum valuein control leaves or roots for stress array experiments; not rep 5genes not represented on the Affymetrix 25K genechip. *s expres-sion restricted to silique development and not mature seed.
had reached an air-dry state (,5% relative water con-tent [RWC]). Leaves were sampled at regular intervalsduring dehydration for procedures described below.They were held in the dry state for a minimum of twoweeks prior to rehydration, which was by soil water-ing.
Seeds of E. nindensis were obtained from the Ag-ricultural Research Council (Pretoria, South Africa)and E. curvula seeds were purchased from SilverhillSeeds (Cape Town, South Africa). E. teff seeds werea kind donation from Mr Z. Ginbot.
Molecular studies and bio-informatics
Sequencing and classification of X. humilis LEAs
The 59 ends of 16 cDNAs described as LEAs, de-hydrins, putative LEAs or LEA-like proteins on thebasis of blastx analysis (Collett et al., 2004) were se-quenced on a MegaBACE 500 sequencer (AmershamBiosciences, Little Chalfont, UK) using the DYEnamicET Dye terminator Cycle sequencing Kit for Mega-BACE (Applied Biosystems). Sequences were ana-lysed by InterProScan (EBI) to identity conservedPfam domains and to assign the LEAS, putative LEAs,LEA-like proteins, and dehydrins to InterPro (IPR) Su-perfamilies (Zdobnov and Apweiler, 2001).
Classification of LEAs from other resurrection plants
Sequences of LEAs that have been reported fromother resurrection plants were obtained from GenBank,analysed by Interproscan, and assigned to IPR Super-families.
Northern blot analysis
The relative abundance of LEA mRNA transcriptsduring a cycle of desiccation and rehydration was ex-amined by northern blot analysis of total RNA samplesextracted from X. humilis leaves at early and late stag-es of desiccation and rehydration, as described by Col-lett et al. (2004).
Microarray analysis
The AGI gene locus number for all 49 ArabidopsisLEA and LEA-like proteins grouped under the differ-ent LEA superfamilies (IPRs) were downloaded fromthe Interproscan website (Table 1). These superfami-lies included the LEA groups defined by Wise (2003)and IPR00839, since several of the genes in this Su-perfamily are listed as encoding LEA-like proteins(e.g., AAD10377 hydrophobic LEA-like protein, Ory-za sativa).
Scaled microarray expression data during seed de-velopment and abiotic stress treatments correspondingto the Arabidopsis locus IDs listed in Table 1, and for71 anti-oxidants listed by Mittler et al. (2004) wasdownloaded from the Genevestigator database(www.genevestigator.ethz.ch) (Zimmerman et al.,2004) (Experiments 90, 120, 121, 122, 124 and 124).These experiments are part of AtGenExpress, a mul-tinational effort to profile the transcriptome of Arabi-

774 N. ILLING ET AL.
dopsis (http://web.uni-frankfurt.de/fb15/botanik/mcb/AFGN/atgenex.htm). 35 out of the 49 ArabidopsisLEAs were present on the Affymetrix 25K chip usedin these experiments. We checked that the distributionof signal intensities for each of the full datasets wassimilar to validate our cross-comparison between theseed development and abiotic stress series.
Average signal values were calculated as the meanfrom the repeats for each data point in the silique/seedand the abiotic stress series. A general signal value foreach gene in control plants was determined by takingthe mean of the control data points for green shoot androot respectively from the abiotic stress series. Foreach gene, each average signal intensity in the siliqueand seed development series and in the abiotic stressexperiments, as well as the general control values wereexpressed as a percentage of the maximum valueacross all the experiments. Genes were manuallygrouped into the following clusters based on their pro-files of expression; A) Seed specific genes: maximumexpression in seed, less than 30% of the maximumexpression in any of the abiotic stress treatments, B)maximum expression in seed and more than 30% ofthe maximum expression in any of the abiotic stresstreatments, C) maximum expression in one of the abi-otic stress treatments as well as more than 30% of themaximum expression during seed development, D)Stress specific genes: maximum expression in one ofthe abiotic stress treatments and less than 30% of themaximum expression during seed development and E)Housekeeping genes: expression in the control plantsis more than 40% of the maximum expression valueof either the seed development or abiotic stress treat-ment series. The average expression in controls wasless than 30% of the maximum expression value ingroups A–D.
Physiological studies
Relative water content (RWC) determination
The RWC was calculated as the water content di-vided by the water content estimated at full turgor. Themeans of the water content of leaves at full turgor wererecorded for each species using more than 20 repre-sentative leaf samples from plants that had been fullyhydrated overnight in plastic bags. Water contentswere gravimetrically determined by oven drying at708C for 48 hr.
Antioxidant enzyme activity
Leaf tissues and seeds were extracted and analysedfor AP, GR and SOD analysis as described previously(Farrant et al., 2003, 2004). Three 250 mg replicatesof leaf tissue were ground in liquid nitrogen and ex-tracted in 3 ml of cold buffer (0.1 M phosphate bufferpH 7.8; 2 mM dithiothreitol [DTT]; 0.1 mM EDTA;1.25 mM PEG 4000; 0.1 g insoluble polyvinylpyrrol-idone [PVP]). The extract was centrifuged at 16,000 gfor 15 min and the supernatant was desalted on a Se-phadex G-25 PD10 column (Pharmacia) equilibrated
with 0.1 M phosphate buffer (pH 7.8). Samples wereeluted with phosphate buffer (0.1 M, pH 7.8) and so-lutions collected for analysis of enzyme activities. APwas measured as described in Wang, et al. (1991), GRby the method of Esterbauer and Grill (1978) and SODwas measured using the method of Giannopolitis andRies (1971) and modified by Bailly et al. (1996).
Sucrose determinationThree 100 mg replicates of leaf tissue and seeds
were ground in liquid nitrogen and samples extractedin cold 100 mM NaOH (50% v/v ethanol/water). Chlo-roform (15% v/v) was added and the samples incu-bated on ice for 10 min. Thereafter the pH was ad-justed to 7.5 with 100 mM HEPES in 100 mM glacialacetic acid. After centrifugation for 20 min at 48C at28,000 g, the supernatant was removed. A repeat ex-traction was performed on the pellets. The superna-tants were pooled and then centrifuged. Sucrose in thesupernatant was calculated from the spectrophotomet-ric measurement of NADPH production using a D-glucose/D-fructose sugar assay kit (Boehringer Mann-heim, Germany).
Measurement of photosynthesisLight-saturated net photosynthesis (A) of leaves of
E. curvula and E. nindensis during desiccation wasmeasured using a Ciras-1 infrared gas analyzer with aParkinson’s Leaf Cuvette and in built illumination unit(PP Systems, Hertfordshire, UK) operated in differ-ential mode at an ambient CO2 concentration of 350ppm and 228C (50% RH). The parameter A was cal-culated according to the equations of von Caemmererand Farquhar (1981) and the data were expressed as apercentage of A measured in control hydrated tissues.Measurements were taken on 5 individual plants andrepeated during at least two cycles of drying.
RESULTS
Clustering expression profiles of Arabidopsis LEAmRNAs during seed development and in response toabiotic stress
LEAs were identified as being up-regulated inleaves in the resurrection plant X. humilis followingdesiccation (Collett et al., 2004). We posed the ques-tion: Are any of the LEA groups restricted to a profileof seed-specific or abiotic-stress specific gene expres-sion in a desiccation-sensitive plant? To this end, wefirst analysed the expression patterns of LEAs from adesiccation-sensitive plant during seed developmentand abiotic stress by mining publicly-available Ara-bidopsis genome and microarray data.
All the AGI gene locus IDs that fall into 10 differentLEA superfamilies, defined on InterProScan, wereidentified (Table 1). Out of 49 Arabidopsis LEAs, 35had specific probes on the 25K Affymetrix gene chipand the expression of these 35 was compared betweensilique and seed development and during abiotic stresstreatments (cold, osmotic, salt and drought stress).
We aimed to distinguish between seed-specific and

775SIGNATURE OF SEEDS IN RESURRECTION PLANTS
stress-specific LEAs on the basis of expression profilesin the selected AtGenExpress dataset. LEA expressionprofiles grouped into different clusters illustrated inFigure 1 and summarized in Table 1. According to thisanalysis, we were able to identify LEA groups that fellonly into Cluster A (i.e., seed specific), but no LEAgroups that were restricted to Cluster D (i.e., abioticstress specific). The Group 1 LEAs (IPR000389) anda Group 9 LEA (PD68804) were identified as beingseed-specific (i.e., cluster A) as there was very lowexpression (,10% of seed expression) in roots orleaves during abiotic stress treatments. Although only3 out of the 6 Group 6 LEAs (Table 1) were repre-sented on the Affymetrix 25K genechip, these werealso only expressed during silique development andfor the latter stages of seed maturation. The maximumabsolute level of expression of the silique-specificAt5g27980 was very low in comparison to the otherexpression profiles reported in this study (,0.5% ofmost abundant LEA, i.e., At5g664400/LEA2, clusterB). Group 7 LEAs (IPR004926), were expressed understress conditions (cluster C and D) or as housekeepinggenes in roots (cluster E). The one LEA from Group8 (IPR004864) that was represented in these experi-ments was expressed at high levels in both maturingseed, and in response to abiotic stress (Table 1, Fig.1). Some members within LEA groups 2, 3, 4, 10 wereseed-specific, whilst others were activated in responseto abiotic stress treatment and seed maturation, or werehousekeeping genes.
What is remarkable is the diversity of LEAs ex-pressed during seed maturation; 74% of the LEAs areexpressed during seed development with 19/35 LEAsbeing expressed at their maximum in seed develop-ment and 7/35 LEAs being expressed at .30% max-imum levels in seeds. In contrast, 46% of the LEAswere expressed in response to abiotic stress, with 10/35 being expressed at maximum level in either leavesor roots and 6/35 LEAs being expressed at .30% lev-el. Expression of homologues of Arabidopsis LEAsfrom Groups 1, 6 and 9 in desiccating vegetative tissueof resurrection plants, could be considered an indicatorthat vegetative DT is due to appropriation of seed DT.This could arise from a change in regulation of genesthat in desiccation-sensitive plants, such as Arabidop-sis, are only expressed during the desiccation phase ofseed maturation.
Sequence analysis of sixteen X. humilis LEAs
To see whether any of the sixteen desiccation up-regulated LEA ESTs identified in the leaves of X. hu-milis (Collett et al., 2004) belonged to the unique seed-specific LEAs we identified in Arabidopsis (i.e.,groups 1, 6 or 9), we sequenced the 59 end of each X.humilis cDNA. The closest homologue for each X. hu-milis putative LEA was identified by a BlastX searchand the nucleotide sequences for each LEA were an-alysed by InterProScan, and grouped into LEA super-families on the basis of their IPR Superfamily classi-fication (Table 2) (Zdobnov and Apweiler, 2001).
When InterProScan failed to identify PFAM domainsin the X. humilis sequence information, LEAs weregrouped on the basis of the InterProScan of their clos-est homologues identified by BlastX analysis (Table2).
With the exception of the Group 1 LEAs(IPR000389) and Group 9 LEAs (PD688044), genesbelonging to all the other LEA groups were identifiedin the 16 LEAs isolated from 400 sequenced X. humiliscDNAs (Table 2). In Arabidopsis, 10 genes are clas-sified in the LEA-2 (IPR000167) and 14 genes areclassified in the LEA-3 (IPR004238) superfamilies(Table 1). These two superfamilies were also well-rep-resented in the X. humilis cDNAs (Table 2). We alsoanalysed LEAs reported as up-regulated during des-iccation in other resurrection plants, namely Crater-ostigma plantigineum, X. viscosa and the moss, Tor-tula ruralis These LEAs can be classified into groups2, 3, 8 and 10 (Table 3). Again, Groups 2 and 3 arepredominant. Only a small number of Arabidopsisgenes are classified in the Group 1 and Group 9 su-perfamilies, and the lack of representation of genes inthese groups from resurrection plants could be due tothe fact that the EST datasets analysed to date do notconstitute the full LEA complement.
The LEA6 cDNA (HC248) was the only LEA in theX. humilis geneset that clearly corresponded to a LEAthat was expressed only in maturing seed in Arabi-dopsis. No conclusions could be drawn on the rolesthat the other X. humilis LEAs play in the evolutionof desiccation tolerance in vegetative tissues, sincethey belong to superfamily groups which include bothseed-specific members as well as members which areexpressed in both seed and abiotic stress in Arabidop-sis (Fig. 1).
Northern blot analysis of X. humilis LEAs
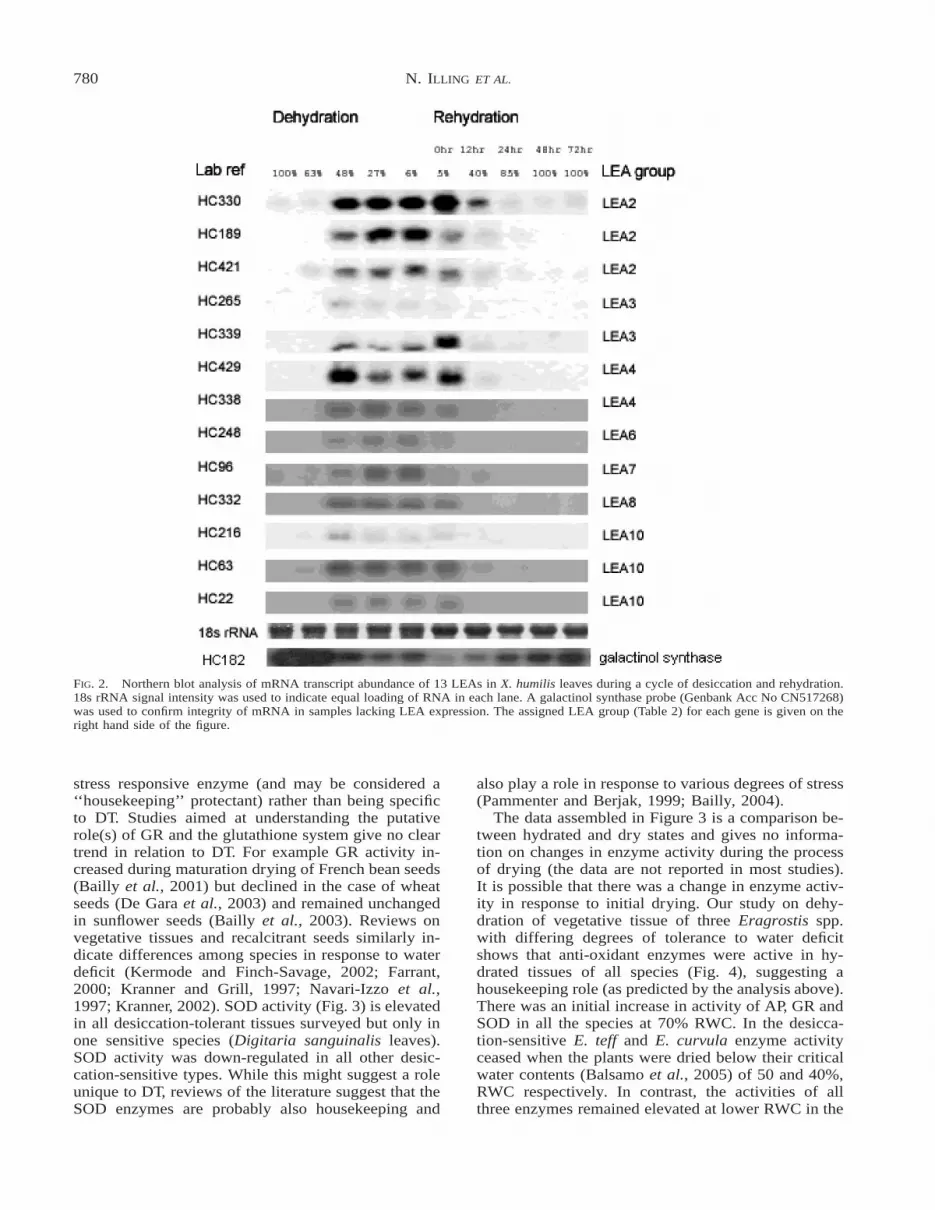
Many of the Group 2 and Group 3 LEAs are up-regulated in response to abiotic stress treatments inArabidopsis (Fig. 1). We tested the hypothesis that X.humilis Group 2 and Group 3 LEAs corresponding tothe stress LEAs from desiccation-sensitive plants,might be up-regulated during the early stages of de-hydration (i.e., .65% RWC), and that a second classof LEAs, which correspond to those expressed duringthe desiccation phase of seed development, are ex-pressed during the later stages of desiccation (,65%RWC). Northern blot analysis of 13 of the 16 LEAs(Fig. 2) clearly showed that the X. humilis LEAs in-vestigated all have similar profiles of expression. Theyare all significantly up-regulated only in the later stag-es of desiccation (i.e., ,65% RWC) in leaves, andmost LEA mRNA transcripts are stably stored in dryleaves (,6% RWC).
Clustering Arabidopsis anti-oxidant mRNAexpression profiles during seed development and inresponse to abiotic stress
We extended our analysis of the selected Arabidop-sis AtGenExpress microarray dataset, posing the ques-

776 N. ILLING ET AL.
FIG. 1. Manual cluster analysis of expression profiles of 35 Arabidopsis LEA genes during silique and seed development and in response toabiotic stress treatments (cold, osmotic, salt and drought) in green shoots (A) and roots (B). 100% represents the highest signal intensity foreach particular gene across these experiments. *1: The maximum intensity signal for these genes was less than 1% of the maximum signal inthe LEA subset of genes. *2: The maximum intensity signal for these genes was less than 2.5% of the maximum signal in the LEA subset ofgenes.

777SIGNATURE OF SEEDS IN RESURRECTION PLANTS
FIG. 1. Continued.
tion: can we identify any anti-oxidant genes that areexpressed exclusively during the desiccation phase ofseed maturation in this desiccation-sensitive plant, andare up-regulated during desiccation in the vegetative
tissue of desiccation tolerant plants? Microarray datafrom the seed development and abiotic stress seriescorresponding to AGI gene locus IDs for 68/71 dif-ferent anti-oxidants listed by Mittler et al. (2004)
778 N. ILLING ET AL.
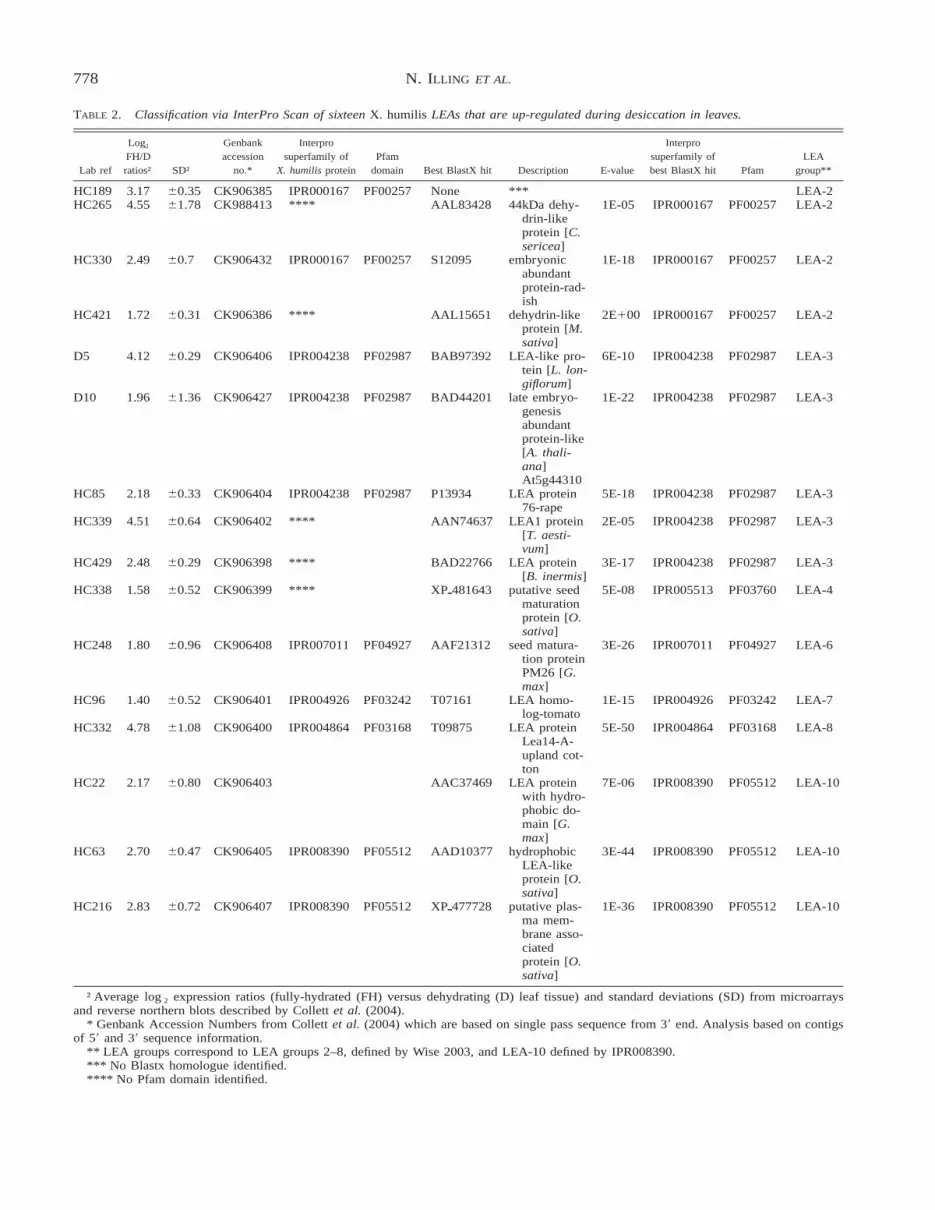
TABLE 2. Classification via InterPro Scan of sixteen X. humilis LEAs that are up-regulated during desiccation in leaves.
Lab ref
Log2
FH/Dratios† SD†
Genbankaccession
no.*
Interprosuperfamily of
X. humilis proteinPfam
domain Best BlastX hit Description E-value
Interprosuperfamily ofbest BlastX hit Pfam
LEAgroup**
HC189 3.17 60.35 CK906385 IPR000167 PF00257 None *** LEA-2HC265 4.55 61.78 CK988413 **** AAL83428 44kDa dehy-
drin-likeprotein [C.sericea]
1E-05 IPR000167 PF00257 LEA-2
HC330 2.49 60.7 CK906432 IPR000167 PF00257 S12095 embryonicabundantprotein-rad-ish
1E-18 IPR000167 PF00257 LEA-2
HC421 1.72 60.31 CK906386 **** AAL15651 dehydrin-likeprotein [M.sativa]
2E100 IPR000167 PF00257 LEA-2
D5 4.12 60.29 CK906406 IPR004238 PF02987 BAB97392 LEA-like pro-tein [L. lon-giflorum]
6E-10 IPR004238 PF02987 LEA-3
D10 1.96 61.36 CK906427 IPR004238 PF02987 BAD44201 late embryo-genesisabundantprotein-like[A. thali-ana]At5g44310
1E-22 IPR004238 PF02987 LEA-3
HC85 2.18 60.33 CK906404 IPR004238 PF02987 P13934 LEA protein76-rape
5E-18 IPR004238 PF02987 LEA-3
HC339 4.51 60.64 CK906402 **** AAN74637 LEA1 protein[T. aesti-vum]
2E-05 IPR004238 PF02987 LEA-3
HC429 2.48 60.29 CK906398 **** BAD22766 LEA protein[B. inermis]
3E-17 IPR004238 PF02987 LEA-3
HC338 1.58 60.52 CK906399 **** XPp481643 putative seedmaturationprotein [O.sativa]
5E-08 IPR005513 PF03760 LEA-4
HC248 1.80 60.96 CK906408 IPR007011 PF04927 AAF21312 seed matura-tion proteinPM26 [G.max]
3E-26 IPR007011 PF04927 LEA-6
HC96 1.40 60.52 CK906401 IPR004926 PF03242 T07161 LEA homo-log-tomato
1E-15 IPR004926 PF03242 LEA-7
HC332 4.78 61.08 CK906400 IPR004864 PF03168 T09875 LEA proteinLea14-A-upland cot-ton
5E-50 IPR004864 PF03168 LEA-8
HC22 2.17 60.80 CK906403 AAC37469 LEA proteinwith hydro-phobic do-main [G.max]
7E-06 IPR008390 PF05512 LEA-10
HC63 2.70 60.47 CK906405 IPR008390 PF05512 AAD10377 hydrophobicLEA-likeprotein [O.sativa]
3E-44 IPR008390 PF05512 LEA-10
HC216 2.83 60.72 CK906407 IPR008390 PF05512 XPp477728 putative plas-ma mem-brane asso-ciatedprotein [O.sativa]
1E-36 IPR008390 PF05512 LEA-10
† Average log2 expression ratios (fully-hydrated (FH) versus dehydrating (D) leaf tissue) and standard deviations (SD) from microarraysand reverse northern blots described by Collett et al. (2004).
* Genbank Accession Numbers from Collett et al. (2004) which are based on single pass sequence from 39 end. Analysis based on contigsof 59 and 39 sequence information.
** LEA groups correspond to LEA groups 2–8, defined by Wise 2003, and LEA-10 defined by IPR008390.*** No Blastx homologue identified.**** No Pfam domain identified.
779SIGNATURE OF SEEDS IN RESURRECTION PLANTS
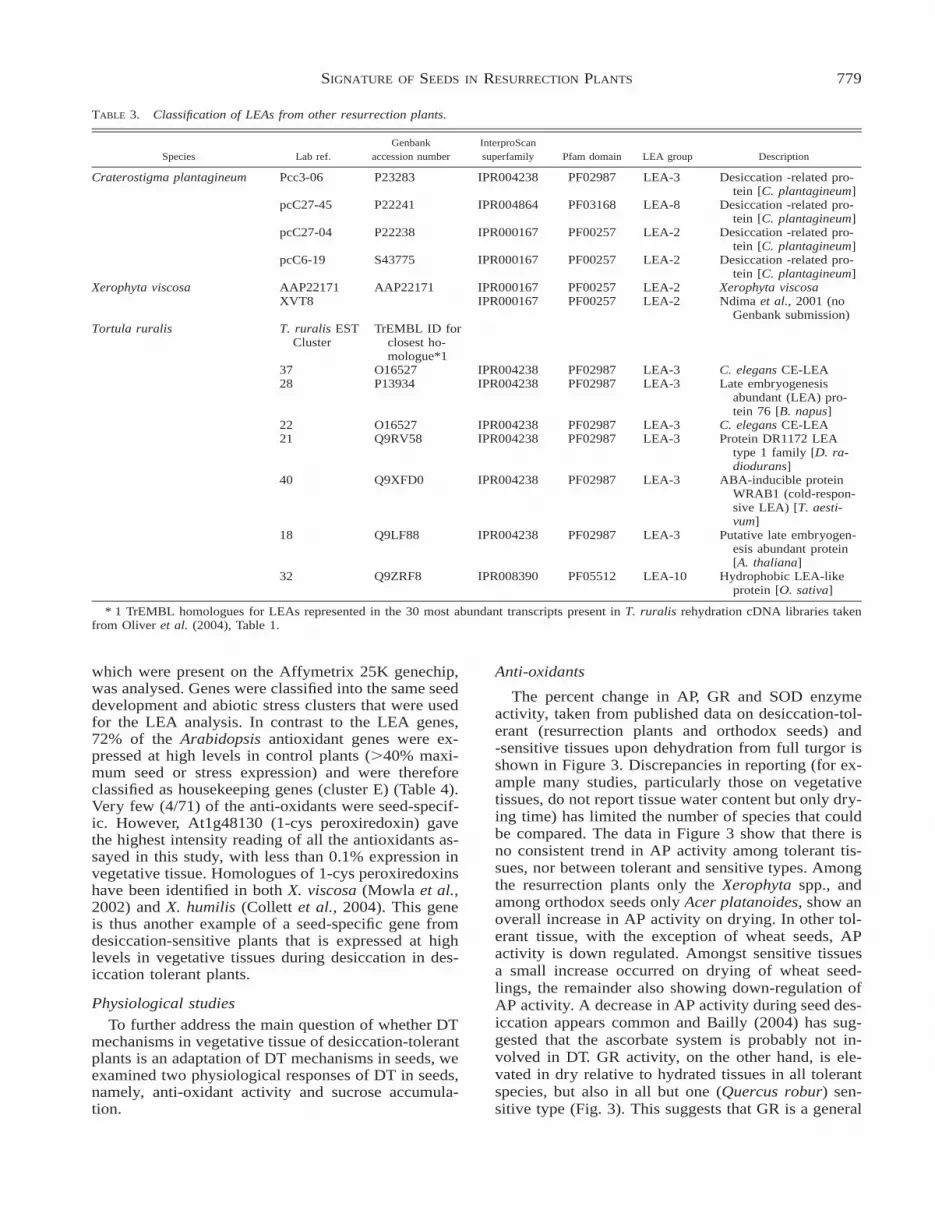
TABLE 3. Classification of LEAs from other resurrection plants.
Species Lab ref.Genbank
accession numberInterproScansuperfamily Pfam domain LEA group Description
Craterostigma plantagineum Pcc3-06 P23283 IPR004238 PF02987 LEA-3 Desiccation -related pro-tein [C. plantagineum]
pcC27-45 P22241 IPR004864 PF03168 LEA-8 Desiccation -related pro-tein [C. plantagineum]
pcC27-04 P22238 IPR000167 PF00257 LEA-2 Desiccation -related pro-tein [C. plantagineum]
pcC6-19 S43775 IPR000167 PF00257 LEA-2 Desiccation -related pro-tein [C. plantagineum]
Xerophyta viscosa AAP22171 AAP22171 IPR000167 PF00257 LEA-2 Xerophyta viscosaXVT8 IPR000167 PF00257 LEA-2 Ndima et al., 2001 (no
Genbank submission)Tortula ruralis T. ruralis EST
ClusterTrEMBL ID for
closest ho-mologue*1
37 O16527 IPR004238 PF02987 LEA-3 C. elegans CE-LEA28 P13934 IPR004238 PF02987 LEA-3 Late embryogenesis
abundant (LEA) pro-tein 76 [B. napus]
22 O16527 IPR004238 PF02987 LEA-3 C. elegans CE-LEA21 Q9RV58 IPR004238 PF02987 LEA-3 Protein DR1172 LEA
type 1 family [D. ra-diodurans]
40 Q9XFD0 IPR004238 PF02987 LEA-3 ABA-inducible proteinWRAB1 (cold-respon-sive LEA) [T. aesti-vum]
18 Q9LF88 IPR004238 PF02987 LEA-3 Putative late embryogen-esis abundant protein[A. thaliana]
32 Q9ZRF8 IPR008390 PF05512 LEA-10 Hydrophobic LEA-likeprotein [O. sativa]
* 1 TrEMBL homologues for LEAs represented in the 30 most abundant transcripts present in T. ruralis rehydration cDNA libraries takenfrom Oliver et al. (2004), Table 1.
which were present on the Affymetrix 25K genechip,was analysed. Genes were classified into the same seeddevelopment and abiotic stress clusters that were usedfor the LEA analysis. In contrast to the LEA genes,72% of the Arabidopsis antioxidant genes were ex-pressed at high levels in control plants (.40% maxi-mum seed or stress expression) and were thereforeclassified as housekeeping genes (cluster E) (Table 4).Very few (4/71) of the anti-oxidants were seed-specif-ic. However, At1g48130 (1-cys peroxiredoxin) gavethe highest intensity reading of all the antioxidants as-sayed in this study, with less than 0.1% expression invegetative tissue. Homologues of 1-cys peroxiredoxinshave been identified in both X. viscosa (Mowla et al.,2002) and X. humilis (Collett et al., 2004). This geneis thus another example of a seed-specific gene fromdesiccation-sensitive plants that is expressed at highlevels in vegetative tissues during desiccation in des-iccation tolerant plants.
Physiological studies
To further address the main question of whether DTmechanisms in vegetative tissue of desiccation-tolerantplants is an adaptation of DT mechanisms in seeds, weexamined two physiological responses of DT in seeds,namely, anti-oxidant activity and sucrose accumula-tion.
Anti-oxidants
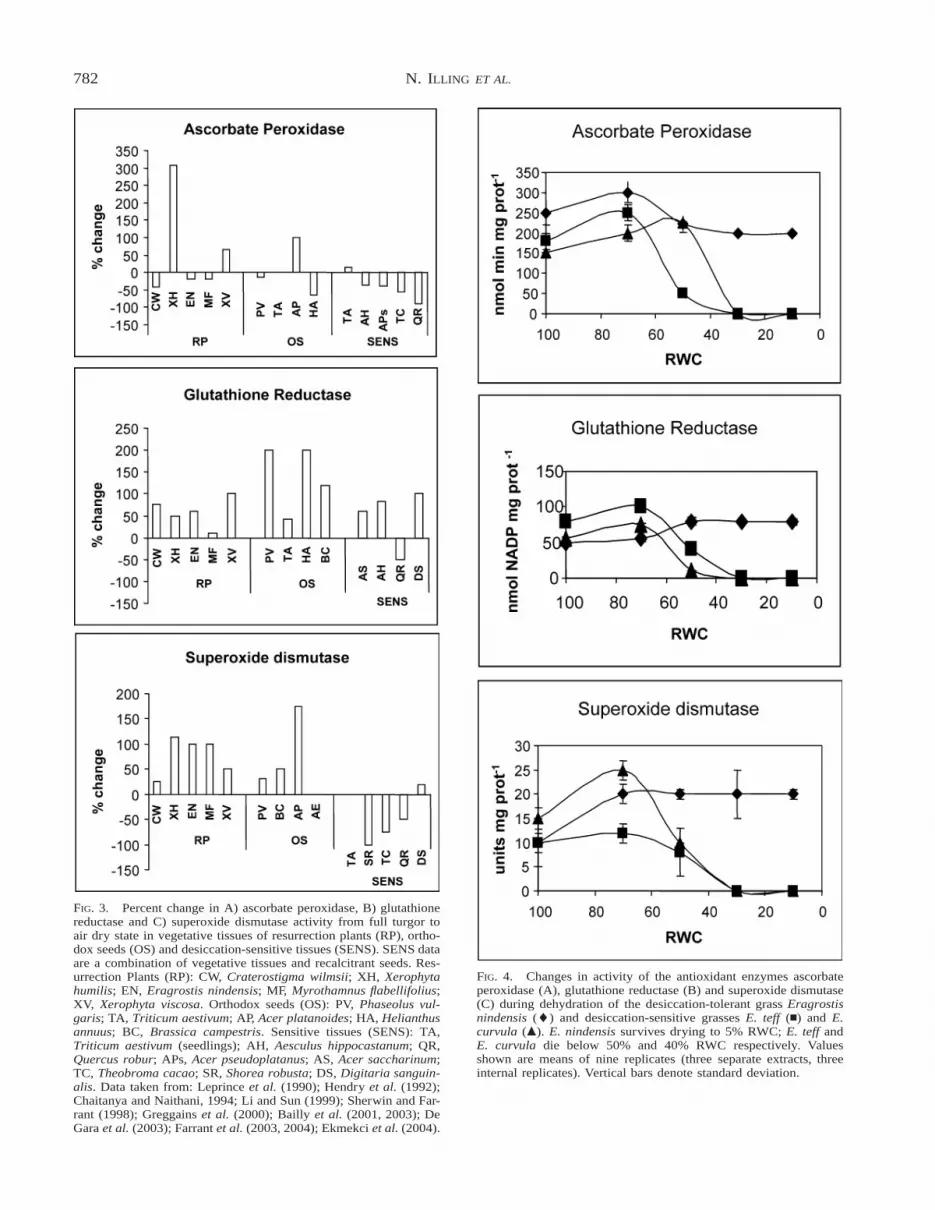
The percent change in AP, GR and SOD enzymeactivity, taken from published data on desiccation-tol-erant (resurrection plants and orthodox seeds) and-sensitive tissues upon dehydration from full turgor isshown in Figure 3. Discrepancies in reporting (for ex-ample many studies, particularly those on vegetativetissues, do not report tissue water content but only dry-ing time) has limited the number of species that couldbe compared. The data in Figure 3 show that there isno consistent trend in AP activity among tolerant tis-sues, nor between tolerant and sensitive types. Amongthe resurrection plants only the Xerophyta spp., andamong orthodox seeds only Acer platanoides, show anoverall increase in AP activity on drying. In other tol-erant tissue, with the exception of wheat seeds, APactivity is down regulated. Amongst sensitive tissuesa small increase occurred on drying of wheat seed-lings, the remainder also showing down-regulation ofAP activity. A decrease in AP activity during seed des-iccation appears common and Bailly (2004) has sug-gested that the ascorbate system is probably not in-volved in DT. GR activity, on the other hand, is ele-vated in dry relative to hydrated tissues in all tolerantspecies, but also in all but one (Quercus robur) sen-sitive type (Fig. 3). This suggests that GR is a general
780 N. ILLING ET AL.
FIG. 2. Northern blot analysis of mRNA transcript abundance of 13 LEAs in X. humilis leaves during a cycle of desiccation and rehydration.18s rRNA signal intensity was used to indicate equal loading of RNA in each lane. A galactinol synthase probe (Genbank Acc No CN517268)was used to confirm integrity of mRNA in samples lacking LEA expression. The assigned LEA group (Table 2) for each gene is given on theright hand side of the figure.
stress responsive enzyme (and may be considered a‘‘housekeeping’’ protectant) rather than being specificto DT. Studies aimed at understanding the putativerole(s) of GR and the glutathione system give no cleartrend in relation to DT. For example GR activity in-creased during maturation drying of French bean seeds(Bailly et al., 2001) but declined in the case of wheatseeds (De Gara et al., 2003) and remained unchangedin sunflower seeds (Bailly et al., 2003). Reviews onvegetative tissues and recalcitrant seeds similarly in-dicate differences among species in response to waterdeficit (Kermode and Finch-Savage, 2002; Farrant,2000; Kranner and Grill, 1997; Navari-Izzo et al.,1997; Kranner, 2002). SOD activity (Fig. 3) is elevatedin all desiccation-tolerant tissues surveyed but only inone sensitive species (Digitaria sanguinalis leaves).SOD activity was down-regulated in all other desic-cation-sensitive types. While this might suggest a roleunique to DT, reviews of the literature suggest that theSOD enzymes are probably also housekeeping and
also play a role in response to various degrees of stress(Pammenter and Berjak, 1999; Bailly, 2004).
The data assembled in Figure 3 is a comparison be-tween hydrated and dry states and gives no informa-tion on changes in enzyme activity during the processof drying (the data are not reported in most studies).It is possible that there was a change in enzyme activ-ity in response to initial drying. Our study on dehy-dration of vegetative tissue of three Eragrostis spp.with differing degrees of tolerance to water deficitshows that anti-oxidant enzymes were active in hy-drated tissues of all species (Fig. 4), suggesting ahousekeeping role (as predicted by the analysis above).There was an initial increase in activity of AP, GR andSOD in all the species at 70% RWC. In the desicca-tion-sensitive E. teff and E. curvula enzyme activityceased when the plants were dried below their criticalwater contents (Balsamo et al., 2005) of 50 and 40%,RWC respectively. In contrast, the activities of allthree enzymes remained elevated at lower RWC in the

781SIGNATURE OF SEEDS IN RESURRECTION PLANTS
TABLE 4. Analysis of expression of anti-oxidant genes during seed development and in response to abiotic stress in A. thaliana.
Cluster A Cluster EMax seed, ,30% stress Control .50% seed or stress
At1g20630 Catalase 1 (Cat1) At1g07890 Ascorbate peroxidase (APX1)At1g48130 1-Cys peroxiredoxin (1-cys-PrxR) At1g08830 Cu/Zn superoxide dismutase (CSD1)At2g28190 Cu/Zn superoxide dismutase (CSD2) At1g09090 NADPH oxidase (RbohB)At3g11050 Ferritin 4* At1g19230 NADPH oxidase (RbohE)At3g56350 Mn superoxide dismutase like At1g20620 Catalase 3 (Cat3)
At1g33660 Ascorbate peroxidase (APX7)Cluster B At1g60740 Peroxiredoxin Type 2 (Type 2 PrxR D)Max seed, .30% stress At1g63460 Glutathione peroxidase 8 (GPX8)
At1g65990 Peroxiredoxin Type 2 (Type 2 PrxR A) At1g63940 Monohydroascorbate reductase (MDAR1)At4g11600 Phospholipid glutathione peroxidase (GPX6) At1g64060 NADPH oxidase (RbohF)At5g64210 Alternative oxidase (AOX2) At1g65980 Peroxiredoxin Type 2 (Type 2 prxR B)
At1g75270 Dehydroascorbate reductase (DHAR3)Cluster C At1g77490 Thylakoid ascorbate peroxidase (Thylakoid-APX)Max stress, .30% seed At2g25080 Glutathione peroxidase 1 (GPX1)
At1g19570 Dehydroascorbate reductase 5 (DHAR5) At2g40300 Ferritin 3At2g31570 Glutathione peroxidase 2 (GPX2) At2g43350 Glutathione peroxidase 3 (GPX3)At3g24170 Glutathione reductase 1 (GR1) At2g48150 Glutathione peroxidase (GPX4)At3g56090 Ferritin 2 At3g06050 Peroxiredoxin 2-cys (2-cys-PrxR F)At5g01600 Ferritin 1 At3g10920 Mn superoxide dismutase (MSD1)
At3g11630 Peroxiredoxin 2-cys (2-cys-PrxR A)Cluster D At3g26060 Peroxiredoxin (PrxR O)Max stress, ,30% seed At3g27620 Alternative oxidase (AOX1C)
At1g32350 Putative alternative oxidase (AOX putative) At3g27820 monodehydroascorbate reductase (MDAR3)At3g09640 Ascorbate peroxidase 2 (APX2) At3g45810 NADPH oxidase J (RbohJ)At3g09940 Monohydroascorbate reductase 2 (MDAR2) At3g52880 Monohydroascorbate reductase (MDAR4)At3g22360 Alternative oxidase 1B (AOX1B) At3g52960 Peroxiredoxin Type 2 (Type 2 PrxR E)At3g22370 Alternative oxidase 1A (AOX1A) At3g54660 Glutathione reductase (GR2)At5g47910 NADPH oxidase (RbohD) At3g63080 Glutathione peroxidase 5 (GPX5)
At4g08390 Stromal ascorbate peroxidase (Stromal-APX)At4g09010 Ascorbate peroxidase (APX4)At4g11230 NADPH oxidase (RbohH0)At4g22260 Alternative oxidase (AOX1B)At4g25090 NADPH oxidase (RbohG)At4g25100 Fe superoxide dismutase (FSD1)At4g31870 Glutathione peroxidase 7 (GPX7)At4g32320 Ascorbate peroxidase (APX6)At4g35000 Ascorbate peroxidase (APX3)At4g35090 Catalase 2 (Cat2)At4g35970 Ascorbate peroxidase 5 (APX5)At5g03630 Monodehydroascorbate reductase 5 (MDAR5)At5g06290 Peroxiredoxin 2-cys (PrxR B)At5g07390 NADPH oxidase (RbohA)At5g16710 Dehydroascorbate reductase (DHAR1)At5g18100 Cu/Zn superoxide dismutase (CSD3)At5g23310 Fe superoxide dismutase (FSD3)At5g36270 Dehydroascorbate reductase (DHAR2)At5g51060 NADPH oxidase (RbohC)At5g51100 Fe superoxide dismutase (FSD3)At5g60010 NADPH oxidase (RbohH)
* Silique specific.
DT E. nindensis compared to the desiccation-sensitivespecies. Mature air-dry orthodox seeds of both E. nin-densis and E. teff retained significant anti-oxidant en-zyme activity (Fig. 5). Since the enzyme assays aredone in vitro this reflects only their potential for ac-tivity and it is unlikely that they are active in situ at5% RWC. Nonetheless, the ability to remain activewhen extracted from dry tissue is evidence that theenzymes had been sufficiently protected in the drystate. The lack of enzyme activity in dry tissues of E.teff and E. curvula is likely to be due to their desic-cation-induced denaturation (rather than down-regu-lation) since the plants did not survive drying below50% RWC (data not shown). Analysis of anti-oxidant
enzyme activity therefore corroborates the conclusionsfrom the Arabidopsis antioxidant expression profiles,in that anti-oxidant activity does not appear to be amechanism specific to DT.
Sucrose
Our survey of changes in sucrose content in re-sponse to drying in tissues of desiccation-tolerant and-sensitive species (Fig. 6) shows that this sugar doesindeed increase, to varying extents, on drying inangiosperm resurrection plants and orthodox seeds.There was also a general increased level in responseto drying in those sensitive tissues from which wecould obtain drying course data. However, all but one
782 N. ILLING ET AL.
FIG. 4. Changes in activity of the antioxidant enzymes ascorbateperoxidase (A), glutathione reductase (B) and superoxide dismutase(C) during dehydration of the desiccation-tolerant grass Eragrostisnindensis (l) and desiccation-sensitive grasses E. teff (n) and E.curvula (m). E. nindensis survives drying to 5% RWC; E. teff andE. curvula die below 50% and 40% RWC respectively. Valuesshown are means of nine replicates (three separate extracts, threeinternal replicates). Vertical bars denote standard deviation.
FIG. 3. Percent change in A) ascorbate peroxidase, B) glutathionereductase and C) superoxide dismutase activity from full turgor toair dry state in vegetative tissues of resurrection plants (RP), ortho-dox seeds (OS) and desiccation-sensitive tissues (SENS). SENS dataare a combination of vegetative tissues and recalcitrant seeds. Res-urrection Plants (RP): CW, Craterostigma wilmsii; XH, Xerophytahumilis; EN, Eragrostis nindensis; MF, Myrothamnus flabellifolius;XV, Xerophyta viscosa. Orthodox seeds (OS): PV, Phaseolus vul-garis; TA, Triticum aestivum; AP, Acer platanoides; HA, Helianthusannuus; BC, Brassica campestris. Sensitive tissues (SENS): TA,Triticum aestivum (seedlings); AH, Aesculus hippocastanum; QR,Quercus robur; APs, Acer pseudoplatanus; AS, Acer saccharinum;TC, Theobroma cacao; SR, Shorea robusta; DS, Digitaria sanguin-alis. Data taken from: Leprince et al. (1990); Hendry et al. (1992);Chaitanya and Naithani, 1994; Li and Sun (1999); Sherwin and Far-rant (1998); Greggains et al. (2000); Bailly et al. (2001, 2003); DeGara et al. (2003); Farrant et al. (2003, 2004); Ekmekci et al. (2004).

783SIGNATURE OF SEEDS IN RESURRECTION PLANTS
FIG. 5. Antioxidant enzyme activity in mature dry seeds of E. nin-densis (white bars) and E. teff (black bars). AP, ascorbate peroxidase(units, nmol·min21·mg protein21); GR, glutathione reductase (units,nmol NADP.mg protein21); SOD, superoxide dismutase (units,Units.mg protein21). Values shown are the means of nine replicates(three separate extracts, three internal replicates). Vertical bars de-note standard deviation.
FIG. 6. Percent change in sucrose content from full turgor to airdry state in vegetative tissues of resurrection plants (RP), orthodoxseeds (OS) and desiccation-sensitive tissues (SENS). SENS data area combination of vegetative tissues and recalcitrant seeds. Resurrec-tion Plants (RP): CW, Craterostigma wilmsii; XH, Xerophyta hu-milis; EN, Eragrostis nindensis; MF, Myrothamnus flabellifolius;BC, Borya constricta; CS, Coleochloa setifera; MK, Microchloakunthii; XV, Xerophyta viscosa; SS, Sporobolus stapfianus. Ortho-dox seeds (OS): PS1, PS2 & PS3, Pisum sativum, genotypes SD1,SD5 and SD7 respectively from Karner et al. (2004); HV, Hordeumvulgaris; PV, Phaseolus vulgaris; TA, Triticum aestivum; BC, Bras-sica campestris; AP, Acer platanoides. Sensitive tissues (SENS): TA,Triticum aestivum (seedlings); QR, Quercus robur; AM, Avicenniamarina; APs, Acer pseudoplatanus; AS, Acer saccharinum; AA, Ar-aucaria angustifolia; CS, Camellia sinensis. Data taken from: Le-prince et al. (1990); Farrant et al. (1992); Finch-Savage and Blake(1994); Steadman et al. (1996); Farrant and Walters, (1997); Ghas-empour et al. (1998); Li and Sun, (1999); Farrant et al. (2003, 2004);Karner et al. (2004); Vander Willigen et al. (2001); Whittaker et al.(2004).
FIG. 7. Changes in photosynthetic assimilation rate (A) as a per-centage of that in control hydrated tissue (lefthand Y axis, closedsymbols) and sucrose content (mmol. mg dw21) (righthand Y axis,open symbols) during dehydration of the desiccation-tolerant grassEragrostis nindensis (m) and desiccation-sensitive grasses E. cur-vula (l) and E. teff (M) (sucrose only). E. nindensis survives dryingto 5% RWC; E. teff and E. curvula die below 50% and 40% RWCrespectively. The values shown are means of three separate extrac-tions. Vertical bars denote standard deviation.
(wheat seedling) of the reports on sensitive tissues dealwith recalcitrant seeds (not desiccation-tolerant) andthe accumulation of sucrose may well be as storagereserve rather than pertaining to water deficit duringmaturation. At least one species (Avicennia marina)does not dry during the terminal stages of developmentand yet considerable levels of sucrose accumulate(Farrant et al., 1992). The work on wheat coleoptilesindicated that these tissues were newly germinated andstill relatively plastic with respect to re-induction ofDT (Farrant et al., 2004) and thus sucrose accumula-tion in that system might well be related to DT.
Our study on Eragrostis spp. provides additional ev-idence that sucrose accumulation is a mechanismlinked to DT. In vegetative tissues, sucrose accumu-lates only in the desiccation-tolerant species E. nin-densis in response to drying (Fig. 7). In contrast, wefind that the mature orthodox seeds of both E. ninden-sis and E. teff accumulate high sucrose levels of 88(612) and 145 (616) mmol·mg dw21 respectively.
The accumulation of sucrose in E. nindensis (Fig.7) is not likely to be due to photosynthesis, as this isshut down early in the drying time course before max-imal sucrose accrual. This is true too of other resur-rection plants studied to date (Farrant, 2000; Mundreeet al., 2002; Whittaker et al., 2001, 2004) and sucroseaccumulation is proposed to be from mobilization ofother storage oligo- and/or polysaccharides such as oc-tulose, stachyose and starch (Schwall et al., 1995; Nor-wood et al., 2000, 2003). In seeds, sucrose is importedfrom the parent plant. Although photosynthesis contin-ues to lower water contents in E. curvula, sucrose isnot accumulated in this species (Fig. 7) and this spe-cies apparently does not have the ability to up-regulatealternative means of sucrose accumulation upon waterdeficit stress. Sucrose accumulation is therefore com-mon to both seed development and DT in vegetative

784 N. ILLING ET AL.
tissues, and we suggest that sucrose accumulation forprotection against desiccation damage is not associatedwith photosynthesis but involves up-regulation of al-ternative pathways, with signals that might be commonto those present during orthodox seed development.
DISCUSSION
This study offers an approach to exploring the ori-gins of DT in angisosperms and resurrection plants.Two possibilities were entertained which are not nec-essarily mutually exclusive: DT in resurrection plantswas acquired via DT of seeds and/or via effective ad-aptation of abiotic stress responses. We have looked atboth molecular and physiological/biochemical re-sponses to desiccation and abiotic stress to ask whetherresponses commonly associated with DT in vegetativetissues are also active in desiccating seeds and/or dur-ing abiotic stress.
LEAs
First we systematically compared the expressionlevels of 35 LEAs represented on an Arabidopsis Af-fymetrix 25K gene chip during seed development, andunder a standardized set of conditions for abiotic stresstreatments. A large diversity of LEAs were expressedduring seed development in comparison to the re-sponse of vegetative tissue to abiotic stress. Whereasthere is no particular class of LEAs that is uniquelyexpressed during abiotic stress, genes belonging toLEA-1, -6 and -9 superfamilies are only significantlyexpressed during seed development. These LEAs maythus be uniquely associated with defense against se-vere water loss such as would occur in desiccation-tolerant angiosperms/resurrection plants or orthodoxseeds.
Many of the abiotic stress-responsive LEAs identi-fied in the expression dataset have also been identifiedin other Arabidopsis microarray studies on abioticstress. These include At1g01470 (LEA-8), At1g20440(LEA-2), At1g52690 (LEA-3), At5g06760 (LEA-4)and At5g66400 (LEA-2) (Bray, 2004). Our compari-son with expression data for seed development showsthat only At1g20440 (LEA-2) is specific to the abioticstress response. Notably, no LEA-1, -6, -7, -9 or -10genes were identified as stress up-regulated in thesestudies.
We have shown that the expression of at least 16different LEA genes, representing the LEA-2, -3 -4,-6 -7, -8 and -10 superfamilies, is activated during des-iccation in X. humilis leaves (Collett et al., 2004).LEA-6 was identified as a ‘seed-specific’ group in Ar-abidopsis. Northern blot analysis has shown that theLEAs investigated are specifically activated during thelate stages of desiccation, and not during the earlystages of water loss, suggesting that they are a unique-ly desiccation-specific set of LEAs. We speculate thatthe simultaneous activation of such a large comple-ment of LEAs under conditions of water loss couldpoint towards the formation of an interacting networknecessary for the stabilization of membranes and the
protection of proteins. An alternative explanationcould be that different LEAs are specifically targetedto different organelles or cellular structures, wherethey play a local role in protecting proteins, nucleicacids and membranes from the effects of water loss.LEAs have been reported to be expressed at high lev-els in several other desiccation tolerant plants includ-ing the bryophyte T. ruralis, and the angiosperms C.plantagineum and X. viscosa (Table 3). These LEAsall represent the LEA-2, LEA-3 LEA-8 and LEA-10superfamilies. Six different LEA-3 ESTs wereamongst the most abundantly represented mRNA tran-scripts in a T. ruralis rehydration cDNA library (Oli-ver et al., 2004) and a LEA-3 gene has been shownto expressed in the anhydrobiotic nematode Aphelen-chus avenae (Browne et al., 2002). The LEA-3 super-family may thus represent very ancient proteins thatplay important roles during desiccation.
The nomenclature used in this study to describe theLEA superfamilies corresponds to the convention de-scribed by Wise (2003), based on differences in pep-tide composition (see Wise and Tunnacliffe, 2004). Wehave included a LEA-10 group to accommodate anunclassified LEA-like protein in the Pfam database. Itremains to be seen whether different functions can beassigned to the different LEA superfamilies. We usedthe terms ‘LEA’ and ‘LEA-like’ to identify superfam-ilies which represent this large class of proteins in thePfam database. It is important to bear in mind that theannotation of sequences as LEAs tends to be arbitrary,originally assigned according to the abundance of atranscript during late embryogenesis. A unifying fea-ture of this group of proteins is their large number ofhomologous repetitive hydrophilic peptide motifs andtheir high percentage of glycine residues. Recent re-ports on functional analysis of recombinant proteins ofAvLEA1, a Group 3 LEA protein from the anhydro-biotic nematode A. avenae, and Em, a group I LEAprotein from wheat (Goyal et al., 2005) have shownthat both these LEAs show anti-aggregation propertiesand protected enzyme activity under conditions of wa-ter loss. These properties were synergistically en-hanced in the presence of sucrose.
Antioxidants
In contrast to the LEAs, mRNA transcripts for mostof the anti-oxidant enzymes were abundant in Arabi-dopsis under control conditions. Very few anti-oxi-dants were seed-specific but notably, one of these, a1-cys-peroxiredoxin, has been previously shown to beabundantly expressed during desiccation in the mossT. ruralis and in the leaves of X. humilis and X. vis-cosa.
Overall, the physiological data on antioxidant en-zymes such as AP, GR and SOD suggest that these are‘housekeeping’ protectants, responsive in the case ofmost abiotic stresses. We propose that they while theyare part of the protection systems in desiccation-tol-erant tissues, they are not unique to them and thus arenot useful in the evaluation of evolution of DT. Only

785SIGNATURE OF SEEDS IN RESURRECTION PLANTS
in true desiccation-tolerant tissues can the activity re-main elevated, but this is likely to be a consequenceof mechanisms that protect the anti-oxidant enzymes,rather than a unique DT mechanism.
However, there are some antioxidants that appear tobe novel to DT, expressed only in maturation dryingof orthodox seeds and desiccation of resurrectionplants. For example, a 1-Cys peroxiredoxin has beenreported to be seed specific (Aalen, 1999; Haslekas etal., 1998) but is induced on drying of the resurrectionplant X. viscosa (Mowla et al., 2002). Interestingly, a1-Cys peroxiredoxin is also expressed during rehydra-tion of the desiccation-tolerant moss T. ruralis (Oliver,1996) and thus might be indicative of the evolutionaryprocess of these antioxidants in DT.
Sucrose
Sucrose appears to be universally present in matureorthodox seeds (Chen and Burris, 1990; Vertucci andFarrant, 1995) and increases in response to desiccationin all angiosperm resurrection plants studied to date(Ghasempour et al., 1998; Scott, 2000; Whittaker etal., 2001, 2004). Although the precise role of sucrosein DT is still unclear, it is generally agreed that theaccumulation of this metabolite is important for pro-tection against desiccation damage (Koster and Leo-pold, 1988; Scott, 2000; Walters et al., 2002). Our datashow that sucrose accumulates in vegetative tissues ofonly the desiccation-tolerant species of Eragrostis and,like other resurrection plants studied to date (Farrant,2000; Mundree et al., 2002; Whittaker et al., 2001,2004), appears unrelated to photosynthesis. Thus whilesucrose itself is a common metabolite, its method ofaccumulation under stress conditions might be an im-portant factor in DT. Breakdown of oligo- and poly-saccharides and their mobilization and resynthesis intosucrose within tissues that become protected againstdesiccation damage is a common phenomenon in seedsand resurrection plants. It is not known whether themechanisms and signals for this accumulation are sim-ilar in seeds and resurrection plants but it is clear thatthey are absent from vegetative tissues of desiccation-sensitive species E. teff and E. curvula.
IN SUMMARY
Some of the LEAs and the majority of anti-oxidantsanalysed in the present study appear to be ‘‘house-keeping’’ or are activated under conditions of abioticstress (cold, osmotic, salt or drought) in desiccation-sensitive and -tolerant tissues. We propose that theseare active and protect only at higher water contents(.65% RWC), but that for true desiccation-tolerancea further repertoire of protectants are necessary andthat these are activated below 65% RWC in desicca-tion-tolerant tissues. The activation of a number of‘seed-specific’ desiccation protection mechanisms,such as sucrose accumulation and expression of aLEA-6 and a 1-cys-peroxiredoxin gene, in the vege-tative tissues of desiccation-tolerant plants points to-wards acquisition of DT from seeds.
ACKNOWLEDGMENTS
We thank Cathal Seoighe and Brigitte Hamman forassistance in compiling the data for the study andBronwen Aken and Zek Ginbot for some of the anti-oxidant and sucrose data generated in the Eragrostisstudy. This research was supported by funding fromthe University of Cape Town, the National ResearchFoundation, South Africa and the National Bioinfor-matics Network, South Africa.
REFERENCES
Aalen, R. B. 1999. Peroxiredoxin antioxidants in seed physiology.Seed Sci. Res. 9:285–295.
Alpert, P. and M. J. Oliver. 2002. Drying without dying. In M. Blackand H. W. Pritchard (eds.), Desiccation and survival in plants:Drying without dying, pp. 3–43. CABI publishing, Wallingfordand New York.
Bailly, C. 2004. Active oxygen species and antioxidants in seedbiology. Seed Sci. Res. 14:93–107.
Bailly, C., A. Audigier, F. Ladonne, M. H. Wanger, F. Ocste, F. Con-bineau, and D. Come. 2001. Changes in oligosaccharide contentand antioxidant enzyme activities in developing bean seeds asrelated to acquisition of drying tolerance and seed quality. J.Exp. Bot. 52:701–708.
Bailly, C., A. Benamar, F. Corbineau, and D. Come. 1996. Changesin malondialdehyde content and in superoxide dismutase, cat-alase and glutathione reductase activities in sunflower seeds asrelated to deterioration during accelerated aging. Physiol. Plant.97:104–110.
Bailly, C., J. Leymarie, S. Rousseau, D. Come, A. Feutry, and F.Corbineau. 2003. Sunflower seed development as related to an-tioxidant enzyme activities. In G. Nicolas, K. J. Bradford, D.Come, and H. W. Pritchard (eds.), The biology of seeds: Recentresearch advances, pp. 69–75. CABI publishing, Wallingfordand New York.
Balsamo, R., C. Vander Willigen, and J. M. Farrant. 2005. Relatingleaf tensile properties to drought tolerance for selected speciesof Eragrostis. Ann. Bot. (In press)
Bewley, J. D. and M. J. Oliver. 1992. Desiccation-tolerance in veg-etative plant tissues and seeds: protein synthesis in relation todesiccation and a potential role for protection and repair mech-anisms. In C. B. Osmond and G. Somero (eds.), Water and life:A comparative analysis of water relationships at the organis-mic, cellular and molecular levels, pp. 141–160. Springer-Ver-lag, Berlin.
Bray, E. A. 1991. Molecular responses to water deficit. Plant Phy-siol. 103:1035–1040.
Bray, E. A. 1997. Plant responses to water deficit. Trends Plant Sci.2:48–54.
Browne, J., A. Tunnacliffe, and A. Burnell. 2002. Plant desiccationgene found in nematode. Nature 416:38.
Chaitanya, K. S. and S. C. Naithani. 1994. Role of superoxide, lipidperoxidation and superoxide dismutase in membrane perturba-tion during loss of viability in seeds of Shorea robusta Gaertnf.New Phytol. 126:623–627.
Chen, Y. and J. S. Burris. 1990. Role of carbohydrates in desiccationtolerance and membrane behaviour in maturing maize seed.Crop Sci. 30:971–975.
Close, T. J. 1997. Dehydrins: A commonality in the response ofplants to dehydration and low temperature. Physiol. Plant. 100:291–296.
Collett, H., A. Shen, M. Gardner, J. M. Farrant, K. J. Denby, andN. Illing. 2004. Towards transcript profiling of desiccation tol-erance in Xerophyta humilis: Construction of a normalized 11kX. humilis cDNA set and microarray expression analysis of 424cDNAs in response to dehyration. Physiol. Plant. 122:39–53.
Crowe, J. H., J. F. Carpenter, and L. M. Crowe. 1998. The role ofvitrification in anhydrobiosis. Ann. Rev. Physiol. 60:73–103.
Dace, H., H. W. Sherwin, N. Illing, and J. M. Farrant. 1998. Use ofmetabolic inhibitors to elucidate mechanisms of recovery from

786 N. ILLING ET AL.
desiccation stress in the resurrection plant Xerophyta humilis.Plant Growth Reg. 24:171–177.
De Gara, L., M. C. de Pinto, V. M. C. Moliterni, and M. G.D’Egidio. 2003. Redox regulation and storage processes duringmaturation in kernels of Triticum aestivum. J. Exp. Bot. 54:249–258.
Dickie, J. B and H. W. Pritchard. 2002. Systematic and evolutionaryaspects of desiccation tolerance in seeds. In M. Black and H.W. Pritchard (eds.), Desiccation and survival in plants: Dryingwithout dying, pp. 239–260. CABI publishing, Wallingford andNew York.
Ekmekci, Y., J. M. Farrant, J. A. Thomson, and S. M. Mundree.2004. Antioxidant response and photosynthetic characteristicsof Xerophyta viscosa Baker and Digitaria sanguinalis L. leavesinduced by high light. Israel J. Plant Sci. 52:177–187.
Esterbauer, H. and D. Grill. 1978. Seasonal variation of glutathioneand glutathione reductase in needles of Picea abies. Plant Phy-siol. 61:119–121.
Farrant, J. M., 2000. A comparison of mechanisms of desiccation-tolerance among three angiosperm resurrection plant species.Plant Ecol. 151:29–39.
Farrant, J. M., C. Bailly, J. Leymarie, B. Hamman, D. Come, andF. Corbineau. 2004. Wheat seedlings as a model to understanddesiccation-tolerance and—sensitivity. Physiol. Plant. 120:563–574.
Farrant, J. M., S. Bartsch, D. Loffell, C. Vander Willigen, and A.Whittaker. 2003. An investigation into the effects of light onthe desiccation of three resurrection plants species. Plant CellEnv. 26:1275–1286.
Farrant, J. M., N. W. Pammenter, and P. Berjak. 1992. Developmentof the recalcitrant (homoiohydrous) seeds of Avicennia marina(Forsk.) Vierh.: Anatomical, ultrastructural and biochemicalevents associated with development from histodifferentiation tomaturation. Ann. Bot. 7:75–86.
Farrant, J. M. and C. Walters. 1997. Ultrastructural and biophysicalchanges in developing embryos of Aesculus hippocastanum inrelation to the acquisition of tolerance to drying. Physiol. Plant.104:513–524.
Finch-Savage, W. E. and P. S. Blake. 1994. Indeterminate develop-ment in desiccation-sensitive seeds of Quercus robur L. SeedSci. Res. 4:127–133.
Gaff, D. F. 1977. Desiccation tolerant plants of southern Africa. Oec-ologia 31:95–109.
Gaff, D. F. 1980. Protoplasmic tolerance of extreme water stress. InN. C. Turner and P. J. Kramer (eds.), Adaptions of plants towater and high temperature stress, pp. 207–230. John Wileyand Sons, New York.
Gaff, D. F. 1987. Desiccation tolerant plants in South America. Oec-ologia 74:133–136.
Gaff, D. F. 1989. Responses of desiccation tolerant ‘‘resurrection’’plants to water stress. In K. H. Kreeb, H. Richter, and T. M.Hinckley (eds.), Structural and functional responses to environ-mental stress: Water shortage, pp. 255–268. SPB AcademicPublishers, The Hague.
Ghasempour, H. R., D. F. Gaff, R. P. W. Williams, and R. D. Gi-anello. 1998. Contents of sugars in leaves of drying desiccationtolerant flowering plants, particularly grasses. Plant GrowthReg. 24:185–191.
Giannopolitis, C. N. and S. K. Ries. 1971. Superoxide dismutases.I. Occurrence in higher plants. Plant Physiol 59:309–314.
Goyal, K., L. J. Walton, and A. Tunnacliffe. 2005. LEA proteinsprevent protein aggregation due to water stress. Biochem. J.388:151–157.
Greggains, V., W. E. Finch-Savage, W. P. Quick, and N. M. Atherton.2000. Putative desiccation tolerance mechanisms in orthodoxand recalcitrant seeds of the genus Acer. Seed Sci. Res. 10:317–327.
Haslekas, C., R. A. Stacy, V. Nygaard, F. A. Culianez-Marcia, andR. B. Aalen. 1998. The expression of a peroxiredoxin antioxi-dant gene, AtPerl, in Arabidopsis thaliana is seed-specific andrelated to dormancy. Plant Molec. Biol. 36:833–845.
Hendry, G. A. F., W. E. Finch-Savage, P. C. Thorpe, N. M. Atherton,S. M. Buckland, K. A. Nilsson, and W. E. Seel. 1992. Free
radical processes and loss of seed viability during desiccationin the recalcitrant species Quercus robur. New Phytol. 122:273–279.
Ingram, J. and D. Bartels. 1996. The molecular basis of dehydrationtolerance in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol.47:377–403.
Kappen, L. and F. Valladares. 1999. Opportunistic growth and des-iccation tolerance: The ecological success of poikilohydrous au-totrophs. In F. I. Pugnaire and F. Valladares (eds.), Handbook offunctional plant ecology, pp. 10–80. Marcel Dekker, New York.
Karner, U., T. Peterbauer, V. Raboy, D. A. Jones, C. L. Hedley, andA. Richter. 2004. myo-Inositol and sucrose concentrations affectthe accumulation of raffinose family oligosaccharides in seeds.J. Exp. Bot. 55:1981–1987.
Kermode, A. R. and W. E. Finch-Savage. 2002. Desiccation sensi-tivity in orthodox and recalcitrant seeds in relation to develop-ment. In M. Black and H. W. Pritchard (eds.), Desiccation andsurvival in plants: Drying without dying, pp.149–184. CABIpublishing, Wallingford and New York.
Knight, H. and M. R. Knight. 2001. Abiotic stress signaling path-ways: Specificity and cross-talk. Trends Plant Sci. 6:1360–1385.
Koster, K. L. and A. C. Leopold. 1988. Sugars and desiccation tol-erance in seeds. Plant Physiol. 88:829–832.
Kranner, I. 2002. Glutathione status correlates with different degreesof desiccation tolerance in three lichens. New Phytol. 154:451–460.
Kranner, I. and D. Grill. 1997. Desiccation and the subsequent re-covery of cryptogamics that are resistant to drought. Phyton 37:139–150.
Leopold, A. C., W. Q. Sun, and I. Bernal-Lugo. 1994. The glassystate in seeds: Analysis and function. Seed Sci. Res. 4:267–274.
Leprince, O., R. Bronchart, and R. Deltour. 1990. Changes in starchand soluble sugars in relation to the acquisition of desiccationtolerance during maturation of Brassica campestris seed. PlantCell Env. 13:539–546.
Li, C. and W. Q. Sun. 1999. Desiccation sensitivity and activities offree radical-scavenging enzymes in recalcitrant Theobroma ca-cao seeds. Seed Sci. Res. 9:209–217.
Mittler, R., S. Vanderauwera, M. Gollery, and F. Van Breusegem.2004. Reactive oxygen gene network of plants. Trends PlantSci. 9:1360–1385.
Mowla, S. B., J. A. Thomson, J. M. Farrant, and S. G. Mundree.2002. A novel stress-inducible antioxidant enzyme identifiedfrom the resurrection plant Xerophyta viscosa. Planta 215:716–726.
Mundree, S. G. and J. M. Farrant. 2000. Some physiological andmolecular insights into the mechanisms of desiccation tolerancein the resurrection plant Xerophyta viscosa Baker. In J. Cherry(ed.), Plant tolerance to abiotic stresses in agriculture: Role ofgenetic engineering, pp. 201–222. Kluwer Academic Publish-ers, The Netherlands.
Mundree, S. G., B. Baker, S. Mowla, S. Peters, S. Marais, C. VanderWilligen, K. Govender, A. Maredza, J. M. Farrant, and J. A.Thomson. 2002. Physiological and molecular insights intodrought tolerance. African J. Biotechnol. 1:28–38.
Navari-Izzo, F., S. Meneguzzo, B. Loggini, C. Vazzana, and C. L.M. Sgherri. 1997. The role of glutathione system during dehy-dration of Boea hydroscopica. Physiol. Plant. 99:23–30.
Ndima, T. B., J. M. Farrant, J. A. Thomson, and S. G. Mundree.2001. Molecular characterization of XVT8, a stress-responsivegene from the resurrection plant Xerophyta viscosa Baker. PlantGrowth Reg. 35:137–145.
Norwood, M., M. R. Truesdale, A. Richter, and P. Scott. 2000. Pho-tosynthetic carbohydrate metabolism in the resurrection plantCraterostigma plantagineum. J. Exp. Bot. 51:159–165.
Norwood, M., O. Toldi, A. Richter, and P. Scott. 2003. Investigationinto the ability of roots of the poikilohydric plant Craterostigmaplantagineum to survive dehydration stress. J. Exp. Bot. 54:2313–2327.
Oliver, M. J. 1996. Desiccation tolerance in vegetative plant cells.Physiol. Plant. 97:779–787.
Oliver, M. J. and J. D. Bewley. 1997. Desiccation-tolerance of planttissues: A mechanistic overview. Hort. Rev. 18:171–214.

787SIGNATURE OF SEEDS IN RESURRECTION PLANTS
Oliver, M. J., S. E. Dowd, J. Zaragoza, S. A. Mauget, and P. R.Payton. 2004. The rehydration transcriptome of the desiccation-toleratant bryophyte Tortula ruralis: Transcript classification andanalysis. BMC Genomics. 2004 Nov 16; 5(1):89.
Oliver, M. J., Z. Tuba, and B. D. Mishler. 2000. The evolution ofvegetative desiccation tolerance in land plants. Plant Ecol. 151:85–100.
Pammenter, N. W. and P. Berjak. 1999. A review of recalcitrant seedphysiology in relation to desiccation-tolerance mechanisms.Seed Sci. Res. 9:13–37.
Piatkowski, D., K. Schneider, F. Salamini, and D. Bartels. 1990.Characterization of five abscisic acid-responsive cDNA clonesisolated from the desiccation-tolerant plant Craterostigma plan-tagineum and their relationship to other water stress genes. PlantPhysiol. 94:1682–1688.
Porembski, S. and W. Barthlott. 2000. Granitic and gneissic outcrops(inselfbergs) as centers of diversity for desiccation-tolerant vas-cular plants. Plant Ecol. 151:19–28.
Proctor, M. C. F. 1990. The physiological basis of bryophyte pro-duction. Bot. J. Linn. Soc. 104:61–77.
Schwall, G., R. Elster, J. Ingram, G. Bernacchia, G. Bianchi, L.Gallagher, F. Salamini, and D. Bartels. 1995. Carbohydrate me-tabolism in the desiccation-tolerant plant Craterostigma plan-tagineum Hochst. In H. Pontis, G. Salerno, and E. Echeverria(eds.), Sucrose metabolism, biochemistry, physiology and mo-lecular biology. Current Topics in Plant Physiology, Vol 14,pp. 245–253. American Society of Plant Physiologists.
Scott, P. 2000. Resurrection plants and the secret of the external leaf.Ann. Bot. 85:159–166.
Sherwin, H. W. and J. M. Farrant. 1996. Difference in rehydrationof three desiccation-tolerant angiosperm species. Ann. Bot. 78:703–710.
Sherwin, H. W. and J. M. Farrant. 1998. Protection mechanismsagainst excess light in the resurrection plants Craterostigmawilmsii and Xerophyta viscosa. Plant Growth Reg. 24:203–210.
Sivamani, E., A. Bahieldrin, J. M. Wraith, T. Al-Niemi, W. E. Dyer,T-H. D Ho, and R. Qu. 2000. Improved biomass productivityand water use efficiency under water deficit conditions in trans-genic wheat constitutively expressing the barley HVA1 gene.Plant Sci. 155:1–9.
Steadman, K., H. W. Pritchard, and P. M. Dey. 1996. Tissue-specificsoluble sugars in seeds as indicators of storage category. Ann.Bot. 77:667–674.
Swire-Clark, G. A. and W. R. Marcotte, Jr. 1999. The wheat LEAprotein Em functions as an osmoprotective molecule in Saccha-romyces cerevisiae. Plant Molec. Biol. 39:117–128.
Vander Willigen, C., N. W. Pammenter, S. G. Mundree, and J. M.Farrant. 2001. Some physiological comparisons between theresurrection grass, Eragrostis nindensis, and the related desic-cation-senstive species, Eragrostis curvula. Plant Growth Reg.35:121–129.
Vertucci, C. W. and J. M. Farrant. 1995. Acquisition and loss ofdesiccation tolerance. In J. Kigel and G. Galili (eds.), Seed de-velopment and germination, pp. 237–271. Marcel Dekker, NewYork.
Vicre, M., O. Lerouxel, J. M. Farrant, P. Lerouge, and A. Driouich.2004. Composition and desiccation induced alterations of thecell wall in the resurrection plant Craterostigma wilmsii Phy-siol. Plant. 120:229–239.
Von Caemmerer, S. and G. D. Farquhar. 1981. Some relationshipsbetween the biochemistry of photosynthesis and the gas ex-change of leaves. Planta 153:376–387.
Walters, C., J. M. Farrant, N. W. Pammenter, and P. Berjak. 2002.Desiccation and Damage. In M. Black and H. W. Pritchard(eds.), Desiccation and survival in plants. Drying without dying,pp. 263–291. CAB International, Oxon.
Wang S. Y., H. J. Jaio, and M. Fuast. 1991 Changes in ascorbate,glutathione, and related enzyme activities during thidiazuron-induced bud break of apple. Physiol. Plant. 82:231–236.
Whittaker, A., A. Bochicchio, C. Vazzana, G. Lindsey, and J. M.Farrant. 2001. Changes in leaf hexokinase activity and metab-olite levels in response to drying in the desiccation-tolerant spe-cies Sporobolus stapfianus and Xerophyta viscosa. J. Exp. Bot.352:961–969.
Whittaker, A., T. Martinelli, A. Bochicchio, C. Vazzana, and J. M.Farrant. 2004. Comparison of sucrose metabolism during therehydration of desiccation-tolerant and desiccation-sensitive leafmaterial of Sporobolus stapfianus. Physiol. Plant. 122:11–20.
Wise, M. J. 2003. LEAping to conclusions: A computational re-analysis of late emryogenesis abundant proteins and their pos-sible roles. BMC Bioinformatics 4:52–71.
Wise, M. J. and A. Tunnacliffe. 2004. POPP the question: What doLEA proteins do. Trends Plant Sci. 9:1360–1385.
Xu, D., X. Duan, B. Wang, B. Hong, T.-H. D. Ho, and R. Wu. 1996.Expression of a late embryogenesis abundant protein gene,HVA1, from barley confers tolerance to water deficit and saltstress in transgenic rice. Plant Physiol. 110:249–257.
Zdobnov, E. M. and R. Apweiler. 2001. InterProScan-an integrationplatform for the signature-recognition methods in InterPro.Bioinformatics 17:847–848.
Zimmermann, P., M. Hirsch-Hoffmann, L. Hennig, and W. Gruis-sem. 2004. GENEVESTIGATOR. Arabidopsis microarray da-tabase and analysis toolbox. Plant Physiol. 136:2621–2632.