the research and management of non-urban koala populations
TRANSCRIPT


Published by Koala Research Centre of Central Queensland Central Queensland University Rockhampton Qld 4702
2001
National Library of Australia Cataloguing-in-publication entry: ISBN I 876674 23 7
All rights reserved. No part of this publication may be reproduced, stored in or introduced into a retrieval system, or transmitted, in any form or by any means (electronic, mechanical, photo-copying, recording or otherwise) without the prior written permission of the author and publisher.
Printed and bound by: University Publishing Unit Central Queensland University Rockhampton Qld 4702
CENTRAL QUEENSLAND UNlVE.RS\TY .. LIBRARY

Contents
List of Reviewers 1 The Koala Research Centre of Central Queensland: An Overview 2 The Central Queensland Koala Volunteers: An Introduction 2
1. Opening Address: workshop on the research and management of 3 non-urban koala Populations Bill Fisher
2. An overview of the understanding of koala ecology: how much more 6 do we need to know? Alistair Melzer and Wayne Houston
3. Landholders address to the koala symposium 46 John Rolfe
4. Community perceptions of koala populations and their management 48 in Port Stephens and Coffs Harbour local government areas, New South Wales Dan Lunney, Dionne Coburn, Alison Matthews and Chris Moon
5. A review of some techniques used to describe koala habitat and its 71 use by koalas with particular reference to low density populations at Eden VicJurskis
6. The Role of Research in Koala Management: Case Studies from 89 South-East Queensland Jim Thompson
7. Koala distribution and density in southeast Queensland: the 105 accuracy and precision of koala surveys. David Dique, Guy Penfold, Jim Thompson, Ros Leslie and Harriet Preece
8. The koala habitat atlas: preliminary results for koala tree species 122 preferences in the city of Ballarat local government area, Victoria Patrick Prevett, Rick Pope, John Callaghan and Lynne Bailey
9. Aspects ofthe ecology of koalas at Blair Athol coal mine 127 Bill Ellis, Peter Hale, Frank Carrick, Moto Hasegawa, Michael Nielsen and David Esser
10. Valuation and management options for koala habitat in the desert 13 9 uplands bioregion of Queensland John Rolfe
11. An investigation into the coprophilous mycobiota ofthe koala 153 (Phascolarctos cinereus, (Goldfuss, 1815)) Tanya Maxwell and Keith Harrower
12. All creatures are equal, though some are more equal than others: the 178 moral and political agency of koalas Robert Kelso

List of Reviewers
Coburn, Dionne Environmental Survey Research NSW National Parks and Wildlife Service
Ellis, William Department of Zoology University of Queensland
Foley, William Botany and Zoology Division Australian National University
Hobson, Peter School ofEducational Studies University ofNew England
Johnson, Chris Department of Zoology and Tropical Ecology James Cook University
Jurskis, Vic Native Forests Division State Forests ofNSW
Kelso, Robert Education and Creative Arts Central Queensland University
Lockie, Stewart Centre for Social Science Research Central Queensland University
Lunney, Dan Vertebrate Ecology NSW National Parks and Wildlife Service
Lyons, Kristen Science Policy Research Centre Griffith University
Mathews, Alison Vertebrate Ecology NSW National Parks and Wildlife Service
McCosker, Julianna Environmental Planning and Biodiversity Environmental Protection Agency
McKillup, Steve Biological and Environmental Sciences Central Queensland University
Menkhorst, Peter Flora and Fauna Branch Department ofNatural Resources and the Environmen*, Victoria
Monypenny, Richard Economics Program James Cook University
Penfold, Guy Conservation Research and Management Unit Environmental Protection Agency
Prevett, Patrick Environmental Management University of Ballarat
Shields, Jim Native Forests Divisional Office State Forests ofNSW
Stratford, Elaine School of Geography and Environmental Studies University ofTasmania
Thompson, Jim Daisy Hill State Forest Queensland Parks and Wildlife Services
1

The Koala Research Centre of Central Queensland
The Koala Research Centre was established in 1993 by the Rockhampton City Council, the Environmental Protection Agency and Central Queensland University. The aim of this Centre is to foster research that will ensure the long-term coexistence of koalas and their habitat throughout the urban, rural and industrial regions of Cental Queensland. In particular, the direction of the Centre's research interests include; the establishment of a data base of koala sightings, development of guidelines for the coexistence of koalas and land use, investigation of methods for the reconstruction of koala habitat and investigation of the methods for optimal management of captive koalas. This work is supported by schools, conservation groups and individuals within the region. In order to optimise the research of the Koala Research Centre, a Scientific Advisory Committee comprised of scientists from the Queensland Environmental Protection Agency, Central Queensland University, the University of Queensland, the Rockhampton City Council and a community representative, has been established to provide peer review.
Central Queensland Koala Volunteers
Koala research in the Central Queensland region is primarily community funded, and therefore relies largely on the contributions of volunteer labour and fund raising for its existence. The Central Queensland Koala Volunteers therefore provide an integral role in ensuring continued research on koalas in this region. In particular, through their work, the Koala Volunteers seek to conserve the koala and other tree living mammals in Central Queensland. This aim is achieved by; supporting research into the koala through the provision of volunteer support to research projects, developing public awareness of the need of koalas and habitat requirements, fostering community support for koalas, assistance in the development of habitat rehabilitation projects, as well as supporting the rehabilitation and release of sick or injured koalas. If you are interested in becoming a member, or would like to receive more information about this organisation, please contact:
Central Queensland Koala Volunteers C/- Alistair Melzer Faculty of Arts, Health and Sciences Central Queensland University Bruce Highway Rockhampton, Qld. 4702 Ph: 07) 4930 9003
2

Opening address: workshop on the research and management of non-urban koala populations
Bill Fisher Queensland Parks and Wildlife Service, Rockhampton, Qld.
Congratulations to the organizers of this workshop, Frank Carrick and David Lamb of the University of Queensland, Alistair Melzer of Central Queensland University (CQU), and John Rolfe, Head of the Emerald campus of CQU and our host. Welcome also to the many speakers and participants from around Australia. May I thank the organisers for the opportunity to be involved with this workshop, on behalf of the Board of the Koala Research Centre of Central Queensland, and my organisation, the Queensland Parks and Wildlife Service.
On receiving the invitation to be involved, I asked myself: What brings so many people to Emerald to talk about koalas? What normally brings crowds together- apart from queues to the football? End of season sales? Lectures on how to make your first million dollars?
I do note that the senate committee on the commercialisation of native wildlife is in Queensland this week. At one stage harvesting koalas was a commercial enterprise in Australia. Is that what the workshop is about?
The issue of attaching an economic value to wildlife has much to offer conservation in at least some situations. Whether farming of emus or kangaroos is a viable proposition I wouldn't like to say. It was an interesting comment I noted the other night that this country wouldn't be too happy if such ventures became hugely successful overseas - with "our" wildlife.
At different times in my career I've been approached with a number of propositions on commercialising wildlife. One of the more interesting when I was based in north Queensland was quite a sincere proposal to set up a cannery for the export of flying foxes to Malaysia. This was pre-morbilli and lyssa viruses. The New Zealanders have talked about an industry based on exporting canned or otherwise prepared possum meat and products - kiwi bear?
Attaching an economic value to wildlife with a return to landholders has been successful with crocodile conservation in the Northern Territory, and to a lesser, and less direct sense in Queensland (not with a return to landholders, rather to crocodile farmers and fauna displays). There have been suggestions as well that the Territorians have been making progress by attaching a value on corellas.
Attaching an economic value to wildlife does appear to be of critical significance for conservation in some developing countries. Illicit trade can be, and is a serious and direct threat to many species in such countries. Some of the African countries with elephant are firmly of the view that the only way to deal effectively with such threats is to have the value, the real dollars, return to the local communities, whether through direct tourism benefits or through a legitimised trade in products, such that there is incentive for local communities to protect against poaching, save habitat and protect waterholes amongst others. I haven't heard the outcome properly, but understood there was a major push on this at a recent IUCN CITES forum.
3

Exporting koalas to zoos in Japan was relatively big business a few years ago. I'm
sure Greg Gordon recalls well the koala kits, the deputations from different
prefectures in Japan, with quite a sales pitch on both sides. While we're not here to consider urban koalas, on the other side of the coin I'm told
that some landholders near Brisbane may be finding koala outcomes an economic
disincentive, with subdivision proposals. Referring back to corellas and fruit bats, it may be that collectively we are not doing
enough about wildlife disincentives - for people who have economic disadvantages
from wildlife, perceived or real. I don't know whether koalas are ravaging too many
cotton crops around here, but farmers do get quite irate about pest fauna. Are we
doing enough for them? Perhaps there is enough empathy for koalas in Australia from
landholders to not require attaching an economic value to them, although this is not
necessarily evident in clearing practices. If the workshop is not- about koala economics, perhaps it is about the conservation
status of koalas? Are koalas a threatened species? Not according to my Department
koalas are listed as "common" on the relevant schedule under the Nature
Conservation Act. Where do koalas fit in this context of rare and/or threatened
species? No doubt many of us drove via Rockhampton to be here for today. It's interesting
that we drove past the only remaining natural occurrence of the bridled nail tail
wallaby to get here, a species previously presumed extinct, and now subject to an
important recovery plan. There is good news with this plan, including understanding
and cooperation from landholders, and early stages on establishing a second colony in
the wild in Idalia National Park. But we're not gathering this number of people to talk
about nail tails. We also drove past a whole host of rare plant species. And a few hours more up the
road is the most endangered mammal in Australia - our Lasiorhinus remains in deep
trouble. But we're not gathering this number of people to talk about northern hairy
nosed wombats either. What is it about koalas? They certainly represent a political minefield. Maybe they
attract research funding? I must admit that from a personal perspective, I do get concerned about the
preoccupation with endangered species when it comes to consideration of nature
conservation, especially in environmental assessment procedures. Whilst it's not as
simple as this, there is almost a trend that says if studies don't pick up any rare or
threatened species or habitats, then everything is okay. Start up the D9 and away we
go. That's not really a fair comment, as some personal concerns regarding those
processes are as much about incremental change and cumulative effects not being
accounted for, which is another story. In any case, there certainly is a deal of
importance necessarily attached to threatened species. Perhaps we could spend rather more energies on ensuring that the common species,
the common habitats and the common communities will still be common in the years
ahead. Maybe that's what the workshop is about - and why koalas and Emerald and
this level of interest all fit together well. Emerald and Central Queensland have certainly been an important focus in the
process of developing tree-clearing guidelines for leasehold lands in Queensland. The
brigalow development scheme showed what can happen when something that is
common is assumed will always be common, and assumes that no special measures
are necessary in the course of development.
4

In the development of tree clearing guidelines, there has been as much to gain in balancing property development and nature conservation in Central Queensland as in any part of the state. The involvement of landholders directly in the development of local guidelines through the Department of Natural Resources has been a major factor in that process. No one would suggest that the guidelines are perfect, but the process of working with the landholders and community groups and developing joint commitments is about as good as it can be.
Researching and managing the common species including, or especially the koala, with its internationally prominent focus, is a vital component of the spectrum of research and management necessary for effective nature conservation outcomes. There is, and can be no one recipe for effective species or nature conservation, and perhaps there is room for greater flexibility in strategies for achieving conservation ends.
We need many recipes to get the results needed. To cook a good stew requires long and low heat. To cook stir-fried vegetables requires the opposite. Both add to our culinary pleasure. We similarly need a variety of approaches to achieving effective nature conservation, and workshops such as this are important in reaching those ends.
May I thank you again for allowing me to be involved in this gathering, and may I wish you all well in achieving a successful workshop, which I now declare officially open.
5

An overview of the understanding of koala ecology: how much more do we need to know?
Alistair Melzer and Wayne Houston Koala Research Centre of Central Queensland, Central Queensland University, Rockhampton, Qld.
Abstract Despite more than a century of community concern for the fate of the koala, its
long-term future appears uncertain. This uncertainty remains despite an extensive research effort over many decades. Do we lack sufficient information to conserve the koala? This paper reviews some of the ecological research and concludes that current ecological knowledge is sufficient for the development of regional koala conservation strategies and local koala action plans.
Introduction Concern for the conservation of the koala has been repeatedly expressed over the
last 100 years. Research into biology and ecology of the species started during the 1920's and 30's. Both conservation concerns and basic research continue today. Despite this, there appears to have been few positive conservation outcomes for the koala. Most koala populations on mainland South Australia and Victoria derived from translocated stock persist, despite periodic intense bushfires and drought. Island populations and some in isolated forest remnants are inherently unstable and overexploit their habitat. Generally, in these states, available habitat limits koala distribution - if not abundance. In New South Wales, despite intense regulatory efforts to manage impacts on koalas and potential koala habitat, and in Queensland where the koala is officially classified as "common", habitat destruction through land clearing is steadily reducing both koala distribution and abundance (Melzer et al. 2000).
In the first quarter of last century community concern focused on the threats of hunting, and this led to the end of the koala fur industry. Hunters were, however, able to exploit other species such as kangaroos. In the last quarter of the 20th century, community concerns have failed to stop land clearing in New South Wales and Queensland. There seems to be no obvious economically viable alternatives to broad acre clearing for farm improvement at the landscape level and for suburban development along the coast. As the extent of habitat is reduced, the koala becomes increasingly vulnerable to extinction. Localised extinctions have already occurred throughout Queensland. If the answer to koala conservation does not lie with community concern, perhaps it lies in the application of outcomes of decades of research.
This paper presents an understanding of koala ecology, and based on this view, presents some principles for practical conservation actions in rural areas.
6

Distribution The koala, Phascolarctos cinereus Goldfuss 1817, is a folivorous arboreal marsupial
occurring naturally in many of the Eucalyptus communities of eastern Australia, feeding predominantly on Eucalyptus leaves. Eucalyptus foliage is high in fibre, tannins and essential oils, and low in available proteins and soluble carbohydrates, and is poor quality fodder. Only four marsupials have managed to exploit Eucalyptus leaves as a major fod<ter source and of these only two, the koala and the greater glider (Petauroides volans), can exist wholly on Eucalyptus foliage (McKay 1995).
Although restricted to Eucalyptus forests and woodlands, the koala exists in a wide range of environments from tropical northern Australia (about latitude 16°) to cool temperate southern Australia Gust below latitude 39°).
In Queensland, at least, koalas are found in moist coastal forests on the eastern seaboard, in the central western subhumid woodlands and some of the Eucalyptus communities fringing watercourses in semiarid western Queensland. The koala, however, is rarely found in wet forests and, in southern latitudes, is generally not found above 600 m (Phillips 1990).
The contemporary distribution of the koala is patchy, and the species is absent from large areas of apparently suitable forest (Phillips 1990). The patchy distribution was noted in the early 1860's (Gould 1863), and remains an enigma today. This is, in part, due to the fluctuation in distribution and abundance of koalas throughout eastern Australia since European settlement. The koala appeared to be rare and of limited distribution at the time of European settlement in 1788 and until about 1830. Gould (1863) found the species difficult to locate and restricted to dense tall Eucalyptus forests. He expressed concern for the future of the species. By 1870, however, the koala was being sighted in lowland open Eucalyptus communities (Parris 1948), and by the 1890's the koala was extremely abundant throughout its range (Phillips 1990). It is postulated that this apparent expansion in regional distribution and increased abundance was due to the removal of aboriginal hunting pressure at the time of European expansion (Parris 1948).
The abundance of the koala during the 1890's led to the development of an intensive hunting industry. From the late 1880's in New South Wales and Victoria, through to the 1930's in Queensland, a series of epidemic disease outbreaks combined with hunting and forest clearing to produce a population crash, and a major reduction in the koala's distribution. By the late 1930's the koala was extinct in South Australia, near extinction in Victoria, and greatly reduced in range in New South Wales. In Queensland the distribution of the koala remained intact, although populations were greatly reduced and fragmented with localised extinctions in some areas. Extensive translocation programs have reintroduced the koala to Victoria, South Australia and parts of southern New South Wales (Finlayson 1934; Parris 1948; Gordon et al. 1990a; Phillips 1990).
Despite these re-introductions, the koala is now considered to have suffered a greater than 50 percent reduction in geographic range since European settlement, "a significant and continuing decline in abundance in more than 50 percent of their former area of occupancy", and is potentially vulnerable (lower risk - near threatened) (Maxwell et al. 1996). This uncertain conservation status is a result of the ongoing destruction of habitat throughout much of the koala's range (Phillips 1990).
Determination of the contemporary distribution of the koala on a national and state wide scale has been made by assessing the presence or absence of koalas in each map sheet in the Australia 1 :250,000 map series (Maxwell et al. 1996) or by analysing
7

responses to community surveys of koala sightings (Phillips 1990; Reed et al. 1990; Menkhorst 1995; Patterson 1996). The first method provides an indication of the extent of the koalas' distribution. The second method also provides a reasonable indication of the extent of distribution, but, as the number of observations increases with the number of observers and access to the environment, it biases any interpretation of relative abundance or habitat utilisation (Ingram and Raven 1991 ).
Phillips (1990) compared the results of the 1985- 1987 National Koala Survey with the results of 1949 and 1975 New South Wales surveys. He concluded that although some populations appeared to have been lost from the southern and western margins of the koala range, "the broad historical distribution of koalas is being maintained". Despite this apparent maintenance of the extent of the koala distribution, Reed et al. (1990) conclude that localised extinctions have occurred within this distribution, especially in the south of the state.
Determinations of distribution at the regional and local scale have also involved community surveys (Lunney et al. 1997) and community surveys in association with direct observation of koalas (Lunney et al. 1996). More commonly, direct methods such as the distribution of koala faecal pellets (Phillips 1995; Phillips and Callaghan 1995; Munks et al. 1996; Pahl 1996; Phillips et al. 2000), call responses to taped koala calls, spotlighting surveys (Jurskis and Potter 1997) and systematic searches for koalas (Hindell 1984; Gordon et al. 1990a; Melzer 1995; Melzer 1996) provide very detailed knowledge of local koala distributions.
In New South Wales these approaches have been used to refine the distribution maps in specific locations with, for example, relatively detailed distribution maps being developed at Iluka in northern New South Wales (Lunney et al. 1996) and in the Eden region of southern New South Wales (Lunney et al. 1997). The latter report, although based on a community survey, is useful because the authors validated over 80 percent of the responses, consequently avoiding many of the biases associated with this type of survey.
Although the koala is not an endangered species, it receives an extremely high degree of State and Commonwealth legislative protection. This is a response to community concern for the welfare of the koala. The koala has always held emotional appeal from the public, and national and international concern over the fate of the koala in Australia forced an end to hunting by 1928 (Phillips 1990). Public concern for the fate of the koala was rekindled in 1985 with fears that epidemics of Chlamydia! blindness, infertility, urinary tract infections and related mortality threatened extinction for the koala (Brown and Carrick 1985). Subsequent research suggested that although Chlamydia spp. may be involved in the decline of some koala populations (Gordon et al. 1990a), the organism is probably endemic in the koala and poses no threat of extinction (Lee and Martin 1988; Ellis et al. 1993). Habitat destruction was highlighted by most contemporary researchers as the principle threat to the conservation of the koala (Lee and Martin 1988; Lunney et al. 1990; Phillips 1990; Melzer et al. 2000) and public concern has recently focused on the need to retain "good" koala habitat.
Taxonomy A species such as the koala with a wide geographic range would be expected to
exhibit geographic variation, with localised populations developing distinctive genotypic races along with phenotypic variations in morphology, physiology, pelage colour, behaviour and habitat preferences (Baker 1968; Kerle 1984).
8

It is not surprising, then, that three subspecies have been described on the basis of body size and fur colour. The Victorian race, Phascolarctos cinereus victor, (type specimen from French Island) is robust, with a broader skull and longer pelage than the other two races. Pelage colour is dark cinnamon to dark brown (Troughton 1935). Mature animal weights range from 9.5 to 13.5 kg for males and 7.0 to 9.8 kg for females (Martin and Handasyde 1995). The Queensland race, Phascolarctos cinereus adustus, (type specimens from Mundubbera) is smaller with a shorter lighter coloured pelage (Thomas 1923). It weighs less than the Victorian race, with males weighing up to 9.7 kg and females growing to 7.6 kg (Melzer 1995). The New South Wales race, Phascolarctos cinereus cinereus, is intermediate in size and colour (Finlayson 1934).
It is assumed that these three races represent a latitudinal cline in body form (Finlayson 1934; Lee and Carrick 1987; McKay 1988; Phillips 1990; Martin and Handasyde 1995), although there has been no formal examination ofthis assumption. Investigations of morphological variation in some Queensland koalas (Melzer, Carrick, Gordon and Lamb pers. com.; Melzer 1995) indicates that morphological variants, in size at least, run contrary to the assumed latitudinal cline. Further, initial studies suggest that this morphological variation, in Queensland at least, may reflect regional genotypic differences (Worthington et al. 1993; Houlden et al. 1996).
Houlden et al. (1996) demonstrated the existence of significant genetic differences amongst koala populations from the Gold Coast, Mutdapilly and Springsure in Queensland; Nowendoc in New South Wales, Kangaroo Island in South Australia and five sites in Victoria. The Victorian and South Australian populations appeared closely related and clearly distinct from the New South Wales and Queensland populations. Each of the Queensland populations and the New South Wales population were clearly distinct from each other. Worthington et al. also detected some "genetic difference" between the Springsure population and south-east Queensland koala populations. Genetic variation in the Victorian and South Australian koalas is much less than in the Queensland and northern New South Wales populations sampled so far, reflecting the recent population bottleneck experienced by P. c. victor (Houlden et al. 1996). To date, no clear boundaries between genetically distinct groups have been identified and there appears to be a broad north - south cline in genetic differentiation (Houlden et al. 1999).
The extent of genetic variation identified in the northern koala populations probably reflects more closely the natural situation. These populations are derived from local stock, unlike the Victorian and South Australian populations, most of which have originated from a very small founder population. For the Victorian koala, at least, it is possible that its ecology and social organisation may be a product of its "unnatural" origins and modern ecological pressures (Melzer 1995). The latitudinal cline and possible regional differences in adaption reflect the ecological amplitude of the species. Regional populations with significant differences in gene frequency can be considered as separate management units. The loss of these units would reduce the species' genetic diversity and threaten the ecological fitness of the species (Sherwin et al. 2000).
Fodder selection Koalas exercise extreme selectivity in fodder choice. They feed predominantly on
the genus Eucalyptus, but will eat other genera to a lesser extent. In any particular area, koalas utilise most species of Eucalyptus but select only one or two species as the predominant fodder source. These staple fodder species represent only a few of the
9

hundreds of Eucalyptus species available to koalas throughout their range (Cork 1987;
Lee and Martin 1988; Phillips 1990) and their selection is apparently unrelated to the
present understanding of Eucalyptus taxonomy (Congreve and Betts 1978). Based on
the National Koala Survey, koalas have been observed in association with at least 69
Eucalyptus species and 30 non-eucalypt species (Phillips 1996). Pahl et al. (1990)
tabulate 55 species of Eucalyptus species that have been recorded in the diet of koalas
and of these, 18 were identified as primary food sources. Non-eucalypt species that
have been recorded as minor dietary components include the following species -
Melaleuca quinquenerva, Melaleuca argentea, Lophostemon suaveolans, L. confertus,
Lisophyllum cunninghami, Acacia cambagei, Eremophila mitchelli and Hakea sp. (U
Nyo Tun 1993; Munks 1993; Hasegawa 1995). Care must be taken in interpreting lists of food tree species. It has been widely
assumed that the tree the koala occupies during the day will be the tree it fed on
during the previous night or will feed on during the next night (Robbins and Russell
1978; Lee and Martin 1988; Hindell and Lee 1990; Pahll996). Hindell et al. (1985)
demonstrated that the assumption is valid for some Victorian koala populations. In
Queensland, at least, the picture is not so clear. Melzer et al. (1995) compared diurnal
tree use with direct observation of the tree species used for feeding. They found that
the species-specific feeding frequency was significantly different from the species
specific diurnal use frequency and that the rank order of species selection for feeding
and species selection for day use were poorly correlated. They concluded that diurnal
tree species selection might only provide an indication of the most frequently selected
fodder species. U Nyo Tun (1993) compared preference in diurnal tree species use and fodder
species selection determined by faecal pellet analysis in some rehabilitated Brisbane
koalas. He found that the most preferred fodder species were the less preferred, or in
the case of Lophostemon confertus, not preferred species for diurnal use.
Hasegawa (1995) examined tree selection by koalas at Point Halloran, near
Brisbane, using faecal pellet counts, radio tracking and faecal pellet analysis. All three
methods identified Eucalyptus tereticomis as the most frequently used tree for diurnal
use and as fodder. Simplistic extrapolation of this finding could be misleading,
however. Hasegawa found that although E. tereticomis was the most frequently
identified tree with faecal pellets beneath it, this species represented only 26 percent
of the total observations. Similarly, the species represented 53 percent of diurnal tree
species use by radio-collared koalas, but occurred in 93 percent of the faecal pellets
examined. Hasegawa (1995) also found that more than 80 percent of the diet of the koalas at
Point Halloran was E. tereticomis. E. crebra comprised up to 6.6 percent, while
Melaleuca quinquenervia varied from about 1 percent to 7 percent when in leaf flush.
Two other species and insect galls from M. quinquenervia formed very small
components of the diet. Hasegawa used Hindell's tree preference index (Hindell et al.
1985) to ascribe a tree preference value (Pi) of 95 to E. tereticomis (Pi = 0 is total
avoidance) (Table I).
10

Table I Tree preference values (Pi where 0 = no preference, >0 = extent of preference Hindell et al. 1985) derived from faecal pellet counts, diurnal tree use and diet analysis at Point Halloran, south east Queensland (Hasegawa 1995).
Tree species Pellet counts Pi Diurnal tree use Pi Faecal pellet analysis Pi
.... !il!:~f!l.YP.~l!:~~~l!~TC! E. tereticornis Melaleuca quinquenervia
18 32 33 17
12 0 19 4
................................................................................... -.... ··················································-··········
63 95 6 1
U Nyo Tun (1993) also examined diurnal tree preferences using Hindell's index (Pi), but used Principal Component Analysis to identify the most preferred food tree species used each month during a limited study of rehabilitated koalas. The most preferred fodder species, as determined by faecal pellet analysis, were E. microcorys, E. propinqua, Lophostemon confertus and E. umbra. This contrasts with the preferences in diurnal tree use summarised in Table II.
Melzer ( 1995) did not assess fodder preferences but did determine preferences in diurnal tree use. He used the Electivity index (E) of Vanderploeg and Scavia (1978) where E=O is random utilisation, E>O is preferential utilisation and E<O is avoidance. The preference rankings (E>O) are summarised in Table III. Melzer found that there was no significant gender difference in preference for diurnal use trees (Table ill).
Table II Preference in diurnal tree use from U Nyo Tun (1993).
Tree species Eucalyptus crebra E. fibrosa fibrosa E. microcorys E. nigra E. propinqua E. resinifera Corymbia trachyphloia E. umbra Acacia aulacocarpa A. leiocalyx A. woodsiana Lophostemon confertus L. suaveolens Melaleuca quinquenervia
Preference index (Pi) 10.37
7.47 8.08 7.01
8.93 3.66
7.47
8.23 6.55
11

Table III Preference in diurnal tree use by central Queensland koalas (Melzer 1995)
denoted by the electivity index E ( -1 = total avoidance, 0 = random use, + 1 = total
preference Vanderploeg and Scavia 1978).
Tree species Gender MeanE
Eucalyptus tereticornis Female 0.56 Male 0.68
Corymbia tessellaris Female 0.41 Male 0.39
E. melanophloia Female 0.49 Male 0.24
E. orgadophila Female 0.66 Male 0.65
E. crebra Female 0.37 Male 0.47
Pahl ( 1996) used faecal pellet distribution and abundance to assess tree usage by
koalas in Logan City, south-east Queensland. He then used Hindell's index (Pi) to
ascribe koala preferences to each species. The most preferred species were E.
microcorys, E. propinqua and E. tereticornis. The results are summarised in Table IV.
Samedi (1995) did not calculate preference indices but stated that the most
frequently used tree species in Brisbane Forest Park near Brisbane, were E. propinqua
(33 percent of sightings), E. microcorys (24 percent of sightings) and E.
drepanophylla (15 percent of sightings). In Victoria, although, E. viminalis and E. ovata seem to be the preferred fodder
species (Hindell et al. 1985; Hindell and Lee 1987), there is some variation among
locations (Hindell et al. 1985), with season (Hindell et al. 1985; Hindell and Lee
1987), across gender (Hindell and Lee 1987) and among individuals (Hindell and Lee
1988). The fodder species preferences (as Hindell's Pi) from the Brisbane Ranges
National Park (Hindell and Lee 1987) are summarised in Table V. Generally, E.
viminalis was preferred, except in summer when the preference switched to E. ovata.
Male koalas tended to prefer E. macrorhyncha and to a lesser extent E. viminalis and
E. ovata. Female koalas consistently preferred E. ovata and E. viminalis.
Fodder tree preference data from New South Wales are scarce. Observations from
the upper Nepean River Valley indicate that E. globoidea and E. sieberi are used
frequently (Tilley and Uebel 1990). There are no data on the relative abundance of
potential food trees in the area so no assessment of preference can be made. Jurskis
and Shields (1995) indicate that koalas in the Eden area generally prefer E.
cypellocarpa and E. seeberi, but provide no indication of how these preferences were
determined. Jurskis and Potter (1997) provide preference data from two locations near
Eden. Preferences were ascribed through the use of the G test to identify
disproportionate use of a tree species. In the escarpment forests E. cypellocarpa and
E. maidenii were preferred, while in the coastal hills and valleys E. consideniana,
E. cypellocarpa and E. longifolia were most preferred.
12
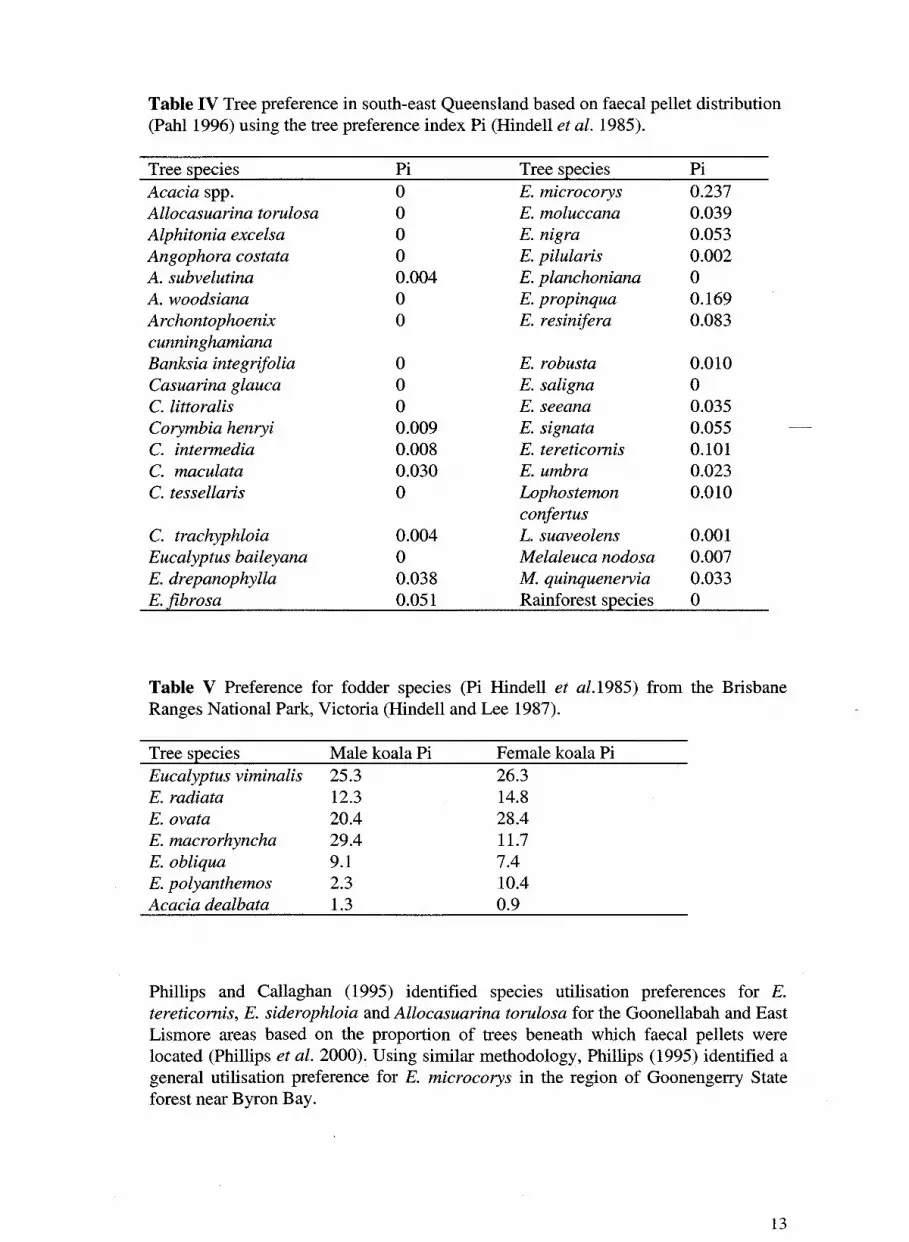
Table IV Tree preference in south-east Queensland based on faecal pellet distribution (Pahl 1996) using the tree preference index Pi (Hindell et al. 1985).
Tree S,Eecies Pi Tree S,Eecies Pi Acacia spp. 0 E. microcorys 0.237 Allocasuarina torulosa 0 E. moluccana 0.039 Alphitonia excelsa 0 E. nigra 0.053 Angophora costata 0 E. pilularis 0.002 A. subvelutina 0.004 E. planchoniana 0 A. woodsiana 0 E. propinqua 0.169 Archontophoenix 0 E. resinifera 0.083 cunninghamiana Banksia integrifolia 0 E. robusta 0.010 Casuarina glauca 0 E. saligna 0 C. littoralis 0 E. seeana 0.035 Corymbia henryi 0.009 E. signata 0.055 C. intermedia 0.008 E. tereticornis 0.101 C. maculata 0.030 E. umbra 0.023 C. tessellaris 0 Lophostemon 0.010
confertus C. trachyphloia 0.004 L. suaveolens 0.001 Eucalyptus baileyana 0 Melaleuca nodosa 0.007 E. drepanophylla 0.038 M. quinquenervia 0.033 E. fibrosa 0.051 Rainforest S,Eecies 0
Table V Preference for fodder species (Pi Hindell et al.1985) from the Brisbane Ranges National Park, Victoria (Hindell and Lee 1987).
Tree s,Eecies Eucalyptus viminalis E. radiata E. ovata E. macrorhyncha E. obliqua E. polyanthemos Acacia dealbata
Male koala Pi 25.3 12.3 20.4 29.4 9.1 2.3 1.3
Female koala Pi 26.3 14.8 28.4 11.7 7.4 10.4 0.9
Phillips and Callaghan (1995) identified species utilisation preferences for E. tereticornis, E. siderophloia and Allocasuarina torulosa for the Goonellabah and East Lismore areas based on the proportion of trees beneath which faecal pellets were located (Phillips et al. 2000). Using similar methodology, Phillips (1995) identified a general utilisation preference for E. microcorys in the region of Goonengerry State forest near Byron Bay.
13

At Tregeagle, seven km south east of Lismore, Faulks (1990) undertook a radio
tracking study of nine koalas and found that E. tereticornis and E. microcorys were the two most frequently preferred tree species in the koala's utilisation of diurnal use
trees. Seven other tree species were preferred less frequently, reflecting variation in the floristics of individual koala home ranges and possibly individual koala
preferences. Preference was expressed as the ratio of observed tree use to expected tree use without selection. Observations of feeding were made but no assessment of
feeding preference was undertaken. E. tereticornis and E. microcorys were the most
frequently eaten species, accounting for almost 79 percent of feeding observations. The results are summarised in Table VI.
Table VI Preferred tree species for diurnal use and frequency of feeding on tree
species near Lismore, New South Wales (Faulks 1990).
Tree species Number of koalas Frequency of preferring the species feeding events for diurnal use %
Eucalyptus 5 52.4 tereticornis E. microcorys 4 26.2 Corymbia intermedia 2 3.6 C. gummifera 1 0 E. grandis 3 2.4 Other Eucalyptus 2 10.7 Cinnamomum 3 4.8 camphora Lophostemon 1 0 confertus Other non eucalypts 3 0
Faulks (1990) also found that there appeared to be a gender difference in diurnal tree
species selection, with male koalas exhibiting a preference for non-eucalypt species. No female koalas exhibited any preference for these species.
Generally, it seems that E. tereticornis, E. microcorys and E. propinqua are the most
consistently preferred fodder species in south-east Queensland and north-east New
South Wales. Other species may be preferred to some extent depending on season, location and gender. Unfortunately, most determinations of preference are based on
diurnal tree use or the distribution of faecal pellets and the data can only be cautiously interpreted as fodder preferences at this stage. In Queensland, Ellis et al. (1998) found
that the best predictor of faecal pellet production was the presence of the koala in a
tree between 6.00pm and midnight. There is a risk, then, that trees used later in the
night or day will be under represented in faecal pellet surveys. An understanding of the allometric constraints to feeding and the digestive strategies
employed by a herbivore should provide the basis for explaining fodder selection by
that herbivore (Milton 1981 ). The relatively small mass of the koala, while allowing mobility in the forest canopy, imposes theoretical, morphological and physiological
constraints on its ability to utilise Eucalyptus foliage as fodder. Eucalyptus foliage is
high in fibre and low in rapidly available energy (Cork 1984). In herbivores, generally high fibre content would be expected to place a limit on the fodder bulk able to be
14

processed each day (McNab 1978), and these constraints should force a relatively small herbivore, such as the koala, to select high nutrient, low fibre fodder such as young foliage, fruit or seeds in their diet (Parra 1978; Demment and Van Soest 1985).
This is the strategy employed by many Australian arboreal herbivores. The brushtail possum (Trichosurus vulpecula), cuscus (Phalanger maculatus) and ringtail possum (Pseudocheirus peregrinus) select a wide variety of young and mature foliage, flowers, fruits, seeds, and on occasion, meat (Kerle 1984; Pahl 1984; Winter and Leung 1995). The greater glider (Petauroides volans) is wholly folivorou, but because of its small size, is able to select young foliage or less fibrous portions of foliage from Eucalyptus species (Cork and Sanson 1990).
Buds and young foliage also form a component of the koalas diet, however mature foliage provides the majority of its diet (Hindell 1984; U Nyo Tun 1993 ). Hindell observed that while koalas appear to prefer young foliage, and consume it first, they then go on to consume large amounts of mature foliage. U Nyo Tun quantified this. He found that mature foliage formed from 50 to 90 percent of the diet of four rehabilitated koalas. Young foliage formed 5 to 35 percent while non-eucalypts represented from 0 to 25 percent of fodder intake. The determination was made through analysis of leaf cuticle fragments in the koala faecal pellets over a year.
Its larger bite size limits the koala's ability to select low fibre portions of foliage so that the bulk of the diet is high in fibre and low in digestible nutrients. The koala, then, appears to be an exception to the constraints expected to apply to small herbivores. Cork and Sanson (1990) suggest that the koala's physiology allow the theoretical feeding constraints to be avoided through reduced metabolic requirements. The energy requirements of the koala are low, requiring a relatively small fodder intake for their maintenance.
The koala appears to utilise a rapid passage of fodder through the gut and the associated digestion and absorption of cell contents, rather than the slower microbial breakdown of cell walls, for the majority of its energy needs. Herbivores that adopt rapid passage of fodder as a digestive strategy would be expected to have a relatively unspecialised hindgut area (Milton 1981). The koala, however, has a highly specialised hindgut, including probably the largest caecum, for body weight, of any herbivore (Lee and Martin 1988). It is not clear exactly what role the caecum has in the koalas physiology, although there is a demonstrative reliance on microbial gut biota, possibly involved in the detannining of cell contents (Smith and Lee 1984), as koalas will die if the biota are removed through antibiotic treatment (Brown and Woolcock 1990).
Herbivores are often seen as adopting digestive strategies favouring either (a) maximum nutritive extraction through long-term retention of fodder in the digestive tract, or (b) rapid passage of fodder through the gut with a consequent reduction in efficiency of nutrient extraction (Milton 1981). The koala appears to have adopted a digestive strategy that incorporates both options. Large leaf fragments and fibre bundles pass rapidly through the gut ( <20 hrs ), while fine particles and solutes are selectively retained in the caecum and proximal colon for long periods (>200 hrs) (Cork and Sanson 1990).
The apparent duel digestive strategy adopted by the koala may explain some of the difficulty in understanding fodder selection in this folivore. Herbivores that rely on slow, efficient nutrient extraction as the principal digestive strategy tend to be more selective feeders, reflecting the greater time commitment to each meal.
15

It would be a disadvantage to fill the gut with relatively indigestible fodder requiring
an increased time commitment to digest. On the other hand, herbivores adapted to
rapid rates of fodder passage can process more fodder per unit time and tend to be less
selective feeders (Bell1971; Van Soest 1982).
As mentioned, the koala is a highly specialised folivore, feeding predominantly on
Eucalyptus and, to a lesser extent, a few other genera. Commonly, at any one location,
one or two Eucalyptus species form the diet staple, while a number of other
Eucalyptus and non-Eucalyptus species are eaten to a lesser extent (Phillips 1990).
Freeland and Janzen ( 197 4) suggest that such a specialised feeder could only develop
in an environment with a reliable, abundant and year round supply of foliage. Such an
environment would allow the evolution of specialist metabolic and behavioural
pathways by the herbivore to deal with the plant defences to herbivory. This appears
to be the case with the koala, which is able to metabolise much of the dietary
Eucalyptus oils (Eberhard et al. 197 5) and possesses a dietary system that allows the
rapid passage of indigestible lignin, while retaining the relatively digestible cell
fractions (Cork and Sanson 1990). The koala, however, is also highly selective about which species, tree or tree
component is eaten. Furthermore, this selection may differ with the gender of koala,
the season or between regions (Lee and Martin 1988; Phillips 1990).
Fodder selection by koalas has been attributed variously to, avoidance of the high
fibre and toxic components or selection for low fibre, high soluble carbohydrate, high
nutrient concentrations (especially nitrogen) and optimal moisture content. Pratt
(1937) proposed dietary choice based on the presence or absence of foliar cyanogenic
glycosides. These glycosides, however, have been found to occur infrequently in
Eucalyptus, and although they may provide a basis for the rejection of some fodder,
they are not thought to provide a general basis for fodder selection (Lee and Martin
1988). Pratt ( 1937) also argued that koalas selected fodder for the Eucalyptus essential oils
phellandrene and cineole as thermoregulatory aids despite asserting that cineole is
probably toxic to koalas. In contrast, Fleay (1937) proposed that koalas select fodder
that contains cineole, which accordingly must be an essential dietary constituent. Betts
(1978) agreed, and considered that an intake of cineole was "necessary for koala
health". Fodder selection, however, was based on an avoidance of the sesquiterpenoid
class of essential oils. Eberhard (1978) and Southwell (1978) found no clear
relationship between essential oil composition of Eucalyptus leaf and fodder selection
by koalas. Fodder selection has been linked both with essential oils (greater than 2
percent of dry matter) and leaf moisture levels (Hume and Esson 1993). They
suggested a role for eucalypt oils as a positive feeding cue and that oils with a
relatively high proportion of volatile monoterpenes may be preferred. The observation
that koalas always smell fodder before selecting a leaf suggests that some volatile
characteristic is involved in determining selection. Further studies are needed to
positively identify this volatile agent. Osawa (1993) found that the sugar content of
foliage influenced fodder selection. Captive koalas in Japanese zoos tended to favour
eucalypt foliage containing only monosaccharides, while avoiding foliage containing
di- and trisaccharides. He speculated that the avoidance of more complex sugars was a
consequence of some volatile compound in the leaf rather than the sugars themselves.
Eberhard (1978) suggested that koalas accept or reject fodder on the basis of its
palatability (Osawa'a sugars perhaps) and nutritional content rather than essential oil
content.
16

This suggestion was supported by Ullrey et al. (1981). They reported the selection of high nitrogen, low fibre fodder by captive koalas. Degabriele (1981; 1983), however, argued a role for foliar nitrogen alone in fodder selection. This was tested by Hindell (1984), who failed to find a general relationship between fodder selection and nitrogen or leaf nutritional status, and he considered this to be consistent with the physiology of koalas. The koala's digestive system allows the rapid passage of fibre and large fragments while retaining cell solutes and fine particulate matter in the proximal colon and caecum. According to Hindell, the caecum may be the site of microbial d'etannining of proteins and it may be the concentration of tannins likely to bind to protein, rather than the leaf nutrient content, which determines fodder preference.
Cork and Sanson (1990) considered it unlikely that nitrogen alone would be limiting in any environment and consider nitrogen to have an uncertain relationship with fodder quality and selection in foliage. This was based on a review of studies on diet selection by koalas and other arboreal marsupials. Subsequently Pahl and Hume (1990) demonstrated a significant increase in koala fodder selection above a threshold of 1.8 percent foliar nitrogen and 65 percent moisture.
Despite this, Cork (1992) argued that the foliar polyphenols, including the tannins, effect the quality of foliage as fodder and helps to explain fodder and habitat choice in koalas and other arboreal, marsupial folivores. The polyphenols, however, must be considered in relation to other leaf constituents such as foliar nitrogen, phosphorous and potassium. Some support for this hypothesis comes from a study of diet selection by captive koalas in New South Wales undertaken by Hume and Esson (1993), who were able to associate condensed tannins (negatively) and the ratio of nitrogen to condensed tannins (positively) to preference for fodder. As yet, no similar relationship has been identified in wild koala populations.
Leaf moisture may have a bearing on fodder selection. Hindell (1984) found significant, but weak, correlations between the water content of foliage from potential fodder species and fodder preference in free ranging koalas. The relationship was not detected in eucalypt species of low fodder preference despite having high foliar moisture contents. Presumably other leaf components can override moisture content as a factor in fodder selection. Phenol-terpene hybrids in eucalypt foliage appear to influence fodder choice in captive folivores (Lawler et al. 1998; Lawler and Foley 2000) but their role in determining habitat quality has not yet been investigated.
Pahl and Hume (1990) demonstrated a significant increase in koala fodder selection above a threshold of 65 percent moisture. Hume and Esson (1993) found the threshold to be about 55 percent moisture, while Melzer (1995) identified two thresholds. One threshold was obtained in November when the threshold was about 63 percent, while the other was identified in September when the threshold was about 51 percent. Melzer (1995) suggested that the two moisture thresholds might reflect a change in fodder utilisation, in response to the moisture demands of thermoregulation in the koala. Hindell ( 1984) had suggested that the koalas were adjusting their fodder selection to maintain moisture intake during drought conditions. Ellis et al. (1995) confirmed this view in a study of water turnover in a population of koalas in sub-humid central Queensland. To maintain water balance with increasing evaporative water loss during dry seasons, both male and female koalas increased their utilisation of the relatively moist Eucalyptus tereticomis, and correspondingly decreased their use of the drier Eucalyptus crebra.
17

The moisture threshold identified by Pahl and Hume ( 1990) is higher than those
reported by Hume and Esson (1993) and Melzer (1995). The season under which their
trials were undertaken was not stated. The animals used were Victorian koalas
transported to Armidale in northern New South Wales. The threshold then, may
reflect the moisture demands of southern koalas in a novel climate at the time of the
trials. The moisture threshold identified by Hume and Esson (1993) was derived from
animals in 13 zoos across about 1,000 km and over a number of seasons.
Consequently, it is not possible to assess the influence of climate on this result.
Despite these problems of interpretation, it appears that generally koalas will change
their selection of fodder to meet moisture requirements.
It appears that fodder selection in koalas is a complex process, related to leaf
chemistry and environmental variables such as climate and soil chemistry. There is, as
yet, no indication as to the olfactory basis for selection. Leaf moisture is important
under certain environmental conditions and provides one means of interpreting shifts
in fodder selection within a population, but does not account for all fodder preferences
or avoidances.
Habitat selection Fodder quality may have a bearing on habitat selection as well as fodder selection
(Cork 1992). Under the "resource availability hypothesis" (Bryant et al. 1992), higher
nutrient soils produce fodder with lower concentrations of digestibility reducing
polyphenols. Consequently, the better quality soils would support a habitat of better
quality for the herbivore. There is some evidence that koala populations tend to be
greater in tree communities on richer soils (Hindell 1984; Phillips 1990: Reed et al.
1990), but as yet there is no direct evidence linking fodder quality with habitat
selection. As the koala has no significant natural predators, apart from human hunters, the
contemporary populations are effectively resource limited (Lee and Martin 1988).
Accordingly, it would be expected that the koala would "adopt" a strategy of optimal
habitat selection (Rosenzweig 1991). As population density increases, habitat use
expands into less optimal habitat. This density dependent habitat.selection, according
to Rosenzweig, results in an "ideal free distribution" where a species has distributed
its individuals proportionally, with regard to available resources, among the habitats.
This theory, developed by Fretwell and Lucas (1970), assumes amongst other things
that the density of species' individuals correlates with resource density within the
various habitats. From the point of view of koalas then, the density of koalas in any habitat should
reflect the quantity of resources at the site, and hence the quality of the habitat.
Consequently, an analysis of a number of habitats supporting a range of koala
densities should provide an understanding of what determines koala habitat quality.
Numerous descriptions of koala habitat have been made (Finlayson 1934; Lee and
Carrick 1987; Hindell and Lee 1987; Cork et al. 1988; Gordon et al. 1990a; Phillips
1990; Reed et al. 1990; Melzer 1995). Those that assess habitat structure are shown in
Table Vll. Unfortunately, the lack of an adequately defined floristic and structural
classification of Eucalyptus communities used by koalas in most of these reports has
hindered the more general applicability of these studies in an explanation of the
patchy distribution of the koala.
18

Finlayson (1934) noted the Central Queensland koala's preference for "open eucalyptus parks" and avoidance of "scrubs", but provided no definition of his terms. Parkland can be described as "a grassland region with isolated or grouped trees" while scrub can refer collectively to an area "covered by low trees or shrubs" or "tall thick rainforest" (Delbridge et al. 1981). These terms are thus too ambiguous to be usefuL
Lee and Carrick (1987) described koala habitat as primarily open forest and woodlands dominated by Eucalyptus species. They note, however, that prior to 1850 koalas were only recorded in the dense forests of eastern Victoria.
Phillips (1990), reporting the results of the 1985 - 1987 National Koala Survey, described the structure of koala habitat in equally general terms. In Victoria and South Australia, koalas were more often recorded in woodland than open forest, and rarely in closed forest. In Queensland most sightings were from open forest, less frequently from woodland and rarely from closed forest. There was no definition of the tree community descriptive terms and no way of testing differences in definition between states or region. It was concluded that koalas preferred more open habitat to dense habitat. The majority of these sightings were associated with the most preferred fodder species in each state and were found in conjunction with better watered and/or more fertile soils.
Reed et al. (1990) provided a similar ecological interpretation of a 1986 - 1987 koala survey in New South Wales. Their conclusions were generally the same as those of Phillips (1990), but with the addition of a structural definition of koala habitat. They found that koalas were most commonly associated with tree communities "characterised by medium to tall Eucalyptus with tree cover between 10 and 70 percent". These structural classifications of koala habitat indicate a koala preference for relatively open Eucalyptus communities. The generalised nature of these descriptions, however, fails to provide any understanding of why koalas are more frequently found in such habitats.
Hindell and Lee (1987), working in the Brisbane Ranges near Melbourne, linked koala habitat utilisation to the floristic and structural aspects of Eucalyptus communities. They found that koala density was higher where the most preferred fodder species was most abundant and had greater height and large canopies. This was reflected in a negative correlation between koala density and tree density. Hindell and Lee (1987) concluded that the large, most preferred fodder trees provided more foliage, thus requiring less frequent moves by the koala, shelter from climatic stress and refuge from predation. Other workers have found a similar relationship with tree girth (Lee and Martin 1988; White 1994; Hasegawa 1995; Melzer 1995). Lee and Martin (1988) observed a relationship between clusters of home ranges and patches of larger trees scattered through the woodland. This was not the case in central Queensland, as home ranges were too large and widespread to be associated with clusters of large trees (Melzer 1995). Norton and Neave (1996) suggest another explanation for preference for larger girthed trees, noting that larger trees have deeper root systems that would better enable them to procure moisture during harsher conditions than smaller sized trees. In the multi layered eucalypt forests of south-east New South Wales, Jurskis and Potter (1997) reported that koalas tended to use trees that had distorted crowns and have been overtopped or restricted by more vigorous growth.
19

Table VII Descriptions of koala habitat structure.
Author Location Habitat Assessment
Method Assessment Findings
Bennet et NE Vic. al. 1991
Hindell and SE Vic Lee 1987
?
floristic classification
Martin 1985b
Vic ?
Faulks 1990
NE NSW floristic classification
ground surveys of 8 habitat types
ground surveys
ground surveys
ground surveys
Reed et al. NSW 1990
proforma sheet to postal proforma public sheets to public
Jurskis and SE NSW Forestry ground surveys
Potter near Eden Commission ( 1997) forest types Paterson Qld proforma sheet to proforma sheet to
1996 public public and field surveys
Samedi SEQld structure (Specht ground surveys
1995 1970) and floristics
Gordon et SE Qld floristics and ground surveys
al. 1990a location wrt creek and slope
Gordon et Central floristics and ground surveys
al. 1990a Qld location wrt creek and slope
20
open forests dominated by E. radiata and E. dives eucalypt forests dominated by E. viminalis and E. ovata open forests dominated by E. obliqua or E. ovata open-forest dominated -by Cinnamomum camphora, E. tereticornis and E. microcorys 68% of sightings in Medium to tall Eucalyptus of 10-70% cover and 24% in medium Eucalyptus >70% cover koalas more frequently used dry sclerophyll than wet sclerophyll coastal Melaleuca forests, tall open forests, open forests and woodlands, riverine forests preferred habitat was open forest (10-30% cover) eucalypt woodlands dominated by E. tereticornis eucalypt woodlands by dominated by E. tereticornis or E. tereticornis I microtheca

Melzer 1995
Munks 1993
Central Qld
Central Qld
structure (Specht ground surveys 1970) and floristics
land system ground surveys (Morgan 1990) and floristics
open woodlands (1 0-30% cover) dominated by E. tereticornis or E. crebra woodlands and open woodlands dominated by E. camaldulensis
Cork et al. (1988) ignored the structural aspects of Eucalyptus forests and used floristic parameters to describe koala habitat. They considered long-term koala populations, near Wedderburn in New South Wales, to be associated with tree communities containing a high frequency of the most preferred fodder species, E. punctata. Apart from the term "extensive forest", no indication of tree community structure was provided.
Samedi (1995) described a significant preference in habitat selection by captive reared koalas released to forests near Brisbane in Queensland. They preferred open forest (Specht 1970)- dry sclerophyll- with a height of 20 to 30m, and a sparse midstratum of low trees and very tall shrubs. The most common tree species were Eucalyptus drepanophylla, E. microcorys, E. propinqua and E. punctata. Beneath the trees and shrubs, the lower mid-stratum and ground stratum was composed of ferns, forbs and grasses as well as shrubs and vines. The species most commonly used by koalas were E. microcorys, E. punctata and E. drepanophylla. This open forest community was located on the southern aspect of ridges in Samedi's study area. A closely related community (also open forest) abutted the preferred community. Generally this community was avoided by the koalas despite the presence of the most utilised tree species (E. microcorys, E. punctata) within it. These species were among the most common tree species in the preferred community, but much less common in the avoided community. Samedi concluded that the presence of individual tree species was not the dominant factor in influencing habitat selection. It appears that in this case, as with Cork et al. (1988), the density of most frequently used tree species was a determining factor.
In the semi-arid woodlands of north Queensland, Munks et al. (1996) found that the occurrence of koalas (by faecal pellet distribution) was positively correlated with proximity to creek lines, total basal area of trees and species richness, but negatively correlated with the basal area of Acacia species.
Melzer (1995) undertook a floristic and structural classification of the habitat of five koala populations with contrasting population density. He found that, as with Hindell and Lee (1987), the highest koala population density was associated with the most open habitat, and despite the presence of fodder species in all habitats, the high population densities were associated with a high frequency and a high relative frequency of most preferred fodder species. These findings differed subtly, but significantly, from the conclusions of Hindell and Lee (1987), Cork et al. (1988) and Samedi (1995). These authors discussed koala habitat selection in terms of the absolute density of most preferred tree species on their sites. Melzer found that the relative values were significant rather than the absolute values. He points out that there were more most preferred trees per koala at sites with a low population density than at sites with a high koala population density. At the low density sites the preferred trees were scattered through a thick tree community composed
21

predominantly of less preferred tree species. Although not stated by Samedi, an
examination of his community descriptions suggest that the relative frequency of most
used tree species was less in the avoided open forest (Upper and mid-stratum species
25) than in the preferred open forest (Upper and mid-stratum species 16). Melzer
argued that where the most preferred tree species occur at highest relative frequency,
the search effort by koalas would be reduced. Consequently there would be a net
energy saving to those koalas over the koalas exploiting habitat where the relative
frequency of preferred trees was low. The foliar nutritional characteristics of Eucalyptus communities might be an
additional habitat attribute to use when describing the distribution of koalas.
Degabriele (1981; 1983) proposed that the abundance and distribution of the koala
was limited primarily by the availability of nitrogen. Cork and Sanson (1990) argued
that it was unlikely for any one parameter to be generally limiting and postulated, on
the basis of data relating to other arboreal marsupials, that koalas prefer Eucalyptus
communities with high concentrations of foliar nitrogen, phosphorous and potassium,
which would reflect the availability of these elements in the soil. Cork (1992) argued
further that the concentration of polyphenolic anti-herbivory compounds in
Eucalyptus foliage increased as foliar nitrogen, phosphorous and potassium
concentrations decreased. He considered that the absence of koalas from some
Eucalyptus communities on poorer soils could be partly explained by relatively high
foliar concentrations of polyphenolic compounds. Soil nutrients were also implicated
in the selection of preferred habitat by liberated captive koalas in south-east
Queensland. Samedi (1995) failed to find any significant relationship between soil
nutrients and koala habitat selection. He did, however, find that the preferred habitat
of the released koalas had soils with relatively (but not significantly) high sodium
concentrations. He suggested that sodium could affect leaf quality and may also have
a role in metabolic water balance in the koala. Melzer ( 1995) found no association
between koala population density and total foliar nitrogen, phosphorus, potassium or
sodium. This was despite finding significant differences among sites and among tree
species. Gordon et al. (1990a) considered that abiotic factors, such as drought, controlled
koala population density and the regional distribution of koalas by acting on the
abundance of suitable fodder. They noted negative correlations between rainfall and
koala population density at Springsure, in central Queensland, and described the
localised reduction of koala range and abundance during drought in southern
Queensland. They concluded that koala populations persisted in habitats that
happened to have appropriate abiotic parameters such as secure underground aquifers.
The relationship between koalas and creek lines in semi-arid north Queensland
suggested by Munks et al. (1996) fits this model. Inherent in this description is the
probability that leaf moisture content declined as a consequence of a lack of rainfall,
and that the koalas responded in some way to the drying fodder. Hindell ( 1984)
suggested that koalas would adjust their fodder selection to maintain moisture intake.
On their site, however, this did not result in a change in tree community. Such a
change in community utilisation did occur with male koalas in response to a drying
environment in Central Queensland (Melzer 1995). Greater use was made of the
relatively drier community type during the winter months than the summer when
water stress was higher and significantly reduced levels of leaf moisture in the dry
community type were observed. Melzer noted that leaf moisture levels in the preferred
fodder tree (E. tereticornis) that formed the creek bank community remained above
22

threshold levels throughout the year. This pattern of habitat use based on leaf moisture levels in the poorer quality habitat concurred with an observation by Gordon et al. (1990a), who described the existence of a relationship between koala density and rainfall, but only in the poor quality habitat. Further support for this hypothesis comes from a close examination of data presented by Hasegawa (1995). He observed that use of E. tereticomis was higher in autumn 1993 than autumn 1994 and that 1993 was drier than 1994.
A widely distributed animal such as the koala which utilises a broad range of environments across many different biogeographic zones, would be expected to have developed regional variants adapted physiologically and behaviourally to the local environment (Baker 1968). This appears to be the case with the brushtail possum that is found across Australia and has developed an extremely variable approach to fodder selection, habitat selection and habitat utilisation (Kerle 1984). The strategies employed by koalas in fodder and habitat selection may also vary among regions, obscuring general principles in habitat selection and compounding the problems associated with generalisations drawn from widely geographically separated studies. Despite this it generally seems that forest structure and floristics influence koala habitat selection and leaf moisture and climate have a bearing on habitat selection.
Habitat utilisation Regional adaptation to local environments should also affect the nature of habitat
utilisation by koalas. It would be expected that habitat utilisation by any herbivore would be influenced by: (a) the climatic conditions acting on the biotic and abiotic characteristics of the
habitat and on the physiology of the herbivore, (b) seasonal changes in floral phenology and the behaviour of the herbivore, (c) social imperatives acting on individual herbivores, and (d) the structural and floristic characteristics of the habitat (Montgomery 1978).
These conditions should also apply to the koala, although there are very few studies in which all such aspects have been examined.
The patterns of habitat utilisation appear to be affected by annual rainfall. Gordon et al. (1990a) found koala population density along a streamline was inversely related to rainfall. Hindell (1984), Ellis et al. (1995) and Melzer (1995) all noted changes in fodder selection and habitat utilisation associated with environmental moisture balances. The leaf moisture threshold in fodder selection identified by Pahl and Hume (1990), Hume and Esson (1993) and Melzer (1995) may partly explain these results, and could provide a measure of seasonal changes in habitat suitability in some regions.
Local climate may also have an effect. Temperature may determine patterns of tree use and on hot days koalas may utilise the dense foliage of non-fodder trees (Lee and Martin 1988). Melzer (1995) found that the frequency of female koalas staying in the same tree overnight or returning to the same roost tree increased as monthly rainfall totals increased. This apparent sedentary behaviour also increased with increasing monthly mean minimum temperatures. Hasegawa (1995) observed an increase in use of non-fodder trees such as Melaleuca for roosting during the hotter summer months. He attributed this to differences in leaf phenology between Melaleuca and Eucalyptus and consequent foliage cover during the hot period of the year.
23

Seasonal as well as climatic factors can influence habitat utilisation and Hindell
(1984) reported seasonal differences in fodder species preference for koalas in the
Brisbane Ranges of Victoria. In contrast, Hasegawa (1995) found that seasonal
changes in tree use were not reflected in diet that did not change during the year. He
suggested that changes in tree roost preference reflect needs other than food, such as
thermoregulation during summer. Melzer (1995) found male koalas moved greater
distances overnight during the breeding season, while female koalas showed a (non
significant) tendency to increase overnight distances moved during the non-breeding
period. Furthermore, seasonal changes in habitat use were observed.
Climatic and seasonal factors influence phenotypic changes in Eucalyptus trees
(Specht and Brouwer 1975) that in turn may then affect habitat utilisation patterns.
White and Kunst (1990) found that species utilisation by a Redland Bay koala
population in south-east Queensland changed with the flowering periods of the
Eucalyptus species. Similarly, the density of female koalas in Victoria's Brisbane
Ranges was associated with parts of the available habitat where preferred Eucalyptus
species were in leaf flush (Hindell 1984; Hindell et al. 1985). U Nyo Tun (1993)
reported a seasonal change in the amount of leaf flush consumed in south-east
Queensland (more during the summer months). Utilisation of a particular habitat by koalas appears also to be affected by the
structure of the forest community. Gordon et al. (1990a) reported that koalas near
Oakey in south-east Queensland appeared to use young regrowth Eucalyptus populnea
stands more intensely than adjacent mature stands. In contrast, Hindell and Lee (1987)
found a preference for portions of habitat with trees of larger girth, while Mitchell
( 1990) reported smaller home range sizes where the density of large trees was high.
In central Queensland, Melzer (1995) found that 72 percent of the diurnal utilisation
of tree species by both male and female koalas involved trees with mean girths
(measured at breast height) greater than the species average.
In north-east New South Wales, Faulks (1990) found that koalas used trees with
diameters at breast height varying from 0.12 m to 1.97 m. The mean diameter of trees
preferred by males and females was 0.70 m and 0.57 m respectively. These
determinations were made from diurnal tree use. Similarly Lunney et al. (1996) found
that over 55 percent of the trees used by koalas at Iluka in northern New South Wales
had diameters from 0.31m to 0.70m, even though the majority of trees in the study
area had diameters from O.llm to 0.30m. In south-eastern New South Wales, Jurskis
and Potter (1997) reported that koalas generally preferred trees with a diameter at
breast height of between 0.50m and 0.90m for diurnal use.
Hasegawa (1995) used both faecal pellet distribution and diurnal sighting to
investigate tree size selection. Generally trees less than 0.50m girth at breast height
(diameter= 0.16m) were infrequently used. Trees with girths of between 0.50m and
l.Om (diameter = 0.32m) were used slightly more frequently than trees with girths
greater than l.Om. The diurnal sightings, however, revealed that the selection of trees
with girths greater than l.Om increased in winter for both male and female koalas.
White (1994) found no correlation between frequency of observation of koalas and
tree density or stem girth. Sites in this study were isolated patches of remnant
vegetation amongst farmland and a significant relationship was found between koala
numbers and patch size indicating a random distribution of koalas. Despite this, the
trees selected were generally larger on average than the mean size of those available.
The diameter at breast
24

height of selected E. tereticomis was generally greater than 0.55m while that of E. crebra was generally greater than 0.36m.
Social behaviour within a koala population may be a major determinant in habitat utilisation. All koalas live in prescribed areas (range) within the available habitat (Eberhard 1978; Robbins and Russell978; Hindelll984; Mitche111990). Male koala ranges may increase in area in the breeding season; during this period male koalas change locations more often and visit more trees each night than female koalas (Mitche111990). Melzer (1995) observed gender differences in habitat use with males more common in a ridge associated community and females more common in a creek associated community. No differences, however, were observed in diet. Only one study (Hindell and Lee 1987), has shown significant gender differences in the diet of koalas.
Young koalas disperse from optimal habitat to sub-optimal habitat or may be displaced by older individuals (Gordon et al. 1988; Mitchell and Martin 1990). This exclusion appears to be strong as young koalas in suboptimal habitat at Mungalalla Creek, south-west Queensland, failed to use adjacent optimal habitat occupied by older males during a severe drought and eventually died (Gordon et al. 1988). The koalas resident in the optimal habitat survived the drought. Martin (1985b) reported that koalas at Walkerville, Victoria, moved to sub-optimal habitat after over-browsing the preferred habitat, presumably displacing the resident male koala population (Lee and Martin 1988).
Comparisons of home range between the southern and northern limits of the species show considerable variation in area (Table VIII). The home ranges of Victorian koalas are small and clustered with extensive overlaps. In contrast, home ranges of central Queensland koalas are widespread, large and have minimal overlap (Melzer, 1995). Mitchell (1990) postulated that range area was inversely related to population density and positively related to the density of preferred trees. This may well apply regionally, but differences in home range size of central Queensland and Victorian range areas are so great that some other factor is likely to be involved (Melzer 1995).
Whether these differences relate to fundamental differences in habitat resource availability, subspecies differences in habitat exploitation patterns or a combination of both is yet to be determined.
Highest population densities were recorded in southern Victoria and north-east New South Wales. Densities of around one per hectare were commonly recorded from favoured koala habitats in all parts of Australia including inland sites ( eg. Gordon et al.1990a). Very low densities (less than 0.1 per hectare) were observed in coastal NSW, coastal Queensland and inland Queensland. It has been assumed that habitats where koalas occur in low densities are indicative of poor quality habitats and populated by subordinate koalas seeking new ranges. As pointed out by Melzer and Lamb (1994), this may not necessarily be so. For example, such low population densities may be representative of the majority of habitats in the Brigalow Bioregion of central Queensland. They suggest that these communities should be thought of as habitats with low carrying capacity rather than "poor quality".
25

Table VIII Estimates of koala population density and home range area.
Author Locality Method Average Average Population female male density range area range area (lha) (ha) (ha)
Eberhard 1972 SA - Kangaroo - 1.5 to 3.0
(cited in Island Mitchell 1990) Mitchell and Vic- Redbill 2.2 to 6.2
Martin 1990 Creek Martin 1985a Vic- 0.7 to 3.0
Walkerville Mitchell1990 Vic- French harmonic mean 1.18 1.7 3.9 to 8.9
Island Mitchell et al. Vic - Raymond - 0.3
1988 Island Sharpe 1980 Vic - Phillip centre of 0.15 to 0.26 to 5.3
(cited in Island •• 1
activity 0.35 1.07
Mitchell 1990) Hindell 1984 Vic - Brisbane harmonic mean 2.08 3.14 0.7 to 1.6
Ranges Hull1985 Vic - Ballarat min. convex 15.2 14.4 not
(cited in 2 recorded areas
Mitchell 1990) Lunney et al. NSW- near Port - 0.05
1995 Stephens Wellwood NENSW-near - 0.1
1995 Pottsville Ga111980 NE NSW- near - 3.8 to 8.0
(cited in Lismore Mitchell and Martin 1990) Faulks (1990) NENSWnear minimum 2.98 12.35 5.6
Lismore convex polygon 0.57 3.32 modified minimum area (a form of neighbour clustering)
Jurskis and SENSWnear minimum 149 (n=3) 316.5 0.006
Potter ( 1997) Eden convex polygon (n=2) harmonic mean 88.33 (n=3) 132.5
(n=2)
UNyoTun SE Qld- near centre of 13.75* 19.67* 0.4 (White
1993 Brisbane (coast) •• 1
activity and Kunst, 1990)
Samedi 1995 SE Qld- near centre of 11 to 42* 16 to not
Brisbane (west) •• 1
activity 210* recorded
26

Hasegawa 1995 SE Qld - near centre of 4.6 to 5.6 3.6 to 4.3 1.9 to 2.5 Brisbane (coast) • • I activity
min. convex 2.5 to 3.6 1.8 to 2.8 2 area
harmonic mean 2.7 to 3.3 1.3 to 3.1 White 1994 SE Qld- near harmonic mean 1.0 to 45.2 4.3 to 0.25
Ipswich 53.2 White and SEQld 0.01 to 0.6 Kunst 1988 (cited in White 1994) Gordon et al. SE Qld- near 0.6 to 1.5 1990a Oakey Gordon et al. Central Qld - 1.0 to 2.5 1990a near Springsure Melzer and Central Qld - 0.01 Lamb 1994 near Capella Melzer 1995 Central Qld - harmonic mean 39.2 86.5 0.02 to 0.4
near Springsure min. convex 37.6 79.5
2 area *study based on released koalas; 1 Jennrich and Turner 1969; 2 Hayne 1949.
A relationship between gender and home range size is apparent in Table Vill, with males generally having larger home ranges than females (with one exception -Hasegawa 1995). Mitchell (1990) observed that home range size of males was greater during the breeding season, indicating home range size of males was related to breeding requirements as well as food. Larger home range sizes would permit contact with more potential mates by overlapping more female home ranges (Hindell and Lee 1988). A number of studies have also identified age-related differences in home range distribution, with sub-adult males found in suboptimal habitats (Gordon et al. 1990a, Mitchell and Martin 1990). A number of authors have reported interactions between subadult males and dominant males resulting in movement of the subadult males to less favourable habitat (Gordon et al. 1988; Lee and Martin 1988; Mitchell and Martin 1990; Melzer 1995). If, as seems likely, lower availability of food resources in suboptimal habitats would result in the need for larger home range sizes, this would also contribute to the relatively larger average home range sizes of males noted above.
A number of studies have reported high dispersal rates, particularly amongst subadult males (Gordon et al. 1990a and b; Mitchell and Martin 1990). Based on studies of Victorian koala populations, Mitchell and Martin (1990) suggested a model for recruitment to koala populations. They noted that older koalas (>3 years) tended to be sedentary and that at some sites females especially tended to remain for more than 4 years. These older individuals are gradually replaced by younger animals - females deriving predominantly from koalas weaned in the area and males predominantly from koalas immigrating from outside the site. Observations of northern koala populations showing high rates of dispersal of subadult males and the existence of a small group of long-term resident males suggest that this model may apply more universally (Gordon et al. 1990b). In northern NSW, Gall (1980) reported the presence of a pool of "nomadic" males. Both Gordon et al. and Melzer (1995) observed young males
27

entering their study sites and establishing permanent home ranges. Gordon et al. note
that duration of residence in an area and social dominance in males appears to be
related. Social dominance enabled immigrant males to occupy the optimal habitat that
conferred benefits in greater food resources and, perhaps, opportunities to mate.
Further, very few males appeared to remain in residence for long, suggesting that few
reached dominant status. The significance of the territorially dominant males may be
in question; genetic work currently underway in the Zoology Department at the
University of Queensland suggests that in one central Queensland koala population at
least, these dominant males may have only a minor role in siring offspring in the local
population. If the research proves correct, then the pool of so called itinerant males in
secondary habitat may be of much greater importance in koala conservation than
previously thought. Dispersing koalas cross open land, including both disturbed environments such as
cleared paddocks, partially cleared land with an occasional tree (White 1994; Prevett
1996; Melzer and Lamb 1996) and naturally bare environments such as saltflats
(Hasegawa 1995). Hasegawa also observed dispersing koalas using a riparian corridor.
In contrast, White (1994) suggested that koalas did not follow corridors but tended to
favour a direct line of travel crossing pastures and cultivation. Similarly, Prevett
( 1996) found little evidence to suggest that koalas used continuous corridors to move
between remnant vegetation. Instead, dispersing koalas appeared to maintain direction
moving across corridors. Distances moved by dispersing koalas include 0.4 to 2km reported by Gordon et al.
(1990b) in southern Queensland, 0.7 to 8.7km by Mitchell and Martin (1990) in
southern Victoria, and 22 to 48km in South Australia (Robinson 1978). Maximum
dispersal distances include 2km in three weeks (Hasegawa 1995), 1 Okm in 4 months
and 21km in 6 months (Melzer 1995), 29km in 12 months (Robinson 1978) to 50km
over 2.5 years in south-east Queensland (Samedi 1995).
Isolation as a result of habitat fragmentation has been reported to prevent dispersal
in Victorian populations. This has been implicated in causing over-population
(apparently the animals do not disperse) and consequential over browsing of food
resources and subsequent population declines (Martin 1985a). It seems, however, that
koalas will attempt to cross most environments. Presumably successful dispersal is
determined by the availability of trees for refuge and food. The appropriate density
and arrangement of such trees within the landscape to allow successful dispersal is not
known. Some information may be inferred from data on overnight distances moved by
koalas. Interpreting patterns of habitat utilisation is complicated by differences in behaviour
among individuals and between genders (Hindell and Lee 1987) and if, as is expected,
unrecognised regional variants exist within koala populations, then patterns of habitat
utilisation would be expected to vary with locally adapted physiological and
behavioural phenology.
Habitat assessment and models Studies of tree preferences (both roost and food) have shown that koalas have
complex habitat requirements that will not be met simply by conserving a few
varieties of food trees (Cork et al. 1990). Their social and roosting requirements must
also be considered, along with the need for dispersing koalas to have access to
suboptimal habitats. Thus a habitat approach is required if koala conservation is to be
successful.
28

Studies in central Queensland have shown that habitats with high densities of food trees are not necessarily "good" koala habitat (Melzer 1995). Rather it is the relative density of favoured trees, in combination with other favoured habitat characteristics such as leaf moisture levels, which appears to be important at the local level. Leaf moisture is related to proximity to creeks, and a significant relationship was found between koala population density and proximity to creeks (Munks et al. 1996). Leaf nitrogen levels were not important in habitat selection in central Queensland at the local level (Melzer 1995; Munks et al. 1996). Melzer found that foliar nitrogen levels exceeded minimum threshold levels for fodder suitability (one percent nitrogen -Cork and Sanson 1990) for all vegetation types within his study area.
Melzer (1995) developed a simple model to explain local changes in habitat use at his study site in central Queensland. In this model, fodder tree preferences and hence habitat preferences were mediated by leaf moisture levels which varied depending on the season. This model was never validated and its applicability in southern Australian populations has not been tested.
Samedi (1995) developed a local model based on the variables; aspect, slope, altitude, rainfall and proximity to creeks, to predict the occurrence and distribution of suitable koala habitat on forested ranges near Brisbane. The model was able to classify correctly more than 80 percent of known prime koala habitat, whilst including about 50 percent of secondary habitat. Nevertheless, it undervalued, as secondary habitat, flat lowlands to the east of the site known to be prime koala habitat. This was a consequence of the model using slope and aspect as the most significant discriminators.
At the regional level other factors become important, such as differences in soil nutrient levels through their effect on leaf nutrient levels (Cork et al. 1990). A number of studies have reported an association with watercourses and more fertile soils (Hindell 1984; Gordon et al. 1990a; Monks et al. 1996; Phillips 1996). Norton and Neave (1996) found that in a systematic survey, 90 percent of sites were on land of less than 20 degree slopes and underlain by fertile soils (i.e. basalt or alluvium) and 70 percent were in the lower half of catchments.
Cork et al. (1990) suggested that a mosaic of vegetation types was also a prerequisite for prime koala habitat. They believe that such a mosaic would ensure that at least some food tree species were growing and providing high quality food resources for a greater proportion of the year. Other authors have suggested that habitat heterogeneity may be promoted by a mixture of young and old trees (Lunney et a/.1996), presumably within a single habitat. The presence of a mosaic of habitat types has a complementary function in that it provides a diversity of habitat types capable of providing suboptimal habitats for dispersing subadult and subdominant individuals (Munks 1993; Melzer 1995). Such a model may explain low koala densities in association with seemingly favourable vegetation types, but of low habitat heterogeneity.
Cork et al. (1990) suggest topography as the driving force behind habitat heterogeneity, but other factors such as soil type may similarly promote small scale differences in vegetation communities and hence habitat complexity. Gordon et al. (1988) observed that habitat heterogeneity was related to "core" and "marginal" habitat along a creek depending on water availability. Fire may also promote heterogeneity (Cork et al. 1990).
Gordon et al. (1988) suggested a model of habitat use for inland Queensland in which koala population size was regulated by rainfall. A rapid increase in numbers
29

was associated with a few good years of rainfall followed by a gradual decline over
poor years in below average rainfall. They noted that poor rainfall years were much
more numerous than good rainfall years. Survival of the population was dependent on
dominant koalas occupying optimal habitats, with high mortality recorded during
drought years by koalas occupying suboptimal or marginal habitats. Melzer (1995)
noted that climate, particularly frequency of drought, may restrict koala distribution
and abundance in Queensland, and suggested that severity of climate may be a more
effective regulator of regional abundance and distribution than reproductive potential.
He contrasted Queensland populations with Victorian populations of koalas that have
been periodically observed to outstrip food resources and thus exceed the carrying
capacity of the local habitat. By inference, Queensland populations are kept below the
carrying capacity as a result of a greater frequency of poor years and availability of
suboptimal or marginal habitats for dispersal. Norton and Neave (1996) have proposed a functional model to assess koala habitat
quality. They suggested four primary criteria on which habitat quality must be based:
• availability of water is within critical limits;
• availability of soil nutrients is above critical limits;
• the disturbance regime is below a critical threshold;
• patch size must be large enough. Such models may need regional calibration as, for example, water availability and
leaf moisture thresholds may differ at a regional level. Melzer (1995) had already
shown a seasonal change in leaf moisture threshold in central Queensland. Jurskis and
Potter (1997) attributed the failure of the Norton and Neave model in south-east New
South Wales, to their prediction that koalas prefer wetter areas. In the Eden area the
wetter areas were closed forests, and in this region koalas preferred the more open
forests, perhaps overriding the moisture relationship.
At this stage, the lack of acceptance of model predictions at large scales and limited
accuracy of models at local scales means that they contribute little to koala
conservation (Cork et al. 2000).
Habitat loss, fragmentation and degradation Habitat loss from clearing and associated habitat fragmentation has been identified
as the greatest threat to successful koala conservation (Phillips 1990: Reed and
Lunney 1990; National Koala Network 1996). Habitat loss is occurring as a
consequence of urban and industrial expansion, creation of road networks and
continued clearing for agricultural purposes (Paterson 1996; Melzer et al. 2000).
In terms of clearing, South Australia and Victoria have suffered the greatest declines
in potential habitat loss of favoured forest types (i.e. medium and tall open forests)
through clearing (ranges of 88-98 percent and 67-77 percent respectively) (Pahl et al.
1990). Clearing has been implicated in range contractions in the western plains and
south-east coast of New South Wales by Reed et al. (1990), who found that in most
areas where koalas were sighted, numbers were low. High numbers of sightings,
indicative of optimal habitats, were restricted to a few localities in north-east New
South Wales. Most of these were near major population centres on the coast,
suggesting that further loss of optimal habitat may occur through urbanisation
(although classification as optimal habitat may also be an artefact of more observers
adjacent to major population centres). Such declines in once abundant urban
associated koala populations have been recorded from the outskirts of Sydney (Reed
et al. 1990; Smith and Smith 1990). Reed et al. (1990) noted that historically high
i. ,: . .'• ./
30

koala populations in the Bega Valley in southern NSW are much reduced and that these areas coincide with high levels of clearing. In Queensland, populations under the greatest threat are in the south-east from urban expansion, and the Brigalow Bioregion from clearing associated with agriculture (Paterson 1996). Comparisons of current distribution patterns with historical patterns suggest the koala still occurs throughout much of its original range, but with some contraction in the north, both along the coast and inland (Paterson 1996). In contrast, a national koala survey in 1986-87 concluded that the koala had disappeared from 50 to 75 percent of its known range in NSW (National Koala Network 1996).
Widespread disease epidemics in koala populations were reported from the late 1800's and early 1900's. Reed and Lunney (1990) attributed the trigger for these epidemics to a dramatic reduction in habitat quality as a result of massive land clearing, combined with a severe drought at the time. In support of this contention, they noted that where habitat fragmentation as a result of clearing was greatest, declines in population were greatest ( eg. western plains of NSW).
Clearly habitat loss impinges on koala survival by reducing potential populationsize. Effects of habitat fragmentation are more esoteric but equally as insidious. Habitat fragmentation impedes the functioning of the available habitat and impairs its ability to support koala populations. Habitat quality deteriorates as a result of small patch size, inappropriate fire regimes, weed invasion, die-back of food trees, soil erosion, construction of roads and stream siltation (Pahl et al. 1990; Paterson 1996). Effects of drought and all causes of natural mortality are exacerbated in fragmented habitats (Reed and Lunney 1990; May and Norton 1996). May and Norton (1996) note that fragmentation results in an increase in the perimeter-to-interior ratio and increases problems associated with edge effects.
Habitat fragmentation may enhance the incidence of dieback in partially cleared habitat. Pahl et al. (1990) note that dieback is more prevalent in disturbed forests or woodlands where greater than 50 percent of the original tree cover has been cleared. They found that all the main food trees of koalas were susceptible to dieback.
Gordon et al. (1990a) note that incidence of cystitis in koalas is likely to be exacerbated by further clearing and habitat fragmentation in south-east Queensland. Factors that reduce patch sizes below threshold levels (supporting less than 50 animals - Smith 1996) may produce genetic bottlenecks from which local populations may not recover (Reed and Lunney 1990). The Victorian experience, however, suggests that, in the short term at least, genetic bottlenecks in koalas have not reduced population vigour.
Disease Jakob Hoff (1993) links a range of diseases with koalas. Infectious diseases include
Chlamydiosis (incorporating keratoconjunctivitis, cystitis, rhinitis/pneumonia and infertility); koala respiratory disease linked to infections of Cryptococcus, Toxoplasma and Bordatella bronchiseptica (a common cause of death in Queensland according to Jakob Hoff). Parasitic disease involves sarcoptic mange caused by Sarcoptes scabiei in free-living koalas. There is a wide range of noninfectious diseases including warts on the face, lips and gums of young koalas; other neoplasms such as serous cystadenomas; adenocarcinoma of the urinogenital tract and lymphocytic leukemia.
31

Chlamydia The most significant diseases of wild koalas are those associated with Chlamydia
spp. (National Koala Network 1996). Chlamydia is associated with a range of
symptoms including eye infections, respiratory and urinary tract infections, and is
capable of causing sterility in female koalas. Chlamydia is considered to be the major
cause of low fertility in koala populations (McColl et al. 1984) and it is this aspect
that is of most concern as it can depress koala reproductive rates (Martin 1981).
Recent studies indicate that two species of this organism (Chlamydia pecorum and C.
pneumoniae) may be involved (Glassick et al. 1995). These authors note that
transmission of Chlamydia between hosts is as yet poorly understood and it seems
likely that if two species are involved that they may have different modes of
transmission. This has important management implications (Sherwin et al. 2000).
There is evidence that Chlamydia is endemic in wild koala populations and it is
found in populations throughout the koala's range (Phillips 1990). Epidemiological
studies in south-east Queensland and Victoria have shown levels of endemicity in
wild koala populations in the order of 70 to 98 percent (McColl et al. 1984; Lee et al.
1988; Wiegler et al. 1988). Symptoms of overt disease were, by comparison,
relatively low with only 9 percent of koalas in south-east Queensland and less than 5
percent in Victoria showing commonly recognised disease symptoms. Similarly, Ellis
et al. (1993) demonstrated that a relatively undisturbed koala population can support a
chlamydial infection without showing overt chlamydia! disease. Wieger et al. noted
that in some cases the organism is present but without any significant pathological
effects. Findings of the National Koala Survey suggest that incidence of koalas
showing overt symptoms of Chlamydia infection (i.e. eye or urinary disorders) is
probably less than 5 percent (Phillips 1990). Fertility rates varied widely (Table IX), probably due to local differences in the level
of reproductive tract disease. Chlamydia-infected populations showed a range in
fertility from 0 to 84 percent and Chlamydia-free populations from 50 to 83 percent.
Chlamydia has been implicated in lowering reproductive rates in some wild koala
populations in Victoria (Martin and Handasyde 1990) and at Oakey in Queensland
(Gordon et al. 1990a). In these populations, low fertility rates were observed in
combination with a relatively high incidence of female sterility as a result of
Chlamydia infection. Gordon et al. found a negative correlation between cystitis
(urinary tract infection by Chlamydia) and both population density and recruitment of
new animals to the population. This indicated that cystitis caused mortality of
breeding females; in turn reduced the pool of potential emigrants, thus reducing
potential breeding still further. They suggested that a combination of sterility lowering
fertility rates and increased mortality due to cystitis could have contributed to a
reduction in population density observed at this site over several years. However, they
went on to say that there was no evidence that survival of the population was
threatened by the disease, and that the population was recovering. Martin and
Handasyde (1990) reached similar conclusions for Victorian populations noting
depressed fertility rates, but suggesting that these rates were high enough to sustain
population growth. Further they suggested that the koala has probably had a long
history of association with Chlamydia.
32
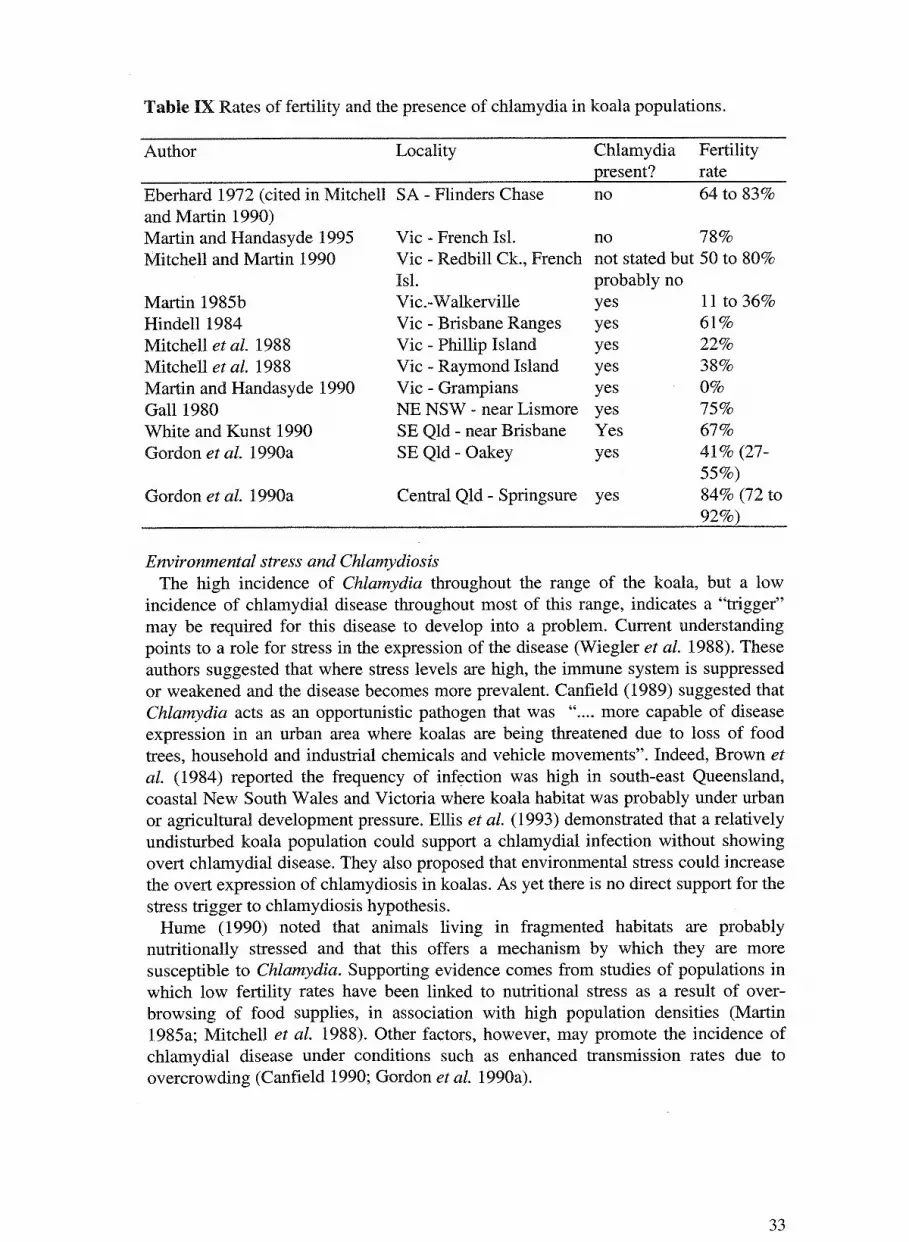
Table IX Rates of fertility and the presence of chlamydia in koala populations.
Author Locality Chlamydia Fertility 12resent? rate
Eberhard 1972 (cited in Mitchell SA - Flinders Chase no 64 to 83% and Martin 1990) Martin and Handasyde 1995 Vic - French Isl. no 78% Mitchell and Martin 1990 Vic - Redbill Ck., French not stated but 50 to 80%
Isl. probably no Martin 1985b Vic.-Walkerville yes 11 to 36% Hindell 1984 Vic - Brisbane Ranges yes 61% Mitchell et al. 1988 Vic - Phillip Island yes 22% Mitchell et al. 1988 Vic - Raymond Island yes 38% Martin and Handasyde 1990 Vic - Grampians yes 0% Gall1980 NE NSW - near Lismore yes 75% White and Kunst 1990 SE Qld - near Brisbane Yes 67% Gordon et al. 1990a SE Qld- Oakey yes 41% (27-
55%) Gordon et al. 1990a Central Qld - Springsure yes 84% (72 to
92%)
Environmental stress and Chlamydiosis The high incidence of Chlamydia throughout the range of the koala, but a low
incidence of chlamydia! disease throughout most of this range, indicates a "trigger" may be required for this disease to develop into a problem. Current understanding points to a role for stress in the expression of the disease (Wiegler et al. 1988). These authors suggested that where stress levels are high, the immune system is suppressed or weakened and the disease becomes more prevalent. Canfield (1989) suggested that Chlamydia acts as an opportunistic pathogen that was " .... more capable of disease expression in an urban area where koalas are being threatened due to loss of food trees, household and industrial chemicals and vehicle movements". Indeed, Brown et al. (1984) reported the frequency of infection was high in south-east Queensland, coastal New South Wales and Victoria where koala habitat was probably under urban or agricultural development pressure. Ellis et al. (1993) demonstrated that a relatively undisturbed koala population could support a chlamydia! infection without showing overt chlamydia! disease. They also proposed that environmental stress could increase the overt expression of chlamydiosis in koalas. As yet there is no direct support for the stress trigger to chlamydiosis hypothesis.
Hume (1990) noted that animals living in fragmented habitats are probably nutritionally stressed and that this offers a mechanism by which they are more susceptible to Chlamydia. Supporting evidence comes from studies of populations in which low fertility rates have been linked to nutritional stress as a result of overbrowsing of food supplies, in association with high population densities (Martin 1985a; Mitchell et al. 1988). Other factors, however, may promote the incidence of chlamydia! disease under conditions such as enhanced transmission rates due to overcrowding (Canfield 1990; Gordon et al. 1990a).
33

Retrovirus infections Canfield et al. (1988) found oncovirus like particles (members of the retrovirus
family) in association with leukemia in koalas. They suggested a link between the
virus and the disease. Later, Worley et al. (1993) linked retrovirus infections to
opportunistic infections, various neoplasms, skull tumours and blood disease in
captive koalas. They suggested that retrovirus infections may have a role to play in the
population dynamics of wild koalas in Australia.
Other mortality factors In most habitats, longevity in koalas is determined ultimately by the rate of tooth
wear. When the teeth wear to a critical point, depending on the nature of the fodder,
the koala is unable to process sufficient food to meet its nutritional needs (Lee and
Martin 1988). Northern Australian populations appear to be less long lived than
southern populations, with Melzer ( 1995) finding a 10 year maximum lifespan in a
relatively disease-free population in inland central Queensland. This contrasts with a
suggested lifespan of more than 10 but less than 20 years for free ranging populations-
in Victoria (Eberhard 1978). Martin and Handasyde (1990) cite ages of between 11
and 17.5 years. Such differences in lifespan, if confirmed, may have significant
implications for differences in demographic parameters such as underlying capacities
of increase between southern and northern koala populations.
Fire, drought and disease are generally considered to be major causes of natural
mortality (Lee and Martin 1988; Gordon et al. 1990a). Seasonally, winter is
considered to stress nutritional reserves and mortality tends to peak during this period
(Butler 1978; Martin 1985b; Lee and Martin 1988; White and Kunst 1990; Melzer
1995). Melzer observed that high mortalities coincided with an unusually wet and
windy autumn-winter period. Butler (1978) suggested a second peak in mortality
might occur in late spring associated with the onset of breeding activity and an
upsurge in reproductive tract pathology. In urban areas, an upsurge in mortalities has
been observed in late spring coinciding with increased movements associated with
breeding (Moon 1995; Nattras and Fiedler 1996).
The effects of drought have been documented in a population in western Queensland
(Gordon et al. 1988). High mortality was associated with a period of extended low
rainfall and very high temperatures. Leaf loss by preferred Eucalyptus species was
extensive and high mortality of koalas was observed. Dominant koalas in the optimal
habitat survived whereas individuals in suboptimal habitats did not. Such mass
mortality events appear to be the exception and were linked to extensive defoliation
under extreme conditions. Other natural agents of mortality include predation and fire. Natural predators
include dingoes and raptors such as the powerful owl and wedge-tailed eagle (Phillips
1990). Several species of ticks have also been implicated in causing occasional koala
deaths either due to anaemia or toxicosis (Stone and Carrick 1996). A number of
studies report koala deaths in bushfires (Tilley and Uebel 1990; Lunney et al. 1995;
National Koala Network 1996) but few data are available on actual mortality levels.
An exception to this was Lunney et al. (1995), who found that out of a population of
134 individuals at Port Stephens in NSW, 37 percent died in a bushfire. Tilley and
Uebel (1995) noted that populations rapidly recovered to pre-fire levels.
The main agents of mortality of koalas near urban areas are vehicles, disease and
dogs (Lee and Martin 1988; Lunney et al. 1996). Lee and Martin (1988) reported that
60 percent of koala mortality on Phillip Island was from road kills. At Coffs Harbour
34

on the north coast of NSW, Moon (1995) found that road kills accounted for 57 percent of deaths, disease 20 percent and dogs 4 percent. White (1994) reported that disease and dogs contributed 33 percent and 5 percent of mortality respectively of a koala population in southern Queensland. Interestingly, he noted that four cases of mortality were a result of accidental falls from trees (19 percent) and one case of a fall due to clearing. In a suburb of Sydney, Smith and Smith (1990) thought that dogs were the most important mortality factor while in Ballarat both cars and dogs were considered to be most important (Prevett 1996).
Prevett et al. (1995) found that road kills occurred where vehicle speeds exceeded 80kph and where wide habitat corridors or linear forests occurred on both sides of the road. At one site they found that koalas established horne ranges that straddled the highway. Road kills at the site were related to the frequency of road crossings by the koalas and individuals were unlikely to survive more than 24 days. They hypothesised that the koalas were immigrants from adjacent populations and that a 'source' and 'sink' model was operating at the site. They further concluded that narrower roads increased koala mortality and that traffic volume, speed and visibility influenced the collision rate.
Reduction of genetic heterogeneity Below a minimum number of individuals the loss of genetic variability and
inbreeding in a population may become a problem for its long-term survival. This minimum number has to be large enough to cope with natural and chance fluctuations in births, deaths and environmental conditions, as well as random changes in genetic composition. The minimum number needed to avoid inbreeding depression is 50. Below this the population has a high chance of becoming extinct as a result of random environmental factors (Smith 1996). Pahl et al. (1990) has used a population size of 500 in planning habitat conservation measures for the koala. Taylor et al. (1991) and Houlden et al. (1996) both identified low levels of genetic diversity in the koalas of Victoria and South Australia. This was attributed to the near extinction and consequent severe genetic bottleneck experienced by the southern koala which only survived extinction through widespread reintroduction of koalas derived from a few animals introduced onto the islands of Western Port Bay near Melbourne. Although this founder population was well below 50 and current genetic diversity is low, koalas in Victoria and South Australia appear vigorous and breed readily where appropriate habitat exists. In the short term at least, low genetic diversity does not seem to be a problem for P.c. victor. Worthington et al. (1993) identified moderate to high levels of inbreeding in a population of captive koalas. Despite this, the only evidence for inbreeding depression was a male biased sex ratio. Although the limited data precluded any firm conclusions, Worthington et al. (1993) hypothesised that koalas have a reduced susceptibility to inbreeding depression as a consequence of their history of small, periodically isolated regional populations. Indeed, Smith (1996) sounds a note of caution. He points out that the minimum viable population number of 50 and 500 is only a general guide in genetic management and "should never be used as a precise rule in deciding a species' fate".
Conclusion Conservation of an animal requires some understanding of its biology and ecology.
Some basic research into aspects of the koala is still lacking (Lunney et al. 1990) in part, reflecting the wide distribution of the species and the contrasting environments
35

occupied by koala populations. In the' light of the failure of community concern to
limit broad acre clearing and the consequential loss of koala habitat, the most pressing
research needs are related to understanding the capacity of the koala to persist within
fragmented landscapes. There is sufficient knowledge, however, for regional conservation strategies to be
developed. The major factors limiting koala populations are known. Fodder species
are documented or are easily determined through dietary studies. The key elements of
koala habitat structure have been described and the principles behind habitat
utilisation are known. The regional conservation strategies should draw on existing
knowledge to: 1. Preserve existing populations and habitats,
2. Re-establish former habitat as part of property planning on participating
properties, 3. Re-introduce populations using koalas with appropriate genotypes, and
4. Develop dispersal environments between habitat fragments.
These conservation strategies should be applied through local action plans that are
linked to LandCare groups and regional communities.
The current understanding of koala ecology allows the statement of a set of general
principles for reconstructing koala habitat and populations. These are:
1. Remnant koala populations, collectively, will form a meta-population with
varying degrees of habitat fragmentation. 2. Reconstructed habitat should contain a relatively high density of at least two
most preferred fodder species; at least some of these trees should have a girth
greater than 30 em at breast height; a range of non-fodder trees and shrubs for
use as day roost trees; landscape complexity and species diversity (for example
ridges supporting E. crebra and valley floors supporting E. terticomis and/or
E. populnea) allowing seasonal, climatic and social variation in tree and
habitat use; a persistent aquifer, and a disperal environment allowing
emigration. 3. Foundation koalas should be drawn from regional genetic management units
and probably from those populations supporting types of Chlamydia
percorum. 4. Sighting of reconstructed koala populations should take account of the major
limiting factors in each region (for example environmental water and moisture
status in central and western Queensland and vehicles and dogs in south-east
Queensland). 5. Restoration programs should take the form of regional strategies and local
action plans, involve local landowners and be multidisciplinary in approach.
References Baker, R. H. (1968). Habitats and distribution. In Biology of Peromyscus (Rodentia.)
(Ed. J. A. King.) pp. 8-126. American Society ofMammalogists, Special
Publication No.2. Bell, R. H. V. (1971). A grazing ecosystem in the Serengeti. Scientific American 225
(1), 86-93. Betts, T. J. (1978). Koala acceptance of Eucalyptus globulus labill. as food in relation
to the proportion of sesquiterpenoids in the leaves. In 'The koala- proceedings of
the Taronga symposium'. (Ed. T. J. Bergin.) pp.75-85. (Zoological Board of
N.S.W.: Sydney.)
36

Brown, A. S., and Carrick, F. N. (1985). Koala disease breakthrough. Australian Natural History 21,314-317.
Brown, A. S., Carrick, F. N., Gordon, G., and Reynolds, K. (1984). The diagnosis and epidemiology of an infertility disease in the female koala, Phascolarctos cinereus (Marsupialia). Veterinary Radiologist 25, 242-248.
Brown, S., and Woolcock, J. (1990). Strategies for the control and prevention of chlamydia! disease in captive koalas. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 295-298. (Surrey Beatty and Sons: NSW.)
Bryant, J.P., Reichardt, P. B., Clausen, T. P., Provenza, F. D. and Kuropat, P. J. (1992). Woody plant- mammal interactions. In 'Herbivores- their interactions with secondary plant metabolites: Ecological and evolutionary processes'. Vol. II. (Eds. G. A. Rosenthal and M. R. Berenbaum.) pp. 344-370. (Academic Press, Inc.: Sydney.)
Butler, R. (1978). Patterns in koala mortality. In 'The koala'. (Ed. T. J. Berjin.) The Zoological Parks Board of New South Wales.
Canfield, P. J. (1989). A survey of urinary tract disease in New South Wales (Australia) koalas. Australian Veterinary Journal66, 103-106.
Canfield, P. J. (1990). Diseases affecting captive and free-living koalas and their implications for management. In 'Koala Summit: managing koalas in New South Wales'. (Eds D. Lunney, C. Urqhart and P. Reed.) pp. 36-38. (NSW National Parks and Wildlife.)
Canfield, P. J., Sabine, J. M., and Love, D. N. (1988). Virus particles associated with leukaemia in a koala. Australian Veterinary Journal65, 327-328.
Congreve, P. and Betts, T. J. (1978). Eucalyptus plantations and preferences as food for a colony of koalas in Western Australia. In 'The koala -proceedings of the Taronga symposium'. (Ed. T. J. Bergin.) pp. 97-105. (Zoological Parks Board of N.S.W.: Sydney.)
Cork, S. J. (1984). Utilisation of Eucalyptus foliage by arboreal marsupials. Proceedings ofthe Nutritional Society of Australia 9, 88-97.
Cork, S. J. (1987). Form and Function in the koala. In 'Koala, Australia's endearing marsupial'. (Ed. L. Cronin.) pp. 8-30. (Reed Books Pty Ltd: N.S.W.)
Cork, S. J. (1992). Polyphenols and the distribution of arboreal, folivorous marsupials in Eucalyptus forests of Australia. In -'Plant Polyphenols'. (Eds. R. W. Hemingway and P. E. Laks.) pp. 653-663. (Plenum Press: New York.)
Cork, S. J., Hume, I. D., and Foley, W. J. (2000). Improving habitat models and their utility in koala conservation. Conservation Biology 14 (3), 660-668.
Cork, S. J., Margules, C. R., and Braithwaite L.W. (1988). 'A survey of koalas and their habitat near Wedderburn N.S.W., suggestions for management and an assessment of the potential effects of a proposed subdivision of four-hectare environmental lots'. Report for Campbelltown City Council.
Cork, S. J., and Sanson, G. D. (1990). Digestion and nutrition in the koala: a review. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 129-144. (Surrey Beatty and Sons: NSW.)
Degabriele, R. (1981). A relative shortage of nitrogenous food in the ecology of the koala (Phascolarctos cinereus). Australian Journal of Ecology 6, 139-141.
Degabriele, R. (1983). Nitrogen and the koala, (Phascolarctos cinereus): some indirect evidence. Australian Journal of Ecology 8, 75-76.
Delbridge, A., Bernard, J. R. L., Blair, D., Ramson, W. S., and Butler, S. (1981). 'The Macquarie Dictionary'. (Macquarie Library Pty Ltd: NSW.)
37

Demment, M. W., and Van Soest, P. J. (1985). A nutritional explanation for body-size
patterns of ruminant and non-ruminant herbivores. American Naturalist 125, 641-
672. Eberhard, I. H. (1978). Ecology of the koala, Phascolarctos cinereus (Goldfuss)
Marsupialia: Phascolarctidae, in Australia. In 'The ecology of arboreal folivores'.
(Ed. G. G. Montgomery.) pp. 315-328. (Smithsonian Institution Press: Washington,
D.C.) Eberhard, I. H., McNamara, J., Pearse, R. J., and Southwell, I. A. (1975). Ingestion
and excretion of Eucalyptus punctata D.C. and its essential oil by the koala,
Phascolarctos cinereus (Goldfuss). Australian Journal of Zoology 23, 169-179.
Ellis, W. A. H., Girjes, A. A., Carrick, F. N., and Melzer, A. (1993). Chlamydia!
infection in koalas under relatively little alienation pressure. Aust. Vet. J. 70, 427-
428. Ellis, W. A H., Melzer, A, Green, B., Newgrain, K., Hindell, M. A., and Carrick, F. N.
(1995). Seasonal variation in water flux, field metabolic rate and food consumption
of free-ranging koalas (Phascolarctos cine reus). Australian Journal of Zoology 43,
59-68. Ellis, W. A. H., Sullivan, B. J., Lisle, A. T., and Carrick, R.N. (1998). The spatial and
temporal distribution of koala faecal pellets. Wildlife Research 25 (6), 663-668.
Faulks, J. (1990). 'The ranging behaviour of koalas'. Honours thesis, University of
New England, Northern Rivers. Finlayson, H. H. (1934). On mammals from the Dawson and Fitzroy Valleys: Central
coast Queensland. Part II. Transactions of the Royal Society of South Australia
55, 218-231. Fleay, D. (1937). Observations ofthe koala in captivity. Successful breeding in
Melbourne Zoo. Australian Zoologist 9 (1), 68-80.
Freeland, W. J., and Janzen, D. H. (1974). Strategies in herbivory by mammals: the
role of plant secondary compounds. American Naturalist 108, 269-289.
Fretwell, S.D., and Lucas, H. L. (1970). On territorial behaviour and other factors
influencing habitat distribution in birds. I. Theoretical development. Acta
Biotheoretica 19, 16-36. Gall, B. C. (1980). Aspects of the ecology of the koala, Phascolarctos cinereus
(Goldfuss), in Tucki Tucki Nature Reserve, New South Wales. Australian Wildlife
Research 7, 167-176. Glassick, T., Giffard, P., and Timms, P. (1995). Chlamydia! infections in koalas: gene
sequencing indicates that two separate chlamydia! species are involved, Chlamydia
pecorum and Chlamydia pneumoniae. In 'Proceedings on a Conference on the
Status of the Koala in 1995'. pp. 154-160. (Australian Koala Foundation:
Brisbane.) Gordon, G., Brown, A. S., and Pulsford, P. (1988). A koala population crash during
drought and heatwave conditions in south-western Queensland. Australian Journal
of Ecology 13, 451-461. Gordon, G., McGreevy, D. G., and Lawrie, B. C. (1990a). Koala population turnover
and male social organisation. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A.
Handasyde and G. D. Sanson.) pp. 189-192. (Surrey Beatty and Sons: NSW.)
Gordon, G., McGreevy, D. G., and Lawrie, B. C. (1990b). Koala populations in
Queensland: major limiting factors. In 'Biology of the Koala'. (Eds. A. K. Lee, K.
A. Handasyde and G. D. Sanson.) pp. 85-95. (Surrey Beatty and Sons: NSW.)
Gould, J. (1863). 'The mammals of Australia'. Taylor and Francis, London.
38

Hasegawa, M. (1995). 'Habitat utilisation by koalas (Phascolarctos cinereus) at Point Halloran, Queensland'. Msc thesis, University of Queensland, St. Lucia, Brisbane.
Hindell, M.A. (1984). 'The feeding ecology of the koala, Phascolarctos cinereus, in a mixed Eucalyptus forest'. Masters Thesis, Department of Zoology, Monash University.
Hindell, M.A., and Lee, A. K. (1987). Habitat use and tree preferences of koalas in a mixed eucalypt forest. Australian Wildlife Research 14, 349-360.
Hindell, M.A., and Lee, A. K. (1988). Tree use by individual koalas in a natural forest. Australian Wildlife Research 15, 1-7.
Hindell, M. A., and Lee, A. K. (1990). Tree preferences of the koalas. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde, and G. D. Sanson.) pp. 117-121. (Surrey Beatty and Sons: N.S.W.)
Hindell, M.A., Handasyde, K. A., and Lee, A. K. (1985). Tree species selection by free-ranging koala populations in Victoria. Australian Wildlife Research 12, 137-144.
Houlden, B. A., Costello, B. H., Sharkey, D., Fowler, E. V., Melzer, A., Ellis, W., Carrick, F., Baverstock, P.R., and Elphinstone, M.S. (1999). Phylogeographic differentiation in the mitochondrial control region in the koala. Phascolarctos cinereus (Goldfuss, 1817). Molecular Ecology 8, 999-1011.
Houlden, B. A., England, P. R., Taylor, A. C., Greville, W. D., and Sherwin, W. B. ( 1996). Low genetic variability of the koala Phascolarctos cine reus in southeastern Australia following a severe population bottleneck. Molecular Ecology 5 (2), 269-281.
Hume, I. D. (1990). Biological basis for the vulnerability of koalas to habitat fragmentation. In 'Koala Summit: managing koalas in New South Wales'. (Eds. D. Lunney, C. Urqhart and P. Reed.) pp. 33-35 NSW National Parks and Wildlife.
Hume, I. D., and Esson, C. (1993). Nutrients, antinutrients and leaf selection by captive koalas (Phascolarctos cinereus). Australian Journal of Zoology 41, 379-392.
Ingram, G. J., and Raven, R. J. (1991). Introduction. In 'An atlas of Queensland's frogs, reptiles, birds and Mammals'. (Eds. G. J. Ingram and R. J. Raven.) pp. 5-14. (Board of Trustees, Queensland Museum: Brisbane.)
Jakob Hoff, R. M. (1993). Diseases of free-living marsupials. In 'Zoo and wild animal medicine: current therapy'. (Ed. M. E. Fowler.) pp. 276-281. (W.B. Saunders Company: Philadelphia.)
Jurskis, V., and Shields, J. (1995). Koalas in south-east New South Wales: Habitat assessment and management across a diverse landscape. In 'Proceedings on a Conference on the Status of the Koala in 1995'. pp. 21-42. (Australian Koala Foundation: Brisbane.)
Jurskis, V., and Potter, M. (1997). 'Koala surveys, ecology and conservation at Eden'. (Forest Research and Development Division, State Forests of New South Wales: Sydney.)
Kerle, J. A. (1984). Variation in the ecology of Trichosurus: Its adaptive significance. In 'Possums and Gliders'. (Eds. A. Smith and I. Hume.) pp. 115-128. (Surrey Beatty and Sons: NSW.)
Lawler, I. R., and Foley, W. J. (2000). Chemical ecology of Eucalyptus essential oils in plant-herbivore interactions. In press. In 'Medicinal and Aromatic Plants -Industrial Profiles: Eucalyptus'. (Ed. J. Coppen. Harwood.) (Academic Press: Victoria.)
39

Lawler, I .R., Foley, W. J., Eschler, B., Pass, D. M., and Handasyde, K. (1998).
Intraspecific variation in secondary metabolites determines food intake by
folivorous marsupials. Oecologia 116, 160-169.
Lee, A., and Martin, R. (1988). 'The Koala- a natural history'. (New South Wales
University Press: NSW.) Lee, A. K., and Carrick, F. N. (1987). Phascolarctidae. In 'Mammalia, Vol. 1B, Fauna
of Australia'. (Eds. D. W. Walton and Richardson, B. J.) pp. 740-754. (Australian
Government Publishing Service: Canberra.)
Lee, A. K., Martin, R. W., and Handasyde, K. A. (1988). Research into Chlamydiosis
in natural populations of the koala in Victoria, Phases 1-3. Report to Australian
National Parks and Wildlife Service.
Lunney, D., Curtin, A., and Matthews, A. (1995). The status of koalas in NSW. In
'Proceedings on a Conference on the Status of the Koala in 1995'. pp. 73-78.
(Australian Koala Foundation: Brisbane.) Lunney, D., Esson, C., Moon, C., Ellis, M., and Matthews, A. (1997). A community
based survey of the koala, Phascolarctos cinereus, in the Eden Region of South
eastern New South Wales. Wildlife Research 24, 111-128.
Lunney, D., Moon, C., and Matthews, A. (1996). A 1990 community based survey of
the koala Phascolarctos cinereus population at Iluka in northern New South Wales.
In 'Koalas- Research for Management. Proceedings of the Brisbane Koala
Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 102-122. (World
Koala Research Inc: Brisbane.) Lunney, D., Urquhart, C. A., and Reed, P. (1990). 'Koala summit- Managing koalas
in New South Wales'. NSW National Parks and Wildlife Service, NSW.
Martin, R. W. (1981). Age-specific fertility in three populations of the koala,
Phascolarctos cinereus (Goldfuss), in Victoria. Australian Wildlife Research 8,
275-283. Martin, R. W. (1985a). Overbrowsing, and decline of a population of the koala,
Phascolarctos cinereus, in Victoria. ll. Population condition. Australian Wildlife
Research 12, 367-375. Martin, R. W. ( 1985b ). Overbrowsing, and decline of a population of the koala,
Phascolarctos cinereus, in Victoria. ill Population dynamics. Australian Wildlife
Research 12, 377-386. Martin, R., and Handasyde, K. A. (1990). Population dynamics of the koala
(Phascolarctos cinereus) in south-eastern Australia. In 'Biology of the Koala'.
(Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp.75-84. (Surrey Beatty and
Sons: NSW.) Martin, R. W., and Handasyde, K. A. (1995). Koala Family Phascolarctidae. In 'The
mammals of Australia'. (Ed. R. Strahan.) pp. 195-198. (Reed Books: Chatswood,
New South Wales.) Maxwell, S., Burbidge, A. A., and Morris, K. (1996). 'The 1996 action plan for
Australian marsupials and monotremes'. For the IUCN/SSC Australasian
Marsupial and Monotreme Specialist Group. Wildlife Australia Endangered
Species Program, Project Number 500. (Environment Australia: Canberra.)
May, S. A., and Norton, T. W. (1996). Influence of fragmentation and disturbance on
the potential impact of feral predators on native fauna in Australian forest
ecosystems. Wildlife Research 23, 387-400.
40

McColl, K. A., Martin, R. W., Gleeson, L. J., Handasyde, K. A., and Lee, A. K. (1984). Chlamydia infection and infertility in the female koala (Phascolarctos cinereus). Veterinary Research 115, 655.
McKay, G. M. (1995). Greater Glider. In 'The mammals of Australia'. (Ed. R. Strahan.) pp. 240-241. (Reed Books: Chatswood, NSW.)
McKay, G. M. (1988). Phascolarctidae. In 'Mammalia', Vol. 5 (Ed. D. W. Walton.) pp. 51-52. Zoological Catalogue of Australia. (Bureau of Flora and Fauna: Canberra.)
McNab, B. K. (1978). Energetics of arboreal folivores: physiological problems and ecological consequences of feeding on an ubiquitous food supply. In 'The Ecology of Arboreal Folivores'. (Ed. G. G. Montgomery.) pp.l53-162. (Smithsonian Institution Press: Washington, D.C.)
Melzer, A. (1995). Aspects of the ecology of the koala, Phascolarctos cinereus (Goldfuss, 1817), in the sub-humid woodlands of Central Queensland. PhD thesis, University of Queensland, St Lucia, Brisbane.
Melzer, A. (1996). 'Assessment of the significance of a proposed 4 ha clearing on habitat utilisation by a population of koalas at the Coolangatta Tweed Golf Club'. A report to AGC Woodward-Clyde Pty Ltd. Koala Research Centre of Central Queensland, Central Queensland University, Rockhampton, Qld.
Melzer, A., Carrick, F., Menkhorst, P., Lunney, D., and StJohn, B. (2000). Overview, critical assessment, and conservation implications of koala distribution and abundance. Conservation Biology 14 (3), 619-628.
Melzer, A., and Lamb, D. (1994). Low density populations of the koala (Phascolarctos cinereus) in Central Queensland. Proceedings of the Royal Society of Queensland 104, 89-93.
Melzer, A., and Lamb, D. (1996). Habitat utilisation by a central Queensland koala colony. In 'Koalas- Research for Management. Proceedings of the Brisbane Koala Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 17-22. (World Koala Research Inc: Brisbane.)
Melzer, A., MacLennan, D., and Lamb, D. (1995). Twenty four hour activity and roost tree use: a guide to fodder selection by Central Queensland Koalas. In 'Proceedings of a conference on the status of the koala in 1995'. pp. 138-144. (Australian Koala Foundation: Brisbane, Qld.)
Menkhorst, P. W. (1995). 'Mammals of Victoria: Distribution, ecology and conservation'. (Oxford University Press.)
Milton, K. (1981). Food choice and digestive strategies oftwo sympatric primate species. American Naturalist 117, 496-505.
Mitchell, P. (1990). The home ranges and social activity of koalas - a quantitative analysis. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 171-187. (Surrey Beatty and Sons: Sydney.)
Mitchell, P. J., Bilney, R., and Martin, R. W. (1988). Population structure and reproductive status of koalas on Raymond Island, Victoria. Australian Wildlife Research 15, 511-514.
Mitchell, P., and Martin, R. (1990). The structure and dynamics of koala populations -French Island in perspective. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp.97-108. (Surrey Beatty and Sons: Sydney.)
Montgomery, G. G. (1978). The ecology of arborealfolivores. (Smithsonian Institution Press: Washington, D.C.)
41

Moon, C. (1995). W.I.R.E.S. koala statistics tell a story. In 'Proceedings on a
Conference on the Status of the Koala in 1995'. pp. 49-54. (Australian Koala
Foundation: Brisbane.) Munks, S. A. (1993). 'Arboreal mammals in the Prairie-Torrens Creek Alluvials
Province of the Desert Uplands: Distribution and Habitat Preference'. Report to the
National Estate Program. Munks, S. A., Corkrey, R., and Foley, W. J. (1996). Characteristics of arboreal
marsupial habitat in the semi-arid woodlands of northern Queensland. Wildlife
Research 23, 185-195. National Koala Network. (1996). 'Draft National Koala Conservation Strategy'.
Australian and New Zealand Environment and Conservation Council, Wildlife
Australia Branch Biodiversity Group, Canberra, ACT.
Nattras, A. E. 0., and Fiedler, K. B. (1996). Koala rescue- the perception and reality.
In 'Koalas -Research for Management. Proceedings of the Brisbane Koala
Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 129-136. (World
Koala Research Inc: Brisbane.)
Norton, T. W., and Neave, H. M. (1996). Koala habitat: a conceptual functional
model. In 'Koalas -Research for Management. Proceedings of the Brisbane Koala
Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 93-101. (World
Koala Research Inc: Brisbane.)
Osawa, R. (1993). Dietary preference of koalas, Phascolarctos cinereus (Marsupialia:
Phascolarctidae) for Eucalyptus spp. with a specific reference to their simple sugar
content. Australian Mammalogy 16 ( 1 ), 85-87.
Pahl, L. (1984). Diet preference, diet composition and population density of the
ringtail possum (Pseudocheirus peregrinus cooki) in several plant communities in
southern Victoria. In 'Possums and Gliders'. (Eds. A. Smith, and I. Hume.) pp.
252-260. (Surrey Beatty and Sons: NSW.)
Pahl, L. (1996). Koala and bushland survey of west and central Logan City. In
'Koalas: research for Management'. Proceedings of the Brisbane Koala
Symposium, 22nd- 23rd September 1990. (Ed. G. Gordon.) pp.82-92. (World
Koala Research Inc: Brisbane, Qld.)
Pahl, L., and Hume, I. D. (1990). Preferences for Eucalyptus species of the New
England Tablelands and initial development of an artificial diet for koalas. In
'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G.D. Sanson.)
pp.123-128. (Surrey Beatty and Sons: Sydney.)
Pahl, L., Wylie, F. R., and Fisher, R. (1990). Koala population decline associated with
loss of habitat and suggested remedial strategies. In 'Koala Summit: managing
koalas in New South Wales'. (Eds. D. Lunney, C. Urqhart and P. Reed). pp. 39-47.
NSW National Parks and Wildlife.
Parra, R. (1978). Comparison of foregut and hindgut fermentation in herbivores. In
'The ecology of arboreal Folivores'. (Ed. G. G. Montgomery.) pp. 205-230.
(Smithsonian Institution Press: Washington, D.C.)
Parris, H. S. (1948). Koalas on the lower Goulburn. Victorian Naturalist 64, 192-193.
Paterson, R. (1996). The distribution ofkoa1as in Queensland, 1986-1989. In 'Koalas
- Research for Management. Proceedings of the Brisbane Koala Symposium,
22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 75-81. (World Koala Research
Inc: Brisbane.)
42

Phillips, B. (1990). 'Koalas- The little Australians we'd all hate to lose'. Australian National Parks and Wildlife Service. (Australian Government Publishing Service: Canberra.)
Phillips, S. (1995). 'An assessment of koalas and their habitat in the Goonengerry State Forest and adjoining freehold lands'. A report for State Forests of NSW. (The Australian Koala Foundation: Brisbane, Qld.)
Phillips, S., and Callaghan, J. (1995). 'Koala management study for Goonellabah and East Lismore'. (Draft report for public discussion). Prepared for Lismore City Council. (The Australian Koala Foundation: Brisbane, Qld.)
Phillips, S., Callaghan, J., and Thompson, V. (2000). The tree species preferences of koalas (Phascolarctos cinereus) inhabiting forest and woodland communities on Quaternary deposits in the Port Stephens area, New South Wales. Wildlife Research 27(1), 1-14.
Phillips, W. (1996). Koala conservation a federal perspective. In 'Koalas- Research for Management. Proceedings of the Brisbane Koala Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 70-74. (World Koala Research Inc: Brisbane.)
Pratt, A. (1937). 'The call of the koala'. (Robertson and Mullens: Melbourne.) Prevett, P. ( 1996). Biology and management of an urban population of koalas at
Ballarat in Victoria. In 'Koalas -Research for Management. Proceedings of the Brisbane Koala Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 137-147. (World Koala Research Inc: Brisbane.)
Prevett, P., Smithyman, S., Pope, R. and Schlagloth, R. (1995). Interaction between koalas and roads on the Midlands Highway and Western Freeway in Victoria. Report to the Australian Koala Foundation, Brisbane. University of Ballarat, Victoria.
Reed, P. C., and Lunney, D. (1990). Habitat loss: the key problem for the long-term survival of koalas in New South Wales. In 'Koala Summit: managing koalas in New South Wales'. (Eds. D. Lunney, C. Urqhart and P. Reed.) pp. 9-31. NSW National Parks and Wildlife.
Reed, P. C., Lunney, D., and Walker, P. (1990). A 1986-1987 survey ofthe koala Phascolarctos cinereus (Goldfuss) in New South Wales and an ecological interpretation of its distribution. In Biology of the Koala. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 55-74. (Surrey Beatty and Sons: NSW.)
Robbins, M., and Russell, E. (1978). Observations on movements and feeding activity of the koala in a semi-natural situation. In 'The Koala- Proceedings of the Taronga symposium'. (Ed. T. J. Bergin.) pp.29-44. The Zoological Parks Board of N.S.W., Sydney.
Robinson, A. C. (1978). The koala in South Australia. In 'The Koala- Proceedings of the Taronga symposium'. (Ed. T. J. Bergin.) pp.132-143. The Zoological Parks Board ofN.S.W., Sydney.
Rosenzweig, M. L. (1991). Habitat selection and population interactions: the search for mechanism. American Naturalist 137, S5-S28.
Samedi I. (1995). 'Modelling habitat suitability for koalas and application of UET methods in planning for koala habitat'. PhD thesis. University of Queensland.
Sherwin, W. B., Timms, P., Wilcken, J., and Houlden, B. (2000). Analysis and conservation implications of koala genetics. Conservation Biology 14 (3), 639-649.
43

Smith, P. (1990). Urban koalas and local government. In 'Koala Summit: managing
koalas in New South Wales'. (Eds. D. Lunney, C. Urqhart and P. Reed.) pp. 130-
136. NSW National Parks and Wildlife.
Smith, R. L. (1996). 'Ecology and field biology'. (Harper Collins College Publishers:
New York.) Smith, A., and Lee, A. (1984). The evolution of strategies for survival and
reproduction in possums and gliders.In 'Possums and Gliders'. (Eds. A. Smith and
I. Hume.) pp.17-33. (Surrey Beatty and Sons: NSW.)
Smith, P., and Smith, J. (1990). Decline of the urban koala (Phascolarctos cinereus)
population in Warringah Shire, Sydney (Australia). Australian Zoologist 26, 109-
129. Southwell, I. A. (1978). Essential oil content of koala food trees. In 'The Koala'. (Ed.
T. J. Bergin.) pp.62-74. The Zoological Parks Board ofN.S.W. Sydney.
Specht, R. L. (1970). Vegetation. In 'The Australian Environment'. (Ed. G. W.
Leeper.) pp. 44-67. CSIRC, Melbourne University Press, Melbourne.
Specht, R. L., and Brouwer, Y. M. (1975). Seasonal shoot growth of Eucalyptus spp.
in the Brisbane area of Queensland (with notes on shoot growth and litter fall in
other areas of Australia). Australian Joumal of Botany 23, 459-439.
Stone, B. F., and Carrick, F. N. (1996). Some effects of ticks on koalas, especially tick
paralysis. In 'Koalas -Research for Management. Proceedings of the Brisbane
Koala Symposium, 22nd-23rd September 1990'. (Ed. G. Gordon.) pp. 65-69.
(World Koala Research Inc: Brisbane.) Taylor, A. C., Marshall-Graves, J. A., Murray, N.D., and Sherwin, W. B. (1991).
Conservation genetics of the koala (Phascolarctos cinereus). II. Limited variability
in minisatellite DNA sequences. Biochemical Genetics 29, 355-363.
Thomas, 0. (1923). On some Queensland Phalangeridae. Annals and Magazine of
Natural History 9, 246-250. Tilley, D., and Uebel, K. (1990). Observations of koala populations within the Sydney
Water Board's Upper Nepean catchment area. In 'Koala Summit: managing koalas
in New South Wales'. (Eds. D. Lunney, C. Urqhart and P. Reed.) pp. 81-84. NSW
National Parks and Wildlife. Troughton, E. (1935). The southern race of the koala. 'Australian Naturalist' 9, 137-
140. U Nyo Tun. (1993). 'Re-establishment of rehabilitated koalas in the wild and their use
of habitat in Sheldon, Redland Shire, South-east Queensland with particular
reference to dietary selection'. Masters Thesis, University of Queensland, St Lucia,
Brisbane. Ullrey, D. E., Robinson, R. T., and Whetter, P. A. (1981). Composition of preferred
and rejected Eucalyptus browse offered to captive koalas, Phascolarctos cinereus
Marsupialia. Australian Joumal of Zoology 29, 839-846.
Vanderploeg, H. A., and Scavia, D. (1979). Two electivity indicies for feeding with
special reference to zooplankton grazing. Joumal of the Fisheries Research Board of Canada 36, 362-
365. Van Soest, P. J. (1982). 'Nutritional ecology of the ruminant'. (0 and B Books:
Corvallis, Oregon.) Wellwood, N. (1995). Koala friendly development- a developers perspective. In
'Proceedings on a Conference on the Status of the Koala in 1995'. pp. 91-111.
(Australian Koala Foundation: Brisbane.)
44

White, N. A. (1994). 'Habitat utilisation and population dynamics of the koala (Phascolarctos cinereus} in the Bremmer River catchment, south-east Queensland'. PhD thesis, University of Queensland, St Lucia, Brisbane.
White, N. A., and Kunst, N.D. (1990). Aspects of the ecology of the koala in southeastern Queensland. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 109-116. (Surrey Beatty and Sons: N.S.W.)
Wiegler, B. J., Girjes, A. A., White, N. A., Kunst, N.D., Carrick, F. N., and Lavin, M. F. (1988). The epidemiology of Chlamydia psittaci infection in a population of koalas (Phascolarctos cinereus) in south-east Queensland, Australia. Journal of Wildlife Diseases 24, 282-291.
Winter, J. W., and Leung, L. K. P. (1995). Common Spotted Cuscus. In 'The Mammals of Australia'. (Ed. R. Strahan.) pp.266-268. (Reed Books: Chatswood, NSW.)
Worley, M., Rideout, B., Shima, A., and Janssen, D. (1993). Opportunistic infections, cancer and hematologic disorders associated with retrovirus infection in the koala (Phascolarctos cinereus). In 'Proceedings of the American Association of Zoo Veterinarians, St Louis, October 1993'. pp. 181-182.
Worthington, W. J .. M., Melzer, A., Carrick, F., and Moritz, C. (1993). Low genetic diversity and inbreeding depression in Queensland koalas. Wildlife Research 20, 177-188.
45

Landholders Address to the Koala Symposium
John Rolfe "Broken Dray", M/S 381, Springsure, Qld.
I live in the Springsure area, and together with my family own and operate a cattle
property on the Nogoa River. It is mostly brigalow country, suitable for clearing so
that better pastures can be established and more cattle run and fattened. This is the
story across the Central Highlands, where substantial amounts of dearing have taken
place and relatively little brigalow country now remains uncleared. Of course, tree
clearing and koalas do not go very well together, and it is a source of mystery to many
people why the clearing continues in some areas. Let me give you some background
on landholders and koalas in the area. For the first hundred years after settlement (in the 1850s), this area was never very
prosperous in comparison to the sheep country of the mitchell grass plains in Western
Queensland. It was not until the 1960s that cattle came to be more profitable than
sheep, and since then a lot of development has occurred in the region.
In the early days, the fur trade was very important to the local economy, and was a
major activity in the region. It was· important for a couple of reasons. Firstly, it
provided a major source of income to the region, and secondly, it provided a very
flexible source of work for people. It appears that there would have been a large
number of people involved in the industry. My grandfather was born in 1900, and in
his memoirs of early years on "Castlevale", a 900 square mile property at the head of
the Nogoa River catchment, he mentions three separate groups of kangaroo shooters
(about eight or nine men in total) that were camped on the property. While these men
would have hunted on other properties as well, it does confirm that the fur trade was a
major industry. These kangaroo shooters would have pursued kangaroos, wallaroos and wallabies
for their skins, as well as koalas and possums. My grandfather recorded that there was
a large trade in koala skins, even when the season had "closed". The skins were salted
and hidden from any stray inspection by police, and were later pressed into wool bales
and consigned to Brisbane. In the 1920s, the koalas virtually died out of the district from the effects of
Chlamydia. My grandfather described the disease in this way in his memoirs:
' ... thousands of bears ... died in much agony between the years 1923 to 1929 from a
disease known to us country people as "Pink Eye". This disease, not the man with a
gun, practically exterminated this little animal from the Central West through to the
east coast of Queensland. This was a cruel disease; they contracted the blindness then
the urinal disorder, to finally die at the butt of trees after lingering for up to six
weeks. They died not only along the rivers and creeks, but back in the range country.
Neither man, dog or dingo would go near them. Shooters wouldn't touch them for fear
of contracting the disease. I never saw one koala in any type of country in the West for
forty five years. ' I remember in the late 1960s that there was some talk of koalas being seen in the
upper tributaries of the Nogoa River again. Sightings seemed to become more
prevalent through the 1970s, and by the early 1980s, koalas were quite often sighted
through our district along major watercourses. But this re-population occurred at the
same time that land clearing reached its peak in our area. In many cases people cleared
46

land where they had no experience of koalas ever using the vegetation. Where koalas were sighted, it was common practice to leave trees along watercourses and in shade clumps for them. But with little understanding of their habitat needs, most of these remnants have proved inadequate.
Additionally, the commercial arguments for clearing trees and running more cattle are generally very powerful. On our property, we have cleared with a great deal of interest in wildlife preservation, and have retained approximately 2,500 hectares of brigalow country, together with a slightly larger area of eucalypt woodland. Because we have not developed that country, we are probably "missing out" on $70,000 or $80,000 of gross income each year. We do not miss that loss of production so much because of our larger property size and because I have an outside income. Not all landholders are in the same position where they can set aside development opportunities.
In setting aside country though, we aretrying not only to conserve koalas. We have colonies of scrub wallabies, scrub turkeys and rock wallabies, each with their own area of scrub set aside. As well, we have a large area of bonewood scrub conserved because of its rarity now in the region. For us, koalas are just one species to consider when these preservation decisions are to be made.
Even so, it is difficult for us to know if we have cleared too much. We did not think in terms of vegetation corridors when we first started, and have some large clumps of scrub that are not very well connected. In some cases we have cleared too close to our main watercourses. What is very unclear is the effect on our koala populations. The recent run of dry years appears to have had a detrimental effect on koala numbers, and whether this has been compounded by the effects of clearing, is uncertain. We are not sure if the change in numbers and usage is more dramatic along creeks that do not have large shelter belts compared to those that have. We are also not sure how resilient koalas are to drought years where substantial numbers of trees die, even along watercourses.
While we are not planning to clear any more country, this type of information is important for people like us to make management decisions to maximise wildlife conservation on their land. The more that scientists can do to collect and disseminate information about koalas and their conservation, the easier it will be for us to make better management choices. For these reasons, the research and activities that this symposium is bringing together in Emerald are very welcome.
47

Community perceptions of koala populations and their management
in Port Stephens and Coffs Harbour Local Government Areas, New
South Wales
Daniel Lunney, Dionne Coburn, Alison Matthews and Chris Moon
NSW National Parks and Wildlife Service, PO Box 1967, Hurstville, NSW.
Abstract Community-based survey questionnaires were distributed in the local government
areas of Port Stephens (May 1992) and Coffs Harbour (October 1990) to identify: 1)
community perceptions of changes in local koala populations; 2) local threats to
koalas; and 3) koala management options which would be supported by the
community. A major fmding was that koala populations were most often perceived as
declining by those respondents who identified changes in the koala populations.
Furthermore, dogs were considered a more significant threat to koalas than cars. The
information gained in these studies was so comprehensive that it enabled draft koala
management plans to be formulated which were used as the basis for State
Environmental Planning Policy No. 44 - Koala Habitat Protection (SEPP 44) and
incorporated into the ANZECC National Koala Conservation Strategy (ANZECC
1998). Community-based surveys are a valuable means of identifying koala
population changes and are advocated where community support is integral to the
adoption and implementation of conservation planning on private land across local
government areas. These surveys are now formally incorporated in New South Wales
in local government area koala management plans under SEPP 44.
Introduction The driving force behind the preparation of the National Koala Conservation
Strategy (ANZECC 1998) was community interest in, and concern for, this uniquely
Australian species. Consequently, a guiding principle of the strategy is the
"recognition that community input and involvement is crucial to the conservation of
koalas". The community has already contributed a wealth of information to the
conservation biology of koalas. The currently accepted distribution of koalas in NSW
(Reed eta!. 1990) was determined from the results of a community-based survey that
led to the conclusion koalas have been lost from 50-75% of their range in the state.
Following this, the koala was listed as "vulnerable and rare" under the National Parks
and Wildlife Act 197 4 as amended by the Endangered Fauna (Interim Protection) Act
1991 and is now listed as "vulnerable" under the Threatened Species Conservation
Act 1995 (Lunney et al. 1996a; 1998a). Community participation was a central theme
at two major public forums on koala conservation in New South Wales (Lunney et al.
1990; Lunney & Matthews 1997a). There is now a growing recognition by wildlife managers and policy makers of the
importance of community knowledge and the necessity for local support in the
implementation of wildlife management strategies if they are to be effective. This
paper details the results of two extensive studies conducted at Port Stephens and Coffs
Harbour in coastal NSW. These surveys examined public perceptions of the status of
48

local koala populations, threats to their survival, and potential management options. Community surveys such as these can be used in the preparation of comprehensive koala plans of management under State Environmental Planning Policy No. 44 -Koala Habitat Protection (SEPP 44), to elicit community concerns and opinions and thus facilitate identification and adoption of appropriate management strategies and planning options. In giving the community a voice on wildlife issues, such surveys offer the community an opportunity to participate in the conservation and management of local wildlife. Without such participation, support and a sense of ownership by the community, local management plans are less likely to succeed in their conservation goals.
Port Stephens and Coffs Harbour were identified at a Koala Summit in 1988 (Lunney & Reed 1990) as local areas worthy of further koala study and management. The authors considered that case studies at a local level were useful in identifYing pressures on koala populations and habitat in NSW (Reed et al. 1990; Lunney et al. 1998a). A pilot study incorporating a community-based survey had been successfully carried out in 1990 on the lluka Peninsula near the mouth of the Clarence River (Lunney et al. 1996b ), so a high return rate of the community questionnaires was expected for these two large local government areas (LGAs) of Port Stephens and Coffs Harbour.
Although the community-based surveys, distributed by post, included both a questionnaire and a map for marking koala locations, this paper reports only on the questionnaire section. In presenting a direct comparison between the two LGAs, this paper gives a rare opportunity to evaluate the community-based survey approach by comparing the response patterns in these two LGAs. This work with koalas provides a practical model for accessing community knowledge and opinion on a broad range of wildlife issues and thereby tests the contribution of community involvement to wildlife management.
Methods Study Areas
Port Stephens: Port Stephens LGA on the New South Wales central coast covers an area of just over 97,000 ha. The population at the 1991 census was 43,719 people and estimated to be growing at around 4% per annum. Major land uses in the study area include farming, defence activities, forestry, tourism and a substantial area of water catchment (Hunter Water Corporation), which contains a large sand-mining operation, with urbanisation and sand-mining activities also occurring outside the water catchment area.
Coffs Harbour: The City of Coffs Harbour (hereinafter referred to as an "LGA" for convenience) is situated on the north coast ofNew South Wales between the Bellinger River Valley, the escarpment of the Great Divide, and the sandstone ridge of the Corindi Plateau/Coast Range, which separates the area from the Clarence Valley. State Forests occupy approximately 43% of the LGA's 94,500 ha, and are managed primarily for timber production, while freehold farmland and urban development areas occupy about 51% of the LGA. The Pacific Highway bisects the coastal plain from north to south, including some of the best koala habitat in Coffs Harbour. Horticultural activity is carried out on 5,140 ha (Coffs Harbour City Council 1992). Banana plantations dominate the slopes of the coastal valleys, particularly those with a northerly aspect. Coffs Harbour has a population of approximately 51,500 people, of whom 36% live in the town of Coffs Harbour, 22% in the Sawtell development area
49

(including Bayldon, Toormina and East Boambee), 20% in Woolgoolga and the
villages of the Northern Beaches and the remainder in rural areas (Australian Bureau
of Statistics 1992).
Postal Survey Port Stephens: In May 1992, a community-based koala survey was distributed by
post to each of the 16,500 residences within the Port Stephens LGA to establish
locations of koala populations. This survey form was a folded A2 sheet with a letter to
residents signed by both the National Parks and Wildlife Service and Port Stephens
Shire Council. Also included was a set of maps and questions aimed at eliciting
information about sightings of koalas, their status, threats to their survival and
potential management options (Appendix I). The survey form was modelled on
previously successful community-based koala surveys at Iluka and Eden (Lunney et
al. 1996b; 1997). In order to facilitate responses to the survey, arrangements were
made with Australia Post for completed koala survey forms to be returned "postage
paid". To further encourage responses, the survey was distributed in conjunction with
an extensive media campaign. In addition, each primary and secondary school within
the LGA was visited to promote completion of the koala surveys by the students'
parents in conjunction with a colouring-in competition for completion by the students
(Callaghan eta!. 1994). Coffs Harbour: The National Parks and Wildlife Service (NPWS) and Coffs
Harbour City Council reached agreement in April 1990 on a joint community-based
koala survey in the Coffs Harbour LGA. It was agreed that a joint approach was more
likely to elicit a broader response from the community because it reflected a co
operative effort to establish the facts before debating management options. State
Forests (formerly the Forestry Commission) research staff also assisted in the design
of the survey. State Forests is a predominant local land manager and its support
further increased the likelihood of a wide community response. In October 1990 a koala survey form, signed by NPWS researchers and the Coffs
Harbour Town Clerk, was delivered to every household in Coffs Harbour LGA
according to a schedule supplied by Australia Post (18,615 forms distributed). The
design of the postal survey was based on the successful pilot survey at Iluka (Lunney
et al. 1996b). It sought answers to questions on koala location, density, change in
numbers, health, death and breeding and the degree of support for various
conservation options. The questionnaire was large, on A3 format, and folded neatly
for postage following the prescribed post office layout (Appendix II). A free-post
return was marked on the survey form and posted. The mailing of the survey was
accompanied by local publicity and substantial support from the local newspaper, the
Coffs Harbour Advocate.
Data Collation All replies from both surveys were entered into a computer database, and summary
tables and cross-tabulations were generated using the program Minitab to present the
information and give a more detailed analysis of the results. All comments recorded
by Coffs Harbour respondents on their survey forms were typed and collated into one
document that, because of its sheer size (143 pages containing 1,011 comments), is
available separately. The full document is lodged with Council, NPWS and in the
Coffs Harbour library.
50

Results Response To The Survey
The response to the survey was higher from Port Stephens, with 2,995 (18%) forms returned, than Coffs Harbour, with 2,018 (11 %) forms returned. Note that totals of responses shown in results and tables below will vary because not all respondents answered all questions.
Frequency of Koala Sightings Of the respondents to the Port Stephens survey, 463 (15%) sighted koalas regularly
(monthly and quarterly), and the majority (57%) had seen koalas at least once during their time in the LGA. That 43% had not seen koalas indicates that there are locations within the LGA that do not have koalas, or where koalas are so rare that few people see them.
In Coffs Harbour, koalas were not often seen frequently, that is, weekly or monthly, in the respondent's local area. Whereas 44% ofthe 1,923 respondents to this question had seen a koala locally, 56% had never seen a koala in their local area. There were 622 respondents (33%) who had seen a koala in their local area within the previous 12 months, and these account for 69% of all respondents who had ever seen a koala.
Who Sees Koalas and Where Koalas were seen in most parts of both LGAs, but there were areas where residents
had a much greater chance of seeing koalas than in other parts of the LGA (Table I). In Port Stephens, Lemon Tree Passage was by far the best area in which to see koalas regularly, with the majority of respondents seeing koalas here monthly or quarterly, and less than 1% never having seen one. Tanilba Bay respondents were also more likely to have seen a koala than not. Reasonable numbers of people in Raymond Terrace, Williamtown, Salt Ash/Bob's Farm and Medowie had seen koalas, although not regularly. In all other areas in Port Stephens, respondents were more likely not to have seen a koala than to have seen one.
Similarly, in Coffs Harbour there was an uneven distribution of koala sightings locally, with some areas carrying the bulk of records. Areas where a majority of the respondents had seen koalas include Ulong!Lowanna (although the response rate was very low), Dairyville/Fridays Creek, Karangi!Coramba!Red Hill and Boambee. In contrast, areas where a majority of the respondents had never seen koalas include Corindi/Woolgoolga, Lower/Central Bucca and Glenreagh/Nana Glen. Recent sightings were more common in the areas of Dairyville/Fridays Creek, Ulong/Lowanna, Karangi/Coramba/Red Hill and Boambee. Recent sightings were uncommon in Corindi/Woolgoolga and Glenreagh/Nana Glen. However, Boambee, Karangi/Coramba!Red Hill, Dairyville/Fridays Creek and Repton/Raleigh are areas where residents commonly see koalas.
In Port Stephens, frequent sightings of koalas were made most often by respondents in the 40 to 69 year age groups (Table II). The lower frequency of sightings (i.e. those that see koalas 1-4 times a year or rarely/once only) was among the younger age groups. The largest numbers of those who had never seen a koala in the LGA were in the oldest age bracket (i.e. 70+). An almost identical pattern occurred in Coffs Harbour where the lower frequency of sightings were made most often by the younger age categories and the age groups most likely never to have seen a koala shifted towards the oldest respondents (70+). However, there did not appear to be a bias
51

towards any of the age groups when it came to people sighting koalas at least once a
month in Coffs Harbour. Unlike the Port Stephens survey form, the Coffs Harbour survey included a request
for the sex of the respondent. Regular sightings of koalas (1-4 times a year) were recorded equally by males and females while more females than males (616 to 392) reported never seeing koalas (Table III). This is likely to indicate that males replied when they commonly saw koalas, but were less inclined than females to reply to the
questionnaire if they had never seen a koala.
Table I How often respondents see koalas in their local area within both Port
Stephens and Coffs Harbour LGAs.
Location At least 1-4 times Rarely or Never TOTAL once/mth a year once only
Port Ste_ehens N % N % N % N % N (%)
Raymond Terrace 40 7 52 9 330 55 175 29 597 (100)
Patterson 0 0 0 0 0 0 1 10 1 (100) 0
Clarencetown 0 0 0 0 0 0 2 10 2 (100) 0
Karuah!Swan Bay 1 2 4 8 18 38 25 52 48 (100)
Williamtown 10 18 10 18 24 43 12 21 56 (100)
Shoal Bay 0 0 1 1 23 18 102 81 126 (100)
Tanilba Bay 59 22 50 18 152 56 12 4 273 (100)
Soldiers Point 1 1 4 2 26 19 106 77 137 (100)
Fingal Bay 0 0 0 0 4 7 61 94 65 (100)
Woodville/Hinton 0 0 0 0 2 12 15 88 17 (100)
Seaham 0 0 0 0 7 12 51 88 58 (100)
Bucketts Way 0 0 1 25 0 0 3 75 4 (100)
Medowie 3 1 28 9 153 55 95 34 277 (100)
Stockton!F em Bay 0 0 0 0 2 4 39 96 41 (100)
Salt Ash/Bobs Farm 22 17 26 59 54 42 27 21 129 (100)
Lemon Tree Passage 135 53 34 13 82 32 3 1 254 (100)
Anna Bay 4 4 2 2 29 32 58 62 93 (100)
Nelson Bay/Corlette 7 2 3 1 73 20 298 78 381 (100)
Other 28 12 22 10 71 30 112 48 233 (100)
Total 310 11 235 8 105 38 1197 43 2792 0 (100)
52

Coffs Harbour N % N % N % N % N (%) Coffs Harbour 25 4 57 10 166 28 345 58 593
(100) Glenreagh/Nana Glen 0 0 2 Boambee 18 17 25
3 13 24 40 24 31 29 32
73 55 (100) 30 106
(100) Lower/Central Bucca 1 Karangi/Coramba!Red 6 Hill
17 0 0 0 0 5 83 6(100) 23 79 (100) 7 21 27 34 43 18
Sawtell/Bayldon/Toor 11 mma Corindi!Woolgoolga 1
Ulong/Lowanna 1 Bonville 7
Korora!Moonee 3
Dairyville/Fridays 3 Creek Repton/Raleigh 5 TOTAL 81
3
0
11 7
2
8
12 4
58 14
5 1
3 33 19 19
29 16
14 37
10 23 243 13
131 31
34 12
4 45 32 31
61 34
13 34
13 29 532 27
220 52 420 (100)
256 87 296 (100)
1 11 9 (100) 44 43 102
(100) 88 48 181
(100) 8 21 38 (100)
16 36 44 (100) 1073 56 1929
100)
Sick I Dead I Breeding Koalas and Koalas in State Forests In Port Stephens, sick koalas were uncommon (Table IV), constituting only 8% of
sightings, whereas koalas with young were much more common, making up nearly half (48%) of all sightings. Records of dead koalas were also common at 34%, with 84% ofthese being dead on a road. State Forests carried 15% of sightings.
In Coffs Harbour, sick koalas represented 13% of sightings, and koalas with young constituted 25% of sightings in the respondents' local areas. Records of dead koalas were 39% of those sighted, with 76% of these being seen dead on the road. State Forests in Coffs Harbour carried 35% of sightings.
Table IT Frequency of koala sightings according to the age of respondents (PS = Port Stephens; CH = Coffs Harbour).
Age 0-19 20-29 30-39 40-49 50-59 60-69 70+
Port Stephens Coffs Harbour At least 1-4 times Rarely or Never At least 1-4 times Rarely or Never once/ a year once only Total once/ a year once only Total mth N %N 3 6 5 23 9 21 50 9 54 53 I4 40 55 17 21 59 I4 35 16 8 16
mth % N % N % N%N %N %N % II 24 50 16 33 48 1 I 9 95 40 100 42 239 10 4 10 211 40 221 4I 536 24 5 II 138 36 147 39 378 21 5 7 109 35 130 4I 315 11 4 8 146 35 180 43 420 9 4 8 66 3I 113 53 211 2 2
14 2I 25 37 28 4I 68 230 499 413
29 13 61 27 130 56 71 14 117 23 287 58 55 I3 132 32 205 50 36 I4 90 34 125 48 262
65 250 73 109
22 9 55 22 164 3 3 24 22 80
Total 259 192 789 907 2147 78 230 504 1019 1831
53
Table ill Frequency of koala sightings compared with the sex of the respondent
(Coffs Harbour only).
Sex of At least 1-4 times a Rarely or Never
Respondent once/mth year once only
N % N % N % N %
Male 44 58 115 50 219 43 392 42 Female 32 42 116 50 290 57 616 58 Total 76 100 231 100 509 100 1008 100
Table IV Numbers of people in Port Stephens and Coffs Harbour LGAs that have
seen sick koalas, koalas with young, dead koalas (and which of these were dead on a
road) and/or koalas in State Forests.
Port Stephens Coffs Harbour
Of those respondents that have seen N % N %
koalas, who has seen ... Sick koalas? 134 8 115 13 Koalas with young? 763 48 217 25 Dead koalas? 543 34 333 39
Of these, how many were dead on 473 84 254 76 a road?
Koalas in a State Forest? 234 15 301 35
Changes in Koala Populations A comparison between the Port Stephens and Coffs Harbour LGAs shows that the
majority of respondents did not know the status of their local koala populations, with
this proportion being greater in Coffs Harbour (Figure I), although this difference was
not found to be significant (X2 = 0.99; df= 3). Ofthose who did have an opinion, few
respondents in either LGA believed that koala populations were increasing. A greater
proportion reported a decline in koala numbers, although "steady" and "decreasing"
were almost equal in Port Stephens. From all areas in Port Stephens LGA, people generally responded that they did not
know the status of local koala populations. Of those who did give a definite response,
only those from Raymond Terrace and Medowie believed numbers to be declining.
The only area that showed a perceived increase in numbers was Tanilba Bay. fu Coffs
Harbour, most respondents (75%) also replied that they did not know whether there
had been changes in koala numbers, especially in the Corindi!W oolgoolga and
Glenreagh/Nana Glen areas. However, there were 286 respondents (15%) who
identified a decrease in numbers, many from the areas of Boambee (23% of local
respondents), Bonville (21 %), Dairyville/Fridays Creek (21 %) and Repton/Raleigh
(21% ), compared to only 34 people (2%) who considered there had been an increase.
When perceptions of koala populations were cross-tabulated with whether
respondents had seen breeding or dead koalas (Table V), it was found that most
people who had seen breeding or dead koalas believed that their numbers were either
54

steady or dropping. Conversely, those people who had not seen breeding or dead koalas generally answered "don't know" to this question.
80r,=======~--------~-----------------------l !!!! Port Stephens
70 D Coffs Harbour
60
% Re 50
sp 40 on ..., .. 30
20
10
0 Increased Remained Decreased Don't
Steady know
In the time you have lived in your local area has the no. of koalas •••.
Figure I Public perception of the status of local koala populations in Port Stephens and Coffs Harbour.
Table V Cross-tabulation of people's perceptions of the status of local koala populations in light of whether or not they have seen a breeding or dead koala (PS =
Port Stephens; CH = Coffs Harbour).
Koalas with Dead Koalas Young PS CH PS CH
Koala populations N % N % N % N % are ... Increasing 69 9 19 9 24 5 6 2 Steady 213 29 29 14 114 22 28 9 Decreasing 267 37 85 40 147 28 94 30 Don't know 181 25 79 37 231 45 183 59 Totals 730 100 212 100 516 100 311 100
The age structure of respondents from both LGAs was similar (Figure II), although there were slightly more responses from older age categories in Port Stephens. When perception of koala status was cross-tabulated with age of respondents (Table VI), the primary deviation from the overall pattern (as shown in Figure II) was that respondents from the older age classes (50-70) were more likely to perceive koala numbers to be increasing or steady (in the case of Coffs Harbour).
The number of respondents generally decreased as length of residence in the LGA increased (Figure III). Perceptions ofthe status of koalas appeared to be related to the number of years respondents had lived in the LGA. In both Port Stephens and Coffs Harbour, residents of between 10 and 20 years made up the greater proportion of respondents who saw koala numbers to be declining (Table VII). Longer-term
55

residents (30 years+) in Coffs Harbour were more likely to perceive koala numbers to
be increasing. This trend occurred to a much lesser extent in Port Stephens.
Table VI Perceptions of changes in local koala populations according to the age of
respondents (PS =Port Stephens; CH = Coffs Harbour).
In the time you have lived in your local area has the number of koalas ....
Increased Remained Steady PS CH PS
Age N % N % N %
0-19 2 3 1 3 6 2 20-29 4 5 1 3 29 8 30-39 14 19 9 27 92 26 40-49 13 18 7 21 69 19 50-59 16 22 12 37 57 16 60-69 16 22 2 6 75 21 70+ 8 11 1 3 27 8 Totals 73 100 33 100 355 100
t1!1! Port Stephens
25 0 Coffs Harbour
!l as 20 +-------] 0
~ 15+-------
~ ..... 0 ~ 10+----
0
0-19 20-29 30-39
CH N % 2 1 16 11 35 24 39 27 31 21 18 12 5 4 146100
40-49
Decreased PS CH
N % N % 12 3 20 7 29 8 36 14 88 24 58 22 72 19 54 20 57 15 46 17 78 21 40 15 35 10 13 5 371100 267100
50-59 60-69
Age ofRespondents (years)
Don't know PS CH
N % N % 25 2 46 4 170) 1 176 13 329 27 387 29 207 17 295 22 160 13 169 13 213 17 172 13 115 10 77 6 1219100 1322100
70+
Figure II Age structure of respondents from Port Stephens and Coffs Harbour koala
surveys.
56

Table VII Perceptions of changes in local koala populations according to the number of years respondents have lived in the LGA (PS = Port Stephens; CH = Coffs Harbour).
Years in the LGA. 1-4 5-9 10- 19 20-29 30+ Totals
"' .... = <li 'e
= Q Q,
"' <li ~ .... Q
~ e
In the time you have lived in your local area has the number of koalas .... Increased Remained Steady Decreased Don't know PS N
19 21 29 10 15 94
35
30
25
20
15
10
5
0
%
20 22 31 11 16 100
CH PS CH PS CH PS N % N % N % N % N % N
7 22 168 37 35 23 97 22 40 14 585 7 22 99 22 38 25 114 25 68 24 335 5 16 124 28 42 27 145 32 110 39 398 3 9 21 5 14 9 42 9 30 10 143 10 31 36 8 25 16 51 12 36 13 132 32 100 448 100 154 100 449 100 284 100 1593
1!1!Port Stephens
E;f----------------Jo Coffs Harbour
1 to 4 5 to 9 10 to 19 20 to 29 30+
No. ofYears liwdin the Shire
%
37 21 25 9 8 100
CH N %
533 39 370 27 300 22 90 7 70 5 1363 100
Figure lll Number of years that respondents have lived in either Port Stephens or Coffs Harbour LGAs.
Threats to Koalas and Options for Their Conservation and Management Respondents from Port Stephens (but not Coffs Harbour) were asked whether they
considered particular activities or developments in the LGA to be serious threats to the long-term survival of koalas (Figure IV). There was a near-unanimous consensus that roving dogs were threatening the survival of koalas in Port Stephens (94% answered "yes"), with similarly high results for both housing development and logging. The threat posed by LGA roads and the Pacific Highway were ranked as fourth and sixth respectively. Beach Mining, i.e., sand mining, was the lowest ranked threat, although there may have been some confusion as to whether the question was referring to all sand mining or sand mining only on beaches.
When asked which management options they would be prepared to support in order to help conserve koalas in their LGA, respondents from both Port Stephens and Coffs Harbour showed the same order of preference (Figure V).
57
The top three options for conservation actions were identified as tree planting
programs, restrictions on dogs and tree preservation orders. These were closely
followed by Environment Protection Zones (i.e. within the Local Environment Plan)
and traffic restrictions. The employment of a wildlife specialist and the use of public
money to buy land for koalas were the least preferred options, although even 71%
support in Coffs Harbour, which was the lowest support for any management option,
was nonetheless a clear majority endorsement. There were a few differences between
the two LGAs, the primary one being a generally higher support for the suggested
management options in Port Stephens. The exception was a slightly higher support in
Coffs Harbour for traffic restrictions, although not significantly higher (X2 = 1.91; df = 6).
For the Port Stephens data, support for management options was cross-tabulated
with threats to ascertain whether respondents' willingness to accept management
actions was influenced by whether or not they perceived activities to be threats to
koalas. Although analysed separately, the results from a cross-tabulation of traffic
restrictions with both the Pacific Highway and LGA roads were very similar. Of those
respondents who perceived either LGA roads or the Pacific Highway to be a threat,
the majority (88% and 89%) were prepared to accept traffic restrictions. In general,
most (80%) were prepared to accept traffic restrictions whether or not they saw the
roads as a threat. A small number of respondents (11-12%) who believed the roads
were a threat were not prepared to accept traffic restrictions, whereas 36-37% of
respondents who did not see roads as a threat did not support restrictions.
Of the 2,3 83 respondents who believed dogs were a threat, 99% were prepared to
support dog restrictions. Of the 145 respondents who didn't see dogs as a threat, more
(66%) supported restrictions than opposed them (34%).
100
90
80
"' 70 ~
?" 60
~
= ;e 50 = 0 Q; 40 "' ~ ~ 30 ~
20
10
0 Roving Dogs Housing Logging Shire Roads Tourist Pacific Beach Mining
Develpmnt Resorts Highway
Are any ofthe following serious threats to the long-term survival
ofthe koala in Port Stephens?
Figure IV Percentage of respondents who perceived the listed activities to be threats
to the long-term survival of koalas in Port Stephens LGA.
58

100
90
80
-"' 70 ~
~ 60
~
= .... 50 "C
= 0 c.. 40 "' ~ ~ 30 '::R ..
20
10
0 Tree Planting Dog Tree Environmental Traffic Employing a Using Public
Programs Restrictions Preservation Protection Restrictions Wildlife Money to
Orders Zones Special~ t Buy Land
Would you support any of the following to help conserve koalas ...
Figure V Percentage of respondents who indicated their support for proposed management options in both Port Stephens and Coffs Harbour.
The majority of respondents who perceived housing developments to be a threat to koalas were prepared to support both Environmental Protection Zones (97%) and the purchase of land with public money, although support for the latter was lower (79%). Those who did not see housing developments as a threat were more prepared to accept Environmental Protection Zones (72%) than the purchase of land ( 4 7%) which had almost equal support for "yes" and "no". There was a modest number (21%) of respondents who, although they believed housing developments to be threatening koalas, did not support land purchase.
In general, most respondents were prepared to accept management strategies regardless of whether or not they perceived logging to be a threat to koalas (Table VIII), the exception being a lower support for the purchase of land with public funds, which was higher for those who saw logging as a threat than for those who did not.
Regardless of whether or not they saw tourist resorts as a threat, most respondents (97%) were prepared to support Environmental Protection Zones. The purchase of public land received much less support, with 21% of those who believed resorts posed a threat unwilling to support this management option.
Support for Environmental Protection Zones and land purchase in the context of whether or not respondents saw beach mining as a threat to koalas was high (98% support for Environmental Protection Zones and 85% support for land purchase). However, those who did not see beach mining as a threat were much less likely to support land purchase (only 58% support) than Environmental Protection Zones (86% support).
59

Table VIll Support for tree planting programs, Tree Preservation Orders,
Environmental Protection Zones and the purchase of land with public money
according to whether or not respondents perceive logging to be a threat to the long
term survival of koalas in Port Stephens LGA.
Is Logging a threat to koalas? Yes No Total
Would you supeort ... N % N % N %
Tree Planting Programs Yes 1745 82 341 16 2086 98 No 14 1 23 1 37 2
Total 1759 83 364 17 2123 100 Tree Preservation Orders Yes 1724 82 301 14 2025 96
No 27 1 56 3 83 4 Total 1751 83 357 17 2108 100 Environmental Protection Yes 1675 81 269 13 1944 94 Zones
No 47 2 81 4 128 6 Total 1722 83 350 17 2072 100
Using Public Money to Buy Yes 1325 67 167 8 1492 75 Land
No 306 16 187 9 493 25 Total 1631 83 354 17 1985 100
In all cases but one, respondents perceived the threats similarly, regardless of how
long they had lived in Port Stephens LGA. The only difference from this trend was the
perceived impact of logging on koalas, where the longer-term residents were less
likely to perceive logging as a threat (although the majority of respondents did).
When support for management options in both LGAs was compared with perceived
status of the local koala population (Table IX), support for both tree planting and dog
restrictions appeared to be unaffected by the respondents' perceptions of their local
koala population. Greatest support for action was generally given by those respondents
who believed koala numbers were decreasing or answered "don't know". In Coffs
Harbour, support for employment of a wildlife specialist and purchase of land with
public funds was lowest from those who saw koala numbers as steady, rather than
those who believed koala numbers to be increasing. Support for management options
appeared to be little affected by whether or not respondents had seen sick koalas or
koalas with young.
60
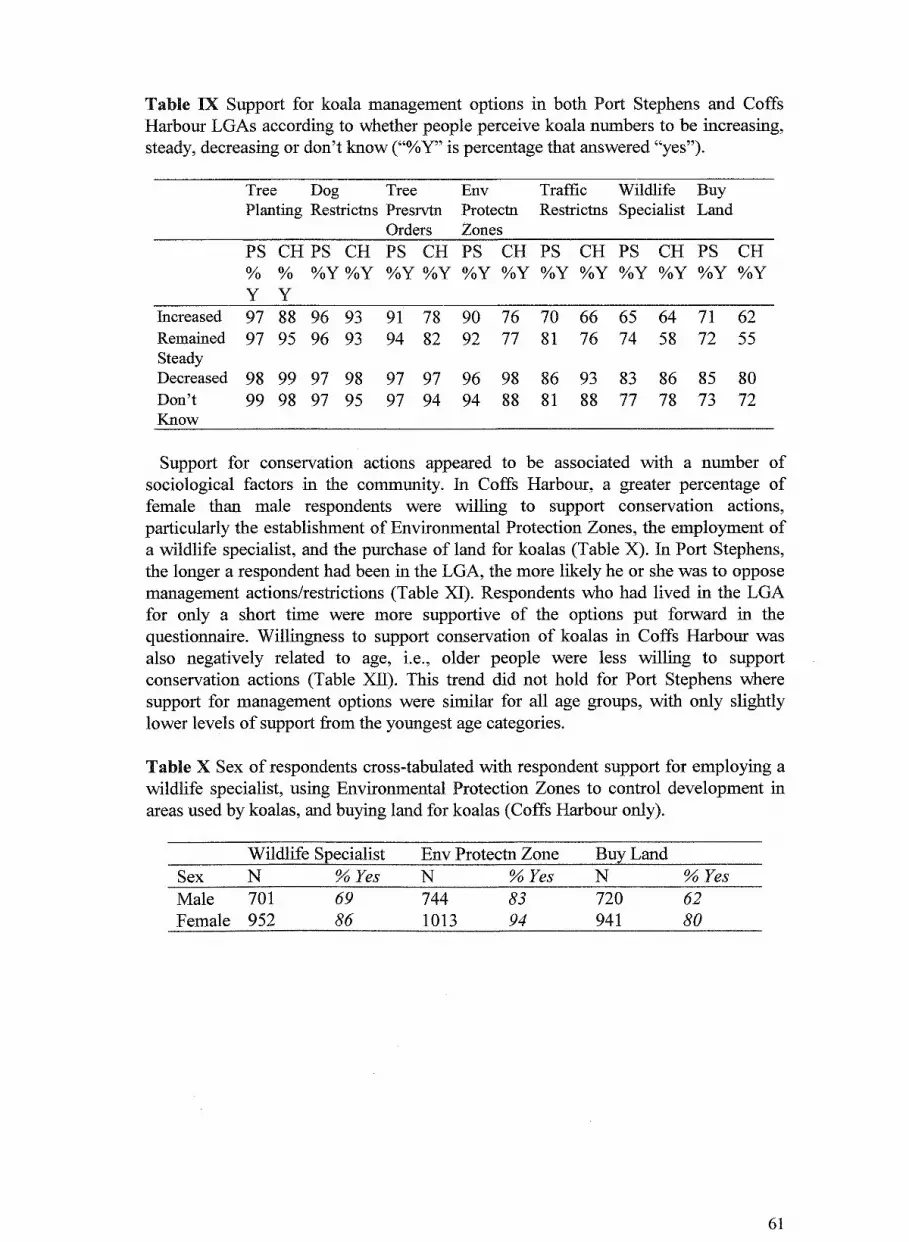
Table IX Support for koala management options in both Port Stephens and Coffs Harbour LGAs according to whether people perceive koala numbers to be increasing, steady, decreasing or don't know ("% Y" is percentage that answered "yes").
Tree Dog Tree Env Traffic Wildlife Buy Planting Restrictns Presrvtn Protectn Restrictns Specialist Land
Orders Zones PS CH PS CH PS CH PS CH PS CH PS CH PS CH % % %Y%Y %Y%Y %Y %Y %Y %Y %Y %Y %Y %Y y y
Increased 97 88 96 93 91 78 90 76 70 66 65 64 71 62 Remained 97 95 96 93 94 82 92 77 81 76 74 58 72 55 Steady Decreased 98 99 97 98 97 97 96 98 86 93 83 86 85 80 Don't 99 98 97 95 97 94 94 88 81 88 77 78 73 72 Know
Support for conservation actions appeared to be associated with a number of sociological factors in the community. In Coffs Harbour, a greater percentage of female than male respondents were willing to support conservation actions, particularly the establishment of Environmental Protection Zones, the employment of a wildlife specialist, and the purchase of land for koalas (Table X). In Port Stephens, the longer a respondent had been in the LGA, the more likely he or she was to oppose management actions/restrictions (Table XI). Respondents who had lived in the LGA for only a short time were more supportive of the options put forward in the questionnaire. Willingness to support conservation of koalas in Coffs Harbour was also negatively related to age, i.e., older people were less willing to support conservation actions (Table XII). This trend did not hold for Port Stephens where support for management options were similar for all age groups, with only slightly lower levels of support from the youngest age categories.
Table X Sex of respondents cross-tabulated with respondent support for employing a wildlife specialist, using Environmental Protection Zones to control development in areas used by koalas, and buying land for koalas (Coffs Harbour only).
Sex Male Female
Wildlife Specialist N %Yes 701 69 952 86
Env Protectn Zone N %Yes 744 83 1013 94
Buy Land N 720 941
%Yes 62 80
61

Table XI Support for conservation actions (employing a wildlife specialist,
implementing Environmental Protection Zones and buying land for koalas) according
to the length of residency (Port Stephens only).
Wildlife Specialist Env Protectn Zone Buy Land
Years N %Yes N %Yes N %Yes
1-4 803 83 863 96 816 79 5-9 531 79 580 94 524 78 10- 19 657 75 689 93 663 72 20-29 203 69 218 93 205 71 30+ 222 66 241 90 228 64 Totals 2416 77 2591 94 2436 75
Table XII Support for employing a wildlife specialist, using Environmental
Protection Zones to protect koala areas, and buying land for koalas in both Port
Stephens and Coffs Harbour according to the age of respondents ("% Y" is percentage
that answered "yes").
Wildlife Specialist Env Protectn Zone Buy Land
Prt Stphns C.ffs Hrbr Prt Stphns C.ffs Hrbr Prt Stphns Cffj Hrbr
Age N %Y N %Y N %Y N %Y N %Y N %Y
0-20 41 68 66 92 42 88 70 100 42 69 64 77 21-30 226 87 223 89 231 96 227 96 222 80 211 83
31-40 491 83 467 84 518 97 491 93 488 81 458 76
41-50 337 77 370 78 362 96 393 88 331 77 371 73
51-60 274 80 223 70 300 94 236 84 278 77 229 65
61-70 355 76 222 74 389 93 244 84 364 74 232 68
70+ 177 76 94 60 191 92 105 78 189 75 104 54
Totals 190 80 166 79 203 95 176 89 191 77 166 72 1 5 3 6 4 9
Discussion We now know a great deal about the koalas, and the community's attitude towards
them and their management, in two LGAs on the north coast of New South Wales.
Prior to this work, community attitudes and the extent of local knowledge was limited
to a small number of individual viewpoints (e.g., Lunney et a!. 1990). This study has
shown that community surveys are a valuable tool for accessing local knowledge and
uncovering information that would otherwise be difficult to obtain. For example, in
these surveys, dogs were perceived to be a major threat. This public acknowledgment
of the danger of dogs to koala conservation can be used as an argument for expansion
of dog control policies and actions implemented by local Councils. Community
surveys have also been recognised as a valuable exercise for involving the community
in the early phases of management strategies, thereby making it more likely that the
community will accept the management options when presented as a formal planning
document.
62

The strength of community survey is further highlighted when it is found to yield similar results to an independent field survey. Guidelines for the preparation of Comprehensive shire-wide Koala Plans of Management (CKPoM) under SEPP 44 state that both community records and field-based data are required to identify koala habitat (Lunney et al. 1997). Thus, in the process of producing CKPoMs for Coffs Harbour (Lunney et al. 1998b) and Port Stephens (in early redraft following the preSEPP plan of Callaghan et al. 1994; Lunney et al. 1998c), field surveys were carried out in both of these areas. This has enabled comparisons to be made between field results and those collected from the community survey which have showed that the two techniques produced strikingly similar results. This is a real bonus for those conducting community surveys, where the concern exists as to whether the results simply reflect population density of observers.
There was considerable similarity between the two LGAs. Points which stand out include: 1) dogs are perceived to be a serious threat to koalas; 2) many members of the community believe that koala numbers are declining and are prepared to support a number of management options in order to see them conserved; and 3) there was particularly strong support for local government to use their available powers to zone land to conserve koalas and their habitat and control the threats, such as by dog control, traffic restrictions and Tree Preservation Orders. A comparison of the results in the two LGAs allows identification of common concerns and local issues, and has ramifications for the design and interpretation of future koala surveys.
Comparison of the Two Local Government Areas A. Similarities 1. Both LGAs show an uneven distribution of koala sightings among local areas
within the LGA. There were markedly different numbers of sightings reported from different parts of the LGA. The striking differences among local areas were also confirmed by fmdings from subsequent field survey (e.g. Lunney et al. 1998b; 1998c ).
2. People in mid-range age brackets in both LGAs reported seeing koalas more often, followed by younger age groups, while koalas were least often seen by the oldest age groups. This result may be influenced by the greater likelihood of people of the mid-range age brackets either having better access to survey forms (e.g. as the head of the household), being more likely to take the time to fill them in, being more aware of the significance of the results once collated or a reflection that this adult age class is an active and community-minded group.
3. A similar proportion of dead koalas was seen in both LGAs (PS 34%; CH 39%), and a similar proportion was seen dead on a road (PS 84%; CH 76%). This fmding suggests that dead-on-road could be used as a local index of koala populations.
4. In general, most people who had seen breeding or dead koalas considered that koala numbers were either steady or dropping. Conversely, those people who had not seen breeding or dead koalas generally answered "don't know" to this question. This suggests that local knowledge of koalas was the basis for a firm response.
5. In both LGAs the age of the respondent was compared with the perception of change in koala populations. There was little influence of age, with the only exception being the older age group, who tended to believe koala numbers were increasing. This pattern was repeated when koala status was compared with the
63

number of years the respondent had lived in the LGA. Those respondents who
believed koala numbers were increasing were more likely to be longer-term
residents. There are a number of possible explanations for this pattern. The longer
term/older residents were more likely to have seen changes in local koala
populations and it may well be that koala numbers were increasing in some
locations. This would suggest that there has been forest regrowth or recovery
:from bush:fires. Neither is apparent on a broad scale :from our reading of the
postwar history of the LGAs, or the few published records (e.g., Yeates 1993; Knott et al. 1999). However, the fact that such a low number of
respondents believed that koala numbers were increasing, including many
respondents who had lived in the LGA for some time, leads us to the view that
there may be other factors influencing this result. The simplest conclusion is that
either clearing of habitat or recent interest in koalas has led to more observations
and hence the registering of an increase. 6. When asked which management options they would be prepared to support in
order to help conserve koalas in their LGA, respondents :from both Port Stephens -
and Coffs Harbour showed the same order of preference. Further, when
management options were compared with the perceived status of the koala
population, the response in both LGAs was very similar: the very high support for
both tree planting and dog restrictions appears to be little affected by the
respondents' perceptions of their local koala population. Greatest support was
generally given by those respondents who believe koala numbers were decreasing
or answered "don't know". Similarly, support for management options in both
LGAs was little affected by whether respondents had seen dead or breeding
koalas. B. Differences 1. Port Stephens had a higher return rate (18% compared to 11%) of the two LGAs.
It is possible that: • the fold-out double A3 size in Port Stephens was more attractive than the
single A3 in Coffs Harbour; • the publicity was better in Port Stephens; • there were more koala interest groups in Port Stephens to raise the profile of
the koala in the community; • koala numbers are greater in Port Stephens and so more people would respond
if they had seen a koala; • koalas had become a significant public interest issue following the publication
of the Koala Summit (Lunney et al. 1990); and
• the formal recognition that the koala had become a "vulnerable and rare"
species in February 1992 following the passage of the Endangered Fauna
(Interim Protection) Act 1991 which amended the National Parks and Wildlife
Act 1974 (Lunney et al. 1996a) had focused attention on the koala so that by
mid 1992 when the Port Stephens survey was launched the idea that the koala
was threatened attracted attention and increased the response rate.
The design of future surveys may be assisted by taking the above points into
consideration. 2. Koalas were seen more :frequently in Port Stephens (11% at least once a month
compared to 4% in Coffs Harbour); more respondents in Port Stephens had seen a
koala at some time (57%) than in Coffs Harbour (44%); and a higher proportion
64

of koalas with young were seen in Port Stephens ( 48%) than in Coffs Harbour (25% ). This suggests that Port Stephens has more koalas over a wider area than does Coffs Harbour and that the Port Stephens koala population is more robust than its Coffs Harbour counterpart. This is consistent with there being a slightly higher proportion of sick koalas seen in Coffs Harbour.
3. Coffs Harbour had a higher proportion of koala sightings in State Forests (35%) than Port Stephens (15%). This is consistent with the greater proportion of area covered by State Forests in Coffs Harbour (approx. 42% ofthe LGA) compared to Port Stephens (approx. 9%).
4. The "don't know" perception of changes in koala populations was slightly higher in Coffs Harbour (75%) than in Port Stephens (62%). However, this response was so high for both that the difference may be trivial.
5. The comparison between LGAs showed differences and similarities in their response to the perception of koala status. The number of respondents who considered that koala numbers were increasing was very low in both LGAs. In Port Stephens the number of respondents who considered that koala numbers were decreasing was equal to the number who considered that they were steady. By contrast, in Coffs Harbour there were twice as many respondents who thought koala numbers were decreasing as thought they were steady. Nonetheless, the proportion of respondents who perceived decreasing koala numbers was greater in Port Stephens.
Appraisal of the Approach The koala summit in 1988 (Lunney et al. 1990) and the koala forum in 1995
(Lunney & Matthews 1997a) were two formal expressions in New South Wales of the need for positive community participation in the process of conserving koalas. The National Strategy for the Conservation of Koalas (ANZECC 1998) gives further resonance to this view. Recent papers published from conferences held by the Australian Koala Foundation (1995; 1996; 1997) discuss the significance of including a wide cross-section of the community when deciding koala matters. The success of the Guidelines and Conditions for Koala Care in New South Wales (Lunney & Matthews 1997b ), which was produced through an extended process of community participation, gives further weight to the significance of community knowledge and concern.
There have been some criticisms of community survey as a method for locating fauna populations and providing information to be used to devise management strategies. A common issue is that community surveys give a biased indication of fauna locations, reflecting areas of high human populations rather than high fauna numbers. For many species this would be the case. However, koalas are one of a group of animals for which this concern is less likely. Koalas preferentially occupy the rich coastal and riverland areas in New South Wales, the very same areas that were first to be settled and continue to be under development pressure by human populations (Lunney & Leary 1988; Reed et al. 1990; Reed & Lunney 1990; Knott et al. 1999). Since koalas preferentially live where people live, it is expected that the two will correlate. This is further supported by the conducting of independent field studies in both Port Stephens and Coffs Harbour which have shown that field results for location of koala populations illustrate a significant overlap with koala habitat areas identified by community survey (Lunney et al. 1998b; 1998c).
65

With regard to the use of community survey questionnaire data as an indication of
community attitudes to conservation issues, some care must be taken in extrapolating
these results to the rest of the community given that respondents represent only a
modest percentage of the population, and it is unknown how representative their
attitudes may be of the entire community. It is possible that responses may come more
commonly from those community members that have an interest in koalas and/or the
environment, and thus responses may be biased in this way. If these limitations are
given due attention when considering the application of data collected from
community surveys, then the many benefits of using this approach will out-weigh the
shortcomings. This will particularly be the case when dealing with a high profile
species, such as the koala, where public decisions, such as by council in relation to
planning and dog control, are likely to be reported in the local papers and to be taken
by the sizable minority who replied to the questionnaire as indicative of council's
concern for co-existence with wildlife. The results of the community surveys at Port Stephens and Coffs Harbour provide
substantial evidence that there is much to be gained by involving the wider community
in conserving koalas. Of particular relevance here is the community's involvement in
the research phase of conservation, particularly in gathering information about koala
distribution, perceived threats and community acceptance of a range of possible
management solutions. A community-based koala survey in the Eden region (Lunney et al. 1997) changed
both government agency and community perception of the koala issues in south-east
New South Wales by establishing where koalas occurred so that management options
could be negotiated. The work in the USA of Kellert (1985) describes the value and
importance of community knowledge to wildlife management, while in Australia the
scientific aspects of the discipline are now becoming part of mainstream science. The
study by Gilfedder and Kirkpatrick (1997) on the influence of owners' intentions on
remnant bush conservation in Tasmania provides a parallel with the work reported in
this paper, in that attitudes were formally sought, grouped and categorised in a way
that managers and decision makers could utilise. The koala conferences in Queensland in 1990 and 1997, entitled Research for
Management (Gordon 1996), and the current set of proceedings (Lyons et al. 2000),
stress the need to involve landholders and land-use decision makers in the process of
managing the long-term conservation of koalas. In this paper we presented and
compared the results oftwo community-based surveys of koalas, analysed community
perceptions of threats to their populations, and canvassed support for potential
solutions. This information enables New South Wales councils to proceed with
preparing draft comprehensive LGA-wide koala plans of management under SEPP 44,
such has been achieved in Coffs Harbour (Lunney et al. 1998b ). There is now little doubt that there is a greater community awareness of the issues,
more support for conservation, and a willingness to see the relevant authorities
exercise their powers to give effect to the community's conservation interests. The
information gained has provided a valuable strand in defining koala habitat across an
entire LGA and provided a long-term perspective on community perceptions of koala
population changes not available from a one-off field study. We conclude that this
approach has much to offer those interested in conserving biodiversity by applying the
contemporary ethic of empowering the local community to support conservation
initiatives. The process is richly deserving of further development and the fmdings
presented here should materially assist in that process.
66

Acknowledgments We gratefully acknowledge the contribution of all respondents to the Coffs Harbour
and Port Stephens surveys, Peggy Eby for Minitab analysis of postal survey results, and funding support from the Sun Herald Koala Fund and the Foundation for National Parks and Wildlife. Thanks go to John Callaghan and Stephen Leathley for their contribution to the Port Stephens work, particularly in Callaghan eta!. (1994), and John Turbill (National Parks and Wildlife Service) for his continuing involvement in the Coffs Harbour plan, especially in seeing it through the public exhibition stage.
References ANZECC (1998). National Koala Conservation Strategy. (Prepared by the National
Koala Network: Canberra.) Australian Bureau of Statistics (1992). 1991 Census of Population and Housing
(Preliminary Data) for Coffs Harbour Local Government Area. (Provided by Coffs Harbour City Council: Coffs Harbour.)
Australian Koala Foundation (1995). Proceedings of a Conference on the Status of the Koala in 1995. Incorporating the Fourth National Carers Conference. (Australian Koala Foundation: Brisbane.)
Australian Koala Foundation (1996). Proceedings of a Conference on the Status ofthe Koala in 1996. (Australian Koala Foundation: Brisbane.)
Australian Koala Foundation (1997). Proceedings of a Conference on the Status of the Koala in 1997. (Australian Koala Foundation: Brisbane.)
Callaghan, J., Leathley, S., and Lunney, D. (1994). Port Stephens Koala Management Plan. Draft for public discussion. Public document as well as report to Department of Planning on a National Estate Grant. Copies available from Port Stephens Shire Council, Raymond Terrace.
Coffs Harbour City Council (1992). Annual Report 1991. (Coffs Harbour City Council: Coffs Harbour.)
Gilfedder, L., and Kirkpatrick, J. (1997). The influence of owners' intentions on remnant bush conservation in Tasmania. In 'Conservation outside Nature Reserves'. (Ed. P. Hale and D. Lamb.) pp. 217-25. (Centre for Conservation Biology, University of Queensland:Brisbane. ).
Gordon, G. (Ed) (1996). Koalas- Research for Management. Proceedings of the Brisbane Koala Symposium, 22nd-23rd September 1990. (World Koala Research Incorporated: Brisbane.)
Kellert, S. R. (1985).Social and perceptual factors in endangered species management. Journal ofWildlife Management 49, 528-36.
Knott, T., Lunney, D., Coburn, D., and Callaghan, J. (1999). An ecological history of koala habitat in Port Stephens Shire and the Lower Hunter on the Central Coast of New South Wales, 1801-1998. Pacific Conservation Biology 4 (4).
Lunney, D., and Leary, T. (1988). The impact on native mammals ofland-use changes and exotic species in the Bega District (New South Wales) since settlement. Australian Journal of Ecology 13, 67-92.
Lunney, D., and Reed, P. (1990). Epilogue: reflections on the Summit. In 'Koala Summit. Managing Koalas in New South Wales'. (Eds. D. Lunney, C. A. Urquhart and P. Reed.) pp. 243-246. (NSW National Parks and Wildlife Service: Hurstville.)
Lunney, D., Urquhart, C. A., and Reed, P. (Eds.) (1990). Koala Summit. Managing Koalas in NSW. In Proceedings of a conference held at the University of Sydney,
67

Lunney, D., Urquhart, C. A., and Reed, P. (Eds.) (1990). Koala Summit. Managing
Koalas in NSW. In Proceedings of a conference held at the University of Sydney,
7-8 November 1988. Pp. 1-268. (NSW National Parks and Wildlife Service:
Hurstville.) Lunney, D., Curtin, A., Ayres, D., Cogger, H. G., and Dickman, C. R. (1996a). An
ecological approach to identifying the endangered fauna ofNew South Wales.
Pacific Conservation Biology 2, 212-231.
Lunney, D., Moon, C., and Matthews, A. (1996b). A 1990 survey ofthe koala
Phascolarctos cinereus population at lluka in northern New South Wales. In
'Koalas: Research for Management'. (Ed. G. Gordon.) pp 102-122. (World Koala
Research Incorporated: Brisbane.) Lunney, D., and Matthews, A. (1997a). The changing roles of State and Local
Government in fauna conservation outside nature reserves: a case study of koalas in
New South Wales. In 'Conservation Outside Nature Reserves'. (Ed. P. Hale and
D.Lamb.) pp. 97-106. (Centre for Conservation Biology, University of Queensland:
Brisbane.) Lunney, D., and Matthews, A. (1997b). Guidelines and Conditions for Koala Care in
New South Wales. (NSW National Parks and Wildlife Service: Hurstville.)
Lunney, D., Esson, C., Moon, C., Ellis, M., and Matthews, A. (1997). A community
based survey of the koala Phascolarctos cinereus in the Eden region of south-east
New South Wales. Wildlife Research 24, 111-128.
Lunney, D., Curtin, A., Ayres, D., Cogger, H. G., Dickman, C. R., Maitz, W., and
Fisher, D. (1998a). The threatened and non-threatened vertebrate fauna ofNew
South Wales: an ecological approach. Environmental and Heritage Monograph
Series. (NSW National Parks and Wildlife Service: Hurstville.)
Lunney, D., Moon, C., Matthews, A., and Turbill, J. (1998b). Coffs Harbour Koala
Plan of Management. A Comprehensive Koala Plan of Management for the City of
Coffs Harbour prepared under State Environmental Planning Policy No. 44- Koala
Habitat Protection. February 1998- Draft for Public Comment. (NSW National
Parks and Wildlife Service: Hurstville.)
Lunney, D., Phillips, S., Callaghan, J., and Coburn, D. (1998c). A new approach to
defming the distribution of Koalas and conserving their habitat: a case study from
Port Stephens Shire on the central coast ofNew South Wales. Pacific Conservation
Biology 4, 186-196. Lyons, K., Melzer, A., Carrick, F., and Lamb, D. (Eds.) (2000). The Management and
Research of Non-urban Koala Populations. (Central Queensland University
Publishing Unit: Rockhampton.) Reed, P., Lunney, D., and Walker, P. (1990). Survey ofthe koala Phascolarctos
cinereus (Goldfuss) in New South Wales (1986-87), with an ecological
interpretation of its distribution. In 'Biology ofthe Koala'. (Eds. A. K. Lee, K. A.
Handasyde and G. D. Sanson.) pp. 55-74. (Surrey Beatty and Sons: Sydney.)
Reed, P., and Lunney D. (1990). Habitat loss: the key problem for the long-term
survival of koalas in NSW. In 'Koala Summit. Managing koalas in NSW'. (Eds. D.
Lunney, C. A. Urquhart and P. Reed.) pp. 9-31. (National Parks and Wildlife
Service: Hurstville.) Yeates, N. (1993). 'Coffs Harbour'. Volume 2: 1946 to 1964. (Bananacoast Printers:
Coffs Harbour.)
68

Appendix I Port Stephens community-based questionnaire survey. This form was distributed through the post to every household in Port Stephens Shire in August 1992.
KOALA SURVEY No Pos~age stamp required
if posted in Austtalia
Dear Rrsident,
Please fold and return to:
Port Stephens Shire Koala Survey Reply Paid No 2116 P.O. BOX270 RAYMOND TERRACE NSW
Ill
2324
We are writing to ask for your co-operation in a koala survey which the National Parks and Wildlife Service is carrying out in Port Stephens Shire in conjuction with the Port Stephens Shire Council and the Hunter Koala Preservation Society. The purpose of the survey is to locate the koala populations in the Shire, to identify their habitat, and to produce a joint NPWS-Council plan to conserve them.
A National Estate Grant to the National Parks and Wildlife Service has provided the primary funding for the survey. Tomago Aluminum has provided funding for the printing and distribution of this questionnaire.
We would like you to fill in this survey from EVEN IF YOU HAVE NEVER SEEN ANY KOALAS. A ·lack of sightings in any area is also important in building the picture of koala distribution. To help us understand the present distribution of koalas, we are asking for any historical information you may have, such as your family's recollections, about koalas in the Shire. We also want to use the opportunity provided by this survey to collect information on some other animals of interest.
We believe that a comprehensive and objective survey is essential for planning koala management in the Shire. For this reason, we are asking all residents to provide detailed and accurate responses to the survey Thank you for taking the time to assist us. ·1:. .• ,
John Callaghan and Dan Lunney (NSW) National Parks and Wildlife Service (049) 873-108 or (02) 585-6489 May 1992
Please circle the nearest answer, or give details as requested.
What is your local area or nearest town? 1) Raymond Terrace 7) Tanilba Bay 2) Paterson · 8) Soldiers Point 3) Oarencetown 9) Fingal Bay 4) Karuah/Swan Bay 10) Woodville/Hinton 5) Williamstown 11) Seabam 6) Shoal Bay 12} Bucketts Way (Limebumers)
19) Other (Please name)
Stephen Leathley Port Stephens Shire Council (049) 830-282
13) Medowie 14) Stockton/Fern Bay 15) Salt Ash/Bobs Farm 16) Lemon Tree Passage 17) Anna Bay 18) Nelson Bay/Corlette
Printed on 100% Recycled paper
69

-..) 0
KOALA SURVEY Please fold and return to:
THE fi HOUSEHOlDER
~ Coffs Harbour Koala Survey
FREEPOST No. 62
IDoet Map
t1ee1e M.rk wHh • (:rOJ'I (d plit.,et
...,.,..,.,o.o,.•ttHfl.elt.,ala,....,edd
Of J~Gulblt) ttle-,uryov savH.
Jr l,..lloela .. u dud, pleue
clrclelhtcronflli"Y)
Coffs Harbour, 2450
\
COFFS HARBOUR KOALA SURVEY OCTOBER 1990
DEAR HOUSEHOLDER . The National Parks and Wildlife Service is carryin9 out a survey of
Koalas in Coffs Harbour with assistance from Coffs Harbour City
Council. the Sun Herald Koala Fund a.nd the Forestry Comnission,
We wa.nt to collect detailed information on Koalas so that future
planning decisions can help to conserve them in Coffs Harbour.
To be successful. the survey needs your support. We a.sk you to fill
out and return this questionnaire. even if you have never seen a
Koala near Coffs Harbour. All replies will be confidential.
Please return the c·omp1eted form to the co11ecHon box at Coffs
Harbour City Council•s Office or post it (no stamp required) to reach
us by 26 October 1990.
Thank you for your help. Chri-s Moon and Dan Lunney - Niltiona..l
Parks and Wildlife Service
PR Harvey - Town Clerk, CHCC
Enquiries phone: 522 SSS
Please circle the closest answer.
1. WHAT IS YOUR LOCAL AREA?
(1) Coffs Harbour (2) Glenreaghl {3) Boambee
Nana Glen
(4) loYer/Central (5) KarangiiCoramba (6) Sawte11/Bay1do~ Bucca /Red Hi11 Toormina
(7) Corindil (8) Ul ong/lowanna (9) Bonville
Woolgoolga (11) Dalryvi11e/ ( 12) Repton/Raleigh
(10) Koronll1oonee Fridays Creek
2. ON AVERAGE, HOW OFTE!I DO YOU SEE KOALAS IN YOUR LOCAL AREA?
(1) Weekly (2) Monthly (3) Quarterly
(4) Yearly (5) Rarely (6} Never
3. HOW MANY KOALAS HAVE YOU SEEN IN YOUR LOCAL AREA IN THE LAST
l1/ELVE MONTHS?
4. IN THE TIHE YOU HAVE LIVED IN YOUR LOCAL AREA HAS THE NU'IIIER OF
KOALAS: (a) Increased (b) Stayed the same (c) Decreased
(d) Oon' t know
If you ha"e noticed a change, what year did this begin? ..•.••••
5. HAVE YOU SEEN SICK KOALAS IN YOUR lOCAl AREA?
hg with infected eyes or a wet. dirty tail)
6. HAVE YOU SEEN KOALAS Willi YOUNG IN YOUR LOCAL AREA?
Yes I No
Yes I No
7. WHERE HAVE YOU EVER SEEN KOALAS IN THE COFFS HARBOUR DISTRICT?
On the large map overleaf. or the inset map on this page. please
show all the- locations and, if possible, the year you nw it.
8. HAVE YOU EVER SEEN /Wt DEAD KOALAS IN THE
COFFS HARBOUR AREA? If Yes, was the Koala dead on a road?
Please mark these on the 11aps.
9. HAVE YOU SEEN KOALAS IN A STATE FOREST NEAR
COFFS HARBOUR? Were you working in that State Forest?
'Jere you 'olisiting or driving through that
State Forest?
Yes I No Yes I No
Yes I No Yes I No
Yes I No
Conserving Koalas in Coffs Hubour might in'ololve some restrictions •
trade-offs or other costs. We would like to know which actions would
have your support.
10. \IOULD YOU SUPPORT /Wt OF THE FOLLOIIING ACTIOIIS TO HELP CONSERVE
KOALAS IN COFFS HARBOUR?
(1) Traffic Restrictions (eg speed limHs
at night in areas with Koalas)
Restrictions on dogs (eg to stop dogs roaming at night)
(2)
(l)
(4)
(5)
(6)
(7)
Tree Preservation Orders (to protect
Koala food trees) Tree-planting PrograiTIII'Ies (to increase
Koalas' food supply)
Employ a Wildlife Specialist in Coffs Har-bour
Environment Protection Zones (to control
development in areas used by Koahs)
Use public money (from rates or taxes)
to buy 1 and for Koa 1 a Reserves
11. HOW MANY YEARS HAVE YOU LIVED IN COFFS HARBOUR?
12. YOUR STREET NAil£
13. STATISTICAL INFORMATION (CONFIDENTIAL)
YOUR AGE SEX:
14. MAY \1£ CONTACT YOU FOR IIJRE DETA!LS?
Name •••. •• • ••• ••••••••
Phone No. Hoose No.
15. DO YOU HAVE /Wt COt91tNTS?
Ves I No
Yts I No
Yes I No
Yes I No
Yes I No
Yes I No
Yes I No
Hale I Female
Yes I No
~ ~
s· 0 ~ g. (1)
e:> IJJ"'' S."'' r:J~
aS. a~·
..... ::::
::OC§.
[ = n 0
\0 og. • (1)
"g
~
~ _E; 0
~
i ::r§ 0 ...... ~ :=q IJ1 I (1) r:J ::r~ 0 IJ1 ,_..(1) o..O.. ...... ..c ::s :::: n(l) 0 ~ H"l ..... tiiig ;I::~ ~ ...... ..... ..... r::;-(1) 0 IJ1
:_~ 0~ £· ;-;1 ~ t;;• (1) § ~ (1) s a~
IJ1

A review of some techniques used to describe koala habitat and its use by koalas with particular reference to low density populations at Eden
Vic Jurskis State Forests of NSW, Southern Research Centre, Eden.
Abstract Koalas occur at low densities in the forests of south eastern New South Wales
near Eden, and this makes it difficult to find koalas and accumulate data describing their habitat and ecology. Standard ecological techniques are briefly outlined and the difficulties in applying them to low density populations are illustrated. Difficulties arise from differences in the scale of koala activity in relation to the scale of environmental variation, small datasets and the inability to establish a measurable range of koala densities.
Three approaches have been used at Eden, ranging from a very fine to a very broad scale. These are: analyses of tree preferences based on variable radius plots around detection points; horne range analyses using minimum convex polygons and harmonic mean isopleths to contrast frequently used and little used areas within horne ranges; and broad comparisons of the distribution of koala records against the pattern of environmental variation in the region. The use of these methods at Eden is compared with alternative approaches described in the literature.
Modelling can potentially be used to test concepts of koala ecology. The density of preferred tree species has been used to model habitat quality. Apparent anomalies suggest that modelling should incorporate a broader perspective. Other environmental factors at the horne range scale as well as the regional scale may be as important as the floristic composition of the forest canopy in determining habitat quality for koalas. Studies of low density populations at Eden have highlighted the need to consider additional factors at broader scales than canopy floristics in order to understand koala distribution and density. Recognition of other factors controlling habitat quality may lead to improved understanding of the variability in carrying capacity of eucalypt ecosystems and the consequential variation in densities of natural koala populations.
Introduction A traditional technique in ecology is to define a study area containing the target
species, stratify the environment in the area and compare the distribution of the target species amongst the strata. Wild koala populations have typically been studied in areas where they are reasonably dense, often in numbers greater than 1 animal per hectare (Table I). In such areas it is possible to relate koala distribution and density to habitat using traditional techniques in relatively small study areas. For example, Mitchell (1992) estimated that there were 444 koalas in a study area of about 700 ha. He estimated that there were about 2 koalas ha-1 in manna gum woodlands, 1 koala ha-
1 in redgurn woodlands and 1 koala per 3 ha in mahogany
71

woodlands. In low density populations the animals are difficult to locate so it is
difficult to accumulate data and to obtain a range of values for koala activity
which can be compared with environmental parameters. Melzer (1994) compared
koala habitats and population densities in the traditional way even though his
study sampled low density populations. Melzer's (1994) study area comprised 5
sites each of 100 ha with a total population of about 63 koalas. Koala densities
ranged from 5 to 40 koalas per 100ha in three woodland sites and 2 or less koalas
per 100ha in two open forest sites (Melzer 1994). Koala densities at Eden are as low as any that have been studied (Table I). The
average density is similar to the minimum density ( <0.02 koalas ha-1) sampled by
Melzer (1994). This is about the limit of measurable density, thus it is difficult to
decide whether koalas are 'present' or 'absent'and a range of densities is not
available for comparison with environmental factors.
Table I Population densities reported in koala studies
Location Koala Density (no. ha-1) Author
French Island, 7.5 Mitchell and Martin
Vic (1990)
Tucki Tucki, 4.0 Gall (1980)
NSW Springsure, 1.5 Gordon et al. ( 1990)
Qld Brisbane Ranges, 1.2 Hindell (1984)
Vic Oakey, Qld 1.1 Gordon et al. (1990)
Gold Coast, 0.4 White and Kunst
Qld (1990)
Springsure, 0.2 Melzer and Lamb
Qld (1994)
Springsure, 0.01 Melzer and Lamb
Qld (1994)
Capella, Qld 0.005 Melzer and Lamb (1994)
Eden,NSW 0.006 Jurskis and Potter (1997)
Harmonic mean (HM) home ranges of eight radio collared koalas at Eden
ranged from about 40 to 500 hectares, and Minimum Convex Polygons (MCPs)
ranged from about 70 to 2000 hectares (Jurskis and Potter 1997). A typical study
site may cover hundreds of hectares with considerable environmental variability
and include only one koala, thus presenting problems for ecological studies. For
example, Figure I is the home range (Harmonic Mean 90% isopleth) of a radio
collared koala at Eden which covered a variety of topographic positions and
aspects. Eight eucalypt species occurred in this koala's range and it was located in
thirteen different forest types (Forestry Commission 1989).
72

1000
• KOALA LOCATIONS
_. _. HARMONIC MEAN 90% ISOPLETH
Figure I Home range of a koala at Eden showing topographic variability
An ecological technique which can be used to overcome difficulties associated
with mobile animals having large home ranges is a regional survey, stratified on a broad scale. Kavanagh et al. (1995) were able to examine the regional distribution of koalas in north east NSW using data from such a survey. Kavanagh and
Bamkin (1995) used a similar approach to study the distribution of owls and arboreal animals at Eden. They sampled 200 sites covering two geological classes
and two logging history classes. Although this approach yielded useful information for many species, only four koalas were detected. A similar regional
survey of 140 sites using playback of taped koala calls only detected four koalas (Jurskis and Potter 1997). Since koalas in low density populations are difficult to detect, regional surveys may not yield sufficient data for meaningful analysis.
73

Koala studies at Eden have examined habitat use at the individual tree scale, the
home range scale and the regional scale. Limited data are available at each of these scales and examination of all these data is required to understand habitat use
by low density koala populations. This paper reviews some of the approaches used at Eden to overcome the difficulties of low densities combined with high
environmental variability, and compares them with alternative approaches. The review indicates that there may be benefits in broadening the focus of koala research and habitat modelling. Although low densities of koalas have sometimes
been regarded as a cause for concern ( eg. Reed et al. 1990), few studies have been
conducted in dense forests as distinct from woodlands/open forests. There is increasing evidence that low density populations are a natural feature of denser
forest environments (White and Kunst 1990; Jurskis and Potter 1997). In future, a broader focus to koala research and modelling may include recognition of a
naturally wide range of carrying capacities for koalas (Jurskis 1996),
corresponding to the wide range of eucalypt dominated structural formations in eastern Australia.
Assessing the use of trees by koalas Since it has not been possible at Eden to measure a range of koala densities and
relate them to environmental variation, analyses have focused on the use of trees
by koalas. At sites where koalas have been detected, plots have been set up around
trees used by koalas and the use of trees compared with their availability in the plots (Jurskis and Potter 1997). The sites have accumulated from incidental
records and a variety of surveys, thus the sample has been obtained 'haphazardly'
rather than randomly or systematically. Trees used by koalas were identified by
the presence of faecal pellets or a radio collared koala and classified by species,
diameter and age/dominance or crown condition (Florence 1996). Surrounding trees were classified in the same way.
Data for particular koalas, environments or geographic areas were then pooled.
Thus a series of data sets was constructed which characterised particular home
ranges, environments or areas according to the average number of trees per hectare
having certain characteristics. The proportional use of each class of tree by koalas was estimated from the sample of used trees. These data were analysed to examine
tree preferences, define vegetation types at sample sites and describe study areas.
Unfortunately the method did not provide data describing environments from which koalas were absent. Thus comparisons could not be drawn between
occupied and unoccupied sites. Phillips et al. (1996) described a similar system for determining tree preferences
using fixed area plots including all trees over 10 em dbh. Jurskis and Potter (1997)
used concentric variable radius plots sampling fixed numbers of trees between 10
em and 29 em dbh in one plot and trees 30 em dbh and over in a second plot. This avoided oversampling of saplings or small trees and ensured adequate sampling of
established trees in the upper canopy. Melzer (1994) sampled vegetation in core
home ranges using the point centred quarter method which allowed equivalent sampling of a range of tree classes. Phillips et al. (1996) distributed their plots
randomly within predetermined biophysical strata, but subsequently analysed only those plots containing koala faecal pellets. Other tree preference studies have
measured trees along transects or around grid points systematically distributed through the study areas. Examples are the studies of Hindell et al. (1985) and
74

White and Kunst (1990). Some studies measured every tree in the study area, for example Lithgow (1980), as cited by Hindell et al. (1985).
The difficulty with applying either random sampling (eg Phillips et al. 1996) or systematic sampling ( eg Hindell et al. 1985) to low density populations is that very few records of trees used by koalas are obtained and a great deal of effort is spent collecting data which cannot be related to koala activities. The haphazard sampling of Jurskis and Potter (1997) is open to criticism because conclusions are potentially biased by factors which made the studied animals apparent (Cork 1995). For example, the data may disproportionately represent drier forests rather than wetter forests because koalas are more detectable in shorter more open stands.
Whatever the sampling strategy, analyses of tree preferences typically rely on a chi square test or G test of the relationship between the proportions of species used by koalas and the proportions of species available to them. If there is a statistically significant difference, preferred trees are those which receive disproportionately high use compared to their availability. Application of this method, in conjunction with systematic samples, assumes a homogenous distribution of tree species across the landscape ( eg Hindell et al. 1985), so that the proportions of species available to koalas are fixed for the whole study area. Although the same assumptions also applied to the haphazard sampling of Jurskis and Potter (1997), their method only sampled parts of the study area that were known to be used by koalas and the levels of use were consistently very low. Thus, floristic homogeneity was not assumed across a wide range of koala densities. An inbuilt assumption of homogeneity may cause difficulties in interpreting data. For example, Hindell et al. (1985) found that a population of koalas exhibited tree preferences, but when Hindell and Lee (1988) studied 20 individual koalas with defined home ranges in the same population, most of the koalas didn't show tree preferences. Preferences were only apparent when the preferred species were in low abundance in the koalas' home range. Similarly, Melzer (1994) provided little evidence of preferential use of eucalypts within the core home ranges of koalas at Norwood Creek. The importance index of White and Kunst (1990) may partially compensate for problems associated with floristic
. heterogeneity since common species are likely to be distributed more uniformly than uncommon species, and they are given more weight in the index. Tree preference indices and importance indices have been constructed and discussed by Hindell and Lee (1987, 1988) and White and Kunst (1990) respectively. The preference index ranks trees according to disproportionate use by koalas. A species which is uncommon at a site may have a high index, but may comprise a small proportion of the resource used by koalas. The importance index uses a weighting for relative abundance to modify the preference ranking so that common preferred species rank higher than uncommon preferred species. Although the analyses have mainly been used to describe use of tree species by koalas, they can also be used to describe use of trees according to size, crown condition or a combination of factors (Jurskis and Potter 1997).
A different approach was taken by Phillips et al. ( 1996) who calculated a strike rate (proportion of trees with faecal pellets) for each tree species across a range of active plots (plots containing faecal pellets) and used non parametric tests to identify differences between species in their strike rate. Preferred tree species were those which had a significantly higher strike rate than other trees in the same
75

stratum. Activity levels (percentage of trees having faecal pellets) were calculated
for each plot and subsequently compared with the presence of preferred trees.
Phillips et al. (1996) did not indicate that activity levels were compared with other
environmental parameters. This would have potential benefits, particularly if plots
with zero activity were included in the analyses. In low density populations at
Eden, plot 'activity levels' are extremely low and zero levels do not occur due to
the impracticality of random sampling. Thus a range of activity levels is not
available to compare with environmental factors. Phillips et al. (1996) used
Spearmans Rank Correlation Procedure to check for relationships between ranked
strike rates and ranked densities. The existence of positive relationships apparently
would be taken to indicate a lack of preference. Phillips et al. (1996) applied a
great deal of statistical rigour to their analyses of species preferences, however
they didn't analyse other factors such as tree size or crown condition which may
also affect preferences (Hindell and Lee 1987; Jurskis and Potter 1997), nor did
they use the plots (including nil plots) to examine relationships between site
factors and activity levels. Although tree species preferences have been reported at Eden, forest stands or
forest types containing high densities of preferred tree species do not support high
densities of koalas (Jurskis 1996; Jurskis and Potter 1997), suggesting that other
environmental factors have an over-riding influence on habitat quality. Melzer
(1994) reported a wide range of koala densities on five sites which all contained
Eucalyptus tereticomis. There was no correlation between koala density and the
density of preferred species (Melzer 1994 ), suggesting that factors other than
floristics were important to koalas. Since the tree plots from low density populations at Eden do not contain a
sufficient range of activity levels (including zero), they can't be used for detailed
comparisons of koala use according to site factors. Nevertheless, plot data can be
used to categorise plots into environmental strata and compare the frequency of
plots within strata against the relative extent of the strata within a study area. For
example, Jurskis and Potter (1997) compared the frequency of koala detections
between dry and wet vegetation types (Table II). This comparison assumed that
the different vegetation types were equally available to each koala. Such
assumptions should be tested. In the case of the koala occupying the home range
shown in Figure I, for example, the assumption appeared reasonable since the
home range included a fairly even spread of both exposed and sheltered aspects as
well as ridge and gully situations, suggesting a reasonably even spread of wet and
dry forest types. In low density populations, different analyses may need to be
applied to data from different sources, according to available information or
assumptions which can be made regarding the environment of the source area.
Plot data from a study area or home range can be pooled to provide a floristic
and/or structural description of the area. Table II is a stand table describing the
forest overstorey within the home range depicted in Figure I. It shows that eight
eucalypt species occur in the home range of which six are reasonably common,
one (E. sieberi) is very common and one (E. smithii) is quite rare. There is a large
number of trees in the smallest size class and a decreasing number of trees in the
larger size classes.
76

Table II Observations of radio collared koalas in Forest Types1
Forest Type - Dry
Koala 63 112 114 121 123 132 150 152 Total Ruth 21 26.5 10.5 1.5 1 60.5 Simon 12.5 3 2.5 1 19 Wayne 1 1 1 4 2 1 10 Bob 17 14.5 8 8 1.5 7 56 Roberta 3 7 13 1 1 25 Robert 1 2 3 2 3 1 12 Michelle 0.5 5 5.5
Total 4 61 66 1 11 26.5 8.5 10 188
Forest Type- Wet
Koala 154 155 156 157 158 165 166 169 Total Ruth 1 2 22.5 25.5 Simon 6 5 1.5 2 3.5 2 20 Wayne 1 1 1 3 Bob 2 5 4.5 5.5 1 7.5 6.5 32 Roberta 10 2 6 18 Robert 2 2 Michelle 3 0.5 3.5 Total 9 11 1 21 33.5 1 11 16.5 104
63 Woolybutt, 112 Silvertop Ash, 114 Silvertop Ash-Stringybark, 121 Blueleaved Stringybark, 123 Coastal Stringybark, 132 Stringybark-Gum, 150 Messmate, 152 Messmate-Gum, 154 Brown Barrel, 155 Brown Barrel-Gum, 156 Brown Barrel/Messmate-Ash, 157 Yellow Stringybark-Gum, 158 Southern Blue Gum, 165 Gully Peppermint, 166 River Peppermint, 169 Yellow Stringybark. 1 Where the floristic composition of plots did not fit a single Forest Type, two FT' s were nominated and a frequency of observation of 0.5 was assigned to each type. (From Jurskis and Potter (1997)
Table ill describes the floristic composition of the forest overstorey in a number of home ranges. It shows that one home range (Allan) is floristically quite different to the other seven which mostly have similar proportions of the four or five common species but could be split into two or three separate groups on the basis of associate species such as E. fastigata, E. elata and E. longifolia.
77
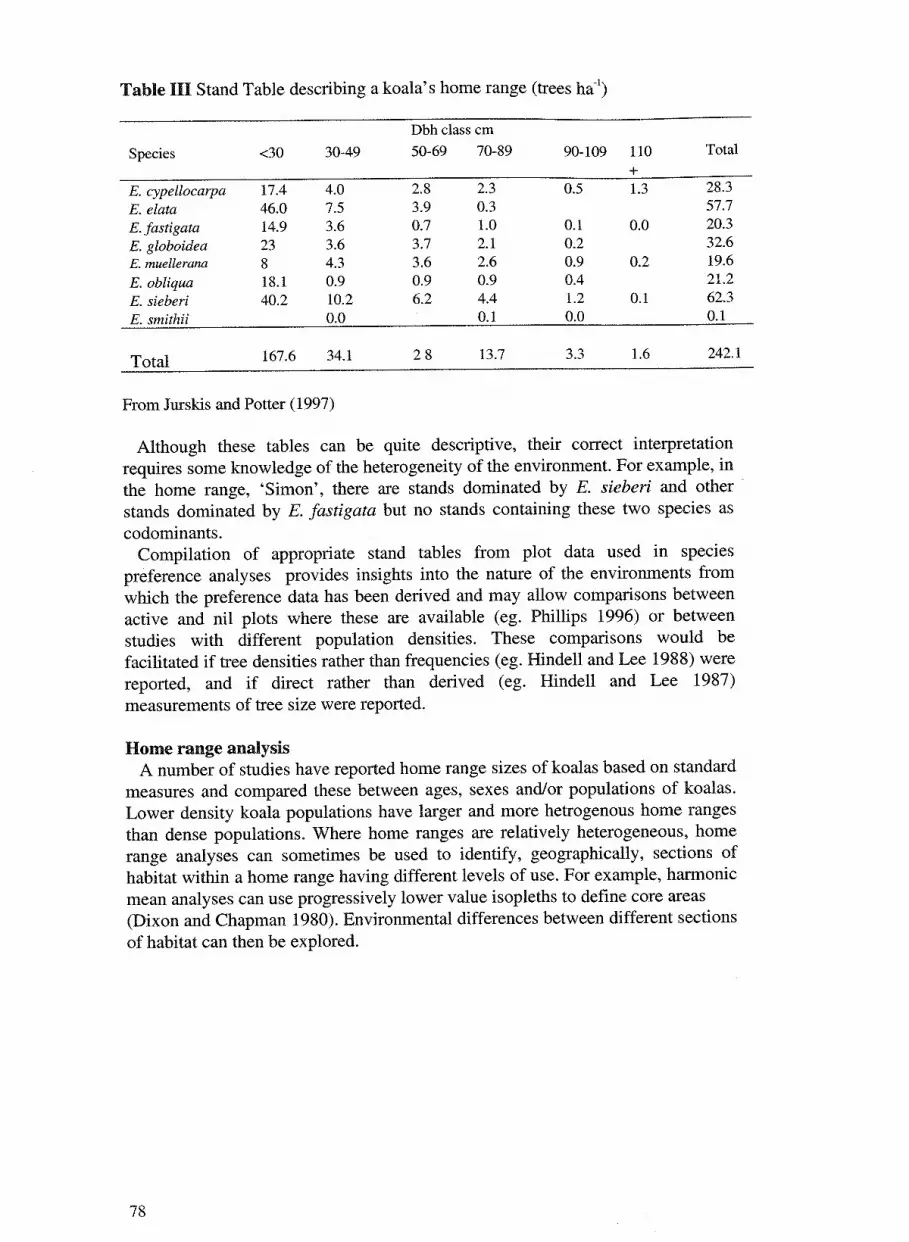
Table HI Stand Table describing a koala's home range (trees ha-1)
Dbhclass em
Species <30 30-49 50-69 70-89 90-109 110 Total
+ E. cypellocarpa 17.4 4.0 2.8 2.3 0.5 1.3 28.3
E. elata 46.0 7.5 3.9 0.3 57.7
E. fastigata 14.9 3.6 0.7 1.0 0.1 0.0 20.3
E. globoidea 23 3.6 3.7 2.1 0.2 32.6
E. muellerana 8 4.3 3.6 2.6 0.9 0.2 19.6
E. obliqua 18.1 0.9 0.9 0.9 0.4 21.2
E. sieberi 40.2 10.2 6.2 4.4 1.2 0.1 62.3
E. smithii 0.0 0.1 0.0 0.1
Total 167.6 34.1 28 13.7 3.3 1.6 242.1
From Jurskis and Potter (1997)
Although these tables can be quite descriptive, their correct interpretation
requires some knowledge of the heterogeneity of the environment. For example, in
the home range, 'Simon', there are stands dominated by E. sieberi and other
stands dominated by E. fastigata but no stands containing these two species as
codominants. Compilation of appropriate stand tables from plot data used in species
preference analyses provides insights into the nature of the environments from
which the preference data has been derived and may allow comparisons between
active and nil plots where these are available ( eg. Phillips 1996) or between
studies with different population densities. These comparisons would be
facilitated if tree densities rather than frequencies ( eg. Hindell and Lee 1988) were
reported, and if direct rather than derived ( eg. Hindell and Lee 1987)
measurements of tree size were reported.
Home range analysis A number of studies have reported home range sizes of koalas based on standard
measures and compared these between ages, sexes and/or populations of koalas.
Lower density koala populations have larger and more hetrogenous home ranges
than dense populations. Where home ranges are relatively heterogeneous, home
range analyses can sometimes be used to identify, geographically, sections of
habitat within a home range having different levels of use. For example, harmonic
mean analyses can use progressively lower value isopleths to define core areas
(Dixon and Chapman 1980). Environmental differences between different sections
of habitat can then be explored.
78

Figure II shows an example of a koala's home range (Harmonic Mean 60% isopleth) compared with the pattern of logged and unlogged coupes. A chi-square analysis of the Minimum Convex Polygon (MCP) home range indicated that this koala 'preferred' logged coupes (Jurskis and Potter 1997). The Harmonic Mean (HM) analysis also indicated that the pattern of logged coupes was shaping the core habitat since there were two core areas centred on logged coupes and the shape of the larger one was extended in the direction of other logged coupes (Jurskis and Potter 1997).
Figure III is an example of a home range (HM 70%) which suggested that the koala avoided ridgetops and/or roads. There are a number of alternative hypotheses which could explain the observed pattern of home range use. For example, the koala may have avoided the noise, dust and human activity associated with the roads; it may have preferred to eat the tree species associated with the gullies or it may have preferred the microclimatic conditions produced by the topography and forest structure in the gullies. Whatever the underlying cause, identification of core home range areas and comparison of these with other parts of home ranges provides for broader consideration of habitat use than tree use analyses alone.
1000 0 1000 Meters
e KOALA LOCATIONS
-- HARMONIC MEAN 70% ISOPLETH
LOGGED COUPE
UNLOGGED COUPE
................. PRIVATE PROPERTY
Figure II Home range of a koala at Eden compared with logging pattern
79

Relative to the volume of literature on the ecology of koalas there seems to have been little attention paid to home range analyses in relation to environmental factors. Mitchell (1990) analysed home ranges of koalas in great detail. Although
he related home range sizes to the density of large trees, he concentrated more on koalas' social activities than on the environmental factors which may have been
shaping home ranges. This is not surprising given the relative uniformity of the study site (which was described as 9 ha of manna gum woodland) compared with
sites carrying low density populations. Mitchell (1990) suggested that larger home
ranges reported in other studies may be a result of environmental heterogeneity and sparsity of preferred trees.
Cork (1995) was concerned that home range analyses at Eden represented habitat use by widely separated individual koalas. He suggested that more
intensive observation of radio collared koalas would result in the detection of
additional koalas and allow better understanding of the spatial use of habitat by populations rather than individuals. Cork's (1995) suggestion did not recognise
that low density populations comprise widely separated individuals, and that
social activity is very limited (Jurskis and Potter 1997). Home range overlap was
observed at Eden and also inferred by the presence of breeding females (Jurskis and Potter 1997). Where home ranges are large, however, home range overlap
does not infer close proximity or social contact between individual koalas.
1000 0
e KOALA LOCATIONS
- - - HARMONIC MEAN 70% ISOPLETH
- - HARMONIC MEAN 90% ISOPLETH
MAJOR ROAD (SEALED)
MINOR ROAD (UNSEALED)
1000 Meters
Figure III Home range of a radio collared koala at Eden compared with
topography and road network
80

Melzer (1995) found that koalas at Norwood Creek did not use their home ranges evenly. Their ranges included nodes of concentrated activity amongst larger areas of infrequently used habitat. Although Melzer (1995) described the floristics and structure of the vegetation within these nodes, he did not describe the vegetation outside the nodes, and therefore was unable to fully explore the factors shaping the core home ranges.
The smaller range of environmental variation in the habitats of the denser koala populations that have typically been studied provides less opportunity for the type of home range analyses that have been reported by Jurskis and Potter ( 1997). Nevertheless home range analyses would potentially provide insights into use of habitat according to factors other than tree species. For example, floristics appeared to have less influence than forest structure on the shape of the core home range shown in Figure II, since the nodes of activity included contrasting floristic associations (pers. obs. author).
Regional surveys Kavanagh et al. (1995) carried out a regional fauna survey covering 291 forest
sites in north eastern New South Wales using a broad geographical and logging history stratification. This survey demonstrated that koalas were associated with heavily logged forests at lower elevations. Unfortunately, sufficient numbers of koalas have not been detected in regional surveys at Eden to allow these types of analyses.
The difficulties in accumulating ecological data relating to the low density Eden koala population have led to the use of incidental records in broad scale comparisons of regional koala distribution and environment. For example, Jurskis and Potter (1997) compared incidental koala records with the distribution of old growth forest, and concluded that koalas at Eden are not associated with old growth forest. Jurskis et al. ( 1994) compared the distribution of incidental records with the habitat predictions of Norton and Saxon (1993), which assumed a preference for wetter areas. As there was virtually no coincidence, Jurskis et al. (1994) concluded that koalas were associated with drier forests. Lunney et al. ( 1997) reported a postal survey of incidental koala records. They compared the regional distribution of koala records with the modelled distribution of vegetation types, and concluded that the greatest numbers of records came from some of the dry forest types. Cork (1995) suggested that apparent associations of koalas with dry forest types at Eden could be an artefact of better detectability in these types. However, taken together, the evidence from plots, home range analyses and incidental records at Eden as well as evidence from other areas (CSIRO 1996) supports an association of koalas with drier forests.
Lunney et al. (1997) also compared incidental koala records against land tenure and, surprisingly, concluded that neither National Parks nor freehold land were major reservoirs of koalas, even though 46% of their records were from these lands. A particular problem with using incidental records to define koala distributions is uneven 'search effort' (Cork et al. 1995). Differences in accessibility, visitation and survey effort can have marked effects on detection rates. Lunney et al. (1997) acknowledged that a relatively high survey effort contributed to the prominence of Tantawangalo State Forest in their koala
81

82
distribution data. In contrast they did not suggest that a lack of access, visitation
and survey was responsible for fewer koala records from National Parks compared
with State Forests. Although comparatively little effort has gone into surveying
National Parks, Jurskis and Potter (1997) reported high detection rates of koalas in
some National Parks during recent surveys by State Forest's staff. This suggests
that the conclusions of Lunney et al. (1997) may have been unduly influenced by
differences in sampling intensity. Many of the problems associated with haphazard sampling of small plots and
potential sampling biases could be overcome if more efficient survey techniques
were available. Increased detection rates achieved recently using modified call
playback techniques (Jurskis and Potter 1997) provide for some optimism that this
will happen. The gathering of data that would allow analysis of koala distribution
at a landscape scale could overcome some of the major difficulties in studying low
density populations.
Habitat modelling It is necessary to describe habitat and the way koalas use it in order to
understand their ecology and develop effective management strategies. One
potential application of such an understanding is to predict the distribution of
koala habitat in a region rather than having to survey the whole region. Even if the
distribution of koalas in a region is well known, construction and testing of a
model can confirm that understanding and give confidence about its application to
management of koalas. Three levels of habitat description have been discussed:
fine scale individual tree use, medium scale patch use in home ranges and broad
scale regional distribution of koalas. Studies of low density populations at Eden
suggest that all three levels need to be considered to fully understand koala
ecology and appreciate its implications for management. There has been a tendency for koala research to focus on tree species as the
primary determinant of koala ecology. However the great variety of preferred
species (both in the wild and in feeding experiments), and the conditional nature
of many conclusions regarding tree preferences, suggest that tree species is not
intrinsically important to koalas, except perhaps in marginal habitats (Jurskis
1996). Rather, tree species may be an indicator of more basic environmental
factors which determine carrying capacity. White and Kunst (1990) found that
seven out of fourteen tree species were preferred in their study of a moderately
dense population (Table I). They suggested that the importance of a few particular
tree species to koalas was overstated. Melzer (1995) found that about three
quarters of koala records at Norwood Creek came from the riverine Eucalyptus
tereticomis community or the E. crebra community on ridges and slopes. In more
than three quarters of the highly utilised tree clusters within home ranges, either
E. tereticomis or E. crebra was the dominant species, but Melzer (1995) provided
little evidence that use of these species was disproportionately high in relation to
their availability in those areas. Similarly Hindell and Lee (1987) demonstrated that high koala densities in the
Brisbane Ranges were associated with stands dominated by E. viminalis, but
Hindell and Lee (1988) found that most individual koalas did not show a species
preference. Of the koalas showing a preference, half preferred a species other than
E. viminalis. These studies suggest not so much that E. fibrosa, E. tereticomis ,

E. crebra and E. viminalis are important species, but that the landform/vegetation units in which they predominate are favourable environments where most eucalypts can produce fodder that is palatable to koalas. It has long been recognised that koalas are primarily a species of woodlands and drier forests (Phillips 1990; CSIRO 1996). Melzer (1995) found a negative correlation between foliage projective cover (FPC) and koala density, suggesting that woodlands have a higher carrying capacity than forests. Melzer (1995) also found that koala densities were correlated with relative density of 'preferred' species but that there was no correlation with absolute density of these species. He suggested that relative densities of 'preferred' species would influence the energy efficiencies of koalas through search effort. However clear evidence of preferences would be needed to support this explanation. Hindell and Lee (1988) only detected preferences when the relative density of the preferred species was low. This does not support the search effort proposition. An alternative explanation is that the floristic composition of a site is an indicator of environmental factors influencing habitat quality. A tree species may occupy a large environmental envelope within which its utility to koalas will vary. For example, at Eden, E. cypellocarpa is a preferred species in drier forests, but there is little evidence that koalas use the species in wet forests (Jurskis 1996). A florisitic association will usually have a smaller environmental envelope than an individual species and will therefore provide a better indication of habitat quality. Martin (1985) suggested that lowland coastal forests in Victoria had a higher carrying capacity for koalas than taller inland forests. Gordon et al. (1990) found higher densities of koalas in bimble box regrowth than in mature woodland of the same species, indicating that vegetation structure as well as floristics could influence koala densities.
Species preferences have not been used as a basis for modelling koala habitat at Eden for several reasons. The major preferred species in the escarpment forests, monkey gum (E. cypellocarpa ), occurs in both wet and dry forests, but seems to be preferred only in drier forests. Structural factors relating to trees and forests often appear to outweigh floristic factors in determining koalas' use of habitat. Species preferences apparently vary amongst different landforms .having different vegetation associations (Jurskis and Potter 1997). With the exception of areas lacking access and visitation, especially some of the National Parks, the broad regional distribution of koalas is reasonably well known (Figure IV). Koalas are dispersed widely throughout the drier forests in very low densities but may occur in slightly higher concentrations in two areas with a long history of disturbance at Tantawangalo-Yurammie and Bermagui-Murrah (Jurskis and Potter 1997). A successful model might predict koala habitat to occur throughout the forests in two density classes, one being measurable and the other too low to measure. In the absence of nodes of dense populations detailed modelling is potentially unproductive. Although a dense population is known from manna gum woodlands on the edge of the Monaro Tableland, none are known or expected to occur in the region's forests (Jurskis and Potter 1997).
Phillips et al. (1996) outlined the approach used by the Australian Koala Foundation (AKF) to model koala habitat. Tree preferences are examined within geomorphological and floristic strata. Although plots are randomly selected, only active plots (those containing koala faecal pellets) are used to examine tree preferences. A vegetation map or model is then used to categorise the regional koala habitat · according to the prevalence of preferred species. Floristic
83

84
associations dominated by preferred species with a density independent strike rate
are classed as primary koala habitat. Paradoxically, Phillips et al. (1996) reported the existence of areas of primary habitat without koalas in close proximity to areas of secondary or marginal habitat containing koalas. There is an element of
circularity in their analyses, since unoccupied plots do not contribute to their
predictive model of habitat but are then used as evidence of unoccupied primary habitat.
The AKF modelling appears to rely on an implicit assumption that the stratification used to guide the sampling incorporated appropriate environmental
factors at an appropriate scale to categorise koala habitat. A possible alternative approach would be to use field measurements of site characteristics to explore
differences between active and inactive plots within or across strata. Phillips et al. ( 1996) attributed the absence of koalas from predicted primary
habitat to localised extinctions and koala's social behaviour. They suggested a propensity for dispersing koalas to leave one aggregation of koalas and join another even if better habitat nearby is unoccupied. However Mitchell (1990) found insufficient social behaviour to describe any clear patterns from about 10,000 instantaneous observations of koalas. He found that most koalas neither sought nor avoided other koalas, but several animals shifted their home ranges, moving into areas from which other animals had been removed. Although Mitchell (1990) suggested that young males may disperse to increase their chances of mating, he also suggested that most associations between koalas were due to the activities of relatively few adult males, particularly dominant males. Lee et al.
( 1990) translocated a socially familiar group and a socially unfamiliar group of koalas. Individuals from both groups dispersed independently without maintaining an association (Lee et al. 1990). A need for social contact does not seem to be a particularly compelling explanation for an absence of koalas from optimal habitats. Further development of the modelling technique may be required to resolve the anomaly.
The impracticality of using random plot locations and the consequent lack of presence/absence data is a problem in studying low density koala populations. Studies of dense populations should use presence/absence data to explore possible relationships between habitat quality and factors other than presence of preferred tree species. Jurskis (1996) argued that some preferred tree species are distributed in tall wet sclerophyll forests as well as in poorer (for timber), drier forests, and that koalas typically prefer them on the drier sites. Prevett et al. (this volume) described differential use of E. obliqua by koalas at Ballarat according to
geological substrate. In plots located on newer volcanics, forty six percent of trees
were used compared to only nine percent from plots on Ordovician substrates.
STATE FORESTS
PINE FORESTS
NATIONAL PARKS
PRESERVED FOREST
PRIVATE PROPERTY
5 0 5 10 15 20 Kilometers ~~~~~~
Figure IV Distribution of koala records at Eden
MAJOR DRAINAGE
-- REGIONAL BOUNDARY
8 KOALA RECORDS
s
85

86
Habitat models should not rely solely on preferred species. Preferences for factors other than species ( eg. growth stage) may be important. Other
environmental and historical relationships should be closely examined, particularly where higher koala densities facilitate such analyses. A review of
faunal habitat modelling in south eastern Australia by Cork and Catling (1996) concluded that adequate modelling for arboreal fauna requires data on forest
structure. An integrated approach combining three levels of habitat assessment is more likely to be successful than an approach based solely on tree preference.
Broader scale regional assessment such as that of Kavanagh et al. (1995) could be used to inform a vegetation based model. Environmental or historical factors other
than localised extinctions may explain some of the apparent anomalies produced
by models which rely heavily on species preferences.
Conclusions Studies of low density koala populations in forests present difficulties. There is
considerable environmental variation within home ranges, data sets are small,
presence or absence of koalas at a site is difficult to determine, and it is not
possible to establish a measurable range of koala densities except, potentially, at a
regional scale. To understand the ecology of koalas in low density forest
populations it is necessary to make the most of limited data and to consider koala ecology at the tree level, the home range level and the regional level.
Studies of higher density populations and habitat models have tended to focus
on tree species as the primary determinant of habitat quality. Some apparent anomalies suggest that more use should be made of data on presence/absence and
differential use of home ranges to explore habitat at a broader scale than
individual trees. Studies of low density populations support this suggestion.
Consideration of broader environmental factors should help to explain the
variability in carrying capacity of eucalypt ecosystems which produces a wide
range of densities amongst koala populations. At Eden there may be an opportunity to increase knowledge of koala ecology at
a regional level if modified playback techniques permit a1;1 effective regional
survey to be carried out.
Acknowledgements Sandra Robbie typed the manuscript. Simon van Holst produced the figures. Pat
Prevett and an anonymous referee provided helpful criticism on an earlier draft of
this paper.
References Austin, I. (1994). A Bibliographical Reference on the Koala. (Australian Koala
Foundation: Brisbane.) Cork, S. (1995). Koala conservation in the south-east forests. Consultancy report.
NSW National Parks and Wildlife Service and State Forests ofNSW. Cork, S., Feary, S., and Mackowski, C. (1995). Koala Conservation in the South
East Forests: Proceedings of an Expert Workshop. NSW National Parks and Wildlife Service and State Forests of NSW.
Cork, S., and Catling, P. (1996). Modelling distributions of arboreal and grounddwelling mammals in relation to climate, nutrients, plant chemical defences

and vegetation structure in the eucalypt forests of southeastern Australia. Forest Ecology and Management 85, 163-175.
CSIRO. (1996). Murwillumbah Management Area Fauna Survey 1995. CSIRO Division of Wildlife and Ecology.
Dixon, K. R., and Chapman, J. A. (1980). Harmonic mean measure of animal activity areas. Ecology 6, 1040-1044.
Florence, R. G. (1996). Ecology and Silviculture of Eucalypt Forests. (CSIRO: Australia.)
Forestry Commission ofNSW. (1989). Forest Types in New South Wales. Research Note No. 17. (Forestry Commission of New South Wales: Sydney.)
Gall, B. C. (1980). Aspects of the ecology of the koala, Phascolarctos cinereus (Goldfuss), in Tucki Tucki Nature Reserve, New South Wales. Australian Wildlife Research 7, 167-176.
Gordon, G., McGreevey, D. G., and Lawrie, B. C. (1990) Koala populations in Queensland: major limiting factors. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 85-95. (Surrey Beatty & Sons Pty. Ltd: Chipping Norton, NSW.)
Hindell, M.A. (1984). The feeding ecology of the koala, Phascolarctos cinereus, in a mixed Eucalyptus forest. M.Sc. Thesis, Monash University. Clayton, Victoria.
Hindell, M.A., Handasyde, A., and Lee, A. K. (1985). Tree Species Selection by Free-Ranging Koala Populations in Victoria. Australian Wildlife Research 12, 137-144.
Hindell, M.A., and Lee, A. K. (1987). Habitat Use and Tree Preferences of Koalas in a Mixed Eucalypt Forest. Australian Wildlife Research 14, 349-360.
Hindell, M.A., and Lee, A. K. (1988). Tree Use by Individual Koalas in a Natural Forest. Australian Wildlife Research 15, 1-7.
Jurskis, V. (1996). Habitat Assessment, Tree Preferences, Environment and History, Where's the Connection? In 'Proceedings from A Conference on the Status of the Koala in 1996'. pp 31-40. (Australian Koala Foundation: Brisbane.)
Jurskis, V., Rowell, D., and Ridley, D. (1994). Survey techniques and aspects of the ecology of the koalas near Eden. Research Paper. No. 22. (State Forests of NSW: Sydney.)
Jurskis, V., and Potter, M. (1997). Koala Surveys, Ecology and Conservation at Eden. Research Paper No. 34. (State Forests ofNSW: Sydney.)
Kavanagh, R., Debus, S., Tweedie, T., and Webster, R. (1995). Distribution of Nocturnal Forest Birds and Mammals in North-eastern New South Wales: Relationships with Environmental variables and Management History. Wildlife Research 22, 359-377.
Kavanagh, R., and Bamkin, K. (1995). Distribution of nocturnal forest birds and mammals in relation to the logging mosaic in south-eastern New South Wales, Australia. Biological Conservation 71,41-53.
Lee, A. K., Martin, R. W. and Handasyde, K. A. (1990). Experimental translocation of koalas to new Habitat. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 299-312. (Surrey Beatty & Sons Pty. Ltd: Chipping Norton, NSW.)
Lithgow, K. A. (1980). Aspects of the feeding ecology of the koala Phascolarctos cinereus, in Victoria. Honors. Thesis, Monash University.
87

Lunney, D., Esson, C., Moon, C. Ellis, M., and Matthews, A. (1997). A
community-based survey of the koala, Phascolaractos cinereus, in the Eden
Region of South-eastern New South Wales. WildlifeResearch 24, 111-128.
Martin, R. W. (1985). Over browsing and decline of a population of the koala,
Phascolarctos cinereus, in Victoria III. Population Dynamics. Australia
Wildlife Research 12, 377-385. Melzer, A. (1995). Aspects of the Ecology of the Koala, Phascolarctos cinereus
(Goldfuss, 1817), in the Sub-humid Woodlands of Central Queensland. Phd
Thesis, University of Queensland. Melzer, A., and Lamb, D. (1994). Low density populations of the koala
(Phascolarctos cinereus) in Central Queensland. Proceedings of the Royal
Society of Queensland 104, 89-93. Mitchell, P. (1990). The home ranges and social activity of koalas- a quantitative
analysis. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G.
D. Sanson.) pp. 171-187. (Surrey Beatty & Sons Pty. Ltd: Chipping Norton,
NSW.) Mitchell, P. (1992). Koalas on Raymond Island. Distribution and Habitat Use.
Report to Australian (Koala Foundation: Brisbane.) Mitchell, P., and Martin, R. (1990). The structure and dynamics of Koala
populations -French Island in perspective. In 'Biology of the Koala'. (Eds. A.
K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 197-208. (Surrey Beatty &
Sons Pty. Ltd: Chipping Norton, NSW.) Norton, T. W., and Saxon, M. J. (1993). Predicted potential habitat of the Koala,
Phascolarctos cinereus in south-east New South Wales and recommendations
for field survey. Consultancy report to NSW National Parks & Wildlife
Service. Phillips, B. (1990). 'Koalas. The little Australians we'd all hate to lose'.
(Australian Government Printing Service Press: Canberra.)
Phillips, S., Callaghan, J., and Staines, D. L. (1996). The Koala Habitat Atlas
Field Survey Guidelines, Data Analysis & Habitat Modelling Details. In
Proceedings from A Conference on the Status of the Koala in 1996. (Australian
Koala Foundation: Brisbane.) Prevett, P. T., Pope, R., Callaghan, J., and Bailey, L. (this volume). The Koala Habitat
Atlas: Preliminary results for koala tree species preferences in the City of Ballarat
Local Government Area, Victoria. In The Management and Research of Non-urban
Koala Populations. (Eds. K. Lyons, A. Melzer, F. Carrick, and D. Lamb.) pp. 94-
97. (Central Queensland University Publishing Unit: Rockhampton.)
Reed, P. C., Lunney, D., and Walker, P. (1990). A 1986-1987 survey of the koala
Phascolarctos cinereus (Goldfuss) in New South Wales and an ecological
interpretation of its distribution. In 'Biology of the Koala'. (Eds. A. K. Lee, K.
A. Handasyde and G. D. Sanson.) pp. 55-74. (Surrey Beatty & Sons Pty. Ltd:
Chipping Norton, NSW.) White, N. A., and Kunst., N.D. (1990). Aspects of the ecology of the koala in
southeastern Queensland. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A.
Handasyde and G. D. Sanson.) pp. 109-116. (Surrey Beatty & Sons Pty. Ltd:
Chipping Norton, NSW.)
88

The Role of Research in Koala Management: Case Studies from South-East Queensland
Jim Thompson Koala Research Unit, Queensland Parks and Wildlife Service, PO Box 5116, Daisy Hill, Qld.
Abstract A comprehensive long-term management program has been established for koalas in
a 400 km2 area in south-east Queensland. This regional approach to management is required as there is a diversity of habitat, a wide variety of threatening processes and a regionally different nature of habitat utilisation by koalas in Queensland. These differences demand the development of localised management strategies. In this context, the role of a research program is to assess the long-term viability of the regional koala population, and develop, monitor and objectively evaluate management strategies. Results to date indicate variation in demographic parameters such as age structure, sex ratios and reproductive and mortality rates between local populations. Case studies reveal that there is insufficient data to assess some management strategies after two years, and highlight the need for long-term data sets and monitoring programs. Population modelling tools should be utilised to assist with assessing and prioritising management strategies and supporting the research of koalas in the region.
Introduction Conservation programs often commence when a population is in decline, when only
a small number of animals remain, or in response to political or public pressure (Caughley and Gunn 1996). In many situations ecological data are inadequate to develop management recommendations, and this can result in poor management decisions (Begon et al. 1996). It is the linking of research and monitoring to management that is fundamental to effective conservation. A successful example is the Lord Howe Island Woodhen, where detailed research and monitoring resulted in the feral pig (not cats, rats or habitat loss, as first thought), being identified as the major threat to the survival of the bird population (Miller and Mullette 1985). Appropriate management programs were then developed. In the longer-term, the benefits of management require assessment through ongoing monitoring programs (McCallum 1997), particularly focused on population trends (Caughley and Sinclair 1994).
In this paper these issues are addressed in relation to the Koala Coast Protection Plan, which was implemented in 1995. This is a comprehensive, government sponsored management program for the conservation of koalas in a region to the south-east of Brisbane. This regional approach to management in Queensland contrasts with the state-wide approach to management adopted in NSW, particularly in reference to land-use planning policies. This is significant because there is currently differing opinion as to the best approach to utilise legislation or planning schemes to protect koala habitat.
89

The major objective of this paper is to evaluate the progress of the Koala Coast
Protection Plan after 2 years, with a focus on the role of research in the management
of the koala in the region. Preliminary research results are reviewed in relation to the
robustness of the data and the timeframe needed for research to contribute to the
effective management of the koala. Case studies are presented, detailing the rationale
behind the adoption of particular management strategies and the role of research in
evaluating these strategies. These case studies address 3 major issues affecting koalas
in this region; (1) loss of habitat, (2) koalas being killed on roads by cars, and (3) the
rehabilitation of sick and injured animals. The management program and research aims are also assessed in the context of
whether they adhere to a recommended framework for threatened species management
(Possingham et al. 1993). The potential contribution of population modelling to the
development of conservation strategies for koalas in this region is discussed.
Regional Environment and Current Management Strategies The region, known as the Koala Coast, is approximately 400 km2 in area and is~
situated to the south-east of Brisbane. The area encompasses the mainland part of
Redland Shire, the western portion of Logan City and the southern part of the
Brisbane City Council area. The region contains large areas of remnant bushland
(open eucalypt forest), town and industrial precincts and significant urban
development. It is considered part of the greater Brisbane area. Records held in the Queensland archives confirm that koalas were abundant in the
area in the early 1900's (Outridge 1927). Koalas are now widely distributed
throughout the region in both urban and bushland areas (unpublished data). Population
estimates of between 3000 and 5000 animals are based on broadscale vegetation and
faecal pellet surveys by Pahl (1993) and population studies undertaken by White and
Kunst (1990) in the Sheldon area. Recent estimates suggest this figure may be as high
as 6000 koalas (unpublished data). Within the region detailed information of local
koala populations is confined to a few specific locations where population studies
have been conducted (White and Kunst 1990; Nyo Tun 1993; Hasegawa 1995).
The major threat to the survival of the koala in this region is loss of habitat due to
land development, and clearing for agriculture or other enterprises. In the non-urban
areas, 47% of the bushland has been cleared or contains only scattered vegetation
(Pahl 1992). Preservation of the existing habitat is the primary conservation focus.
Other documented threats to the koala population include the dangers posed by motor
vehicles and dog attacks. Previous studies, coupled with 8 years of incidental records of koala sightings
(Nattrass, unpublished data), local community knowledge of koalas, and hospital
records from sick and injured animals, provided a sound knowledge base for the
development of the current management program. This program, the Koala Coast
Protection Plan, was implemented by the Queensland Parks and Wildlife Service
(QPWS) and is a comprehensive package of initiatives, including:
• a research program targeted at the distribution, density and population dynamics
of the koala; • a geographic information system for mapping and modelling koala distribution in
the region and for assisting in the development of land-use planning policies;
• a region specific State Planning Policy addressing local land planning issues;
• the acquisition of selected bushland areas; • extension and public education programs;
90

• programs to reduce the numbers of koalas killed on roads; and • koala rehabilitation, involving the care and treatment of sick and injured koalas.
The greatest value of each of these initiatives lies in their integration with the other components of the Plan (Thompson 1996).
The Linking of Research and Management There are typically two approaches to linking research and management. The first is
to conduct research and develop management recommendations based on the results. The second is to implement 'best bet' management strategies based on available information, and utilise research to monitor the outcomes and redirect management programs where required. It is the second approach that has been adopted here.
Despite a sound knowledge of koalas in the region, the lack of comprehensive population data has resulted in some components of the Koala Coast Protection Plan being based more on the precautionary principle, and major public expectations, than on an objective assessment of management options. Consequently, the research program that runs parallel with the management program has a dual aim. Firstly, to monitor and evaluate existing management strategies, and secondly, to better understand the dynamics of koala populations to enable the development of further long-term management options.
Is there a more effective approach to conservation planning that objectively assesses management options, yet incorporates political and public expectations? Possingham et al. (1993) argue there is, and detail five steps for threatened species management: 1. collate existing information on a species; 2. generate, list and cost management options; 3. assess each management option using population viability analysis; 4. conduct sensitivity analyses to estimate the critical parameters in the system; 5. monitor the population and the impact of management programs.
The key process in this framework for threatened species management is population viability analysis (PVA), and research should be targeted at providing information for this process. PV A is a process to assess the likelihood of a population becoming extinct within a specified time and under particular circumstances (Shaffer 1981; Possingham et al. 1993; Boyce 1995). This approach is similar to that advocated by Lindenmayer et al. (1993), who state that the limitations of PYAs are far outweighed by the benefits. They believe PV As to be underutilised as a component of wildlife research and management programs in Australia. Possingham et al. (1993) believe that this approach to management need not be confined to threatened species and has application for all well studied species, those regarded as keystone or indicator species and species of special cultural value. However, Possingham et al. (1993) concede that in some circumstances management strategies may need to be implemented before several of the steps are initiated.
91

Research Methods and Preliminary Results The primary focus of the research program to date has been to establish baseline
data against which long-term management programs can be assessed. Monitoring
programs (ie. measuring the response of a population to particular management strategies) are liable to misinterpretation unless adequate baseline data have been
collected. The three fields of study commenced are distribution and density, comparative ecology and population dynamics. The following is a brief review of
results from 1996 and 1997.
Distribution and Density Three large-scale surveys of up to 24 bushland sites in the Koala Coast Region were
conducted in 1996 and 1997 (for methodology see Dique et al. this volume). The density of koalas recorded at all sites ranged from 0- 1.32 koalas ha-1
• On sites where
a zero density was recorded, koalas were found in subsequent surveys. These density
estimates are conservative as it is thought that approximately 16% of animals are missed during a survey (Dique et al. this volume). Density estimates are high, relative
to much of central and western Queensland (Melzer and Lamb 1994), but low when
compared with sites in Victoria where densities between 1 and 9 koalas per ha-1 have
been recorded (Martin 1985; Every 1986; Mitchell and Martin 1990). A detailed
analysis of the survey data and the methods employed can be found in Dique et al.
(this volume). Koalas have been found at all sites indicating that they remain widely
distributed throughout the region.
Comparative Ecology and Population Dynamics An intensive investigation of the ecology and population dynamics commenced in
1996. Koalas were captured at 3 sites and sexed, weighed and a number of
morphometric measurements taken. Details of the catching methodology are found in Dique et al. (this volume). Blood samples and chlamydia! swabs were taken from
some animals for genetic and disease testing. Each independent koala was eartagged
and fitted with a radio-collar made by Titley Electronics. Koalas were tracked regularly (at least once each fortnight), using hand held receivers (Telonics TR-2) and
aerials (Titley Electronics and Sirtrack Electronics) to provide information on
mortality, dispersal, home range size and tree usage. A total of 62 animals were captured and 60 animals radio-collared at 3 sites in June
November 1996. Clinical signs of Chlamydia were evident in 11 of the 21 animals
(52%) found at Site 3. This was higher than the incidence of Chlamydia at sites 1 and
2, which recorded 15% (n=20) and 0% (n=21) respectively. The mortality rate of the 60 radio collared animals until June 1997 was 17% (n=lO).
All animals were recaptured at Site 1 in May/June 1997. A total of 35 adult koalas
have now been captured at Site 1 during the two capture periods. During these 2 capture periods, 22 pouch or back young were found. The sex ratio of the dependent
young is 68% (n=15) male and 32% (n=7) female. The sex ratio of the adults is 34%
(n=12) male and 66% (n=23) female. Fecundity differs markedly between sites although caution must be taken interpreting these data due to the small sample sizes (Table I).
All animals have been regularly tracked for approximately 1 year. From the 60
radio-collared animals a total of 1200 tracking records have been obtained. Home range determinations for all animals are premature, as there have been too few
92
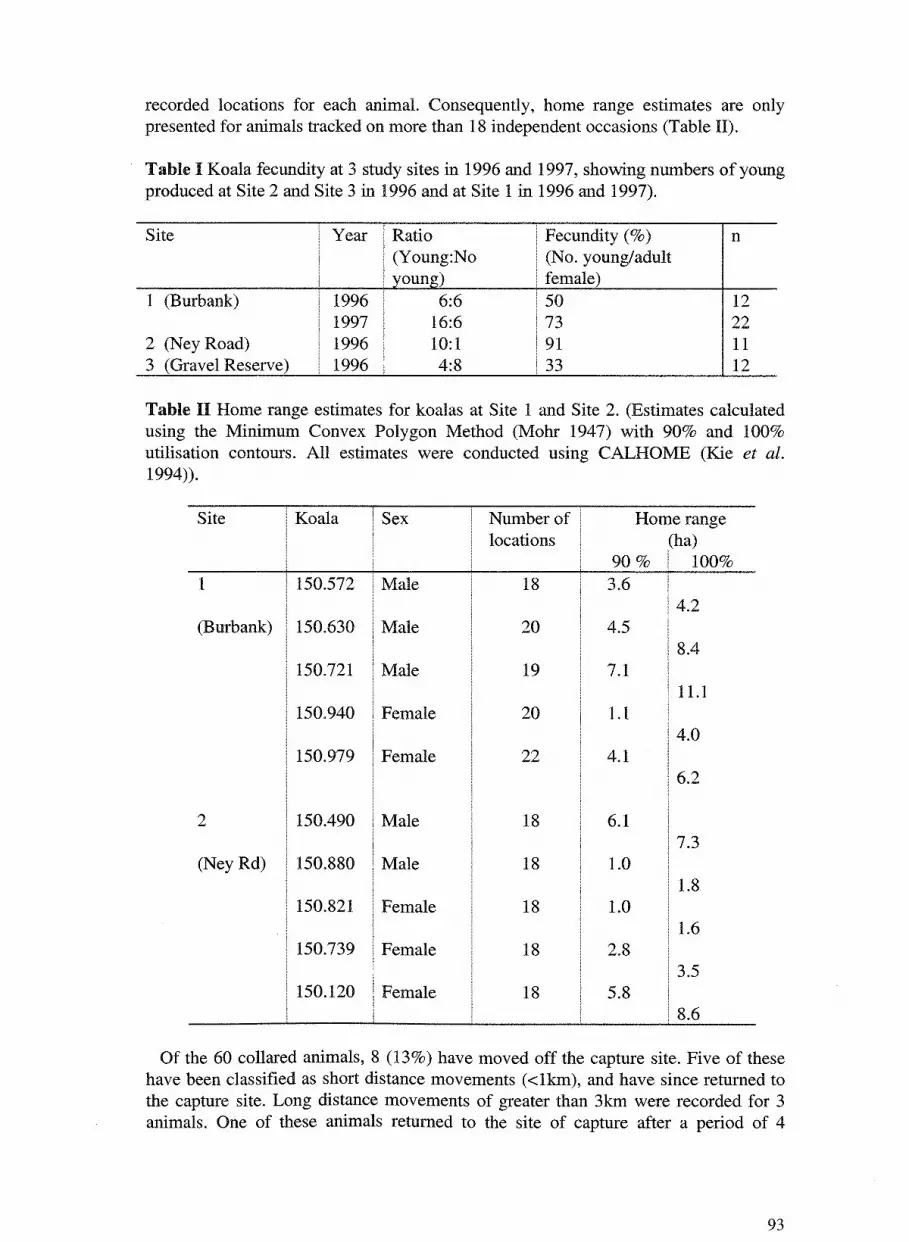
recorded locations for each animal. Consequently, home range estimates are only presented for animals tracked on more than 18 independent occasions (Table II).
Table I Koala fecundity at 3 study sites in 1996 and 1997, showing numbers of young produced at Site 2 and Site 3 in 1996 and at Site 1 in 1996 and 1997).
Site Year Ratio Fecundity (%) n (Young:No (No. young/adult young) female)
1 (Burbank) 1996 6:6 50 12 1997 16:6 73 22
2 (Ney Road) 1996 10:1 91 11 3 (Gravel Reserve) 1996 4:8 33 12
Table II Home range estimates for koalas at Site 1 and Site 2. (Estimates calculated using the Minimum Convex Polygon Method (Mohr 1947) with 90% and 100% utilisation contours. All estimates were conducted using CALHOME (Kie et al. 1994)).
Site Koala Sex Number of Home range locations (ha)
90% 100% 1 150.572 Male 18 3.6
4.2 (Burbank) 150.630 Male 20 4.5
8.4 150.721 Male 19 7.1
11.1 150.940 Female 20 1.1
4.0 150.979 Female 22 4.1
6.2
2 150.490 Male 18 6.1 7.3
(Ney Rd) 150.880 Male 18 1.0 1.8
150.821 Female 18 1.0 1.6
150.739 Female 18 2.8 3.5
150.120 Female 18 5.8
~
Of the 60 collared animals, 8 (13%) have moved off the capture site. Five of these have been classified as short distance movements ( <lkm), and have since returned to the capture site. Long distance movements of greater than 3km were recorded for 3 animals. One of these animals returned to the site of capture after a period of 4
93

months, 1 animal has died (drowned) and one has moved to a new area 7km from the
point of capture.
Case Studies In the following section the major management strategies of the Koala Coast
Protection Plan are evaluated as case studies. This evaluation is based on:
• the reasons for developing these strategies; • the role of research in evaluating these strategies in the short and long term;
• whether results collected to date are sufficient to assess outcomes; and
• whether these strategies are consistent with the framework outlined earlier and
described by Possingham et al. (1993).
Case Study 1 Habitat Conservation The loss of remaining habitat is regarded as the most critical conservation issue
facing the koala in the Koala Coast Region (Pahl1993). The need to conserve habitat
appears self evident, yet the process to address this need is problematic. Protection of
habitat can be achieved through property acquisition, conservation agreements and/or
planning controls (Thompson 1996). Property acquisition has considerable appeal and
certainty, and significant areas in the Koala Coast Region have been recently
purchased by State and Local Governments as part of koala and bushland conservation
programs. Similarly, the long-term protection of privately owned bushland through
conservation agreements has commenced. However, the acquisition or protection by
agreement of all suitable land is impractical and can be both slow and prohibitively
expensive. Consequently, planning controls were needed. State Planning Policy (SPP) 1/95, released in December 1995, was reviewed and re
released as SPPl/97 in October 1997. The intention of this policy is to conserve and
protect extensive koala habitat from inappropriate land use and development
(Queensland Government 1997). The policy is used to provide direction and support
to governments and others making planning decisions that impact on koala habitat
(Thompson 1996). The role of research in monitoring the effectiveness of land use planning policies is
both short and long term. The principal test of SPPl/97 in the Koala Coast Region is
the long-term protection of koala habitat. This can be measured by comparing
available habitat in 1995 with available habitat at different times in the future and
assessing whether the rate of land development, or the loss of good quality habitat, has
slowed. This is principally a GIS based investigation that requires a classification of
land and habitat types and is currently underway. A further long-term assessment of impact of planning schemes is the impact on
koala numbers. If the viability of the koala population is closely linked to available
habitat, there should be a corresponding decrease in the koala population following
the removal of habitat. This is best measured by extensive and detailed knowledge of
population distribution and density over a long period. However, population estimates
may also be affected by other factors, including disease risks, habitat quality and local
variation in habitat use. Detailed surveys, as outlined by Dique et al. (this volume), provide the baseline data
for comparison with further surveys that would need be conducted in 5, 10 and 20
years time. These surveys provide the best quantifiable method for determining
population trends. Previous phone-in or questionnaire surveys conducted in the region
by local community groups and anecdotal data may provide indicators of distribution
94

change or range contraction. However, it is difficult to quantify these changes, as there has been no standardised or replicated approach taken to survey methodology.
In the short-term research can, and has, assisted by prioritising areas for conservation. Pressure for release of land for development continues, and by determining the conservation value of available habitat, informed decisions can be made regarding the development of some areas. At this point conservation value is being crudely measured in the short term by koala density and population size. Based on surveys conducted across the region, during the two years of this project, it has been possible to prioritise areas for conservation using these two estimates. Put simply, where a requirement is made to assess two or more areas, the area with the greatest number of koalas is given the highest conservation value. Care should be taken when evaluating koala habitat in this manner, as population density can be a misleading indicator of habitat quality (Van Horne 1983). In the longer term, vegetation surveys, koala movement behaviour, and an understanding of population change (ie. measured as rate of increase), are needed to properly rank the conservation value of a range of areas.
This approach to linking management with research is similar to that advocated by Possingharn et al. (1993) and Boyce (1992). However, PVAs were not utilised to prioritise habitat conservation over other management strategies, and the challenge now is to collect information for this process. A comprehensive assessment requires PV A modelling to incorporate the spatial structure of the koala and its habitat (Akcakaya 1994). This information is currently incomplete as vegetation surveys are still to be conducted and no koala surveys have occurred in urban or semi-urban areas.
In summary, research programs that focus on collecting robust information on the distribution and density of species at a range of sites can have an immediate role in ongoing and reactive habitat conservation issues. However, for longer term impacts data collected over two years provides baseline density estimates only and does not provide the detail regarding population change or population demographics necessary for informed habitat protection.
Case Study 2 Koala Protection on Roads (Speed Zones) Approximately 300 koalas in the Koala Coast region are hit on roads by cars each
year. Eighty percent of these animals do not survive (Nattrass, unpublished data). It is postulated that reducing traffic speed in areas where many koalas are hit by cars will lower traffic-related mortality of koalas (Thompson 1996). Records collected by QPWS from 1986-1994 indicated that the majority of road accidents occurred at night between August and December each year. Consequently, a trial was established to assess the impact of differential speed limits (ie. lower night-time speed limit). This trial began in 1995, and ran between August and December until1999.
This management strategy is an example of an attempt to combat a known threat to koalas in the region. This strategy is supported by the general public, as it addresses an issue of frequent public concern. From a public education perspective, it further reinforces the message that koalas and humans are occupying a shared resource that must be managed for both species. From the database of road accident locations from 1990-1994, 4 sub-arterial roads with high koala mortalities were selected for the speed zone trials. Koala mortality is also being closely monitored at an additional 4 roads as controls. Should this trial prove successful, differential speed zones could be established on a number of other roads in the region.
95

The role of research is twofold. Firstly, in the short term to assess if the introduction of speed zones reduces the numbers of koalas killed on individual roads and in the region. Secondly, to assess the impact on the regional population of approximately 300 additional deaths each year. Over the first three years of the trial (95-97) there was no significant change in koala mortality on the treatment roads when compared with figures from 1994 (Table III). This was despite an increase over this period in the number of koalas killed on roads throughout the region (Figure I), suggesting that the results from the treatment roads were inconsistent with regional trends. However, results from control roads also showed no change, indicating that the higher regional koala mortality figures were the result of more deaths on other roads in the region.
Table HI Numbers of koalas hit by cars on speed zone roads and control roads between 1994 and 1997 (speed zones implemented in 1995)
(General Linear Model - Repeated Measures Analysis of Variance (SAS Institute 1988) showed no Year*Speed interaction (F=0.49, p>0.74, df=4,24) nor Year (F=0.56, p>0.69, df=4,24) or Speed (F=0.52, p>0.49, df=1,6) effects.
Roads 1 Year I 1994 1995 1996 1997
Speed Zone Roads Delancy St 1 3 0 0 Finucane Rd 14 10 12 9 Old Cleveland Rd, 3 1 0 8 East Sturgeon St 3 4 2 4 Total 21 18 14 21
Control Roads Boundary Rd 8 8 7 14 NeyRd 4 3 2 2 Redland Bay Rd 9 8 11 8 Wellington St 8 3 4 0 Total 29 22 24 24
Results from traffic monitoring showed that the impacts of the differential speed zones on traffic speeds were also negligible (Main Roads 1998). There was no overall significant decrease in vehicle speeds due to the speed zones, despite the statutory requirement to adhere to the posted speed limits.
At this point these results are inconclusive. There appears to be no decrease in koala mortality on the selected roads, although figures from these roads are inconsistent with regional trends. Additional information is required on which roads in the region have exhibited higher koala mortality and under what conditions motorists will adhere to differential speed limits. Alternative ways need to be investigated to slow cars down. Other factors that also confound the results of this experiment include; increased traffic volume on the roads, changes in vegetation surrounding the roads, and weather conditions.
For the management of the koala in the longer term, the impact of road accident deaths on the koala population needs to be clarified. Can the regional population
96

continue to sustain approximately 300 deaths per year or are these animals surplus to the population needs? This question may be best answered by incorporating these figures in a PVA assessment as advocated by Possingham et al. (1993), or in other population growth models. A potential reduction in deaths can then be prioritised as a management option against other strategies. However, unless over the next 2 years of this trial koala deaths can be reduced on these roads, this will not remain a viable management option. If car speeds and koala deaths can be reduced, consideration must be given to the number of roads suitable for this program. It may not be practical, or the best use of resources, to implement speed zones on enough roads to significantly reduce annual mortality rates. As it is, only approximately 20 deaths occur annually on the selected treatment roads.
N o. of K o a Ia s h it by cars
A
'"
Year
Figure I Number of koalas hit by cars on roads in the Koala Coast Region between 1994 and 1997.
Finally, caution must be taken when implementing management strategies such as differential speed zones, as the results of these trials can be difficult to assess. Consistent and standardised records must be obtained for many years to measure the success of these programs. Ideally, this would include before and after records and similar assessments of nil-treatment areas. As the number of animals killed or injured on individual roads in the Koala Coast Region is mostly less than 20 per annum, many years of data are required before the success or failure of the program can be established.
Case Study 3 Koala Rehabilitation The rehabilitation of sick and injured koalas from the Koala Coast has been ongoing
for many years. Much of this work has been undertaken by wildlife carers, under the direction of staff from QPWS. QPWS recognised the need to provide a facility to treat these koalas and established the Moggill Koala Hospital in 1990. Five hundred and fifty one koalas from the Koala Coast Region were treated in 1995, and 659 in 1996 (Nattrass, unpublished data). These numbers have continued to increase since 1996 (unpublished data), resulting in the known loss of approximately 550-600 koalas per year from this region. The major focus of this work is to return as many koalas to the wild as possible.
The rescue and movement of koalas within the Koala Coast is undertaken by QPWS staff and volunteers from the Logan City and Redland Shire areas. The koala ambulance provides a 24 hour service and is jointly funded by State and Local Government as part of the Koala Coast Protection Plan.
97

All sick, injured and orphaned koalas are transported to the Moggill Wildlife Hospital.
The number of animals treated is beyond the capabilities of volunteer or carer groups
and in many cases the treatment is specialised. How can we assess the value of this program beyond the significant public demand
and expectation for such work? Firstly, this significant investment has a measurable
outcome in terms of the survival rate of otherwise doomed animals. In 1995-96,
approximately 24% of all koalas (286 animals over both years) survived. This would
appear to be a significant contribution to the local population, although the long-term
fate of rehabilitated animals requires further investigation. To prioritise resources, the
cost per surviving animal can be compared with the costs of other management
options, including the future expansion of speed zones. Secondly, can the regional koala population sustain 500-600 deaths per year in the
long-term? Preliminary results showed a wide variation in fecundity rates between
local populations. If we assume a population size of 5000 animals in the region and an
even sex ratio, then 20% of all females must breed successfully to replace the known
number of dead animals. Based on preliminary results that show fecundity rates of
between 33% and 91%, this seems possible. However, it is not known how repeatable
these figures are, why there are differences between local populations, and what
proportion of these females are sexually mature. These data provide a good
understanding of the major causes of mortality throughout the region, but only a basic
insight into the differences in mortality between areas. The relationship between the losses described above and the total losses from the
population per annum, is unknown. Research indicates that in some areas (particularly
bushland), less than 10% of all mortalities are reported to QPWS, as the bodies of
dead animals are not found. To fully assess the impact of mortality on the population
more accurate estimates of regional mortality rates are required.
As with the previous 2 case studies, 2 years data is inconclusive to properly assess
the effectiveness of koala rehabilitation. The wide variation in parameter estimates
between sites precludes simple calculations to determine the future of the population.
The question of long-term sustainability is best answered by continuing to collect
detailed population information from a number of sites over a number of years. This
information is the basis for modelling as recommended by Possingham et al. (1993).
Without this detail, modelling using PV As may fail to properly represent the regional
situation, particularly between local sites. When prioritising management strategies, certain weighting must be given to animal
welfare issues and the concerns of the public. Objective assessments or economic
analyses of the contribution of koala rehabilitation to conservation in this region
ignore public and political expectations. These assessments can also fail to recognise
the educational value of some of this work.
Discussion Scale of Management
The unique public appeal of the koala ensures that the status and management of the
species are the focus of ongoing debate. Much of the debate centres on the scale of
management and whether koalas should be managed in a local, regional or national
context. At the present time koalas are widely distributed in Queensland. They are
classified as common wildlife under the Nature Conservation Act 1992, and are not in
danger of extinction on a statewide basis. However, some populations throughout
Queensland (such as the population found in the Koala Coast), are locally threatened,
98

and specific management programs are needed in these areas. This differs from New South Wales where the koala is listed as vulnerable across the state under the Threatened Species Conservation Act 1995, and conservation issues have a statewide focus.
What then is the appropriate scale of management? The aim of the statewide approach in New South Wales is to ensure long-term survival over its present range (Department of Planning 1995), whereas in Queensland the focus is on the protection of locally threatened populations, as the overall distribution is regarded as relatively secure. I believe both approaches to habitat conservation are valid, yet for different reasons. The statewide approach used in New South Wales was developed because only general information on koala distribution and density is available. Consequently, a process was developed whereby all areas are assessed, and potentially valuable koala habitat identified before development proceeds. This does not restrict development in areas of little koala value, but provides a safeguard where there is little knowledge of the local koala population. The end point of this process is the development of a local management plan.
In areas such as the Koala Coast Region of Queensland, basic information on the koala population is known, and the major or potential threats to the koala have been identified. Management action cannot be delayed while further research is undertaken. Planning policies can be drafted which accurately delineate areas of critical habitat and are compatible with local government planning schemes. The starting point for this process is in effect a local management plan. In these areas management programs, coupled with further research and monitoring, provide the best hope for long-term conservation.
The approach in Queensland acknowledges that koalas face a wide range of different threats, and that the pressure on regional populations similarly varies, hence a regional approach best meets the requirements of these disparate groups. Given this regional approach, it is necessary to determine an appropriate size for a management unit. Management units can be delineated on the basis of genetic differentiation from other koala populations (Houlden et al. 1996), or by geographic fragmentation. In this case, the population in the Koala Coast is effectively isolated from other major populations by physical barriers such as rivers, highways and suburban development.
Development, Monitoring and Assessment of Management Programs The ecology and distribution of the koala is partly documented (White and Kunst
1990; Pahl 1993; Hasegawa 1995), however this work needs replication across the region and basic information on koala density and population dynamics is required. Management strategies based solely on anecdotal or incomplete evidence can be ill directed or impossible to monitor effectively (Caughley and Gunn 1996). Therefore, the objective for koala management in the Koala Coast should be to establish a program whereby natural resource management and scientific experimentation are integrated and progress concurrently. Fundamental to this is an understanding of trends in abundance, and the impact of changes in demographic parameters such as recruitment, dispersal and mortality (Brawn and Robinson 1996). These trends in abundance or population rates of change are necessary to properly evaluate the effects of perturbations or management programs (Walsh et al. 1995).
Management strategies based on an objective assessment and prioritisation of options developed from sound research, ignore the human dimension that is inextricably linked to the management of high profile animals, such as the koala. This
99

requires a balance between objectively targeted management options and public
expectation and ethical demands. The demand for action to address a well
documented, but poorly understood process, often dictates the management agenda.
This is partly the case for koalas in the Koala Coast Region. Given these difficulties, can a framework for threatened species management as
outlined by Possingham et al. (1993) be applied to koalas in the Koala Coast Region,
and is it practical to do so? I would argue that such a framework can be applied, albeit
with an altered order of implementation, as some management strategies are already in
place. The current management program incorporates many of the steps for threatened
species management outlined by Possingham et al. (1993), although little use has been
made of population modelling techniques. To progress, the current Koala Coast
management program requires a means to objectively assess and rank existing and
new management options. This process should utilise population modelling
techniques ( eg. PV As) and proceed concurrently with the existing program. The
critical step is to recognise how best to harness these modelling techniques to improve
conservation programs. Despite a growing number of advocates for the use of PV As
in wildlife management, others view their use with caution. Caughley and Gunn
( 1996) believe PV As should not be used as a diagnostic tool and that their output has
a substantial margin of error. Therefore their use is limited to a restricted set of
circumstances. Others acknowledge that PV As are not a panacea, but can be utilised
as a forecasting tool and as a means of identifying important parameters within a
system (Lindenmayer et al. 1993). This can then fee<;! back into the design of research
programs. The existing management strategies employed in the Koala Coast Protection Plan,
as outlined in the case studies presented, target obvious threatening processes. The
dilemma with developing management strategies using the framework outlined by
Possingham et al. (1993) is the time required to obtain sound scientific data on which
to assess options. Caution must be exercised, as much of the information required for
population modelling and viability analyses of koalas remains equivocal. Many would
argue that in the case of habitat loss, the problems are self evident, and remedial
action should be immediate. However, even when populations are on the brink of
extinction (particularly in local areas), the processes that have driven them there are
often unrecorded or unknown (Caughley and Sinclair 1994). Habitat destruction is
often blamed, but there can be a range of factors, many of which are overlooked.
The best way to assess the impact of a management program is through an
experimental manipulation with before and after monitoring at replicated treatment
and control sites. However, with small populations or protected species it is often
impossible to achieve this for both logistic and ethical reasons. Inadequate sample
sizes plague investigations into conservation biology (Caughley and Gunn 1996).
Consequently, most studies focus on the observation or monitoring of a number of
existing populations. Even adequate replication of sites can be difficult. This
immediately places caveats on the interpretation of the results and large data sets are
needed to elucidate any trends.
Data Collection and Estimation of Demographic Parameters
The focus of many previous studies both within and outside the Koala Coast has
been on the ecology, anatomy and diseases of the koala, and little attempt has been
made to document the population dynamics or viability of this species.
100

The exception to this has been in Victoria, where a number of detailed studies have produced a significant body of data from which to determine population status and change ( eg. Martin 1981; Martin 1985; Martin and Handasyde 1990; Mitchell and Martin 1990), and in central western Queensland where Gordon et al. (1990a and 1990b) studied koala populations over a 16 year period. However, caution should be taken when extrapolating this information from one area to another. Results to date indicate that even between local sites significant differences in parameter estimates are recorded.
The research conducted to date is limited in its direct application to the development of alternative management options, as it is short-term data based on small sample sizes. Population trends or differences recorded in demographic parameters between or within areas are speculative, and this work requires replication over time. While it is difficult to determine a minimum timeframe required to collect robust baseline data as this depends on the annual variation within each parameter, it is possible to say that two years is generally insufficient to meet these needs.
The data collected to date is relevant to the day-to-day management of specific sites where detailed information is available on local population size and structure, tree and habitat use and the health and reproductive status of the koalas. Knowledge of the distribution and density of koalas across the region also has relevance to comparative assessments between sites for issues such as development applications. However, density information from a range of different sites suggests that predictions of regional population sizes based on single site studies (Pahl 1992) are flawed.
Finally, a cautionary note on the research of high profile species. There is a public expectation that research and monitoring will provide an immediate assessment of management programs. This can create pressure on the conduct of a research program that results in a refocussing of effort into areas of short-term gain (ie. surveys in new areas). This must be avoided if long-term benefits, which come from detailed studies, are to be achieved.
Conclusion The management of existing populations of wildlife should be commenced while the
population remains relatively stable, and management strategies - not recovery plans - can be implemented. To date there has been insufficient time to conduct the necessary research to properly assess management programs. This research program should provide, after 5 years, a useful study of population dynamics of koalas for south-east Queensland. This timeframe should allow for seasonal or unexpected impacts to be properly evaluated.
The work will benefit greatly from the application of population viability analysis as a means of assessing and prioritising management strategies (Boyce 1992). These techniques can assist with, and have been suggested as, a fundamental component of a framework for improving the management of threatened species (Possingham et al. 1993). These techniques should now be utilised to support the management and research of koalas in the Koala Coast.
101

Acknowledgments I thank all members of the Koala Research Unit (David Dique, Guy Penfold, Ros
Leslie, Deidre de Villiers and Harriet Preece) for their input into the current research program, including the collection, collation and analysis of data and for discussions
related to many of the topics raised in this paper. I thank Ric Nattrass for access to unpublished data and Greg Gordon for reviewing the manuscript. Comments from two anonymous referees greatly improved the manuscript.
References Akcakaya, H. R. (1994). GIS enhances endangered species conservation efforts. GIS
World 7(11), 36-39. Begon, M., Harper, J. L., and Townsend, C. R. (1996). 'Ecology'. (Blackwell Science:
Boston.) Boyce, M. (1992). Population viability analysis. Annual Review of Ecological Systems
23, 481-506. Boyce, M. (1995). Population viability analysis exemplified by models for the
northern spotted owL In 'Statistics in Ecology and Environmental Monitoring'. pp 3-18.
Brawn, J.D., and Robinson, S. K. (1996). Source-sink population dynamics may
complicate the interpretation oflong-term census data. Ecology 77(1), 3-12. Caughley, G., and Gunn, A. (1996). 'Conservation Biology in Theory and Practice'.
(Blackwell Science: Cambridge.) Caughley, G., and Sinclair, A. R. E. (1994). 'Wildlife Ecology and Management'.
(Blackwell Science: Cambridge.) Department of Planning. (1995). State Environmental Planning Policy No 44- Koala
Habitat Protection. Circular No B35, 22 March. Dique, D., Penfold, G., Thompson, J., Leslie, R., and Preece, H. (this volume). Koala
distribution and density in south-east Queensland: The accuracy and precision of koala surveys. In 'The Management and Research of Non-urban Koala Populations'. (Eds. K. Lyons, A. Melzer, F. Carrick, and D. Lamb.) pp. 81-93. (Central Queensland University Publishing Unit: Rockhampton.)
Every, K. R. (1986). Evaluation of a decline in a population of the koala, Phascolarctos Cinereus (Goldfuss) in Ventnor Reserve, Phillip Island, Victoria, by
means of a triple count technique. Australian Wildlife Research 13,517-525.
Gordon, G., McGreevy, D. G., and Lawrie, B. C. (1990a). Koala populations in Queensland: major limiting factors. In 'Biology of the Koala'. (Eds A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 85-95. (Surrey Beatty and Sons Pty Ltd: Sydney.)
Gordon, G., McGreevy, D. G., and Lawrie, B. C. (1990b). Koala population turnover and male social organization. In 'Biology ofthe Koala'. (Eds A. K. Lee, K. A.
Handasyde and G. D. Sanson.) pp. 189-192. (Surrey Beatty and Sons Pty Ltd: Sydney.)
Hasegawa, M. (1995). Habitat Utilisation by Koalas (Phascolarctos cinereus) at Point
Halloran, Queensland. Unpublished Masters Thesis, University of Queensland.
102

Houlden, B. A., England, P. R., Taylor, A. C., Greville, W. D., and Sherwin, W. B. (1996). Low genetic variability of the koala (Phascolarctus cinereus) in southeastern Australia following a severe population bottleneck. Molecular Ecology 5, 269-281.
Kie, J. G., Baldwin, J. A., and Evans, C. J. (1994). CALHOME Electronic Users Manual. July 1994. U.S. Forest Service, Albany, California.
Lindenmayer, D. D., Clark, T. W., Lacy, R. C., and Thomas, V. C. (1993). Population viability analysis as a tool in wildlife conservation policy: with reference to Australia. Environmental Management 17, 745-758.
Main Roads. (1998). Koala Speed Zones 1998 Report. Internal report prepared by the Traffic Engineering Section, Transport Technology Division, Main Roads: Brisbane.
Martin, R. W. (1981). Age-specific fertility in three populations of the koala, Phascolarctos cinereus (Goldfuss), in Victoria. Australian Wildlife Research 8, 275-283.
Martin, R. W. (1985). Overbrowsing and decline of a population of the koala, Phascolarctos cinereus in Victoria. ill. Population dynamics. Australia Wildlife Research 12, 377-385.
Martin, R. and Handasyde, K. (1990). Population dynamics ofthe koala (Phascolarctos cinereus) in southeastern Australia. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 75-84. (Surrey Beatty and Sons Pty Ltd: Sydney.)
McCallum, H. I. (1997). Rock-wallaby biology and management: Synthesis and directions for future research. Australian Mammalogy 19, 319-324.
Melzer, A., and Lamb, D. (1994). Low density populations of the koala (Phascolarctos cinereus) in Central Queensland. Proc. R. Soc. Qld 104, 89-93.
Miller, B., and Mullette, K. J. (1985). Rehabilitation of an endangered Australian bird: the Lord Howe Island woodhen Tricholimnas sylvestris (Sclater). Biological Conservation 34, 55-95.
Mitchell, P., and Martin, R. (1990). The structure and dynamics ofkoala populations French Island in perspective. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 97-108. (Surrey Beatty and Sons Pty Ltd: Sydney.)
Mohr, C. 0. (1947). Table of equivalent populations ofNorthAmerican small mammals. Am. Midi. Nat. 37,223-249.
Nyo Tun, U. (1993). Re-establishment of rehabilitated koalas in the wild and their use of the habitat in Sheldon, Redland Shire, Southeast Queensland with particular reference to dietary selection. Unpublished MSc. Thesis. University of Queensland.
Outridge, R. (1927). Letter from the Queensland Producers' Association to the Honourable Acting Premier of Queensland. 20th July 1927. Queensland Archives.
Pahl, L. (1992). Assessment of impact on koalas of a proposed transport corridor in the Logan and Redland area. Occasional Paper 1992 1:1-50. University of Queensland, Gatton College.
103

Pahl, L. (1993). Final Report, Joint Regional Koala Habitat Project. Unpublished
Report.Possingham, H. P., Lindenmayer, D. B., and Norton, T. W. (1993). A
framework for the improved management of threatened species based on
Population Viability Analysis (PVA). Pacific Conservation Biology 1, 39-45.
Queensland Government. (1997). State Planning Policy 1/97 Conservation of koalas
in the Koala Coast. Department of Local Government and Planning. Commenced 3
October 1997. SAS Institute. (1988). SAS/STAT User's Guide. Release 6.03 Edition. SAS Institute
Inc. Cary, NC. Shaffer, M. L. (1981 ). Minimum population sizes for species conservation BioScience
31, 131-134. Thompson, J. (1996). Koala conservation in the Koala Coast- A government's role.
Proceedings from A Conference on the Status ofthe Koala in 1996 at Coolangatta
Queensland. Australian Koala Foundation, Brisbane. pp 91-96. Van Home, B. (1983). Density as a misleading indicator ofhabitat quality. J. Wild
Manage. 47(4), 893-901. Walsh, N. E., Griffith, B., and McCabe, T. R. (1995). Evaluating growth of the
Porcupine Caribou herd using a stochastic model. J Wild!. Manage. 59(2), 262-
272. White, N. A., and Kunst, N.D. (1990). Aspects of the ecology of the koala in
southeastern Queensland. In 'Biology ofthe Koala'. (Eds. A. K. Lee, K. A.
Handasyde and G. D. Sanson.) pp 109-116. (Surrey Beatty and Sons Pty Ltd: Sydney.)
104

Koala distribution and density in southeast Queensland: the accuracy and precision of koala surveys
David Dique, Guy Penfold, Jim Thompson, Ros Leslie and Harriet Preece Koala Research Unit, Queensland Parks and Wildlife Service, PO Box 5116, Daisy Hill, Qld.
Abstract The distribution and density of koalas in the Koala Coast Region, southeast
Queensland, were assessed as part of the ongoing management of the species. Koala surveys were conducted at 24 sites during two survey periods, February/March 1996 and August/September 1996. Daytime counts along strip transects were used to provide estimates of koala density. Density estimates from the surveys ranged from 0 to 1.3 koalas per hectare. Mark-resight studies conducted at two study sites suggested that daytime koala counts underestimate the size of the population by approximately 16%. Differences in density estimates between surveys appeared to reflect seasonal population changes at one site. Computer simulations of koala populations suggested that failing to detect koalas during a census has a small but noticeable effect on the precision of koala density estimates. Precision was lower when randomly placed transects were used to sample extremely clumped koala populations. Sampling intensity must be higher in dense populations compared to less dense populations if a similar level of precision is required.
Introduction In 1995, the Queensland Department of Environment established a Koala Research
Program to determine the conservation requirements of the koala (Phascolarctos cinereus) in the Koala Coast Region, 20km southeast of Brisbane. The objective of the research program was to provide information for effective conservation of the koala in the Region. Determining the distribution and density of koalas in the Koala Coast Region is an essential component of the research program, as knowledge of the size or density of a population is often essential to its effective management (Caughley and Sinclair 1994).
Many census techniques are available for conducting surveys of koalas. Such techniques include the use of faecal pellet surveys (Phillips and Callaghan 1996), questionnaire surveys (Phillips 1990), transect counts (Mitchell et al. 1988; Gordon et al. 1990; White and Kunst 1990; Melzer 1995), mark-resight estimates (Hasegawa 1995) and spotlight counts (Smith and Andrews 1997). For this project, transect counts were identified as the most appropriate method for counting koalas for two reasons; firstly, an estimate of absolute density was deemed more useful than an estimate of relative density, and secondly, the method had been commonly used in other studies enabling comparisons to be made to available data. However, there has been no attempt to quantify the errors associated with this technique.
Estimation of errors has two components; accuracy and precision. Accuracy is a measure of bias error and refers to the closeness of an estimate to the true value. Precision is a measure of sampling error or the repeatability of estimates (Caughley
105

and Sinclair 1994). To allow comparisons between koala populations at different sites and monitor koala population changes over time, both accuracy and precision should be considered.
The accuracy of density estimates obtained from koala surveys was quantified using
mark-resight methodology. Mark-resight is a modification of mark-recapture whereby a sample from a population is marked and released, and a subsequent survey is
undertaken to estimate the ratio of marked to unmarked animals in the population (Caughley and Sinclair 1994). The advantages ofmark-resight methods over recapture
techniques include efficiencies of costs and time for research teams and additionally, mark-resight is a relatively less invasive procedure for the study animals (Minta and
Mangel 1989). Given the sedentary nature of koalas during the day, mark-resight
methodology provides a useful technique for quantifying the accuracy of density estimates from koala surveys.
Computer simulations have been used to measure the precision of koala density
estimates by modelling the effects of parameter manipulations on density estimates. Different sampling strategies were applied to various koala distributions to construct a
distribution of density estimates. The flexibility given by simulations has enabled the
evaluation of the effects of manipulating various parameters (such as failure to detect
koalas and sampling intensity) on the precision of density estimates. In summary, the objectives of this study were to:
a) describe the distribution and density of koalas on a number of bushland sites in
the Koala Coast Region; b) use mark-resight methodology to determine the accuracy of koala surveys and;
c) use computer simulations to investigate the precision of density estimates
obtained during koala surveys.
Methods The Study Area
The Koala Coast Region covers an area of approximately 375km2 and is located
20km southeast of Brisbane. The Region is bordered by the Gateway Arterial and
Manly Road to the north, Logan River to the south, Moreton Bay to the east and the Pacific Highway to the west (Figure I). The Koala Coast Region contains areas of
extensive urban and industrial development, agricultural lands and remnant bushland.
The majority of the remnant vegetation in the Region has been classified as Nerang
Beenleigh open-forest alliance (Elsol and Dowling 1978). This vegetation type
characteristically contains a mixture of tree species, most commonly Corymbia
citriodora, Eucalyptus siderophloia, E. carnea, E. tindaliae, E. microcorys, C.
intermedia, E. resinifera and E. propinqua.
Koala Surveys Surveys for koalas were conducted in Summer/ Autumn 1996 and Winter/Spring
1996. Twenty-four sites, 8 remnant bushland sites and 16 bushland sites, were selected and surveyed for koalas. Aerial photos were used to select each site to ensure
an adequate geographical spread of sites across the region. The entire area of smaller sites (<40ha) was searched while larger sites (40-400ha) were sampled. Transects
were arranged systematically at all sites so that a minimum of 30% of each site was
sampled. Observers, equipped with a compass and binoculars, were spaced 15m apart, walked a fixed bearing and searched all trees within each transect for koalas.
106

Accuracy- Mark-resight Accuracy of koala surveys was assessed at two sites, Gravel Reserve (site 3 - 53ha)
and Burbank (site 13- 157ha) (Figure I). Koalas were captured throughout the Gravel Reserve, however search and capture was restricted to an area of approximately 50-60ha at Burbank. Koalas were detected at Gravel Reserve and Burbank using the same techniques as those employed during surveys. Once detected, koalas were captured, marked and released. Koalas were captured using a modified version of the 'noose and flag' technique (Bali and Delaney 1997). An 11 metre extension ladder and rock climbing equipment were used to ascend trees where koalas were located. A 6 metre telescopic aluminum pole, with a flag (plastic bag or cloth) was waved above the head of the koala encouraging it to descend. Once the koala was on the main stem of the tree, a lockable noose was placed over the animals' head, stopping it from reascending the tree. Koalas were captured at the base of the tree and then placed in a wire cage. Each captured koala was fitted with a radio transmitter attached to a synthetic collar (Titley Electronics, NSW).
Three surveys were conducted on consecutive days in February 1997 at each of the two sites where all koalas encountered were recorded. Koalas fitted with radio collars were recorded as 'marked' and those without were recorded as 'unmarked'. At the completion of each survey, all koalas at each site were radio tracked to determine the number of marked koalas present within the search area, and therefore, the proportion of marked koalas that were not detected during the survey. The population estimate for the search area at both sites was then calculated using the Peterson estimate (Caughley and Sinclair 1994).
Precision There are five factors that could affect the precision of koala density estimates
(Table I). Simulations were run for low, medium and high density koala populations, sampling either randomly or systematically, and at a range of different sampling intensities (5-90% ). Populations were either clumped or randomly distributed and the failure to detect koalas during field surveys was examined with different probabilities of detecting koalas. During the simulations, the probability of detecting a koala was specified and only koalas that were likely to be seen were included in the tally for koalas.
Five steps were involved in simulating the effects of sampling intensity and undetected koalas on the precision of density estimates (Table II). In step one, the simulations used parameter values which reflected the field surveys as closely as possible. In step two, either a random or clumped distribution was specified. Coordinates for random koala distributions were produced by using random number tables. To produce clumped distributions, the site was first overlaid into a defmed grid. Coordinates for clumped koala distributions were obtained by manipulating the proportion of koalas for each grid cell (Figure II). Within each grid cell the koalas were randomly distributed. Using this system it was possible to simulate koala populations with three degrees of aggregation. In step three, the number of transects used to sample koalas was based on sampling intensities ranging from 5-90%.
107

Table I Summary of factors affecting precision which have been incorporated into the
simulations
Factor
Koala density
Sampling strategy (systematic vs. random sampling)
Probability of koalas being undetected
Sampling intensity
Koala distribution (random/clumped)
Expected effect on precision
Precision is lower at high densities (Caughley & Sinclair 1994)
Systematic sampling increases precision (Caughley & Sinclair 1994)
Any variability in the percentage of koalas seen will tend to decrease precision
Increased sampling intensity will increase precision Clumping will tend to decrease precision
Table II Steps involved in simulating the effects of sampling intensity and undetected
koalas on the precision of density estimates
(1) Specify fixed parameters: - specify site area - specify koala numbers - koala distribution (random/clumped) - specify probability of not detecting koalas - sampling strategy (systematic vs. random)
(2) Set up koala population: - assign coordinates to each koala
(3) Count the number of koalas which fall within transects and are detected
(4) Repeat steps (2) and (3) 1000 times (5) Repeat steps (2)- (4) for a range of sampling intensities (e.g. 5%- 90%)
108

Figure I The Koala Coast Region is located 20km south-east of Brisbane. The site number is shown for each of the areas that have been surveyed for koalas.
109

Koala Density Fluctuations Two methods were used to assess differences in density estimates from repeated
surveys for the Commonwealth Land (site 11). The first method, the ratio method, can
be used to calculate the standard error associated with density estimates obtained from sampling units of unequal size (e.g. transects of unequal area) (see Caughley and
Sinclair 1994 ). This standard error can then be used to calculate confidence limits for the density estimate. The second method used simulations to determine the range of
koala densities for which the probability of obtaining the field estimate is greater than or equal to 5%. This was done by running simulations (Table II) for the
Commonwealth Land with a known field estimate. An additional step was introduced to the simulation to determine the probability of obtaining the field estimate for a
range of test koala densities above and below the field estimate. This produced a
normal distribution with the field estimate at the centre. From this type of distribution it is possible to define the range of koala densities for which the probability of
obtaining the field estimate is greater than or equal to 5%. By estimating these ranges
for the Commonwealth Land at different survey times, it can be determined whether
or not differences between field estimates are likely to be due to sampling errors.
Results Koala Surveys
A total of 24 bushland and urban remnant bushland sites were surveyed in the Koala
Coast Region. Nineteen sites were surveyed in survey one (Summer/ Autumn 1996)
and 22 sites in survey two (Winter/Spring 1996). The range of densities obtained for survey one and two are 0-1.3 koalas per hectare and 0-1.2 koalas per hectare
respectively (Table III). Although density ranges were similar between surveys, the
majority of sites had different estimated densities in surveys one and two. For
example, the Commonwealth Land (site 11) recorded densities of 0.64, and 0.19
koalas per hectare for surveys one and two respectively. Koalas were found on all sites, even those sites that recorded densities of 0 koalas per hectare (koalas were
located outsidet ransects).
Accuracy - Mark-resight In mid-late 1996, a total of 40 koalas were captured and collared at Gravel Reserve
and Burbank (20 koalas at each). Based on density estimates from the koala surveys, collared koalas accounted for approximately 60-70% of koalas present at each site.
Both sites were surveyed on three consecutive days in February 1997. Seventeen of
the 20 collared koalas were present on Gravel Reserve during the surveys. On day three, 16 collared koalas were present. At Burbank, 10 of the 20 collared koalas were
present in the search area (approximately 50ha of the total 157.2ha) on days one and
three. On day two of the survey, 9 collared koalas were present in the search area. On
average, 16% of collared koalas were not detected during the three surveys at the two sites (Table IV).
110

0.1 0.1 0.8
• ............ •• • • 0.8 • • • • • ......
• 0.1
• • 0.1
(c) Extremely Clumped
0.1 0.3 0.6
• ... • • ~· . ' • 0.6 • .. • • .... •
• •• • • ... • 0.3
• •
0.1 •
(b) Moderately Clumped
• • • • • • • • • • • ,. • • • • • • • • • • • I • • •
• • • • ..
(a) Random
Figure H Examples of the three koala distributions used in the simulations. For the clumped distributions, the probability of a koala being placed in each row or column is shown.
111
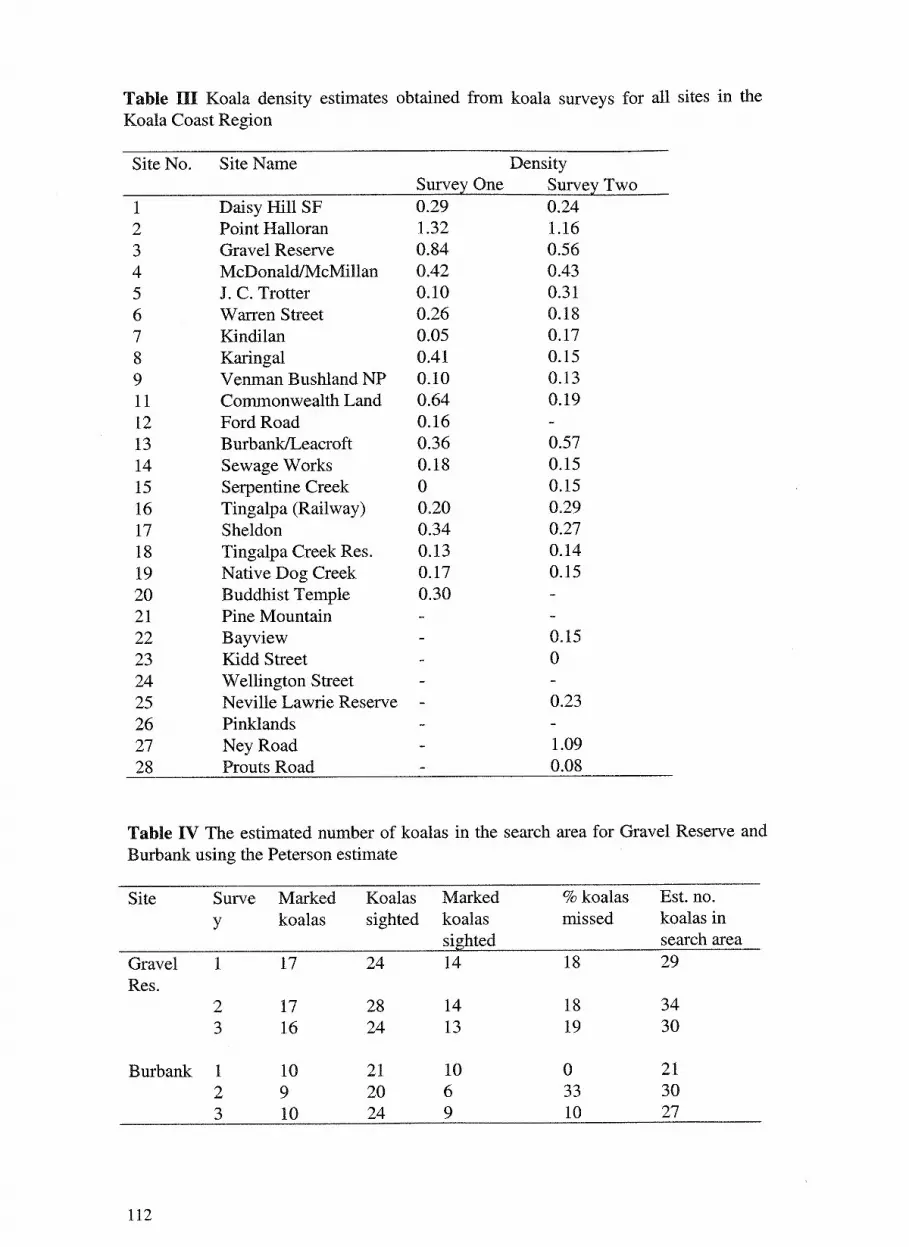
Table Ill Koala density estimates obtained from koala surveys for all sites in the Koala Coast Region
Site No. Site Name Density Survey One Survey Two
1 Daisy Hill SF 0.29 0.24 2 Point Halloran 1.32 1.16 3 Gravel Reserve 0.84 0.56 4 McDonald/McMillan 0.42 0.43 5 J. C. Trotter 0.10 0.31 6 Warren Street 0.26 0.18 7 Kindi1an 0.05 0.17 8 Karin gal 0.41 0.15 9 Venman Bushland NP 0.10 0.13 11 Commonwealth Land 0.64 0.19 12 Ford Road 0.16 13 B urbank/Leacroft 0.36 0.57 14 Sewage Works 0.18 0.15 15 Serpentine Creek 0 0.15 16 Tingalpa (Railway) 0.20 0.29 17 Sheldon 0.34 0.27 18 Tingalpa Creek Res. 0.13 0.14 19 Native Dog Creek 0.17 0.15 20 Buddhist Temple 0.30 21 Pine Mountain 22 Bayview 0.15 23 Kidd Street 0 24 Wellington Street 25 Neville Lawrie Reserve 0.23 26 Pinklands 27 NeyRoad 1.09 28 Prouts Road 0.08
Table IV The estimated number of koalas in the search area for Gravel Reserve and Burbank using the Peterson estimate
Site Surve Marked Koalas Marked %koalas Est. no. y koalas sighted koalas missed koalas in
sighted search area
Gravel 1 17 24 14 18 29 Res.
2 17 28 14 18 34 3 16 24 13 19 30
Burbank 1 10 21 10 0 21 2 9 20 6 33 30 3 10 24 9 10 27
112

0.20
0.00
0
\ \
v\ · .. \ . ··.~ . v\ .
..... ,
20
-o- No koalas missed - -v · 20% probability of missing koala --o - 50% probability of missing koala
~ ~
.............. ~ ---- ---o
- "V ••• . --------
40 60 80 100
Percentage of site surveyed
Figure III Standard deviation of koala density estimates (1000 replicates) for Daisy Hill State Forest. Simulations were run with randomly distributed koalas and systematic sampling.
113
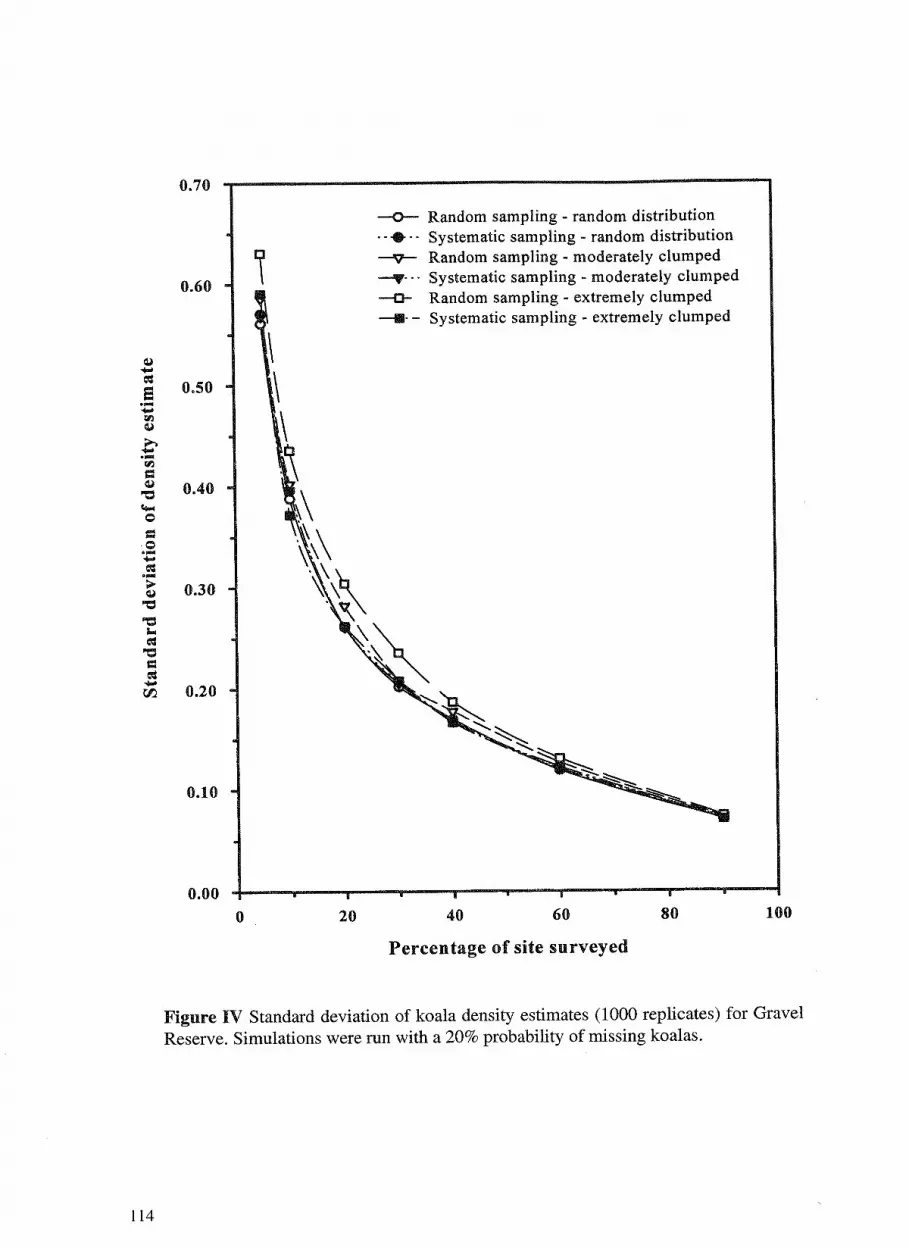
~ -~ e ..... -riJ ~
;;.-. -..... riJ
= ~ "'0 ~ Q
= Q .... -~ .... ... ~
"'0 "'0
"" ~ "'0 = ~ -00
114
0.70
0.60
0.50
0.40
0.30
0.20
0.10
0.00
0 20
-o-- Random sampling - random distribution
·- • · · Systematic sampling - random distribution
-v- Random sampling - moderately clumped
--y- · · Systematic sampling - moderately clumped
-o- Random sampling - extremely clumped
-IIIII·- Systematic sampling - extremely clumped
40 60 80
Percentage of site surveyed
100
Figure IV Standard deviation of koala density estimates (1000 replicates) for Gravel
Reserve. Simulations were run with a 20% probability of missing koalas.

Precision Approximately 16% of the koalas on each site were missed during surveys.
Simulations assumed that 20% of koalas on a site remained undetected during surveys. The effect of missing koalas on the precision of koala density estimates is illustrated for Daisy Hill State Forest (site 1) (Figure III). A 20% probability of missing koalas produced a small increase in the standard deviation of density estimates which was more or less constant across a wide range of sampling intensities. The effect of different koala distributions and sampling strategies on the precision of density estimates was minimal (Figure IV). With systematic sampling, koala distribution appeared to have no effect on the standard deviation of density estimates, even when koalas were extremely clumped. However, there was a small decrease in precision when randomly placed transects were used to sample extremely clumped koala populations. This effect was most pronounced at low sampling intensities.
Failure to detect koalas had a small but noticeable effect on the standard deviation of koala density estimates, therefore, subsequent simulations assumed a 20% undetected koala rate. Koala distribution had no significant effect on precision when sampling was systematic, therefore, all subsequent simulations assumed randomly distributed koalas.
The simulated results of the effect of sampling intensity on the precision of koala density estimates at a low, medium and high density site, suggested that higher density sites require greater sampling intensity compared to low density sites to maintain a similar level of precision (Figure V). Venman Bushland National Park (site 9- a low density site) was surveyed with a sampling intensity ranging from 17% to 25%. The simulations suggested that the sampling intensity at this site provided a precise estimate. There was little increase in precision beyond a sampling intensity of about 20%. Koala density estimates for Daisy Hill State Forest (site 1 - medium density site) used samples ranging from 11% to 18% of the site. The simulations suggested that at least 20% of the site should have been sampled. A further small increase in precision would be achieved by increasing the sample size to as high as 40% of the site. The precision of koala density estimates for Gravel Reserve (site 3-high density site) was much lower than for the previous two sites. At this site, a much higher sampling intensity would be needed to provide a comparable level of precision. During the field surveys, between 40% and 100% of Gravel Reserve had been sampled. The simulations suggested that increasing the sampling intensity above 40% would increase the precision of the koala density estimate. However, there would be little increase in precision beyond a sampling intensity of about 60%.
Koala Density Fluctuations The koala density estimates for the Commonwealth Land for surveys one and two
were 0.64 and 0.19 koalas per hectare respectively. Results of simulations and the ratio method suggested that the actual densities varied on the site between surveys and that the differences were not a result of sampling error (Figure VI). As the ranges of estimates for the two survey periods did not overlap, it would seem extremely unlikely that the actual density of koalas on the site was the same for both sampling periods.
115

1.6
----o; .c: .. ... 1.2 c. '-'
.£ "' c ...
"0 0.8 o;
-; 0
..:.:: "0 ... .... o;
0.4 e 0 ·.;:
~~--®-·f··*·····t··-···-··*•• "' IJ;l
0.0
0 20 40 60 80 100
Percentage of site surveyed
1.6
'2 .c: .. ...
1.2 c. '-'
£ "' = ...
"0 o; 0.8 ;;; 0 0 ..:.::
"0
~~-l~- -~ ----- ~--------- *--... ..... o;
0.4 e ·.;: "' IJ;l
0.0
0 20 40 60 80 100
Percentage of site surveyed
1.6 ,-.. 0 o; .c: .. ... ,e 1.2
I 0
..... . ..: 0 "' =
r -~-+-- -l-------1-...
'!;l
'-'1 0.8 -; 0
..:.:: '!;l ...
I .... "' 0.4 e .: ..,
0 IJ;l
0.0 0 20 40 60 80 100
Percentage of site surveyed
Figure V The distribution of koala density estimates (1000 replicates) for a low (top),
medium (middle) and high density site (bottom). The dashed line indicates the koala
density used for the simulations. Simulations were run with random koala distributions and
systematic sampling. The centre line of the box plots indicates the median of the 1000
density estimates. The top and bottom of the box represent the 75th and 25th percentiles. The
bars on either side of the box represent the 90th and 1Oth percentiles, and the open circles
represent the 95th and 5th percentiles.
116
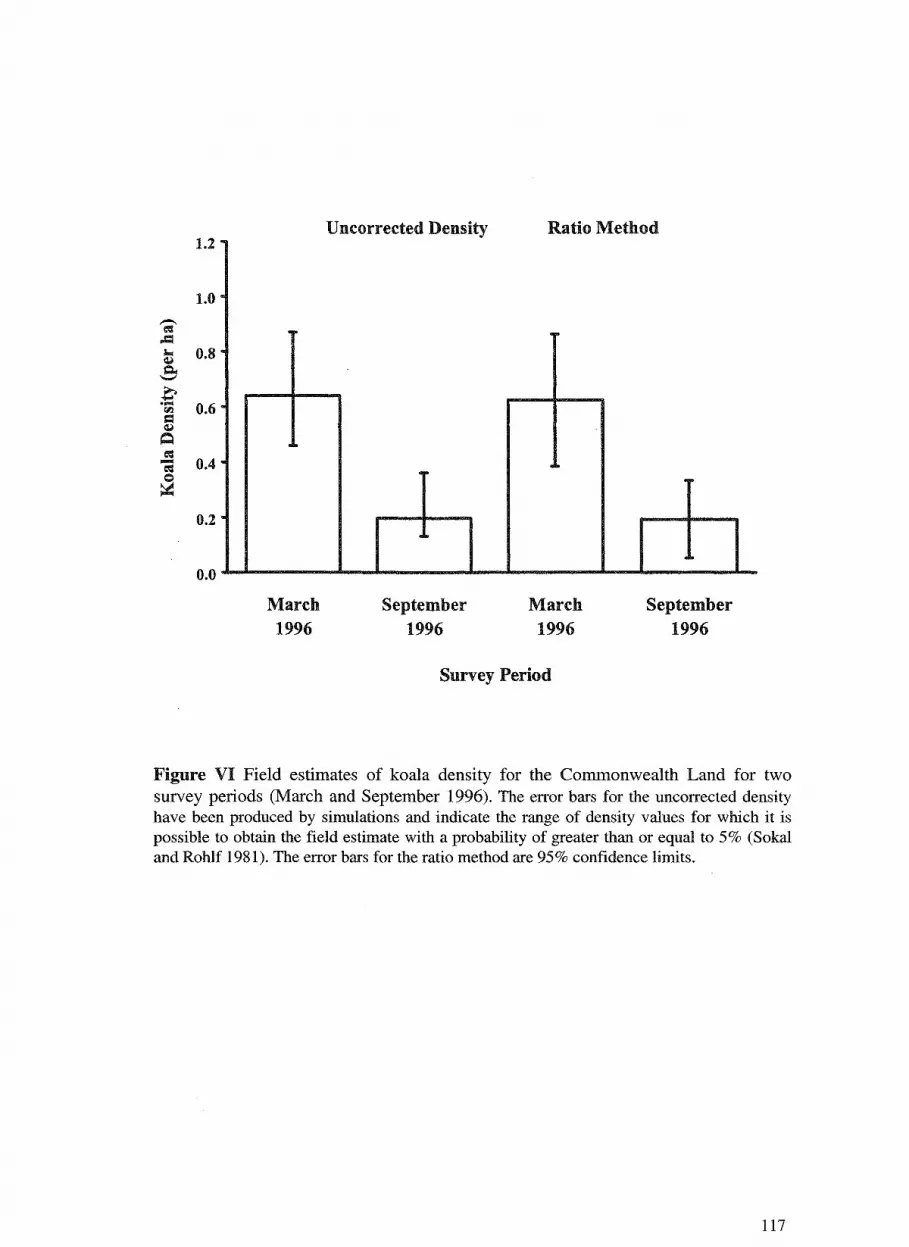
1.2
1.0
'2 ..::=
loot 0.8 ~ !:l.. "-'
.e-..... 0.6 1"-l
= ~
~ ~ - 0.4 ~
~ 0.2
0.0
.
March 1996
Uncorrected Density
September 1996
Ratio Method
March 1996
September 1996
Survey Period
Figure VI Field estimates of koala density for the Commonwealth Land for two survey periods (March and September 1996). The error bars for the uncorrected density have been produced by simulations and indicate the range of density values for which it is possible to obtain the field estimate with a probability of greater than or equal to 5% (Sokal and Rohlf 1981). The error bars for the ratio method are 95% confidence limits.
117

Discussion Koala Surveys
The koala density estimates obtained from bushland koala surveys in the Koala
Coast Region of south-east Queensland of 0-1.3 koalas per hectare are comparable to
koala densities from other areas in Queensland (Table V). However, koalas in the
Region must co-exist with humans in a heavily urbanised environment, and therefore
must be managed differently to other populations in Queensland. Urban development
in the Koala Coast Region is destined to increase as southeast Queensland is currently
experiencing the fastest rate of population growth of any region in Australia (Catterall
and Kingston 1993). The consequence of such population growth is that remaining
bushland areas will further diminish if current planning and land-use practices
continue (Catterall and Kingston 1993). However, it is difficult to recommend that
current planning and land-use practices be modified for koala conservation without
accurate and precise baseline data on koala distribution and abundance in the Region.
Table V Koala densities recorded from koala populations in Queensland
Location Central Queensland South-east Queensland Southern Queensland South-east Queensland
Accuracy
Density (koalaslha) 0.02-0.4 0.4 0.1-2 1.9-2.5
Author Melzer (1995) White and Kunst (1990) Gordon et al. (1990) Hasegawa (1995)
The mark-resight method has been used in one other study in the Koala Coast
Region for calculating the proportion of koalas missed during surveys. Hasegawa
(1995) found that 28.6% of koalas were not detected during a daytime search using
the transect method. This is slightly higher than the 16% recorded from this study. It
would be expected that the percentage of koalas not detected during surveys would
vary across different habitat types, however, Hasegawa (1995) had seven koalas
collared and conducted only one survey which suggest that our results (Table IV) may
be a little more robust. It is extremely important to obtain accurate density estimates for koala populations
in the Koala Coast Region as information on koala abundance is required for
development applications. A flora and fauna assessment prepared by Dames and
Moore (1994) for Redland Shire Council for a proposed subdivision development of a
60ha bushland site on Ney Road (site 27) found that koalas did not reside on the site.
The development was approved based on this report. Surveys conducted in survey two
(winter/spring 1996) suggest that Ney Road has one of the highest koala densities of
the 24 sites surveyed (Table ill). This highlights the importance of obtaining accurate
distribution and abundance data in the Koala Coast Region. It has been documented that the mark-resight method for population estimation has
clear advantages over many other methods of population estimation. Pollock (1981)
considers capture resight to be a design that has great potential for many species and
circumstances. In many cases, capture-resight is cheaper than capture-recapture as it
reduces the field effort after an initial intense capture and marking period, and requires
fewer and less trained personnel for resighting. Also, more data is able to be gathered
in less time and re-sightings are able to take place while achieving other research
objectives. In general, the method is less disruptive to the organisms involved and the
118

environment (Minta and Mangel 1989). This particular benefit is extremely useful in conducting surveys in areas where the koala has an extremely high public profile, as in the Koala Coast Region.
Precision Computer simulations provided a useful method for assessing the adequacy of
sampling schemes to determine koala density. They were able to assess the effects of many factors on the precision of koala density estimates, and therefore provide koala researchers and managers with an important tool when designing koala censusing programs. It is important, however, to determine the proportion of koalas that fail detection during a survey prior to running the simulations because it produces a small but noticeable effect on precision (Figure ill). Results from the mark-resight experiment in this study suggest that approximately 16% of koalas are not detected during surveys. This of course varies slightly across sites on a local scale but may vary to a greater extent across habitat types as variation is almost entirely due to differences in visibility (e.g. floristics, vegetation structure etc.) (Gordon 1998 pers. comm.).
Sampling strategy and simulated koala distribution had little effect on the precision of koala density estimates except when koalas were extremely clumped and randomly sampled during a censusing event (Figure IV). Koalas are solitary animals and are rarely encountered in pairs, even in areas of high koala density (Mitchell 1989). However, koalas are rarely randomly distributed and in general tend to be heterogeneously distributed depending on climate and vegetation structure and floristics (Lee and Martin 1988). It has been documented that in areas such as southwest Queensland during times of drought, koala distribution tends to be concentrated around vegetation associated with permanent water (Gordon et al. 1988). To minimise the effects of clumping on the precision of koala density estimates, sampling can be done systematically (Caughley and Sinclair 1994).
Another reason why simulations are an important tool to researchers and managers is that simple graphical presentation of results allows the value of increasing sampling intensity to be assessed. This study looked at the effects of sampling intensity on precision of koala density estimates at low, medium and high density sites (Figure V). The graphical representation of the results of simulations for these three densities made it easy to assess what percentage of a site should be sampled to achieve a desired level of precision. It is not logistically feasible to gain this type of understanding through repeated sampling in the field. The results of the simulations suggested that precision increases with sampling intensity and the standard deviation increases with density (Caughley and Sinclair (1994) suggest that this is almost always the case). From a management point of view, precision should be maximised so that repeated counts reflect the current status of a population.
Density Fluctuations A number of studies have suggested that seasonal movements, changes in habitat
utilisation, and winter mortality are causes of density fluctuations. White and Kunst's (1990) study in the Koala Coast Region noted seasonal trends in certain tree species utilisation owing to flowering phenology. Gordon et al. (1990), from a study in Oakey in southern Queensland, and Melzer (1995) from a study in Springsure in Central Queensland, reported environmentally induced changes in community utilisation by koalas. These studies noted that changes in fodder quality (Eucalyptus productivity,
119

leaf concentrations of water, nitrogen, potassium and phosphorus) linked with
environmental parameters such as rainfall and maximum temperature accounted for
changes in community utilisation. White and Kunst (1990), Martin (1985) and
Eberhard (1972) recorded higher mortalities of koalas in winter and early spring and
suggested that winter conditions imposed stresses on nutritional reserves. Any one or
a combination of these factors may have been responsible for the density fluctuations
at the Commonwealth Land (Figure VI).
Conclusion Providing baseline data on the distribution and abundance of organisms is for many
management programs the most important step (Caughley and Sinclair 1994). The
results from this study provide that baseline data for koalas in the Koala Coast
Region. The mark-resight study showed that to obtain accurate results from a koala
census, population underestimation needs to be considered. Computer simulations can
be used to provide information on factors affecting the precision of density estimates
that could not have been obtained by conventional sampling methods. By obtaining--
accurate and precise density estimates objective assessment of the conservation status
of areas can be made. Thompson (this volume) highlights the importance of the role of
research in identifying areas most deserving of conservation, so that effective
conservation planning and management can be put into place. The ability to obtain
accurate and precise distribution and abundance information is therefore important for
koala conservation in the Koala Coast Region.
Acknowledgments We are grateful to numerous volunteers who have assisted with the field surveys
undertaken. Thanks also to landholders in the study area for access to their properties.
We thank Greg Gordon for reviewing the manuscript.
References Bali, R., and Delaney, R. (1997). A Review of Koala Radio-collaring Research.
Consultancy report for the South East Forests Koala Research Committee.
Catterall, C. P., and Kingston, M. (1993). Remnant Bushland of South-east
Queensland in the 1990's: It's distribution, loss, ecological consequences and
future prospects. Griffith University and Brisbane City Council.
Caughley, G., and Sinclair, A. R. E. (1994). 'Wildlife Ecology and Management'.
(Blackwell Science: Cambridge.) Dames and Moore. (1994). Flora and Fauna Assessment. Proposed subdivision
development - Ney Road, Capalaba. Report prepared for Redland Shire Council.
Eberhard, I. H. (1972). Ecology of the koala, Phascolarctos cinereus (Goldfuss), on
Flinders Chase, Kangaroo Island. PhD Thesis, The University of Adelaide,
Adelaide. Elsol, J. A., and Dowling, R. M. (1978). Moreton Region Vegetation Map Series:
explanatory booklet for Beenleigh sheet scale 1:100 000. Botany Branch,
Queensland Department of Primary Industries, Brisbane.
Gordon, G., Brown, A. S., and Pulsford, T. (1988). A koala (Phascolarctos cinereus
Goldfuss) population crash during drought and heatwave conditions in south
western Queensland. Australian Journal of Ecology 13, 451-461.
120

Gordon, G., McGreevey, D. G., and Lawrie, B. C. (1990). Koala populations in Queensland: major limiting factors. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 189-192. (Surrey Beatty & Sons: Sydney.)
Hasegawa, M. (1995). Habitat utilisation by koalas (Phascolarctos cinereus) at Point Halloran, Queensland. MSc Thesis, University of Queensland.
Lee, A., and Martin, R. (1988). 'The Koala: a natural history'. (University of New South Wales Press: Sydney.)
Martin, R. W. (1985). Overbrowsing and decline of a population of the koala, Phascolarctos cinereus, in Victoria II. Population condition. Australian Wildlife Research 12, 367-375.
Melzer, A. (1995). Aspects of the ecology of the koala, Phascolarctos cinereus (Goldfuss, 1817), in the sub-humid woodlands of central Queensland. PhD Thesis, University of Queensland.
Minta, S., and Mangel, M. (1989). A simple population estimate based on simulation for capture-recapture and capture-resight data. Ecology 70(6), 1738-1751.
Mitchell, P. J., Bilney, R., and Martin, R. W. (1988). Population structure and reproductive status of koalas on Raymond Island, Victoria. Australian Wildlife Research 15,511-514.
Mitchell, P. (1989). The social organisation of koalas. PhD Thesis, Monash University, Clayton, Victoria.
Phillips, B. (1990). 'Koalas- The little Australians we'd all hate to lose'. Australian National Parks and Wildlife Service. (Australian Government Publishing Service: Canberra.)
Phillips, S., and Callaghan, J. (1996). Koala Habitat Atlas- Project No.5: Campbelltown Local Government Area- DRAFT. Australian Koala Foundation.
Pollock, K. H. (1981). Capture-recapture models: a review of current methods, assumptions, and experimental design. Studies in Avian Biology 6, 426-435.
Smith, A. P., and Andrews, S. (1997). 'Koala habitat, abundance and distribution in the Pine Creek study area'. A report to State Forests ofNSW.
Sokal, R. R., and Rohlf, F. J. (1981). 'Biometry- The principles and practice of statistics in biological research'. Second Edition. (W. H. Freeman and Company: New York.)
White, N. A., and Kunst, N.D. (1990). Aspects of the ecology ofthe koala in southeastern Queensland. In 'Biology of the Koala'. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) pp. 189-192. (Surrey Beatty & Sons: Sydney.)
121

The koala habitat atlas: preliminary results for koala tree species preferences in the city of Ballarat local government area, Victoria.
Patrick Prevett1, Rick Pope 2 , John Callaghan 3 and Lynne Bailey 4
1 Centre for Environmental Management, University of Ballarat, PO Box 663, Victoria.
2 State Data Centre, Central Square, PO Box 2117, Ballarat, Victoria.
3 Australian Koala Foundation, PO Box 9899, Brisbane, Queensland.
4 Centre for Environmental Management, University of Ballarat, PO Box 663, Victoria.
Abstract The Spot Assessment Technique for assessment of koala habitat quality is a central
tool in the preparation of Koala Habitat Atlas Maps produced by the Australian Koala Foundation. This technique was used extensively in the City of Ballarat Local Government Area as the basis for the identification of eucalypt communities and assessment of habitat quality used by koalas. Thirty nine primary plots and a total of 1898 trees have been assessed. Sample plots comprised 10 species of eucalypts and 3 species of non-eucalypts. All 39 primary plots displayed evidence of utilisation by koalas with faecal pellets recorded beneath all 10 species of eucalypts and 3 species of non-eucalypts. Less than 10% of native forests in the Ballarat Local Government Area occurred on the fertile Newer Volcanic soils and these showed high levels of usage by koalas. Eucalyptus viminalis sub species viminalis and E. obliqua were the dominant species on these soils but only E. viminalis sub species viminalis was the subject of preferential utilisation by koalas.
Introduction A Koala Habitat Atlas map is a multilayered Geographic Information System that
provides information on koala habitat quality in map format (Phillips et al. 1996). The purpose of the map is to provide planners and decision makers with a science-based and user friendly planning tool with which to make realistic and effective decisions to conserve wild koalas (Sharp and Phillips 1997). Prior to the establishment of Atlas methodology in eastern Australia there was no systematic approach to koala habitat assessment and therefore the importance of any specific block of land to koalas was largely unknown. Maps generated by the Atlas project will rectify this situation. Data sets which can be incorporated into the GIS include vegetation, geology, soils and terrain data including slope, aspect, elevation, drainage lines, roads and land tenure.
The AKF recognises the need for the Atlas to continually evolve, particularly as new technology becomes available (Sharp and Phillips 1996). It is also recognised that vegetation survey together with underlying soil data collection is extremely time consuming and costly. Therefore in Ballarat the opportunity was taken initially to assess the possibility of using remote sensed Landsat imagery to produce the vegetation map. Unfortunately Landsat imagery did not provide the level of resolution required to distinguish between the different eucalypt communities found in the region. For example, Eucalyptus obliqua open forest could not be distinguished from Eucalyptus obliqua-Eucalyptus radiata-Eucalyptus rubida open forest on remote
122

sensed images. Distinction between map units such as these was essential in order to define koala habitat preferences.
Previous work by Prevett (1991) has shown that koalas are widespread in the urban and rural fringes of Ballarat. Prevett recognised the significance of native vegetation remnants for koalas and showed that koalas were able to move between remnant patches within the fragmented forest landscape. This study has extended his work and provided evidence of the uneven distribution of koalas in the region and koala tree species preferences in relation to soil type.
Vegetation mapping and koala habitat preferences were based on the Spot Assessment Technique. A spot assessment is a sampling procedure used to provide a reliable means of determining the significance of a given area of habitat for koalas. In this study the Spot Assessment Technique was used to determine the significance of different vegetation communities for koalas (Phillips and Callaghan 1995).
Until this project was undertaken, no vegetation map with sufficient detail to represent koala habitat use was available. The principal outcomes of the project will be Koala Atlas Maps for the City of Ballarat Local Government Authority, an area of approximately 70,000 ha. The purpose of this paper is to quantify tree preferences and habitat utilisation by koalas particularly with reference to the range of soil types found in the Ballarat region. This information, when the data sets are complete, will be used to produce a vegetation map to show details of eucalypt communities present and a map that identifies koala habitat quality within the Ballarat LGA.
Methods A map, produced by classification of a Landsat TM image was overlaid by
geomorphological units for the area. Spot assessment localities were randomly generated for each vegetation unit within each geomorphological unit.
Sites were selected which sampled the full range of geomorphological and floristic variation within the study region. Care was taken to select sites from within core areas of vegetation where possible and so avoid major habitat disturbances such as major roads, urban developments or recent bush fires. Field site coordinates for the selected sites were entered into a Global Positioning System (GPS) to physically locate these sites on the ground. At the site (identified by GPS coordinates) a spot assessment for koala scats was conducted as follows. A representative area of the vegetation community to be assessed was selected and all trees within 20m of the tree chosen to be at the centre of the plot were marked so that at least 25 trees were included. If there were less than 25 trees within this initial area, the radius was extended until 25 trees were included and marked. A tree was defined as having a stem greater than 100 mm diameter at breast height (DBH). A search was conducted within a 1m radius around the base of all marked trees for approximately two person minutes or until one koala scat was found, whichever happened first. The search was disciplined with care taken to search methodically to detect any scats beneath the litter layer. Data recorded included GPS coordinates (stored for post processing where possible), tree species present, tree species where scats were found, koala sightings, DBH, canopy density, understorey species and density, ground cover species and density and land unit type (eg. ridge top, gully, mid slope), date, recorder and place name.
The results were presented in the form of proportional strike rates. A proportional strike rate (SR) is the number of trees of a species with koala pellets divided by total number of trees of the same species within the plot. These quotients were calculated for all sites where koala scats were detected, (active sites).
123

Results Eucalyptus species present and distribution in the Ballarat LGA
From the spot assessments a total of 11 species of the genus Eucalyptus were identified in native forests of the Ballarat LGA, including E. yarraensis and E. melliodora. Both of these species appear to have a limited distribution in the area and require further sampling to adequately assess use by koalas (Table I). Of the 11 species, there was evidence of use of 10 species by koalas.
It was found that about 70% of native forests of the region occur on impoverished low nutrient soils of Ordovician origin
Tree species utilised by koalas according to soil type Preferential tree usage by koalas on Ordovician, Granitic and Newer Volcanic soils
are shown in Table ll. Tree species utilised by koalas on Ordovician soils included E.
dives (SR 0.10), E. radiata (SR 0.07), E. aromaphloia (SR 0.09), E baxteri (0.13), E. obliqua (SR 0.09), E. viminalis ssp. viminalis (SR 0.43), E. ovata (SR 0.25), E. rubida (SR 0.07), and E. melliodora (SR 0.38). Of these species only E. viminalis ssp. viminalis was considered to be preferentially used by koalas. Less than 10% of the native forests occur on the fertile Newer Volcanic soils with fertile soils. These showed high usage by koalas. E. viminalis ssp. viminalis and E. obliqua were the dominant species. However only E. viminalis ssp. viminalis was the subject of preferential utilisation by koalas. The strike rates for E. obliqua varied considerably according to soil types. On Ordovician soils the strike rate was 0.09 whilst on the Newer Volcanics the strike rate was 0.46.
Discussion The findings suggest that the more fertile Newer Volcanic soils provided the higher
quality koala habitat, subject to the occurrence and relative density of the preferred tree species. Forests on the Ordovician derived soils at Creswick, Invermay and Canadian State Forests and the Garribaldi block provided less significant koala habitat and had a lower koala carrying capacity. These latter forests occupy substantial tracts in the east of the study region, forming an almost continuous corridor of native vegetation. Further work is required to determine the ways in which koalas (and other species) use the mosaic of habitat types that occur in a matrix of predominantly residential areas and farmlands.
The differential use of Eucalyptus obliqua by koalas in the region is of interest. It suggests either an increased palatability of this species on nutrient rich soils, or increased use by koalas as a result of codominance with Eucalyptus viminalis ssp. viminalis. The level of conditional independence of these species on each other requires further assessment. Further work is also required to determine whether koala tree species preferences as indicated by different species strike rates are due to the nutrient status of different soils or other soil characteristics, such as water availability influencing the palatability of leaves.
124

Table I Number of eucalypts and non-eucalypts assessed from active sites in relation to soil types.
Eucalyptus Species Ordovician Granitic Newer Volcanics
E. viminalis sub sp. 0 61 0 cygnatensis E.obliqua 623 211 50 E. rubida 61 1 0 E. aromaphloia 202 0 0 E. baxteri 23 0 0 E. dives 357 0 0 E. radiata 138 0 0 E. ovata 28 0 0 E. viminalis sub sp. 21 0 7 viminalis E. melliodora 8 0 0 Non-Eucalyptus Species Acacia melanoxylon 21 19 2 Acacia meamsii 2 6 0 Exocarpos cupressifonnis 2 1 0 Total number of trees 1486 299 59 accessed
Table II Strike rates assessed from active sites on soils derived from different rock types.
Eucalyptus Species Ordovician Granitic Newer Volcanics
E. viminalis sub sp. * 0.87 * cygnatensis E. obliqua 0.09 0.20 0.46 E. rubida 0.07 * * E. aromaphloia 0.09 * * E. baxteri 0.13 * * E. dives 0.10 * * E. radiata 0.07 * * E. ovata 0.25 * * E. viminalis sub sp. 0.43 * 0.67 viminalis E. melliodora 0.38 * * Non-Eucalyptus Species Acacia melanoxylon 0.33 0.11 1.0 Acacia meamsii 0 0 0 Exocarpos cupressifonnis 0 0 0
* no tree of this species were found in the sample
125

The next stage in the preparation of the Koala Habitat Atlas Maps for the Ballarat
Local Government Authority will be to use the spot assessment data to model koala
habitat across the entire region. To do this, the mapped vegetation communities
weighted according to the density of preferred and or secondary browse species will
be intersected with other layers of the GIS, including soil attributes and topography
(drainage, slope, elevation and aspect). Earlier in the project it was hoped that remote sensing would make the process of
mapping vegetation more efficient. However, it soon became apparent that subtle
differences in floristic composition and structure of the forests significant to koala
distribution could not readily be distinguished by remote sensing. Further work is
required before this technique can be applied to mapping koala habitat preferences in
the eucalypt communities that occur in the Ballarat region.
Conclusions Koalas are widespread within the Ballarat LGA and are present at higher densities
on the Newer Volcanic soils. Koala faecal pellets were found beneath all ten species
of eucalypt. Whether E. yarraensis is used by koalas has not yet been established.
Only E. viminalis spp viminalis is considered to be preferentially used by koalas.
The Koala Habitat Atlas project will make a significant contribution to our
understanding of nature conservation in a production environment. It will integrate
information on the distribution of soil landscapes, vegetation communities and the
nature and quality of koala habitat in the Ballarat LGA. The Atlas maps, with
associated information on soils and vegetation, when complete, will provide the City
of Ballarat LGA with a significant planning and management tool for the future. It
will also provide baseline reference material to guide continuing koala research in the
region. In addition, it is expected that the publicity associated with the project will
encourage establishment of further koala habitat mapping at other locations in
Victoria with subsequent benefits to this species and other forest fauna.
Acknowledgements The authors gratefully acknowledge the Australian Koala Foundation who provided
the initiative and funding support for the project and the methodology for the koala
assessments. I would also like to thank the City of Ballarat for major funding support,
and I am particularly grateful for the untiring efforts of the Australian Trust for
Conservation Volunteers, who toiled long and tedious hours in the field collecting scat
data.
References Phillips, S., and Callaghan, J. (1995). 'The Spot Assessment Technique for
Determining the Significance of Habitat Utilisation by Koalas'. (Draft Report.)
(Australian Koala Foundation: Brisbane, Queensland.) Phillips, S., Callaghan, J., and Thompson, V. (1996). 'The Koala Habitat Atlas Project
No 6: Port Stephens Local Government Area. Draft Report Prepared for Port
Stephens Council'. (Australian Koala Foundation: Brisbane, Queensland.)
Prevett, P.T. (1991). Movement paths of koalas in the urban-rural fringes of Ballarat,
Victoria: Implications for Management. In 'Nature Conservation 2: The Role of
Corridors'. (Ed. D. A. Saunders and R. J. Hobbs.) pp. 259-273. (Surrey Beatty
and Sons: Chipping Norton, NSW.) Sharp, A., and Phillips, S. (1997). Koalas, Science and Conservation. In 'Ecosystem
Management: The Legacy of Science'. (Ed. C. and D. Lewis.) (Halstead Press:
Sydney, NSW.
126

Aspects of the ecology of koalas at Blair Athol Coal Mine.
Bill Ellis\ Peter Hali, Frank Carrick1, Moto Hasegawa1
, Michael Nielsen3 and David Esser4
1 Koala Study Program, The University of Queensland, Brisbane. 2 Centre for Conservation Biology, The University of Queensland, Brisbane. 3 School of Land and Food Sciences, The University of Queensland, Brisbane. 4 Blair Athol Coal Mine, Clermont, Qld.
Abstract The diet and breeding ecology of koalas at Blair Athol Coal Mine were evaluated for
consistency with theoretical predictions based on previous studies. Faecal cuticle analysis revealed that koalas utilise food and roosting tree species in different proportions at this site. Dispersing koalas may be able to modify the proportion of dietary components eaten and utilise species not available in the area from where they dispersed. DNA microsatellite analysis revealed multiple paternity within sets of offspring from individual females and confirmed the role of non-resident koalas in the breeding ecology of the local group. Sex ratio and home range dynamics resembled those reported for other sites except that ranges appear larger than those of more southern populations.
Introduction Occupying perhaps the highest public profile of Australia's fauna, the koala has
historically been the subject of considerable public speculation as well as scientific investigation. As a result, dual "arms" of knowledge have developed, fuelled by public interest on the one hand, and scientific endeavour on the other. The koala is subject to media interest and is the object of several research articles in the scientific literature each year. It is a flagship species for conservation.
As a result, management of the habitat of the koala is undertaken in an environment pervaded by public dissemination of information. This places land managers under pressure to accommodate publicly perceived koala habitat requirements, without a base of data to underpin management decisions. At Blair Athol, several paradigms of koala ecology are being investigated, prior to and as part of, the implementation of a program designed to ensure the long-term security of koala habitat at the site. Key issues in the program include the identification of an accurate indirect survey method to estimate diet and investigation of the behaviour of koalas involved in the long-term breeding success at the site. Information describing the impact of disturbance on local groups of koalas will be used to modify management procedures.
Several recent reports (Tun 1993; Hasegawa 1995) suggest indirect methods of diet estimation (Hindell et al. 1985; Achurch 1989) to be inaccurate predictors of koala diets. Nonetheless, these methods have been widely employed (Eberhard 1978; Hindell 1979; Robbins & Russell 1978; Lithgow 1980; Martin 1985; Hindell et al. 1985; Hindell and Lee 1987; Phillips and Callaghan 1995; Ellis et al. 1995). These methods may be applicable in some areas, but not others. There is limited scientific information available to land managers regarding the effect of disturbance on koala ecology (Melzer and Lamb 1992; Melzer and Lamb 1996), though there are suggestions in the popular press (Sharp
127

1995; Beeh 1997). Added to this, some reports suggest that social dynamics influence
population viability for koalas (Sharp 1995; Beeh 1997). Koalas in Central Queensland occur in areas under threat of habitat disturbance for
agricultural purposes. However, few signs of disease have been reported (Ellis et al.
1993), and koalas occur across a broad area, providing an optimistic prognosis for the
long-term survival of this group. Nevertheless, management strategies still require
accurate information about the habitat requirements of koalas in such an environment.
This report describes the results of initial investigations designed to develop and test
methods for interpreting habitat use and the breeding dynamics of koalas at Blair Athol
Coal Mine, using radio relocation, faecal cuticle examination and hypervariable
microsatellite DNA analysis.
Method Site description
The Blair Athol study site comprises some 6000 hectares including the Blair Athol Coal
Lease (3000 ha) and surrounding woodland, 25 kilometres north west of Clermont
(22.49S 147.39E) within the brigalow belt of Central Queensland. Open-cut coal mining
occurs in the north west comer of the study site. The adjacent areas consist of Eucalyptus
woodland with grazing and crop production occurring on the north east boundary of the
site. The woodlands of the study site are dominated by poplar box (Eucalyptus populnea)
and narrow leaved iron bark (E. crebra), with an understorey dominated by black wattle
(Acacia leiocalyx) and ironwood (A. excelsa). The most prevalent shrub species in the
area are currant bush (Carissa ovata) and hop bush (Dodonea viscosa). The two most
common types of plant community within the study site are rosewood (Acacia
rhodoxylon) I narrow-leaved ironbark and poplar box I silver leaved ironbark (E.
melanophloia) associations. Patches of forest red gum (E. tereticornis) are present in both
associations. The climate of the Clermont region is sub-monsoonal with a long-term average annual
precipitation rate of 671 mm. The mean yearly maximum and minimum temperatures are
30°C and 15°C.
Location and capture of koalas Koalas were located by systematic surveys of all areas within the site, and captured
with the assistance of a high-lift bucket-truck (Simon, Australia), by conventional
methods (Lee and Martin 1990) or with the koala trap developed by Hasegawa (1995).
Koalas were fitted with collar-mounted radio transmitters (Titley Electronics, Australia)
which enabled their movements to be monitored. Each transmitter emitted an individual
frequency between 150 and 152 MHz. A Regal 2000 receiver and 3 element Y agi antenna
(Titley Electronics, Australia) were used to radio-track koalas. Dependent young (aged less than one year) were not collared since they generally
occupied the same tree as their mother and were observed when their mother was located.
Sub-adult koalas (aged between one and two years) were fitted with modified collars
containing an elastic section to prevent discomfort during growth.
Tree use and home range analysis Trees in which koalas were sighted during weekly radio tracking procedures were
marked using individually numbered stainless steel tags. The position of the tree was
recorded using a hand held Global Positioning System unit (Trimble Navigation,
Australia); the tree height, girth and species were also recorded.
128

Harmonic mean horne range size was calculated using the Ranges V Analysis System (Kenward and Hodder, Institute of Terrestrial Ecology, Dorset, U.K.). Statistical analyses were performed using SAS statistical software (SAS Institute, Cary, N.C., U.S.A.).
Diet analysis Diet analysis was carried out using faecal pellets according to the method of Tun (1993;
for review see Ellis et al. 1999). This method has been used in other areas within Queensland (Tun 1993; Hasegawa 1995) to determine local diet preferences for koalas. A local reference collection for leaf cuticle identification was prepared and analysis of faecal cuticle remains was used to determine the species eaten by koalas.
Parentage Analysis- DNA extraction Tissue was taken from non-anaethsetised koalas with a sterile leather punch that
removed a 3mm-diarneter section of the outermost portion of one ear. The sample was kept refrigerated in 20% dimethyl sulphoxide in saturated NaCl or stored in ethanol until processed.
DNA was extracted from tissue samples that had been ground in liquid nitrogen or from huffy coat using standard protocols (Sarnbrook et al. 1989). After digestion with Proteinase K, contaminating proteins were removed by a phenol/chloroform extraction procedure and the DNA precipitated in salt and ethanol, washed twice in 70% ethanol, resuspended in buffer containing ltnM EDTA and stored frozen.
Genetic analyses Genotypes at six dinucleotide rnicrosatellite loci used previously for koala population
studies (Houlden et al. 1996) were obtained using standard procedures. Six pairs of oligonucleotide primers were used to amplify loci by PCR (20 s denaturing at 94°C, 30 s annealing at 50°C, 40 s extension at 72°C) with one of the primers in the reaction labelled with gamma 33P. PCR products were electrophoresed on 6% acrylarnide denaturing gels
at 45 volts/em for 1.5- 3 h. Gels were dried under vacuum at 80°C and exposed to x-ray film for 2 - 6 days. Allele sizes were scored with reference to a sequence ladder of known size.
Mother-offspring status was established from behavioural observations. The exchange of pouch or back-young or the fostering ofback young has not been confirmed in reports of behavioural studies of free ranging koalas. Males were excluded as potential sires if a match with the non-maternal allele was not provided at any of the six loci analysed.
Results Tree use and diet
Koalas at Blair Athol utilise several distinct Eucalyptus communities across the site. These were 1) E. populnea open forest, found in much of the central on-site area and including stands of C. intermedia, E. cambageana and creek systems supporting Melaleuca bract eat a; 2) E. crebra open forest with association between E. crebra, Acacia shirlei and A. rhodoxylon ; 3) E. melanophloia open forest in association with mixed brigalow (A. harpophylla) open scrub; 4) E. cambageana open forest, in association with C. tessellaris and C. dallachiana and 5) mixed Eucalyptus forest, which was comprised of several species including, C. dallachiana E. tereticornis, C. citriodora and E. melanophloia in a matrix of woodland.
Although trees occupied during the day accurately indicated species heavily represented in the diet, several species which koalas were not observed to use for day-time roosting were also (Table 1).
129
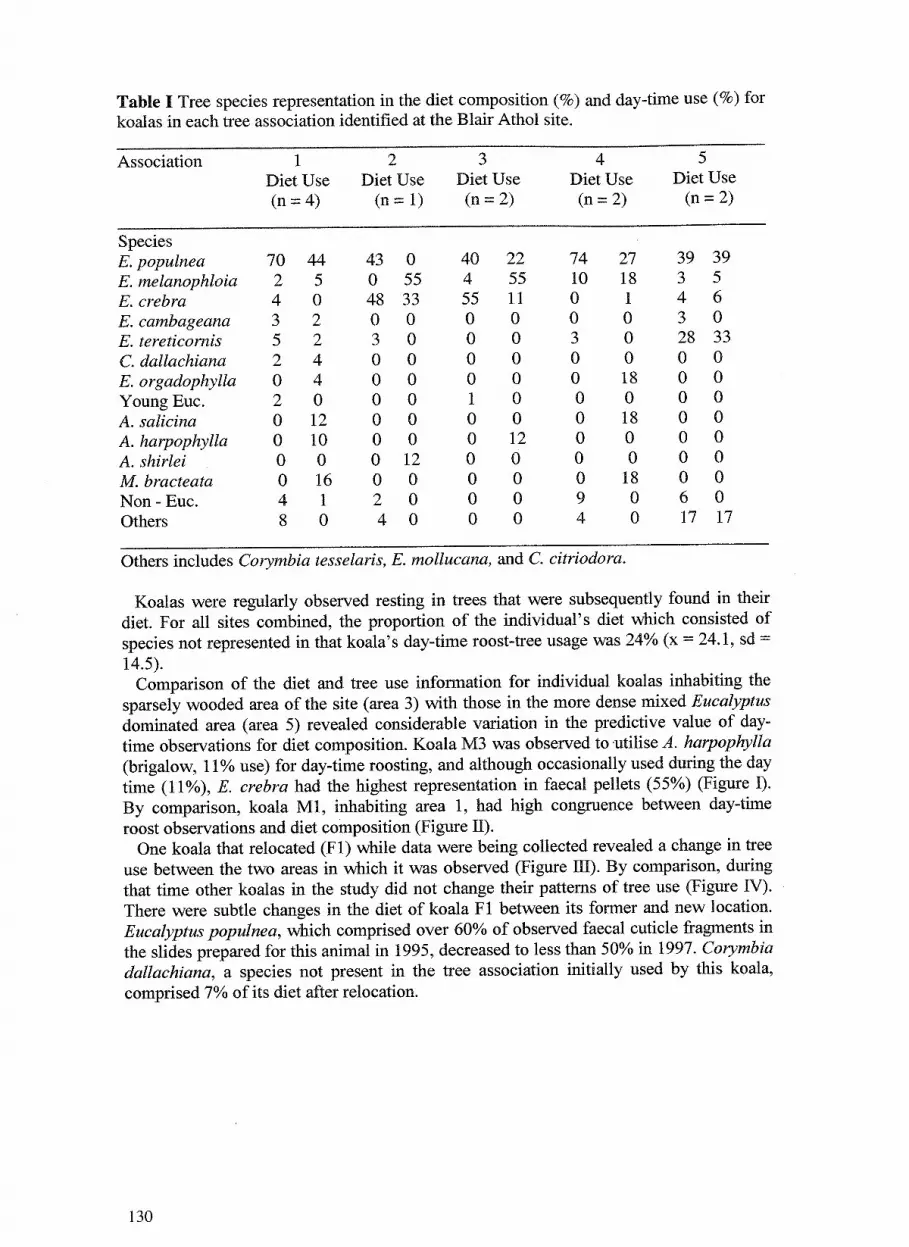
Table I Tree species representation in the diet composition (%) and day-time use (%) for koalas in each tree association identified at the Blair Athol site.
Association 1 2 3 4 5 Diet Use Diet Use Diet Use Diet Use Diet Use (n= 4) (n = 1) (n = 2) (n= 2) (n= 2)
Species E. populnea 70 44 43 0 40 22 74 27 39 39 E. melanophloia 2 5 0 55 4 55 10 18 3 5 E. crebra 4 0 48 33 55 11 0 1 4 6 E. cambageana 3 2 0 0 0 0 0 0 3 0 E. tereticornis 5 2 3 0 0 0 3 0 28 33 C. dallachiana 2 4 0 0 0 0 0 0 0 0 E. orgadophylla 0 4 0 0 0 0 0 18 0 0 YoungEuc. 2 0 0 0 1 0 0 0 0 0 A. salicina 0 12 0 0 0 0 0 18 0 0 A. harpophylla 0 10 0 0 0 12 0 0 0 0 A. shirlei 0 0 0 12 0 0 0 0 0 0 M. bracteata 0 16 0 0 0 0 0 18 0 0 Non- Euc. 4 1 2 0 0 0 9 0 6 0 Others 8 0 4 0 0 0 4 0 17 17
Others includes Corymbia tesselaris, E. mollucana, and C. citriodora.
Koalas were regularly observed resting in trees that were subsequently found in their
diet. For all sites combined, the proportion of the individual's diet which consisted of
species not represented in that koala's day-time roost-tree usage was 24% (x = 24.1, sd =
14.5). Comparison of the diet and tree use information for individual koalas inhabiting the
sparsely wooded area of the site (area 3) with those in the more dense mixed Eucalyptus
dominated area (area 5) revealed considerable variation in the predictive value of day
time observations for diet composition. Koala M3 was observed to ·utilise A. harpophylla
(brigalow, 11% use) for day-time roosting, and although occasionally used during the day
time (11 %), E. crebra had the highest representation in faecal pellets (55%) (Figure I).
By comparison, koala M1, inhabiting area 1, had high congruence between day-time
roost observations and diet composition (Figure II). One koala that relocated (F 1) while data were being collected revealed a change in tree
use between the two areas in which it was observed (Figure III). By comparison, during
that time other koalas in the study did not change their patterns of tree use (Figure IV).
There were subtle changes in the diet of koala F1 between its former and new location.
Eucalyptus populnea, which comprised over 60% of observed faecal cuticle fragments in
the slides prepared for this animal in 1995, decreased to less than 50% in 1997. Corymbia
dallachiana, a species not present in the tree association initially used by this koala,
comprised 7% of its diet after relocation.
130

A. harpophylla
E. crebra
E.popu/nea
E. me/anophloia
Other E uca !yptus
1!!1% Use 0 10 20 30 40 50 60 70
1!!1% Diet
Figure I Tree use and diet composition for koala M3 at Blair Athol in 1995
E. orgadophylla
E. populnea
E. tereticomis p E. papuana ~ (C.dallachiana) ~~%Use I
C%Diet E. melanophloia
~
E. cambageana p Non-Eucalyptus p
Other Eucalyptus p 0 10 20 30 40 50 60 70 80
Figure ll Tree use and diet composition for koala M1 at Blair Athol in 1995
Population structure and dynamics Between November 1993 and April 1997, 52 individual koalas utilising the site were
captured: 9 adult males, 12 adult females, 3 sub-adult males (less than two years but not accompanied by an adult), 5 sub-adult females and 23 pouch or back young. Six pouch young were deemed too small at capture for tissue to be sampled and data for these individuals are not included in the current analysis. Daily radio tracking was carried out for two-week periods at two-month intervals during the year, in between which koalas were radio tracked on a weekly basis.
Koalas were considered to be residents for year 1 of the study if they were re-captured on-site in year 2. Thereafter, koalas were considered to be residents once there was
131
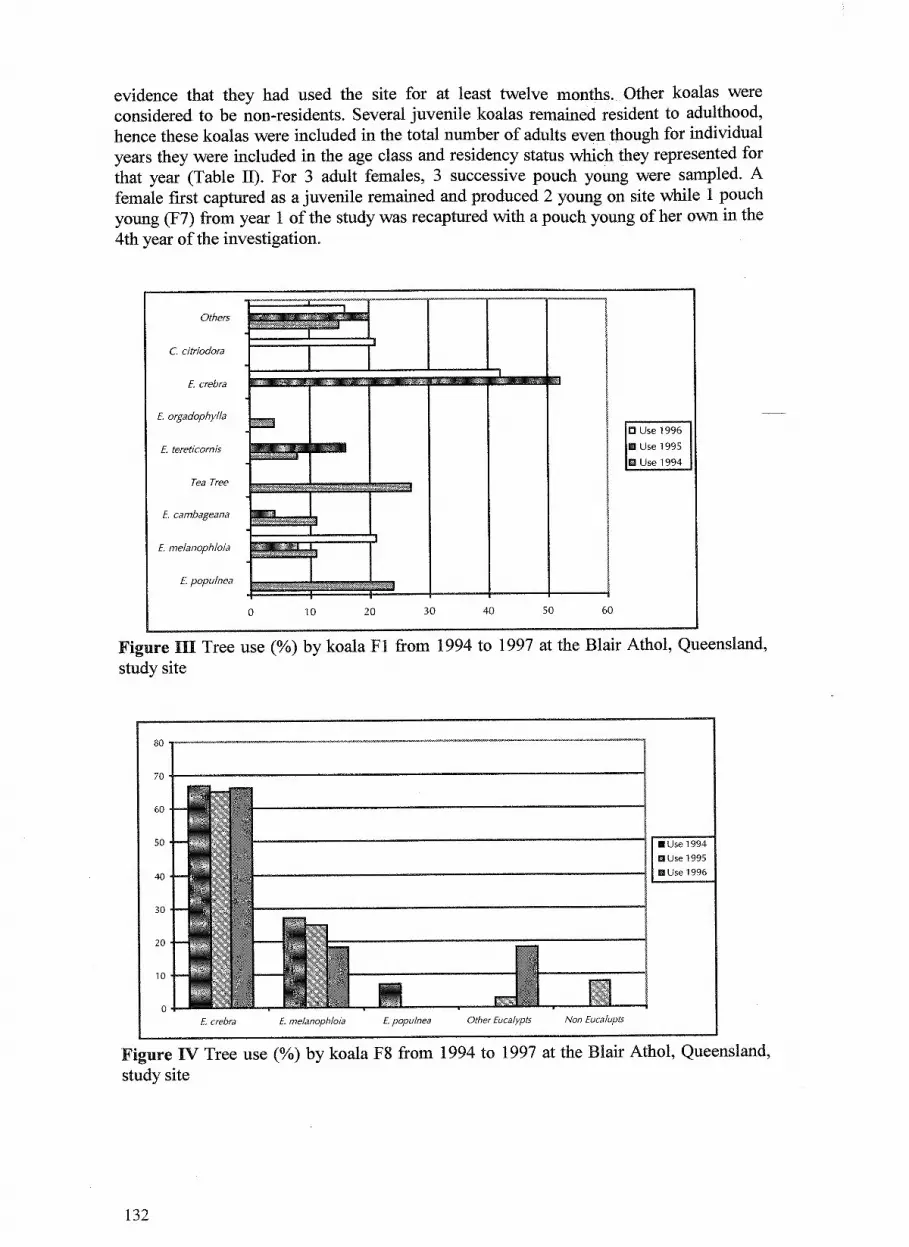
evidence that they had used the site for at least twelve months. Other koalas were considered to be non-residents. Several juvenile koalas remained resident to adulthood, hence these koalas were included in the total number of adults even though for individual years they were included in the age class and residency status which they represented for that year (Table II). For 3 adult females, 3 successive pouch young were sampled. A female first captured as a juvenile remained and produced 2 young on site while 1 pouch young (F7) from year 1 of the study was recaptured with a pouch young of her own in the 4th year of the investigation.
Others
C. citriodora
E. crebra
E. orgadophy//a
E. tereticornis
Tea Tree
E. cambageana
E. me/anophloia
E. populnea
0 10 20 30 40 50 60
IJ Use 1996
111Use1995
Ill Use 1994
Figure ill Tree use(%) by koala Fl from 1994 to 1997 at the Blair Athol, Queensland, study site
80
70
60
50
40
30
20
10
0 E. crebra E. melanophloia E.populnea Other Eucalypts Non Eucalupts
Ill Use 1994
Ill Use 1995
1111 Use 1996
Figure IV Tree use (%) by koala F8 from 1994 to 1997 at the Blair Athol, Queensland, study site
132
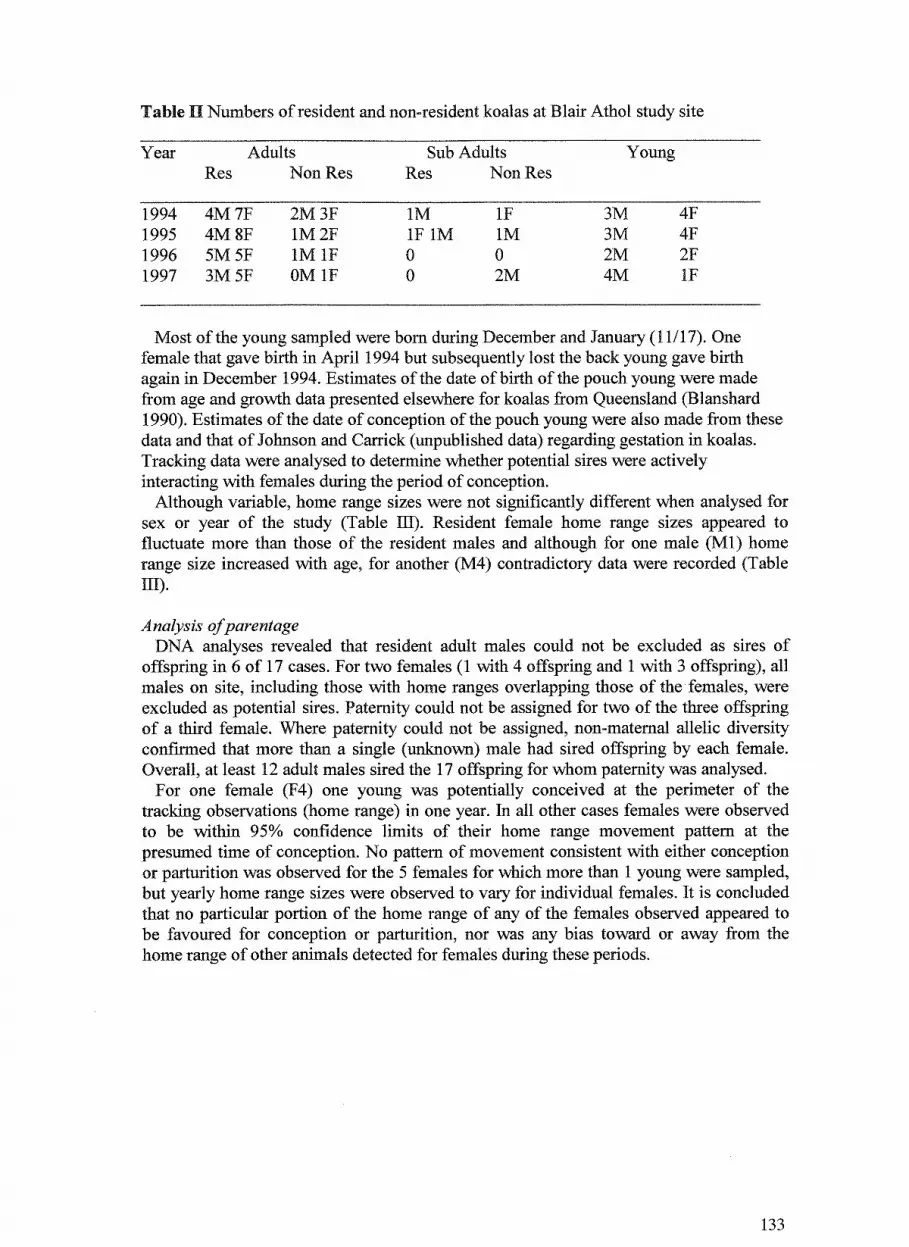
Table ll Numbers of resident and non-resident koalas at Blair Athol study site
Year Adults Sub Adults Young Res Non Res Res Non Res
1994 4M7F 2M3F 1M 1F 3M 4F 1995 4M8F 1M2F IF 1M 1M 3M 4F 1996 5M5F 1M 1F 0 0 2M 2F 1997 3M5F OM1F 0 2M 4M 1F
Most of the young sampled were born during December and January (11117). One female that gave birth in April 1994 but subsequently lost the back young gave birth again in December 1994. Estimates of the date of birth of the pouch young were made from age and growth data presented elsewhere for koalas from Queensland (Blanshard 1990). Estimates of the date of conception of the pouch young were also made from these data and that of Johnson and Carrick (unpublished data) regarding gestation in koalas. Tracking data were analysed to determine whether potential sires were actively interacting with females during the period of conception.
Although variable, home range sizes were not significantly different when analysed for sex or year of the study (Table ill). Resident female home range sizes appeared to fluctuate more than those of the resident males and although for one male (M1) home range size increased with age, for another (M4) contradictory data were recorded (Table III).
Analysis of parentage DNA analyses revealed that resident adult males could not be excluded as sires of
offspring in 6 of 17 cases. For two females (1 with 4 offspring and 1 with 3 offspring), all males on site, including those with home ranges overlapping those of the· females, were excluded as potential sires. Paternity could not be assigned for two of the three offspring of a third female. Where paternity could not be assigned, non-maternal allelic diversity confirmed that more than a single (unknown) male had sired offspring by each female. Overall, at least 12 adult males sired the 17 offspring for whom paternity was analysed.
For one female (F4) one young was potentially conceived at the perimeter of the tracking observations (home range) in one year. In all other cases females were observed to be within 95% confidence limits of their home range movement pattern at the presumed time of conception. No pattern of movement consistent with either conception or parturition was observed for the 5 females for which more than 1 young were sampled, but yearly home range sizes were observed to vary for individual females. It is concluded that no particular portion of the home range of any of the females observed appeared to be favoured for conception or parturition, nor was any bias toward or away from the home range of other animals detected for females during these periods.
133
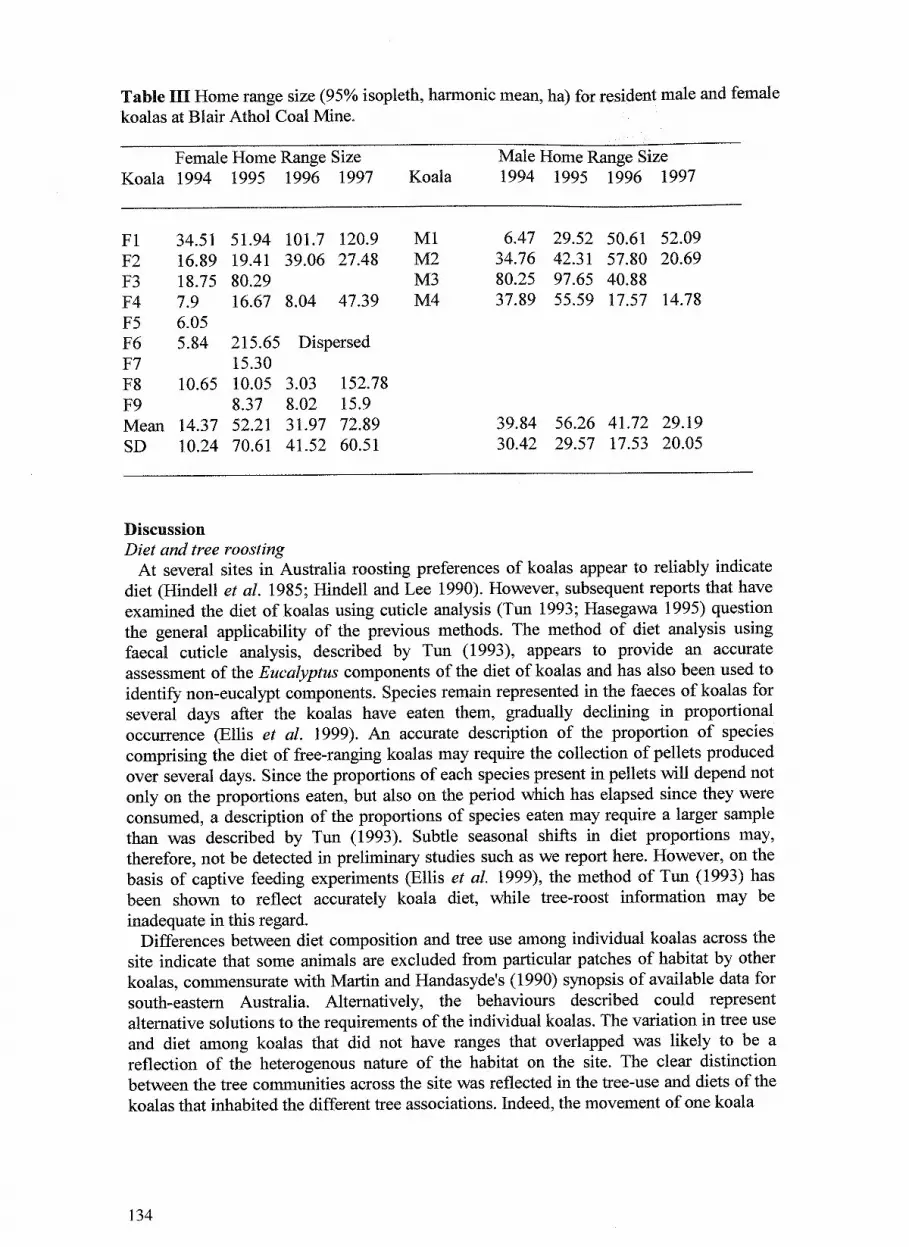
Table lll Home range size (95% isopleth, harmonic mean, ha) for resident male and female koalas at Blair Athol Coal Mine.
Female Home Range Size Male Home Range Size Koala 1994 1995 1996 1997 Koala 1994 1995 1996 1997
F1 34.51 51.94 101.7 120.9 M1 6.47 29.52 50.61 52.09 F2 16.89 19.41 39.06 27.48 M2 34.76 42.31 57.80 20.69 F3 18.75 80.29 M3 80.25 97.65 40.88 F4 7.9 16.67 8.04 47.39 M4 37.89 55.59 17.57 14.78 F5 6.05 F6 5.84 215.65 Dispersed F7 15.30 F8 10.65 10.05 3.03 152.78 F9 8.37 8.02 15.9 Mean 14.37 52.21 31.97 72.89 39.84 56.26 41.72 29.19 SD 10.24 70.61 41.52 60.51 30.42 29.57 17.53 20.05
Discussion Diet and tree roosting
At several sites in Australia roosting preferences of koalas appear to reliably indicate diet (Hindell et al. 1985; Hindell and Lee 1990). However, subsequent reports that have examined the diet of koalas using cuticle analysis (Tun 1993; Hasegawa 1995) question the general applicability of the previous methods. The method of diet analysis using faecal cuticle analysis, described by Tun (1993), appears to provide an accurate assessment of the Eucalyptus components of the diet of koalas and has also been used to identify non-eucalypt components. Species remain represented in the faeces of koalas for several days after the koalas have eaten them, gradually declining in proportional occurrence (Ellis et al. 1999). An accurate description of the proportion of species comprising the diet of free-ranging koalas may require the collection of pellets produced over several days. Since the proportions of each species present in pellets will depend not only on the proportions eaten, but also on the period which has elapsed since they were consumed, a description of the proportions of species eaten may require a larger sample than was described by Tun (1993). Subtle seasonal shifts in diet proportions may, therefore, not be detected in preliminary studies such as we report here. However, on the basis of captive feeding experiments (Ellis et al. 1999), the method of Tun (1993) has been shown to reflect accurately koala diet, while tree-roost information may be inadequate in this regard.
Differences between diet composition and tree use among individual koalas across the site indicate that some animals are excluded from particular patches of habitat by other koalas, commensurate with Martin and Handasyde's (1990) synopsis of available data for south-eastern Australia. Alternatively, the behaviours described could represent alternative solutions to the requirements of the individual koalas. The variation in tree use and diet among koalas that did not have ranges that overlapped was likely to be a reflection of the heterogenous nature of the habitat on the site. The clear distinction between the tree communities across the site was reflected in the tree-use and diets of the koalas that inhabited the different tree associations. Indeed, the movement of one koala
134

between two associations appeared to be accompanied by a change in dietary components.
The variation between the species composition of the diet of some koalas, and the composition of their roost tree selections indicates that factors other than food choice can influence the latter. In some cases, koalas were eating species that were not used for day time roosting, indicating positive tree-selection by these koalas. This observation is consistent with that of Ellis et al. (1995), who found that tree species selection by koalas varied between winter and summer. This may have been reflected in the diet of those koalas because in summer, trees with higher leaf moisture were more often selected for day time roosting. However, no diet analysis was performed during that study. The tree associations in the different areas of the current study site varied considerably, and this was reflected in the tree use and diet composition data compared between each site (Table I). Hence, even within the comparatively localised area of the current study, variations of habitat type (with respect to tree species) may result in quite different patterns of tree use and diet selection for the resident koalas. Some species with high occurrence in the diet of koalas in one area of the study were used and/or eaten less in other areas of the study.
Koalas at Blair Athol may have relatively larger home ranges than their southern counterparts (Table III) and our preliminary data bear little resemblance to those of Eberhard (1978) who observed some koalas to be in the same tree on over 60% of observations. We cannot exclude the possibility that the ecology of koalas at our site is significantly different to that of some previous reports. The study site of Mitchell ( 1989) (Redbill Ck, French Island) was some 9 ha in size, with 3.9 to 8.9 resident koalas per ha, compared to the over 6000 ha of the site of our study. The population studied by Eberhard (1978) was also an introduced island group (Kangaroo Island); both areas are the subjects of site-specific management issues at present. Hence, management decisions based on an understanding of koala ecology developed in those forest environments may have a limited application to woodlands areas such as this study.
Genetic analysis of the Blair Athol group of koalas Assertions as to the role of dominant males in the breeding ecology of koala (Sharp
1995; Beeh 1997) are suggested by Mitchell (1990) to be counter-intuitive, since they imply that fathers and daughters will mate. The description of koala mating systems has generally recognised the lack of genetic data. A loose dominance-based hierarchical breeding system has been proposed (Mitchell 1990) that is supported by several studies of free ranging koala populations (for review see Martin and Handasyde 1990). Mitchell (1989) found that male koalas living in an area develop dominance hierarchies, with the dominant males moving most frequently and widely, initiating and dominating encounters and having the highest frequencies of encounters with conspecifics. Mitchell (1990) concluded that if dominance status amongst male koalas is related to mating success, then ranging activity could be associated with that success. Mitchell's report (Mitchell 1990) presents a reasonable synopsis of field observations of koalas in southern Australia, some of which are consistent with the observations made so far in this study. An aim of the current project was, therefore, to detennine whether particular animals and areas (or individual trees) could be key factors in reproduction at the site.
Our data reveal that the majority of offspring (11/17) born at the site were sired by transient or unknown males. Unknown males were unlikely to be residents, or they would have been located due to the frequent nature of the surveys and intensive tracking procedures. It is possible that they resided outside the logistical boundaries of the study, with home ranges overlapping those of some of the on-site females. However, for this to
135

be the case in a dominance-based breeding system, one would expect single males to sire
numerous offspring, even if the males were not identified in tracking data. Their DNA
"signature" would be apparent in the genotypes of offspring. In fact, the data identify that
several non-resident males, moving undetected through the site, sired many of the
offspring. Our data suggest the social arrangement does not influence mating success in the
expected manner, in which an "alpha" male dominates. Males do not appear to
monopolise access to females, as evidenced by the paternity analysis. Mitchell (1990)
observes that alpha males can only interfere with breeding by subordinates if they are in
the vicinity of such events. It is difficult to envisage this as a breeding strategy in
woodland ecosystems such as Blair Athol, due to the spatial scale of home ranges.
Although the results of the present study indicate that mating success does not appear to
be related to the maintenance of discrete home ranges by males at the site, the social
structure resembles that proposed by Mitchell (1990). Therefore, there is no reason to
presume that untoward external factors influenced our results. Our preliminary
conclusion is that the spatial arrangement of free-ranging koalas "resident" in a
prescribed area does not necessarily reveal information about the breeding ecology of this
animal. Our data do not support the model of a dominance-based social structure
describing the breeding ecology of koalas. A role for the "travelling salesman" mating
strategy in breeding is supported by the preliminary data presented here. The role of both
resident and transient males in the breeding ecology of koalas at the site appears
significant. The model may not apply to other sites and further studies in other
representative koala habitat will help to clarify this.
Acknowledgments Blair Athol Coal Mine funded this research. Bronwyn Houlden assisted with provision
of DNA microsatellite primers. James Sullivan, Andrew Thompson and Andrew Pearce
assisted with field logistics. Nyo Tun, Andrew Veary, Petra Lundgren and Sonya
Cudmore assisted with fieldwork. Michael Cunningham assisted with laboratory
techniques. Edgar Dumpit, Tim McDonald, Ron Elms and Mark Salmon assisted with
koala capture. Bill and Glen Harrison are gratefully acknowledged for permitting access
to private property.
References Achurch, H. M. (1989). Tree utilisation by koala (Phascolarctos cinereus) populations
with regard to their geographic distribution within close proximity to the City of
Brisbane. Honours Thesis. The University of Queensland. Beeh, P. (1997). How koalas really live. GEO Australasia 19, 72-82. Blanshard, W. H. (1990). Growth and development ofthe koala from birth to weaning. In
'Biology ofthe koala'. (Eds A. K. Lee, K. A. Handasyde, and G.D. Sanson). pp. 193-202. (Surrey Beatty and Sons: Sydney.)
Eberhard, I. H. (1972). Ecology ofthe koala Phascolarctos cinereus (Goldfuss) on
Flinders Chase, Kangaroo Island. Ph.D Thesis, The University of Adelaide, Adelaide.
Eberhard, I. H. (1978). Ecology of the koala, Phascolarctos cinereus (Goldfuss) Marsupialia: Phascolarctidae, in Australia. In 'The Ecology of Arboreal Folivores'.
(Ed. G. G. Montgomery). pp 315-327. (Smithsonian Institution Press: Washington, D.C., U.S.A.)
136

Ellis, W., Carrick, F. N., Lundgren, P., Veary, A., and Cohen, B. (1999). The use of faecal cuticle examination to determine the dietary composition of koalas. Australian Zoologist 31, 127-133.
Ellis, W. A. H., Girjes, A. A., Carrick, F. N., and Melzer, A. (1993). Chlamydia! infection in koalas under relatively little alienation pressure. Australian Veterinary Journal 70, 427-428.
Ellis, W. A. H., Melzer, A., Green, B., Newgrain, K., Hindell, M. A., and Carrick, F. N. (1995). Seasonal variation in water flux, field metabolic rate and food consumption of free-ranging koalas (Phascolarctos cinereus). Australian Journal of Zoology 43, 59-68.
Hasegawa, M. (1995). Habitat utilisation by koalas (Phascolarctos cinereus) at Point Halloran, Queensland. M. Sc. Thesis. The University of Queensland.
Hindell, M.A. (1979). Diet selectivity in the koala. Honours Thesis, Monash University, Victoria.
Hindell, M.A., Handasyde K. A., and Lee A. K. (1985). Tree selection by freeranging koala populations in Victoria. Australian Wildlife Research 12, 137-144.
Hindell, M.A., and Lee, A. K. (1987). Habitat use and tree preferences of koalas in a mixed eucalypt forest. Australian Wildlife Research 14, 349-360.
Hindell, M.A., and Lee, A. K (1990). Tree preferences of the koala. In 'Biology of the koala'. (Eds A. K. Lee, K. A. Handasyde, and G. D. Sanson.) pp. 117-121. (Surrey Beatty and Sons: Sydney.)
Houlden, B. A., England, P., and Sherwin, W. B. (1996). Paternity exclusion in koalas using hypervariable microsatellites. Journal of Heredity 87, 149-152.
Lee, A., and Martin, R. (1990). Life in the slow lane. Australian Natural History 8, (90) 34-42.
Lithgow, K. A. (1980). Aspects of the feeding ecology ofthe koala Phascolarctos cinereus, in Victoria. Honours Thesis. Monash University, Victoria.
Martin, R. W. (1985) Overbrowsing, and decline of a population of the koala, Phascolarctos cinereus, in Victoria. I. Food preference and food tree defoliation. Autralian Wildlife Research 12, 355-365.
Martin, R. W., and Handasyde, K. A. (1990). Population dynamics of the koala (Phascolarctos cinereus) in southeastern Australia. In 'Biology of the koala'. (Eds. A. K. Lee, K. A. Handasyde, and G. D. Sanson.) pp 75-83. (Surrey Beatty and Sons: Sydney.)
Melzer, A. and Lamb, D. (1992). Koala habitat in Central Queensland: Implications for management. In 'Fitzroy Catchment Symposium Proceedings'. (Eds. L. J. Duivenvoorden, D. F. Yule, L. E. Fairweather, and A. G. Lawrie.) (Central Queensland University: Rockhampton.)
Melzer, A., and Lamb, D. (1996). Habitat utilisation by a Central Queensland koala colony. In 'Koalas- research for management'. Proceedings of the Brisbane Koala Symposium, 22-23 September 1990. (Ed. G. Gordon.) pp 17-22. (World Koala Research Incorporated: Brisbane.)
Mitchell, P. J. (1989). The social organisation ofkoalas. Ph.D. Thesis, Monash University, Clayton, Victoria.
Mitchell, P. (1990). The home ranges and social activity of koalas- a quantitative analysis. In 'Biology of the koala'. (Eds A. K. Lee, K. A. Handasyde, and G. D. Sanson.) pp. 171-187 (Surrey Beatty and Sons: Sydney.)
Phillips, S., and Callaghan, J. (1995). The spot assessment technique for determining the significance of habitat utilisation by koalas. Australian Koala Foundation.
137

Robbins, M., and Russell, E. (1978). Observation on movements and feeding activity of the koala in a semi-natural situation. In 'The koala: Proceedings of the Taronga symposium on koala biology, management and medicine'. (Ed. T. G. Bergin.) pp 29-41. (Zoological Parks Board ofN.S.W.: Sydney.)
Sambrook, J., Fritsch, E., and Maniatis, T. (1989). 'Molecular Cloning: A Laboratory
Manual'. 2nd edition. (Cold Spring Harbour Laboratory: Cold Spring Harbour.)
Sharp, A. (1995). The koala book. (Bateman: Auckland, New Zealand.) Tun, N. (1993). Re-establishment of rehabilitated koalas in the wild and their use of
habitat in Sheldon, Redland Shire, South East Queensland with particular reference to dietary selection. M. Sc. Thesis. The University of Queensland.
138

Valuation and Management Options for Koala Habitat in the Desert Uplands Bioregion of Queensland
John Rolfe Central Queensland University, P.O. Box 197, Emerald.
Abstract Conservation of koala populations in semi-arid landscapes of Queensland is
problematic. In many cases, these semi-arid rangeland areas are suffering from problems of overgrazing and land degradation, and are susceptible to clearing pressures as graziers attempt to improve the productivity of their land. These pressures are likely to impact over the long term on koala populations.
A closer look at the issues involved can be gained from a case study of the Desert Uplands, one of 13 biogeographic regions of Queensland. This region of scattered woodland country covers approximately 4% of Queensland, straddling the low tablelands of the Great Dividing Range from a little south of Jericho to a touch north of Torrens Creek. The region supports scattered but relatively stable populations of koalas, particularly on the northwestern side.
While current tree clearing guidelines limit clearing on leasehold land to approximately 80% of many vegetation types, and protect many rare and isolated vegetation communities, there is still large potential for wide-scale tree clearing to occur. Almost no restrictions occur over freehold land, and few mechanisms exist to signal the wishes of the wider community for biodiversity conservation.
In this paper, some of the philosophical and ethical issues underlying species and biodiversity protection are outlined, and then compared to the current incentive structure facing landholders and resource managers in the Desert Uplands. The inadequacy of current institutional and land tenure systems is presented as a major cause of land use conflicts, together with some mechanisms to address these deficiencies. Among the first steps is development of non-market valuation techniques to estimate public demands for conservation of koala habitat.
Introduction Habitat destruction is seen as the greatest single threat to koala populations in
Queensland, and habitat retention is one of the simplest solutions to conservation issues (Melzer and Lamb 1992). Yet clearing rates are still high across Queensland, particularly in the acacia scrub and eucalypt woodland areas that characterise much of Central Queensland (McCosker and Cox 1996). In many areas, koala populations are sporadic and scattered, little is known of them, and the immediate impacts of clearing on populations are often not fully apparent. However, as vegetation areas dwindle, and as habitats become more fragmented, impacts are likely to be larger.
The impact of broadscale tree clearing in one particular area of Queensland, the Desert Uplands bioregion, illustrates some of the issues. This region of semi-arid woodland country in central-western Queensland supports scattered populations of koalas across much of the region (Melzer and Lamb 1992; Munks et al. 1996). While vegetation along the watercourses is usually the optimum habitat for these populations, they also utilise a broad range of the various woodland communities (Munks et al. 1996). These communities coexist with the pastoral use of that land, mostly for the purposes of grazing cattle.
139

In an effort to improve the productivity of their land, graziers in the region have been clearing the trees and introducing improved pasture. Initially these activities were confined to the small areas of relatively fertile soil, but the last few years has seen wide scale clearing in the less fertile country, particularly the open eucalypt woodlands of ironbark and box (McCosker and Cox 1996). While some doubts exist about the long term profitability of these developments, the overwhelming reason why clearing occurs is because pastoralists are attempting to improve their economic viability in the face of changing and deteriorating market conditions (Rolfe et al. 1997).
However, the wider Australian community is almost a bystander to this process. On the one hand, their demands for agricultural produce are reflected through market signals, providing clear incentives for pastoralists to improve their productivity. On the other hand, Australians value the conservation of biodiversity, including the preservation of koalas, one of the best known of the Australian mammals. These demands for preservation options are not being transmitted very effectively to the pastoralists and other land managers.
The principal mechanism for recognising conservation and preservation demands occurs through the institutional and regulatory process. Recent changes to tree clearing guidelines in Queensland provide an example of how this institutional process might work. However, institutional mechanisms are not necessarily efficient or effective at balancing demands. In many cases, faulty institutions are an important explanation of why environmental problems occur in the first place (Wills 1997). As well, most institutions, such as land tenure conditions, are simultaneously targeting a number of goals, and thus do not necessarily provide the right incentives for careful environmental management (Holmes 1996).
Environmental economics can address these, and related issues, in two broad ways. First, it can assess the social welfare implications of particular resource use issues by quantifying and comparing the various outcomes of the alternatives. For the issue of interest, koala populations affected by tree clearing in the Desert Uplands bioregion, a comparison might be made of the social welfare changes attached to both preservation and tree clearing options for an area of koala habitat.
The second broad application of environmental economics is in evaluating the effectiveness of management and institutional instruments. Environmental problems generally occur as a result of inappropriate incentives and signals that are being transmitted to decision makers and resource users. In some cases these occur because poor property rights lead to open access and depletion, or where property rights do not recognise the interests of all affected parties. In other cases, the problem of 'missing markets' means that some wishes of society are not transmitted to resource users. Evaluating the incentives and signals that people face provides a pathway to optimising resource use mechanisms.
This paper explores the relevance of environmental valuation techniques to a particular case study, the conservation of koala populations in the Desert Uplands bioregion. The paper is organised as follows. In the next section, a description of the Desert Uplands bioregion is presented, followed by an analysis of the tree clearing debate and the current impacts of tree clearing in the region. Following this is an overview of the various legislative and regulatory mechanisms that may impact on conservation of koalas. Discussion and conclusions will then focus on the potential for economic analysis to provide some insight into the efficient allocation of resources in society.
140

Koala Populations in the Desert Uplands Bioregion The Desert Uplands is a band of scattered woodland country straddling the low
tablelands of the Great Dividing Range from a little south of Jericho on the Capricorn Highway to a little north of Torrens Creek on the Flinders Highway. One of thirteen terrestrial bioregions of Queensland, it lies between the drier mitchell grass plains to the west, and the wetter brigalow (Acacia harpophylla) country to the east. The bioregion is almost the same size as Tasmania, covering approximately 4% of Queensland.
The climate in the Desert Uplands can generally be classified as semi-arid (Smith and Rowland 1991). Rainfall generally declines from east to west across the region, varying from approximately 450mm per annum on the western side to 600mm per annum on the eastern side. Evaporation rates in the region are high, and are often in excess of 3100mm per year. Together with high temperatures and the rainfall variability, this means that seasonal conditions range from being lush to extended periods of dry conditions and droughts.
The Desert Uplands has been divided into three ecological regions, the PrairieTorrens Creek province to the north west, the Cape-Campaspe province to the northeast, and the Alice Tablelands province in the centre and south of the region. The region forms part of the watershed between the Burdekin catchment and the Lake Eyre catchment, with a small portion of the region to the north draining to the Gulf of Carpentaria. There are two notable internal drainage basins to Lake Buchanan and Lake Galilee.
The Desert Uplands have poor soils relative to other areas of Queensland. Much of the region represents the remnants of ancient, deeply weathered plateaus together with associated alluvial fans. Much of the Alice Tablelands province is covered by red and yellow earths that vary between sandy and loamy textures. These are termed 'massive' soils, as distinct from 'structured' soils. These massive earths tend to be infertile, with, for example, very low levels of phosphorous (Ahern et al. 1994 ).
Vegetation types can be classified into three broad groups. The most dominant are the eucalypt woodlands of box (E. populnea, E. brownii) and ironbark (E. crebra, E. whitei, E. melanophloia), together with communities of bloodwoods (eg Corymbia lamprophylla) and yellowjacks (eg E. similis) (Anderson 1993; Brooker and Kleinig 1994; Rolfe et al. 1997). Smith and Rowland (1991) estimate that these communities cover 86.2% of the Desert Uplands region.
The second most common vegetation communities occur on the pockets of fertile clay soils through the region. These are often the acacia communities of brigalow, gidgee and blackwood, although it also includes some areas of box on alluvial flats, as well as other species associated with these. In the third group are vegetation areas on skeletal soils associated with rocky outcrops and escarpments. Within these very broad classifications, a number of different ecosystems may be distinguished. For example, Morgan (1997) has categorised at least 58 different ecosystems for the bioregion.
Scattered populations of koalas occur through the region, but appear to be most common in the Prairie-Torrens Creek province. The 1967 survey of the Wildlife Preservation Society of Queensland indicated that koala populations were located in both the southern and northern section of the region. The National Koala Survey indicated that most koala populations were located in the southern part of the region, with small numbers in the Prairie-Torrens Creek province (Phillips 1990). Environmental Protection Agency data indicates a similar pattern (Melzer and Lamb 1992). However, Munks et al. (1996) suggest that the Prairie-Torrens Creek province is the most favoured area in the Desert Uplands for koalas (as well as for brushtailed
141

possums and a glider). Their survey work indicated that koalas were using ten of the eleven land types surveyed in that province, with populations favouring watercourse areas. They suggest that
koalas are more widely distributed in the semi-arid woodlands of the Desert Uplands than previously indicated by more broad-scale surveys (Munks et al. 1996 p.192).
These discrepancies in the reported distributions of koalas in the region can be explained principally by the low levels of information available. The Desert Uplands region is sparsely settled, and until recently, little effort had been made to assess the environmental resources of the area (Rolfe et al. 1997). There are low levels of understanding about the occurrence and interrelationships between fauna and vegetation types, largely reflecting the lack of scientific data available. In part, this lack of previous interest has been justified on the basis that grazing activities have been low impact relative to other areas of Queensland1
• However, as broadscale tree clearing intensifies in areas of the Desert Uplands, concerns about potential environmental impacts have increased interest in the scientific assessment of the region's natural resources.
Melzer and Lamb (1992) report that at a study site in the Springsure region, koalas frequented vegetation communities along both the streamlines and surrounding hills, favouring areas of more fertile soils. In a similar result, the study of Munks et al. (1996) indicated that koalas and possums made extensive, but intermittent, use of a wide range of land-use types. In the Prairie-Torrens Creek province, the vegetation on these land use types included gums (E. coolabah ), ironbark (E. camaldulensis, E. whitei), bloodwood (C. terminalis), box (E. brownii), gidgee (A. cambagei) and blackwood (A. argyrodendron).
Melzer and Lamb (1992) noted that koalas in the Springsure community were observed eating E. tereticomis, E. crebra, E. melanophloia, E. orgadophylla and E. exserta, and were probably utilising other species. All of these communities are present in the Desert Uplands region. Other vegetation types that may be important for koalas that occur in the Alice Tablelands province include C. tessellaris, C. citriodora, E. camaldulensis and E. populnea2
•
While these studies show that koalas use a broader range of land types than those along watercourses, they also highlight the patchiness of koala distributions. This selected distribution appears common across eastern Australia (Munks et al. 1996). The census figures reported in Phillips (1990) shows that the semi-arid zones of Queensland exhibit similar patchwork characteristics of population distribution, with extensive areas where koalas are absent or extremely rare. What is not always clear are the reasons why some areas are favourable for koala communities and others are apparently not (Melzer and Lamb 1992, Munks et al. 1996).
It appears that some linkages exist between population distributions and the fertility and nutrient levels of the corresponding soil types. Melzer and Lamb (1992) report higher densities of koala populations occurred on the more fertile basaltic clay soils at their Springsure study site, while lowest density populations occur on the least fertile, sandy soils. Monks et al. (1996) hypothesis that this general relationship explains why the Prairie-Torrens Creek province, with its more fertile clay soils, supports more extensive koala populations than other parts of the Desert Uplands.
Both Melzer and Lamb (1992) and Monks et al. (1996) report strong correlations between koala populations and creek lines. This appears to be because foliar moisture
1 In some areas of the region, lack of surface water and the presence of poison heartleaf, a small bush poisonous to stock, has restricted the impact on the region by domestic animals. 2 See also Gordon et al. (1990).
142

levels tend to be higher along watercourses, although tree size and density also tends to increase. The latter may be important in terms of available food and shade (Monks et al. 1996). As well, there may be some relationship between toxin levels in eucalyptus leaves and both moisture levels and nutrient levels that explains population distributions (Cork 1995; Munks et al. 1996). In a semi-arid environment, it is not surprising that moisture availability should be a major determinant of habitat choice. Mo.nks et al. (1996) report that water availability rather than the level of soil nutrients tends to be the major indicator of preferred habitat in the Desert Uplands.
Tree Clearing in the Desert Uplands The principal threat to koala populations in Queensland is habitat destruction
(Phillips 1990). This is generally because koala populations tend to concentrate in areas of higher soil fertility, which are also the most favoured areas for development (Melzer and Lamb 1992; Cork 1995). The same tradeoffs that have occurred in southeastern Australia are now starting to occur in the Desert Uplands, where landholders are becoming more interested in developing their better soils for pasture purposes.
The effect of tree clearing on koala populations is not always easy to ascertain. In~ general terms, the high levels of tree clearing that has occurred in Queensland, particularly in the brigalow region, has reduced areas of koala habitat (Melzer and Lamb 1992). For example, Sattler (1993) estimates that less than 1% of fertile soils along Van Dyke Creek (in the Springsure district) remain uncleared. Compared to the brigalow region and other land types in south-eastern Queensland, the Desert Uplands is still relatively intact. McCosker and Cox (1996) indicate that nearly 40% of the south-eastern side of the Desert Uplands has been cleared. There has been little clearing in the north-east, and moderate clearing in the north-west and south-west of the region. Overall clearing rates for the Desert Uplands are likely to lie between 15 and 20%3
•
The effect of tree clearing in specific cases relates to the actual habitats used by koalas. There are large areas of the Desert Uplands where koalas are not present and where subsequent tree clearing will have little or no impact on populations. However, tree clearing tends to be more concentrated in areas of fertile soils, and this is where potential for conflict exists between development and conservation options. As the available acacia communities become depleted, there is increased interest in clearing eucalypt woodland areas and areas of alluvial soils along watercourses throughout the Central Queensland region (McCosker and Cox 1996). While the brigalow and gidgee communities did support low density koala populations (Melzer and Lamb 1992; Munks et al. 1996), some of the eucalypt communities may be more significant habitats. This implies that significant tradeoffs currently exist between clearing and conservation options.
While the financial returns from clearing in areas of highly fertile soil are usually high,- the evidence about the financial returns from clearing in the Desert Uplands is mixed (Rolfe et al. 1997); The principal reason for clearing is to introduce improved pasture for beef cattle. Landholders clear vegetation because they expect the costs will be returned in the form of increased production. In the better quality clay soils that supported acacia scrubs, these improvements have been relatively successful.
In the poorer quality soils, particularly those supporting the eucalypt woodlands of the Alice Tablelands province, some doubt exists about the long-term viability of clearing. In part the problems relate to low levels of soil fertility, particularly extremely low levels of phosphorous. In many areas, significant re-growth of eucalypt
3 Estimation is not straightforward because of the lack of knowledge about the extent of pre-European vegetation and the subsequent occurrence of woody weeds.
143

seedlings often occurs, which may be impossible, for financial reasons, to control. As well, the porous nature of many of the soils means that lower levels of available moisture are present compared to the clay soils. Over the longer term, some improved pastures have been found to be no more productive for grazing than the naturally occurring spinifex pastures (Lorimer, Environmental Protection Authority, pers comm).
Legislative and Regulatory Controls There are two principal legislative controls that impact on the conservation of koala habitat in Queensland. These are the Land Act (1994) and the Nature Conservation Act (1992). Under the Land Act (1994), pastoralists on leased land (pastoral holdings and grazing homesteads) need to apply for permission for broadscale tree clearing. Approximately 80% of the Desert Uplands region is held as leasehold land, with some areas of freehold land being located in the western and southern sections of the region. This means that most tree clearing in the region only occurs after a permit to clear has been issued from the Department of Natural Resources. There are no legislative or policy controls over clearing on freehold lands.
The Land Act in Queensland has historically been focused on improving the State's productive base. For these purposes, it has been used to sub-divide holdings for closer settlement, promote land ownership through conversion to homestead and freeholding titles, and to improve productivity through the application of lease conditions. Approximately 68% of Queensland is still held under leasehold title, mostly in the arid and semi-arid rangelands area (Sattler 1993; Holmes 1996).
Freehold title does have some restrictions on the property rights of landholders. Generally, landowners do not have title to mineral resources, surface and sub-surface waters, or to wildlife. Lease tenures are more restricted in scope. Among the important attributes of leasehold title are limitations on land use, being generally confined to pastoral purposes, as well as limitations on the clearing and removal of trees. Many of the development requirements of leases, relating to stocking rates, capital investment and residence, are now obsolete. More recently, attention has focused on issues of sustainability and property planning, and this "duty of care" emphasis is becoming more pronounced (Holmes 1996).
These trends can be illustrated in relation to the tree clearing guidelines. Prior to the 1990s, tree clearing targets and subsequent re-growth control were often specified within the lease conditions of landholders. These conditions were aimed at improving the productive land base, and although some minimum conditions applied, these were usually ignored and not enforced. Among the minimum conditions were recommendations for a 10% vegetation retention rate, and restrictions on clearing within 40 metres of a watercourse (Sattler 1993). Emphasis on production has resulted in very high rates of clearing in the more fertile soils of Queensland. Some vegetation communities, such as specialised brigalow, softwood and coastal woodland areas, have less than 1% of original cover remaining (Rolfe et al. 1997).
The 1990 Wolfe Report into Land Policy and Administration in Queensland flagged the change in emphasis from development goals to wider goals of sustainability and public interest. This report noted that leasehold land needed to be better managed, and that community wealth did not simply relate to the grim determination to satisfy the more immediate and pressing of fiscal demands (p.lO). However, while the Report recommended that: As a general principle, Crown land which is unique, especially valuable, arid or semi-arid, has environmental value or is vulnerable to degradation, or presents other special problems should not be freeholded (p.i), few other specific recommendations relating to these goals were made.
144

In March 1995, the Queensland Government released its Guidelines For Broadscale Tree Clearing On Leasehold And Other State Lands In Queensland.. The policy recognised:
that to meet the State's economic development policy objective the clearing of trees is often required. When doing so the State's interests in the protection of specific on-site and off-site values such as habitat protection and biodiversity need to be recognised (p.l).
The need for the Policy was explained on the basis that tree clearing, is now increasingly encroaching on the more marginal land types and the remnants or pockets of uncleared areas in the extensively developed parts of the State (p.5).
Action was seen to be required in order to avoid irreversible environmental losses and costly recovery and rehabilitation programs where impacts are reversible. The Draft State guidelines outlined that the
protection of wildlife corridors, particularly riparian zones along rivers, creeks and gullies is recognised as the singularly most important action to guarantee the distribution and abundance of wildlife throughout the landscape. . ..... the protection of riparian corridors for wildlife habitat purposes requires substantially larger buffers than are required for bank stability and erosion control. In this respect, they are as significant to the maintenance of wildlife as coastal and estuarine mangroves .... Poorly managed tree clearing also leads to soil erosion, salinity, weed invasion, damage to streams and rivers, loss of water quality and a range of other land degradation problems, as well as direct costs to affected industries and urban communities (p5).
Partly in response to criticisms of the draft by landholders (Rolfe et al. 1997), the Queensland Government issued the Preliminary Tree Clearing Policy (PTCP) on the 18th of December 1995 as a reference document for the development of local guidelines for broadscale tree clearing, replacing the earlier Draft State Guidelines. The local guidelines were seen as a format for landholders to have some input into the policy for their region, as well as ensuring that some flexibility would be generated within the overall policy framework. The main aims of the policy (consistent with principles outlined in the Land Act) were to:
maintain the productivity of the land; allow the development of the land; prevent degradation of the land
0 maintain biodiversity 0 maintain the environmental and amenity values of the landscape, and
maintain the scientific, recreation and tourism values of the land (p.2). The guidelines outlined in the PTCP divide Queensland into 7 proposed clearing
zones, based on native pasture communities. The Desert Uplands forms one of those zones, with the following description attached:
There is considerable activity in relation to tree clearing in this zone, with sustainable development occurring. The region consists of infertile eucalypt woodlands interspersed with small areas of fertile clay soils. Regional wildlife movement considerations make this zone an important one in which to maintain woodland cover (p.lO).
The PTCP states that the local guidelines within each clearing zone are to be developed through a process of local and community consultation. Applications for tree clearing permits will then be evaluated against these local guidelines once they become statutory additions to the Land Act.
145

Compliance with the guidelines is to be encouraged through the progressive identification and mapping of sensitive and critical areas, and through on-site inspections. On a broader scale, monitoring of tree clearing in the state is to be assessed through effective remote sensing.
The Land Act does not generally relate tree clearing permits to the protection of specific items of fauna and flora. In part, this is because the Nature Conservation Act (1992) outlines a number of mechanisms by which environmental resources may be protected. Under that Act, the range of recognised protected areas includes national parks, conservation parks, resources reserves, nature refuges, coordinated conservation areas and wilderness areas. These represent a spectrum of mixes between conservation and other uses, including Aboriginal and Islander use, as well as pastoral use. Nature refuges and coordinated conservation areas are designed to co-exist with pastoral land holders.
As well, the Nature Conservation Act includes provisions for interim conservation orders that may be used to halt development activities that impact on endangered wildlife. These orders have been applied in northern Queensland to areas of habitat for the mahogany glider. The Act also allows for the compulsory declaration of nature~ refuges where the area is of major interest or a critical habitat. In general though, the Act envisages nature refuges to be voluntary agreements between landholders and the government, where the landholders refrain from certain activities. A conservation covenant may be balanced by compensation payments to the landholder.
In practice, nature conservation in Queensland has been dichotomous rather than being multi-faceted in application. The emphasis has been on increasing the proportion of the State held in national parks, rather than on developing the joint use approaches implied in the nature refuges and coordinated conservation area mechanisms. While the thrust of increasing the representation of national parks in Queensland is consistent with the current low status of representation in many of the biogeographic regions of Queensland (Walsh 1996), the ramifications of the policy are that increased resources are necessary for purchase and subsequent management of these lands.
In conclusion, there have been substantial changes in the legislative and regulatory mechanisms in Queensland that focus much more specifically on sustainability issues and the protection of environmental resources. However, these changes have limited application to koala populations in the Desert Uplands. The conservation of vulnerable and endangered vegetation types will assist koala populations to the extent that they use these vegetation types. As well, the conservation of vegetation along watercourses will aid koala populations. However, the provisions will still allow substantial amounts of tree clearing to occur in the Desert Uplands, and substantial permits for this clearing have already been issued (Rolfe et al. 1997). As well, there are few mechanisms under the Land Act for considerations about koalas and other non-endangered species of wildlife to be incorporated into decisions about permits for tree clearing. This is particularly the case in decisions about buffer zones and vegetation strips.
The application of the Nature Conservation Act also offers little protection to scattered populations of koalas. The thrust of nature protection in Queensland has been to increase the area and representation of national parks, rather than to explore opportunities for joint resource use that combine production and conservation goals. The use of interim conservation orders to stop specific acts of tree clearing appears to be confined in practice to endangered species.
146

This means that while the Land Act and the Nature Conservation Act help to set out some minimum standards of conservation, they do not necessarily meet society's preferences for actual tradeoffs. To explain this difference, an economic overview of preferences is set out in the following section.
An Economic Assessment of Conservation Options Resource use decisions are often difficult because of the variety of competing
interests. The complexity of many decisions, coupled with the long term nature of many outcomes, means that it is often difficult to fully assess the consequences of outcomes. Environmental losses, such as a reduction in koala populations, are one example of a problem that no one sets out to cause deliberately, but occur as a result of side-consequences of other choices.
In the Desert Uplands, there are clear signals from the Australian community about the benefits of increased productivity through market signals for beef and wool. However, Australian people also have preferences about other factors. In some cases, these are for the resources to be directly used for some other purpose, such as for tourism. The market is one mechanism for deciding which direct use offers the most benefit to society, and acts, for example, to allocate land between wool and beef production.
There is also a range of indirect benefits that the Desert Uplands may provide (such as acting as a carbon sink against greenhouse gases) that are more difficult to quantify. These indirect benefits (and costs) are more difficult to include within market signals, and there is usually a role for government to play to ensure that resource users take some account of the wider range of benefits.
Australian people also hold a range of non-use values for environmental resources which are even more difficult to quantify. These include existence values, which reflect the benefits that an individual may receive from simply knowing that a species exists, even if they never visit or see it. The benefits that people receive may relate to their wishes for other people and future generations to be able to value and enjoy vicarious consumption of these environmental assets. Thus existence value may incorporate substantial components of bequest values. Option values and quasi-option values relate to the opportunities for future use and for better decisions that may result from current conservation of an environmental asset.
Government intervention in market processes is also warranted where no account may be taken of non-use values. Divergences between public and private benefits mean that individual resource users may face incentives that are not very consistent with the public interest, creating a market failure problem. The government may intervene in a number of ways, such as through the reallocation of property rights, the imposition of regulations, and the use of taxation and other pricing mechanisms.
Koala conservation in the Desert Uplands is a clear example of a market failure problem. Koalas are a species that the general public has high values for, both in terms of indirect use (tourism, vicarious consumption), and preservation or non-use values (existence value, bequests for future generations). In the Desert Uplands, most values are likely to be for preservation in terms of existence value. Yet tree clearing decisions are made with almost no consideration for the public wishes for preservation of koala habitat.
The current permits for tree clearing take no account of whether arboreal marsupials are present or not in the areas to be cleared. While the protection of endangered and vulnerable vegetation communities and riparian zones will have some positive effects on koala communities, there is still no mechanism to differentiate land clearing permits for koala habitat.
147

This means that while society may have preferences for development in those areas that koalas do not inhabit, rather than for vegetation communities that koalas currently use, there is no mechanism to transfer those wishes to resource users. In particular, this means that landholders do not receive -the full range of incentives that would promote efficient resource use. Landholders who currently conserve koala habitat get no recognition or compensation for their efforts, while those who clear vegetation benefit from increased production from their introduced pastures.
Among the main reasons why there are no clear mechanisms to reflect society's wishes are problems in the quantification of these indirect and non-use values. Because non-use values are not transmitted through market mechanisms, there are no clear signals to provide estimates of their magnitude. For this purpose, economists have been developing a range of valuation techniques to assess indirect and non-use values (Morrison et al. 1996). Many of these rely on the quantification of values that people state they have, rather than values that may be revealed through market operations. Economists set up hypothetical market tradeoffs involving a good of particular interest and ask people to state what their tradeoff would be. The techniques are described as non-market stated preference techniques.
Contingent valuation, the principal stated preference technique used by economists, has been used to value the existence of a variety of rare and threatened species including grizzly bears, bottlenose dolphins and whooping cranes (Pearce 1996). However, there are some difficulties in applying the contingent valuation technique to situations that survey respondents have little understanding or knowledge of. It is likely that biases occur in the results because the information given causes problems of framing and scoping to occur4
• Framing problems relate to the difficulties that a respondent may have in structuring the particular issue in question in relation to other issues of relevance. Scoping problems suggest that respondents are indifferent between different amounts of the environmental good of interest.
Using the contingent valuation technique to value koala habitat in the Desert Uplands region would be likely to generate problems involving both scoping and framing issues because the Desert Uplands region is remote and unknown to most Australians. As well, the contingent valuation method is really a 'one-shot' technique useful for capturing the value difference between the status quo and a development option, but is not so useful when a range of possible options are to be considered. The latter is the case in the Desert Uplands where tradeoffs between preservation and development vary across properties, ecosystems and vegetation types.
The Choice Modelling technique has been developed to address some of these difficulties (Morrison et al. 1996), and appears more appropriate for estimating preservation values for koala habitat than does the contingent valuation method. The technique operates by presenting the issue in question as the function of a number of attributes. By giving respondents choices that are variations of these attributes, and repeating that process, information is generated about the contribution of each different attribute to the choices. By including an attribute that involves cost or price (such as a tree levy), it is possible for the researcher to estimate how respondents tradeoff other attributes against that cost attribute. A multi-nominal logit estimation procedure is used for this purpose. In this way, the Choice Modelling technique can generate information about how respondents perceive the value of changes in different attributes and, depending on the application, the value of particular overall scenarios.
4 There are a range of other biases that may be associated with the contingent valuation technique (Mitchell and Carson 1989).
148

The outcomes of a Choice Modelling experiment are useful to a policy analyst because value differences between a large range of policy options can normally be generated. As well, the technique often gives some indication of the choice processes used to make decisions, and of the marginal values associated with particular attributes. Rolfe and Bennett (1995, 1996) and Rolfe et al.(1997) report on the application of the technique to valuing overseas rainforests. Rolfe et al. (1997) and Blarney et al. (1997) report on the potential application of the technique to valuing remnant vegetation in the Desert Uplands region of Queensland.
The overall assessment of tree clearing options in an area where koala habitat is involved would require the use of a technique such as cost-benefit analysis. Under this framework, the benefits flowing from a development option would be compared with the costs to determine if net benefits can be generated. Many of the production benefits and the direct costs of development can be taken from relevant market information. However, impacts on environmental factors (such as the loss of koala habitat) should also be included, as should other social factors (eg. loss of regional employment). Non-market valuation techniques can be used to add this 'missing' information. The policy analyst can then determine the overall value to society of proposed developments, and the potential for government involvement to address problems of market failure.
Policy Implications of Valuing Koala Habitat in the Desert Uplands From a policy perspective, the issue of interest is the extent to which the Australian
community holds higher values for vegetation areas that are used by koalas over other vegetation areas. Information about these values can be used in two main ways.
First, the relevant value of tree clearing options can be more accurately addressed by decision makers. For example, the marginal production of some areas of the Desert Uplands means that tree clearing and pasture improvement has limited commercial benefits. This is particularly the case for areas of less fertile soils. In some cases the inclusion of non-use values for preservation options in a total economic framework will be sufficient to demonstrate that the community derives greater benefit from conservation options that retain koala habitat than from tree clearing and pasture development options. In some cases decision makers may be able to use information about conservation values to develop specific management options. These may include options such as the extension of riparian buffer zones in areas of koala habitat and the purchase of significant environmental areas for preservation purposes.
The second principal use of information is in assessing values for compensation and transfer of rights purposes. There are perhaps three main areas where this may be applicable.
(a) Under the Nature Conservation Act 1992, provision is made for agreements to be made between landholders and the government that combine both preservation and production goals. These provisions involve a transfer of rights to the Government through some form of covenant that is binding on the landholder and subsequent title holders. In return, landholders may receive some form of compensation under section 45 of the Act. Production losses would probably be the minimum level that such compensation amounts could be set at. For landholders to receive an incentive to enter into such a voluntary agreement, though, compensation amounts would need to be higher. At the other end of the scale, preservation values that society may hold will form the upper limit to such compensation agreements.
149

(b) Provision is made under the Nature Conservation Act 1992 for compulsory imposition of conservation orders and agreements. As well, it would be possible for further amendments to occur to the Lands Act 1994 that may give greater protection to environmental assets. The introduction of controls on freehold title that related to vegetation and wildlife conservation would be one possible form of such an amendment. These changes would effectively transfer property rights from the landholders to the Government through the use of changes to tenurial or regulatory instruments. While the effect will vary according to whether changes involve proprietary power (changes in tenure and conditions) or sovereign power (changes in the rules and regulations governing society), the transfer of rights will involve issues of compensation (Holmes 1996). The valuation of preservation benefits will provide one means of assessing such compensation amounts.
(c) Preservation and conservation activities are not exclusively in the public domain. Private individuals and markets can also operate to preserve ecosystems and species. Bennett (1995) provides some evidence about the success of private protection in both Australia and overseas locations. Current proposals by Birds Australia to purchase Gluepot Station in South Australia for $350,000 and raise the money through public donations is a fine example of private conservation at work.
The Choice Modelling technique may provide very useful information into this process of private conservation in several ways. First, it can provide estimates about the willingness to pay by Australians for preservation issues. Second, through the collection of socio-economic data, it can identify the groups that have the highest interest in paying for conservation. Thirdly, through the valuation process, it can identify the scenarios and tradeoffs that people find most appealing.
Conclusion Issues of broadscale tree clearing in the Desert Uplands bioregion of Queensland
illustrate many of the complexities of koala management in non-urban environments. Scattered populations of koalas exist in the region, particularly in the Prairie-Torrens Creek province. Their presence appears to be related to both aspects of soil fertility and moisture availability. This means that their preferred habitats appear to be along watercourses in areas of higher soil fertility.
The Desert Uplands bioregion is a band of scattered woodland country where agricultural production is limited by relatively low rainfall, poor soils and pasture that is not very palatable for domestic stock. In recent years landholders have been clearing their country in an attempt to improve productivity. While changes to tree clearing regulations now protect endangered and vulnerable vegetation communities, they will allow extensive clearing of some eucalypt woodland communities. There is no provision in the regulations for koala habitat to impact on clearing choices.
The use of non-market valuation techniques such as Choice Modelling provides the potential for decision makers to assess the demands of the wider community for preserving koala habitat. This information can be used in two main ways. In the first, it can be incorporated within resource use decisions to assess the most efficient use of resources. In the second, it can assess compensation amounts that may be involved in voluntary agreements under the Nature Conservation Act, the forced transfer of property rights, or the establishment of private conservation initiatives. The application of non-market valuation techniques such as Choice Modelling to issues of koala habitat preservation should be the next step in improving information about preservation options.
150

References Ahern, C. R., Shields, P. G., Enderlin, N. G., and Baker, D. E. (1994). 'The Soil
Fertility of Central and North-East Queensland Grazing Lands'. Report prepared for the Meat Research Corporation and the Queensland Department of Primary Industries, QDPI, Brisbane.
Anderson, E. (1993). 'Plants of Central Queensland'. (Department of Primary Industries: Brisbane.)
Bennett, J. W. (1995). Protecting Nature ... Privately. Policy 11(3), 11-15. Blarney, R. K., Rolfe, J. C., Bennett, J. W., and Morrison, M.D. (1997).
'Environmental Choice Modelling: Issues and Qualitative Insights'. Choice Modelling Research Report No. 4, University College. (The University of New South Wales: Canberra.)
Brooker, M. I. H., and Kleinig, D. A. (1994). 'Field Guide to Eucalypts. Volume 3: Northern Australia'. (Inkata Press: Sydney.)
Cork, S. (1995). Life in a Salad Bowl? Australia Nature 25(2), 30-37. Gordon, G., Brown, A. S., and Pulsford, T. (1988). A Koala (Phascolarctos cinereus
Goldfuss) population crash during drought and heatwave conditions in southwestern Queensland. Australian Journal of Ecology 13, 451-461.
Gordon, G., McGreevy, D. G. and Lawrie, B. C. (1990). Koala Populations in Queensland: major limiting factors. (Eds. A. K. Lee, K. A. Handasyde and G. D. Sanson.) 'Biology of the Koala'. (Surrey Beatty and Sons: Sydney.)
Holmes, J. (1996). The policy relevance of the State's proprietary power: lease tenures in Queensland. Australian Journal of Environmental Management 3, 240-256.Land Administration Commission (Wolfe Report). (1990). 'Report of a Review of Land Policy and Administration in Queensland'. (Queensland Government: Brisbane.)
McCosker, J. C., and Cox, M. J. (1996). 'Central Brigalow Bioregional Conservation Strategy Report'. (Australian Nature Conservation Agency: Canberra.)
Melzer, A., and Lamb, D. (1992). 'Koala Habitat in Central Queensland: Implications for Management'. Paper presented to the Fitzroy Catchment Symposium, (University of Central Queensland: Rockhampton.)
Mitchell, R. C., and Carson, R. T. (1989). 'Using Surveys to Value Public Goods: The Contingent Valuation Method'. (Resources for the Future: Washington.)
Morrison, M.D., Blarney, R. K., Bennett, J. W., and Louviere, J J. (1996). A Comparison of Stated Preference Techniques for Estimating Environmental Values. 'Choice Modelling Research Report No 1.' (University College: University of New South Wales, Canberra.)
Munks, S. A., Corkrey, R., and Foley, W. J. (1996). Characteristics of Arboreal Marsupial Habitat in the Semi-arid Woodlands of Northern Australia. Wildlife Research 23, 185-195.
Pearce, D. (1996). Global Environmental Value and the Tropical Forests: Demonstration and Capture. In 'Forestry, Economics and the Environment.' (Eds. W. L. Adamowicz, P. C. Boxall, M. K. Luckert, W. E. Phillips and W. A. White.) (CAB International: Wallingford.)
Phillips, B. (1990). 'Koalas, The Little Australians We'd All Hate to Lose'. (Australian National Parks and Wildlife Service: Canberra.)
151

Rolfe, J. C., and Bennett, J. W. (1995). International Demand for Rainforests: Using Focus Groups to Establish Valuation Frameworks. Vanuatu Forest Conservation Research Report No 9. (University College, University of New South Wales: Canberra.)
Rolfe, J. C., and Bennett, J. W. (1996). Valuing International Rainforests: A Choice Modelling Approach. Vanuatu Forest Conservation Research Report No 12. (University College: University of New South Wales, Canberra.)
Rolfe, J. C., Bennett, J. W., and Louviere, J. J. (1997). 'Choice Modelling- An Exercise in Environmental Valuation'. paper presented to the 41st annual conference of the Australian Agricultural and Resource Economics Society, Gold Coast, January.
Rolfe, J. C., Blarney, R. K., and Bennett, J. W. (1997). Remnant Vegetation and Broadscale Tree Clearing in the Desert Uplands Region of Queensland. Choice Modelling Research Report No 3. (University College: University of New South Wales, Canberra.)
Sattler, P. (1993). Riparian Zone Management in Queensland and the Northern Territory: Policy and Practice. In 'Ecology and Management of Riparian Zones in Australia'. (Eds. S. E. Bunn, P. J. Pusey and P. Price.) LWRRDC Occasional Paper Series No:05/93, Canberra.
Smith, J. R., and Rowland, M. J. (1991). 'The Desert Uplands Biogeographical Zone (Queensland): A Heritage Resource Assessment'. Report to the Department of Environment and Heritage, Brisbane, and the Australian Heritage Commission, Canberra.
Walsh, K. (1996). Futures in Agricultural Production and the Environment. In 'Futures for Central Queensland'. (Eds. D. Cryle, G.Griffin and D.Stehlik.) (Central Queensland University: Rockhampton.)
Wills, I. R. (1997). 'Economics and the Environment: A Signalling and Incentives Approach'. (Allen and Unwin: Sydney.)
152

An investigation into the coprophilous my co biota of the koala (Phascolarctos cinereus, (Goldfuss, 1815))
Tanya Maxwell and Keith Harrower Biology Department, Faculty of Applied Science, Central Queensland University, Rockhampton, Queensland.
Introduction This paper reports some aspects of the work undertaken as part of an Honours
project on the coprophilous mycobiota of koala dung carried out at Central Queensland University during 1997. Firstly, it is essential to define some of the key terminology used in this paper. The term coprophilous comes from the Greek 'copros' meaning dung or excrement and 'philous' meaning loving. This term is used for an ecologically specialised group of fungi that may require their spores to be treated during the progression through the digestive tract before they will germinate (Webster 1970). For some species of coprophilous fungi the enzyme pancreatin is the trigger for germination. Mycobiota is a term used to collectively describe the fungi (or in this case the microfungi) that occupy an area, region or substrate. Simply put, the subject of this paper is a detailed study of the microfungi that are present in or on the faeces of koalas (Phascolarctos cinereus).
Even though dung has passed through the digestive tract of an animal, it retains many nutrients which can be utilised and liberated by the microbial inhabitants of that substrate. The establishment of coprophilous fungi on voided faeces generally requires the prior passage of their spores through the digestive tract of an animal (Cooke & Rayner 1984). Therefore, the life cycle of coprophilous fungi closely adheres to the ecological cycle that occurs between dung - herbage - animal gut - dung (Webster 1970). This process has been studied for a very large range of herbivores including rabbits (Harper & Webster 1964; Angel & Wicklow 1973; Gopal & Lodha 1982; Anson et al. 1985; Glocking & Dick 1994), sheep (Richardson 1972; Parker 1979), cattle (Dickinson & Underhay 1977; Khan & Krug 1989; Mahoney & Strongman 1994) and macropods (Harrower & Nagy 1979). Even though dung from any animal will yield fungi, that of herbivorous animals is by far the best source of coprophilous fungi. This is because the dung of either omnivores or carnivores is most likely to be decomposed by the action of bacteria rather than coprophilous fungi (Bell 1983).
The presence of coprophilous fungi on the dung of arboreal animals, including the koala, has not yet been investigated. This lack of recorded investigations of the coprophilous fungi that may be present in or on koala dung makes this an interesting subject for study. Unlike other herbivorous mammals, the koala is a fastidious eater, only feeding on the foliage of a select number of Eucalyptus spp. The consumption of eucalyptus by koalas has been an area of moderate interest due to the ingestion of essential oils contained in the foliage. These oils are not only toxic when ingested but are also known for their antimicrobial effects. A few studies have investigated the processing of these ingested oils to determine what happens to them during their passage through the animal. Eberhard et al. (1975) found that 7-30% of the volatile oils ingested were present in the faeces and 1% were in the urine, but some components that were present in the faeces were different to those ingested. This can be explained by the possible transformation of the oils in the gut. It has been stated that elimination of some of the essential oils in the gut occurs through the skin and surface of the lungs and that most of the oil is absorbed before it reaches the site of
153

microbial fermentation in the caecum and proximal colon (Eberhard et al. 1975). However, it is reasonable to conclude that the coprophilous fungi of koala dung would have to be able to tolerate these substances in order to survive the digestion process and reproduce in the voided faeces.
Methods The first aspect of the project involved a preliminary study aimed at investigating
whether or not koala dung supported a community of coprophilous fungi. This was essential because there are no documented investigations into the incubation of koala dung. When it was established that this substrate did indeed support a microfungal community, although quite different from other herbivore dung, experimentation was carried out to document the succession of these fungi over a period of time.
The Preliminary Study This experiment was conducted using twenty-four faecal pellets collected from one
identifiable koala at Kroombit Tourist Park near Biloela (150°45'E, 24°27'S). Six
pellets were placed in each of four glass petri dishes which contained filter paper kept
moist with reverse osmosis (RO) water. The pellets were examined daily for signs of
mycelial growth and for the presence of reproductive structures. A number of
microfungal species were isolated onto suitable media and incubated at 28°C.
The Main Project The main project was fundamentally the same as the preliminary study, however,
more pellets were used and each pellet was incubated in isolation from all others to allay any concerns of cross contamination. In order to complete this project a large and readily available supply of koala faecal pellets was required. Subsequently, it was decided that pellets would be collected from the koala population at Kroombit Tourist Park near Biloela and also from captive koalas living in the Rockhampton Botanical Gardens.
From 21 to 23 of February 1997, a field trip was undertaken at Kroombit Tourist Park to collect faecal pellets from known individual koalas. Using a radio tracking device, koalas were located by tuning to identifying frequencies for different individuals. Once an individual was located in a particular tree its identity was confirmed by examining the combination of coloured tags which were located on the individuals ear. Each koala had a unique identifier, which was a three digit number, which referred to its radio tracking frequency, followed by listing the first letter of each of the three coloured tags in the koalas ear (for example, coloured tags of red, orange and blue would be recorded as ROB while colours tags of green, orange and light blue would be recorded as GOLB). Once the koala was identified, a tarpaulin, which was sterilised by spraying it with 70% ethanol, was placed beneath the tree in an attempt to catch faecal pellets. Faecal pellets were collected from five koalas in the area. Each tarpaulin was checked after approximately 12 hours, when any freshly voided pellets were collected using sterilised forceps, placed into paper bags and sealed. These pellets constituted the "fresh" faecal pellet samples. Faecal pellets which were freshly voided but had either missed the tarpaulin or rolled off onto the ground were also collected from the same site. These constituted the "ground" samples. Since there was an obvious visual difference between fresh and aged pellets, it was easy to determine whether pellets collected from the ground were voided within the previous 12 hour period. These two sets of samples were collected in order to determine whether or not faecal pellets were contaminated by the soil. In addition to
154

the collection of faecal pellets, the tree species in which each koala was located when the faecal pellets were collected was also recorded.
While at Kroombit Tourist Park the base of known feeding trees were examined for old koala pellets. Observations were made of their state of decomposition and if there were any microfungi present. Photographs were taken to compare this natural state of pellet decomposition to the laboratory incubated pellets.
From the collected samples it was decided that the pellets from three koalas would be used for the laboratory incubation experiment as these were the only three individuals who had deposited an adequate number of replicate pellets for both the "fresh" and "ground" samples. Once in the laboratory the pellets were weighed, had their lengths measured and were then placed in experimental chambers; Figure I illustrates the experimental chamber. For each koala, ten "fresh" pellets and ten "ground" pellets were used, giving a total sample size of sixty pellets. As for the preliminary experiment, each chamber contained filter paper that was kept moist by periodic addition of RO water. One pellet was placed in each chamber with a glass microscope slide overlaying the pellet but not touching it in order to collect any ballistically discharged spores which the fungi might produce. The use of a glass slide -was based on the fact that some coprophilous fungi have a phototropic response and also that their spores are often mucilaginous and stick to vegetation upon impact. Any spores collected in this way could be readily isolated and used to initiate axenic cultures.
Chamber ---~~~o--
Glass Slide
Faecal Pellet Moistened Filter Paper
Figure I Diagram of an Experimental Chamber
All sixty chambers were set up in the laboratory which was maintained at a constant temperature (22°C), in a position that exposed them to the natural light fluctuations of day and night.
Chambers were observed daily for a period of 83 days, with the appearance of fungal reproductive structures documented in order to examine any patterns of coprophilous fungal succession. As reproductive structures appeared, aseptic techniques were used to isolate fungal species onto a number of different media. Media used included half strength Potato Dextrose Agar (HPDA), Czapek Dox Agar (CDA), Malt Extract Agar (MEA), V8 Vegetable Juice Agar (V8A), Sabouraud Dextrose Agar (SDA) and Rose-Bengal Agar (RBA). In addition to the commercially available media, excess koala faecal pellets were used to make an agar medium called Koala Dung Agar (KDA). All of the fungal ~ecies that appeared were inoculated onto a range of media types and incubated at 28 C in the dark. Those species which were unable to sporulate under these conditions were transferred to a position under a near ultra-violet light source (Phillips 40 Watts/08TL) in order to stimulate sporulation, as performed by Carlile (1965).
In addition to the sixty experimental chambers which were originally set up, another set of samples were incubated due to concerns raised about availability of 02 and the build up of volatile oils which may be leaching from the pellet into the chamber.
155

To eliminate these concerns another incubation chamber was designed. For this experiment pellets were collected from the Rockhampton Botanical Gardens with samples being taken from both koalas and Agile Wallabies (Macropus agilis) which were kept in relatively close proximity to one another. Faecal pellets were collected from the ground but it was not determined which individuals they came from. This experiment was set up during April 1997 and involved the use of twenty koala and twenty wallaby pellets, with ten of each pellet type being incubated in two different chamber types. The first chamber type was identical to the chamber used to incubate the original sixty samples, illustrated in Figure I. The second type of chamber, represented in Figure II, was a rectangular container made of clear plastic which contained the base of two glass petri dishes each with moistened filter paper (using RO water) and one dung pellet. In each plastic container the two glass petri dishes contained the same type of dung pellet, ie. two koala pellets in a container and two wallaby pellets in a container. The lid of the plastic container was only placed loosely on top thus allowing 02 to diffuse in and any volatile oils to diffuse out. Species
present were noted and isolated.
Plastic Container_.,.. ~Lid
Faecal Pellet Filter Paper
Glass Petri Dishes
Figure II Diagram of second experimental chamber, allowing entrance of 02 and
escape of volatile oils
Photographs were taken of both incubated pellets and reproductive structures of many of the isolates and used as a reference base for the process of fungal identification. A number of methods were utilised in an attempt to correctly identify all fungal species. A combination of the prepared axenic cultures and numerous taxonomic keys were used as well as different identifying media (Raper et al. 1965; Rifai 1969; Pitt 1979; Carmichael et al. 1980; Pitt & Hocking 1985). A JEOL Scanning Electron Microscope (SEM) - 5300L V was also utilised as a tool to gather information about reproductive structures, such as spore, conidiophore and phialide dimensions, in order to use the taxonomic keys. Fungal samples were prepared by attaching them to specimen stubs using double-sided adhesive and sputter coating these samples with 5nm of gold in a Polaron Plasma Beam Coating Unit. Images were then collected and fungal characteristics measured using the Oxford Instruments EXL2 Image analysis and x-ray analysis package.
156

Results The Preliminary Study
From the preliminary study it was found that koala dung supported a fungal community, but one which was very different from coprophilous communities previously recorded for other herbivores. The main observations noted from this study were that, in general terms, there was a paucity of recognised coprophilous species present, and also each pellet contained a different combination of species. That is, no two pellets had the exact same species composition.
The Main Project Observations conducted at Kroombit Tourist Park revealed no coprophilous fungi
on old koala dung. The major differences between the two pellet types were the colour, compactness of the pellet material and the amount of apparent moisture and oil, in that new pellets appeared to glisten in the sunlight. Furthermore, the fresh pellets had a strong scent of Eucalyptus oil while aged pellets did not.
The collection of koala faecal pellets from known individuals at Kroombit Tourist Park yielded over 230 samples and Table I provides a summary of the data collected. The sixty pellets chosen for incubation came from three koalas; two females (370 BRR and 329 BOO) and one male (268 LBOO). The average weights and sizes of these sixty incubated pellets are displayed in Table II.
Table I Sample data collected from Kroombit Tourist Park, 21-23 February 1997
Date Time Bag Koala Sex Tree Area Pellet Pellet Type No. Count
21/2/97 22:30 1 370 E. crebra Flats 6 Ground BRR
648 S?
E. crebra Ridge 10 Ground GOO
22/2/97 12:40 2
268 cf E. crebra Ridge 19 Fresh LBOO
" 13:30 4
350 S?
E. crebra Ridge 7 Fresh DBRR
" 14:00 7
370 S?
E. crebra Flats 13 Fresh BRR
" 16:40 8
370 S?
E. crebra Flats 5 Ground BRR
" 16:45 9
329 S?
E. Flats 7 Fresh BOO dichromophloia
" 18:30 10
329 S?
E. Flats 14 Fresh BOO dichromophloia
" 18:30 11
329 S?
E. Flats 9 Ground BOO dichromophloia
" 18:35 12
329 S? E. Flats 8 Aged
BOO dichromophloia " 18:35 13
157

" 18:35 14 ? ? E. Flats 27 Aged dichromophloia
" 18:45 15 ? ? E. Flats 24 Aged dichromophloia
23/2/97 10:00 16 329 ~ E. Flats 13 Fresh BOO dichromophloia
" 10:00 17 329 ~ E. Flats 21 Ground BOO dichromophloia
" 11:30 18 370 ~ E. crebra Flats 4 Fresh BRR
" 11:30 19 370 ~ E. crebra Flats 19 Ground BRR
" 14:47 A 648 ~ E. crebra Ridge 7 Ground GOO
" 15:25 20 268 a E. crebra Ridge 8 Fresh LBOO
" 15:25 21 268 a E. crebra Ridge 10 Ground LBOO
Incubation of the selected sixty koala faecal pellets emphasised the uniqueness of the fungal community noted from the preliminary study. With regular inspection it was clear that even pellets isolated from the same individual on the same day and at the same time had a different species composition. For example, three of the ten incubated "fresh" faecal pellets of koala 370 BRR at day 53 displayed different species compositions. Pellet 1 supported the reproductive structures of Aspergillus carneus, A. terricola, Cladosporium sp. and Fusarium sporotrichioides; pellet 2 had Penicillium decumbens and Sordaria macrospora while pellet 3 had P. decumbens, P. purpurogenum and another Penicillium sp .. The vast majority of incubated pellets were different from one another in species composition, which is different from any other documented observations of herbivore dung.
From the sixty incubated koala pellets a total of thirteen fungal species were noted, of which eleven species were successfully isolated into axenic cultures. Of all isolated species only one, Sordaria macrospora, is considered to be a true coprophilous species, in the classic sense. The remainder of the isolates belong to the Deuteromycotina which are not usually considered to be members of the 'classic' coprophilous habitat. The following species were present : Aspergillus terricola Marchal Penicillium decumbens Thorn Penicillium purpurogenum Stoll Aspergillus carneus Blochwitz Sordaria macrospora * Cladosporium sp Aspergillus parasiticus Speare Papulaspora sp Fusarium sporotrichioides Sherb Isolate Ten Penicillium sp
158
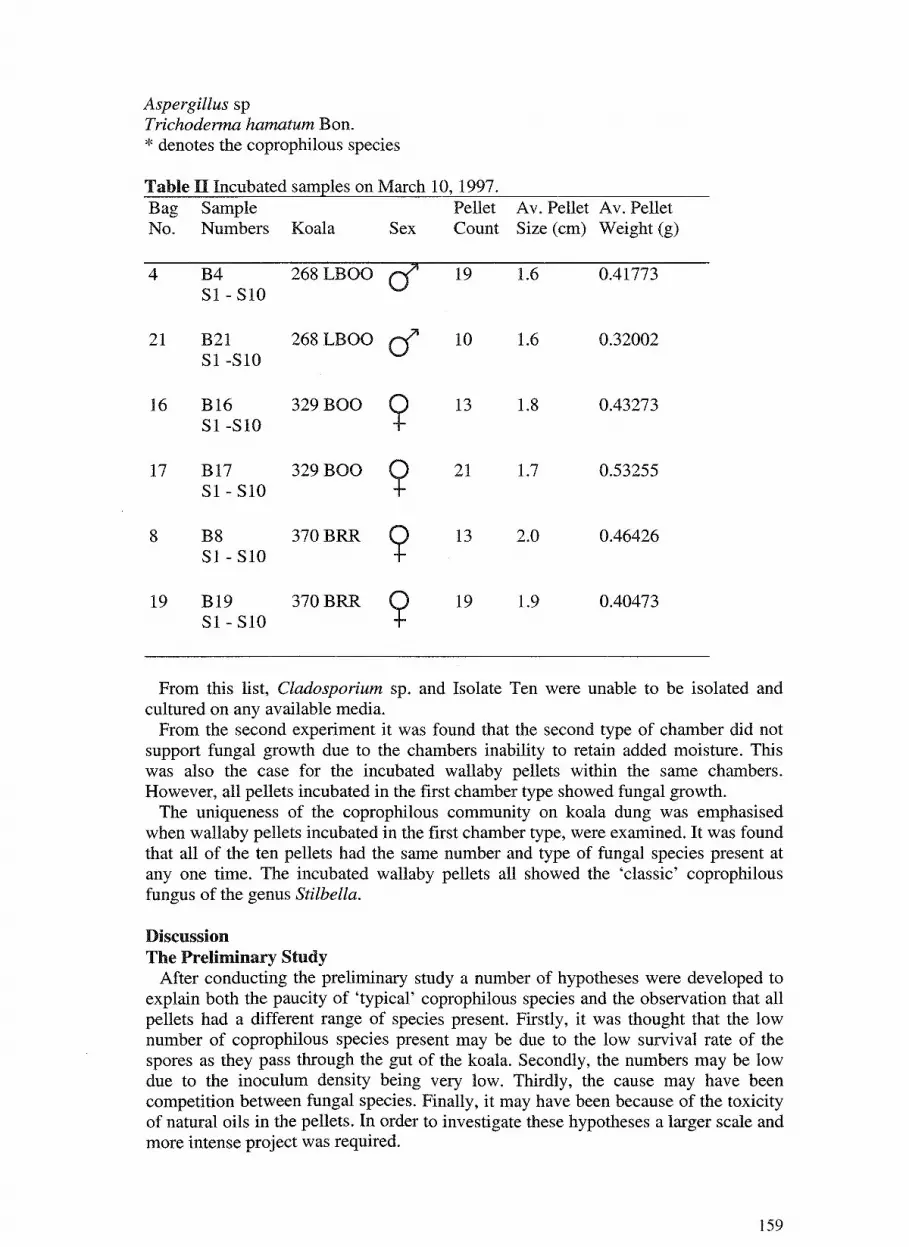
Aspergillus sp Trichoderma hamatum Bon. * denotes the coprophilous species
Table II Incubated samples on March 10, 1997. Bag Sample Pellet A v. Pellet A v. Pellet No. Numbers Koala Sex Count Size (em) Weight (g)
4 B4 268LBOO 0 19 1.6 0.41773 S1 - SIO
21 B21 268LBOO cf 10 1.6 0.32002 Sl-S10
16 B16 329BOO S? 13 1.8 0.43273 S1-S10
17 Bl7 329BOO S? 21 1.7 0.53255 S1- SIO
8 B8 370BRR S? 13 2.0 0.46426 Sl- S10
19 B19 370BRR S? 19 1.9 0.40473 S1- SlO
From this list, Cladosporium sp. and Isolate Ten were unable to be isolated and cultured on any available media.
From the second experiment it was found that the second type of chamber did not support fungal growth due to the chambers inability to retain added moisture. This was also the case for the incubated wallaby pellets within the same chambers. However, all pellets incubated in the first chamber type showed fungal growth.
The uniqueness of the coprophilous community on koala dung was emphasised when wallaby pellets incubated in the first chamber type, were examined. It was found that all of the ten pellets had the same number and type of fungal species present at any one time. The incubated wallaby pellets all showed the 'classic' coprophilous fungus of the genus Stilbella.
Discussion The Preliminary Study
After conducting the preliminary study a number of hypotheses were developed to explain both the paucity of 'typical' coprophilous species and the observation that all pellets had a different range of species present. Firstly, it was thought that the low number of coprophilous species present may be due to the low survival rate of the spores as they pass through the gut of the koala. Secondly, the numbers may be low due to the inoculum density being very low. Thirdly, the cause may have been competition between fungal species. Finally, it 111ay have been because of the toxicity of natural oils in the pellets. In order to investigate these hypotheses a larger scale and more intense project was required.
159

The Main Project It may be suggested that the lack of observed coprophilous fungi growing on koala
dung under field conditions may be as a result of the extreme dry conditions in the field. It is noted that in order for fungi to grow and ultimately produce reproductive structures they require a certain amount of water to be available in the substrate. Ultimately, at this point it should be noted that the experimentation which was carried out by the incubation of koala pellets in the laboratory under controlled conditions with the addition of adequate moisture and limitation of fungal predators such as insects is not a reflection of what occurs in nature. Instead, this manipulation is simply a representation of the potential of the koala faecal pellet if it is provided with optimum conditions to support and encourage fungal growth and reproduction.
On completion of the 83 days of incubation of sixty koala faecal pellets, it was noted that the lack of consistency observed in the preliminary study had again occurred. These pellets lacked the normal range of 'classical' coprophilous species, such as members of the Ascomycotina, the Mucorales (the so-called 'sugar' fungi) and many others. Instead, pellets had an unusually high proportion of species from the Deuteromycotina including Aspergillus, Penicillium, Cladosporium and Fusarium with each pellet displaying an individual pattern of species succession.
It may be postulated that the Aspergillus and Penicillium species (7 out of 13 species) observed on the incubated dung are contaminants from airborne spores from the surrounding environment, and that these spores had landed on the surface of the voided pellet and produced reproductive structures. This is a justified hypothesis knowing that these fungal species are frequently isolated from the air and soil (Lodha 1974). The hypothesis is further enhanced by a study conducted by Wicklow (1981) which found that potential fungal contaminants are, for the most part, unable to survive gut passage. Other studies have, however, found that several members of the Deuteromycotina (namely Aspergillus, Cladosporium and Trichoderma) are able to survive gut passage and are deposited in faeces (Lodha 1974). Bonner & Fergus (1959) recorded these species in both the feed and faeces of cattle in India. Therefore, it can be assumed that if these fungal spores are in the air and are able to be randomly deposited on faecal pellets they would also have the same likelihood of being randomly deposited on ingested foliage.
Since the majority of studies carried out on herbivore dung have been on animals which primarily graze on foliage close to the ground, it is easy to explain the inoculation of foliage. However, koalas feed on the foliage from the branches of tall Eucalyptus spp. and their faecal pellets are voided onto the ground below. Thus, inoculation of the foliage with fungal spores may be less frequent. Fungi have developed many modes of spore dispersal in order to achieve successful regeneration. Coprophilous fungi are known, in general, for their ballistic release of spores as a method of spore liberation which then subsequently allows the spores to be dispersed by the wind (Gregory 1966). In order to achieve successful dispersal a number of physical requirements must be met, including air movement caused by differences in pressure and temperature in different parts of the atmosphere. In order for the atmosphere to be suitable for dispersal of spores the movement of the wind, turbulence, viscosity, layering and convection must all be taken into account together with subsidiary factors such as the wind gradient near the ground, pattern of atmospheric circulation and solar radiation (Gregory 1966). In effect, if all of these conditions are 'optimal', effective air dispersal of fungal spores will take place. An example of just how effective this dispersal can be is in the study carried out by Craigie (1945) when fungal spores were located on the underside of an aircraft flying at 14 000 feet and the concentration of spores at 1000 feet was calculated at 1000
160

spores/m3. With this in mind it is not difficult to expect that spores of coprophilous fungi are able to reach the foliage of tall eucalypts, reaching heights of 40 to 50m (Cremer 1990).
Not only may air movement play a role in inoculation of foliage, but other vectors such as insects may be responsible for spore dispersal (Jennings & Lysek 1996). The outer surface of arthropods can be covered with fungal spores as they brush past the reproductive structures of a fungus. These animals may then climb the eucalypts, perhaps to eat the foliage, and in doing so inadvertently deposit these spores on the foliage. Finally, koalas may also ingest fungal spores with soil, which they are known to occasionally eat (Carrick 1997). It is hypothesized, therefore, that by whatever means, the koala is able to ingest a reasonable quantity of fungal spores.
The question then arises as to why these spores are unable to survive digestion. It is hypothesised that the 'classical' coprophilous species, although able to survive digestion, are not resistant to the effects of essential oils and tannins. The diet of the koala is based solely on foliage which contains a high level of these oils and also tannins which the koala is able to process by absorption into the blood and subsequent processing in the liver to compounds which are considered non-toxic (Phillips 1990). This process occurs before the site of microbial fermentation in the caecum and proximal colon thereby ensuring the microbiota on which the koala depends for symbiotic digestion of foliage is not eliminated. However, the effect of these oils on fungal spores may take place in the stomach before detoxification. If so, it would then be expected that no fungal species would occur in the faeces of the koala. However, it is known that just as some animals are able to detoxify toxins to render them harmless, some microorganisms are able to cope with high levels of tannins in unique ways (Scalbert 1991).
This is especially true for members of the Deuteromycotina (namely Aspergillus and Penicillium) which are often found growing on the surface of liquid tannery pits (Scalbert 1991 ). Many microorganisms secrete polymers which bind to tannins in order to render them unavailable for binding with other essential molecules such as microbial enzymes (Scalbert 1991). With this in mind it is possible to assume that members of the Deuteromycotina (11 out of the 13 species recorded) are able to survive both the process of digestion and the effect of tannins and be present in the voided faecal pellet of the koala and thus be able to grow on this substrate.
Although there is little mention of the Deuteromycotina in any of the literature dealing with coprophilous fungi, it does not mean that they are absent. In fact, they are often abundant on the dung of most animals and they are often neglected due to them not being considered 'classical' or 'true' coprophilous fungi (Seifert et al. 1983). However, it may be noted that several species of the Deuteromycotina are found exclusively on the dung of some herbivores. Furthermore, when dung is typically incubated for subsequent examination it is moistened thoroughly. However, members of the Deuteromycotina tend to be expressed on dung that is less than saturated ( eg. koala pellets) (Seifert et al. 1983). Since koala pellets are relatively dry, this may also be the reason for the longer than normal incubation period to achieve complete fungal expression. It may also be due to interspecific competition between fungal species occupying the one pellet.
Although the observations of fungal succession on koala dung are not what is normally recorded this does not render them insignificant. These differences that were observed can be explained due to the koala's diet being unlike that of 'typical' herbivores such as rabbits, sheep and cattle, all of which have been studied extensively. However, in contrast to other herbivore dung, which all show the same species on almost all faecal pellets when isolated from the same animal, that of koalas
161
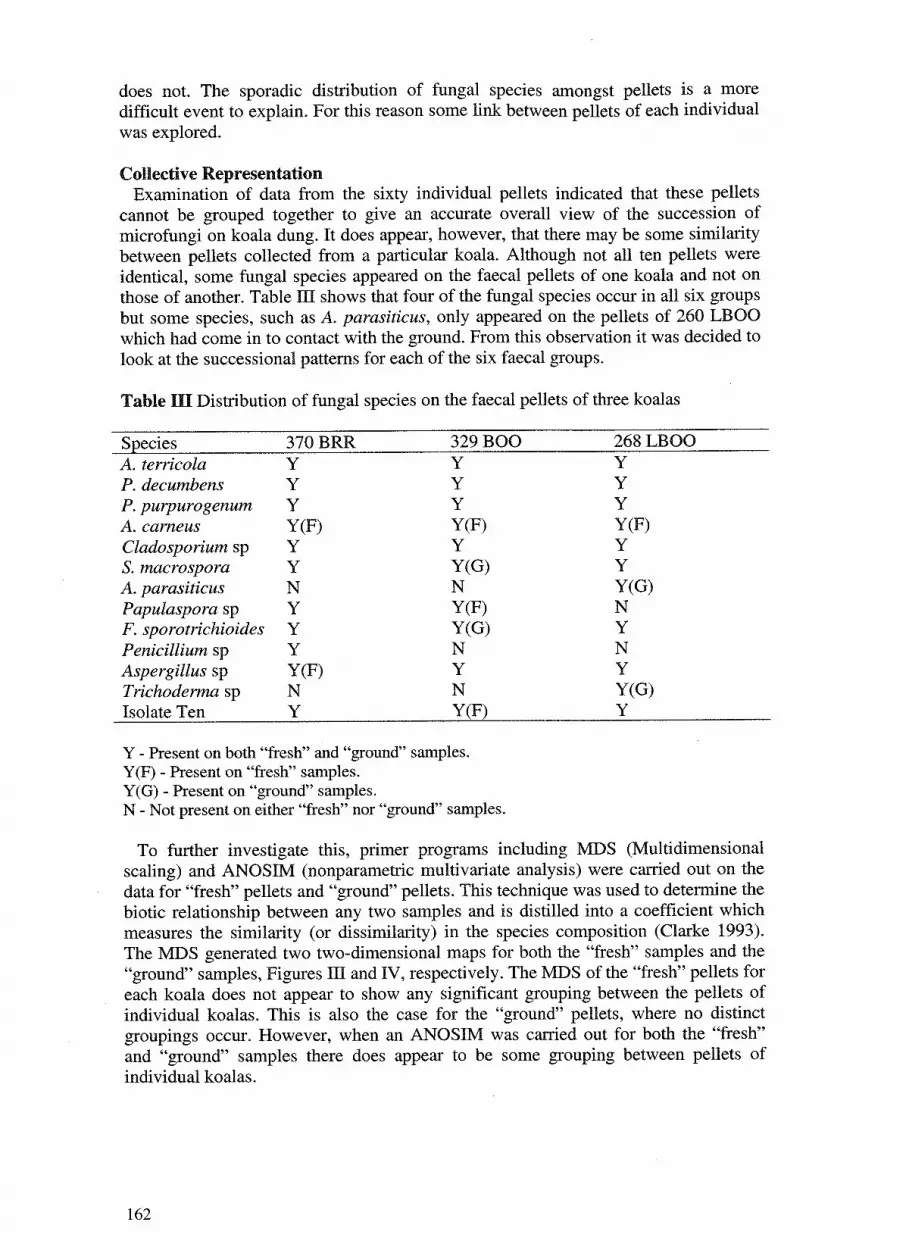
does not. The sporadic distribution of fungal species amongst pellets is a more difficult event to explain. For this reason some link between pellets of each individual was explored.
Collective Representation Examination of data from the sixty individual pellets indicated that these pellets
cannot be grouped together to give an accurate overall view of the succession of microfungi on koala dung. It does appear, however, that there may be some similarity between pellets collected from a particular koala. Although not all ten pellets were identical, some fungal species appeared on the faecal pellets of one koala and not on those of another. Table III shows that four of the fungal species occur in all six groups but some species, such as A. parasiticus, only appeared on the pellets of 260 LBOO which had come in to contact with the ground. From this observation it was decided to look at the successional patterns for each of the six faecal groups.
Table Ill Distribution of fungal species on the faecal pellets of three koalas
Species 370BRR 329BOO A. terricola y y
P. decumbens y y
P. purpurogenum y y
A. cameus Y(F) Y(F) Cladosporium sp y y
S. macrospora y Y(G) A. parasiticus N N Papulaspora sp y Y(F) F. sporotrichioides y Y(G) Penicillium sp y N Aspergillus sp Y(F) y
Trichoderma sp N N Isolate Ten y Y(F)
Y - Present on both "fresh" and "ground" samples. Y(F)- Present on "fresh" samples. Y(G)- Present on "ground" samples. N - Not present on either "fresh" nor "ground" samples.
268LBOO y y y Y(F) y y Y(G) N y N y Y(G) y
To further investigate this, primer programs including MDS (Multidimensional scaling) and ANOSIM (nonparametric multivariate analysis) were carried out on the data for "fresh" pellets and "ground" pellets. This technique was used to determine the biotic relationship between any two samples and is distilled into a coefficient which measures the similarity (or dissimilarity) in the species composition (Clarke 1993). The MDS generated two two-dimensional maps for both the "fresh" samples and the "ground" samples, Figures III and IV, respectively. The MDS of the "fresh" pellets for each koala does not appear to show any significant grouping between the pellets of individual koalas. This is also the case for the "ground" pellets, where no distinct groupings occur. However, when an ANOSIM was carried out for both the "fresh" and "ground" samples there does appear to be some grouping between pellets of individual koalas.
162
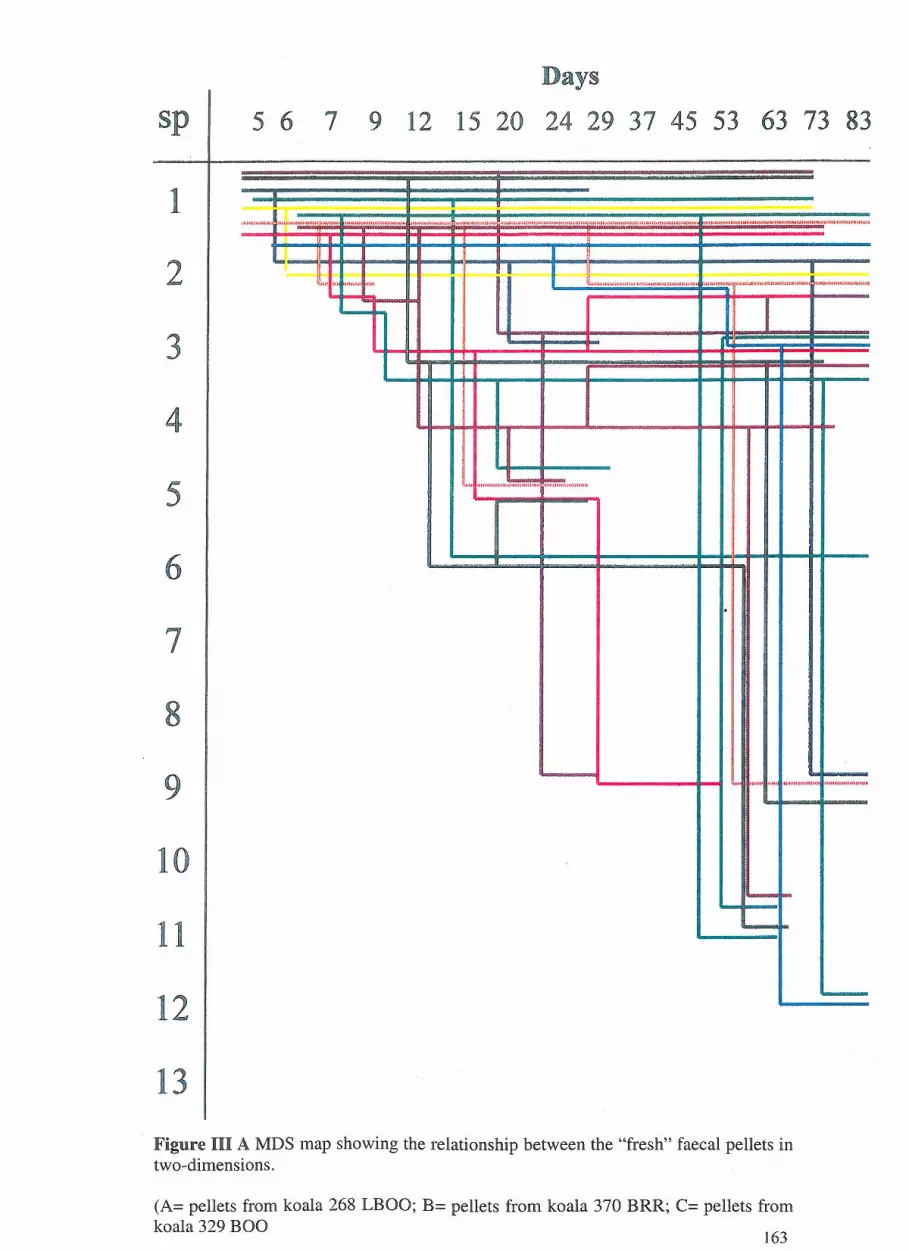
Days
sp s 6 7 9 12 15 20 24 29 37 45 53 63 73 83
1 ,. ... ,-~, i.:r:;·~"''''~"'" ··-~""'"'"'·"'~· ·~·""·~··~,.~ ..... ,,'"'"'"''" ~ ''"''''"""'~'·•·»~~···~·· ···· ''"'~'~"·~·····'''"' ''"""'"·"···· ··· ··~ ... ., ........................ ~-........................... .
"'
2 -· ·-
3
4
5
6
7
8
9
10
11
12
13
Figure III A MDS map showing the relationship between the "fresh" faecal pellets in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from koala 329 BOO
163
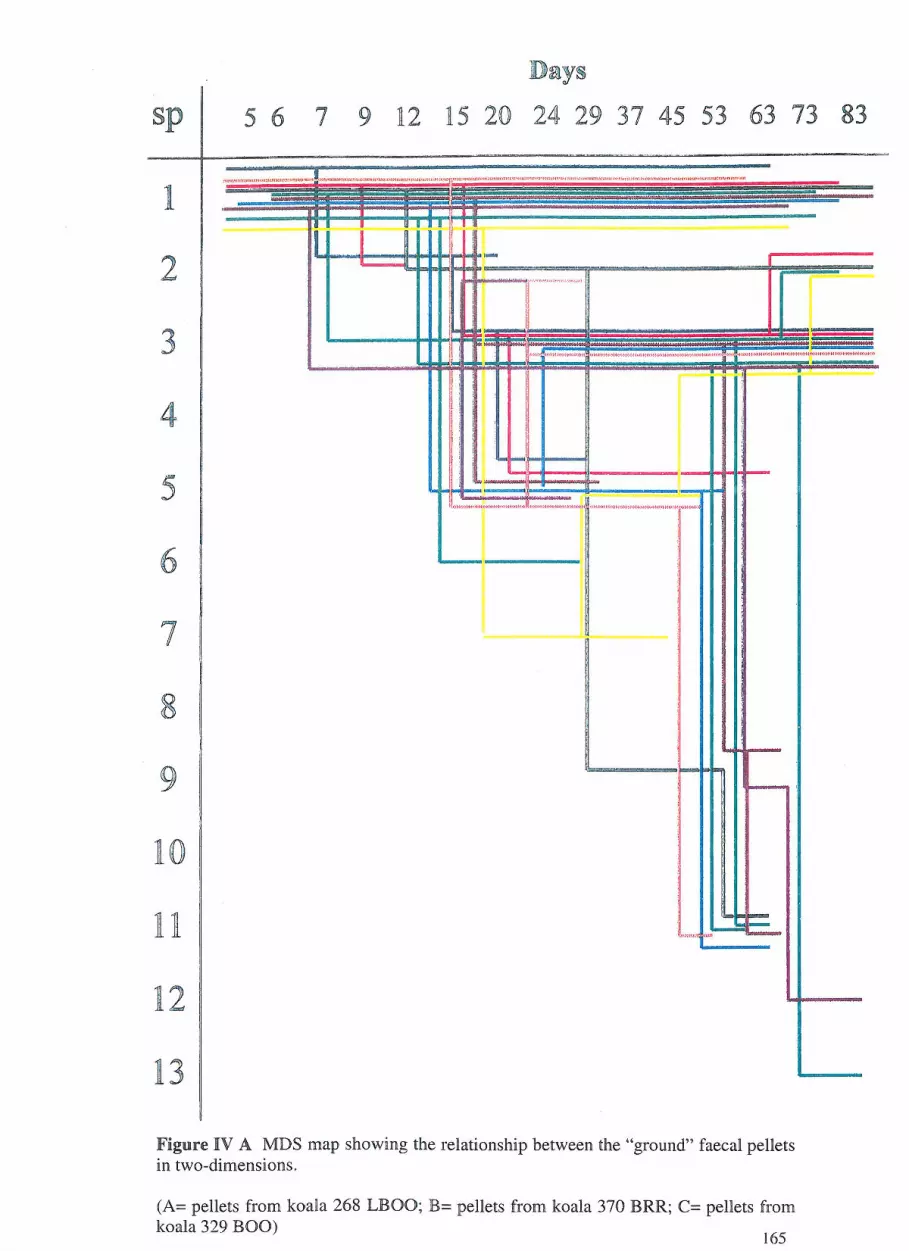
Day§
sp s 6 7 9 12 15 20 24 29 37 45 53 63 73 83
1
2 i ,:I
3 I I l
4
5 j :
6
7
8
10
11
12
13
Figure IV A MDS map showing the relationship between the "ground" faecal pellets in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from koala 329 BOO)
165

Day§
sp 5 6 7 9 12 15 20 24 29 37 45 53 63 73 83
1 I
2
3 r~~~ ~ " ~ l~<l' r '"'"""~ '"l!'ll'l•>.'llt•:mU : I ! - I !IIH•~!U 'U•!•"IIIU"tl :1"" "1\! 1 ~1~ 11 ~~. ~~l! l i ' IU ~'~· .... ,l\.11iliUHIII'I~il"!lt<J ~WIUUU"II'!I I Iti!I!!~U~'II"' ~J!II! II I
~ : I 4
5
6
7
8
9
10
11
12
13
Figure III B MDS map showing the relationship between the "fresh" faecal pellets in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from koala 329 BOO 167
.I
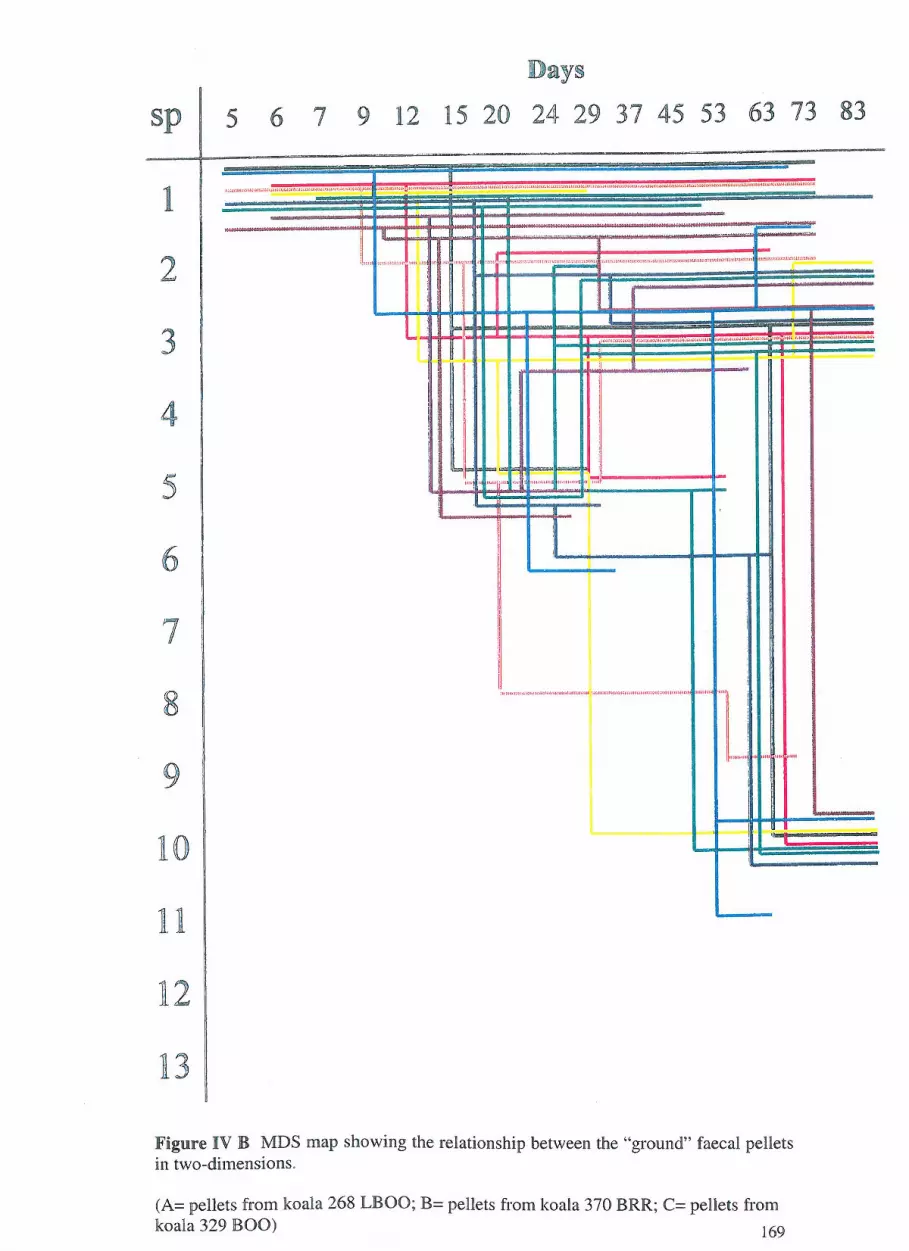
Day§
sp s 6 7 9 12 15 20 24 29 37 45 53 63 73 83
1 I
. II
2
3 ""' : ,_ 1." -
4 . I '
i •
5
6 I
7
\lrAi~OJI' Ill 11~'1' :•JI<, <~1114
' 9
10
1.1
12
13
Figure IV B MDS map showing the relationship between the "ground" faecal pellets
in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from
koala 329 BOO) 169

sp 5 6 7 9 12 15 20 24 29 37 45 53 63 73 83
. I.
1 , . I II ' . il l . '" § ,.,,.., ·' . - "'"'"'"·'" --=== i ~ ;- ' ;;- ~
2 l ;!
1': :•i • I• •• : •i• •i•l •l .•l •i• •i "• I• •i '• • •I ·; •\•' io •I i ;•i l•'· t :• I . ,, ' · rl
•'
:
3 : .JI
I! !
"= r ,_
4 I II
' I
5 ,.~~J 6
7
8
9
10
11 -
12
13
Figure III C MDS map showing the relationship between the "fresh" faecal pellets in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from koala 329 BOO 171
I •I ' ' I Jo \r
~
!-..
,, .
Day§
sp 5 6 7 9 12 15 20 24 29 37 45 53 63 73 83
1
2
3
4
5
6
7
9
10
11
12
13
Figure IV C MDS map showing the relationship between the "ground" faecal pellets in two-dimensions.
(A= pellets from koala 268 LBOO; B= pellets from koala 370 BRR; C= pellets from koala 329 BOO) 173

Statistical results for the "fresh" samples, as listed in Table IV, indicate that the pellets from each individual could be grouped into three distinct groups, with a clustering of pellets belonging to each animal. This was determined by examining the level of significance between the pellets of two koalas. If the significance is below 5 % then this means that the two sets of pellets are significantly different. When examining these results for the "ground" pellets, Table V, the ANOSIM shows that while the pellets from koala 370 BRR are significantly different from the pellets of both 268 LBOO and 329 BOO, the pellets of the later two koalas are not significantly different, even though these individuals were found feeding on different tree species. This result cannot be explained by gender, as one animal was female and the other was male. Furthermore, the koalas were found in different areas of Kroombit Tourist Park, and their pellets were collected on different days.
Table IV Statistical Results of ANOSIM of "fresh" pellets
Koalas Significance
268 LBOO - 370 BRR 1.4%
268 LBOO - 329 BOO 0.2%
370 BRR - 329 BOO 1.7%
There appears to be no general pattern of species composition over the entire incubation of sixty koala faecal pellets. However, the statistical data indicate otherwise. From this analysis it is possible to group the thirty "fresh" pellets into three distinct groups pertaining to the three individual koalas. For the "ground" pellets, however, it is only possible to group the pellets of one individual from the rest of the pellets.
Table V Statistical Results of ANOSIM of "ground" pellets
Koalas Significance
268 LBOO - 370 BRR 0.2%
268 LBOO - 329 BOO 6.7%
370 BRR - 329 BOO 0.1%
The results of this study indicate that the microfungi found on koala dung is unique to that observed from other herbivores. Many factors may have attributed to the patterns observed and further investigation is needed before any definite conclusions can be drawn about the differences seen. Future study should be directed toward answering the questions raised in this paper and also investigating the dung of other arboreal herbivores to develop an overview of the mycobiota of the dung of less 'typical' herbivores.
175

Acknowledgments Thanks to Robyn Willis for help with collection of field samples. Also thanks to
many people in the Faculty of Applied Science at Central Queensland University for their advice and assistance.
References Angel, K., and Wicklow, D. T. (1973). Microflora in energy flow schemes. Journal of
Ecology 62, 429-437. Anson, A. E., Fisher, P. J., and Kuthubutheen, A. J. (1985). Interactions between
Pilobolus crystallinus and Pseudomonas paucimobilis isolated from rabbit dung. Transactions of the British Mycological Society 85 (1), 161-164.
Bell, A. (1983). 'Dung Fungi: An illustrated guide to coprophilous fungi in New Zealand'. (Victoria University Press: Wellington.)
Bonner, R. D., and Fergus, C. L. (1959). The fungus flora of cattle feeds. Mycologia 51, 55- 86.
Carlile, M. J. (1965). The photobiology of fungi. Annual Review of Plant Physiology 16, 175 - 202.
Carmichael, J. W., Kendrick, W. B., Conners, I. L., and Sigler, L. (1980). 'Genera of Hyphomycetes' .(The University of Alberta Press: Edmonton.)
Carrick, F. N. (1997). Pers. Comm. Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community
structure. Australian Journal of Ecology 18, 117- 143. Cooke, R. C., and Rayner, A. D. (1984). 'Ecology of Saprotrophic Fungi'.! (Longman
Group Ltd: U.S.A.) Craigie, J. H. (1945). Epidemiology of stem rust in Western Canada. Scientific
Agriculture 25, 285-401. Cremer, K. W. (1990). 'Trees for Rural Australia'. (Inkata Press: Melbourne.) Dickinson, C. and Underhay, V. (1977). Growth of Fungi in Cattle Dung.
Transactionsofthe British Mycological Society 69(3), 47-477. Eberhard, I. H., McNamara, J., Pearse, R. J., and Southwell, I. A. (1975). Ingestion
and Excretion of Eucalyptus punctata D.C. and its Essential Oil by the Koala, Phascolarctos cinereus. Australian Journal of Zoology 23, 169-179.
Glockling, S. L., and Dick, M. W. (1994). Five new species of Rotiferophthora isolated from dung. Mycological Research 98(8), 833-844.
Gopal, K., and Lodha, B. C. (1982). Studies on coprophilous fungi VI. Chaetomium. Transactions of the British Mycological Society Japan 23, 259-271.
Gregory, P. H. (1966). Dispersal. In 'The fungi: An advanced treatise'. (Eds. G. C. Ainsworth and A. S. Sussman.) pp. 709-732. (Academic Press: New York.) Harper, J., and Webster, J. (1964). An experimental analysis of the coprophilous fungal succession. Transactions of the British Mycological Society 47(4), 511-530.
Harrower, K. M., and Nagy, L.A. (1979). Effects of nutrients and water stress on growth and sporulation of coprophilous fungi. Transactions of the British Mycological Society 72(3), 459-462.
176

Jennings, D. H., and Lysek, G. (1996). 'Fungal Biology: Understanding the fungal lifestyle'. (BIOS Scientific Publishers: Oxford.)
Khan, R. S., and Krug, J. C. (1989). New records of the Sordariaceae from East Africa. Mycologia 81(6), 862-869.
Lodha, B. C. (1974). Decomposition of digested litter. In 'Biology of Plant Litter Decomposition'. (Eds. I. Dickinson and G. Pugh.) pp. 231-241. (Academic Press: New York.)
Mahoney, C. J., and Strongman, D. B. (1994). Nematophagous fungi from cattle manure in four states of decomposition at three sites in Nova Scotia, Canada. Mycologia 86(3), 371-375.
Parker, A. (1979). Associations between coprophilous ascomycetes and fecal substrates in Illinois. Mycologia 71, 1206-1214.
Phillips, B. (1990). 'Koalas: The little Australians we'd hate to lose'. (Australian Government Publishing Service: Canberra.)
Pitt, J. I. (1979). 'The Genus Penicillium and its teleomorphic states Eupenicillium and Talaromyces'. (Academic Press: London.)
Pitt, J. I., and Hocking, A. D. (1985). 'Fungi and Food Spoilage'. (Academic Press: Sydney.)
Raper, K. B., Fennell, D. I., and Austwick, P. K. C. (1965). 'The Genus Aspergillus'. (Robert E. Krieger Publishing Company: Florida.)
Richardson, M. J. (1972). Coprophilous Ascomycetes on different dung types. Transactions of the British Mycological Society 58(1), 39-48.
Rifai, M.A. (1969). A revision of the genus Trichodenna. Mycological Papers 116, 1-56.
Scalbert, A. (1991). Antimicrobial Properties of Tannins. Phytochemistry 30(12), 3875 -3883. Seifert, K. A., Kendrick, B., and Murase, G. (1983). A key to Hyphomycetes on dung. (Department of Biology, University of Waterloo: Cananda.)
Webster, J. (1970). Coprophilous fungi. Transactions of the British Mycological Society 54(2), 161-180.
Wicklow, D. T. (1981). The coprophilous fungal community: a mycological system for examining ecological ideas. In 'The Fungal Community: Its organisation and role in the ecosystem'. (Eds. D. T. Wicklow and G. C. Carroll.) pp.47-75. (Marcel Dekker; New York.)
177

All creatures are equal, though some are more equal than others: the moral and political agency of koalas
Robert Kelso Lecturer in Philosophy, Central Queensland University, Rockhampton, Qld.
Introduction Koalas occupy a unique place in the national psyche. The dominant image is one of
warm, furry 'teddy bear' creatures that spend much of their lives quietly browsing in the tree-tops harming no-one. For many international tourists, koalas, along with kangaroos, present the quintessential image of Australia. From one perspective, "they are our greatest ambassadors" (AKF Newsletter 1996-1997). Whether those images are accurate or not does not seem to be a critical issue for those tourist promoters and others with a direct economic relationship to koalas, but they do raise problems for others. Those charged with the responsibility for ensuring the well-being of koalas and their habitat are now subjected to considerable scrutiny, their activities present unique problems and require justification to governments, scientific and public audiences. The nature of justification extends beyond scientific and technical reasoning.
The first part of this paper addresses some of those problems from philosophical perspectives which attempt to answer the questions of whether animals (and thus koalas) have moral agency and rights, and if so, how we should act towards them. The second part of the paper links those moral concerns to practical-ethical questions such as whether koalas should be relocated or culled; and whether habitat protection is of equal importance to koala protection. The final part of the paper considers some ethical, political and economic issues which have made koalas such a prominent part of the contemporary Australian social landscape.
Aldridge (1998) argues that the public, and probably most scientists, would not be familiar with all of the arguments which represent the diverse views about the moral agency of animals. The classical humanist approach is to distinguish between animals and humans by virtue of human rationality and human intelligence, and this has given rise to two mutually exclusive models of moral reasoning. The first model is deontological, an idealised or universal set of moral understandings which are available to all rational beings. A succinct deontological critique of cruelty to animals might rely on nothing more than to say that it is wrong. The second model is teleological, for example, utilitarian, and would argue that cruelty is wrong because it creates pain and suffering. However, short term pain and suffering may be necessary for long term happiness, in which case cruelty may be a relative term. By comparison to the classical humanist approaches which privilege humankind, the biocentric approach accords all living things equal value. Thus, a deformed human foetus may be of no lesser value than a healthy animal. The biocentric approach often underpins radical 'green' politics and some animal liberation arguments. Given the diversity of views, a compelling case can be put that there is presently no general community consensus about the issues involved (Rose 1989). But, regardless of the lack of consensus about the moral status of animals in general, there does appear to be moral concern about koalas, and, to a lesser extent, their habitats.
The moral argument pursued in this paper adopts the second of three distinct orientations as described by Garner (1993). The first is that animals have no moral
178

status, this being the exclusive right of human beings. The second argument is described as the current orthodox view from which animals, as sentient beings, have some moral status but are inferior to humans. The third argument or position, put by writers such as Regan and Singer and briefly outlined below, challenges the first two in proposing that all animals (including humans) have equal standing.
The moral agency of animals The late 20th century recognition of environmental and animal rights may appear to
be novel, even embarrassing (Midgley 1989), however, it has a long history in Western philosophy (Regan 1975; Sorabji 1993). Humankind's relationship to animals was redefined by the rise of humanism, particularly in the post-Aristotelian debate and subsequent Christian tradition (Linzey 1989; Sorabji 1993) and most notably in the 'speciesism' evident in humans' preference for other humans (Ryder 1975). Singer (1975 p.185), presents the outcome as, " ... the rule of the human animal over other animals". Those now arguing for the equal standing of animals with humans are asking us to identify the roots of the problem of animal rights and to recognise that:
While-ever we continue to perceive the world from a standpoint which places humans at the centre of creation and relegates other forms of life to a secondary position, according to this kind of argument, we will simply perpetuate the attitudes to nature which have led to our past mistakes. What is required, it is often suggested, is a transformation of the anthropocentric perspective in our culture to one which gives equal significance to all forms of life in the workings of our world (Wells 1993 p.515).
Singer has argued that recognition of animal rights will require a shift towards the, " ... fundamental moral principles of equality of consideration of interests that ought to govern our relations with all beings" (Singer 1975 p.213), and this clearly includes animals. As Darwin made explicit in The Origin of the Species, man is merely an animal. But, how can this problem be constructed as a moral problem rather than simply as an ethical disagreement? Regan (1983 p.240) identifies a fundamental issue when he argues that:
Morality will not tolerate the use of double standards when cases are relevantly similar. If we postulate inherent value in the case of moral agents and recognise the need to view their possession of it as being equal, then we will be rationally obliged to do the same in the case of moral patients. All who have inherent value thus have it equally, whether they be moral agents or moral patients. All animals are equal, when the notions of 'animal' and 'equality' are properly understood, 'animal' referring to all (terrestial, at least) moral agents and patients, and 'equality' referring to their equal possession of inherent value.
The distinction between morality and ethics Many individuals and most governments do not accept the moral position which
accords equal status to all sentient beings, including animals and humans. This position becomes even more difficult when attempts are made to reconcile, "the individualistic nature of moral rights with the more holistic view of nature" and the environs in which the animals live (Regan 1975 p.361). The most commonly accepted contemporary approach on animal rights is relativist in that it recognises that animals have limited rights as moral patients (reactive),
179

rather than moral agents (active and autonomous). Given the influence and regulation of professional bodies and governments, researchers and scientists must justify use (or abuse) of an animal even though that use may be based on ethically acceptable clinical practice. There is now a strong body of public opinion which accepts that animals have rights (Garner 1993) and that arguments for research involving animals be rigorously evaluated (Balls and Sou thee 1989 p.111 ).
The argument for humans having pre-eminence over animals is based on the concept of autonomy which:
requires a being to be self-governing; to have beliefs and preferences and to be able to act according to them, changing behaviour when the evidence suggests that beliefs can no longer be supported. Such an autonomous being - with thoughts, intentions and beliefs - can be held morally responsible for his actions and can recognise when a wrong is inflicted on him (Gamer 1993 p.13).
For the proponents of animal rights (Singer, Regan), the reliance on moral autonomy as the critical distinguishing feature between humans and other animals is simply an example of anthropocentric bias. The biocentric view does not include that humans be treated differently to animals and thus, " ... reduces people from social beings to a simple species - to zoological entities that are interchangeable with bears, bisons, deer, or, for that matter, fruit flies and microbes" (Bookchin 1988 p.18). If we were to take a biocentric view to its extreme though logical ethical consequence, a chlamydia cell, a koala, the trees they eat, or, for that matter, a human being are of equal value. However, there are winners and losers in the competition for survival, and humans, as the dominant animal, are uniquely placed to decide who the winners and losers are. Not all humans get a say. In our contemporary society it is the politicians, scientists and technical experts, not moral philosophers, to whom we turn for answers, but there is no guarantee of consistency.
Rose (1989 p.126) has recognised that, "Animal research is an activity of science, and any consideration relating especially to its regulation should take into account the nature and conduct of the scientific process". Two eminent philosophers of science addressed this 'problem' from different perspectives. They both recognised the need for individuals to engage in significant introspection and debate about what they are doing. Sir Karl Popper (1959 p.l11) argued that "The empirical basis of objective science has nothing absolute about it. Science does not rest upon solid bedrock. The bold structure of its theories rises, as it were above a swamp. It is like a building erected on piles". In an equally devastating attack upon scientific 'objectivity', In The Structure of Scientific Revolutions, Thomas Kuhn (1962) argued that in periods of what he called "normal science", scientists are deeply committed to established theories and methods. This view fundamentally challenged the rationality of science and suggested that scientists are committed to their theories and explanations even, at times, to the point of being closed-minded. Established paradigms or theories, ideas and methods remain 'true' only until a rival paradigm attracts more adherents within the scientific community. The implications flowing from both views is that absolute empirical truths are rare, if not impossible, and that some aspects of scientific practice are not so much objective, value-free inquiry, as equivalent to political action.
Ethical reasoning is an activity grounded in rational, moral and political theorising; it is a uniquely human activity. Ethical reasoning is an element of the anthropocentric view of the natural world, one in which everything exists in terms of human experience and values, thus the existence of humankind is considered to be the central fact of the universe. It could therefore be argued that ethical reasoning about the natural environment and the living things within it, presupposes a biased view.
180

From such a standpoint it would be impossible for humans to make decisions about other living things on any basis other than human interests and understanding.
Prior to regulation by government and public opinion, scientists, researchers, farmers, public sector managers and other interested professionals have been able to make decisions about koalas and their habitats without undue scrutiny. That situation has changed; there is now intense scrutiny of matters to do with science, research and the application of technologies (New Scientist 1998 p.l). Special interest groups now 'speak on behalf of the environment, koalas, and their interests; their attitudes are illustrated in the following ways. "Humans created the situation ... We blame them [koalas]. We discuss shooting them. Who is at fault here?'' (The Australian Koala Foundation Newsletter Dec 1996/Jan 1997 p.S). Thus, the ethics of human-koala interaction are firmly on the public agenda and any person working in the area needs to be aware of the major issues and be able to respond appropriately; but, we must recognise that whatever ethical approach we take it cannot be anything other than human-centred.
The modernist predilection for socially constructed knowledge has meant that the distinction between morality and ethics is not universally accepted. Morality has adeontological character with reference to universal values whereas ethics need be no more than an acceptance of customary practice. Therefore, it can be noted that while some activities (such as culling animals) are seen as ethical they can just as easily be constructed by others as 'immoral'. Furthermore, it has been suggested by Rose (1989 p.125) that, "To avoid consideration of the complexity and difficulties of the moral issues involved would ... be morally irresponsible". Most scientists and field workers, like other professionals, are inclined towards the kinds of values and practices which their training has instilled in them.
It could be considered a 'natural' reaction for those scientists and field workers who are in immediate contact with koalas to develop a detailed understanding, even sympathy for them. Some may adopt an advocacy role in relation to them. But not all scientists and field workers would agree on the solutions to problems confronting koalas. It is also likely that the disciplinary base from which they work will provide the dominant values upon which opinions are formed and decisions are made. Some of those values and opinions may be influenced by the length of time and the amount of contact those workers will have had with koalas and with other members of their profession. To caricature different approaches, field workers may tend towards the view that nature should take its course while laboratory based workers may be more inclined to accept and/or propose technical intervention. Those workers with a primary responsibility for the physical environment may place a higher value on the quality of trees and other vegetation and diminish the value of the koalas that feed on them. The different values placed on each of the objects in the equation would require different answers to ethical questions such as; "Should we relocate koalas from highdensity population areas?" It may be more humane to simply kill enough koalas to relieve the grazing pressure on the food trees as relocation creates as many problems as it solves. In this case, a qualified veterinarian could be on hand to euthanaise dying animals to reduce their inevitable suffering. This approach would certainly save trees from destruction. Alternatively, we could allow nature to take its course (as we do with other wild animal populations) and wait for starvation to reduce fertility and increase morbidity.
However, if all animals are equal, then similar issues are faced when dealing with animals or humans; if humans are starving and something can be done about it, something usually is. Thus, if we are to follow the radical animal rights philosophy to its logical conclusion then we could never entertain any possible moral justification
181

for 'culling', that is, deliberately killing, koalas. Likewise, the grounds for the decision to euthanaise a koala would be applied equally in the case of a human 'suffering' a similar level of starvation, venerial infection, loss of reproductive capacity and 'enjoyment of life'. Interestingly, human euthanasia is practiced in particular circumstances, where disease is considered to have eroded the quality of life to such an extent that recovery is not possible and quality of life cannot be regained. While human euthanasia is accepted in limited cases it is more widely accepted for animals.
The following example illustrates how different ethical positions affect outcomes and produce judgments. One of the final sentences of a veterinary report described how, "The decision to euthanaise the young female koala was based on ethical grounds due to the severity of the clinical signs, and also upon her inability even if treated, to regain reproductive function". The koala had been examined on capture with obvious signs of disease (Chlamydia) noted, removed to a clinic for assessment, " ... and was subsequently euthanaised". The report and its contents would have been unremarkeable if it were about domestic cats or ringtail possums, but the decision to euthanaise a koala created difficulties for the attention which it might attract from environmental interest groups who were active in the area. The scientist in question was sufficiently aware of the potential difficulties flowing from the decision and so the report was sent to another colleague for comment. If it had occurred in an isolated non-urban area then the same political pressures would not have been as pressing.
What kinds of ethical arguments might be put forward which would justify or explain the decision to euthanaise? Given the nature of the report and the author's request to ask for collegial evaluation, it could be concluded that the decision to euthanaise was based on a number of factors including: the available scientific evidence; professional judgment formed over many years of contact with koalas; and due consideration of the political nature and public concern of activities logically associated with such work. Support for the decision to euthanaise can also be found in the National Health and Medical Research Committee Animal Welfare Committee Guidelines (1994). The decision can clearly be seen as an example of utilitarian ethics in which the quality of pleasure is directly balanced against the quantity of pain; at some point a quick death is preferable to a painful existence.
A veterinarian may also apply the kinds of ethics which we normally associate with human health care workers. Thus, the veterinarian as a health care provider may not distinguish between a human and an animal's right to life, relief from pain or unnecessary surgical procedure and may even act as advocate and agent on the animal's behalf (Porter 1989). Gamer (1993) makes the point that the adoption of a 'scientific' approach may elicit a more objective, dispassionate reaction. The shift in values from 'vet' to veterinary scientist may be subtle but profound. Returning the koala to the wild may have resulted in a slow painful death, or the subsequent infection of other koalas, but it would have restored the 'natural balance' as nature took its course. However, vicarious suffering would also ensue from humans who may be aware of the koala's precarious state, so it could be argued that the greatest happiness for all concerned was to euthanaise the koala. But how much consideration should we give to the human's discomfort ?
It is impossible for humans to communicate with koalas or consider their interests in the same manner in which they communicate and interact with other humans. Therefore, decisions made by humans about koalas and their habitats cannot be made on any basis other than human interests and understanding; such anthropocentric views of the natural world and of other sensate entities, are central to our consciousness. A moral equivalency argument would claim that chlamydia is a living
182

entity and deserving of equal status to the koala. Chlamydia kills koalas just as koalas kill trees. On Kangaroo Island, koalas too can be seen as a destructive feral organism. However, an anthropocentric-scientific explanation would assume a hierarchy based on the grounds that a koala, having certain similarities with, and being an object of interest for, human beings, is considerably more valuable than chlamydia, and therefore has a higher claim to existence. The value of the tree presumably falls somewhere in between. If the scientist responsible for the decision to euthanaise the koala had been working
in a scientifically literate society, or a society which had not experienced some of the diverse effects of green politics, then the decision to euthanaise would not have attracted a great deal of attention. However, scientists don't work in a social and political vacuum, and koala research cannot be simply defended as the disinterested pursuit of knowledge. Koalas have become 'big business', they have also been reified. The dual effects of reification by animalist, green, and environmentalist movements, and valorisation by big business, has created competing, possibly mutually exclusive paradigms of value, function, use and management. As one koala advocate puts it, "Where there is a profit motive, there will always be corruption and I have been around the Koala world long enough to know that people are capable of creating this sort of furore to position themselves to look like white knights in shining armour coming along to save these poor creatures" (The AKF Newsletter Dec 1996/Jan 1997p.6). These pressures need to be resolved but there is no guarantee that the outcome will be acceptable to all interested parties.
An anthropocentric approach to science has allowed, possibly encouraged, the characteristic human drive to control and manipulate all the 'things' in the natural environment to the humans' best interests. Fortunately for koalas, they are of considerable interest to humans and therefore attract much 'research' attention. When human intervention manifests as both scientific and technological activity, it is notionally based on objective facts, even though it still appears to be subject to all of the usual human vagaries of political opportunism. The following statement illustrates the point:
The AKF is so disillusioned by the way that government has called science in, ostensibly to solve the problems and then once its political objectives have been met, science has been distorted, discredited, discarded and forgotten. Science can answer the Kangaroo Island dilemma as it can the problems of overpopulation in certain localised parts of Victoria, as long as political interference doesn't get in the way (The Australian Koala Foundation Newsletter Dec 1996/Jan 1997 Editorial, p.2).
However, the politics of koala research are not always clear-cut. Although quality science is the best ally of endangered koala populations and their habitats, the possible misuse or manipulation of environmental or population characteristics in the long term detriment remains.
183

Koala politics In the past 60 years the koala has enjoyed a steady transition from economic
resource to internationally recognised cultural icon. Management practices employed up to the 1930s (hunting to the point of eradication) would now be abhorrent to the general population. However, at the same time that the koala was gaining increased protection its habitat came under increased threat. Modernisation and progress in the form of land clearing and urbanisation along the east coast has reduced the easily accessible koala habitats to small isolated pockets. A generation ago, children lined up at zoos for the obligatory koala-cuddling snap-shot. Nowadays, many thousands of visitors expect to be able to handle koalas; besides the threat of cars and dogs, there is a now real possibility of some koalas being literally cuddled to death.
The nature of popular press reporting of the politics surrounding koalas and their habitats is both irrational and anti-scientific, and frequently based on 'naturalist' aesthetics. The emergence of 'green' politics, in tandem with a range of 'environmental movements', has resulted in the social (re)construction of particular koala habitats as "sacred sites". The political prominence afforded to koala habitat can now be appropriated for other purposes as the following example demonstrates:
Up in Nullum State Forest, back of Mullumbimby, a tallowwood comes down and the green group descends. Finding koala droppings at its base, a cry goes up. "We've got to have an army here," says someone. "Get the network happening. We were too late. We could have saved this tree from being dropped." ... The protest began ovemight, but attracted no-one with "the [feral] look" that day. A few tumed up in the next few days, and I don't know if it was because the phone tree takes a bit longer to reach them or whether they were still recovering from the doof which finished only the day before. But the greens won anyway. The minister put a halt to logging in the compartment before any pictures of hairy folk sitting on tripods hit the city television screens (Whittaker 1996 p.50).
The emergence of 'green politics' in Australia started most noticeably with the battles over Lake Pedder and the Franklin River in Tasmania. Over the past twenty years the mainstream political parties have reacted to environmentalist demands with varying levels of commitment. At best the major parties saw the environment as just another element to be juggled and placed in the already crowded political landscape.
The Australian Democrats identified the environment as a central issue in their policy framework, but with the inevitable compromises necessary for economically sustainable growth, have failed to satisfy some of the more committed greens' demands. Independent 'Green' senators (Brown, Tasmania and Chamarette, W.A) held the balance of power in the parliament until the 1996 federal election in which the Democrats and Green parties split the environmentalist vote. This trend followed the pattern set in previous state and federal elections. In the July 1996 Queensland election the Labor Party lost six metropolitan seats in which the preservation of koala habitat was a major issue. The green movement has been credited with the Labor Party's loss of up to six seats in metropolitan Brisbane in the 1995 Queensland election (Young Inquiry Report). But it is doubtful whether the electors who sent a protest vote to the Goss government were making judgments from some clearly defined ethical or political position; it was more than likely a 'NIMBY' (Not In My Back Yard) reaction to the construction of the South Coast Motorway which was described in The ALP Queensland Branch Committee of Inquiry into the 1995 State Election Result, (Young Inquiry 1995 p.9) as " ... the most talked about issue arising
184

out of the election". Ironically, it is those same members of the voting public who the koala population can rely on for support against developers and, potentially, scientists.
The shift to 'green' or 'naturalist' values will not necessarily prove too difficult for politicians, they will simply count the potential votes and act accordingly. Nor would the shift present too many problems for ethicists as aesthetics is an allied discipline and anyone arguing from an aesthetic position should be able to understand how political, economic and scientific understandings are constructed. The shift will, however, complicate the work of scientists and others, as it fundamentally challenges aspects of their worldviews in ways described briefly above by Kuhn and Popper. If that shift takes on the more extreme postmodem rhetoric of nihilism then rational science will increasingly come to be seen by a significant portion of the population as 'irrational'. Perhaps a return to 'common sense' would be the best solution? The recurring problem that it runs into, however, is that;
'anthropocentrism'- human-centred values and understandings - is really so basic to our culture and so destructive of the environment, and if it must be replaced with a biocentric or ecocentric ethos in order to create the conditions which will allow us to save the world from ecological devastation, then we need to go far beyond the traditional politics of human welfare for solutions to our dilemmas (Wells 1993 p.517).
Allied to that recurring problem is that scientists and other technologists expect or require rational informed consent as a precondition to public practice. The precondition for this is that kind of rationality peculiar to scientific reasoning, which is predicated upon,
the possibility that human beings can regulate their beliefs by rational self-reflection, that they can free themselves from interest, passion, tradition, prejudice and autonomously "rule" their own thoughts, and that they can determine their actions as a result of self-reflection and rational evaluation, an evaluation the conclusions of which ought bind any rational agent (Pippin 1991 p.l3).
The radical greens and biocentrists would, of course, disagree. What is scientifically or ethically rational,
" ... can today, in the age of a wide-spread defeatism with regards to reason, no longer be taken seriously. Whoever mentions "rationality" provokes normative dissent" (Schnadelbach 1990 p.273).
We presume that rational thought underpins the actions of scientists or field workers when they intervene (interfere?) in the existence of koalas and all the other organisms in their environs (such as chlamydia). Thus, on the above arguments a number of possible stances can be adopted. The solutions follow the moral positions outlined earlier in the paper. For those who recognise koalas, and other animals, as moral agents deserving of equal respect, then our higher order human rationality places considerable responsibility upon us to act in their best interests. The logical consequence of this position is to not harm or exploit them, or any other animal for that matter, in any way. The obvious option is to become vegetarian. An allied argument is presented by the radical biocentrists; for them, the only ethically acceptable course of action is to not interfere, 'let nature take its course'. The middle course was to accord animals limited moral status in a world dominated by human interests. Where this can be justified as part of a wider scientific world-view in which there is no morality independent of human ethics then scientists will have to accept
185

constant interference and/or participation in their activities from politicians, environmentalists, the business sector and other interested parties. If nothing else, this should make their professional lives interesting indeed.
Moral Obligations and the Scientist Moral respect for koalas requires active human intervention on behalf of the diverse
koala populations and their environs. This would be justified from the orthodox moral position in which humans recognise their superior position to other animals, but also that this position brings responsibilities. While this does not accept the strict equality of moral agents it may be beneficial in providing greater protection for moral patients - that is, a greater duty of care may be exercised when koala needs and habitat protection is argued for. The fallacy that a 'natural' outcome is morally superior would be seen as moral cowardice - merely as an excuse to do nothing. Given humans' greater rational powers and their complicity in endangering koalas and their habitat, it is incumbent upon them that intervention be positive and clearly beneficial. Therefore scientists have a moral duty to ensure that the community has access to factual information about the distribution, conservation and management of koalas at national, State and local levels.
There is a clear need for scientists to argue for direct intervention as management based on the best available evidence about koalas and their habitats. However, this will require resolution of a number of issues including, culling and euthanasia, fertility control and translocation. Intervention raises significant ethical and moral problems, but sitting back and letting nature take its course is not acceptable compromise. The reality is that human intervention, regardless of the motives, is disruptive and potentially dangerous to the wellbeing of the koalas. Perhaps the only solution is to ensure that whatever harm occurs is minimised.
References Australian Koala Foundation Newsletter, December 1996-January 1997. Attfield, R. and Dell, K. (Eds.) (1996). 'Values, Conflict and the Environment'.
(A verbury: Aldershot.) Balls, M., and Southee, J. (1989). Reducing Animal Experiments By Reducing Their
Necessity. In 'The Status of Animals: Ethics, Education and Welfare'. (Eds. D. Paterson and M. Palmer.) (Wheatons: Oxon.)
Bookchin, M. (1988). Social Ecology Versus Deep Ecology. Socialist Review 18(3),9-29.
Garner, R. (1993). 'Animals, Politics and Morality'. (Manchester University Press: Manchester.).
Kuhn, T. (1962). 'The Structure of Scientific Revolutions'. (Chicago University Press: Chicago.)
Linzey, A. (1989). Reverence, Responsibility and Rights. In 'The Status of Animals: Ethics, Education and Welfare'. (Eds. Paterson, D. and Palmer, M.) (Wheatons: Oxon.)
Midgley, M. (1989). Practical Solutions. In 'The Status of Animals: Ethics, Education and Welfare'. (Eds. Paterson, D. and Palmer, M.) (Wheatons: Oxon.)
New Scientist. (1998). Ethical Dilemmas. Inside Science No.114, New Scientist October, 1-4.
Pippin, R. (1991). 'Modernism as a Philosophical Problem'. (Blackwell: Oxford.) Popper, K (1959). 'The Logic of Scientific Discovery'. (Hutchinson: London.)
186

Regan, T. (1983). 'The Case For Animal Rights'. (UCLA Press: Los Angeles.) Rose, M. (1989). Regulation of Animal Research: The Australian Experience. In 'The
Status of Animals: Ethics, Education and Welfare'. (Eds. D. Paterson and M. Palmer) (Wheatons: Oxon.)
Ryder, R. (1975). 'Victims of Science: The Use of Animals in Research'. (DavisPoynter: London.)
Schnadelbach, H. (1990). Remarks about Rationality and Language. In 'The Communicative Ethics Controversy'. (Eds. Benhabib, S. and Dallmayr, F.) (MIT Press: Cambridge.)
Sorabji, R. (1993). 'Animal Minds and Human Morals'. (Cornell University Press: New York.)
Wells, D. (1993). Green Politics and Environmental Ethics: A Defence of Human
Welfare Ecology. Australian Journal of Political Science, 28, 515-527. Whittaker, M (1996). Just don't call us Ferals. The Australian Magazine, Weekend
Australian, February 17-18, p.50. Young Inquiry. (1995). 'The ALP Queensland Branch Committee of Inquiry into the
1995 State Election Result', October 1995, Chairman, Mick Young.
187
