the photosystem stoichiometry in thylakoids of some australian shade-adapted plant species
TRANSCRIPT

Atist. J. Plant Physiol., 1990, 17, 665-74
The Photosystem Stoichiometry in Thylakoids of Some Australian Shade-adapted Plant Species
W. S. chowA, Jan M. ~ n d e r s o n A and A. el is^ A Division of Plant Industry, CSIRO, GPO Box 1600, Canberra, ACT 2601, Australia.
Department of Plant Biology, University of California, Berkeley, CA 94720, USA.
Abstract
The concentrations of functional photosystem I1 (PSII) reaction centres in leaves and photosystem I reaction centres (P700) in thylakoids isolated from comparable leaves of Australian shade-adapted plant species of diverse taxa, life-forms and habitats were compared. The concentrations of PSII were determined directly in leaves by the oxygen yield per single-turnover flash in the presence of far-red background illumination. The concentrations of P700 were determined by the light-induced absorbance change of thylakoid membranes at 703 nm. On a chlorophyll basis, the amounts of both functional PSII and P700 were lower in shade species than in sun species. The PSII/PSI reaction centre stoichi- ometries of the shade species ranged from 1.2 to 1 .9 indicating that (i) shade-adapted species do not have a fixed 1: 1 ratio; and (ii) their PSWPSI ratios are usually lower than those of sun species (1.7-1.8). We conclude that shade plants display variable photosystem stoichiometry. The results are discussed in terms of the interplay between the adjustment of photosystem stoichiometry and that of the light-harvesting chlorophyll antenna size of each photosystem in the thylakoid membrane of shade species.
Introduction
The light environment (both light quantity and quality) has a profound influence on the composition, function and structure of the photosynthetic apparatus (Lichtenthaler and Meier 1984; Anderson and Osmond 1987). Plants growing in the deep shade on the floor of a Queensland rain forest receive less than 0.5% of the photosynthetically active radiation incident on the canopy. Their light environment comprises very low intensity diffuse light (-60%) which is greatly enriched in far-red light, and direct intense sunflecks (-40%) of duration from 15 s to 5 min (Bjorkman and Ludlow 1972). Deep shade is therefore characterised by low levels of PAR and by a marked increase in the proportion of far-red light which is absorbed selectively by photosystem I (PSI)*.
*Abbreviations used: CPa, core PSII complex; CPla and CPl , PSI complexes, with CPla containing more of the PSI light-harvesting complex than CPl ; DCMU, 3-(3,4-dicholorpheny1)-1,l-dimethylurea (diuron); LCHII, light-harvesting complex of PSII; MES, 2-(N-morpholino)ethanesulfonic acid; PSI, 11, photosystem I , 11; P700, photosystem I reaction centre chlorophyll; SDS, sodium dodecyl sulfate; TES, N-tris(hydroxymethyl)methyl-2-amino ethanesulfonic acid; Tris, tris(hydroxymethy1) aminomethane.

W. S. Chow et a1
The well-established lower Chl a/Chl b ratios and higher xanthophyll/p-carotene ratios of shade plants reflect variations in the complement of their pigment-protein complexes (Lichtenthaler and Meier 1984). Shade species contain greater amounts of the Chl a/b- proteins of PSII and PSI, and relatively lesser amounts of the Chl a-proteins of the core of both photosystems relative to sun species (see Anderson et al. 1988). These variations in pigment-protein complexes in the chloroplasts of shade and sun species imply also differ- ences both in the amounts of P680 and P700, as well as in the light-harvesting antennae of each photosystem. Indeed, a striking feature of the sun/shade response is a marked modulation in the photosystem reaction centre stoichiometry (Melis and Brown 1980; Wild et al. 1986; Anderson and Osmond 1987; Anderson et al. 1988). In earlier studies, this PSII/PSI ratio was either lower (Boardman et al. 1972; Chu and Anderson 1984; Anderson et al. 1988) or higher (Melis and Harvey 1981) in shade species than in sun species. This discrepancy in data could have been due to the species investigated and their differing habitats, or the methods used for quantitation, particularly that for the PSII reaction centres, since different values for the concentration of PSII reaction centres have been obtained by various methods. In any case only a few shade-adapted species have been studied, and obviously it is necessary to examine a greater number of such species before any generalisation can be made regarding the stoichiometry of the two photosystems.
Our aims in this study were threefold: (1) to determine the amounts of functional PSI and PSII reaction centres on a chlorophyll basis in a variety of shade species growing in a deep shade environment; (2) to compare the photosystem stoichiometry of shade species with those of sun species, and species grown at high and low irradiance; and (3) to compare the effects of light quality and quantity on a selected species, peas.
Materials and Methods
Plant Material Leaves were collected from a variety of shade plants growing on the floor of an extensive shade
gully in the Australian National Botanic Gardens, Canberra (Table 1). The leaves were collected in
Table 1. List of rainforest and/or wet sclerophyll forest species, their family, geographical origin, region of occurrence in Australia and life form
ACT, Australian Capital Territory; NSW, New South Wales; Q, Queensland; SA, South Australia; T, Tasmania; V, Victoria
Species Family Origin Region Life form
Aristotelia australasica
Blechnum wattsii
Cordyline stricta Dicksonia
antarctica Doodia aspera
Hedycarya angustifolia
Helm holtzia glaberrima
Pollia crispata
Tasmannia purpurascens
Elaeocarpaceae
Blechnaceae
Agavaceae Cyatheaceae
Blechnaceae
Monimiaceae
Philydraceae
Commelinaceae
Winteraceae
New Enland National Park, NSW
-
Arakoon, NSW Tidbinbilla, ACT
Mallacoota, NSW
Brindabella Range, ACT/NSW border
Lamington National Park, Q
Mount Cambewarra, Central Coast NSW
Barrington Tops National Park, NSW
NSW
NSW
Shrub
Fern (low)
Shrub Fern
(arborescent) Fern (low)
Tree
Herb
Herb
Shrub

Photosystem Stoichiometry in Shade-adapted Species
early mornings in April and May, 1989 from the most shaded position of each plant; in every case the irradiance was less than 15 pmol photons m-2 s- ' even at mid-day.
Pea plants (Pisum sativum L. cv. Greenfeast) were grown in a glasshouse in natural light, together with spinach and barley plants. For low irradiance studies, pea plants were also grown in vermiculite/ perlite mixtures in a growth cabinet: the light source was white fluorescent tubes (Philips TLD 36W/86) operated with a 12 h light (45 pmol photons m-2 s-')/12 h dark cycle. The temperature was 24'C throughout. For light quality studies, peas were grown with a 16 h light/24OC; 8 h dark/l4OC cycle. The growth light was either fluorescent illumination filtered by yellow Plexiglas (PSII-light, - 69 pmol photons m-2 s-l, 520-695 nm, or incandescent illumination filtered by red Plexiglas (PSI-light, 48 pmol photons m-2 s- ' from 580 to 695 nm, plus 47 pmol photons m-2 s- ' from 695 to 740 nm). Because of the lower quantum yield obtainable with the far-red component of PSI-light, these two light treatments gave approximately equivalent irradiance. The plants were supplied with Hoagland's solution once weekly and with water daily. The irradiance below 695 nm was measured with a Li-Cor model LI-185A quantum photometer and the relative spectral irradiance of eight light quality regime measured by a spectroradiometer (SR3000A, Macam Photometrics Ltd, Livingstone, Scotland).
Chloroplasts were isolated by a gentle rapid method (Chow and Hope 1987). Leaf tissue was homogenised at O°C in a Waring blendor for the minimal time required to chop the tough leaves (<30 s) in 130 mL of isolation buffer containing 50 mM MES-NaOH (pH 6.5), 400 mM sorbitol, 5 mM MgC12, 10 mM NaCI, 0.2% sodium ascorbate, 0.2% bovine serum albumin, and 1% polyvinyl- pyrrolidone (PVP-40T, Sigma). The homogenate was filtered through three layers of Miracloth and immediately centrifuged for 1 min at 1000 x g. The chloroplasts were washed three times with the isolation buffer minus sodium ascorbate and polyvinylpyrrolidone and kept on ice, or promptly frozen in liquid N2.
Determination of PSI, PSII and Chlorophyll-Proteins
The concentration of P700 was determined from the absorbance changes at 703 nm induced by a 0.25 s flash of blue-green light (Corning 4-72, -700 pmol photons m-2 s-') in a flash photometer constructed by Professor W. Haehnel (Chow and Hope 1987). Thylakoids (25 p~ Chl) were suspended in ( m ~ ) : 50 TES-NaOH (pH 7 . 9 , 400 sucrose, 5 MgC12, 10 NaC1, 1 sodium ascorbate, 0 .1 methyl viologen, 0.03 dicholorophenolindophenol, 0.002 DCMU and 0.05% (v/v) nonidet. Under the above experimental conditions, maximum oxidation of P700 was achieved well within the duration of the actinic flash, with a typical half-time of about 10 ms, while the half-time for re-reduction in the dark was typically 1 s. An extinction coefficient of 64 m ~ - ' cm-' was used for P700 (reduced minus oxidised) at 703 nm (Hiyama and Ke 1972).
The concentration of PSII was measured in leaf pieces by the O2 evolution induced by a total of approximately 2000 repetitive flashes (duration 3 ps, at half-height) generated by a zenon flash lamp (Stroboslave type 1539-A) at 4 Hz (Chow et al. 1989). Leaves were dark-adapted for 15 min in a Hansatech (King's Lynn, UK) 0 2 electrode chamber thermostated at 25°C; the C 0 2 concentration was 1%. Continuous background far-red light ( - 17 pmol photons m-2 s-'; 700-730 nm) was present during flash illumination to ensure that electron transport by PSI was not limiting (Chow et al. 1989).
DCMU-binding was determined by the method of Tischer and Strotmann (1977), as modified by Chow et al. (1989). Thylakoids (1 mL, 50 p~ Chl), in 50 mM TES-NaOH (pH 7.5), 400 mM sucrose, 10 mM NaC1, 5 mM MgC12, were mixed with [ ' 4 C ] ~ C ~ ~ , (Amersham) stock (50 p~ in ethanol) and the ethanol concentrations were adjusted to be the same in the diuron concentration series from 50 to 400 nM. After equilibration in the dark for 3 min, the mixtures were centrifuged in an Eppendorf 5414 centrifuge for 3 min, and 0 .7 mL of the supernatant was mixed with Beckman Ready-Solv EP scintillant and counted. Standards were processed in the same way, except for the absence of the thylakoid membranes. The amount of DCMU bound was assumed to be given by the difference between the concentration in the supernatant with and without thylakoid membranes in the assays.
Leaf discs were finely chopped and ground with acetone and quantitatively made up to 80% acetone/2.5 mM Na-phosphate (pH 7.8). Chl content and Chl a/Chl b ratios were calculated according to Arnon (1949).
Thylakoid membranes were solubilised at O°C in 50 mM Tris (pH 8.0) containing 30% (v/v) glycerol and 0.75% (w/v) SDS to give a SDS/Chl (w/w) ratio of 7.5. Samples were applied immediately to SDS-PAGE tube gels made and run as described previously (Chu and Anderson 1984). The percentages of chlorophyll in individual bands were calculated from gel scans measured at 675 nm and 650 nm (Chu and Anderson 1984).
Tab
le 2
. C
hl a
/Chl
b r
atio
s an
d th
e ph
otos
yste
ms
reac
tion
cen
tre
stoi
chio
met
ry o
f sh
ade-
adap
ted
spec
ies
Fun
ctio
nal
[PS
II]
was
det
erm
ined
by
repe
titi
ve s
ingl
e tu
rnov
er f
lash
es i
n th
e pr
esen
ce o
f ba
ckgr
ound
far
-red
lig
ht.
P70
0 or
PS
I co
ncen
trat
ion
was
de
term
ined
as
the
ligh
t-in
duce
d ab
sorb
ance
cha
nge
at 7
03 n
m,
usin
g 30
p~
redu
ced
dich
loro
phen
olin
doph
enol
as
elec
tron
don
or.
Mea
n +
s.e.
m.
(n)
Spe
cies
C
hl a
C
hl a
+ b
Fun
ctio
nal
PS
I1
P70
0 F
unct
iona
l D
CM
U s
ites
D
CM
U s
ites
C
hl b
(p
mol
m--
2)
[mm
ol (
mol
Chi
)-'1
P
SI1
[m
mol
F
unct
iona
l P
SI1
P
SI
(mol
Chi
)-'1
Ari
stol
elia
aus
tral
asic
a B
lech
num
wat
tsii
C
ordy
lin
e st
rict
a
Dic
kson
ia a
nta
rcti
ca
Doo
dia
aspe
ra
Hed
ycar
ya a
ngu
stif
olia
Hel
mh
oltz
ia g
labe
rrir
na
Pol
lia
cris
pata
T
asm
anni
a pu
rpur
asce
ns

Photosystem Stoichiometry in Shade-adapted Species
Results
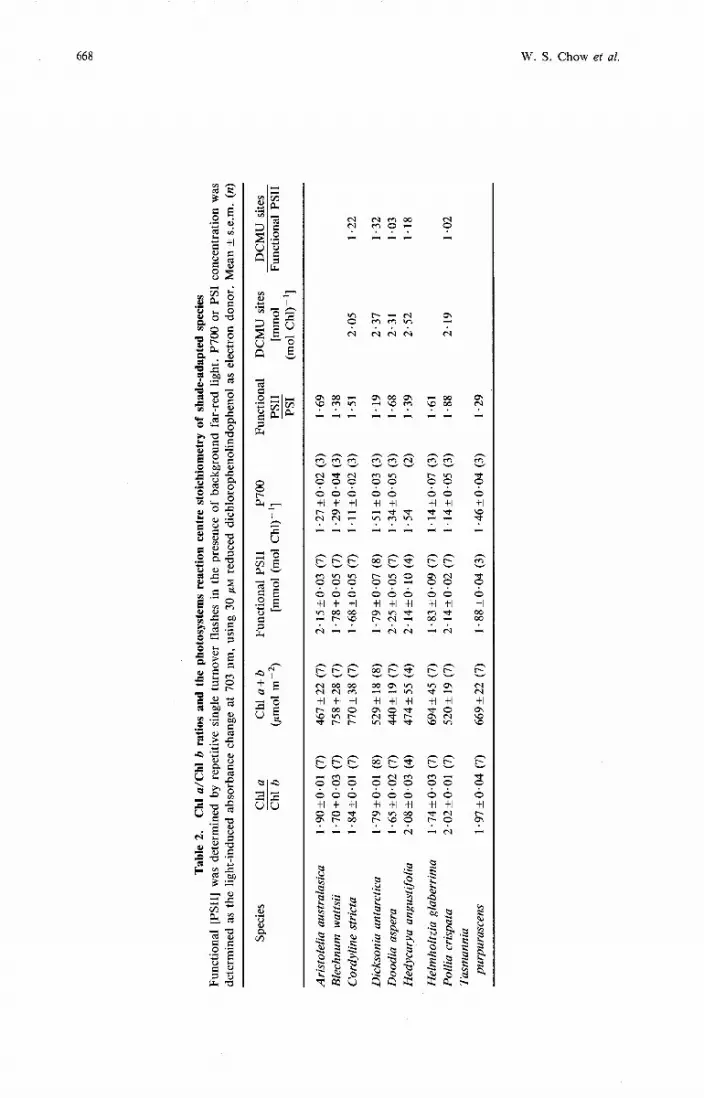
All the Chl a/Chl b ratios of the Australian shade species (Table 1) were very low, ranging from 1.7 to 2.0 (Table 2). It is well known that shade species have lower Chl a/ Chl b ratios than sun species, whose values are typically 2.8-3.3 (see Anderson and Osmond 1987). On a leaf area basis, their chlorophyll content was rather high (Table 2) a common but not universal characteristic of shade plants (Boardman 1977). Electron microscopic examination of the shade leaves and chloroplast structures showed that all the leaves had many more chloroplasts towards their upper surfaces, and the mesophyll chloroplasts were all larger than those of sun plants (data not shown). These large chloroplasts were almost filled with thylakoid membranes. As previously observed, each granum had many thylakoid compartments. The grana stacks were not only much wider (800 nm or more) but also the interconnecting stroma thylakoids were typically shorter than observed in sun species. These are the characteristic morphological features of most shade species (Goodchild et al. 1972).
The P700 content of these shade species ranged from 1.1 to 1.54 mmol P700 (mol ~ h 1 ) - ' (Table 2), so that the Chl/P700 ratios varied from about 650 to 900. On a Chl basis, the amounts of P700 are lower than those of sun species whose Chl/P700 ratios from many determinations are typically 610 + 40 (e.g. Melis 1984; Wild et al. 1986; Chow et al. 1988a; Jursinic and Pearcy 1988). These results confirm the findings of the first study to compare the P700 content of shade and open habitat species (Anderson et al. 1973), namely, that shade species had a lower P700 content.
The concentration of the functional PSII reaction centres (on a chlorophyll basis) in plant thylakoid membranes is still controversial, with different values being obtained with several methods. Recently, a method was developed to measure PSII directly in leaves, thereby avoiding any inactivation of functional PSII during thylakoid isolation or subsequent measurements (Chow et al. 1988a; Chow et al. 1989). The concentration of functional PSII centres in vivo was determined by the O2 yield of leaf discs exposed to 1% C02, and repetitive flashes at 4 Hz (xenon lamp, flash of 3 ps at half-height) in the presence of background far-red light to ensure no limitation in P700 turnover. The concentrations of functional PSII in the shade species determined with repetitive flashes range from 1.68 to 2.25 mmol (mol ~ h 1 ) - ' , i.e. Chl/PSII ratios from 444 to 595 (Table 2). Like the P700 content, the PSII content (on a chlorophyll basis) of shade species is also generally lower than that observed for sun species (Table 3). For example, peas, spinach and barley have 3.0-3.2 mmol PSII (mol Ch1)-' as determined by the flash O2 yield method in leaves grown in a glasshouse in spring/summer. Comparable values have also been observed in thylakoids from the same species by Melis and Brown (1980), Melis and Harvey (1981) and Melis (1985), who assayed PSII by the absorbance change of the primary quinone electron acceptor, QA, of PSII at 320 nm.
The method of repetitive single-turnover flashes for quantifying the active PSII content in isolated thylakoids in the presence of suitable PSII acceptors is taken as valid by many researchers (Eckert et al. 1984; Graan and Ort 1986; Wild et al. 1986; Jursinic and Dennenberg 1985). Using spinach which is well-suited for obtaining isolated thylakoids with minimal damage to the photosynthetic electron transport system, we found that the content of PSII estimated by repetitive flash yield of leaf discs was comparable to that of isolated thylakoids in the presence of ferricyanide (with or without 2,6-dichloro-p-benzoquinone): the PSII contents of leaf discs were no more than 8% higher than that of isolated thylakoids.
The estimation of PSII reaction centres by the number of herbicide-binding sites (particularly diuron and atrazine) is also considered valid for measuring the total PSII content of isolated thylakoid membranes (Tischer and Strotmann 1977; Renger et al. 1981; Jursinic and Dennenberg 1985; Graan and Ort 1986). We also measured the DCMU-binding capacity of isolated thylakoids from five shade species (Table 2); the values obtained are on average about 14% higher than those obtained with leaf discs (Table 2). Previously, we found the same differences between the in vivo PSII content measured by repetitive single-

W. S. Chow et al.
turnover flashes and the in vive DCMU-binding-sites of isolated thylakoids (Chow et al. 1988a, 1989).
Given that the number of binding sites for diuron and atrazine in a single batch of thylakoids may differ by as much as 20% (Chow et al. 1990), we consider that the method of repetitive flashes applied to leaf discs is at least as good as herbicide-binding sites or flash illumination of isolated thylakoids. This method using repetitive flashes was particularly useful for the shade species used in this study because their leaves were generally too tough for the isolation of active thylakoids. Measurement of PSII in vivo has also been reported independently by Jursinic and Pearcy (1988).
Table 3. Photosystem stoichiometry in sun species, and pea plants grown under high and low irradiance or yellow (favouring PSII) and
red plus far-red (favouring PSI) light The PSII content was assayed by repetitive single-turnover flashes except for pea grown in yellow or red plus far-red light, where PSII
was assayed by DCMU-binding
Chl a PSI1 PSI PSI1 - - Chl b [mmol (mol ~h1)- '1 PSI
Sun species grown in a glasshouse
Spinach 2 .8 2.98 1.71 1.74 Barley 3 .3 2.89 1.94 1.49 Pea 3 .3 2.93 1.64 1.79
Growth under low irradiance Pea 2 .6 2.15 1.69 1.27
Growth under yellow (mainly PSZZ) and red plus far-red (mainly PSI) light
Pea (yellow) 3 .0 1.75 1.73 1.01 Pea (red plus far-red) 2.5 2.02 1 .04 1.95
The stoichiometries of PSII/PSI reaction centres demonstrate a range of values obtained for the shade species examined from 1.19 to 1.88 (Table 2). Most of these values are lower than the 'consensus' PSII/PSI ratio of 1.7-1.8 found in a wide variety of sun species, but some are comparable. The results support earlier findings (Boardman et al. 1972; Anderson et al. 1988) that the PSII/PSI reaction centre stoichiometries of shade species are generally lower than those observed for sun species.
To examine the light-harvesting antenna size of PSII and PSI qualitatively, we also compared the amounts of chlorophyll present in the different thylakoid pigment-proteins resolved by non-denaturing gel electrophoresis (Chu and Anderson 1984). The relative amounts of chlorophyll associated with the various pigment-protein complexes of the thylakoid membranes of shade species are roughly comparable (Table 4). However, these values are consistently different from the values obtained with a typical sun species, spinach. The shade species had only 8.7-11.6% of the total chlorophyll associated with the core PSII complex, CPa, compared with 14.570 in sun species. With the decline in CPa content in shade species, there is a concomitant increase in the relative amount of LHCII. The amount of total chlorophyll associated with PSII (CPa and LHCII) relative to PSI (CPla and CPl) is greater in the shade species compared to spinach (Table 4). The LHCII/CPa ratios were 4.7-6.9 for the shade species examined, in contrast to 3 . 3 in spinach (Table 4). These data indicate that the apparent light-harvesting antenna size of PSII is much larger in the shade species relative to spinach, due to an increase in peripheral LHCII relative

Photosystem Stoichiometry in Shade-adapted Species
Table 4. Chlorophyll associated with the PSI and PSII pigment-protein complexes in the thylakoid membranes of shade species and a sun species (spinach)
Abbreviations: CPla and CP1 are photosystem I complexes with CPla containing more of the PSI light-harvesting complexes, LHCI, than CP1; LHCII is the light-harvesting complex of photosystem I1 resolved as individual bands, LHCP', LHCP' and LHCP~ (Chu and Anderson 1984); CPa is the core PSII complex. PSII Chl is the sum of LHCII and CPa; PSI Chl is the sum of CP la and CP1.
Each value is the average of at least three independent measurements
Species Chl a Percentage of chlorophyll contained PSI1 Chl LHCII Chl b in pigment-protein complexes PSI Chl CPa
CPla CP l LHCII CPa FP
Aristotelia australasica 1 .82 3.7 15.6 60.0 10.7 10.0 3.2 5.6 Dicksonia antarctica 1.88 9.7 10.7 60.0 8 .7 10.9 3 .4 6.9 Doodia aspera 1.79 9 .4 10.4 61.9 9 .1 8.2 3.7 6.8 Pollia crispata 1.85 4.9 16.5 55.3 11.6 12.2 3 . 1 4.8 Spinacia oleracea 2.82 7 .9 18.8 48.4 14.5 10.4 3.3 3.3
to the core PSII complex. Furthermore the amount of total chlorophyll associated with the PSI complexes (CPla and CP1) is lower for the shade species compared to spinach (Table 4).
To compare the effects of either light quantity or quality on the adjustment of the photosystem reaction centre stoichiometry in the same plant, pea plants were acclimated in a glasshouse under natural high irradiance or in a controlled growth cabinet under low irradiance, as well as yellow or red light. The amounts of PSII reaction centres of leaf discs were then estimated by the repetitive single-turnover O2 flash yields, and PSI content by the light-induced oxidation of P700 in isolated thylakoids as was the case for the shade species (Table 2). While the amount of P700 on a chlorophyll basis was not influenced by irradiance, there was more PSII in high-light than in low-light pea thylakoids (Table 3). Hence the PSII/PSI reaction centre ratio was greater in high than in low irradiance (Table 3), mimicking the differences observed in sun versus shade. This effect of irradiance on the photosystem reaction centre stoichiometry is already well-established (Leong and Anderson 1984; Wild et al. 1986; Chow and Hope 1987; Evans 1987a; Chow et al. 1989), although a smaller difference (maximum 29%) between the PSII/PSI reaction centre ratios has been reported for the pea cultivar, Progress 9, grown under high and low irradiances (Lee and Whitmarsh 1989). For comparison, when peas were grown under red plus far-red light which preferentially excited PSI only, there was more PSII reaction centres and less PSI reaction centres, compared to peas grown under yellow light which mainly excites PSII (Table 3). These compensatory effects mean that peas grown under red plus far-red light have higher PSII/PSI reaction centre ratios than those grown under yellow light (Table 3). The results of these light-quality treatments are comparable to earlier observations of Glick et al. (1986) with peas and Deng et al. (1989) with spinach.
Discussion
The shade light environment, even at a single location, may give rise to different amounts of PSII and PSI reaction centres in plant species growing in the same shaded habitat (Table 2). The PSII/PSI reaction centre ratios of shade species are generally lower than those of sun species (1.7-1.8), although some are comparable. Significantly, the PSII/PSI stoichiometries are not fixed at any value than can be ascribed to 'shade-adapted' species in general. Rather, the results show significant differences in the PSII/PSI ratio among diverse shade-adapted species.
None of the present species studied had the high PSII/PSI ratios of three shade plants previously observed by Melis and Harvey (1981), but agreed rather with other results which

W. S. Chow et al.
showed lower PSII/PSI reaction centre stoichiometries in shade-tolerant plants (Boardman et al. 1972; Chow et al. 1988b). Thus, the observation made by Melis (Melis and Harvey 1981; Melis 1984) that the more extensive grana of shade species correlated with higher PSWPSI ratios is not confirmed. The lower PSWPSI reaction centre ratios in shade plants compared with sun plants are due to a greater decrease in PSII content than PSI content on a chlorophyll basis (Table 2). The lower PSII content is consistent with the extensive appressed membranes of the grana stacks having fewer PSII complexes per unit membrane area. This difference in sun/shade responses is also evident in the distribution of freeze- fracture particles of sun and shade thylakoids, with shade species having fewer EF, large particles (PSII core) per unit area (Goodchild et al. 1972).
The apparent lack of uniform response among shade species cannot be attributed to any single factor. In addition to the unique light quality and light intensity in a shade environment, there may be other parameters that exert control on the PSII/PSI ratio in shade species. Among them, one could cite the ability of each species to adjust the light- harvesting antenna size of PSII versus PSI. Not only has the photosystem stoichiometry been regulated to various extents as part of the shade or low light response, but there is also less PSII and P700 on a chlorophyll basis compared to sun species (Tables 2 and 3). In the shade species examined, the lower content of the PSII reaction centres per unit chlorophyll implies a larger PSII light-harvesting antenna (Table 2). This is comfirmed by the qualitative distribution of chlorophyll amongst the chlorophyll-protein complexes (Table 4). With more of the total chlorophyll in the light-harvesting Chl a/b-proteins of PSII (LHCII) and less in the core Chl a-proteins, the light-harvesting antenna sizes of PSII in shade species are larger than those of sun species. This increase in the light-harvesting antenna of PSII in shade species would be expected from their lower Chl a/Chl b ratios. As suggested earlier (Anderson et al. 1988), on a chlorophyll basis, shade plants have fewer, but larger PSII units relative to PSI, in order to harvest effectively the meagre light, while sun plants have more, but smaller PSII units relative to PSI to maximise photosynthetic electron transport capacity and minimise photoinhibition at saturating light.
Despite the different composition and structure of shade and low-light chloroplasts compared to sun and high-light chloroplasts, all non-stressed C3 plants have the same quantum yield of O2 evolution under light-limiting conditions (Bjorkman and Demmig 1987; Evans 1987b). We suggest that this remarkable constancy of quantum yield is achieved by the light regulation (via the interplay of light quality and quantity) of both the relative amounts of PSII and PSI, and the light-harvesting antenna size of each photosystem. If, for example, only the amounts of PSII reaction centres (but not the antenna size) were to change in response to light, there would be an imbalance of excitation of the two photo- systems, resulting in a loss of quantum yield of O2 evolution.
Compared with the exposed canopy, the light environment experienced by plants growing on the forest floor is a unique blend of markedly attenuated light which is enriched in far-red irradiance absorbed mainly by photosystem I, and short, intense bursts of direct sunlight (Bjorkman and Ludlow 1972). To compare whether light quality or quantity might be the predominant regulatory factor in shade light, we compared the effects of alterations of either light quantity or light quality in the same species, pea, using the same methods as we had used to determine the photosystem reaction centre stoichiometry in shade species (Table 2). It is already well-known that light quantity alone regulates photosystem reaction centre stoichiometry, with plants grown in low irradiance having lower PSII/PSI ratios (due to a lower PSII content) than those grown in high irradiance (cf. Anderson and Osmond 1987; Table 3).
Light quality alone also regulates the photosystem reaction centre stoichiometry. Plants grown in light mainly absorbed by PSI (either in far-red enriched or red light) had relatively less PSI and more PSII based on a chlorophyll basis, than those grown in light absorbed by both photosystems or mainly PSII (either far-red deficient or yellow light) (Table 3; Melis and Harvey 1981; Glick et al. 1986; Deng et al. 1989). This modulation of PSII/PSI

Photosystem Stoichiometry in Shade-adapted Species
reaction centre ratios by light quality is a compensatory strategy designed to help correct the marked imbalance in the light excitation energy received by the two photosystems.
In conclusion, it is now clear that both light quantity and light quality can regulate the amounts of PSII and PSI reaction centres on a chlorophyll basis as well as the photosystem stoichiometry (Tables 2 and 3). This regulation is not surprising, since shade canopy light is both greatly attenuated and also altered in light quality, and in nature changes in irradiance are usually accompanied by changes in light quality. Thus, in the canopy shade environment, the lower irradiance will tend to decrease the amount of PSII reaction centres on a chlorophyll basis (Table 3). However, the concomitant increase in far-red light, absorbed preferentially by PSI, will tend to increase the amount of PSII reaction centres on a chlorophyll basis (Glick et al. 1986, also Table 3) in order to balance the input of excitation energy to both photosystems. We conclude that it is the opposing interplay of both light quantity and quality that modulates the photosystem reaction centre ratio and, concomitantly, the size of the light-harvesting antennae of PSII. Since the PSII reaction centre content on a chlorophyll basis is usually lower in shade species (Table 2) compared to sun species (Table 3) it seems that, in a shaded terrestrial habitat, the marked decrease in light quantity may play a greater role than the altered light quality in the adjustment of PSII reaction centre content and photosystem stoichiometry with land plants.
Acknowledgments
We are very grateful to Mr G. Butler of the Australian National Botanic Gardens, Canberra, for permission and help in the collection of plant material, Mr L. A. Craven of the Australian National Herbarium, Canberra, for assistance in taxonomy, Drs N. K. Boardman and J . R. Evans for helpful discussion, and to Stephanie Hossack-Smith and Celia Miller for their excellent technical assistance.
References
Anderson, J . M., and Osmond, C. B. (1987). Shade-sun responses: compromises between accliniation and photoinhibition. In 'Photoinhibition'. (Eds D. J . Kyle, C. B. Osmond and C. J. Arntzen.) Topics in Photosynthesis, Vol. 9, pp. 1-38. (Elsevier: Amsterdam.)
Anderson, J . M., Chow, W. S. and Goodchild, D. J . (1988). Thylakoid membrane organisation in s u d s h a d e acclimation. Australian Journal of Plant Physiology 15, 11-26.
Anderson, J . M., Goodchild, D. J., and Boardman, N. K. (1973). Composition of the photosystems and chloroplast structure in extreme shade plants. Biochimica et Biophysica Acta 325, 573-85.
Arnon, D. I. (1949). Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Beta vulgaris. Plant Physiology 24, 1-15.
Bjorkman, O., and Demmig, B. (1987). Photon yield of Oz evolution and chlorophyll fluorescence characteristics at 77K among vascular plants of diverse origins. Planta 170, 489-504.
Bjorkman, O., and Ludlow, M. M. (1972). Characterization of the light climate on the floor of a Queensland rainforest. Carnegie Institution of Washington Year Book 71, 85-94.
Boardman, N. K. (1977). Comparative photosynthesis of sun and shade plants. Annual Review of Plant Physiology 28, 355-77.
Boardman, N. K., Anderson, J . M., Thorne, S. W., and Bjorkman, 0 . (1972). Photochemical reactions of chloroplasts and components of the photosynthetic electron transport chain in two rainforest species. Carnegie Institution of Washington Year Book 71, 107-14.
Chow, W. S., and Hope, A. B. (1987). The stoichiometries of supramolecular complexes in thylakoid membranes from spinach chloroplasts. Australian Journal of Plant Physiology 14, 21-8.
Chow, W. S., Anderson, J. M., and Hope, A. B. (1988~). Variable stoichiometrics of photosystem I1 to photosystem I reaction centres. Photosynthesis Research 17, 277-81.
Chow, W. S., Hope, A. B., and Anderson, J. M. (1989). Oxygen per flash from leaf discs quantifies photosystem 11. Biochimica et Biophysica Acta 973, 105-8.
Chow, W. S., Hope, A. B., and Anderson, J. M. (1990). A reassessment of the use of herbicide binding to measure photosystem I1 reaction centres in plant thylakoids. Photosynthetis Research 24, 109-13.

W. S. Chow et al.
Chow, W. S., Qian, L., Goodchild, D. J., and Anderson, J. M. (1988b). Photosynthetic acclimation of Alocasia macrorrhiza (L.) G. Don to growth irradiance: structure, function and composition of chloroplasts. Australian Journal of Plant Physiology 15, 107-22.
Chu, Z.-X., and Anderson, J . M. (1984). Modulation of the light-harvesting assemblies in chloroplasts of a shade plant, Alocasia macrorrhiza. Photobiochemistry and Photobiophysics 8, 1-10.
Deng, X.-W., Tonkyn, J . C., Peter, G. F., Thornber, J . P., and Gruissem, W. (1989). Post-trans- criptional control of plastid mRNA accumulation during adaptation of chloroplasts to different light quality environments. The Plant CeN 1, 645-54.
Eckert, H.-J., Renger, G., and Witt, H. T. (1984). Reduction kinetics of the photo-oxidized chlorophyll aIIC in the nanosecond range. Measurements of the absorption changes at 688 nm under repetitive flash excitation. Federation of European Biochemical Societies Letters 167, 316-20.
Evans, J . R. (1987a). The relationship between electron transport components and photosynthetic capacity in pea leaves grown at different irradiances. Australian Journal of Plant Physiology 14, 157-70.
Evans, J . R. (1987b). The dependence of quantum yield on wavelength and growth irradiance. Australian Journal of Plant Physiology 14, 69-79.
Glick, R. E., McCauley, S. W., Gruissem, W., and Melis, A. (1986). Light quality regulates expression of chloroplast genes and assembly of photosynthetic membrane complexes. Proceedings of the National Academy of Sciences, U.S.A. 83, 4287-91.
Goodchild, D. J., Bjorkman, O., and Pyliotis, N. A. (1972). Chloroplast ultrastructure, anatomy, and content of chlorophyll and soluble protein in rainforest species. Carnegie Institution of Washington Year Book 71, 102-7.
Graan, T., and Ort, D. R. (1986). Detection of oxygen-evolving photosystem I1 centres inactive in plastoquinone reduction. Biochimica et Biophysica Acta 852, 320-30.
Hiyama, T., and Ke, B. (1972). Difference spectra and extinction coefficients of P700. Biochimica et Biophysica Acta 267, 160-71.
Jursinic, P . A., and Dennenberg, R. (1985). Reconciliation of the absorption change at 325 nm and other flash-induced determinations of concentrations of active vhotos~stem I1 centres. Archives o f Biochemistry and Biophysics 241, 540-9.
Jursinic, P . A., and Pearcy, R. W. (1988). Determination of the rate limiting step for photosynthesis in a nearly isonuclear rapeseed (Brassica napus L.) biotype resistant to atrazine. Plant Physiology 88, 1195-200.
Lee, W. J., and Whitmarsh, J. (1989). Photosynthetic apparatus of pea thylakoid membranes. Plant Physiology 89, 932-40.
Leong, T. Y., and Anderson, J. M. (1984). Adaptation of the thylakoid membranes of pea chloroplasts to light intensities. 11. Regulation of electron transport capacities, electron carriers, coupling factor (CF1) activity and rates of photosynthesis. Photosynthesis Research 5, 117-28.
Lichtenthaler, H. K., and Meier, D. (1984). Regulation of chloroplast photomorphogenesis by light intensity and light quality. In 'Chloroplast Biogenesis'. (Ed R. J. Ellis.) pp. 261-81. (Cambridge University Press: Cambridge.)
Melis, A. (1984). Light regulation of photosynthetic membrane structure, organization and function. Journal of Cell Biology 24, 271-85.
Melis, A. (1985). Functional properties of photosystem IIB in spinach chloroplasts. Biochimica et Biophysica Acta 808, 334-42.
Melis, A., and Brown, J . S. (1980). Stoichiometry of system I and system I1 reaction centers and of plastoquinone in different photosynthetic membranes. Proceedings of the National Academy of Sciences, U.S.A. 77, 4712-16.
Melis, A., and Harvey, G. W. (1981). Regulation of photosystem stoichiometry, chlorophyll a and chlorophyll b content and relation to chloroplast ultrastructure. Biochimica et Biophysica Acta 637, 138-45.
Renger, G., Fromm, R., and Hagemann, R. (1981). The modification of atrazine binding by the redox state of the endogenous high-spin iron and by specific proteolytic enzymes in photosystem I1 membrane fragments and intact thylakoids. Biochimica et Biophysica Acta 935, 173-83.
Tischer, W., and Strotmann, H. (1977). Relationship between inhibitor binding by chloroplasts and inhibition of photosynthetic electron transport. Biochimica et Biophysica Acta 460, 113-25.
Wild, A., Hopfner, M., Riihle, W., and Richter, M. (1986). Changes in the stoichiometry of PSI1 components as an adaptive response to high-light and low-light conditions during growth. Zeitschrift fur Naturforschung 41C, 597-603.
Manuscript received 22 May 1990, accepted 13 July 1990