the organizing principles of neuronal avalanches: cell assemblies in the cortex?
TRANSCRIPT

The organizing principles of neuronalavalanches: cell assemblies in thecortex?Dietmar Plenz and Tara C. Thiagarajan
Section of Neural Network Physiology, National Institute of Mental Health, Porter Neuroscience Research Center, Building 35,
room 3A-100, 35 Convent Drive, Bethesda, MD 20892, USA
Review TRENDS in Neurosciences Vol.30 No.3
Neuronal avalanches are spatiotemporal patterns ofneuronal activity that occur spontaneously in superficiallayers of the mammalian cortex under variousexperimental conditions. These patterns reflect fastpropagation of local synchrony, display a richspatiotemporal diversity and recur over several hours.The statistical organization of pattern sizes is invariantto the choice of spatial scale, demonstrating that thefunctional linking of cortical sites into avalanches occurson all spatial scales with a fractal organization. Thesefeatures suggest an underlying network of neuronalinteractions that balances diverse representations withpredictable recurrence, similar to what has been theo-rized for cell assembly formation. We propose that ava-lanches reflect the transient formation of cell assembliesin the cortex and discuss various models that providemechanistic insights into the underlying dynamics,suggesting that they arise in a critical regime.
IntroductionThat memories, and indeed behaviors, are represented bythe participation of distributed groups of neurons in a cellassembly [1,2] has been one of the most powerful andenduring ideas in neuroscience. The synchronization ofneuronal activity observed during various cognitive andbehavioral tasks has been proposed as one mechanism bywhich neuronal participation in a cell assembly might beidentified [3,4]. Moreover, the propagation of synchrony inthe form of ‘synfire chains’ [5] has been suggested as atheoretical construct for the dynamic selection of neuronalgroups into cell assemblies [6,7]. However, the organiz-ation of the dynamics giving rise to cell assemblies has yetto be identified. If distributed patterns of synchronizedneuronal activity are indeed the physical correlate ofmemories and behaviors [8,9], then cortical activity mustproduce these synchronized patterns in a dynamic regimethat is highly flexible but yet enables a certain degree ofpredictability. Dynamics that give rise to cell assemblieswould have to be capable of producing a rich diversity ofpatterns of various sizes, many of which must be stablyreconstructed over time [10–14].
Here, we discuss recent findings on spontaneouslyarising synchronized activity patterns in superficial
Corresponding author: Plenz, D. ([email protected]).Available online 1 February 2007.
www.sciencedirect.com 0166-2236/$ – see front matter � 2006 Elsevier Ltd. All rights reserve
cortical layers – i.e. ‘neuronal avalanches’ [12,15,16], inthe context of cell assembly formation. Although thedefinition of a ‘cell assembly’ has been associated withvarious mechanisms such as attractor neural networks,here we define a cell assembly as a group of neurons whoseactivity is transiently linked together by an active processintrinsic to the network. In addition, we define ‘synchro-nized’ simply as sufficiently temporally proximal so as toenable superposition in the extracellular field. We showthat neuronal avalanches are attractive from a cell assem-bly perspective because of the large diversity of avalanchepatterns found at many different scales, their stable recur-rence in time with high precision and the interestingproperties of their statistics that indicate rich internaldynamics with long-range correlations capable of complexoutcomes. We further discuss how this form of activitycould reflect a ‘critical state’ in the network, drawing fromvarious models on complex systems dynamics.
Neuronal avalanches represent spatially irregularpatterns of propagated synchronyNeuronal avalanches are a previously unknown but robustform of synchronous activity in the cortex that was firstmeasured in organotypic somatosensory cortex culturesfrom rat, grown on microelectrode arrays (MEA). Theactivity arose spontaneously after maturation of the cul-tures and depended on fast inhibition [15]. It was alsofound that neuronal avalanches could be induced in acutesomatosensory and medial prefrontal cortex slices fromadult rats with a combination of N-methyl-D-aspartate(NMDA) and dopamine or a dopamine D1 receptor agonist[15,16] (Figure 1a,b). In these experimental paradigms, theactivity that was measured at each electrode was a localfield potential (LFP). The MEA configuration used forthese in vitro recordings was an 8 � 8 grid of electrodes,uniformly spaced at 200 mm, enabled measurements ofnonoverlapping LFPs across �2.6 mm2 of cortex(Figure 1a). The LFP activity displayed frequent negativecomponents which took the characteristic shape of a popu-lation spike: a sharp (10–50 ms) negative deflection oftenfollowed by a longer-lasting positive deflection, a featurelargely restricted to the superficial layers of the cortex [16](Figure 1a,b). The negative deflection of the LFP (nLFP) isof particular interest because, in vitro, where the thintissue slice is submerged in grounded medium, it is likely
d. doi:10.1016/j.tins.2007.01.005
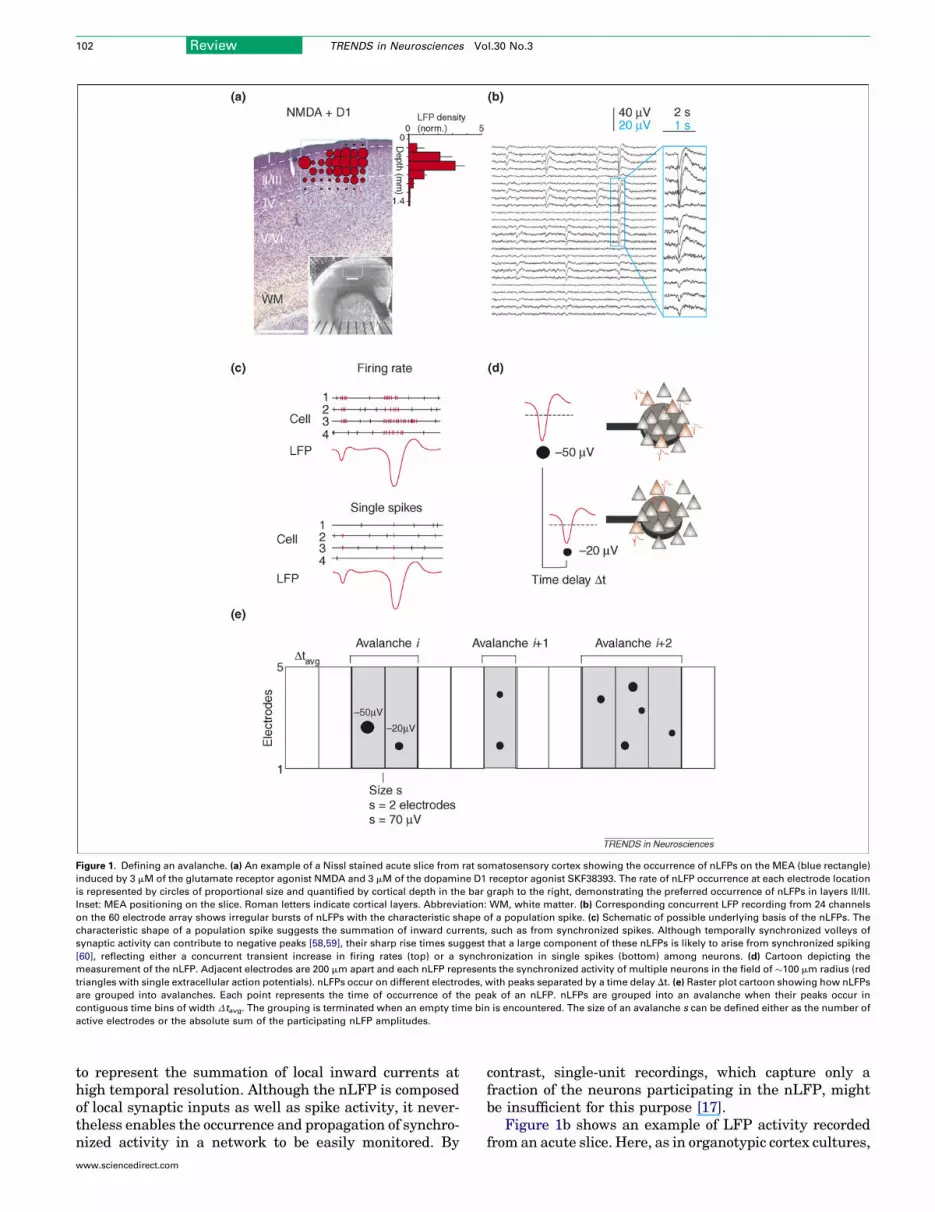
Figure 1. Defining an avalanche. (a) An example of a Nissl stained acute slice from rat somatosensory cortex showing the occurrence of nLFPs on the MEA (blue rectangle)
induced by 3 mM of the glutamate receptor agonist NMDA and 3 mM of the dopamine D1 receptor agonist SKF38393. The rate of nLFP occurrence at each electrode location
is represented by circles of proportional size and quantified by cortical depth in the bar graph to the right, demonstrating the preferred occurrence of nLFPs in layers II/III.
Inset: MEA positioning on the slice. Roman letters indicate cortical layers. Abbreviation: WM, white matter. (b) Corresponding concurrent LFP recording from 24 channels
on the 60 electrode array shows irregular bursts of nLFPs with the characteristic shape of a population spike. (c) Schematic of possible underlying basis of the nLFPs. The
characteristic shape of a population spike suggests the summation of inward currents, such as from synchronized spikes. Although temporally synchronized volleys of
synaptic activity can contribute to negative peaks [58,59], their sharp rise times suggest that a large component of these nLFPs is likely to arise from synchronized spiking
[60], reflecting either a concurrent transient increase in firing rates (top) or a synchronization in single spikes (bottom) among neurons. (d) Cartoon depicting the
measurement of the nLFP. Adjacent electrodes are 200 mm apart and each nLFP represents the synchronized activity of multiple neurons in the field of �100 mm radius (red
triangles with single extracellular action potentials). nLFPs occur on different electrodes, with peaks separated by a time delay Dt. (e) Raster plot cartoon showing how nLFPs
are grouped into avalanches. Each point represents the time of occurrence of the peak of an nLFP. nLFPs are grouped into an avalanche when their peaks occur in
contiguous time bins of width Dtavg. The grouping is terminated when an empty time bin is encountered. The size of an avalanche s can be defined either as the number of
active electrodes or the absolute sum of the participating nLFP amplitudes.
102 Review TRENDS in Neurosciences Vol.30 No.3
to represent the summation of local inward currents athigh temporal resolution. Although the nLFP is composedof local synaptic inputs as well as spike activity, it never-theless enables the occurrence and propagation of synchro-nized activity in a network to be easily monitored. By
www.sciencedirect.com
contrast, single-unit recordings, which capture only afraction of the neurons participating in the nLFP, mightbe insufficient for this purpose [17].
Figure 1b shows an example of LFP activity recordedfrom an acute slice. Here, as in organotypic cortex cultures,
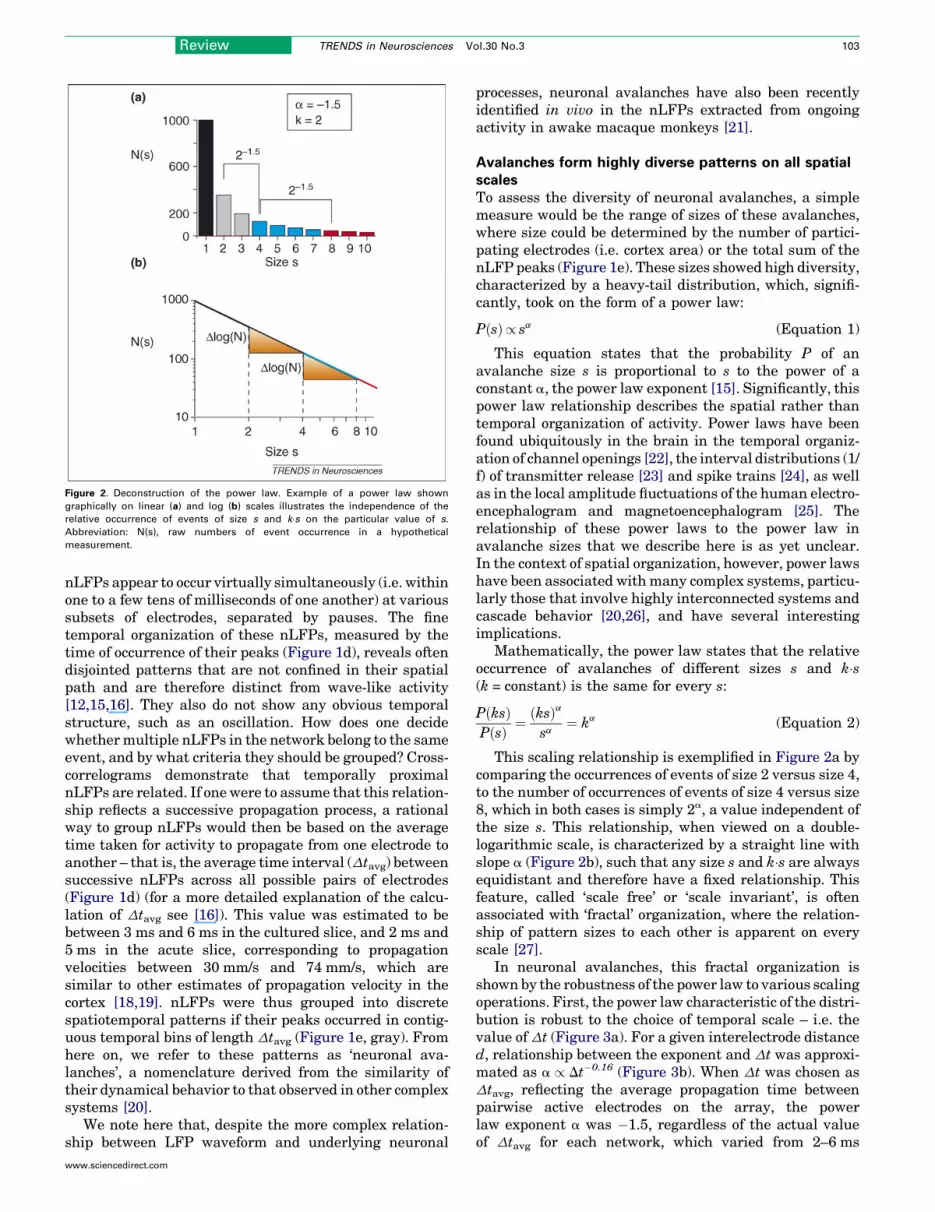
Figure 2. Deconstruction of the power law. Example of a power law shown
graphically on linear (a) and log (b) scales illustrates the independence of the
relative occurrence of events of size s and k�s on the particular value of s.
Abbreviation: N(s), raw numbers of event occurrence in a hypothetical
measurement.
Review TRENDS in Neurosciences Vol.30 No.3 103
nLFPs appear to occur virtually simultaneously (i.e. withinone to a few tens of milliseconds of one another) at varioussubsets of electrodes, separated by pauses. The finetemporal organization of these nLFPs, measured by thetime of occurrence of their peaks (Figure 1d), reveals oftendisjointed patterns that are not confined in their spatialpath and are therefore distinct from wave-like activity[12,15,16]. They also do not show any obvious temporalstructure, such as an oscillation. How does one decidewhethermultiple nLFPs in the network belong to the sameevent, and by what criteria they should be grouped? Cross-correlograms demonstrate that temporally proximalnLFPs are related. If one were to assume that this relation-ship reflects a successive propagation process, a rationalway to group nLFPs would then be based on the averagetime taken for activity to propagate from one electrode toanother – that is, the average time interval (Dtavg) betweensuccessive nLFPs across all possible pairs of electrodes(Figure 1d) (for a more detailed explanation of the calcu-lation of Dtavg see [16]). This value was estimated to bebetween 3 ms and 6 ms in the cultured slice, and 2 ms and5 ms in the acute slice, corresponding to propagationvelocities between 30 mm/s and 74 mm/s, which aresimilar to other estimates of propagation velocity in thecortex [18,19]. nLFPs were thus grouped into discretespatiotemporal patterns if their peaks occurred in contig-uous temporal bins of length Dtavg (Figure 1e, gray). Fromhere on, we refer to these patterns as ‘neuronal ava-lanches’, a nomenclature derived from the similarity oftheir dynamical behavior to that observed in other complexsystems [20].
We note here that, despite the more complex relation-ship between LFP waveform and underlying neuronal
www.sciencedirect.com
processes, neuronal avalanches have also been recentlyidentified in vivo in the nLFPs extracted from ongoingactivity in awake macaque monkeys [21].
Avalanches form highly diverse patterns on all spatialscalesTo assess the diversity of neuronal avalanches, a simplemeasure would be the range of sizes of these avalanches,where size could be determined by the number of partici-pating electrodes (i.e. cortex area) or the total sum of thenLFP peaks (Figure 1e). These sizes showed high diversity,characterized by a heavy-tail distribution, which, signifi-cantly, took on the form of a power law:
PðsÞ/ sa (Equation 1)
This equation states that the probability P of anavalanche size s is proportional to s to the power of aconstant a, the power law exponent [15]. Significantly, thispower law relationship describes the spatial rather thantemporal organization of activity. Power laws have beenfound ubiquitously in the brain in the temporal organiz-ation of channel openings [22], the interval distributions (1/f) of transmitter release [23] and spike trains [24], as wellas in the local amplitude fluctuations of the human electro-encephalogram and magnetoencephalogram [25]. Therelationship of these power laws to the power law inavalanche sizes that we describe here is as yet unclear.In the context of spatial organization, however, power lawshave been associated with many complex systems, particu-larly those that involve highly interconnected systems andcascade behavior [20,26], and have several interestingimplications.
Mathematically, the power law states that the relativeoccurrence of avalanches of different sizes s and k�s(k = constant) is the same for every s:
PðksÞPðsÞ ¼
ðksÞa
sa¼ ka (Equation 2)
This scaling relationship is exemplified in Figure 2a bycomparing the occurrences of events of size 2 versus size 4,to the number of occurrences of events of size 4 versus size8, which in both cases is simply 2a, a value independent ofthe size s. This relationship, when viewed on a double-logarithmic scale, is characterized by a straight line withslope a (Figure 2b), such that any size s and k�s are alwaysequidistant and therefore have a fixed relationship. Thisfeature, called ‘scale free’ or ‘scale invariant’, is oftenassociated with ‘fractal’ organization, where the relation-ship of pattern sizes to each other is apparent on everyscale [27].
In neuronal avalanches, this fractal organization isshown by the robustness of the power law to various scalingoperations. First, the power law characteristic of the distri-bution is robust to the choice of temporal scale – i.e. thevalue of Dt (Figure 3a). For a given interelectrode distanced, relationship between the exponent and Dt was approxi-mated as a / Dt�0.16 (Figure 3b). When Dt was chosen asDtavg, reflecting the average propagation time betweenpairwise active electrodes on the array, the powerlaw exponent a was �1.5, regardless of the actual valueof Dtavg for each network, which varied from 2–6 ms
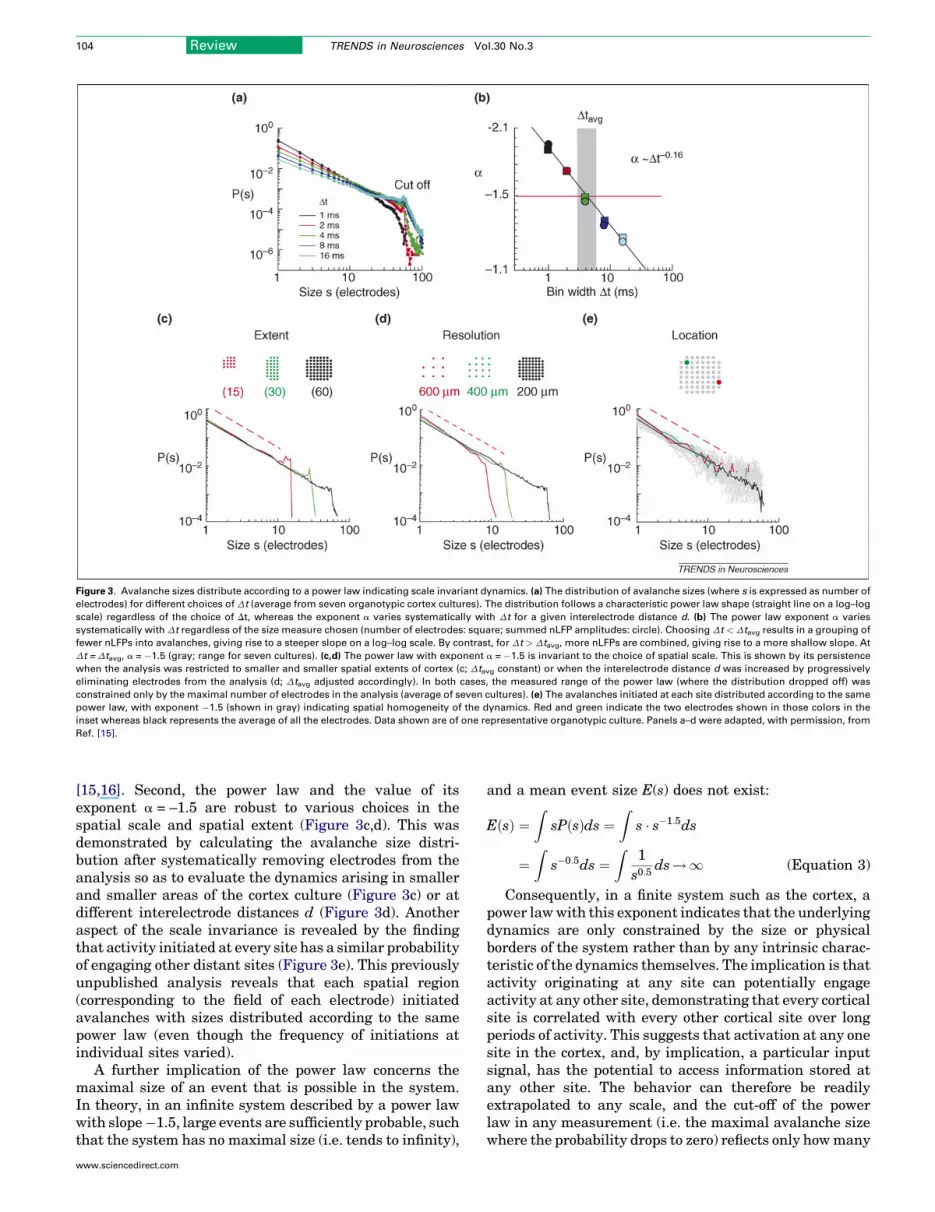
Figure 3. Avalanche sizes distribute according to a power law indicating scale invariant dynamics. (a) The distribution of avalanche sizes (where s is expressed as number of
electrodes) for different choices of Dt (average from seven organotypic cortex cultures). The distribution follows a characteristic power law shape (straight line on a log–log
scale) regardless of the choice of Dt, whereas the exponent a varies systematically with Dt for a given interelectrode distance d. (b) The power law exponent a varies
systematically with Dt regardless of the size measure chosen (number of electrodes: square; summed nLFP amplitudes: circle). Choosing Dt < Dtavg results in a grouping of
fewer nLFPs into avalanches, giving rise to a steeper slope on a log–log scale. By contrast, for Dt > Dtavg, more nLFPs are combined, giving rise to a more shallow slope. At
Dt = Dtavg, a = �1.5 (gray; range for seven cultures). (c,d) The power law with exponent a = �1.5 is invariant to the choice of spatial scale. This is shown by its persistence
when the analysis was restricted to smaller and smaller spatial extents of cortex (c; Dtavg constant) or when the interelectrode distance d was increased by progressively
eliminating electrodes from the analysis (d; Dtavg adjusted accordingly). In both cases, the measured range of the power law (where the distribution dropped off) was
constrained only by the maximal number of electrodes in the analysis (average of seven cultures). (e) The avalanches initiated at each site distributed according to the same
power law, with exponent �1.5 (shown in gray) indicating spatial homogeneity of the dynamics. Red and green indicate the two electrodes shown in those colors in the
inset whereas black represents the average of all the electrodes. Data shown are of one representative organotypic culture. Panels a–d were adapted, with permission, from
Ref. [15].
104 Review TRENDS in Neurosciences Vol.30 No.3
[15,16]. Second, the power law and the value of itsexponent a = –1.5 are robust to various choices in thespatial scale and spatial extent (Figure 3c,d). This wasdemonstrated by calculating the avalanche size distri-bution after systematically removing electrodes from theanalysis so as to evaluate the dynamics arising in smallerand smaller areas of the cortex culture (Figure 3c) or atdifferent interelectrode distances d (Figure 3d). Anotheraspect of the scale invariance is revealed by the findingthat activity initiated at every site has a similar probabilityof engaging other distant sites (Figure 3e). This previouslyunpublished analysis reveals that each spatial region(corresponding to the field of each electrode) initiatedavalanches with sizes distributed according to the samepower law (even though the frequency of initiations atindividual sites varied).
A further implication of the power law concerns themaximal size of an event that is possible in the system.In theory, in an infinite system described by a power lawwith slope�1.5, large events are sufficiently probable, suchthat the system has no maximal size (i.e. tends to infinity),
www.sciencedirect.com
and a mean event size E(s) does not exist:
EðsÞ ¼Z
sPðsÞds ¼Z
s � s�1:5ds
¼Z
s�0:5ds ¼Z
1
s0:5ds!1 (Equation 3)
Consequently, in a finite system such as the cortex, apower lawwith this exponent indicates that the underlyingdynamics are only constrained by the size or physicalborders of the system rather than by any intrinsic charac-teristic of the dynamics themselves. The implication is thatactivity originating at any site can potentially engageactivity at any other site, demonstrating that every corticalsite is correlated with every other cortical site over longperiods of activity. This suggests that activation at any onesite in the cortex, and, by implication, a particular inputsignal, has the potential to access information stored atany other site. The behavior can therefore be readilyextrapolated to any scale, and the cut-off of the powerlaw in any measurement (i.e. the maximal avalanche sizewhere the probability drops to zero) reflects only howmany
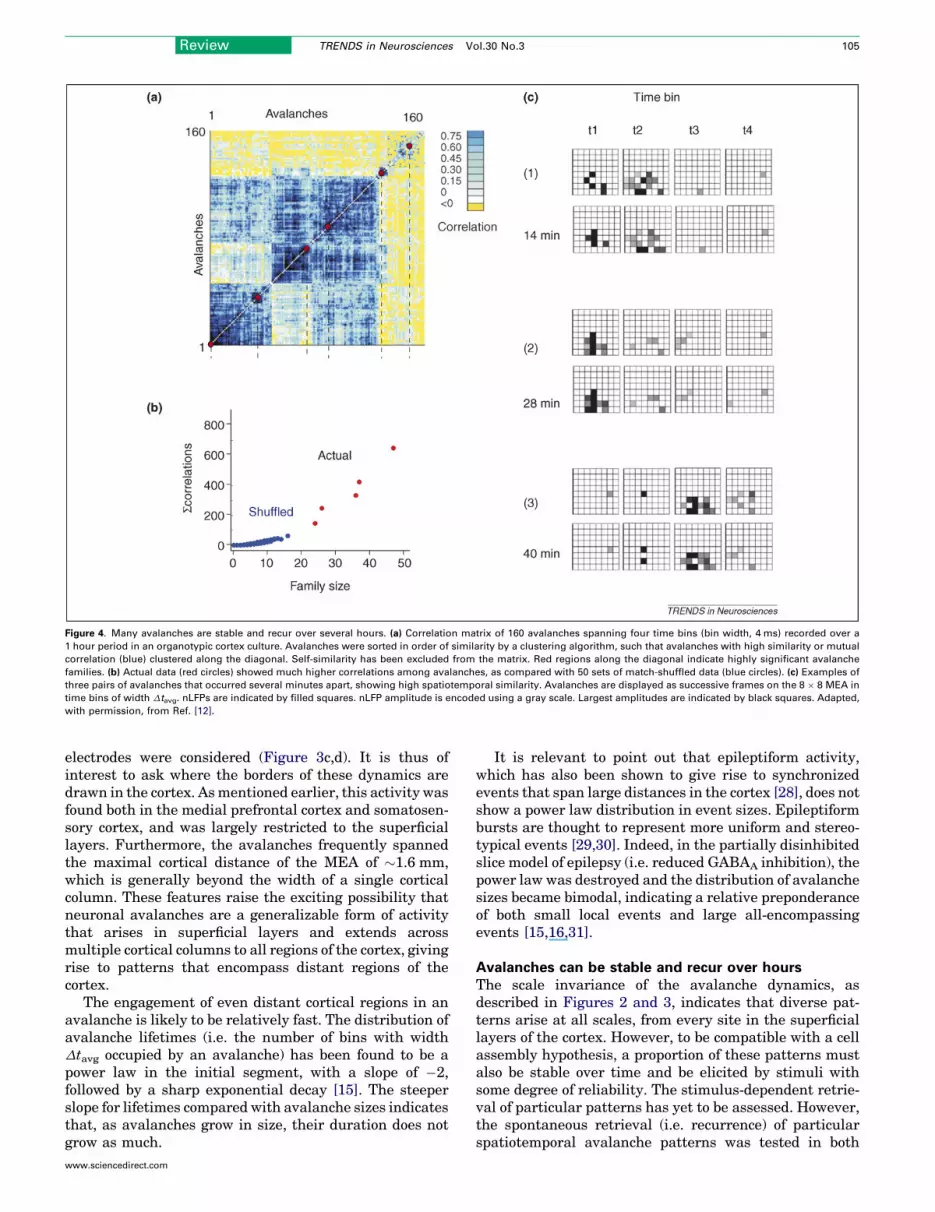
Figure 4. Many avalanches are stable and recur over several hours. (a) Correlation matrix of 160 avalanches spanning four time bins (bin width, 4 ms) recorded over a
1 hour period in an organotypic cortex culture. Avalanches were sorted in order of similarity by a clustering algorithm, such that avalanches with high similarity or mutual
correlation (blue) clustered along the diagonal. Self-similarity has been excluded from the matrix. Red regions along the diagonal indicate highly significant avalanche
families. (b) Actual data (red circles) showed much higher correlations among avalanches, as compared with 50 sets of match-shuffled data (blue circles). (c) Examples of
three pairs of avalanches that occurred several minutes apart, showing high spatiotemporal similarity. Avalanches are displayed as successive frames on the 8 � 8 MEA in
time bins of width Dtavg. nLFPs are indicated by filled squares. nLFP amplitude is encoded using a gray scale. Largest amplitudes are indicated by black squares. Adapted,
with permission, from Ref. [12].
Review TRENDS in Neurosciences Vol.30 No.3 105
electrodes were considered (Figure 3c,d). It is thus ofinterest to ask where the borders of these dynamics aredrawn in the cortex. Asmentioned earlier, this activity wasfound both in the medial prefrontal cortex and somatosen-sory cortex, and was largely restricted to the superficiallayers. Furthermore, the avalanches frequently spannedthe maximal cortical distance of the MEA of �1.6 mm,which is generally beyond the width of a single corticalcolumn. These features raise the exciting possibility thatneuronal avalanches are a generalizable form of activitythat arises in superficial layers and extends acrossmultiple cortical columns to all regions of the cortex, givingrise to patterns that encompass distant regions of thecortex.
The engagement of even distant cortical regions in anavalanche is likely to be relatively fast. The distribution ofavalanche lifetimes (i.e. the number of bins with widthDtavg occupied by an avalanche) has been found to be apower law in the initial segment, with a slope of �2,followed by a sharp exponential decay [15]. The steeperslope for lifetimes compared with avalanche sizes indicatesthat, as avalanches grow in size, their duration does notgrow as much.
www.sciencedirect.com
It is relevant to point out that epileptiform activity,which has also been shown to give rise to synchronizedevents that span large distances in the cortex [28], does notshow a power law distribution in event sizes. Epileptiformbursts are thought to represent more uniform and stereo-typical events [29,30]. Indeed, in the partially disinhibitedslice model of epilepsy (i.e. reduced GABAA inhibition), thepower law was destroyed and the distribution of avalanchesizes became bimodal, indicating a relative preponderanceof both small local events and large all-encompassingevents [15,16,31].
Avalanches can be stable and recur over hoursThe scale invariance of the avalanche dynamics, asdescribed in Figures 2 and 3, indicates that diverse pat-terns arise at all scales, from every site in the superficiallayers of the cortex. However, to be compatible with a cellassembly hypothesis, a proportion of these patterns mustalso be stable over time and be elicited by stimuli withsome degree of reliability. The stimulus-dependent retrie-val of particular patterns has yet to be assessed. However,the spontaneous retrieval (i.e. recurrence) of particularspatiotemporal avalanche patterns was tested in both
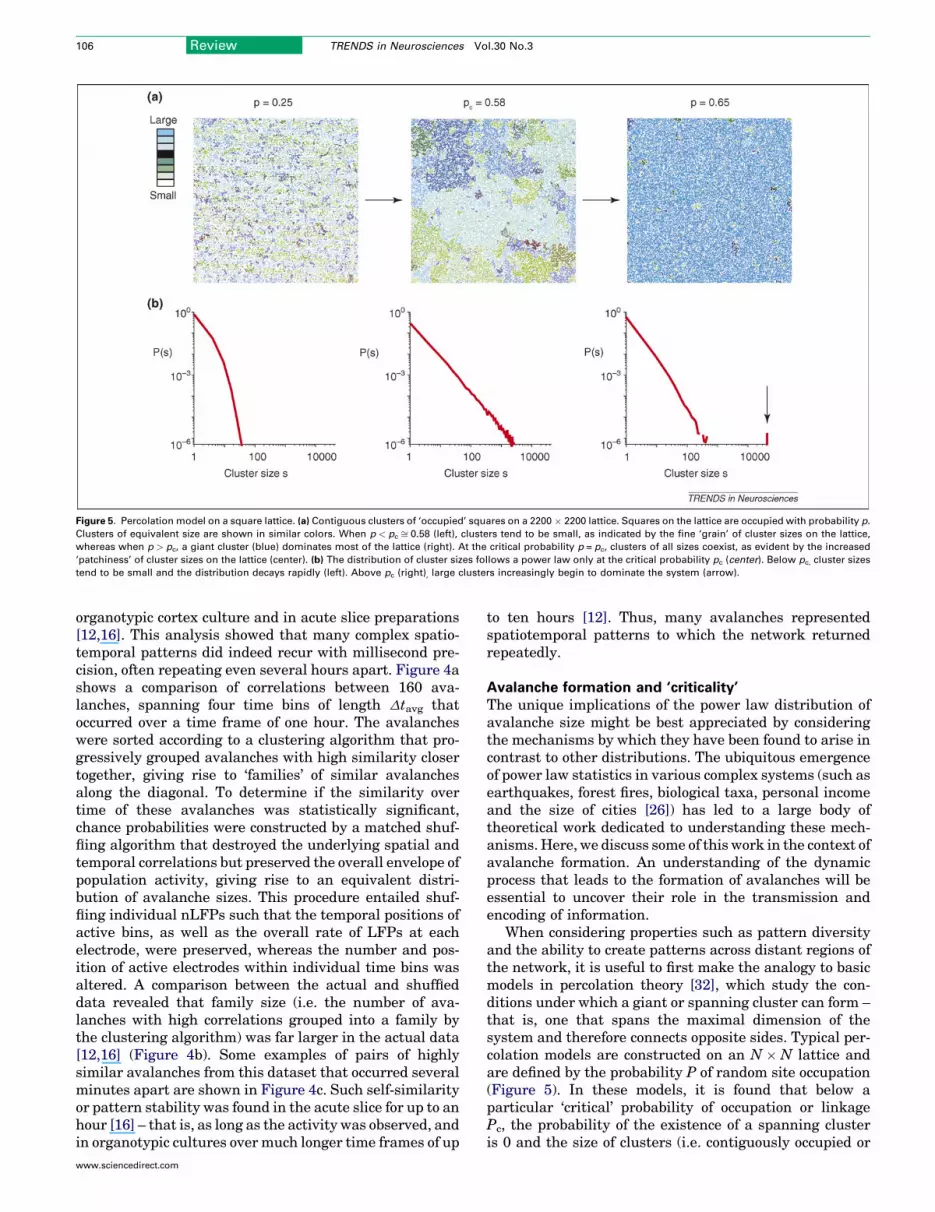
Figure 5. Percolation model on a square lattice. (a) Contiguous clusters of ‘occupied’ squares on a 2200 � 2200 lattice. Squares on the lattice are occupied with probability p.
Clusters of equivalent size are shown in similar colors. When p < pc ffi 0.58 (left), clusters tend to be small, as indicated by the fine ‘grain’ of cluster sizes on the lattice,
whereas when p > pc, a giant cluster (blue) dominates most of the lattice (right). At the critical probability p = pc, clusters of all sizes coexist, as evident by the increased
‘patchiness’ of cluster sizes on the lattice (center). (b) The distribution of cluster sizes follows a power law only at the critical probability pc (center). Below pc, cluster sizes
tend to be small and the distribution decays rapidly (left). Above pc (right), large clusters increasingly begin to dominate the system (arrow).
106 Review TRENDS in Neurosciences Vol.30 No.3
organotypic cortex culture and in acute slice preparations[12,16]. This analysis showed that many complex spatio-temporal patterns did indeed recur with millisecond pre-cision, often repeating even several hours apart. Figure 4ashows a comparison of correlations between 160 ava-lanches, spanning four time bins of length Dtavg thatoccurred over a time frame of one hour. The avalancheswere sorted according to a clustering algorithm that pro-gressively grouped avalanches with high similarity closertogether, giving rise to ‘families’ of similar avalanchesalong the diagonal. To determine if the similarity overtime of these avalanches was statistically significant,chance probabilities were constructed by a matched shuf-fling algorithm that destroyed the underlying spatial andtemporal correlations but preserved the overall envelope ofpopulation activity, giving rise to an equivalent distri-bution of avalanche sizes. This procedure entailed shuf-fling individual nLFPs such that the temporal positions ofactive bins, as well as the overall rate of LFPs at eachelectrode, were preserved, whereas the number and pos-ition of active electrodes within individual time bins wasaltered. A comparison between the actual and shuffleddata revealed that family size (i.e. the number of ava-lanches with high correlations grouped into a family bythe clustering algorithm) was far larger in the actual data[12,16] (Figure 4b). Some examples of pairs of highlysimilar avalanches from this dataset that occurred severalminutes apart are shown in Figure 4c. Such self-similarityor pattern stability was found in the acute slice for up to anhour [16] – that is, as long as the activity was observed, andin organotypic cultures over much longer time frames of up
www.sciencedirect.com
to ten hours [12]. Thus, many avalanches representedspatiotemporal patterns to which the network returnedrepeatedly.
Avalanche formation and ‘criticality’The unique implications of the power law distribution ofavalanche size might be best appreciated by consideringthe mechanisms by which they have been found to arise incontrast to other distributions. The ubiquitous emergenceof power law statistics in various complex systems (such asearthquakes, forest fires, biological taxa, personal incomeand the size of cities [26]) has led to a large body oftheoretical work dedicated to understanding these mech-anisms. Here, we discuss some of this work in the context ofavalanche formation. An understanding of the dynamicprocess that leads to the formation of avalanches will beessential to uncover their role in the transmission andencoding of information.
When considering properties such as pattern diversityand the ability to create patterns across distant regions ofthe network, it is useful to first make the analogy to basicmodels in percolation theory [32], which study the con-ditions under which a giant or spanning cluster can form –that is, one that spans the maximal dimension of thesystem and therefore connects opposite sides. Typical per-colation models are constructed on an N � N lattice andare defined by the probability P of random site occupation(Figure 5). In these models, it is found that below aparticular ‘critical’ probability of occupation or linkagePc, the probability of the existence of a spanning clusteris 0 and the size of clusters (i.e. contiguously occupied or
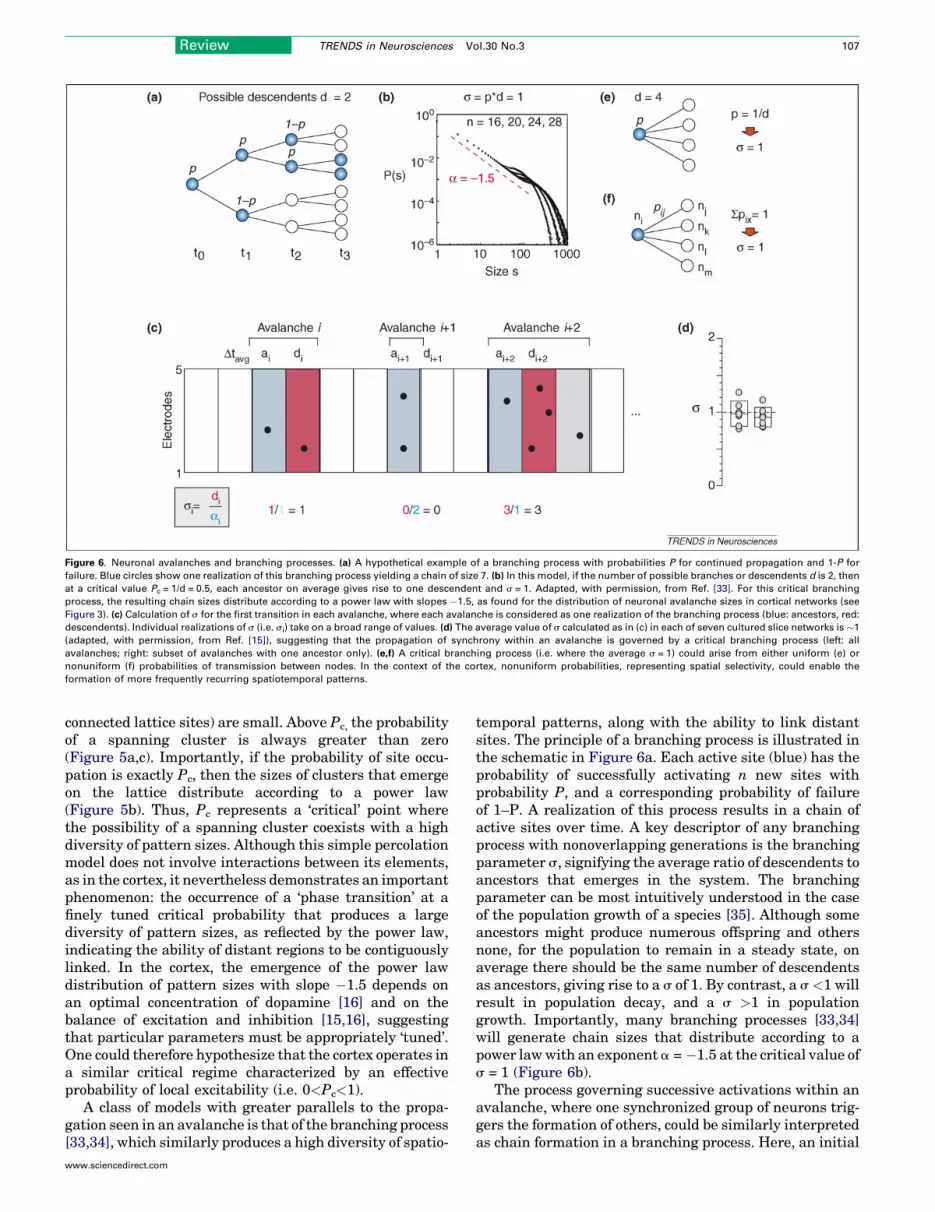
Figure 6. Neuronal avalanches and branching processes. (a) A hypothetical example of a branching process with probabilities P for continued propagation and 1-P for
failure. Blue circles show one realization of this branching process yielding a chain of size 7. (b) In this model, if the number of possible branches or descendents d is 2, then
at a critical value Pc = 1/d = 0.5, each ancestor on average gives rise to one descendent and s = 1. Adapted, with permission, from Ref. [33]. For this critical branching
process, the resulting chain sizes distribute according to a power law with slopes �1.5, as found for the distribution of neuronal avalanche sizes in cortical networks (see
Figure 3). (c) Calculation of s for the first transition in each avalanche, where each avalanche is considered as one realization of the branching process (blue: ancestors, red:
descendents). Individual realizations of s (i.e. si) take on a broad range of values. (d) The average value of s calculated as in (c) in each of seven cultured slice networks is �1
(adapted, with permission, from Ref. [15]), suggesting that the propagation of synchrony within an avalanche is governed by a critical branching process (left: all
avalanches; right: subset of avalanches with one ancestor only). (e,f) A critical branching process (i.e. where the average s = 1) could arise from either uniform (e) or
nonuniform (f) probabilities of transmission between nodes. In the context of the cortex, nonuniform probabilities, representing spatial selectivity, could enable the
formation of more frequently recurring spatiotemporal patterns.
Review TRENDS in Neurosciences Vol.30 No.3 107
connected lattice sites) are small. Above Pc, the probabilityof a spanning cluster is always greater than zero(Figure 5a,c). Importantly, if the probability of site occu-pation is exactly Pc, then the sizes of clusters that emergeon the lattice distribute according to a power law(Figure 5b). Thus, Pc represents a ‘critical’ point wherethe possibility of a spanning cluster coexists with a highdiversity of pattern sizes. Although this simple percolationmodel does not involve interactions between its elements,as in the cortex, it nevertheless demonstrates an importantphenomenon: the occurrence of a ‘phase transition’ at afinely tuned critical probability that produces a largediversity of pattern sizes, as reflected by the power law,indicating the ability of distant regions to be contiguouslylinked. In the cortex, the emergence of the power lawdistribution of pattern sizes with slope �1.5 depends onan optimal concentration of dopamine [16] and on thebalance of excitation and inhibition [15,16], suggestingthat particular parameters must be appropriately ‘tuned’.One could therefore hypothesize that the cortex operates ina similar critical regime characterized by an effectiveprobability of local excitability (i.e. 0<Pc<1).
A class of models with greater parallels to the propa-gation seen in an avalanche is that of the branching process[33,34], which similarly produces a high diversity of spatio-
www.sciencedirect.com
temporal patterns, along with the ability to link distantsites. The principle of a branching process is illustrated inthe schematic in Figure 6a. Each active site (blue) has theprobability of successfully activating n new sites withprobability P, and a corresponding probability of failureof 1–P. A realization of this process results in a chain ofactive sites over time. A key descriptor of any branchingprocess with nonoverlapping generations is the branchingparameter s, signifying the average ratio of descendents toancestors that emerges in the system. The branchingparameter can be most intuitively understood in the caseof the population growth of a species [35]. Although someancestors might produce numerous offspring and othersnone, for the population to remain in a steady state, onaverage there should be the same number of descendentsas ancestors, giving rise to a s of 1. By contrast, a s <1 willresult in population decay, and a s >1 in populationgrowth. Importantly, many branching processes [33,34]will generate chain sizes that distribute according to apower lawwith an exponent a = �1.5 at the critical value ofs = 1 (Figure 6b).
The process governing successive activations within anavalanche, where one synchronized group of neurons trig-gers the formation of others, could be similarly interpretedas chain formation in a branching process. Here, an initial

108 Review TRENDS in Neurosciences Vol.30 No.3
group of active electrodes (the ancestors) in one time bin‘gives rise’ to a group of descendents in the next generationor successive time bin. Because there might be more thanone ancestor in a time bin, it is not possible to ascertainwhich descendent arose from which ancestor. Thus, thebranching parameter for each individual avalanche si wasapproximated as the ratio of the total descendents in timebin t+1 to the total ancestors in time bin t. si was obtainedin this manner for the first generation for every avalanche(Figure 6c) [15]. By this method, the branching parameterfor the system was found to be close to 1 (0.90 � 0.19 for allavalanches; 1.04 � 0.19 for avalanches initiated by a singleancestor; Figure 6d), indicating that a local synchronousgroup on average triggers one other local synchronousgroup.
Several aspects of propagation in the cortex extendbeyond the simplicity of these models. First, in their mostsimple form, branching processes assume that each suc-cessive generation is independent of previous generations.This is not likely to be the case in neuronal networks,where the activation of one site might alter its futureparticipation in avalanche formation owing to refractori-ness or short-term plasticity. Second, uniform branchingprobabilities across all sites do not favor the recurrence ofparticular chains [36] (Figure 6e). Thus, for preferredspatiotemporal patterns to emerge from a branching pro-cess, the probabilities must differ between branches orpaths (Figure 6f). Such selectivity in the sequentialengagement of sites could reflect the wide range of synapticweights encountered at the single-neuron level.
Nonetheless, even if a branching process were to giverise to preferred spatial paths, a stochastic process mightnot be appropriate for the degree of predictability requiredfor cortical function. A third class of models that can befully deterministic in nature and provides an alternativesolution is that of self-organized criticality (SOC) [20].These models demonstrate a critical regime characterizedby a power law distribution of event sizes that evolves outof the system dynamics rather than fine-tuning of a globalcritical parameter, such as the value of Pc as discussedearlier. In SOC models, a feedback loop between a slowprocess that increases energy at lattice sites throughexternal inputs, and a fast, threshold-dependent processthat dissipates energy in the form of cascades or ava-lanches, pushes the system to a critical density of activesites, where a power law of avalanche sizes emerges [37].These models can yield power law exponents of �1.5 inaccordance with the findings on neuronal avalanches [38].When applied to the cortex, one can think of the ‘density’ asthe fraction of neurons that are active, the threshold of thefast process as the firing threshold, and the interactionsbetween sites as the rules governing transmission [38–41].In addition, several specific mechanisms have been ident-ified that enable neuronal networks to reach and maintainthe critical state. Changes in synaptic connectivityregulated by Hebbian covariance plasticity can stabilizea network of excitatory and inhibitory neurons in thecritical regime [42]. Short-term plasticity can expand thecritical regime in networks of nonleaky integrate-and-fireneurons [43]. Importantly, specific wiring rules, imple-mented either directly in the network design [44], by
www.sciencedirect.com
removing or depressing activated synapses that produceundesirable outputs (‘learning from mistakes’ [45], or aris-ing from local homeostatic regulation of neuronal inter-actions [46], can achieve critical neuronal dynamics.Finally, electrically coupled neuronal networks can alsoshow critical behavior [47]. Although these various modelssuggest how, in principle, criticality could be achieved inthe brain, their relationship with respect to neuronalavalanches has yet to be determined.
Other theoretical models of power law formationAlthough we focus here on ‘critical state’ models, we pointout the existence of other classes of models that also giverise to power law distributions of event sizes across a widerange of parameters. In these examples, the power lawsarise in different contexts that are not analogous to thedynamical process of avalanche formation but might none-theless have some conceptual underpinnings that arerelated.
One example is the Yule process [48], a class of modelsthat has been used to explain various power law distri-butions of event sizes such as in the population sizes ofcities and number of species in a genus [26]. Central tothese processes is a growth rule of ‘rich get richer’ [49],where the probability of growth of an entity or object in thesystem is proportional to its size. Such a process is notreadily applicable to dynamics because it is designedspecifically for the growth of persistent entities (such asa city) and relies on an interdependence of these entities ineach growth cycle. For example, in networks growingthrough a ‘preferential attachment’ rule [50], existingnodes compete with each other for newly formed links withincoming nodes. Importantly, Yule processes result inpower laws for event size distributions with exponentssteeper than �2, which is different from the exponent ofa = �1.5 observed for neuronal avalanches. For a moredetailed discussion of power laws as they might relate tocortical anatomy, see [51–53].
Another example is a class of models called highlyoptimized tolerance (HOT), which approaches the emer-gence of power law distributions uniquely from theperspective of system design [54,55]. HOT models arepredominantly focused on the optimization of system out-come (such as the growth of an entity) and robustness todamage, arising, for example, from environmental chal-lenges. At the optimum, the entity is often distributedaccording to a heavy tail function, such as a power law.Thesemodels require heterogeneous external initiations tocreate internal diversity, and result in systems that are notself-similar – that is, have design components that differ atdifferent spatial scales. This is different from avalanchedynamics in the cortex, where similar power law behavioremerges from each site of initiation. However, concep-tually, one could imagine that such design optimizationmight be at play in the cortex, influencing unexploredaspects of avalanche formation, such as the spatial selec-tivity of propagation.
Summary and conclusionsWe have described several features of the organizing prin-ciples of neuronal avalanches in the cortex that suggest

Review TRENDS in Neurosciences Vol.30 No.3 109
that they reflect the dynamics underlying cells assemblyformation. First, neuronal avalanches represent a richdiversity of spatiotemporal patterns with long-rangespatial correlations, as implied by their power law organ-ization. Second, these patterns are stable beyond chanceand recur with millisecond precision over many hours.Third, they arise in the superficial layers of multiplefunctional regions and form across multiple cortical col-umns with a scale-invariant organization, suggesting thatthey represent a dynamical process that can be scaled tolink distant regions. Finally, neuronal avalanches arisefrom the dynamics underlying the propagation of localsynchrony. Thus, we suggest that neuronal avalancheactivity in the cortex is a strong candidate for the repres-entation of cell assemblies in the cortex, and warrantsfurther investigation in this context. Questions of immedi-ate interest include the significance of avalanches in infor-mation processing and their relevance in vivo, particularlywith respect to cognitive outcome.
The various models described here provide insight intothe origins of these dynamics. Several models have beenproposed as explanations for the power laws that emerge innature, of which critical state models are the most relevantto avalanche dynamics. For instance, the finding of abranching parameter of 1 hints at the possibility of acritically balanced branching process that has been shownto optimize at least some aspects of information trans-mission in feedforward networks [15,56]. Alternatively,SOC and related models provide examples of how thedynamics of a system could enable it to ‘evolve’ into acritical state, possibly positioning the system for maximalcomputational ability [57]. However, to get deeper insightinto the process and its relevance to cell assembly for-mation, newmodelsmust account for additional features ofneuronal avalanches that are not captured in thesemodels,such as the stable recurrence of particular spatiotemporalpatterns and the conditions under which these patternscan be retrieved.
AcknowledgementsWe thank Drs. Jeff Diamond, David Leopold, Barry Richmond, DanteChialvo, Sinisa Pajevic, Michael Buice, and David Alderson for theirhelpful comments and discussion during the writing of this article andAgatha Monzon and Tim Bellay for critical proofreading. This work wassupported by the Intramural Research Program of the NIH, NIMH.
References1 Hebb, D.O. (1949) The Organization of Behavior, Wiley2 Lashley, K.S. (1958) Cerebral organization and behavior. Res. Publ.
Assoc. Res. Nerv. Ment. Dis. 36, 1–43 Singer, W. (1999) Neuronal synchrony: a versatile code for the
definition of relations? Neuron 24, 49–654 Varela, F. et al. (2001) The brainweb: phase synchronization and large-
scale integration. Nat. Rev. Neurosci. 2, 229–2395 Abeles, M. (1992) Corticonics, Cambridge University Press6 Diesmann, M. et al. (1999) Stable propagation of synchronous spiking
in cortical neural networks. Nature 402, 529–5337 Cateau, H. and Fukai, T. (2001) Fokker-Planck approach to the pulse
packet propagation in synfire chain. Neural Netw. 14, 675–6858 Singer, W. et al. (1990) Formation of cortical cell assemblies. Cold
Spring Harb. Symp. Quant. Biol. 55, 939–9529 deCharms, R.C. and Zador, A. (2000) Neural representation and the
cortical code. Annu. Rev. Neurosci. 23, 613–64710 Harris, K.D. et al. (2003) Organization of cell assemblies in the
hippocampus. Nature 424, 552–556
www.sciencedirect.com
11 Ikegaya, Y. et al. (2004) Synfire chains and cortical songs: temporalmodules of cortical activity. Science 304, 559–564
12 Beggs, J.M. and Plenz, D. (2004) Neuronal avalanches are diverse andprecise activity patterns that are stable for many hours in cortical slicecultures. J. Neurosci. 24, 5216–5229
13 Segev, R. et al. (2004) Hidden neuronal correlations in culturednetworks. Phys. Rev. Lett. 92, 118102
14 Eytan, D. and Marom, S. (2006) Dynamics and effective topologyunderlying synchronization in networks of cortical neurons.J. Neurosci. 26, 8465–8476
15 Beggs, J.M. and Plenz, D. (2003) Neuronal avalanches in neocorticalcircuits. J. Neurosci. 23, 11167–11177
16 Stewart, C.V. and Plenz, D. (2006) Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficiallayers of rat prefrontal cortex. J. Neurosci. 26, 8148–8159
17 Bedard, C. et al. (2006) Does the 1/f frequency scaling of brain signalsreflect self-organized critical states? Phys. Rev. Lett. 97, 118102
18 Wu, J.Y. et al. (1999) Propagating activation during oscillations andevoked responses in neocortical slices. J. Neurosci. 19, 5005–5015
19 Timofeev, I. et al. (2000) Origin of slow cortical oscillations indeafferented cortical slabs. Cereb. Cortex 10, 1185–1199
20 Bak, P. (1996) How Nature Works: The Science of Self-organizedCriticality, Copernicus, Springer-Verlag
21 Petermann, T. et al. (2006) Neuronal avalanches in vivo. Society forNeuroscience Abstracts
22 Toib, A. et al. (1998) Interaction between duration of activity and timecourse of recovery from slow inactivation in mammalian brain Na+channels. J. Neurosci. 18, 1893–1903
23 Lowen, S.B. et al. (1997) Quantal neurotransmitter secretion rateexhibits fractal behavior. J. Neurosci. 17, 5666–5677
24 Teich, M.C. et al. (1997) Fractal character of the neural spike train inthe visual system of the cat. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 14,529–546
25 Linkenkaer-Hansen, K. et al. (2001) Long-range temporal correlationsand scaling behavior in human brain oscillations. J. Neurosci. 21,1370–1377
26 Newman, M.E. (2006) Power laws, pareto distributions and Zipf’s law.arXiv: cond-mat 0412004 v3, 1-28 http://arxiv.org/PS_cache/cond-mat/pdf/0412/0412004.pdf
27 Mandelbrot, B.B. (1982) The Fractal Geometry of Nature,W.H.Freeman and Company
28 Albowitz, B. and Kuhnt, U. (1993) Spread of epileptiform potentials inthe neocortical slice: recordings with voltage-sensitive dyes. Brain Res.631, 329–333
29 Pinto, D.J. et al. (2005) Initiation, propagation, and termination ofepileptiform activity in rodent neocortex in vitro involve distinctmechanisms. J. Neurosci. 25, 8131–8140
30 Tsau, Y. et al. (1998) Initiation of spontaneous epileptiform activity inthe neocortical slice. J. Neurophysiol. 80, 978–982
31 Plenz, D. (2005) Comment on ‘Critical branching captures activity inliving neural networks and maximizes the number of metastablestates’. Phys. Rev. Lett. 95, 219801
32 Stauffer, D. and Aharoney, A. (1994) Introduction to PercolationTheory, Taylor and Francis
33 Zapperi, S. et al. (1995) Self-organized branching processes: mean-fieldtheory for avalanches. Phys. Rev. Lett. 75, 4071–4074
34 Harris, T. (1963) The theory of branching processes, Springer-Verlag35 Kimmel, M. and Axelrod, D. (2002) The Galton-Watson process, In
Branching Processes in Biology, pp. 33–61, Springer-Verlag36 Vogels, T.P. et al. (2005) Neural network dynamics. Annu. Rev.
Neurosci. 28, 357–37637 Bak, P. et al. (1988) Self-organized criticality. Phys. Rev. A. 38, 364–37438 Eurich, C.W. et al. (2002) Finite-size effects of avalanche dynamics.
Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 66, 06613739 Corral, A. et al. (1995) Self-organized criticality and synchronization in
a lattice model of integrate-and-fire oscillators. Phys. Rev. Lett. 74,118–121
40 Chen, D. et al. (1995) Self-organized criticality in a cellular automatonmodel of pulse-coupled integrate and fire neurons. J. Phys. A Math.Gen. 28, 5177–5182
41 Herz, A.V. and Hopfield, J.J. (1995) Earthquake cycles and neuralreverberations: collective oscillations in systems with pulse-coupledthreshold elements. Phys. Rev. Lett. 75, 1222–1225

110 Review TRENDS in Neurosciences Vol.30 No.3
42 Bienenstock, E. and Lehmann, D. (1998) Regulated criticality in thebrain? Adv. Complex Syst. 1, 361–384
43 Levina, A. et al. (2006) Dynamical synapses give rise to a power-lawdistribution of neuronal avalanches.Adv. Neur. Inf. Proc. Syst. 18, 771–778
44 Teramae, J-N. and Fukai, T. (2006) Local cortical circuit model inferredfrom power-law distributed neuronal avalanches. J. Comput. Neurosci,DOI: 10.1007/s10827-006-0014-6
45 Bak, P. and Chialvo, D. (2001) Adaptive learning by extremal dynamicsand negative feedback. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 63,031912
46 Abbott, L.F. and Rohrkemper, R. A simple growth model constructscritical avalanche networks. Prog. Brain Res. (in press)
47 de Arcangelis, L. et al. (2006) Self-organized criticality model for brainplasticity. Phys. Rev. Lett. 96, 028107
48 Yule, G.U. (1925) A mathematical theory of evolution based on theconclusions of Dr. J. C. Willis. Philos. Trans. R. Soc. Lond. B Biol. Sci.213, 21–87
49 Simon, H.A. (1955) On a class of skew distribution functions.Biometrika 42, 425–440
50 Barabasi, A.L. and Albert, R. (1999) Emergence of scaling in randomnetworks. Science 286, 509–512
51 Sporns, O. et al. (2004) Organization, development and function ofcomplex brain networks. Trends Cogn. Sci. 8, 418–425
Five things you might no
1.Elsevier is a founder member of the WHO’s HI
developing countries to gain free access to scientif
the Trends and Current Opinion collections and D
charge or at significan
2.The online archive of Elsevier’s premier Cell Pres
January 2005. Free access to the recent archive, i
Biology, is available on ScienceDirect and the Cell
first publ
3.Have you contributed to an Elsevier journal, book o
entitled to a 30% discount on books and stand-a
more information, cal
+1 800 782 4927 (USA) or +1 800 460 3110
or +44 (0)1865 474 010
4.Elsevier has a long tradition of liberal copyright pol
posting of preprints on public servers and the pos
Elsevier has extended its author posting policy to
their articles free of charge on their personal webs
5.The Elsevier Foundation is a knowledge-centered f
throughout the world. A reflection of our culturall
for example, funded the setting up of a video lib
provided storybooks to children in Cape Town, sp
Visiting Professorship at Brigham and Wome
International Conference on Childre
www.sciencedirect.com
52 Buzsaki, G. et al. (2004) Interneuron Diversity series: Circuitcomplexity and axon wiring economy of cortical interneurons.Trends Neurosci. 27, 186–193
53 Binzegger, T. et al. (2005) Axons in cat visual cortex are topologicallyself-similar. Cereb. Cortex 15, 152–165
54 Carlson, J.M. and Doyle, J. (1999) Highly optimized tolerance: amechanism for power laws in designed systems. Phys. Rev. E Stat.Phys. Plasmas Fluids Relat. Interdiscip. Topics 60, 1412–1427
55 Carlson, J.M. and Doyle, J. (2002) Complexity and robustness. Proc.Natl. Acad. Sci. U. S. A. 99 (Suppl. 1), 2538–2545
56 Kinouchi, O. and Copelli, M. (2006) Optimal dynamical range ofexcitable networks at criticality. Nat. Phys. 2, 348–351
57 Legenstein, R. and Maass, W. (2007) What makes a dynamical systemcomputationally powerful? In New Directions in Statistical SignalProcessing: From Systems to Brain (Haykin, S. et al., eds), pp. 127–154, MIT Press
58 Plenz, D. and Aertsen, A. (1993) Current source density profiles ofoptical recording maps: a new approach to the analysis of spatio-temporal neural activity patterns. Eur. J. Neurosci. 5, 437–448
59 Kamondi, A. et al. (1998) Dendritic spikes are enhanced by cooperativenetwork activity in the intact hippocampus. J. Neurosci. 18, 3919–3928
60 Bedard, C. et al. (2004) Modeling extracellular field potentials and thefrequency-filtering properties of extracellular space. Biophys. J. 86,1829–1842
t know about Elsevier
NARI and AGORA initiatives, which enable
ic literature. More than 1000 journals, including
rug Discovery Today, are now available free of
tly reduced prices.
s journal collection became freely available in
ncluding Cell, Neuron, Immunity and Current
Press journal sites 12 months after articles are
ished.
r series? Did you know that all our authors are
lone CDs when ordered directly from us? For
l our sales offices:
(Canada, South and Central America)
(all other countries)
icies and for many years has permitted both the
ting of final articles on internal servers. Now,
allow authors to post the final text version of
ites and institutional repositories or websites.
oundation that makes grants and contributions
y rich global organization, the Foundation has,
rary to educate for children in Philadelphia,
onsored the creation of the Stanley L. Robbins
n’s Hospital, and given funding to the 3rd
n’s Health and the Environment.