the onset of incubation in birds: can females control hatching patterns?
TRANSCRIPT

Anim. Behav., 1998, 55, 1043–1052
The onset of incubation in birds: can females control hatching patterns?
KAREN L. WIEBE, JU}RGEN WIEHN & ERKKI KORPIMA}KILaboratory of Ecological Zoology, Department of Biology, University of Turku
(Received 4 February 1997; initial acceptance 20 May 1997;final acceptance 6 August 1997; MS. number: 7843)
Abstract. We investigated intraspecific variation in incubation behaviour of wild Eurasian kestrels,Falco tinnunculus, in Finland and tested whether patterns of hatching asynchrony could be predictedfrom patterns of incubation. The timing of the onset of incubation varied considerably for 17 femalekestrels. Eggs generally hatched in the order they were laid, and both total hatching span of the clutchand the pattern of eggs hatching on certain days corresponded well with incubation behaviour. Thisresult was consistent with the idea that females have much control over hatching patterns. In themajority (65%) of cases, the proportion of daily incubation increased monotonically with the layingsequence, a pattern described previously in other birds. Unusual patterns of incubation (35%) were mostcommon in females with poor body condition during incubation and may be the result of energyconstraints during laying. ? 1998 The Association for the Study of Animal Behaviour
Because embryonic development does not typi-cally begin in avian eggs until the parent incubates(Drent 1975), birds have the ability to influencehatching patterns by varying the onset of incu-bation during laying. If incubation does not beginuntil the clutch is complete, eggs should hatchsynchronously. Conversely, incubation before thelast egg, which is common in many taxa (Clark &Wilson 1981; Ricklefs 1993), will give the earlyeggs a developmental head start so that hatchingoccurs over a period of days. Explaining theadaptive significance of hatching asynchrony hasgenerated a steady stream of hypotheses andexperiments over the last three decades (reviewsin Magrath 1990; Stoleson & Beissinger 1995;Stenning 1996). Nearly all hypotheses that pro-pose asynchrony is adaptive assume that femalescan control hatching patterns by incubationbehaviour, yet this has rarely been tested. Exper-imental manipulations of hatching asynchronyin the field have focused on the age (or size)difference between the first and last nestlingsonly, while often ignoring differences betweenintermediate nestlings in the brood.
Correspondence and present address: Karen Wiebe,Department of Biology, University of Saskatchewan,112 Science Place, Saskatoon, Saskatchewan, CanadaS7N 5E2 (email: [email protected]). J. Wiehnand E. Korpimaki are at the Department of Biology,University of Turku, FIN-20014, Turku, Finland.
0003–3472/98/041043+10 $25.00/0/ar970660 ? 1
104
It is also assumed, but infrequently confirmed,that in most avian species eggs hatch in the orderthey are laid. Hatching patterns may bear littleresemblance to incubation patterns because ofunequal distribution of heat within a clutch(Bortolotti & Wiebe 1993), or because last-laideggs require less time to hatch than early eggs(Vinuela 1991). Embryos themselves may useauditory cues to accelerate or delay their ownhatching times (Vince 1964; Schwagmeyer et al.1991). It is often inferred from activity of thefemale or from hatching times of eggs that someincubation occurs before the clutch is complete(e.g. Stouffer & Power 1990). Direct measurementof the onset of incubation in a few species,obtained by placing thermometers in nests or bylooking at embryonic development, has confirmedthat incubation usually increases graduallythroughout the laying period with partial incu-bation of early eggs (e.g. Zerba & Morton 1983;Kennamer et al. 1990; Meijer 1990; Meijer &Siemers 1993). Only two studies, however, havediscussed intraspecific variation in incubation andhow this directly relates to subsequent hatchingpatterns (see Haftorn 1981; Bortolotti & Wiebe1993).
A female bird may vary the degree of hatchingasynchrony by changing the onset of incubationduring the laying sequence and/or by changing therate at which incubation is increased during laying
998 The Association for the Study of Animal Behaviour
3
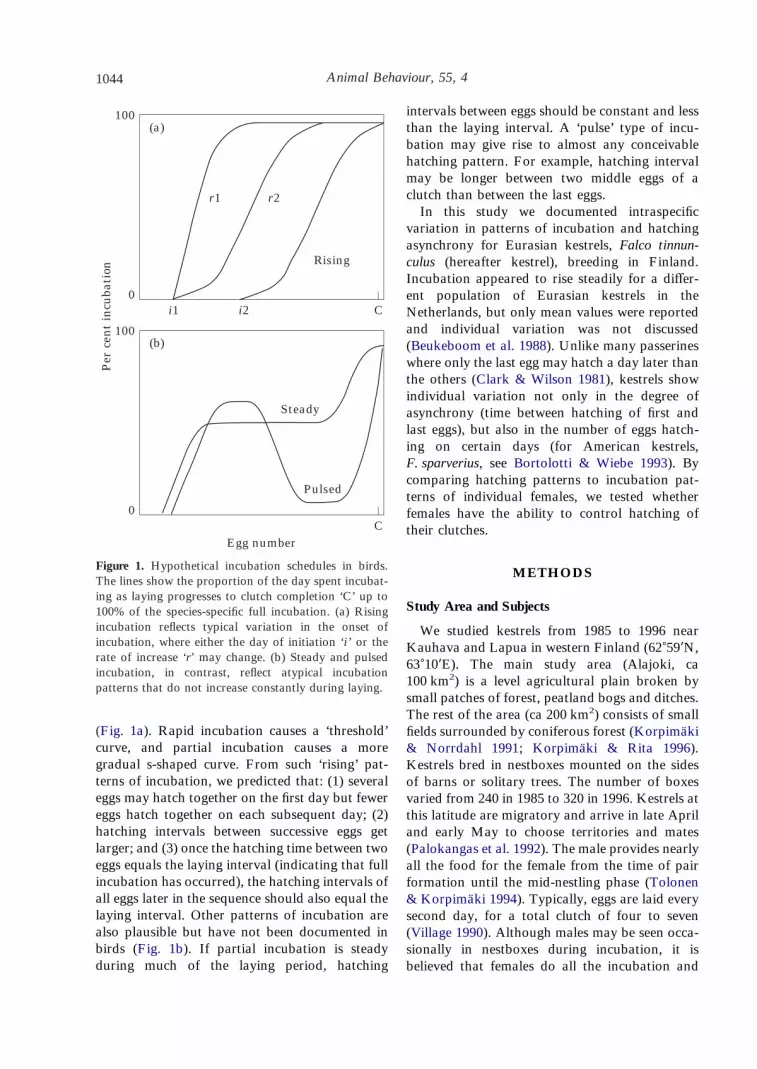
Animal Behaviour, 55, 41044
(Fig. 1a). Rapid incubation causes a ‘threshold’curve, and partial incubation causes a moregradual s-shaped curve. From such ‘rising’ pat-terns of incubation, we predicted that: (1) severaleggs may hatch together on the first day but fewereggs hatch together on each subsequent day; (2)hatching intervals between successive eggs getlarger; and (3) once the hatching time between twoeggs equals the laying interval (indicating that fullincubation has occurred), the hatching intervals ofall eggs later in the sequence should also equal thelaying interval. Other patterns of incubation arealso plausible but have not been documented inbirds (Fig. 1b). If partial incubation is steadyduring much of the laying period, hatching
intervals between eggs should be constant and lessthan the laying interval. A ‘pulse’ type of incu-bation may give rise to almost any conceivablehatching pattern. For example, hatching intervalmay be longer between two middle eggs of aclutch than between the last eggs.
In this study we documented intraspecificvariation in patterns of incubation and hatchingasynchrony for Eurasian kestrels, Falco tinnun-culus (hereafter kestrel), breeding in Finland.Incubation appeared to rise steadily for a differ-ent population of Eurasian kestrels in theNetherlands, but only mean values were reportedand individual variation was not discussed(Beukeboom et al. 1988). Unlike many passerineswhere only the last egg may hatch a day later thanthe others (Clark & Wilson 1981), kestrels showindividual variation not only in the degree ofasynchrony (time between hatching of first andlast eggs), but also in the number of eggs hatch-ing on certain days (for American kestrels,F. sparverius, see Bortolotti & Wiebe 1993). Bycomparing hatching patterns to incubation pat-terns of individual females, we tested whetherfemales have the ability to control hatching oftheir clutches.
METHODS
Study Area and Subjects
We studied kestrels from 1985 to 1996 nearKauhava and Lapua in western Finland (62)59*N,63)10*E). The main study area (Alajoki, ca100 km2) is a level agricultural plain broken bysmall patches of forest, peatland bogs and ditches.The rest of the area (ca 200 km2) consists of smallfields surrounded by coniferous forest (Korpimaki& Norrdahl 1991; Korpimaki & Rita 1996).Kestrels bred in nestboxes mounted on the sidesof barns or solitary trees. The number of boxesvaried from 240 in 1985 to 320 in 1996. Kestrels atthis latitude are migratory and arrive in late Apriland early May to choose territories and mates(Palokangas et al. 1992). The male provides nearlyall the food for the female from the time of pairformation until the mid-nestling phase (Tolonen& Korpimaki 1994). Typically, eggs are laid everysecond day, for a total clutch of four to seven(Village 1990). Although males may be seen occa-sionally in nestboxes during incubation, it isbelieved that females do all the incubation and
C
100
0
Egg number
Per
cen
t in
cuba
tion
(b)
Steady
Pulsed
C
100
0
(a)
r1
Rising
r2
i2i1
Figure 1. Hypothetical incubation schedules in birds.The lines show the proportion of the day spent incubat-ing as laying progresses to clutch completion ‘C’ up to100% of the species-specific full incubation. (a) Risingincubation reflects typical variation in the onset ofincubation, where either the day of initiation ‘i ’ or therate of increase ‘r’ may change. (b) Steady and pulsedincubation, in contrast, reflect atypical incubationpatterns that do not increase constantly during laying.

Wiebe et al.: Incubation in kestrels 1045
brooding of young (Village 1990). The breedingperformance of kestrels in our area is determinedmainly by the abundance of their major prey,Microtus voles (Korpimaki 1986), whose densitiesfluctuate more than a 100-fold in 3- to 4-yearcycles (Korpimaki & Norrdahl 1991).
From 1993 to 1996, we trapped adult femalesin the nestbox during early incubation, about3–5 days after the last egg was laid. In all years, wealso trapped about 80–95% of adults during themid-nestling period (15 days after hatching) usinga swing-door trap attached to the front of thenestbox. We banded the parents with an individ-ual combination of colour bands, and recordedtheir mass, and tarsus, tail and wing length.Adults were aged as yearling or older based on thecolour of plumage (Village et al. 1980). For anindex of body condition, we used the residualsfrom a regression of body mass and wing length(Robb et al. 1992).
Measuring Incubation and Hatching
In 1996, we placed temperature data-loggers(‘HOBO’, Onset Instruments, Massachusetts,U.S.A.) in 17 nests during the laying period. Thesesmall, electronic loggers were programmed torecord temperature every 5 min and stored 1800temperature readings on a microchip which latercould be downloaded directly into a computer.We hid the data loggers in the straw lining belowthe nest-cup so the female kestrels could not easilymove them. Because the data-logger was in thenest material, it recorded a lower temperaturethan that of the incubating female, typically about29)C. Thus, we used the steady temperaturerecorded after the last egg was laid to defineoperationally the criterion for incubation. Wecalculated the time of incubation temperature(to the nearest 30 min) for each day of thelaying period for each nest. We also placed threedata-loggers in nestboxes at different locations onthe study area as controls measuring ambienttemperature. Nests were visited for 5 min everyday or every second day during laying to recordwhen eggs were laid and to number the eggs.Because we were observing the behaviour of thefemales as part of another study, we could timeour visits to coincide with periods the birds werenaturally off the nest. Thus, the visits should nothave had much impact on natural incubationrhythms.
At the time of hatching in 1996, we visited thenests daily to record when each egg hatched. Bylooking at the degree of cracking of the eggs andthe wetness of newly hatched nestlings, we esti-mated hatching intervals to the nearest half-day(12 h; Stokland & Amundsen 1988; Vinuela 1996).When analysing hatching patterns from previousyears, we estimated days of hatching (24-h inter-vals) from the wing lengths of chicks measuredduring the first 5 days after the hatching of thelast chick, excluding nests with hatching failure.During this early period, wing growth is linearand is not influenced by gender or hatching order(Dijkstra et al. 1990), thus, wing length is areliable indicator of hatching to within 24 h.Sometimes it was difficult to represent graphicallyin the tables the precise day of hatching if, forexample, four nestlings hatched equally spacedover 48 h. In this case, we assigned two of thenestlings to ‘day 1’ and two to ‘day 2’. However,this graphic imprecision did not affect calculationsof incubation and hatching times, which weredone on a per egg basis, nor the ‘inferred incu-bation schedules’, which were based on exact winglengths. We performed statistical tests usingSPSS (1992).
RESULTS
Laying Intervals and Hatching Order
We observed 52 laying intervals at 34 nests in1996, 50 of these intervals (96%) were 2 days inlength. One interval was 3 days between the lasttwo eggs of the clutch, and another interval was7 days between the third and fourth eggs of asix-egg clutch. The latter case might be betterconsidered an interruption in laying, but the earlyeggs of the clutch remained viable and hatched inthe correct order and in a typical hatching pattern(Nestbox 866, Table I). Despite daily nest checksduring hatching at 16 nests with numbered eggs,we were usually unable to distinguish the hatchingorder of the first three eggs, which typicallyoccurred in close succession. At the five nestswhere we could determine the hatching order ofthe first three eggs, hatching order matched layingorder at two nests. At the other three nests, theeggs hatched on the same day but with the first orsecond egg hatching after the third. At all 16 nests,the hatching order of eggs 4–7 matched theirlaying order.

Animal Behaviour, 55, 41046
Patterns of Incubation
The pattern of onset of incubation during lay-ing varied among the 17 females with temperatureloggers in their nests. Sometimes full day andnight incubation began abruptly during the layingsequence consistent with the ‘threshold’ model ofincubation, but other birds showed partial incu-bation either during the day (0600–1800 hours) orduring the night. The time during the 24-h periodthat a given female incubated was not regular.Bouts of incubation tended to be longer on dayswhen eggs were laid, but incubation also occurredon days between laying. Occasionally, during boththe day and night, the temperature of eggs wasraised above ambient, but it was cooler than thenormal incubation temperature, implying thatbirds were on or near the eggs without fullyincubating them. Females with five-egg clutchesbegan incubation at a slightly earlier stage duringlaying (median stage of initiation=egg 3, N=8)than females with six-egg clutches (medianstage of initiation=egg 3.5, N=8, Mann–WhitneyU-test, P=0.047). Females typically incubated95–100% of the time (full incubation) on, orthe day after, the penultimate egg was laid buteven this varied. Four of the 17 females incubated
less than 75% of the time until the day after thelast egg.
The time spent incubating each egg duringlaying was also highly variable; we found patternsthat matched each of the hypothetical incubationschedules (Figs 1 and 2). A pattern of risingincubation on each successive egg was common(64% of females; Table I) but this rise was oftennot monotonic during the laying period (Nestbox560, Fig. 2). Such an ‘irregular rising’ curve wasusually associated with incubation on the day anegg was laid, and reduced incubation on thefollowing day when no egg was laid. When thetotal amount of incubation was summed for eachegg during the laying sequence, however, theproportion of time females spent incubatingincreased monotonically; thus, we consideredboth types of curves to be ‘rising’ incubation.Three of the 17 females had a ’steady’ onset ofincubation and two had ‘pulsed’ incubation(Table I, Fig. 2).
Incubation Versus Hatching Patterns
For the 17 females, we constructed predictedhatching patterns of eggs by summing the total
Table I. Predicted and observed hatching patterns in clutches of the Eurasian kestrel in Finland
NestboxClutch
size Incubation
Predicted pattern†
Match‡
Observed pattern†
Day 1 Day 2 Day 3 Day 4 Day 1 Day 2 Day 3 Day 4
271 5 R 3 1 0 1 * 3 1 0 1840 5 S 2 1 1 1 * 2 1 1 1560 5 R(I) 3 1 0 1 * 3 1 0 1619 5 R 2.5 1.5 0 1 ** 3 1 0 1665 5 R(I) 3 1 1 ** 2.5 1.5 1697 5 R 3 1 0 1 * 3 1 0 1425 5 R 3 1 0 1 — Hatching failure490 5 S 3 1 1 — Hatching failure454 6 S 4 1 1 * 4 1 1267 6 R 3.5 1.5 0 1 ** 3 1 1 0484 6 R 4.5 0.5 1 * 4.5 0.5 1499 6 R(I) 4 1 0 1 * 4 1 0 1699 6 R 4 1 0.5 0.5 ** 3.5 1.5 0 1866 6 S 3 1 1 1 * 3 1 1 1967 6 P 3 2 0 1 * 3 2 0 1277 6 R(I) 4 1 0 1 *** 3 1 1 1861 7 P 4 2 0 1 * 4 2 0 1
R: monotonic rising; R(I): irregular rising; S: steady; P: pulse (see text, and Fig. 2).†Numbers refer to the number of eggs hatching on a given day (day 1 of hatching is the leftmost column in eachcase). An egg hatching between 2 days is shown by ‘0.5’.
‡*Exact match, **one egg off by half a day, ***one egg off by 1 day.

Wiebe et al.: Incubation in kestrels 1047
amount of time spent incubating each egg prior tothe last egg and rounding this value off to thenearest half day. For the 15 nests where all eggshatched, the hatching pattern matched exactly thepredicted pattern in 10 of 15 (67%) cases (Table I).In four other cases, eggs hatched half a day earlieror later than expected, and in only one case did anegg hatch on a different day than predicted. Thetotal incubation time (rounded to the nearesthour) on eggs laid prior to the last egg was alsoclosely related to the hatching span (estimated tothe nearest half day) of the clutch (r2=0.91,P<0.001; Fig. 3). In general, hatching times wereslightly longer than those predicted by incubationtimes.
Assuming laying intervals of 2 days, and thathatching patterns were based on incubationpatterns during laying, we classified observedpatterns of hatching from 1985 to 1996 intothree inferred incubation patterns: ‘rising’,‘steady’ and ‘pulsed’ (see Introduction; Table II).Chi-square tests showed no difference in thefrequency of different types of inferred incu-bation between the three most common clutchsizes, four-, five- and six-egg clutches (÷2
4=4.62,N=147, P=0.32) or between the three phases ofthe vole cycle (÷2
4=2.76, N=147, P=0.59).
Smaller or inexperienced females may have haddifficulty warming all eggs equally, but inferredincubation patterns were not related to femalesize (wing length) (F2,133=0.13, P=0.87) or toage, classified as yearling versus older (÷2
2=0.47,N=134, P=0.79). Three females in the data setwere recorded in 2 years; each had ‘rising’ incu-bation in both years. The onset of incubationmight also be related to a female’s body reservesduring laying. We found a significant differencein body condition in early incubation for femaleswith different patterns of inferred incubation(F2,44=4.41, P=0.017). Bonferroni tests foundno difference in body condition between femaleswith either ‘rising’ or ‘steady’ incubation butfemales with ‘pulsed’ incubation were in poorercondition (Fig. 4). By the time females wereweighed again in the mid-nestling period, therewas no longer a relationship between theseincubation patterns and body condition(F2,127=0.99, P=0.37).
The condition of females in early incubationwas also negatively correlated with laying date(r2= "0.35, N=45, P=0.02) and laying datetended to differ between the three hatching pat-terns (F2,135=2.72, P=0.07). In particular, birdsthat showed ‘steady’ incubation tended to haveearlier laying dates than birds showing ‘rising’ or
6
100
02
Egg number
Per
cen
t in
cuba
tion
80
60
40
20
3 4 5
Figure 2. Some observed patterns of incubation forEurasian kestrels breeding in Finland. The proportion ofthe day spent incubating is shown for each day of thelaying sequence. Each nestbox had five eggs except for967 which had six. Nestbox 967 had ‘pulsed’ incubation,nestbox 840 had ‘steady’ incubation and the othernestboxes had ‘rising’ incubation. ——: Nestbox 560;· · ·: nestbox 271; - - -: nestbox 619; — — —: nestbox840; — - —: nestbox 967.
3.5
3.5
1Incubation (days)
Hat
chin
g sp
an (
days
)
3
2.5
2
1.5
1.5 2 2.5 3
Figure 3. The relationship between the total amount ofincubation prior to the last egg and the hatching spanfor 15 clutches of Eurasian kestrels. The solid linerepresents a perfect correspondence between incubationand hatching times.

TE
R(
S(
P(
OP
Animal Behaviour, 55, 41048
‘pulsed’ incubation patterns. The relationshipbetween body condition and incubation patternwas probably not a result of laying date per se,however, because when the ‘steady’ incubationgroup was removed from the analysis, there wasno difference in mean laying date between ‘rising’and ‘pulse’ incubation patterns but there was stilla significant difference in body condition betweenthese two groups.
To see possible effects of hatching patterns weanalysed nestling mortality. At the 17 nests in1996, a poor food year with harsh springweather, at least one nestling died at all nests.In the larger data set of inferred incubationpatterns, 80% of nests with ‘rising’ incubationpatterns had mortality compared with only 55and 50% of ‘steady’ and ‘pulse’ nests, respect-ively. A logistic regression indicated that of thethree incubation patterns, there was a greaterprobability that at least one nestling would diein nests with ‘rising’ incubation (Wald=4.55,df=1, P=0.032).
DISCUSSION
Only a few studies have examined intraspecificvariation for incubation during laying, but itseems to exist for altricial (Morton & Pereyra1985) and semi-altricial birds (Bortolotti & Wiebe1993). There was considerable variation in thepattern of onset of incubation between individualkestrels in our population. Both the onset ofincubation and the rate of increase of incubationduring the laying sequence differed betweenfemales, even with the same clutch size (Fig. 2).Some females even delayed full incubation until1–2 days after clutch completion. Althoughfemales in our population with six-egg clutchesinitiated incubation 1 day later, on average, rela-tive to the laying of the first egg than females withfive-egg clutches, hatching spans increased withclutch size (unpublished data). This result suggeststhat the onset of incubation in birds is not necess-arily fixed relative to the laying of the last egg(cf. Beukeboom et al. 1988).
able II. Frequency of the most common hatching patterns in five- (N=52) and six-egg (N=74) clutches of theurasian kestrel from 1985 to 1996
Day of hatching for:
Clutch size of 6 Clutch size of 5
1 2 3 4 5 1 2 3 4 5
ising55%) (56%)
12.1 ,,, ,, — , — 11.5 ,, ,, , — —8.1 ,,,, , — , — 9.6 ,, ,, — ,8.1 ,,, ,, , — — 7.7 ,,, — , — ,6.7 ,,, , , — , 7.7 ,,,, , — — —5.4 ,,,,, , — — — 5.7 ,,, , — , —2.7 ,, ,, , , — 5.7 ,,,, — , — —
teady16%) (27%)
9.5 ,,, , , , — 19.2 ,,, , , — —4.1 ,,,, , , — — 7.7 ,, , , , —2.7 ,, , , , ,
ulse28%) (17%)
4.1 ,, ,,, , — — 13.0 ,,, ,, — — —4.1 ,,, , — , ,2.7 ,,, — ,, — ,2.7 ,, ,,, — , —2.7 ,,,, ,, — — —
nly patterns that were observed more than once are shown, so percentages do not necessarily sum to total per cent.atterns are classified as ‘rising’, ‘steady’ or ‘pulse’ (see Methods, Fig. 1).

Wiebe et al.: Incubation in kestrels 1049
Proximate Effects on Incubation
For the majority of female Eurasian kestrels(55–65%; Tables I and II), incubation timeincreased as successive eggs were laid, which isconsistent with the pattern observed in otherspecies (e.g. Parsons 1972; Haftorn 1978, 1981;Meijer 1990). The onset of incubation in birds isprobably controlled, at least at a coarse level, byhormones. An increase in prolactin during layingseems to initiate broodiness and increase incu-bation (Goldsmith & Williams 1980; Balthazart1983) but the role of prolactin or other hor-mones is not clear and may vary between species(Goldsmith 1983). We also found some unusualincubation patterns (‘steady’ or ‘pulsed’ duringlaying), that have not been previously describedfor birds. Such intraspecific variation in the onsetof incubation suggests that hormonal controls arenot rigid (cf. Mead & Morton 1985) but may beinfluenced by physical characteristics of thefemale, or by the environment. Thus, for ourpopulation, it is unlikely that the act of incubationitself is the fine-tuned proximate mechanism thatterminates laying as proposed by Meijer et al.(1990), although a general relationship mayexist between rising prolactin levels, increasingincubation and clutch termination.
Incubation behaviour may be influenced bypatterns of food supply during laying. In
Fennoscandia, numbers of voles cycle with 50- to150-fold differences in density between the highand low phases (Hanski et al. 1993). In theNetherlands where prey densities within andbetween years are more stable (Dijkstra et al.1988, figure 6), the relatively predictable con-ditions may facilitate regular and predictablelaying and incubation patterns (see Meijer et al.1990). The year that we measured incubation inFinland (i.e. 1996) was a low vole year with harshspring weather, so energy constraints during lay-ing may have been particularly severe, limiting theability of females to incubate without interrup-tion. Similarly, in food supplementation exper-iments, the timing of the onset of full incubationrelative to the last egg in blue tits, Parus caeruleus,was constrained by energy and depended on foodsupply (Nilsson 1993). For species such as thekestrel where the male feeds the female duringlaying (Village 1990), the male’s hunting perform-ance may be the proximate factor most closelylinked to incubation rhythms.
Females’ Control of Hatching Patterns
Hatching patterns are the result of both layingand incubation schedules; the laying interval isthe maximum possible hatching time betweeneggs. Laying intervals may vary within species(Beissinger & Waltman 1991; Watson et al. 1993)and may be longer in individuals with a poor foodsupply during laying (Nilsson & Svensson 1993;Wiebe & Martin 1995). Laying intervals in ourpopulation, even in a low vole year when femalesmay have had difficulty gathering resources forforming eggs, were almost invariably 2 days. Thisresult is consistent with other reports for thespecies (Newton 1979; Village 1990), but Aparicio(1994) found more variation in Spanish kestrels;only 75% of 66 intervals were 2 days. Althoughfood may affect the speed at which eggs are laid,laying rates do not necessarily affect subsequenthatching patterns. Females in our population didnot incubate continuously during laying gaps;consequently, eggs laid 3 and 7 days apart hatchedonly 1 day apart.
Females appeared to have a good ability tocontrol both the hatching span (asynchrony) oftheir clutch (Fig. 3) and the specific pattern ofeggs hatching on certain days (Table I); eggsgenerally hatched in the order they were laid,except for some minor shifts between the first
Steady
30
–20Pulse
Type of incubation
Fem
ale
con
diti
on
Rising
20
10
0
–10
11
29
5
Figure 4. Mean& body condition of female Eurasiankestrels in early incubation in relation to the hatchingpattern of their clutches (1993–1996). Sample sizes areshown above the bars.

Animal Behaviour, 55, 41050
three eggs, which hatched on the same day. Half-day discrepancies between predicted and observedhatching patterns in Table I were within theamount of error for our estimates of egg-layingtimes. Our recorded incubation times (Fig. 3) mayhave been slightly less, on average, than the actualhatching times because some development mayhave occurred at temperatures below the incu-bation temperature. Perhaps there was also somelag time before high temperatures were recordedby the temperature data-loggers. In a closelyrelated species, the American kestrel, even eggslaid late in the laying sequence do not alwayshatch in the order they are laid, and females thatare smaller than average have irregular incubationpatterns like the ‘pulse’ patterns observed inthis study (Bortolotti & Wiebe 1993). Americankestrels have the highest ratio of clutch volume tofemale mass (11.6%) of any raptor species inNewton’s (1979) extensive review and higherthan Eurasian kestrels (9.5%). Based on theseresults, Bortolotti & Wiebe (1993) and Wiebe& Bortolotti (1993) hypothesized that body sizeconstrained hatching patterns for Americankestrels. We did not find such a relationshipbetween hatching patterns and female body size inthis study, so size is unlikely to be a constraint forEurasian kestrels.
Instead, food reserves during incubation mayaffect hatching patterns of Eurasian kestrels.Females with irregular hatching patterns consist-ent with ’pulsed’ incubation during laying werein the poorest condition shortly after completingtheir clutch, suggesting that a shortage of energy(food) during laying limited their ability toachieve uninterrupted incubation. Unlike manypasserines (e.g. Slagsvold & Lifjeld 1989; Nilsson1993), female kestrels do almost no hunting forthemselves during laying (Village 1990), so thereshould be little direct conflict about whether toincubate or to forage. It may simply be tooenergetically costly to incubate and form eggsat the same time. Based on values reported inMeijer et al. (1989), adding the cost of eggformation to the cost of incubation alone isan increase of about 24% in daily energyexpenditure.
Significance of Hatching Patterns
Nearly all hypotheses and field experiments onhatching asynchrony have considered only the
time interval between the hatching of first andlast egg while ignoring possible consequences ofdifferent hatching patterns. We speculate thathatching patterns may have different, adaptivefunctions related to growth and survival of nest-lings. For example, ‘steady’ incubation whichspaces nestlings evenly, might be most relevantfor the peak-load reduction hypothesis (Hussell1972). When considering the brood-reductionhypothesis (Lack 1954), hatching four nestlings onthe same day and one nestling 2 days later mayhave different consequences for the efficiency ofbrood reduction than hatching three nestlings onthe same day and one nestling on each subsequentday. We predict that the last nestling would diemore readily in the first case because there is moretime between the ultimate and penultimatenestling. Logistic regression showed that the over-all probability of nestling mortality was greaterfor ‘rising’ incubation patterns compared with‘pulse’ incubation patterns perhaps because of thegreater spacing between last nestlings and a largegroup of older nestlings. However, the results arepreliminary as we were unable to control for otherimportant factors such as food supply andbody condition of the adults while provisioningyoung.
It is also important to remember that hatchingpatterns may not be adaptive but that incubationrhythms may be selected during the laying periodfor other reasons including energy savings or eggviability (Wiebe & Bortolotti 1994; Stoleson &Beissinger 1995). Some eggs in one nest with‘rising’ incubation and one nest with ‘steady’incubation failed to hatch (Table I), but no con-clusions about the effect of incubation rhythm onhatching success can be made with this smallsample.
Although a strong relationship existed betweenincubation patterns and hatching patterns in ourkestrel population, it is still unclear whether suchpatterns were adaptive. Hatching spans increasedwith female age and decreased in low voleyears (unpublished data) but the specific patternsof hatching did not vary with the vole cycle orwith the age of the parents. Constraints duringlaying seem to be the most parsimonious expla-nation for ‘pulsed’ incubation patterns, but thecauses and consequences of the wide variety of‘rising’ and ‘steady’ incubation patterns in kestrelsand in other birds needs to be investigatedexperimentally.

Wiebe et al.: Incubation in kestrels 1051
ACKNOWLEDGMENTS
Seppo Neuvonen, Steven Beissinger, PatriciaSchwagmeyer and an anonymous referee madevaluable comments on the manuscript. We aregrateful to Kathy Martin for lending us thedata-loggers used in this study. We also thankPasi Tolonen, Mikko Hast, Mikko Hanninen,Ossi Hemminki and Timo Hyrsky for help incollecting data and Sakari Ikola for logisticalassistance in the field. The study was funded bythe Academy of Finland (to E.K.), the Center forInternational Mobility, Ministry of Education,Finland (to K.W.) and the Emil AaltonenFoundation (to J.W.).
REFERENCES
Aparicio, J. M. 1994. The effect of variation in the layinginterval on proximate determination of clutch size inthe European Kestrel. J. Avian Biol., 25, 275–280.
Balthazart, J. 1983. Hormonal correlates of behaviour.In: Avian Biology, Vol. 7 (Ed. by D. S. Farner, J. R.King & K. C. Parkes), pp. 221–365. New York:Academic Press.
Beissinger, S. R. & Waltman, J. R. 1991. Extraordinaryclutch size and hatching asynchrony of a neotropicalparrot. Auk, 108, 863–871.
Beukeboom, L., Dijkstra, C., Daan, S. & Meijer, T.1988. Seasonality of clutch size determination in thekestrel, Falco tinnunculus: an experimental study.Ornis Scand., 19, 41–48.
Bortolotti, G. R. & Wiebe, K. L. 1993. Incubationbehaviour and hatching patterns in the Americankestrel Falco sparverius. Ornis Scand., 24, 41–47.
Clark, A. B. & Wilson, D. S. 1981. Avian breedingadaptations: hatching asynchrony, brood reduction,and nest failure. Q. Rev. Biol., 56, 253–277.
Dijkstra, C., Daan, S., Meijer, T., Cave, A. J. & Foppen,R. P. B. 1988. Daily and seasonal variation in bodymass of the kestrel in relation to food availability andreproduction. Ardea, 76, 127–140.
Dijkstra, C., Bult, A., Bijlsma, S., Daan, S., Meijer, T. &Zijlstra, M. 1990. Brood size manipulations in thekestrel (Falco tinnunculus): effects on offspring andparent survival. J. Anim. Ecol., 59, 269–285.
Drent, R. 1975. Incubation. In: Avian Biology, Vol. 5(Ed. by D. S. Farner & J. R. King), pp. 333–420. NewYork: Academic Press.
Goldsmith, A. R. 1983. Prolactin in avian reproductivecycles. In: Hormones and Behaviour in Higher Verte-brates (Ed. by J. Balthazart, E. Prove & R. Gilles),pp. 375–387. Berlin: Springer–Verlag.
Goldsmith, A. R. & Williams, D. M. 1980. Incubation inmallards (Anas platyrhynchos): changes in plasmalevels of prolactin and luteinizing hormone. J. Endo-crinol., 86, 371–379.
Haftorn, S. 1978. Egg laying and regulation of eggtemperature during incubation in the goldcrestRegulus regulus. Ornis Scand., 9, 2–21.
Haftorn, S. 1981. Incubation during the egg layingperiod in relation to clutch size and other aspects ofreproduction in the great tit (Parus major). OrnisScand., 12, 169–185.
Hanski, I., Turchin, P., Korpimaki, E. & Henttonen, H.1993. Population oscillations of boreal rodents: regu-lation by mustelid predators leads to chaos. Nature,Lond., 364, 232–235.
Hussell, D. J. T. 1972. Factors affecting clutch size inarctic passerines. Ecol. Monogr., 42, 317–364.
Kennamer, R. A., Harvey, W. F. & Hepp, G. R. 1990.Embryonic development and nest attentiveness ofwood ducks during egg laying. Condor, 92, 587–592.
Korpimaki, E. 1986. Predation causing synchronousdecline phases in microtine and shrew populations inwestern Finland. Oikos, 46, 124–127.
Korpimaki, E. & Norrdahl, K. 1991. Numerical andfunctional responses of kestrels, short-eared owls andlong-eared owls to vole densities. Ecology, 72, 814–826.
Korpimaki, E. & Rita, H. 1996. Effects of brood sizemanipulations on offspring and parental survivalin the European kestrel under fluctuating foodconditions. Ecoscience, 3, 264–273.
Lack, D. 1954. The Natural Regulation of AnimalNumbers. Oxford: Clarendon Press.
Magrath, R. D. 1990. Hatching asynchrony in altricialbirds. Biol. Rev., 95, 587–622.
Masman, D., Daan, S. & Beldhuis, J. A. 1988. Ecologi-cal energetics of the kestrel: daily energy expenditurethroughout the year based on the time-budget, foodintake and doubly labelled water methods. Ardea, 74,64–81.
Mead, P. S. & Morton, M. L. 1985. Hatching asyn-chrony in the mountain white-crowned sparrow(Zonotrichia leucophrys oriantha): a selected orincidental trait? Auk, 102, 781–782.
Meijer, T. 1990. Incubation development and clutch sizein the starling. Ornis Scand., 21, 163–168.
Meijer, T. & Siemers, I. 1993. Incubation developmentand asynchronous hatching in junglefowl. Behaviour,127, 309–322.
Meijer, T., Masman, D. & Daan, S. 1989. Energetics ofreproduction in female kestrels. Auk, 106, 549–559.
Meijer, T., Daan, S. & Hall, M. 1990. Family planningin the kestrel (Falco tinnunculus): the proximate con-trol of covariation of laying date and clutch size.Behaviour, 114, 117–136.
Morton, M. L. & Pereyra, M. E. 1985. The regulation ofegg temperatures and attentiveness patterns in thedusky flycatcher (Empidonax oberholseri). Auk, 102,25–37.
Newton, I. 1979. Population Ecology of Raptors.Vermillion, South Dakota: Buteo Books.
Nilsson, J.-Ar . 1993. Energetic constraints on hatchingasynchrony. Am. Nat., 141, 158–166.
Nilsson, J.-Ar . & Svensson, E. 1993. The frequency andtiming of laying gaps. Ornis Scand., 24, 122–126.
Palokangas, P., Alatalo, R. V. & Korpimaki, E. 1992.Female choice in the kestrel under different avail-ability of mating options. Anim Behav., 43, 659–665.

Animal Behaviour, 55, 41052
Parsons, J. 1972. Egg size, laying date and incubationperiod in the herring gull. Ibis, 114, 536–541.
Ricklefs, R. E. 1993. Sibling competition, hatchingasynchrony, incubation period and life span inaltricial birds. Curr. Ornithol., 11, 199–276.
Robb, L. A., Martin, K. & Hannon, S. J. 1992. Springbody condition, fecundity and survival in femalewillow ptarmigan. J. Anim. Ecol., 61, 215–223.
Schwagmeyer, P. L., Mock, D. W., Lamey, T. C. &Beecher, M. D. 1991. Effects of sibling contact onhatch timing in an asynchronously hatching bird.Anim. Behav., 41, 887–894.
Slagsvold, T. & Lifjeld, J. T. 1989. Constraints onhatching asynchrony and egg size in pied flycatchers.J. Anim. Ecol., 58, 837–845.
SPSS 1992. SPSS for Windows: Base System User’sGuide. Release 5.0. Chicago: SPSS.
Stenning, M. J. 1996. Hatching asynchrony, broodreduction and other rapidly reproducing hypotheses.Trends Ecol. Evol., 11, 243–246.
Stokland, J. N. & Amundsen, T. 1988. Initial sizehierarchy in broods of the shag: relative significanceof egg size and hatching asynchrony. Auk, 105,308–315.
Stoleson, S. H. & Beissinger, S. R. 1995. Hatchingasynchrony and the onset of incubation in birds,revisited: when is the critical period? Curr. Ornithol.,12, 191–270.
Stouffer, P. C. & Power, H. W. 1990. Density effectson asynchronous hatching and brood reduction inEuropean starlings. Auk, 107, 359–366.
Tolonen, P. & Korpimaki, E. 1994. Determinants ofparental effort: a behavioral study in the Eurasiankestrel, Falco tinnunculus. Behav. Ecol. Sociobiol., 4,435–441.
Village, A. 1990. The Kestrel. London: Poyser Press.Village, A., Marquiss, M. & Cook, D. C. 1980. Moult,
aging and sexing of kestrels. Ring. Migr., 3, 53.Vince, M. A. 1964. Social facilitation of hatching in the
bobwhite quail. Anim. Behav., 12, 531–534.Vinuela, J. 1991. Ecologia reproductiva del milano
negro, Milvus migrans, en el Parque Nacional deDonana. Ph.D. thesis, Universidad Complutense deMadrid.
Vinuela, J. 1996. Establishment of mass hierarchies inbroods of the black kite. Condor, 98, 93–99.
Watson, M. D., Robertson, G. J. & Cooke, F. 1993.Egg-laying time and laying interval in the CommonEider. Condor, 95, 869–878.
Wiebe, K. L. & Bortolotti, G. R. 1993. Brood patchesof American kestrels: an ecological evolutionaryperspective. Ornis Scand., 24, 197–204.
Wiebe, K. L. & Bortolotti, G. R. 1994. Food supplyand hatching spans of birds: energy constraints orfacultative manipulation? Ecology, 75, 813–823.
Wiebe, K. L. & Martin, K. 1995. Ecological and physio-logical effects on egg laying intervals in ptarmigan.Condor, 97, 708–717.
Zerba, E. & Morton, M. L. 1983. The rhythm ofincubation from egg laying to hatching in mountainwhite-crowned sparrows. Ornis Scand., 14, 188–197.