the intraparietal cortex: subregions involved in fixation, saccades, and in the visual and...
TRANSCRIPT

The Intraparietal Cortex: Subregions Involved in Fixation,Saccades, and in the Visual and Somatosensory Guidance
of Reaching
*Georgia G. Gregoriou and *†Helen E. Savaki
*Department of Basic Sciences, Division of Medicine, School of Health Sciences, University of Crete, and †Institute of Appliedand Computational Mathematics, Foundation for Research and Technology, Hellas, Greece
Summary: The functional activity of the intraparietal cortexwas mapped with the [14C]deoxyglucose method in monkeysperforming fixation of a central visual target, saccades to visualtargets, reaching in the light during fixation of a central visualtarget, and acoustically triggered reaching in the dark while theeyes maintained a straight ahead direction. Different subregionsof the intraparietal cortical area 7 were activated by fixation,saccades to visual targets, and acoustically triggered reachingin the dark. Subregions in the ventral part of the intraparietalcortex (around the fundus of the intraparietal sulcus) were ac-tivated only during reaching in the light, in which case visual
information was available to guide the moving forelimb. Incontrast, subregions in the dorsal part of the intraparietal cor-tical area 5 were activated during both reaching in the lightand the dark, in which cases somatosensory information wasthe only one available in common. Thus, visual guidance ofreaching is associated with the ventral intraparietal cortex,whereas somatosensory guidance, based on proprioceptive in-formation about the current forelimb position, is associatedwith dorsal intraparietal area 5. Key Words: [14C]deoxyglu-cose method—Arm movements—Eye movements—Monkey—Parietal cortex.
Accurate reaching toward an object of interest requiresa precise neural representation of target location relativeto the body. Visual and somatosensory information isused to compute the spatial location of objects, whereasinformation about our own movements constantly up-dates the distance between the object and the movingforelimb. Elucidation of the brain areas involved in pro-cessing sensorimotor information used for the guidanceof reaching may provide an insight into the strategies thebrain uses to guide voluntary movements.
Hierarchically located in between the visual and motorareas, the posterior parietal lobe receives convergent in-put from different sensory modalities as well as efferencecopy signals from motor areas to guide eye and forelimbmovements (Kalaska, 1996; Andersen et al., 1997; Colby
and Goldberg, 1999). Neurophysiologic and metabolicstudies have demonstrated that area 7, in the lateral bankof the intraparietal sulcus (IPs), displays visual activity(Blatt et al., 1990; Colby et al., 1995) and helps programsaccades to visual, memorized, and auditory targets(Gnadt and Andersen, 1988; Andersen et al., 1990;Colby et al., 1995; Mazzoni et al., 1996). Area 5, in themedial bank of IPs, displays visual (Colby and Duhamel,1991; Savaki et al., 1993), somatosensory (Savaki et al.,1993; Duhamel et al., 1998), and skeletomotor activity(Georgopoulos and Massey, 1988; Kalaska et al., 1990)and guides reaching by maintaining a representation ofthe forelimb position based on proprioceptive feedbackand motor efference information (Kalaska et al., 1983;Georgopoulos et al., 1984).
Goal-directed reaching is usually accomplished by aninitial saccadic eye movement to foveate the target ofinterest, followed by a subsequent arm movement towardthe same target. One aim of the current study was todissociate the cortical regions in IPs which are involvedin the control of eye movements from those involved inthe control of forelimb movements. To this end, the[14C]deoxyglucose (14C-DG) quantitative autoradio-graphic method (Sokoloff et al., 1977) was used to mapthe metabolic activity in the brain of monkeys perform-
Received November 20, 2000; final revision received January 25,2001; accepted January 25, 2001.
Supported by the Human Frontier Science Program grantRG0039/1988-B. G. Gregoriou was supported by the Greek GeneralSecretariat of Research and Technology, grant ∏ENE� 95E�24.
Address correspondence and reprint requests to Helen E. Savaki,Department of Basic Sciences, Division of Medicine, School of HealthSciences, University of Crete, P.O. Box 1393, GR-71110, Iraklion,Crete, Greece.
Journal of Cerebral Blood Flow and Metabolism21:671–682 © 2001 The International Society for Cerebral Blood Flow and MetabolismPublished by Lippincott Williams & Wilkins, Inc., Philadelphia
671

ing fixation of a visual target, saccades to visual targets,and reaching of a peripheral target in the light duringfixation of a central visual target. The results of the firsttwo experiments demonstrated that distinct subregions inthe lateral intraparietal area 7 (7IP) are related to visualfixation and to saccadic eye movements. The results ofthe third experiment demonstrated that subregions in thedorsal part of the medial intraparietal area 5 (5DIP) andin the ventral intraparietal cortex around the fundus ofIPs (areas 5VIP and 7VIP) are related to reaching withone forelimb in the light.
Another important problem is the dissociation be-tween visual and somatosensory information used toguide reaching movements. In previous experiments, theauthors used the 14C-DG method to map the local cere-bral metabolic activity, during the performance of a vi-sually cued reaching task, in visually intact monkeys andin monkeys with one hemisphere deprived of all knownvisual input by optic chiasm section and commissuroto-my (Savaki et al., 1993). The different activations in-duced by the same behavior in the visually intact and thevisually deprived monkeys led the authors to proposethat the dorsal part of intraparietal area 5 is associatedwith the proprioceptive guidance of the moving forelimb,whereas the ventral part of the same area is related to thevisual guidance of reaching. To test this hypothesis, theauthors performed an additional experiment in the cur-rent study, in which the monkey had to reach from acentral to a peripheral position in complete darknesswhile its eyes maintained a straight-ahead direction.Only somatosensory information is available to guide theforelimb during reaching in the dark (RD). In contrast,both visual and somatosensory information is availableto guide the forelimb during reaching in the light (RL).Therefore, comparison of the distribution of activity inthe IPs of the RD and the RL monkeys should allow fordissociation between subregions responsible for the so-matosensory and the visual guidance of reaching. Ac-cording to the authors’ earlier proposal (Savaki et al.,1993), they predicted that only the dorsal intraparietalarea 5 (5DIP) would be activated in the RD monkey,whereas the ventral intraparietal cortex (areas 5VIP and7VIP) would be activated only in the RL monkey. In thecurrent article, the authors provide evidence to supportthese predictions and consequently their earlier hypoth-esis. Furthermore, the distinct intraparietal cortical sub-regions that are involved in visual fixation, visuallyguided saccades, reaching in the light, and reaching inthe dark are demonstrated for the first time in metabolicmaps of high resolution.
MATERIALS AND METHODS
Experiments were performed in accordance with institutionalguidelines complied with Greek and EU laws for the Care andUse of Laboratory Animals.
SubjectsFive adult female Macaca mulatta monkeys weighing be-
tween 3 and 4 kg were used. One monkey was an untrainedcontrol, whereas the other four monkeys were trained to per-form four different tasks. All five monkeys had their headsfixed and a water delivery tube attached close to their mouth.For immobilization of the head, a metal bolt embedded in den-tal cement was surgically implanted on the head of each mon-key with the use of mandibular plates that were secured on thebone by titanium screws (Synthes). Eye movements in the fourexperimental monkeys were recorded with implanted scleralsearch coils (Robinson, 1963). All surgical procedures wereperformed under general anesthesia (ketamine hydrochloride20 mg/kg, intramuscularly) using aseptic techniques. Eye po-sition was digitized online at a sampling rate of 500 Hz usingthe Spike2 software (Cambridge Electronics Design; Cam-bridge, U.K.). Saccades and reaching movements were made toa video (touch-sensitive) screen placed 23 cm in front of eachmonkey. Monkeys could not move their forelimbs during thefixation and saccade tasks and had to maintain fixation duringthe reaching tasks. Visual targets were red round illuminatedspots of 1.5° diameter. Successful completion of each trial wasrewarded with water.
Behavioral tasksTo use the smallest possible number of monkeys, the five
behavioral tasks used in the current study were designed so thatcontrolled behavioral parameters were gradually added. Thisway, all tasks were complementary to each other, and the majormetabolic findings in each monkey per task were verified bythose in the monkey of the following, more complex task (seeResults for more detailed explanation).
The first monkey was an untrained control, seated in front ofthe nonfunctioning behavioral apparatus and receiving neithervisual stimuli nor liquid reward during the 14C-DG experiment.
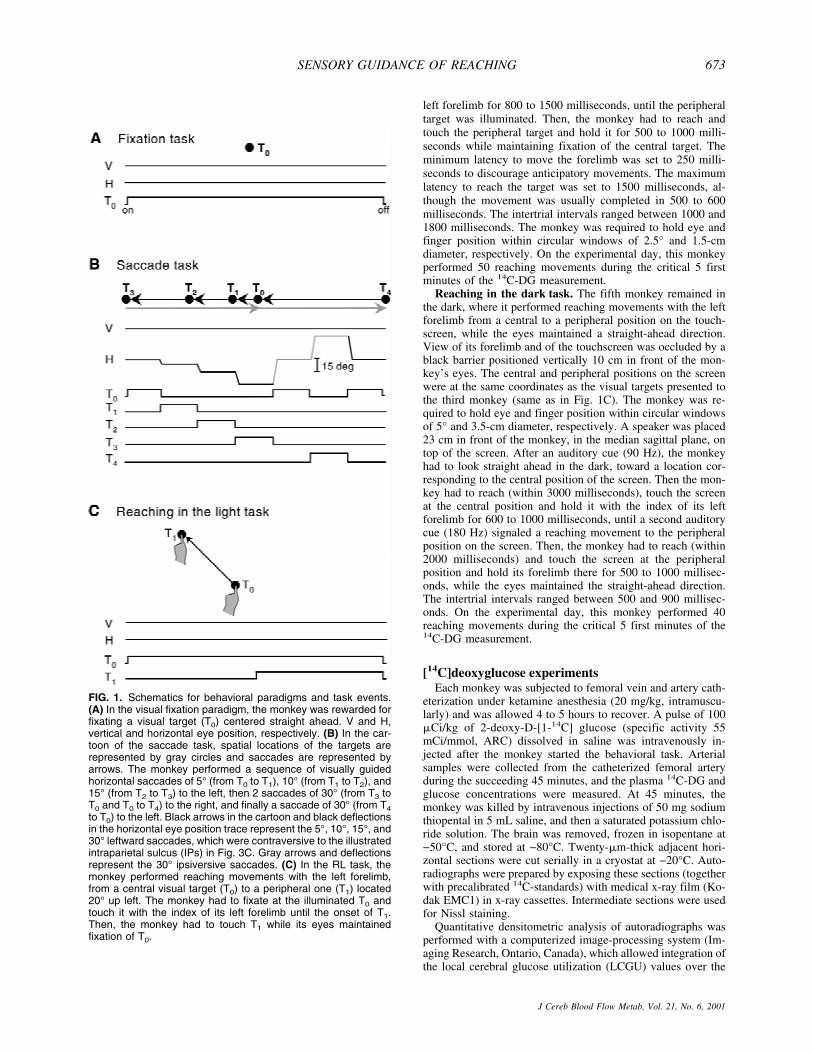
Fixation task. The second monkey had to fixate a centralvisual target located straight ahead (Fig. 1A). The monkey hadto maintain fixation during the whole period of illuminationof the target (4 seconds per trial) within a circular window of2.5° diameter and was allowed to move the eyes only duringthe intertrial intervals (200 to 300 milliseconds). On the ex-perimental day, this monkey maintained fixation for 75% ofthe time.
Saccade task. The third monkey was required to perform asequence of visually guided horizontal saccades of 5°, 10°, and15° to the left, then 2 saccades of 30° to the right, and finallya saccade of 30° to the left, as illustrated in Fig. 1B. Themonkey had to fixate each illuminated target for 300 to 600milliseconds until it disappeared and the next one was illumi-nated signaling a saccadic movement to the new location. Fixa-tion was maintained within a circular window of 2.5° diameter.The minimum latency to move the eyes after onset of eachtarget was set to 100 milliseconds to discourage anticipatorymovements. The intertrial intervals ranged randomly between500 and 800 milliseconds. On the experimental day, this mon-key performed 55 leftward saccades of each amplitude (5°, 10°,15°, and 30°) during the critical 5 first minutes of the 14C-DGmeasurement.
Reaching in the light task. The fourth monkey performedreaching movements in the light with the left forelimb, from acentral to a peripheral visual target, during central fixation (Fig.1C). The central target was located in the median sagittal planeat shoulder height, whereas the peripheral one was located 20°up left on a line inclined by 45° with respect to the horizontaland the midsagittal planes. The monkey was required to look atthe illuminated central target and touch it with the index of its
G. GREGORIOU ET AL.672
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001
left forelimb for 800 to 1500 milliseconds, until the peripheraltarget was illuminated. Then, the monkey had to reach andtouch the peripheral target and hold it for 500 to 1000 milli-seconds while maintaining fixation of the central target. Theminimum latency to move the forelimb was set to 250 milli-seconds to discourage anticipatory movements. The maximumlatency to reach the target was set to 1500 milliseconds, al-though the movement was usually completed in 500 to 600milliseconds. The intertrial intervals ranged between 1000 and1800 milliseconds. The monkey was required to hold eye andfinger position within circular windows of 2.5° and 1.5-cmdiameter, respectively. On the experimental day, this monkeyperformed 50 reaching movements during the critical 5 firstminutes of the 14C-DG measurement.
Reaching in the dark task. The fifth monkey remained inthe dark, where it performed reaching movements with the leftforelimb from a central to a peripheral position on the touch-screen, while the eyes maintained a straight-ahead direction.View of its forelimb and of the touchscreen was occluded by ablack barrier positioned vertically 10 cm in front of the mon-key’s eyes. The central and peripheral positions on the screenwere at the same coordinates as the visual targets presented tothe third monkey (same as in Fig. 1C). The monkey was re-quired to hold eye and finger position within circular windowsof 5° and 3.5-cm diameter, respectively. A speaker was placed23 cm in front of the monkey, in the median sagittal plane, ontop of the screen. After an auditory cue (90 Hz), the monkeyhad to look straight ahead in the dark, toward a location cor-responding to the central position of the screen. Then the mon-key had to reach (within 3000 milliseconds), touch the screenat the central position and hold it with the index of its leftforelimb for 600 to 1000 milliseconds, until a second auditorycue (180 Hz) signaled a reaching movement to the peripheralposition on the screen. Then, the monkey had to reach (within2000 milliseconds) and touch the screen at the peripheralposition and hold its forelimb there for 500 to 1000 millisec-onds, while the eyes maintained the straight-ahead direction.The intertrial intervals ranged between 500 and 900 millisec-onds. On the experimental day, this monkey performed 40reaching movements during the critical 5 first minutes of the14C-DG measurement.
[14C]deoxyglucose experimentsEach monkey was subjected to femoral vein and artery cath-
eterization under ketamine anesthesia (20 mg/kg, intramuscu-larly) and was allowed 4 to 5 hours to recover. A pulse of 100�Ci/kg of 2-deoxy-D-[1-14C] glucose (specific activity 55mCi/mmol, ARC) dissolved in saline was intravenously in-jected after the monkey started the behavioral task. Arterialsamples were collected from the catheterized femoral arteryduring the succeeding 45 minutes, and the plasma 14C-DG andglucose concentrations were measured. At 45 minutes, themonkey was killed by intravenous injections of 50 mg sodiumthiopental in 5 mL saline, and then a saturated potassium chlo-ride solution. The brain was removed, frozen in isopentane at−50°C, and stored at −80°C. Twenty-�m-thick adjacent hori-zontal sections were cut serially in a cryostat at −20°C. Auto-radiographs were prepared by exposing these sections (togetherwith precalibrated 14C-standards) with medical x-ray film (Ko-dak EMC1) in x-ray cassettes. Intermediate sections were usedfor Nissl staining.
Quantitative densitometric analysis of autoradiographs wasperformed with a computerized image-processing system (Im-aging Research, Ontario, Canada), which allowed integration ofthe local cerebral glucose utilization (LCGU) values over the
FIG. 1. Schematics for behavioral paradigms and task events.(A) In the visual fixation paradigm, the monkey was rewarded forfixating a visual target (T0) centered straight ahead. V and H,vertical and horizontal eye position, respectively. (B) In the car-toon of the saccade task, spatial locations of the targets arerepresented by gray circles and saccades are represented byarrows. The monkey performed a sequence of visually guidedhorizontal saccades of 5° (from T0 to T1), 10° (from T1 to T2), and15° (from T2 to T3) to the left, then 2 saccades of 30° (from T3 toT0 and T0 to T4) to the right, and finally a saccade of 30° (from T4
to T0) to the left. Black arrows in the cartoon and black deflectionsin the horizontal eye position trace represent the 5°, 10°, 15°, and30° leftward saccades, which were contraversive to the illustratedintraparietal sulcus (IPs) in Fig. 3C. Gray arrows and deflectionsrepresent the 30° ipsiversive saccades. (C) In the RL task, themonkey performed reaching movements with the left forelimb,from a central visual target (T0) to a peripheral one (T1) located20° up left. The monkey had to fixate at the illuminated T0 andtouch it with the index of its left forelimb until the onset of T1.Then, the monkey had to touch T1 while its eyes maintainedfixation of T0.
SENSORY GUIDANCE OF REACHING 673
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

entire extent of each area of interest. The definition of bordersof cortical subregions was based primarily on their positionrelative to the identifiable fundus and crowns of the IPs (Fig.2A and 2B). LCGU was calculated as in the authors’ previousexperiments (Savaki et al., 1993, 1996), from the appropriatekinetic constants for the monkey (Kennedy et al., 1978), by theoriginal operational equation of the 14C-DG method (Sokoloffet al., 1977). Normalization of LCGU values was based on theaverage unaffected gray matter value pooled across all mon-
keys. Side-to-side percent differences within each monkey(%Dif in Table 1) were calculated as %Dif � (Contra −Ipsi)/Ipsi × 100, where Contra represents the LCGU averagevalue in each cortical area of interest of the hemisphere con-tralateral to the moving forelimb, and Ipsi represents the re-spective ipsilateral LCGU value. Contra to Ipsi LCGU valueswere compared for statistical significances by the Kolmogorov-Smirnov test. Moreover, percent differences between monkeys(%DIF in Results) were calculated as either %DIF � (RD −
FIG. 2. Reconstruction of the medialand lateral banks of intraparietal sul-cus (IPs). (A) Schematic representa-tions of three horizontal sectionsthrough the right hemisphere of amonkey brain at different dorsoventrallevels (top, dorsal sections; bottom,ventral sections). In the first and thirdsections, the light and dark dotted ar-eas outline the cortex of the medial(dorsal, 5DIP; ventral, 5VIP) and thelateral (7DIP and 7VIP) banks of IPs,respectively. (middle) The striped areaindicates the caudorostral extent ofthe reconstructed cortex. The solidcircle (b) within the striped area indi-cates the point of alignment of adja-cent sections, which corresponds tothe intersection of the IPs with the pa-rietoccipital sulcus (POs). Letters a toc indicate the same landmarks in Aand B. As, arcuate sulcus; Cs, centralsulcus; Ls, lunate sulcus; STs, supe-rior temporal sulcus. (B) Schematicrepresentation of two-dimensionalmaps. Zero caudorostral level (line b)corresponds to the most posterior andmedial part of IPs, used for alignmentof adjacent sections. Solid line in themiddle of the map indicates the fundusof the IPs. Dotted lines a and c indicatethe crowns of the medial and lateralbanks of IPs, respectively. Differentshades represent caudal and rostralsubregions. DIP, dorsal intraparietalcortex; VIP, ventral intraparietal cor-tex.
TABLE 1. Metabolic effects in subregions of the intraparietal cortex
Area (n)
C F S RL RD
Contra Ipsi Contra %Dif Ipsi Contra %Dif
5DIP-rostral (105) 38 ± 2 42 ± 2 46 ± 2 43 ± 2 50 ± 5 16* 45 ± 4 55 ± 5 22*5DIP-rostralmost (45) 44 ± 2 58 ± 2 32* 46 ± 1 64 ± 3 39*5DIP-medial (29) 42 ± 1 52 ± 3 24* 43 ± 1 56 ± 2 30*
5DIP-caudal (65) 39 ± 2 40 ± 1 42 ± 3 42 ± 6 46 ± 4 10* 40 ± 2 42 ± 2 55VIP-rostral (43) 38 ± 1 42 ± 1 46 ± 1 47 ± 2 53 ± 3 13* 46 ± 3 46 ± 2 05VIP-caudal (43) 41 ± 1 44 ± 1 48 ± 3 52 ± 2 57 ± 6 10* 45 ± 1 45 ± 3 07DIP-rostral (103) 43 ± 4 48 ± 3 51 ± 3 46 ± 4 47 ± 6 2 46 ± 2 45 ± 3 −2
PFR (18) 43 ± 1 63 ± 2 74 ± 4 59 ± 2 49 ± 27DIP-caudal (66) 41 ± 2 41 ± 2 43 ± 3 39 ± 5 38 ± 3 −2 42 ± 5 40 ± 3 −57VIP-rostral (42) 42 ± 1 47 ± 1 57 ± 4 50 ± 3 56 ± 5 12* 49 ± 2 49 ± 1 07VIP-caudal (42) 44 ± 1 43 ± 1 50 ± 5 46 ± 2 50 ± 3 8* 46 ± 2 48 ± 4 4
Values represent the normalized mean of glucose utilization (LCGU) expressed in �mol 100 g−1 min−1 ± SD obtained from one control (C) andfour experimental monkeys performing one of the following tasks: fixation (F), saccades (S), reaching in the light (RL), and reaching in the dark (RD).n, number of sets of five adjacent horizontal sections used in obtaining mean LCGU values for each region in each hemisphere; %Dif, contra-to-ipsipercent difference calculated as (Contra − Ipsi)/Ipsi × 100; PFR, parietal fixation region.
* All side-to-side differences greater than 5% were statistically significant by the Kolmogorov-Smirnov test at P < 0.001.
G. GREGORIOU ET AL.674
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001
RL)/RL × 100 or %DIF � (RL − RD)/RD × 100. For thesecalculations, the average LCGU value in each cortical area ofinterest contralateral to the moving forelimb in the RD and theRL monkeys was used. Regression analysis was performed tocorrelate the intensity of the fixation effect (LCGU values) ineach hemisphere with the following behavioral parameters: (1)duration of fixation, (2) number of small amplitude contraver-sive saccades, (3) total number of small amplitude (contraver-sive and ipsiversive) saccades, (4) number of small amplitudeipsiversive saccades, (5) total number of contraversive (smalland large amplitude) saccades, and (6) percentage of success-ful trials.
Two-dimensional reconstruction of corticalmetabolic maps
Two-dimensional reconstruction of the spatio-intensive pat-tern of metabolic activity (LCGU) within the rostrocaudal andthe dorsoventral extent of the unfolded IPs was generated ineach hemisphere, as previously described (Dalezios et al.,1996; Savaki et al., 1997). In summary, the distribution ofactivity in the caudorostral extent in each section was deter-mined by measuring LCGU values pixel by pixel (resolution 45�m/pixel) along a line parallel to the surface of the cortex,covering all cortical layers. Each data array (a series of LCGUvalues in each horizontal section), resulting from image seg-mentation in the caudorostral direction, was aligned with thearrays obtained from adjacent horizontal sections (the total of550 sections of 20-�m thickness which contained the wholeextent of IPs) in the dorsoventral dimension of the brain. Thecaudalmost part of the IPs (the intersection of IPs with the
parietoccipital sulcus, POs) was used for the alignment of ad-jacent data arrays (point b in Fig. 2A, and line b in Fig. 2B).The plotting resolution of both the caudorostral and the dorso-ventral dimensions was 100 �m. Accordingly, each verticalline at a given dorsoventral level on the left half of the meta-bolic maps (Figs. 3 and 4) represents the distribution of theaverage activity in the medial bank along the rostrocaudal di-mension in five serial sections. The vertical line at the samedorsoventral level on the right half of the metabolic maps rep-resents the rostrocaudal distribution of the average activity inthe lateral bank within the same five sections. Occasional miss-ing data arrays in the dorsoventral dimension were filled usinglinear interpolation between neighboring values.
The maps of hemispheric differences (Fig. 5) were obtainedby subtracting the metabolic map of the ipsilateral IPs cortex(Fig. 4C) from the map of the IPs contralateral to the movingforelimb (Fig. 4D) in the RD monkey and by subtracting themetabolic map of the IPs contralateral to the moving forelimbin the RD monkey (Fig. 4D) from the map of the IPs contra-lateral to the moving forelimb in the RL monkey (Fig. 4B). Thefundus as well as the crowns of IPs were used for alignment ofthe maps used for subtraction.
RESULTS
The 14C-DG method allows the visualization of pat-terns of brain activity in awake behaving monkeys, with-out invading them and their cerebral processes. The au-thors used this quantitative method to map the distribu-tion of metabolic activity in the IPs of one control and
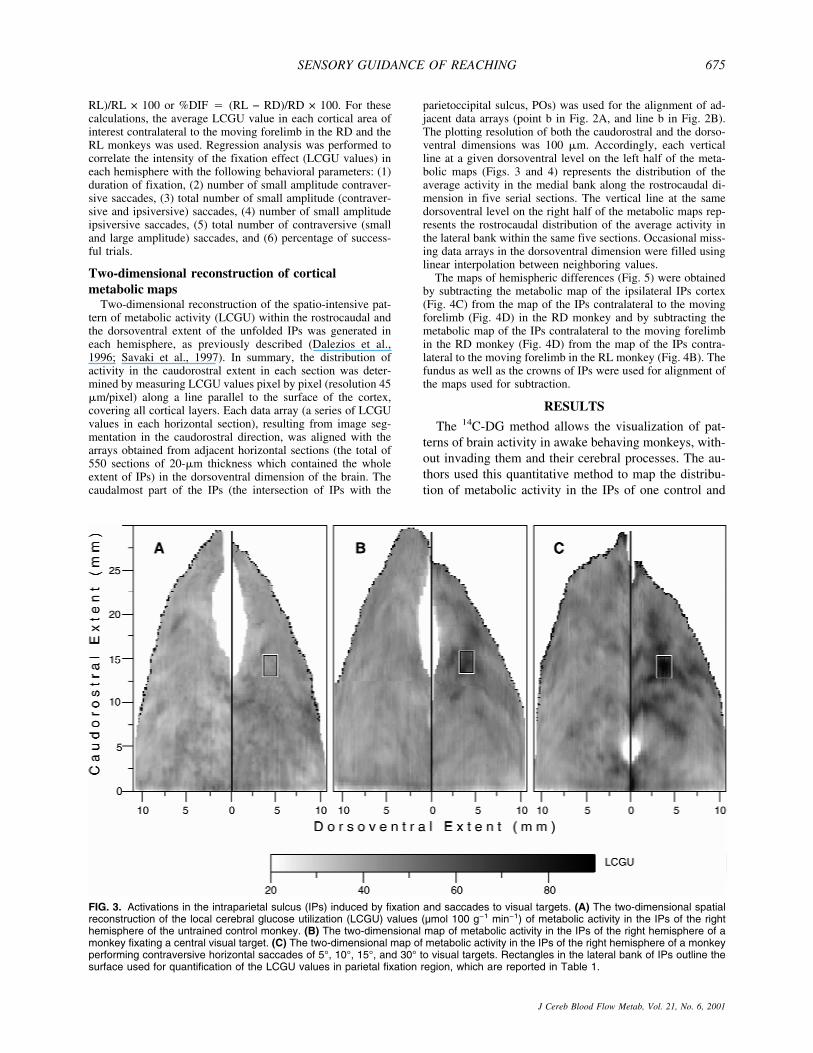
FIG. 3. Activations in the intraparietal sulcus (IPs) induced by fixation and saccades to visual targets. (A) The two-dimensional spatialreconstruction of the local cerebral glucose utilization (LCGU) values (µmol 100 g−1 min−1) of metabolic activity in the IPs of the righthemisphere of the untrained control monkey. (B) The two-dimensional map of metabolic activity in the IPs of the right hemisphere of amonkey fixating a central visual target. (C) The two-dimensional map of metabolic activity in the IPs of the right hemisphere of a monkeyperforming contraversive horizontal saccades of 5°, 10°, 15°, and 30° to visual targets. Rectangles in the lateral bank of IPs outline thesurface used for quantification of the LCGU values in parietal fixation region, which are reported in Table 1.
SENSORY GUIDANCE OF REACHING 675
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

four experimental monkeys performing one of four tasks:(1) fixation of a central visual target, (2) horizontal sac-cades of different amplitudes to visual targets, (3) reach-ing in the light during fixation of the central visual target,and (4) reaching in the dark while eyes maintained astraight-ahead direction. The distribution of metabolicactivity in the entire caudorostral and dorsoventral extentof the IPs (Fig. 2A) was represented as a two-dimensional map in each one of the ten hemispheres ofthe present study. The medial and the lateral banks of IPswere subdivided into different subregions on the basis ofthe activations induced by the performance of the differ-
ent tasks (Fig. 2B). This parcellation has been used inprevious studies (Savaki et al., 1997; Savaki and Dale-zios, 1999) and is now presented in detail, on the basis offunctional criteria provided by the current study. Thenomenclature used to describe this parcellation differsfrom what other authors have reported in recent litera-ture. However, the use of this nomenclature is necessaryto present quantitative results in activated regions thatextend beyond the already reported ones. Area 5DIP cov-ers the dorsalmost three fifths of the medial bank. Itsrostral subregion (5DIP-rostral) is defined as the rostral-most three fourths of the entire rostrocaudal extent of the
FIG. 4. Activations in the intrapari-etal sulcus (IPs) induced by reachingin the light and reaching in the darkduring fixation. (A) Cortical two-dimensional map of metabolic activ-ity in the IPs of the left hemisphere,ipsilateral to the moving forelimb, ina monkey reaching from a central toa peripheral visual target in the lightduring fixation of the central target.(B) The two-dimensional map of cor-tical activity in the IPs of the righthemisphere, contralateral to themoving forelimb, in the same mon-key as A. r, rostralmost; m, medial-activated regions, outlined in the me-dial bank of IPs. (C) The two-dimensional map of cortical activityin the IPs of the left hemisphere, ip-silateral to the moving forelimb, in amonkey that remained in completedarkness and performed acousticallytriggered reaching from a central to aperipheral position while the eyesmaintained a straight ahead direc-tion. (D) The two-dimensional map ofcortical activity in the IPs of the righthemisphere, contralateral to themoving forelimb, in the same mon-key as C. LCGU, local cerebral glu-cose utilization.
G. GREGORIOU ET AL.676
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001
medial bank, reaching the fundus in the rostral tip of thebank. The remaining caudalmost one fourth (5DIP-caudal) extends caudally up to the intersection of the POswith the IPs. The delineation of areas 5DIP-rostral and5DIP-caudal is based on the fact that the former areadisplayed major activations in contrast to the latter one inthe reaching monkeys (Fig. 5). Area 5VIP covers theventralmost two fifths of the medial bank (4 mm adjacentto the fundus) extending rostrally for 21 mm startingfrom the IPs-POs intersection. The parts correspondingto 5VIP-rostral and 5VIP-caudal cover the rostralmosttwo thirds and the caudalmost one third of area 5VIP,respectively. The same spatial relations are valid forthe homologous subregions in the lateral bank of theIPs, that is, for areas 7DIP-rostral, 7DIP-caudal, 7VIP-rostral, and 7VIP-caudal. The delineation of rostral andcaudal regions within the lateral bank is based on thedifferential activations induced by eye movements inthese regions (Fig. 3). In most of the illustrated hemi-
spheres (Figs. 3C, 4A to 4D), the fundus of the IPs isalmost parallel to the horizontal plane of sectioning, andthus overlaps with the line of zero dorsoventrality. How-ever, in 2 of the IPs maps (Fig. 3A and 3B), the rostralpart of both banks 5 and 7 deviates from the line of zerodorsoventrality, apparently because of a different geom-etry of these sulci.
Fixation and saccade related regions in the IPs ofthe monkey brain
The two-dimensional reconstructed map of metabolicactivity in the IPs of the right hemisphere in the monkeyperforming fixation of a central visual target displays arestricted region of enhanced metabolic activity in thelateral bank (Fig. 3B), which is not present in the respec-tive IPs map of the untrained control monkey (Fig. 3A).This activation is located almost in the middle of thedorsoventral extent of the lateral bank (3 to 4 mm fromthe fundus) and rather rostrally (10 to 12 mm from its
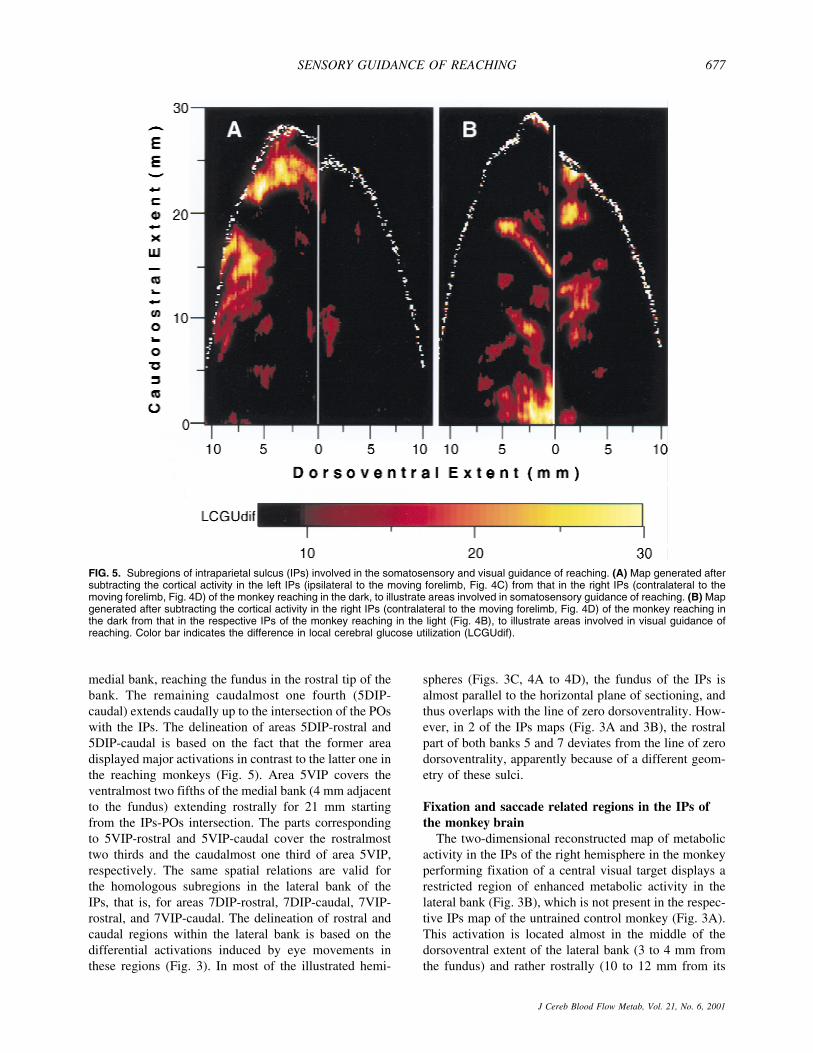
FIG. 5. Subregions of intraparietal sulcus (IPs) involved in the somatosensory and visual guidance of reaching. (A) Map generated aftersubtracting the cortical activity in the left IPs (ipsilateral to the moving forelimb, Fig. 4C) from that in the right IPs (contralateral to themoving forelimb, Fig. 4D) of the monkey reaching in the dark, to illustrate areas involved in somatosensory guidance of reaching. (B) Mapgenerated after subtracting the cortical activity in the right IPs (contralateral to the moving forelimb, Fig. 4D) of the monkey reaching inthe dark from that in the respective IPs of the monkey reaching in the light (Fig. 4B), to illustrate areas involved in visual guidance ofreaching. Color bar indicates the difference in local cerebral glucose utilization (LCGUdif).
SENSORY GUIDANCE OF REACHING 677
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

anterior tip). It lies within both areas 7DIP-rostral and7VIP-rostral. The same activation was observed in theIPs map of the left hemisphere of the fixating monkeyand not in the respective map of the untrained controlsubject (maps similar to those of the right hemispheres inFig. 3B and 3A, respectively, and thus not illustrated).The absence of such activation in the IPs of the untrainedcontrol monkey indicates that the activated region inFig. 3B may be related to either the fixation itself or tothe performance of small amplitude saccades (0° to 2.5°)around the point of fixation. This area is referred to asthe parietal fixation region (PFR). Quantification ofthe average LCGU value within the activated PFR inthe fixating monkey (outlined by rectangle in Fig. 3B)and the corresponding region in the control monkey re-veals a 47% increase in metabolic activity within theactivated PFR.
The right hemisphere of the monkey performing con-traversive horizontal saccades of 5°, 10°, 15°, and 30°displays activations within an extended region in the lat-eral bank of IPs (Fig. 3C). This region is referred to asthe parietal saccade region (PSR). The activations withinPSR appear as bands of increased metabolic activity thatcover part of areas 7DIP-rostral and 7VIP-rostral. Themost active subregion within the PSR of the monkeyperforming saccades (outlined by rectangle in Fig. 3C)overlaps topographically with the PFR of the fixatingmonkey, and is most probably related to the multipleshort fixations of all visual targets for saccades requiredby the task.
The activation in 7IP of the left hemisphere in the RLmonkey (Fig. 4A), which lies approximately 4 mm fromthe fundus and 12 mm from the anterior tip of the lateralbank, overlaps topographically with the PFR of the fix-ating monkey. PFR is activated in the RL monkey be-cause this animal performed reaching during central vi-sual fixation. Thus, by using complementary tasks, theactivated PFR is demonstrated not only in the two hemi-spheres of the fixating monkey but also in the two hemi-spheres of the saccading and the RL monkeys. Rect-angles of the same dimensions positioned at correspond-ing coordinates to those indicated in Fig. 3 were used forquantification of the LCGU within the PFR of all theanalyzed hemispheres. Of interest is the gradual decreaseof the PFR activation in the hemispheres of (1) the sac-cading (S) monkey, (2) the left and (3) right hemispheresof the fixating (F) monkey, (4) the ipsilateral and (5)contralateral hemispheres of the RL monkey, as well as(6) the control (C) monkey (74, 64, 63, 59, 49 and 43�mol 100 g−1 min−1, respectively). Regression analysiswas performed to reveal any possible correlation be-tween the intensity of the PFR activation and severaldifferent behavioral parameters. The intensity of PFRactivation in the different hemispheres is not correlatedwith the duration of fixation (2.8 minutes for the S mon-
key, 3.6 minutes for the F monkey, and 2.9 minutes forthe RL monkey) during the critical 5 first minutes of the14C-DG experiment (regression coefficient, r � 0.08,P > 0.1). In contrast, the intensity of the PFR activationis well correlated with the number of small amplitudecontraversive saccades performed around the point offixation (168 for the S monkey, 82 for the left and 138for the right hemisphere of the F monkey, 63 for theipsilateral and 18 for the contralateral RL hemispheres)during the 5 first minutes of the experiment (r � 0.92,P < 0.05). When the total number of small amplitude(contraversive and ipsiversive) saccades is used for thecomparison, the r drops to 0.76 (P > 0.1). This dropof the regression coefficient is because of the inclusionof small amplitude ipsiversive saccades (43 for theS monkey, 138 for the left and 82 for the right hemi-sphere of the F monkey, 18 for the ipsilateral and 63 forthe contralateral RL hemisphere), which are not corre-lated with the intensity of the PFR activation (r � 0.04,P > 0.1). Similarly, when the total number of contraver-sive (small and large amplitude) saccades (345 for theS monkey, 121 for the left and 178 for the right hemi-sphere of the F monkey, 197 for the ipsilateral and 143for the contralateral RL hemisphere) is used for the com-parison the r drops to 0.72 (P > 0.1). Finally, the authorsexamined the relation between the intensity of the PFRactivation and the state of attention of the animal. Agree-ably, the percentage of successful trials could reflect thestate of attention of the monkey. Regression analysisdemonstrates that the intensity of the PFR activation inthe different hemispheres is not correlated with the per-centage of successful trials (80 for the S monkey, 45 forthe F monkey, and 70 for the RL monkey) during the5 first minutes of the experiment (r � 0.06, P > 0.1). Inconclusion, the results of the linear regression analysisindicate that 85% of the variance of the dependent vari-able (intensity of the PFR activation) can be accountedfor by the number of small amplitude contraversive sac-cades performed around the point of fixation.
In summary, the metabolic effects induced by the per-formance of visual fixation and saccades to visual targetswere confined in the intraparietal area 7 of the lateralbank in the IPs. The absence of any remarkable activa-tion in the medial bank of the monkeys performing thefixation and saccade tasks indicates that intraparietal area5 has no major involvement in the visual and saccadicaspects of these tasks.
Reaching related regions in the IPs of themonkey brain
The medial bank of the IPs was activated morestrongly in the monkeys performing the reaching tasks,and mainly in the hemisphere contralateral to the movingforelimb (Fig. 4B and 4D).
G. GREGORIOU ET AL.678
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

Figure 4A and 4B represent the distribution of meta-bolic activity in the hemispheres ipsilateral and contra-lateral to the moving forelimb, respectively, of the mon-key reaching from a central to a peripheral target in thelight during fixation of the central visual target (RL).Both areas 5DIP and 5VIP are strongly activated in thehemisphere contralateral to the moving forelimb (Fig.4B). Two activated regions are focused within area5DIP-rostral in the contralateral hemisphere of the RLmonkey (r and m outlined in Fig. 4B). The first one (r),which is located in the rostralmost part of area 5DIP, ismore pronounced and displays 32% greater activity inthe contralateral as compared with the ipsilateral hemi-sphere (Table 1, 5DIP-rostralmost). The second one (m),which is located caudally within the limits of area 5DIP-rostral, is greater by 24% in the contralateral than in theipsilateral hemisphere (Table 1, 5DIP-medial). Severalactivations within area 5VIP of the RL monkey werebilateral, with the most pronounced ones running almostparallel to each other and covering the entire rostrocau-dal extent in the hemisphere contralateral to the reachingforelimb. Some activations within area 7VIP of the RLmonkey also were bilateral, with the most intense andextended ones running parallel to each other in the con-tralateral 7VIP-rostral.
Figure 4C and 4D represent the distribution of meta-bolic activity in the hemispheres ipsilateral and contra-lateral to the moving forelimb, respectively, of the mon-key performing acoustically triggered reaching move-ments from a central to a peripheral position, in completedarkness, while its eyes maintained a straight-ahead di-rection (RD). Area 5DIP-rostral in the RD monkey dis-plays two activated regions, the 5DIP-rostralmost and the5DIP-medial, in the hemisphere contralateral to the mov-ing forelimb (outlined in Fig. 4D), same way as in the RLmonkey. Thus, by using complementary tasks, the acti-vated regions 5DIP-rostralmost and 5DIP-medial aredemonstrated not only in the RL but also in the RDmonkey. These activations in the RD monkey displaygreater glucose consumption in the contralateral hemi-sphere as compared with the corresponding regions ofthe ipsilateral hemisphere by 39% and 30%, respectively.Comparison of these two effects in 5DIP-rostral in theRD and RL monkeys demonstrated that the activationswere slightly greater in the RD monkey (Table 1). Incontrast, area 5VIP of the RD monkey (Fig. 4D) did notdisplay the pronounced activations observed in the RLmonkey (Fig. 4B). A spatially restricted, moderately ac-tivated band is present in the lateral bank of the IPs in theRD monkey, bilaterally. Given that reaching was per-formed in complete darkness while the eyes remainedstationary, this activated band cannot be attributed tovisual stimulation or to voluntary saccades. This activa-tion may be related to eye drifting during fixation in thedark. However, it cannot be excluded that this activation
may be related to the auditory signals used as cues forreaching, because neuronal responses to auditory signalsused as cues for saccades have been reported in the lat-eral bank of the IPs (Mazzoni et al., 1996).
The map generated after subtracting the cortical activ-ity in the IPs ipsilateral to the moving forelimb (Fig. 4C)from that in the IPs contralateral to the moving forelimb(Fig. 4D) in the RD monkey is shown in Fig. 5A. In thismap, the two activated regions 5DIP-rostralmost and5DIP-medial are clearly delineated and demonstrate theunilaterality of the effect induced by RD. The outlines ofthese two activations in Fig. 5A were used to delimitthe corresponding regions (r and m) in Fig. 4B and 4D.In the absence of visual input, these activations can onlybe attributed to somatosensory information used to guidethe forelimb in the dark.
Both somatosensory and visual information is used toguide reaching in the light, whereas only somatosensoryinput is available to guide reaching in the dark. To revealthe regions associated with the visual guidance of reach-ing, the authors subtracted the map of the contralateralIPs in the RD monkey (Fig. 4D) from the equivalent mapin the RL monkey (Fig. 4B). The generated map illus-trates activated zones mainly in the ventral intraparietalcortex around the fundus of IPs (Fig. 5B). Quantitativecomparison of the effects in the different subregions ofthe ventral intraparietal cortex demonstrated activationsgreater by 15% in area 5VIP-rostral, 27% in 5VIP-caudal, and 14% in 7VIP-rostral in the RL as comparedwith the RD monkey.
Based on the results of the two subtraction maps, theauthors conclude that the dorsal part of intraparietal area5 is involved in the somatosensory guidance of the reach-ing forelimb, whereas the ventral intraparietal cortex isimplicated in the visual guidance of reaching.
DISCUSSION
The current results demonstrate that distinct subre-gions within the monkey intraparietal cortex are involvedin different aspects of goal directed movements. This isthe first time that high-resolution maps of the spatialdistribution of metabolic activity in the entire intrapari-etal cortex have been generated to reveal the specificregions involved in visual fixation, voluntary saccades,reaching in the light, and reaching in the dark.
Subregions in the lateral bank of IPs involved infixation and saccades
Parietal saccade region, a region associated with sac-cades to visual targets, was found in the lateral bank ofIPs. The PSR is located in the rostral sector of 7IP andcovers part of area LIP, as this has been defined on thebasis of single unit recording (Andersen et al., 1990;Barash et al., 1991b) and microstimulation (Thier andAndersen, 1996), and the part of area VIP lying in the
SENSORY GUIDANCE OF REACHING 679
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

lateral bank of IPs close to the fundus (Colby et al.,1993). The activation of PSR in the current study isconsistent with the well established saccadic (Barash etal., 1991a; Colby et al., 1995; Thier and Andersen, 1996)and visual (Blatt et al., 1990; Colby et al., 1993, 1995;Gottlieb et al., 1998) properties of areas LIP and VIP.However, the PSR in the current study may cover only apart of the saccade related intraparietal area because itsactivation was induced by single directional (only hori-zontal) saccades of different amplitudes. Indeed, prelimi-nary observations in the authors’ laboratory indicate thatsaccades to targets on a line inclined by 45° with respectto the horizontal plane activate a region that extendscaudal to the presently illustrated PSR (Gregoriou GG,Moschovakis AK, Savaki HE, unpublished observa-tions). The full extent of the saccade related intraparietalarea will be revealed by the performance of saccades ofdifferent amplitudes in several directions.
Parietal fixation region, a region associated with fixa-tion of a visual target, was found in the rostral sector ofthe lateral bank of IPs. The PFR lies in the anterior partof PSR, and posterior to AIP as this has been defined onthe basis of neurophysiologic and anatomic studies(Sakata et al., 1995; Luppino et al., 1999; Murata et al.,2000). The location of PFR appears to be in register withthe recently reported foveal representation in the rostralpart of intraparietal area 7 (Ben Hamed et al., 1999). Itsactivation was consistent in both hemispheres of themonkey performing fixation of a visual target as well asin both hemispheres of the RL monkey. The PFR wasalso active in the monkey performing sequential horizon-tal saccades of different amplitudes, whereas it was notactive in the control monkey. Because the intensity of thePFR activation varied in the different experimental hemi-spheres, the authors used linear regression analysis toexamine whether this activation was correlated with cer-tain behavioral parameters. No significant correlationswere obtained with (1) duration of fixation, (2) totalnumber of small amplitude (contraversive and ipsiver-sive) saccades, (3) number of small amplitude ipsiversivesaccades, (4) total number of contraversive (small andlarge amplitude) saccades, and (5) percentage of success-ful trials. Taking into account that only the number ofsmall amplitude contraversive saccades was significantlycorrelated with the intensity of the PFR activation in thedifferent hemispheres, the authors suggest that the PFRactivation is associated with the performance of smallamplitude contraversive saccades around the point offixation. Therefore, PFR may be the rostral part of PSRthat is associated with small amplitude saccades.
In conclusion, this is the first time that a high-resolution map of functional activity induced by visuallyguided saccades of different amplitudes is generated, anda region related to fixation of a visual target is mapped inthe lateral intraparietal area 7.
Subregions in the IPs involved in the visual andsomatosensory guidance of reaching
The rostral part of the ventral intraparietal cortex (ar-eas 5VIP-rostral and 7VIP-rostral) in the current studylargely overlaps with area VIP as this has been definedon the basis of single unit recording (Colby et al., 1993).The current data demonstrate that area 5VIP (including5VIP-rostral and 5VIP-caudal) is activated by visuallyguided reaching (Fig. 4B) and not in response to visualstimuli used as targets for saccades (Fig. 3C). Given thatcells in area VIP display visual properties (Colby andDuhamel, 1991; Colby et al., 1993), the current findingindicates that visual information in this area is used toguide reaching and not to code target location per se.However, the activation of area 7VIP during both visu-ally guided forelimb movements (Fig. 4B) and visuallyguided eye movements (Fig. 3C) indicates that this areaencodes visual information about the location of stimuliused as targets for motor acts, whatever the effector used.
In the reaching tasks, area 5DIP-rostral was activatedin the hemisphere contralateral to the moving forelimb inboth the RL and RD monkeys. These two monkeys hadin common only proprioceptive-somatosensory informa-tion available to guide the forelimb. Therefore, the acti-vation of 5DIP-rostral should be attributed to the somato-sensory guidance of reaching. Moreover, this activationwas greater in the RD monkey, in which only somato-sensory (and no visual) information was available toguide the forelimb. The higher activation of area 5DIP-rostral in the RD monkey as compared with that in theRL monkey may act as a compensatory mechanism tothe absence of any visual information, in accordancewith the authors’ previous findings and suggestion (Sa-vaki et al., 1993). The ventral intraparietal cortex (in-cluding both 5VIP and 7VIP) was activated mainly in theRL monkey, which was the only one with visual infor-mation available to guide the forelimb. Therefore, theactivation of the ventral intraparietal cortex should beattributed to the visual guidance of reaching. Indeed, asimilar activation to that around the fundus of IPs in theRL monkey also was observed in the IPs of a monkeyperforming reaching and grasping under visual fixationin the light (Raos VC, Papadaki P, Savaki HE, unpub-lished observations). The less lateralized effect in theventral intraparietal cortex associated with visual pro-cessing as compared with that in dorsal intraparietal area5 associated with somatosensory processing is consistentwith previous findings (Colby et al., 1993; Savaki andDalezios, 1999).
The functional dissociation of the dorsal intraparietalarea 5 related to somatosensory guidance of reachingfrom the ventral intraparietal cortex related to visualguidance proves the authors’ earlier hypothesis regardingthe differential contribution of IPs cortical subregions in
G. GREGORIOU ET AL.680
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

the control of reaching (Savaki et al., 1993; Savaki andDalezios, 1999). Moreover, this dissociation is consistentwith the following anatomic data. Area VIP receives vi-sual input from areas MT, MST, FST (Maunsell and VanEssen, 1983; Ungerleider and Desimone, 1986; Bous-saoud et al., 1990), and PO (Colby et al., 1988), andprojects to the ventral premotor cortex (Luppino et al.,1999). More dorsally located areas MIP and PEip receivesomatosensory input (Pandya and Seltzer, 1982; Pearsonand Powell, 1985) and project to the dorsal premotorcortex (Matelli et al., 1998). Area MIP has been de-scribed as the middle one third (Colby et al., 1988; Ma-telli et al., 1998) of the caudal half of the medial bank.Accordingly, area MIP must cover part of the areas 5DIPand 5VIP, a fact consistent with the reported conver-gence of visual and somatosensory input (Pandya andSeltzer, 1982; Colby et al., 1988; Caminiti et al., 1996).Area PEip occupies the dorsalmost one third of the cau-dal half, and the whole rostral part of the medial bank(Matelli et al., 1998), thus overlapping with most of thearea 5DIP-rostral. Two neuronal populations were la-beled in PEip after injection in the dorsal premotor F2arm field (Matelli et al., 1998). These two neuronalpopulations correspond topographically to subregions5DIP-rostralmost and 5DIP-medial, which were bothmarkedly activated in the current study (Fig. 5A).
Furthermore, the functional dissociation of the dorsalintraparietal area 5 related to somatosensory guidance ofreaching from the ventral intraparietal cortex related tovisual guidance is complemented by the following elec-trophysiologic data. Many neurons in VIP respond tovisual stimuli located in the near extrapersonal space(Duhamel et al., 1998). Area MIP displays a range ofproperties from purely visual to bimodal to purely so-matosensory, as the electrode moves from the fundus tothe crown of the medial bank (Colby and Duhamel,1991). The dorsal intraparietal area 5 responds to so-matosensory stimuli (Mountcastle et al., 1975) and con-trols arm movements (Kalaska et al., 1990). Cells withmovement-related activity associated with the executionof reaching are located around the crown of the medialbank of IPs, whereas cells with set- and signal-relatedactivity associated with the planning of reaching are lo-cated deeper in the medial bank (Johnson et al., 1996).Finally, the current data are consistent with the results ofa lesion study that has demonstrated that removal of thedorsal part of the medial bank causes misreaching in thedark, but has little effect on reaching in the light (Rush-worth et al., 1997).
In conclusion, quantitative high-resolution maps offunctional activity in the intraparietal cortex demon-strate, for the first time, the spatially distinct neuronalpopulations that are associated with the visual and thesomatosensory guidance of reaching.
Acknowledgment: The authors are indebted to Adonis Mos-chovakis for help and advice at all stages of the project.
REFERENCES
Andersen RA, Bracewell RM, Barash S, Gnadt JW, Fogassi L (1990)Eye position effects on visual, memory, and saccade-related activ-ity in areas LIP and 7a of macaque. J Neurosci 10:1176–1196
Andersen RA, Snyder LH, Bradley DC, Xing J (1997) Multimodalrepresentation of space in the posterior parietal cortex and its usein planning movements. Annu Rev Neurosci 20:303–330
Barash S, Bracewell RM, Fogassi L, Gnadt JW, Andersen RA (1991a)Saccade-related activity in the lateral intraparietal area. I. Tempo-ral properties; comparison with area 7a. J Neurophysiol 66:1095–1108
Barash S, Bracewell RM, Fogassi L, Gnadt JW, Andersen RA (1991b)Saccade-related activity in the lateral intraparietal area. II. Spatialproperties. J Neurophysiol 66:1109–1124
Ben Hamed S, Bihouee A, Deneve S, Duhamel J-R (1999) Neuralcorrelates of visual fixation in the dorsal lateral intraparietal area ofthe macaque (LIPd). Society for Neuroscience Abstracts 25:1163
Blatt GJ, Andersen RA, Stoner GR (1990) Visual receptive field or-ganization and cortico-cortical connections of the lateral intrapa-rietal area (LIP) in the macaque. J Comp Neurol 299:421–445
Boussaoud D, Ungerleider LG, Desimone R (1990) Pathways for mo-tion analysis: cortical connections of the medial superior temporaland fundus of the superior temporal visual areas in the macaque.J Comp Neurol 296:462–495
Caminiti R, Ferraina S, Johnson PB (1996) The sources of visual in-formation to the primate frontal lobe: a novel role for the superiorparietal lobule. Cereb Cortex 6:319–328
Colby CL, Gattass R, Olson CR, Gross CG (1988) Topographicalorganization of cortical afferents to extrastriate visual area PO inthe macaque: a dual tracer study. J Comp Neurol 269:392–413
Colby CL, Duhamel J-R (1991) Heterogeneity of extrastriate visualareas and multiple parietal areas in the macaque monkey. Neuro-psychologia 23:517–537
Colby CL, Duhamel J-R, Goldberg ME (1993) Ventral intraparietalarea of the macaque: anatomic location and visual response prop-erties. J Neurophysiol 69:902–914
Colby CL, Duhamel J-R, Goldberg ME (1995) Visual, presaccadic, andcognitive activation of single neurons in monkey lateral intrapari-etal area. J Neurophysiol 76:2841–2852
Colby CL, Goldberg ME (1999) Space and attention in parietal cortex.Annu Rev Neurosci 22:319–349
Dalezios Y, Raos VC, Savaki HE (1996) Metabolic activity pattern inthe motor and somatosensory cortex of monkeys performing avisually guided reaching task with one forelimb. Neuroscience 72:325–333
Duhamel J-R, Colby CL, Goldberg ME (1998) Ventral intraparietalarea of the macaque: congruent visual and somatic response prop-erties. J Neurophysiol 79:126–136
Georgopoulos AP, Caminiti R, Kalaska JF (1984) Static spatial effectsin motor cortex and area 5: quantitative relations in a two-dimensional space. Exp Brain Res 54:446–454
Georgopoulos AP, Massey JT (1988) Cognitive spatial-motor pro-cesses. 2. Information transmitted by the direction of two-dimensional arm movements and by neuronal populations in pri-mate motor cortex and area 5. Exp Brain Res 69:315–326
Gnadt JW, Andersen RA (1988) Memory related motor planning ac-tivity in posterior parietal cortex of macaque. Exp Brain Res 70:216–220
Gottlieb JP, Kusunoki M, Goldberg ME (1998) The representation ofvisual salience in monkey parietal cortex. Nature 391:481–484
Johnson PB, Ferraina S, Bianchi L, Caminiti R (1996) Cortical net-works for visual reaching: physiological and anatomical organiza-tion of frontal and parietal lobe arm regions. Cereb Cortex 6:102–119
Kalaska JF, Caminiti R, Georgopoulos AP (1983) Cortical mechanismsrelated to the direction of two-dimensional arm movements: rela-
SENSORY GUIDANCE OF REACHING 681
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001

tions in parietal area 5 and comparison with motor cortex. ExpBrain Res 51:247–260
Kalaska JF, Cohen DAD, Prud’homme M, Hyde ML (1990) Parietalarea 5 neuronal activity encodes movement kinematics, not move-ment dynamics. Exp Brain Res 80:351–364
Kalaska JF (1996) Parietal cortex area 5 and visuomotor behavior. CanJ Physiol Pharmacol 74:483–498
Kennedy C, Sakurada O, Shinohara M, Jehle J, Sokoloff L (1978)Local cerebral glucose utilization in the normal conscious macaquemonkey. Ann Neurol 4:293–301
Luppino G, Murata A, Govoni P, Matelli M (1999) Largely segregatedparietofrontal connections linking rostral intraparietal cortex (areasAIP and VIP) and the ventral premotor cortex (areas F5 and F4).Exp Brain Res 128:181–187
Matelli M, Govoni P, Galetti C, Kutz GF, Luppino G (1998) Superiorarea 6 afferents from the superior parietal lobule in the macaquemonkey. J Comp Neurol 402:327–352
Maunsell JHR, Van Essen DC (1983) The connections of the middletemporal visual area (MT) and their relationship to a cortical hi-erarchy in the macaque monkey. J Neurosci 3:2563–2586
Mazzoni P, Bracewell RM, Barash S, Andersen RA (1996) Motorintention activity in the macaque’s lateral intraparietal area. I. Dis-sociation of motor plan from sensory memory. J Neurophysiol76:1439–1456
Mountcastle VB, Lynch JC, Georgopoulos A, Sakata H, Acuna C(1975) Posterior parietal association cortex of the monkey: com-mand functions for operations within extrapersonal space. J Neu-rophysiol 38:871–908
Murata A, Gallese V, Luppino G, Kaseda M, Sakata H (2000) Selec-tivity for the shape, size, and orientation of objects for grasping inneurons of monkey parietal area AIP. J Neurophysiol 83:2580–2601
Pandya DN, Seltzer B (1982) Intrinsic connections and architectonicsof posterior parietal cortex in the rhesus monkey. J Comp Neurol204:204–210
Pearson RCA, Powell TPS (1985) The projection of the primary so-
matic sensory cortex upon area 5 in the monkey. Brain Res Rev 9:89–107
Robinson DA (1963) A method of measuring eye movement usinga scleral search coil in a magnetic field. IEEE Trans Biomed Elec-tron 10:137–145
Rushworth MES, Nixon PD, Passingham RE (1997) Parietal cortex andmovement. I. Movement selection and reaching. Exp Brain Res117:292–310
Sakata H, Taira M, Murata A, Mine S (1995) Neural mechanisms ofvisual guidance of hand action in the parietal cortex of the monkey.Cereb Cortex 5:429–438
Savaki HE, Kennedy C, Sokoloff L, Mishkin M (1993) Visually guidedreaching with the forelimb contralateral to a ‘blind’ hemisphere:a metabolic mapping study in monkeys. J Neurosci 13:2772–2789
Savaki HE, Kennedy C, Sokoloff L, Mishkin M (1996) Visually guidedreaching with the forelimb contralateral to a ‘blind’ hemisphere inthe monkey: contribution of the cerebellum. Neuroscience 75:143–159
Savaki HE, Raos VC, Dalezios Y (1997) Spatial cortical patterns ofmetabolic activity in monkeys performing a visually guided reach-ing task with one forelimb. Neuroscience 76:1007–1034
Savaki HE, Dalezios Y (1999) 14C-Deoxyglucose mapping of the mon-key brain during reaching to visual targets. Prog Neurobiol 58:473–540
Sokoloff L, Reivich M, Kennedy C, Rosiers MHD, Patlak CS, Petti-grew KD, Sakurada O, Shinohara M (1977) The [14C]deoxyglu-cose method for the measurement of local cerebral glucose utili-zation: theory, procedure, and normal values in the conscious andanesthetized albino rat. J Neurochem 28:897–916
Thier P, Andersen RA (1996) Electrical microstimulation suggests twodifferent forms of representation of head-centered space in theintraparietal sulcus of rhesus monkeys. Proc Natl Acad Sci U S A93:4962–4967
Ungerleider LG, Desimone R (1986) Cortical connections of area MTin the macaque. J Comp Neurol 248:190–222
G. GREGORIOU ET AL.682
J Cereb Blood Flow Metab, Vol. 21, No. 6, 2001