dynamics of horizontal vergence movements: interaction with horizontal and vertical saccades and...
TRANSCRIPT

Vision Research 38 (1998) 3943–3954
Dynamics of horizontal vergence movements: interaction withhorizontal and vertical saccades and relation with monocular
preferences
Anna F. van Leeuwen a,c,*, Han Collewijn a,c, Casper J. Erkelens b,c
a Department of Physiology, Faculty of Medicine, Erasmus Uni6ersity, Rotterdam, PO Box 1738, 3000 DR Rotterdam, The Netherlandsb Department of Physics of Man, Utrecht Uni6ersity, Utrecht, The Netherlands
c Helmholtz Institute, The Netherlands
Received 3 November 1997; received in revised form 23 February 1998
Abstract
We studied the dynamics of pure vergence shifts and vergence shifts combined with vertical and horizontal saccades. It is knownfrom earlier studies that horizontal saccades accelerate horizontal vergence. We wanted to obtain a more complete picture of theinteractions between version and vergence. Therefore we studied pure version (horizontal and vertical), pure vergence (divergenceand convergence) and combinations of both in five adult subjects with normal binocular vision and little phoria (B5°). The visualtargets were LED’s in isovergence arrays presented at two distances (35 and 130 cm) in a dimly lit room. Two targets werecontinuously lit during each trial and gaze-shifts were paced by a metronome. The two subjects with a strong monocularpreference made vergence eye movements together with small horizontal saccades during pure vergence tasks. The other subjects,who did not have a strong monocular preference, made pure vergence movements (without saccades). These findings suggest thatmonocular preferences influence the oculomotor strategy during vergence tasks. Vergence was facilitated by both horizontal andvertical saccades but vergence peak-velocity during horizontal saccades was higher than during vertical saccades. © 1998 Publishedby Elsevier Science Ltd. All rights reserved.
Keywords: Human; Binocular; Eye movements; Oculomotor; Scleral coils
1. Introduction
The switching of binocular fixation between objectsof interest is usually accomplished by a combination ofversion and vergence eye movements. The versionalcomponent, a saccade, accounts for the fast directionalshift and the vergence component of the movementre-aligns the eyes to compensate for changes in dis-tance. In natural environments, targets normally differin direction and in distance. In experimental conditionsit is theoretically possible to isolate version and ver-gence eye movements to study them separately.
Collewijn et al. [1,2] found that saccades betweenisovergent targets were accompanied by a transientchange in vergence. Horizontal and upward verticalsaccades were accompanied by a divergence-conver-
gence sequence while downward saccades showed initialconvergence. From the latter, they concluded thatchanges in vergence during vertical saccades could not(as previously suggested) be caused by a temporary lossof the vergence signal. What causes these changes invergence during saccades is still unclear. Collins et al.[3] measured a 30% greater eye rotation stiffness in thenasal than in the temporal direction. On the otherhand, they showed a 40% greater maximum active forcefor the medial rectus muscle than for the lateral rectusmuscle. We do not know if these maximum forces arerepresentative of the forces during normal eye move-ments. The difference in stiffness could, in contrast tothe force profiles, explain transient divergence duringhorizontal saccades. Enright suggested [4] that the up-down transient vergence asymmetry could be explainedby co-contraction of the vertical recti and (at the sametime) changes in tension of the superior oblique musclesduring vertical saccades. Whether transient vergence
* Corresponding author. Tel.: +31 10 4087558; fax: +31 104367594; e-mail: [email protected].
0042-6989/98/$19.00 © 1998 Published by Elsevier Science Ltd. All rights reserved.
PII: S0042-6989(98)00092-3

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543944
has a useful purpose during 3-D gaze shifts is unclear.Slow non-conjugate eye movements can occur when
two targets are, for instance, aligned in front of one ofthe eyes [5,6]. If only one eye moves, a change invergence and a small change in version result. This typeof movement does fit neither into the definition ofversion nor the definition of vergence. If one wants to fitthe movement into Hering’s Law, one should describe itas a combination of symmetrical vergence and slowversion [6]. In this paper we assume that this type ofmovement is functionally a vergence movement and wewill refer to this movement and other slow non-conju-gate horizontal movements as ‘asymmetrical vergence’.
The historical view, that combined gaze-shifts aresimply the result of summed outputs of the saccadic andthe vergence subsystem, has been gradually abandonedand transformed into a view of interaction between thesubsystems. Enright [7] reported that both horizontaland vertical saccades were effective in mediating largefractions of intended vergence changes. He also re-ported that residual (post-saccadic) vergence was usu-ally asymmetrical or even monocular. This complexbehaviour of vergence and version movements wasdifficult to fit into simple summation of the twomovements.
Enright [5] studied asymmetrical horizontal vergenceby aligning two targets in front of the non-preferred eye.He found that these targets elicited both vergence andsaccadic movements. He proposed that the step compo-nent for each eye depended only on that eye’s visualinput; and that the pulse components generated for eacheye depended on weighted averaging of visual stimuli ofboth eyes. In 1996 Enright found that symmetrical andasymmetrical convergence tasks could be accomplishedwithout saccades [6]. He concluded that convergencemovements could not be accounted for by a singlegenerator of binocular symmetrical input to the eyemuscles but that convergence consisted of two syn-chronous monocular components.
Erkelens et al. [8] Zee et al. [9] and Collewijn et al.[10,11] all found that combining vergence and versionaccelerated vergence and slowed down version. Theyproposed the existence of strong interactions betweenthe saccadic and the vergence subsystem. They rarelyobserved pure vergence movements. Most subjects madehorizontal small saccades when targets were set to elicitonly vergence. A possible function of the occurrence ofsmall horizontal saccades during pure vergence taskscould be the enhancement of vergence so that a newtarget is fixated more quickly. Another explanationcould be that the small saccades bring one of the eyesclose to or even on the target while the fellow eyefollows later. Furthermore, small saccades are simplynecessary if targets are not aligned exactly in a lineprotruding from the cyclopean eye (the point centrallybetween the eyes on the isovergence circle through the
eyes) in any direction. Therefore, experimental settingsto elicit pure vergence eye movements need great preci-sion.
A well-known interaction exists between accommoda-tion and vergence eye movements [12,13]. Duringmonocular viewing, accommodation of the viewing eyeproduces accommodation in the occluded eye and avergence movement. Enright [7] showed, by comparingmonocular and binocular gaze-shifts, that about onequarter of intra-saccadic vergence could be the result ofaccommodation cues. Han et al. [14] compared dynam-ics between the accommodative vergence movementstriggered by the preferred eye and the fellow eye ofsubjects. They found a different accommodation-ver-gence relationship when the non-preferred eye was fixat-ing with the other eye occluded, compared to theopposite. We did not investigate this relationship in thepresent study but accommodative vergence plays a rolein the vergence shifts that we measured. The assumptionthat this accommodative vergence component remainedconstant within subjects during the experiment justifiescomparing gaze shifts within subjects.
Considering previously reported asymmetries [15–18,28] related to eye preferences, we hypothesised thatalso the dynamics of vergence movements could relateto these preferences. A preference for one of the eyescan be detected in about 90 % of the population [19].This preference is often called ‘monocular preference’ or‘sighting dominance’. We use the terms ‘preference’ and‘preferred eye’ in this paper to avoid confusion with theclinical term ‘dominant eye’, the counterpart of the‘amblyopic eye’.
In 1997 Collewijn and colleagues [20] described gaze-shift trajectories and found pre-saccadic vergence move-ments while directional changes did not start until thesaccade began. From this they concluded that control ofthe vergence and version components of a gaze-shift canbe dissociated to some degree. Ocular vergence andversion systems could, therefore, process target vergenceand target direction separately. They proposed a stronginteraction between the two oculomotor activitieswhenever they occurred at the same time. Comparingthe vertical version-horizontal vergence relationship tothe horizontal version-vergence relationship within oneexperiment could show the interactive mechanisms moreclearly.
In our present experiment, we looked at version andvergence eye movements during gaze-shifts betweentargets positioned such as to elicit pure vergence, verti-cal version, horizontal version and combinations ofthese types of movements. From our results it seemslikely that there are separate systems for horizontalvergence and saccades with a strong interaction. Hori-zontal and vertical saccades both facilitate vergence butnot necessarily in the same way. Furthermore, we foundsupport for a relation between eye movement

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–3954 3945
Table 1Main subject characteristics are summarised in this table
Age (years) Correction PhoriaSubject Tube-testSex Ring-test IPD (mm) Experience
33 −4.5/−4.5 Eso1 R/RM R 66 ++30 None Exo L/R L 632 ++F30 None Exo L/RM ?3 65 ++
M4 22 None Eso L/L L 64 −5 F 31 None Exo L/R R 65 +
For the ‘tube-test’, L/R means that there was a symmetrical response and, therefore, no strong monocular preference. For the ‘ring-test’, L (lefteye) or R (right eye) indicates the preferred eye (fixating six to eight times out of eight), ‘?’ indicates inconclusive test-results (no preferencedetected).
asymmetries and monocular preferences during ver-gence tasks.
2. Methods
2.1. Subjects
Five adult subjects participated in our experiments.Four of them had previous experience with visual tasksin search coil measurements. All subjects underwentophthalmic and orthoptic examinations in the Rotter-dam Eye Hospital and additional tests in our depart-ment. The main results are shown in Table 1. They allhad a visual acuity of at least 20/20 in each eye and astereo-acuity of 40 s according to the Titmus stereo-test. All were emmetropic except subject 1 who wore hiscontact lenses during the measurement. None of thesubjects had a phoria larger than 5° (at near or farfixation) as measured with the cover test. Each subjectshowed alternating dominance of the eyes when testedwith the prism-test. Subject 1 had a slight leftwardrotation and tilt of his head of which he had never beenaware. Because of his completely normal binocularfunctions, we accepted this as a normal variation anddid not exclude him from our experiment.
To detect if subjects had a monocular preference, wedesigned a ‘tube-test’, which was comparable to classi-cal preference tests, such as described by Barbeito [15].In our test we gave the subjects two tubes, one in eachhand. We then asked them to look at a target througheach tube consecutively. We used two tubes instead ofone because we wanted to prevent subjects from usingtheir dominant hand each time they looked trough thetube. Subjects were instructed to keep each tube in thehand that we gave it in and were told that it did notmatter which eye they used for fixating the target.Looking through both tubes with the same eye wastaken as a sign of strong preference for that eye.Putting the left tube before the left eye and the righttube before the right eye was taken as absence of astrong preference. To detect weaker preference, wedesigned a separate test that we called the ‘ring-test’. In
this test, subjects had to fixate a self-chosen distantobject through a ring (diameter, 3 cm) that was fixed tothe window of our office on the fifteenth floor (distancebetween circle and eyes, 50 cm). We instructed thesubjects to keep both eyes open and to select a distantobject that fitted completely in the circle. Then wecovered one of the eyes and asked if the object was stillin the circle. This test was repeated eight times; fourtimes standing in front of the circle, twice standing leftof the circle and twice standing right of the circle.Because of the large distance, even subjects who couldnormally choose between suppressing the left and theright image, partly suppressed one of the ring-images. Ifthe subject used the same eye to fixate an objectthrough the ring at least six times, we concluded thatthere was a mild monocular preference for that eye.The results of both tests are shown in Table 1. Two ofthe subjects (1 and 4) showed a strong monocularpreference, two a mild preference (2 and 5); in subject3, we could not detect any preference with these tests.
2.2. Visual conditions
We used vertical and horizontal isovergent arrays ofreal LED targets. The central targets were straight-ahead at distances of 35 and 130 cm from the eyes. Wechose LED combinations to elicit versional saccades,pure vergence or a combination of both (Fig. 1). Eachtarget combination consisted of two continuously litLED’s, presented in dimly lit surroundings. The re-quired vergence shift between the two isovergence ar-rays was 7.7° for an inter-pupillary distance (IPD) of6.5 cm (7.1° for IPD of 6 cm, 8.2° for IPD of 7 cm).The near and far LED’s were perceived equally lumi-nous and comparable in angular size. In this way weminimised convergence-divergence differences due totarget inequality. The distant targets were not occludedby the nearby isovergence array.
2.3. Experimental procedure
Target combinations elicited saccades of 20 and 30°,symmetrical around the centre. We used target combi-
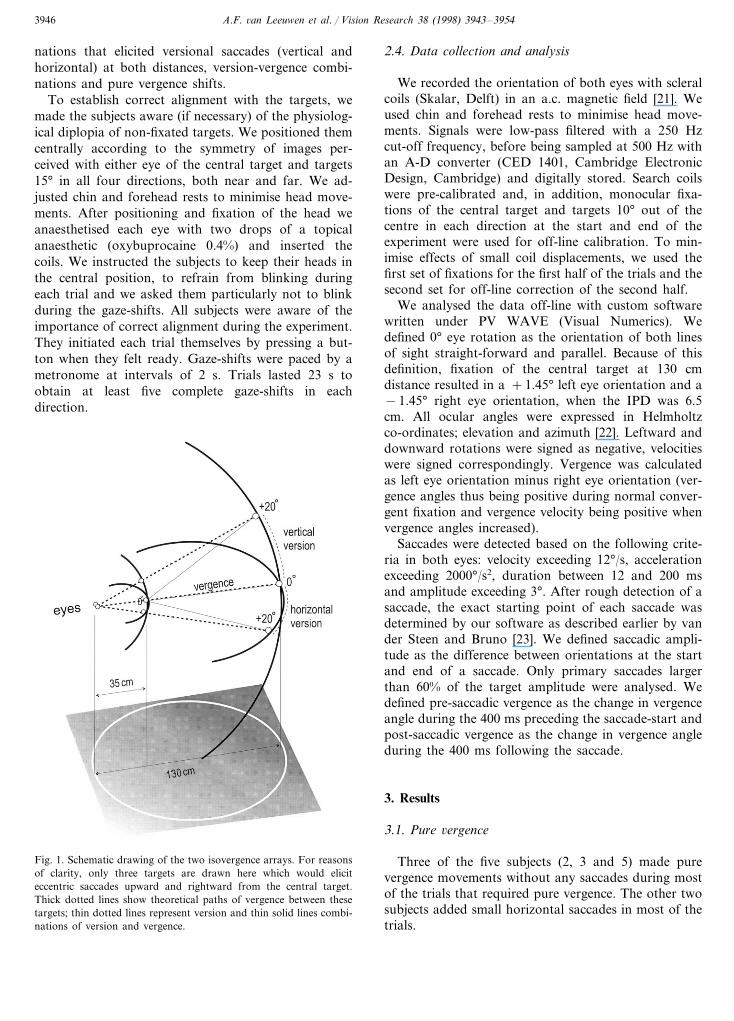
A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543946
nations that elicited versional saccades (vertical andhorizontal) at both distances, version-vergence combi-nations and pure vergence shifts.
To establish correct alignment with the targets, wemade the subjects aware (if necessary) of the physiolog-ical diplopia of non-fixated targets. We positioned themcentrally according to the symmetry of images per-ceived with either eye of the central target and targets15° in all four directions, both near and far. We ad-justed chin and forehead rests to minimise head move-ments. After positioning and fixation of the head weanaesthetised each eye with two drops of a topicalanaesthetic (oxybuprocaine 0.4%) and inserted thecoils. We instructed the subjects to keep their heads inthe central position, to refrain from blinking duringeach trial and we asked them particularly not to blinkduring the gaze-shifts. All subjects were aware of theimportance of correct alignment during the experiment.They initiated each trial themselves by pressing a but-ton when they felt ready. Gaze-shifts were paced by ametronome at intervals of 2 s. Trials lasted 23 s toobtain at least five complete gaze-shifts in eachdirection.
2.4. Data collection and analysis
We recorded the orientation of both eyes with scleralcoils (Skalar, Delft) in an a.c. magnetic field [21]. Weused chin and forehead rests to minimise head move-ments. Signals were low-pass filtered with a 250 Hzcut-off frequency, before being sampled at 500 Hz withan A-D converter (CED 1401, Cambridge ElectronicDesign, Cambridge) and digitally stored. Search coilswere pre-calibrated and, in addition, monocular fixa-tions of the central target and targets 10° out of thecentre in each direction at the start and end of theexperiment were used for off-line calibration. To min-imise effects of small coil displacements, we used thefirst set of fixations for the first half of the trials and thesecond set for off-line correction of the second half.
We analysed the data off-line with custom softwarewritten under PV WAVE (Visual Numerics). Wedefined 0° eye rotation as the orientation of both linesof sight straight-forward and parallel. Because of thisdefinition, fixation of the central target at 130 cmdistance resulted in a +1.45° left eye orientation and a−1.45° right eye orientation, when the IPD was 6.5cm. All ocular angles were expressed in Helmholtzco-ordinates; elevation and azimuth [22]. Leftward anddownward rotations were signed as negative, velocitieswere signed correspondingly. Vergence was calculatedas left eye orientation minus right eye orientation (ver-gence angles thus being positive during normal conver-gent fixation and vergence velocity being positive whenvergence angles increased).
Saccades were detected based on the following crite-ria in both eyes: velocity exceeding 12°/s, accelerationexceeding 2000°/s2, duration between 12 and 200 msand amplitude exceeding 3°. After rough detection of asaccade, the exact starting point of each saccade wasdetermined by our software as described earlier by vander Steen and Bruno [23]. We defined saccadic ampli-tude as the difference between orientations at the startand end of a saccade. Only primary saccades largerthan 60% of the target amplitude were analysed. Wedefined pre-saccadic vergence as the change in vergenceangle during the 400 ms preceding the saccade-start andpost-saccadic vergence as the change in vergence angleduring the 400 ms following the saccade.
3. Results
3.1. Pure 6ergence
Three of the five subjects (2, 3 and 5) made purevergence movements without any saccades during mostof the trials that required pure vergence. The other twosubjects added small horizontal saccades in most of thetrials.
Fig. 1. Schematic drawing of the two isovergence arrays. For reasonsof clarity, only three targets are drawn here which would eliciteccentric saccades upward and rightward from the central target.Thick dotted lines show theoretical paths of vergence between thesetargets; thin dotted lines represent version and thin solid lines combi-nations of version and vergence.

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–3954 3947
Fig. 2. Representative examples of horizontal eye orientations during pure vergence tasks of two subjects. The thin lines represent the left eye andthe right eye, the thick line vergence. Upward going traces represent either rightward movements or convergence movements. The lower panelshows the vergence velocities. Subject 1, who has a preferred right eye, makes small saccades during vergence shifts. During the divergence shift,his right eye is on the target first. Subject 2, who has no strong monocular preference, makes smooth pure vergence movements.
Convergence was faster than divergence in three sub-jects while the other two subjects showed no consistentdifference in peak-velocity between convergence anddivergence. The duration of pure vergence shifts rangedbetween 150 and 650 ms, was slightly shorter for con-vergence than for divergence and was not dependent onthe vergence peak-velocity. Fig. 2 shows a representa-tive example of two subjects’ vergence shifts in a trialwith pure vergence targets. Subject 1 usually addedsmall saccades while subject 2 usually did not.
Subject 1 made rightward saccades of around 3°during pure divergence tasks in all directions (0, 10 and15° eccentric, left, right, up and down). Only in the 15°leftward direction were his divergence movements pure.The small disjunctive saccades put his preferred eye onor within 1° of the target. Convergence movementswere executed mostly without any saccades by thissubject. Subject 4 showed leftward saccades duringdivergence shifts and rightward saccades during conver-gence shifts in pure vergence tasks in all directions. Inthis case the disjunctive saccades (of 2–3° version)usually aligned the preferred eye within 1° of the target.These small disjunctive direction-dependent saccadesoccurred consistently in these two subjects.
The other subjects (2, 3 and 5) made small saccadesonly occasionally during pure vergence tasks. Thesesaccades were usually smaller, more conjugate andmore variable in direction. They seemed of a moredirectionally corrective nature, comparable to smallsaccades during fixation. The subjects showed idiosyn-cratic changes in vertical vergence that were dependenton direction and horizontal vergence angle; these
changes never exceeded 2° and were not furtheranalysed.
Vergence angles during steady fixation were not thesame for all subjects. Part of this variation between thesubjects could be explained by differences in IPD. Weassume that some of the inaccuracy was due to the sizeof the stimuli and to small variations in head position.Small fixation errors are normal and not perceivedbecause of sensory fusion.
3.2. ‘Pure’ 6ersion
During saccades between isovergent targets, therewas always a transient divergence component as foundin earlier studies [1,2]. The magnitude and timing ofthis transient divergence was strongly idiosyncratic andvaried also with direction. Fig. 3 shows vergence-ver-sion plots during 20° horizontal (left panel) and vertical(right panel) isovergent saccades. Multiple gaze-shifts offive subjects for two isovergence angles (2.9 and 10.5°)are plotted. Vergence-version traces did not differ muchduring saccades at the two distances. We found slightlymore rightward versus leftward asymmetry in horizon-tal saccades between the nearby targets than betweenthe distant targets. Subjects 1, 2 and 3 had on averagea longer duration of 30° horizontal saccades at 10.5°isovergence than at 2.9° isovergence with, on average,equal peak-velocities. These differences in durationwere almost statistically significant (t-test) in subject 2(PB0.1), significant in subject 3 (PB0.05) and highlysignificant in subject 1 (PB0.01). The other two sub-jects showed a large variability in saccade duration.

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543948
Fig. 3. 2-D Plots of horizontal vergence and version during 20° symmetrical saccades between isovergent LED’s for all five subjects. The left panelshows horizontal saccades at the two distances, the right panel vertical saccades at the two distances. In this figure divergence is depicted positiveand convergence negative. In this way the plots resemble top-view trajectories. Leftward and downward version are depicted negative, rightwardand upward version are depicted positive. The wrong start and end-position during horizontal saccades at 10.5° isovergence of subject 1 areprobably due to an accidental misplacement of the targets.
Vertical saccades at the two distances also showedslight (usually non-significant) differences in durationbut these were more subject- and direction specific.
During horizontal saccades, all subjects showed ini-tial divergence starting together with the saccade andfollowed by compensatory convergence. At the end of asaccade, some subjects had already re-attained the re-quired vergence angle whereas others needed post-sac-cadic convergence to fixate the target binocularly.Upward saccades showed approximately the same tran-sient vergence sequence but downward saccades usuallyshowed different transient vergence traces. The be-haviour of subject 4, transient convergence with up-ward saccades and transient divergence with downwardsaccades, was compliant with the findings of Collewijnet al. [2,11] and Enright [4]. The other four subjectsshowed transient divergence with all vertical saccadesbut of a smaller magnitude during upward than duringdownward saccades [9]. The transient vergence during30° vertical and horizontal saccades had typically thesame characteristics as for 20° saccades for each sub-ject, being only slightly larger during larger saccades(see Fig. 7).
As described previously [1,2], saccades symmetricalabout the mid-position of these sizes were usually faster
in the horizontal than in the vertical direction. Unlikeprevious results, all subjects showed peak-velocity dif-ferences between symmetrical upward and downwardsaccades of equal amplitudes. Subjects 1, 2, 3 and 4 hadsignificantly faster and shorter upward than downwardsaccades, but subject 5 had much faster downward thanupward saccades.
3.3. Horizontal saccades with horizontal 6ergence
Fig. 4 shows representative traces for two subjects of20° horizontal saccades with vergence shifts (of about7° amplitude). The binocular saccades were unequal inamplitude, yet none of the saccades produced the de-manded vergence angles for fixation of the target. Afterthe saccade the vergence angle was corrected by a puresymmetrical or asymmetrical vergence movement and/or small disjunctive corrective saccades. Fig. 4 alsoshows that vergence velocities often had a double peakduring saccades.
The left panel of Fig. 5 shows vergence-version plotsfor each subject during horizontal saccades with ver-gence shifts. When a convergence shift was requiredwith a saccade some initial divergence, as during ver-sional saccades, was present. Also, this transient diver-

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–3954 3949
Fig. 4. Representative examples of horizontal eye orientations during 20° horizontal saccades between a nearby right target and a distant left targetof two subjects. The thin lines represent the left eye and the right eye, the thick line vergence. Upward going traces represent either rightwardmovements or convergence movements. The lower panel shows the vergence velocities. Notice the presence of pre-saccadic vergence in bothexamples.
gence component was larger during 30° than during 20°convergent saccades (Fig. 7). When a gaze-shift wasdivergent, the transient divergence seemed to be incor-porated in the total vergence shift during both 20 and30° saccades. As a consequence, all subjects showed afairly rectilinear version-vergence trace during divergenthorizontal saccades and a more curved version-ver-gence trace during convergent saccades.
Fig. 5 shows that usually a substantial part of therequired vergence was accomplished after the initialsaccade. The post-saccadic divergence shifts weresmaller after 30° saccades than after 20° saccades.During the larger saccades, the combined version-ver-gence movements lasted longer and more divergencewas accomplished during the saccade. This can be seenin Fig. 7, that shows disconjugate gaze-shifts of 20 and30° version of one subject. Post-saccadic convergencewas not smaller after larger saccades, probably due tothe larger transient divergence that had to be overcome.
Similar to findings by Takagi et al. [24] and byCollewijn et al. [20], we found small vergence move-ments in the required direction preceding the horizontalsaccades; so-called pre-saccadic vergence. We foundpre-saccadic divergence prior to all divergent saccadesin subjects 2, 3 and 5. Subject 1 only showed consistent
pre-saccadic divergence preceding rightward divergentsaccades and subject 4 showed no consistent pre-sac-cadic vergence before horizontal gaze-shifts. The aver-age pre-saccadic divergence movements per subjectaveraged 0.1–0.6° in magnitude. Pre-saccadic conver-gence was usually much smaller (subject averages 0.02–0.2°) but, nevertheless, consistently present in subject 1,2 and 3. The asterisks in Fig. 5 point out all occasionsof consistent pre-saccadic vergence in the requireddirection.
As expected, all the subjects had higher vergencepeak-velocities during horizontal combined version-ver-gence gaze-shifts than during pure vergence shifts of thesame magnitude. As can be seen in Fig. 8, divergencepeak-velocity became at least twice the pure divergencepeak-velocity. The difference between pure convergencepeak-velocity and convergence peak-velocity duringhorizontal saccades was smaller. Divergence and con-vergence peak-velocities were usually around 100°/sduring combined gaze-shifts.
Saccadic peak-velocity was smaller in the eye thatmade the smaller saccade; that is the abducting eyeduring a convergent shift or the adducting eye during adivergent shift. The fellow eye made slightly largersaccades than during pure version but saccadic peak-

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543950
Fig. 5. 2-D plots of combined version-vergence shifts of all five subjects during horizontal and vertical 20° saccades in both directions. The leftand right panel show horizontal and vertical saccades at the two distances. In this figure, divergence is depicted positive and convergence negative.Rightward and upward version are depicted positive. The arrows indicate the direction of the gaze-shifts. The asterisks indicate the occasions ofconsistent pre-saccadic vergence. Especially in subject 3, the pre-saccadic divergence is visible as a small initial non-directional shift at the startof the horizontal version-divergence trace.
velocity remained equal. As a consequence, versionpeak-velocity during horizontal disjunctive gaze-shiftswas lower than during horizontal isovergent gaze-shifts, as described previously by Collewijn and col-leagues [11]. Version velocity and duration had alarger variability during combined horizontal gaze-shifts than during isovergent horizontal saccades.
3.4. Vertical saccades with horizontal 6ergence
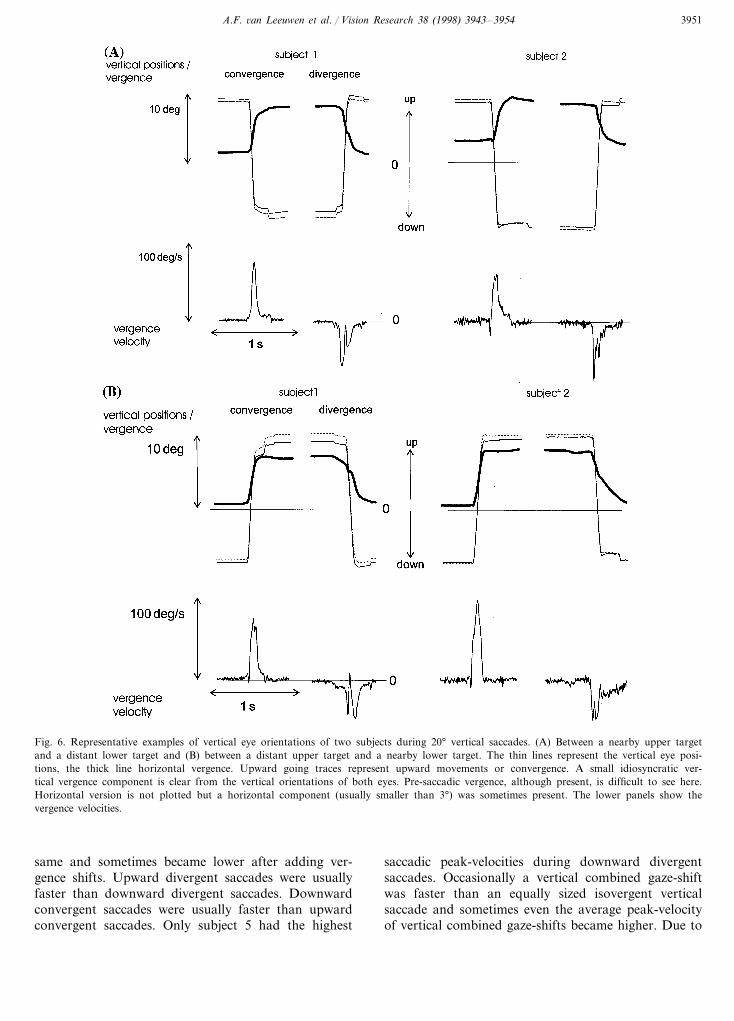
Fig. 6 shows typical traces for two subjects’ 20°vertical saccades with the convergence shift with theupward saccade and the divergence shift with thedownward saccade (A) and vice versa (B). In thisfigure vertical eye orientations and horizontal ver-gence are plotted. As during horizontal gaze-shifts,the vergence velocity often showed a double peak.
The version-vergence relations during vertical sac-cades, some of which are shown in the right panel ofFig. 5, were highly idiosyncratic. The asterisks againpoint out consistent pre-saccadic vergence in the re-quired direction. This pre-saccadic vergence was ofcomparable magnitude as the pre-saccadic vergence
observed preceding horizontal gaze-shifts and consis-tently present in each subject in at least one direction.
The version-convergence traces were not curvedconsistently more than version-divergence traces. Dur-ing convergent vertical saccades, initial divergence wasabsent in subjects 1 and 4 in both directions. Subject3 showed absence of transient divergence duringdownward convergent saccades only. When we com-pare these version-vergence traces with the corre-sponding traces during version shifts, it seems thatthe transient vergence during vertical version was in-corporated in the gaze-shifts whenever it was in theappropriate direction.
In four subjects (1, 2, 3 and 5) vergence peak-ve-locity was higher during vertical gaze-shifts than dur-ing pure vergence shifts but never as high as duringhorizontal gaze-shifts (Fig. 8). Subject 4 showed al-most the same vergence velocities after combinationwith vertical saccades as during pure vergence tasks.Divergence peak-velocity seemed linearly related tosaccadic peak-velocity; convergence peak-velocityseemed independent of saccadic peak-velocity.
Vertical saccadic peak-velocity often stayed the
A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–3954 3951
Fig. 6. Representative examples of vertical eye orientations of two subjects during 20° vertical saccades. (A) Between a nearby upper targetand a distant lower target and (B) between a distant upper target and a nearby lower target. The thin lines represent the vertical eye posi-tions, the thick line horizontal vergence. Upward going traces represent upward movements or convergence. A small idiosyncratic ver-tical vergence component is clear from the vertical orientations of both eyes. Pre-saccadic vergence, although present, is difficult to see here.Horizontal version is not plotted but a horizontal component (usually smaller than 3°) was sometimes present. The lower panels show thevergence velocities.
same and sometimes became lower after adding ver-gence shifts. Upward divergent saccades were usuallyfaster than downward divergent saccades. Downwardconvergent saccades were usually faster than upwardconvergent saccades. Only subject 5 had the highest
saccadic peak-velocities during downward divergentsaccades. Occasionally a vertical combined gaze-shiftwas faster than an equally sized isovergent verticalsaccade and sometimes even the average peak-velocityof vertical combined gaze-shifts became higher. Due to

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543952
Fig. 7. 2-D Plots of version-vergence shifts of one subject (3) during 20° and 30° horizontal (left panel) and vertical (right panel) saccades betweenisovergent targets (upper panel) and between the two distances (lower panel). In this figure, divergence is positive and convergence negative.Rightward and upward version are depicted positive. Notice the larger transient divergence during 30° saccades than during 20° saccades.
the larger variability of combined vertical gaze-shiftscompared to isovergent vertical saccades in all subjects,these differences were not statistically significant.
3.5. Comparison of 6ertical and horizontal non-conjugategaze-shifts
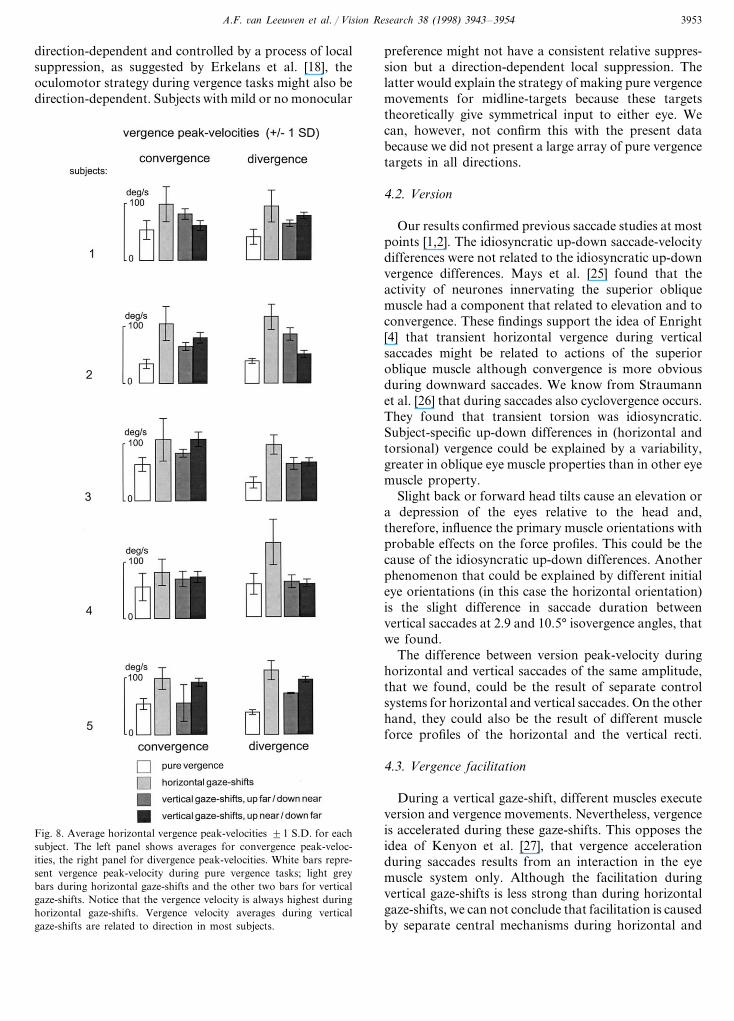
Pre-saccadic vergence was observed more often incombination with vertical saccades than with horizontalsaccades. The percentage of total vergence achieved dur-ing a saccade was larger during divergent than duringconvergent horizontal gaze shifts. During vertical sac-cades, the difference between divergent and convergentgaze-shifts was less clear and sometimes more vergencewas achieved during convergent than during divergentshifts. Fig. 8 shows the average vergence peak-velocitiesof each subject during all target combinations that re-quired a vergence shift. From this figure, clearly vergencepeak-velocity was consistently higher with horizontalthan with vertical saccades. Average vergence velocitiesduring vertical gaze-shifts were related to the directionin most subjects and are, therefore, depicted separately.
The vergence-version plots of convergent horizontalsaccades were consistently more curved than those ofdivergent horizontal saccades. During vertical gaze shifts,differences in version-vergence traces were more idiosyn-cratic. The more rectilinear traces seemed more efficient
in terms of interaction between vergence and version butthese traces did not always have the highest peak-veloc-ities for vergence and version.
4. Discussion
4.1. Pure 6ergence
Our findings suggest that monocular preferences playa role in the oculomotor strategy of subjects duringvergence tasks. Barbeito et al. [16] proposed that individ-ual dynamic asymmetries were related to the functionallocation of the cyclopean eye. Peli and McCormack. [17]found asymmetrical vergence movements after coveringone of the eyes. Uncovering the eye usually led to asaccadic response when the uncovered eye was thepreferred eye and to asymmetrical vergence when it wasthe non-preferred eye. This result can also be explainedby the strategy of initial target-fixation with the preferredeye and subsequent correction of the vergence angle.Subjects with this strategy might have a temporaryrelative suppression of the image of the non-preferredeye. Enright [6] found that most subjects had moresaccade-free trials during a vergence task when targetswere aligned in the midline than when targets werealigned with one of the eyes. If monocular preferences are
A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–3954 3953
direction-dependent and controlled by a process of localsuppression, as suggested by Erkelans et al. [18], theoculomotor strategy during vergence tasks might also bedirection-dependent. Subjects with mild or no monocular
preference might not have a consistent relative suppres-sion but a direction-dependent local suppression. Thelatter would explain the strategy of making pure vergencemovements for midline-targets because these targetstheoretically give symmetrical input to either eye. Wecan, however, not confirm this with the present databecause we did not present a large array of pure vergencetargets in all directions.
4.2. Version
Our results confirmed previous saccade studies at mostpoints [1,2]. The idiosyncratic up-down saccade-velocitydifferences were not related to the idiosyncratic up-downvergence differences. Mays et al. [25] found that theactivity of neurones innervating the superior obliquemuscle had a component that related to elevation and toconvergence. These findings support the idea of Enright[4] that transient horizontal vergence during verticalsaccades might be related to actions of the superioroblique muscle although convergence is more obviousduring downward saccades. We know from Straumannet al. [26] that during saccades also cyclovergence occurs.They found that transient torsion was idiosyncratic.Subject-specific up-down differences in (horizontal andtorsional) vergence could be explained by a variability,greater in oblique eye muscle properties than in other eyemuscle property.
Slight back or forward head tilts cause an elevation ora depression of the eyes relative to the head and,therefore, influence the primary muscle orientations withprobable effects on the force profiles. This could be thecause of the idiosyncratic up-down differences. Anotherphenomenon that could be explained by different initialeye orientations (in this case the horizontal orientation)is the slight difference in saccade duration betweenvertical saccades at 2.9 and 10.5° isovergence angles, thatwe found.
The difference between version peak-velocity duringhorizontal and vertical saccades of the same amplitude,that we found, could be the result of separate controlsystems for horizontal and vertical saccades. On the otherhand, they could also be the result of different muscleforce profiles of the horizontal and the vertical recti.
4.3. Vergence facilitation
During a vertical gaze-shift, different muscles executeversion and vergence movements. Nevertheless, vergenceis accelerated during these gaze-shifts. This opposes theidea of Kenyon et al. [27], that vergence accelerationduring saccades results from an interaction in the eyemuscle system only. Although the facilitation duringvertical gaze-shifts is less strong than during horizontalgaze-shifts, we can not conclude that facilitation is causedby separate central mechanisms during horizontal and
Fig. 8. Average horizontal vergence peak-velocities 91 S.D. for eachsubject. The left panel shows averages for convergence peak-veloc-ities, the right panel for divergence peak-velocities. White bars repre-sent vergence peak-velocity during pure vergence tasks; light greybars during horizontal gaze-shifts and the other two bars for verticalgaze-shifts. Notice that the vergence velocity is always highest duringhorizontal gaze-shifts. Vergence velocity averages during verticalgaze-shifts are related to direction in most subjects.

A.F. 6an Leeuwen et al. / Vision Research 38 (1998) 3943–39543954
vertical saccades. If vergence facilitation is caused by onecentral mechanism during both horizontal and verticalsaccades, the resulting vergence peak-velocities do nothave to be of the same magnitude. The vergence facilita-tion during horizontal saccades could be the result of thesame central facilitation as during vertical saccades butwith an extra facilitation because of activation of thesame muscles twice, by two different efferent mecha-nisms.
The pre-saccadic vergence that we found precedingboth horizontal and vertical saccades supports the sug-gestion of different subsystems for vergence and version.It does not support the gating model by Zee et al. [9],unless the two movements had different reaction times.Our results strongly support the ideas of Collewijn et al.[20], that the vergence system and the saccadic system actseparately, but interact with each other whenever theyoccur at the same time. If we assume the existence of aversion oculomotor system with subsystems for horizon-tal and vertical version and a vergence oculomotorsystem with subsystems for convergence and divergence,we can explain asymmetries within subjects. Variationsin subsystem characteristics together with variations inorbital anatomy, muscle insertion and muscle stiffnessseem a reasonable explanation for the differences be-tween subjects.
5. Final conclusions
Our results support the existence of different oculomo-tor systems for version and vergence with a centralinteraction between the two when both systems are activeat the same time. Furthermore, our results suggest thatmonocular preferences influence oculomotor strategiesduring vergence tasks.
Acknowledgements
We thank the subjects for their patience, the Rotter-dam Eye Hospital for orthoptic evaluation and D. van‘t Ent and J. van der Steen for their useful suggestionsand software-support.
References
[1] Collewijn H, Erkelens CJ, Steinman RM. Binocular co-ordinationof human horizontal saccadic eye movements. J Physiol London1988;404:157–82.
[2] Collewijn H, Erkelens CJ, Steinman RM. Binocular co-ordinationof human vertical saccadic eye movements. J Physiol London1988;404:183–97.
[3] Collins CC, Carlson MR, Scott AB, Jampolsky A. Extraocularmuscle forces in normal subjects. Invest Ophthalmol Vis Sci1981;20:652–64.
[4] Enright JT. Convergence during human vertical saccades: proba-ble causes and perceptual consequences. J Physiol London1989;410:45–65.
[5] Enright JT. The remarkable saccades of asymmetrical vergence.Vis Res 1992;32:2261–76.
[6] Enright JT. Slow-velocity asymmetrical convergence: a decisivefailure of ‘Hering’s Law’. Vis Res 1996;36:3667–84.
[7] Enright JT. Facilitation of vergence changes by saccades: influ-ences of misfocused images and of disparity stimuli in man. JPhysiol London 1986;371:69–87.
[8] Erkelens CJ, Steinman RM, Collewijn H. Ocular vergence undernatural conditions. II Gaze shifts between real targets differing indistance and direction. Proc R Soc London B 1989;236:441–65.
[9] Zee DS, Fitzgibbon EJ, Optican LM. Saccade-vergence interac-tions in humans. J Neurophysiol 1992;68:1624–41.
[10] Collewijn H, Erkelens CJ, Pizlo Z, Steinman RM. Binocular gazemovements: coordination of vergence and version. In: Ygge J,Lennerstrand G, editors. Eye Movements in Reading. Oxford:Pergamon, 1994:97–115.
[11] Collewijn H, Erkelens CJ, Steinman RM. Voluntary binoculargaze-shifts in the plane of regard: dynamics of version andvergence. Vis Res 1995;35:3335–58.
[12] Ciuffreda KJ, Kenyon RV. Accommodative vergence and accom-modation in normals, amblyopes and strabismics. In: Schor CM,Ciuffreda KJ, editors. Vergence Eye Movements: Basic andClinical Aspects. Woburn, USA: Butterworth, 1983:101–73.
[13] Eadie AS, Carlin PJ. Evolution of control system models of ocularaccommodation, vergence and their interaction. Med Biol EngComput 1995;33:517–24.
[14] Han Y, Seideman M, Lennerstrand G. Dynamics of accommoda-tive vergence movements controlled by the dominant and non-dominant eye. Acta Ophthalmol Scand 1995;73:319–24.
[15] Barbeito R. Sighting dominance: an explanation based on theprocessing of visual direction in tests of sighting dominance. VisRes 1981;21:855–60.
[16] Barbeito R, Tam WJ, Ono H. Two factors affecting saccadicamplitude during vergence: the location of the cyclopean eye anda left–right bias. Ophthal Physiol Opt 1986;6:201–5.
[17] Peli E, McCormack G. Dynamics of cover test eye movements.Am J Optom Physiol Opt 1983;60:712–24.
[18] Erkelens CJ, Muijs CE, Van Ee R. Binocular alignment in differentdepth planes. Vis Res 1996;36:2141–7.
[19] Purves D, White LE. Monocular preferences in binocular viewing.Proc Natl Acad Sci USA 1994;91:8339–42.
[20] Collewijn H, Erkelens CJ, Steinman RM. Trajectories of thehuman binocular fixation point during conjugate and non-conju-gate gaze-shifts. Vis Res 1997;37:1049–69.
[21] Robinson DA. A method of measuring eye movements using ascleral coil in a magnetic field. IEEE Trans Biomed Eng1963;BME-10:137–45.
[22] Carpenter RHS. Movements of the Eyes, 2nd Edition. London,England: Pion, 1988:139–81.
[23] Van der Steen J, Bruno P. Unequal amplitude saccades producedby aniseikonic patterns: effects of viewing distance. Vis Res1995;35:3459–71.
[24] Takagi M, Frohman EM, Zee DS. Gap-overlap effects on latenciesof saccades, vergence and combined vergence-saccades in humans.Vis Res 1995;35:3373–88.
[25] Mays LE, Zhang Y, Thorstad MH, Gamlin PD. Trochlear unitactivity during ocular convergence. J Neurophysiol 1991;65:1484–91.
[26] Straumann D, Zee DS, Solomon D, Lasker AG, Roberts DC.Transient torsion during and after saccades. Vis Res1995;35:3321–34.
[27] Kenyon RV, Ciuffreda KJ, Stark L. Unequal saccades duringvergence. Am J Optom Physiol Opt 1980;57:586–94.
[28] Porac C, Coren S. Sighting dominance and egocentric localisa-tion. Vis Res 1986;26:1709–13.