the inheritance of intraerythrocytic sodium level
TRANSCRIPT

American Journal of Medical Genetics 29193-203 (1988)
The Inheritance of lntraerythrocytic Sodium Level
Sandra J. Hasstedt, Steven C. Hunt, Lily L. Wu, and Roger R. Williams
Departments of Human Genetics (S. J. H.), Medicine (S. C. H., R. R. W.), and Pathology (L.L. W.), University of Utah, Salt Lake City, Utah
Intraerythrocytic sodium level (RBC Na) was measured on 1,800 normotensive members of 64 Utah pedigrees ascertained through hypertensive or normotensive probands, sibs with early stroke death, or brothers with early coronary disease. Likelihood analysis provided evidence that RBC N a was determined by four alleles at a single locus. Each allele was recessive to all alleles associated with a lower mean level. The four resultant distributions occurred in the frequencies: 0.8%, 89.3%, 9.7%, and 0.2% with corresponding means (mmol/l RBC) of 4.32, 6.67, 9.06, and 12.19, respectively. The major locus explained 29.0% of the variance in RBC Na; polygenic inheritance explained another 54.6%. A higher frequency of the genotypes for high RBC N a in pedigrees when the proband was hypertensive than normotensive provided evidence that this major locus increases susceptibility to hypertension.
Key w o r k hypertension, genetic, pathophysiology, epidemiology
INTRODUCTION
Intracellular sodium level and the rate at which it is transported through the membrane by various systems differ between persons with essential hypertension and controls [Hilton, 19861. Therefore, a defect in sodium transport may contribute to a predisposition to essential hypertension. Such a defect, once identified, would provide clues to the underlying pathophysiology of the disease, as well as identifying persons predisposed to a specific form of essential hypertension [Blaustein, 19841. Sodium-lithium counter- transport (SLC) is one transport system for which higher rates are seen in hypertensives [Canessa et al., 19801. We previously reported evidence of recessive inheritance of elevated SLC associated with an increased prevalence of hypertension [Hasstedt et al., 19871. However, since many presumed homozygotes for this recessive allele were normotensive, this locus qualifies as a susceptibility gene, as opposed to a disease gene, for hypertension.
Received for publication May 16, 1987; revision received August 7, 1987.
Address reprint requests to Sandra J. Hasstedt, PhD, Department of Human Genetics, 501 Wintrobe Bldg., University of Utah Medical Center, Salt Lake City, UT 84132.
0 1988 Alan R. Liss, Inc.

194 Hasstedtetal.
The sodium content of the cell is another variable that may identify gene-carriers susceptible to hypertension. Elevation of intraleukocytic sodium is a consistent finding in hypertensives [Hilton, 19861; elevation of intraerythrocytic sodium (RBC Na) has been found in some studies [Losse et al., 198 1; Fadeke Aderounmu and Salako, 1979; D’Amico, 1958; Clegg et al., 1982; Wessels and Zumkley, 1980 Montanari et al., 19801, but not in others [Weller, 1959; Bramley et al., 1986a,b; Simon and Conklin, 19861. Small sample sizes may explain why some studies found an insignificant elevation [Hilton, 19861, or failure to adjust or control for age may explain why some studies found a significant elevation [Bramley et al., 1986a,b]; since higher levels were seen when the hypertensive patients had a family history of hypertension [Clegg et al., 19821, heterogeneity within the hypertensives may have obscured the difference in some studies.
For RBC Na to qualify as a preclinical marker, elevated levels must occur in some normotensive persons at risk of hypertension. Elevated RBC Na has been reported in normotensive first degree relatives of hypertensive individuals [ Henningsen and Nelson, 1980; Wessels, 1980; Losse et al., 1981; Herlitz et al., 1983; Lijnen et al., 1984; Gudmundsson et al., 19841, but was not found in males under 30 years [Wessels, 19801, after salt loading [Gudmundsson et al., 19841, or in offspring of mothers with hypertension in pregnancy who later developed sustained essential hypertension [Svensson and Sigstrom, 19861. RBC Na is highly familial in normotensives. Sib-sib and parent-offspring correlations exceed 0.30 [Dadone et al., 1987; Hunt et al., 19871, and heritability was estimated as 0.47 and as 0.58 in adults only, after allowing for familiality due to living together in the same household and attending clinic on the same day [unpublished results].
Here we present an analysis of 1,800 persons in 64 pedigrees ascertained through hypertensive and normotensive probands, sibs who died of strokes, or brothers with early coronary disease. Likelihood analysis was used to test for evidence that alleles at a major locus determine RBC Na.
MATERIALS AND METHODS
Ninety-six pedigrees were ascertained through two or three sibs who died of stroke before age 74, hypertensive and normotensive probands from the Salt Lake Center of the Hypertension Detection and Followup Program, or two brothers with onset of coronary disease before age 55. About 2,500 members of the pedigrees visited the Utah Cardiovas- cular Genetics Clinic twice to participate in studies of the genetic epidemiology of hypertension [Williams et al., 19841 and coronary heart disease [Williams et al., 19791. Each local relative who agreed to participate completed a questionnaire detailing lifestyle and medical information, donated blood and urine for laboratory tests, and was examined for cardiovascular disease. The interval between visits was two to three years.
As part of the second screening cycle, about one-fourth of the sample were selected for and agreed to follow a salt-loading protocol by taking 12 g of sodium chloride as tablets during the 36 h prior to a 12 h overnight urine collection and eating a salty breakfast before visiting the clinic. During the first year of screening, all healthy subjects aged 12-50 were asked to follow this protocol; during subsequent years, nuclear families living in the same household were randomized and half were selected for salt-loading.
RBC Na was measured by atomic absorption spectroscopy [Hentschel et al., 19861. SLC was measured as the rate of sodium-dependent lithium efflux from lithium-loaded red blood cells by the method of Canessa et al. [1980] as adapted by Smith et al. [1982].

The Inheritance of RBC Na 195
The number of ouabain binding sites was measured by the method of Deluise et al. [ 19801 as modified by Ash et al. [1983] to increase the binding and decrease the experimental error. The sodium-potassium cotransport rate constant was measured as described by Smith et al. [1984].
The sample included 155 medicated hypertensives who were measured for RBC Na, but excluded from the analysis because diuretics lower RBC Na. Linear regression of RBC Na on sex and powers of age, performed separately on juveniles (age < 18 years, N =
753) and normotensive adults (N = 1047), allowed explanation of 4% and 696, respectively, of the variance of RBC Na. Levels on each person were adjusted for sex and age. Significant skewness (g = 0.892, P < 0.01) was removed by natural logarithm transformation. After the additional exclusion of any person without a measured relative, the sample consisted of 1,800 normotensive members of 64 pedigrees. Twenty-eight pedigrees contained fewer than 10 members; 21 pedigrees contained 11 to 50 members; 12 pedigrees contained 51 to 100 members; and 3 pedigrees contained over 100 members.
Likelihood analysis [Elston and Stewart, 19711 was used to test for evidence that an allele at a major locus determines extreme levels of RBC Na. Likelihoods were computed using PAP [Hasstedt and Cartwright, 19811 and the maxima obtained with GEMINI [Lalouel, 19791. No ascertainment correction was made, since the pedigrees were ascertained through characteristics other than, although related to, the variable used in the analysis. Hypothesis testing was performed by comparing the likelihood of a submodel to the likelihood of a more general model. Under certain conditions, negative two multiplied by the natural logarithm of the ratio of the likelihoods has an asymptotic chi-square distribution. Although all the conditions may not be met in this application of likelihood analysis [Cannings et al., 19801, chi-square statistics are presented, but must be interpreted with caution.
The mixed model [Elston and Stewart, 197 1; Morton and MacLean, 19741 was used in the analysis. The model assumed the phenotype was independently determined by a major locus with a large effect, additive polygenes each with a small effect, and random environmental factors. The major locus was assumed to have n, alleles, where n, = 2,3, or 4. The polygenic and random environmental components were assumed to be normally distributed, Hardy-Weinberg equilibrium determined the genotypic frequencies. The 2-allele model included the transmission probabilities as parameters [Boyle and Elston, 1979; Lalouel et al., 19831. Likelihoods of the mixed model were approximated [Hasstedt, 19821.
The parameters of the model were pi, the frequency of allele i = 1, . . . , n,, yij, mean of the major locus genotype composed of alleles i and j, i 5 j, H,, the plygenic heritability, and u, the phenotypic standard deviation within major locus genotypes. H, is the proportion of the variance within each major locus genotype attributed to polygenes. The parameters T ~ ~ , T , ~ , T ~ ~ , parent-to-offspring transmission probabilities for the three genotypes, were also parameters when n, = 2.
Major locus genotypic probabilities were computed for each pedigree member. The likelihood of the genetic model was broken into components, each conditioned on a different genotype for that person and the relative value of each component taken as his/her genotypic probability. In this computation, the parameters of the genetic model were fixed at their maximum likelihood estimates.
Estimates of parameters within major locus genotypes were computed using the genotypic probabilities as weighting factors as described in Hasstedt and Moll [ 19871. The parameters estimated included genotypic means of RBC Na in the original scale and of

196 Hasstedt et al.
the sodium transport variables, the genotypic frequency within subgroups of the sample, and the correlation due to the major locus between RBC Na and other variables.
RESULTS
Table I presents means and standard deviations of RBC Na and SLC within subgroups of our sample. Unadjusted means are presented separately for males and females, by age group in normotensives, and in hypertensives. Mean RBC Na was higher in medicated hypertensive than in normotensive adults despite the fact that other studies have shown that diuretics lower levels. Nonetheless, medicated hypertensives were excluded from the analysis. Although salt-loading may lower RBC Na in some subgroups, the mean level of adjusted RBC Na did not differ significantly (t = 0.71, P > 0.05), and salt-loading status was ignored in this analysis. Hypertensive subjects with high SLC (> 0.4 mmol/l RBC/hr) had mean RBC Na lower than in the other hypertensive subjects but higher than in normotensives.
TABLE I. Means and Standard Deviations of Unadjusted RBC Na and SIX3 Within Submow of the %In&
RBC Na (mmol/l RBC) SLC (mmol/l RBC/hr)
Males Females Males Females
Subaour, N Z s N K s N K s N S I S
Juveniles i 12 yrs Juveniles 12-1 8 yrs
Not salt-loaded Salt-loaded
Normotensive adults Not salt-loaded Salt-loaded
Hypertensives SLC > 0.4 SLC < 0.4
231 6.98 1.35 211 6.88 1.45 195 0.255 0.085 185 163 6.88 1.31 148 6.30 1.31 158 0.265 0.106 144 86 6.91 1.51 71 6.56 1.47 82 0.256 0.096 68 77 6.84 1.06 77 6.06 1.09 76 0.275 0.115 76
529 7.09 1.45 518 6.44 1.28 511 0.296 0.101 502 341 7.09 1.53 337 6.48 1.28 330 0.303 0.104 325 188 7.08 1.30 181 6.35 1.28 181 0.283 0.095 177 79 7.74 1.78 76 7.58 1.52 78 0.350 0.134 71 25 7.54 1.50 17 7.37 1.47 53 7.79 1.89 54 7.71 1.55
0.248 0.248 0.243 0.252 0.260 0.266 0.250 0.339
0.087 0.075 0.076 0.074 0.095 0.098 0.089 0.153
The effect of a major locus on a quantitative phenotype is to produce a phenotypic distribution which is a mixture of two or more components. Likelihood analysis suggested that the RBC Na distribution consisted of a mixture of component distributions. The rarer of two components in a mixture was designated the minor component and contrasted to the other. The three different minor components in these data were: (1) one with a higher mean and a frequency around 10% (designated common), (2) one with a higher mean and a frequency around .2% (designated rare), and (3) one with a lower mean and a frequency around 3% (designated low). Chi-square statistics testing the presence of an allele responsible for each minor component are given in Table 11. The hypotheses of no major locus and of dominant inheritance were rejected in all cases; recessive inheritance was also rejected for the rare minor component. Therefore, the estimates of the transmission probabilities given in Table I11 assumed recessive inheritance for the common and low minor components and dominan t inheritance for the rare minor component. For the common and low minor components, environmental transmission, but not Mendelian transmission, could be rejected, providing support for major locus inheritance. For the rare

The Inheritance of RBC Na 197
TABLE II. chi-square Statistics to Test Two Allele Major Locus Hypotheses by Type of Minor Component
Hypothesis" Common Rare LOW d.f.
No major locus (pI = 1) 55.08' 54.49' 21.29' 3 Recessivity (pll = H ~ ~ ) 0.18 34.83" 0.01 1 Dominance ( B , ~ = pu) 7.4Ob 54.49" 16.76' I Mendelian (ill = 1, T~~ = .5, T~~ = 0) 2.32 14.63' 0.22 3 Environmental (pl = T~~ = iI2 = i2J 17.89' 17.78' 17.36' 3
'See text for definition of the parameters. b P < 0.01, 'P < 0.005.
TABLE m. Estimates with Standard Errors of the Transmission Probabilities for the Three Alleles, bv Tvoe of Minor Comwnent
Minor component Transmission ____ probability Common Rare LOW'
71 I 0.978 * .031 0.991 + .004 0.618 712 0.592 ,066 0.610 + .081 0.546 7 2 2 0.000 + .098 1 .o 0.000
'The likelihood surface was too flat to obtain standard errors.
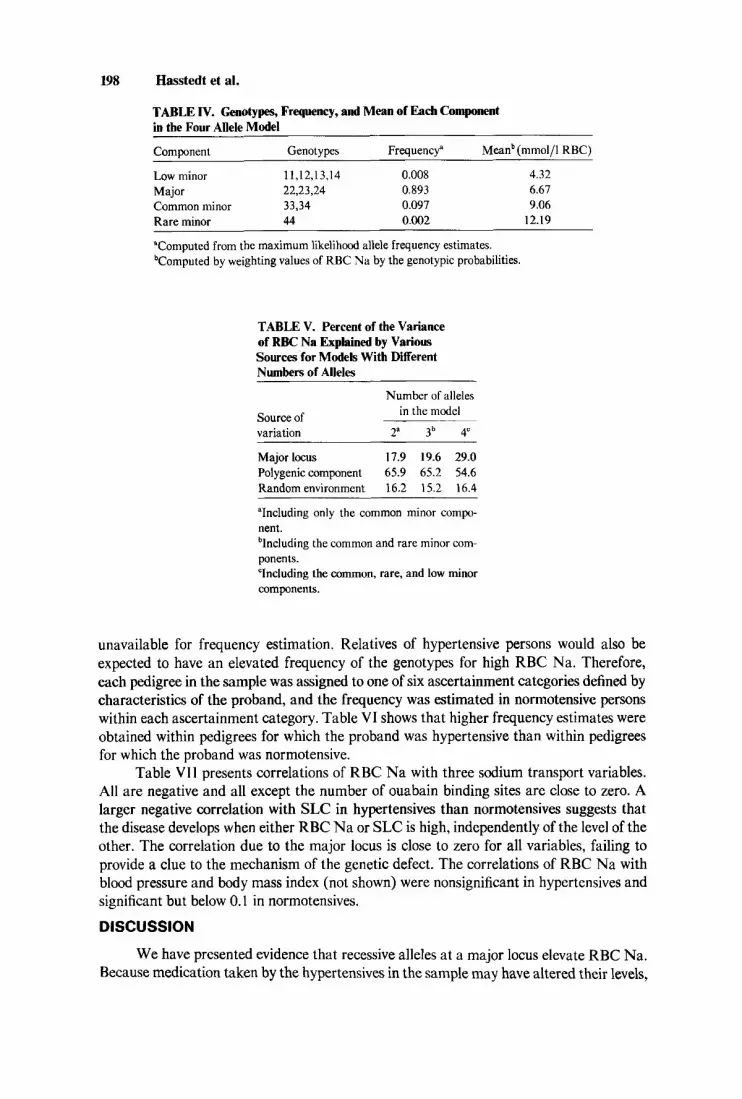
minor component, both Mendelian and environmental transmission could be rejected, indicating that the genetic model did not fit. The mean and frequency estimated for heterozygotes in this model were similar to the mean and frequency of the common minor component. But the common minor component appeared to consist of homozygotes since recessive inheritance was indicated. If the common and rare components were due to different recessive alleles, that would explain why the codominant model did not fit. Therefore, a three allele model was formulated in which two recessive alleles elevated RBC Na, one to the mean of the common minor component, and the second to the mean of the rare minor component. With the three allele model as the alternate hypothesis, the two allele recessive submodels were rejected for the common minor component (xz, = 13.48, P < .005), and the rare minor component (x& = 48.73, P < .005). To include the low minor component as well, a four allele model was formulated with each allele recessive to all alleles associated with a lower mean. With the four allele model as the alternate hypothesis, the three allele submodel was rejected (x& = 22.10, P < 0.005). For the four allele model, the allele frequencies were estimated as p, = .004 f .003, @z = .68 1 * .040, jj3 = .267 f .043, and the polygenic heritability was estimated as Hp = ,769 * .037. Table IV gives the estimated mean and frequency of each distribution. Table V shows that the polygenic component accounted for the majority of the variance in RBC Na, regardless of the number of alleles in the model.
Alleles which increase RBC Na may also increase susceptibility to hypertension. In that case, hypertensive persons would have an elevated frequency of the genotypes for high RBC Na. Since hypertensive persons were excluded from the sample, they were
198 Hasstedt et al.
TABLE IV. Genotypes, Frequency, and Mean of Each Component in the Four Allele Model
Component Genotypes Frequency" Meanb (mmol/l RBC)
Low minor 11,12,13,14 0.008 4.32 Major 22,23,24 0.893 6.61 Common minor 33,34 0.097 9.06 Rare minor 44 0.002 12.19 ~
"Computed from the maximum likelihood allele frequency estimates. bComputed by weighting values of RBC Na by the genotypic probabilities.
TABLE V. Percent of the Variance of RBC Na Explained by Various Sources for Models With Different Numbers of Alleles
Number of alleles in the model Source of
variation 2a 3b 4"
Major locus 17.9 19.6 29.0 Polygenic component 65.9 65.2 54.6 Random environment 16.2 15.2 16.4
"Including only the common minor compo- nent. blncluding the common and rare minor com- ponen ts. "Including the common, rare, and low minor components.
unavailable for frequency estimation. Relatives of hypertensive persons would also be expected to have an elevated frequency of the genotypes for high RBC Na. Therefore, each pedigree in the sample was assigned to one of six ascertainment categories defined by characteristics of the proband, and the frequency was estimated in normotensive persons within each ascertainment category. Table VI shows that higher frequency estimates were obtained within pedigrees for which the proband was hypertensive than within pedigrees for which the proband was normotensive.
Table VIl presents correlations of RBC Na with three sodium transport variables. All are negative and all except the number of ouabain binding sites are close to zero. A larger negative correlation with SLC in hypertensives than normotensives suggests that the disease develops when either RBC Na or SLC is high, independently of the level of the other. The correlation due to the major locus is close to zero for all variables, failing to provide a clue to the mechanism of the genetic defect. The correlations of RBC Na with blood pressure and body mass index (not shown) were nonsignificant in hypertensives and significant but below 0.1 in normotensives.
DISCUSSION
We have presented evidence that recessive alleles at a major locus elevate RBC Na. Because medication taken by the hypertensives in the sample may have altered their levels,

The Inheritance of RBC Na 199
TABLE VI. Estimated Summed Frequency (as Percent) of the Three Genotypes Associated With High Levels Classiied by Ascertainment Category
Number of
Ascertainment category Pedigrees Persons Frequency
Hypertensive, SLC > .4 6 101 17.08 Coronary, hypertensive 5 233 13.56 Hypertensive, SLC i .4 25 306 13.54 Stroke 8 393 10.34 Normotensive 7 48 8.35 Coronary, normotensive 13 719 6.10 Total 64 1800 9.93
TABLE W. Correlations of RBC Na With Sodium Transport Variables in Hypertensives (HBP), Normotensives (NBP), and Between the Major Locus Means in Normotensives (ML)
Variable HBP N BP ML'
Number of ouabain binding sites -0.37b -0.43b -0.12 Sodium-potassium cotransport rate -0.05 -0.08b -0.05 SLC -0.15" - 0.04" -0.02
"P < 0.05. bP < 0.001. T h e correlations between the means for the major locus genotypes.
we could not obtain direct evidence that the same alleles increase susceptibility to essential hypertension. Indirect evidence came from a higher frequency of genotypes for elevated RBC Na in members of pedigrees ascertained through hypertensive probands than those ascertained through normotensive probands. Considering that some of the pedigrees were large, and that the inheritance was recessive, the trend was surprisingly strong.
The genetic model used to test for a major locus is usually assumed to have one locus and two alleles. This model can be extended to either a three allele model or a two locus model. Both may improve the likelihood, regardless of which better repregents the genetic transmission. Since one is not a submodel of the other, their likelihoods cannot be directly compared. The rejection in this analysis of two and three allele models in favor of a four allele model does not rule out a multiple locus model which was not tested. Since a four allele model has 10 genotypic means, testing all possible equivalences to determine dominance reltionships is cumbersome. Of the many possible dominance relationships, only the one suggested by other analyses was tested, as in previous analysis with the three allele model for Lp(a) [Hasstedt and Williams, 19861. The hypothesis of Mendelian transmission was not tested in the four allele model because of the large number of possible transmission probabilities.
Since blood pressure is usually considered to be a polygenic trait, hypertension, occurring upon exceeding a blood pressure threshold, would also exhibit polygenic inheritance [Ward, 19831. However, alleles at two major loci, expressed as elevated RBC

200 Hasstedt et al.
Na (from this analysis) and as elevated SLC [Hasstedt et al., 19871, appear to increase susceptibility to hypertension. Both are incompletely penetrant for hypertension, both have large polygenic components for the quantitative trait, and RBC Na apparently has four alleles. These two loci, possibly along with two or three more major susceptibility loci for hypertension, could imitate polygenic inheritance.
Genotypes at the RBC Na locus and the SLC locus [Hasstedt et al., 19871 could increase risk of hypertension independently, or interactively. In the first case, one subgroup of hypertensives would have high SLC and normal RBC Na, and another would have high RBC Na and normal SLC. In the second case, persons with high SLC and hypertension would also have high RBC Na and vice versa, ie, the second locus would affect penetrance. Our findings which provide information about the relationship are (1) a negative correlation between RBC Na and SLC in hypertensive subjects, (2) elevated RBC Na in hypertensives with high SLC, and (3) a high frequency of the elevated RBC Na genotypes in pedigrees ascertained through hypertensive probands with high SLC. The most consistent explanation is an interaction between the two loci, as well as other genetic or environmental factors.
A defect in sodium transport must be responsible for the variation in RBC Na determined by the major locus, but our data did not reveal the transport system. The largest correlation was with the number of ouabain binding sites. Ouabain binding (reflecting the number of ATPase binding sites) and Na+-K+ ATPase enzymatic activity both contribute to the rate of the sodium pump (net ouabain-sensitive efflux) [Schmalzing et al., 1981; Beutler et al., 19831. Other studies have found RBC Na inversely correlated with the number of ouabain binding sites or pump units [Schmalzing et al., 1981; Webster et al., 1984; Lasker et al., 19851, ouabain-sensitive efflux [Cumberbatch and Morgan, 198 11, Na+-K+-ATPase activity, and furosemide-sensitive Na efflux and the ouabain- resistant fraction of 86Rb-uptake [Lijnen et al., 19841. Although other studies found RBC Na correlated with blood pressure [Webster et al., 19841, weight [Bramley et al., 1986a,b], and suprailiac skinfold thickness [Dadone et al., 19871, we found only weak correlations with blood pressure and body mass index in normotensives.
Salt-loading did not affect the mean level of RBC Na, in agreement with variance components analysis of the same data [unpublished results]. However, familial correla- tions on these data differed depending on salt-loading status [Hunt et al., 19871. Similarly, the literature on dietary sodium contains mixed findings. Sodium loading has produced higher levels of RBC Na in normotensives and hypertensives [Myers et al., 19831, no change whether the person was salt-sensitive or not (increase from 100 mEQ to 300 mEQ/day) [Saito et al., 19851, a decrease in young men with a family history of hypertension, but no significant change in controls without a family history (1 2 g/day for 4 weeks) [Herlitz et al., 1983; Gudrnundsson et al., 19841. Similarly, dietary sodium reduction lowered levels in high-school students (about 70% reduction) [Trevisan et al., 19811 and normotensives (20 mmol/day for 4 days) [Stokes et al., 19861, and had no significant effect in high school students (1 10 to 40 mEQ/day) [Cooper et al., 19841 or in moderately obese middle-aged male hypertensives (120 mmol/day for 4 to 6 weeks) [Fagerberg et al., 19861 or hypertensives (20 mmol/day for 4 days) [Stokes et al., 19861.
ACKNOWLEDGMENTS
This research was supported by National Institutes of Health grant HL 2485548.

The Inheritance of RBC Na 201
REFERENCES
Ash KO, Smith JB, Kemp JW, Lynch MB, Moody FG, Raymond JL, McKnight MR, Williams RR (1983): The erect of diet on ouabain binding to erythrocytes from obese subjects. Clin Physiol Biochem
Beutler E, Kuhl W, Sacks P (1983): Sodium-potassium-ATPase activity is influenced by ethnic origin and not by
Blaustein MP (1984): Sodium transport and hypertension. Where are we going? Hypertension 6:445453. Boyle CR, Elston RC (1979): Multifactorial genetic models for quantitative traits in humans. Biometrics
3 5: 5 5-68. Bramley PM, Paulin JM, Millar JA (1986a): Influence of age, sex and weight on measurements of intracellular
Na+ and K+ in essential hypertension. Clin Exp Pharmacol Physiol 13:347-351. Bramley PM, Paulin JM, Millar JA (1986b): Intracellular cations and transmembrane cation transport in
essential hypertension: The importance of controlled clinical observations. J Hypertension 439-596. Canessa M, Adragna N, Solomon HS, Connolly TM, Tosteson DC (1980): Increased sodium-lithium
countertransport in red cells of patients with essential hypertension. New Engl J Med 302772-776. Cannings C, Thompson EA, Skolnick MH (1980): Pedigree analysis of complex models. In Mielke JH,
Crawford MH (eds): “Current Developments in Anthropological Genetics,” Vol2. New York Plenum,
Clegg G, Morgan DB, Davidson C (1982): The heterogeneity of essential hypertension. Relation between lithium efflux and sodium content of erythrocytes and a family history of hypertension. Lancet 2891-894.
Cooper R, Trevisan M, Van Horn L, Larbi E, Liu K, Nanas S, Ueshima H, Sempos C, Ostrow D, Stamler J (1984): Effect of dietary sodium reduction on red blood cell sodium concentration and sodium-lithium countertransport. Hypertension 6:73 1-735.
Cumberbatch M, Morgan DB (1981): Relations between sodium transport and sodium concentration in human erythrocytes in health and disease. Clin Sci 60555-564.
Dadone MM, Smith JB, Wu L, Ash KO, Williams RR (1987): Evidence for genetic determination of intraerythrocytic sodium concentration in Utah pedigrees. Am J Med Genet 27:3944.
D’Amico G (1958): Red cell Na and K in congestive heart failure, essential hypertension and myocardial infarction. Am J Med Sci 236:156-161.
DeLuise M, Blankburn GL, Flier JS (1980): Reduced activity of the red-cell sodium-potassium pump in human obesity. New Engl J Med 303:1017-1022.
Elston RC, Stewart J (1971): A general model for the genetic analysis of pedigree data. Hum Hered
Fadeke Aderounmu A, Salako LA (1979): Abnormal cation composition and transport in erythrocytes from hypertensive patients. Eur J Clin Invest 9369-375.
Fagerberg B, Isaksson B, Herlitz H, Andersson OK (1986): Body composition, intraerythroqte sodium content, volume regulation and blood pressure during moderate sodium restriction in hypertensive men. Acta Med S a n d 219:371-379.
Gudmundsson 0, Herlitz H, Jonsson 0, Hedner T, Andersson 0, Berglund G (1984): Blood pressure and intraerythrocyte sodium during normal and high salt intake in middle-aged men: Relationship to family history of hypertension, and neurogenic and hormonal variables. Clin Sci 66:427433.
Hasstedt SJ (1982): A mixed model likelihood approximation on large pedigrees. Comp Biomed Res
Hasstedt SJ, Cartwright PE (1981): PAP: Pedigree Analysis Package, Rev 2. Technical Report No. 13,
Hasstedt SJ, Moll PP (1987): Estimation of genetic model parameters for variables correlated with a
Hasstedt SJ, Williams RR (1986): Three alleles for quantitative Lp(a). Genet Epidemiol3:53-55. Hasstedt SJ, Wu LL, Ash KO, Kuida H, Williams RR (1987): Hypertension and sodium-lithium countertrans-
port in Utah pedigrees: Evidence for major locus inheritance. Submitted to Am J Hum Genet. Henningsen NC, Nelson D (1980): Red cell metabolism of sodium in relatives to patients with an established
essential hypertension. In Zumkley H, Losse H (eds): “Intracellular Electrolytes and Arterial Hyperten- sion. I. International Symposium Miinster.” Stuttgart: Georg Thieme Verlag, pp 205-212.
Hentschel WM, Wu LL, Tobin GO, Anstall HB, Smith JB, Williams RR, Ash KO (1986): Erythrocyte cation transport activities as a function of cell age. Clin Chem Acta 157:3344.
Herlitz H, Gudmundsson 0, Jonsson 0, Naucler J, Berglund G (1983): Changes in intraerythrocyte content and transmembrane fluxes of sodium during salt loading in subjects with and without family history of hypertension. Acta Med Scand (suppl) 677:69-72.
11293-299.
obesity. New Engl J Med 309:756-760.
pp 251-298.
21 523-542.
151295-307.
Department of Medical Biophysics and Computing, University of Utah, Salt Lake City.
quantitative phenotype exhibiting major locus inheritance, in preparation.

202 Hasstedt et al.
Hilton PJ (1986): Cellular sodium transport in essential hypertension. New Engl J Med 314222-229. Hunt SC, Dadone MM, Williams RR, Wu LL, Smith JB, Kuida H, Ash KO (1987): Familial correlations from
genes and shared environment for urine, plasma, and intraerythrocytic sodium. Am J Med Genet 2 1 24%255.
Lalouel J-M (1979): GEMINI-A computer program for optimization of general nonlinear functions. Technical Report No. 14, Department of Medical Biophysics and Computing, University of Utah, Salt Lake City.
Lalouel JM, Rao DC, Morton NE, Elston RC (1983): A unified model for complex segregation analysis. Am J Hum Genet 35:816-826.
Lasker N, Hopp L, Grossman S, Bamforth R, Aviv A (1985): Race and sex differences in erythrocyte NA+, K+, and Na+-K+-adenosine triphosphatase. J CIin Invest 75:1813-1820.
Lijnen P, M’Buyamba-Kabangu J-R, Fagard RH, Groeseneken DR, Staessen JA, Amery AK (1984): Intracellular concentration and transmembrane fluxes of sodium and potassium in erythrocytes of white normal male subjects with and without a family history of hypertension. J Hypertens 2:25--30.
Losse H, Zidek W, Zumkley H, Wessels F, Vetter H (1981): Intracellular Na+ as a genetic marker of essential hypertension. Clin Exp Hypertens 3:627440.
Montanari A, Borghi L, Canali M, Curti A, Perinotto P, Novarini A, Borghetti A (1980): Altered sodium efflux in red blood cells from essential hypertensive subjects. In Zumkley H, Losse H (eds): “Intracellular Electrolytes and Arterial Hypertens. I. International Symposium Miinster.” Stuttgart: Georg Thieme Verlag, pp 135-144.
Morton NE, MacLean CJ (1974): Analysis of family resemblance 111. Complex segregation of quantitative traits. Am J Hum Genet 26:489-503.
Myers JB, Morgan TO, Edwards K (1983): Effect of sodium intake on intraerythrocyte sodium concentration in normotensive subjects and patients with untreated mild hypertension. Clin Exp Pharmacol Physiol 1036 1-364.
Saito K, Furuta Y, Sano H, Okishio T, Fukuzaki H (1985): Abnormal relationship between dietary sodium intake and red cell sodium transport in salt-sensitive patients with essential hypertension. Clin Exp Hypertens 7:1217-1232.
Schmalzing G, Pfaff E, Breyer-Pfaff U (1981): Red cell ouabain binding sites, Na+K+-ATPase, and intracellular Na+ as individual characteristics. Life Sci 29:371-381.
Simon G , Conklin DJ (1986): In vivo erythrocyte sodium concentration in human hypertension is reduced, not increased. J Hypertens 4:71-75.
Smith JB, Price AL, Williams RR, Hentschel WM, Sprowell W, Hunt SC, Ash KO (1982): A reproducible sodium-lithium countertransport assay: The outcome of changing key laboratory parameters, Clin Chim Acta 122327-335.
Smith JB, Ash KO, Hentschel WM, Sprowell WL, Williams RR (1984): A simplified method for simultaneously determining countertransport and cotransport in human erythrocytes. Clin Chim Acta 137:169-177.
Stokes GS, Monaghan JC, Middleton AT, Shirlow M, Marwood J F (1986): Effects of dietary sodium deprivation on erythrocyte sodium concentration and cation transport in normotensive and untreated hypertensive subjects. J Hypertens 4:35-38.
Svensson A, Sigstrom L (1986): Blood pressure, erythrocyte sodium and potassium concentrations and Na+K+ ATPase activity in children with hypertensive mothers. J Hypertens 4:269-272.
Trevisan M, Cooper R, Ostrow D, Miller W, Sparks S, Leonas Y, Allen A, Steinhauer M, Stamler J (1981): Dietary sodium, erythrocyte sodium concentration, sodium-stimulated lithium efflux and blood pressure. Clin Sci 61:29~-32s.
Ward RH (1983): Genetic and sociocultural components of high blood pressure. Am J Phys Anthropol 62:9 1-1 05.
Webster DP, Van Winkle LJ, Karratt JJ (1984): Erythrocyte ouabain binding and intracellular Na+ in normotensive obese women and obese women receiving medication for hypertension. Biochem Med 32:232-241.
Weller JM (1959): The acid-base balance and sodium distribution of the blood in essential hypertension. J Lab Clin Med 53553-556.
Wessels F (1 980): Genetic aspects of sodium metabolism in RBC. In Zumkley H, Losse H (eds): “Intracellular Electrolytes and Arterial Hypertension. I. International Symposium Miinster.” Stuttgart: Georg Thieme Verlag, pp 221-227.
Wessels F, Zumkley H (1980): Sodium metabolism of RBC in hypertensive patients. In Zumkley H, Losse H

The Inheritance of RBC Na 203
(eds): “Intracellular Electrolytes and Arterial Hypertension. I. International Symposium Miinster.” Stuttgart: Georg Thieme Verlag, pp 59-68.
Williams RR, Skolnick M, Carmelli D, Maness AT, Hunt SC, Hasstedt S, Reiber GE, Jones RK (1979): Utah pedigree studies: Design and preliminary data for premature male CHD deaths. In Sing CF, Skolnick M (eds): “Genetic Analysis of Common Diseases: Applications to Predictive Factors in Coronary Disease.” New York Alan R Liss, Inc, pp 7 I 1-729.
Williams R, Dadone MM, Hunt SC, Jorde LB, Hopkins PN, Smith JB, Ash KO, Kuida H (1984): The genetic epidemiology of hypertension: A review of past studies and current results for 948 persons in 48 Utah pedigrees. In Rao DC, Elston RC, Kuller LH, Feinleib M, Carter C, Havlik R (eds): “Genetic Epidemiology of Coronary Heart Disease. Past, Present, and Future.” New York, Alan R Liss, Inc., pp 4 19442.
Edited by James F. Reynolds