the escherichia coli dam dna methyltransferase modifies dna in a highly processive reaction
TRANSCRIPT

The Escherichia coli Dam DNA MethyltransferaseModifies DNA in a Highly Processive Reaction
Sabine Urig1, Humaira Gowher1, Andrea Hermann1, Carsten Beck1
Mehrnaz Fatemi1, Andeas Humeny2 and Albert Jeltsch1*
1Institut fur BiochemieFachbereich BiologieJustus-Liebig-UniversitatHeinrich-Buff-Ring 58, 35392Giessen, Germany
2Institut fur BiochemieUniversitatErlangen-NurnbergFahrstrasse 17, 91054 ErlangenGermany
The Escherichia coli dam adenine-N6 methyltransferase modifies DNA atGATC sequences. It is involved in post-replicative mismatch repair, con-trol of DNA replication and gene regulation. We show that E. coli damacts as a functional monomer and methylates only one strand of theDNA in each binding event. The preferred way of ternary complexassembly is that the enzyme first binds to DNA and then to S-adenosyl-methionine. The enzyme methylates an oligonucleotide containing twodam sites and a 879 bp PCR product with four sites in a fully processivereaction. On l-DNA comprising 48,502 bp and 116 dam sites, E. coli damscans 3000 dam sites per binding event in a random walk, that on averageleads to a processive methylation of 55 sites. Processive methylation ofDNA considerably accelerates DNA methylation. The highly processivemechanism of E. coli dam could explain why small amounts of E. colidam are able to maintain the methylation state of dam sites during DNAreplication. Furthermore, our data support the general rule that solitaryDNA methyltransferase modify DNA processively whereas methyltrans-ferases belonging to a restriction-modification system show a distributivemechanism, because processive methylation of DNA would interferewith the biological function of restriction-modification systems.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: DNA methylation; DNA replication; enzyme mechanism;processivity; protein–DNA interaction*Corresponding author
Introduction
DNA from most prokaryotes and eukaryotescontains methylated bases, i.e. 4-methylcytosine,5-methylcytosine and 6-methyladenine. This modi-fication is introduced after DNA replication byDNA methyltransferases (MTases) which employS-adenosylmethionine (AdoMet) as donor for anactivated methyl group. DNA methylation playsan important role in eukaryotes where it is usedas a general tool to repress gene expressioninvolved in genomic imprinting, X-chromosomeinactivation, protection of the genome against self-ish genetic elements and epigenetic gene regu-lation (reviews1 – 7). In addition, alterations of theDNA methylation pattern are frequently observedin human cancers (reviews8,9). In prokaryotes,
DNA methylation contributes to DNA repair,phage protection, gene regulation and control ofDNA replication (reviews1 – 7). Most bacterial DNAMTases modify the DNA at palindromic recog-nition sequences in both strands of the DNA.Thereby, they impress a strain specific methylationpattern to the DNA which allows the distinctionof self and non-self DNA and is the basis of restric-tion-modification systems (RM-systems). RM-systems occur ubiquitously in bacteria and consti-tute an efficient protection system against bacterio-phage infection (reviews10,11). They consist of arestriction endonuclease (reviews12,13) that cleavesDNA at defined recognition sites and a corre-sponding methyltransferase that recognizes thesame DNA sequence and modifies the DNA withinthe site. Since modified DNA is protected againstendonucleolytic attack, the cellular DNA is notcleaved by the restriction enzyme. Invading phageDNA, however, is not modified and, therefore,liable to cleavage.
In addition, many bacteria express DNA MTasesthat are not accompanied by restriction enzymes.
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
E-mail address of the corresponding author:[email protected]
Abbreviations used: AdoMet, S-adenosylmethionine;MTase, DNA methyltransferase; RM-system, restriction-modification system.
doi:10.1016/S0022-2836(02)00371-6 available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 319, 1085–1096
Examples include the dam MTase (GATC) ing-proteobacteria,14,15 and the CcrM MTase(GANTC) in a-proteobacteria.16 These systems areinvolved in post-replicative mismatch repair, thesynchronization of DNA replication and cell cycleand the control of gene expression (reviews17 – 20).In the dam system, the DNA is modified in bothstrands at palindromic GATC recognition sites.During DNA replication hemimethylated damsites are formed which persist for two to fourseconds.21 During this period mismatched base-pairs that arose by errors of the DNA polymerasecan be recognized by the mutHLS system(reviews22,23). Hemimethylation of the DNA allowsto distinguish the parental strand and the newlysynthesized daughter stand which is a prerequisitefor the directed repair of replication errors. In con-trast to bulk DNA, the origin of replication stayshemimethylated for approximately 20 minutes,24,25
because the SeqA protein binds to it and,thereby, prevents dam methylation. As hemi-methylated origins of replication are inactive,this mechanism inhibits premature re-initiationof DNA replication. Whereas the dam MTase isnot required for growth in Escherichia coli,26 theCcrM MTase is an essential protein ina-proteobacteria.27 dam methylation also hasimportant functions in bacterial pathogenicityin E. coli,28 Salmonella thyphimurium,29 – 32 andNeisseria meningitidis33 such that specific inhibitors
of dam methylation could be useful for anti-bacterial therapy.34
The E. coli dam MTase is a monomeric enzymecomprising 278 amino acid residues. dam MTasesfrom different g-proteobacteria form a largemultigene family which contains atleast oneMTase that belongs to an RM-system, theM.Eco RV MTase.35 –38 One important mechanisticparameter of DNA MTases is the processivity ofthe methylation reaction, i.e. the question if afterone turnover the enzyme can slide along the DNAin a process of one dimensional (or linear) diffu-sion and carry out subsequent turnovers on thesame substrate. This mechanism is feasible,because DNA MTases in general are able to slideon the DNA by linear diffusion to locate theirtarget sites.39 – 41 Here, we have investigated theprocessivity of DNA methylation by the damenzyme, because this issue is not clear in the litera-ture: Herman & Modrich42 could not detect proces-sive methylation of DNA using a substrate withtwo sites separated by 2455 bp. In contrast,Bergerat et al. found indirect evidence for proces-sive methylation of up to three dam sites on a plas-mid DNA substrate that are separated by 11 and9 bp, respectively.43 To clarify this importantmechanistic question for the dam MTase, we havecloned, overexpressed and purified the enzymeand investigated processivity of DNA methylationwith different substrates.
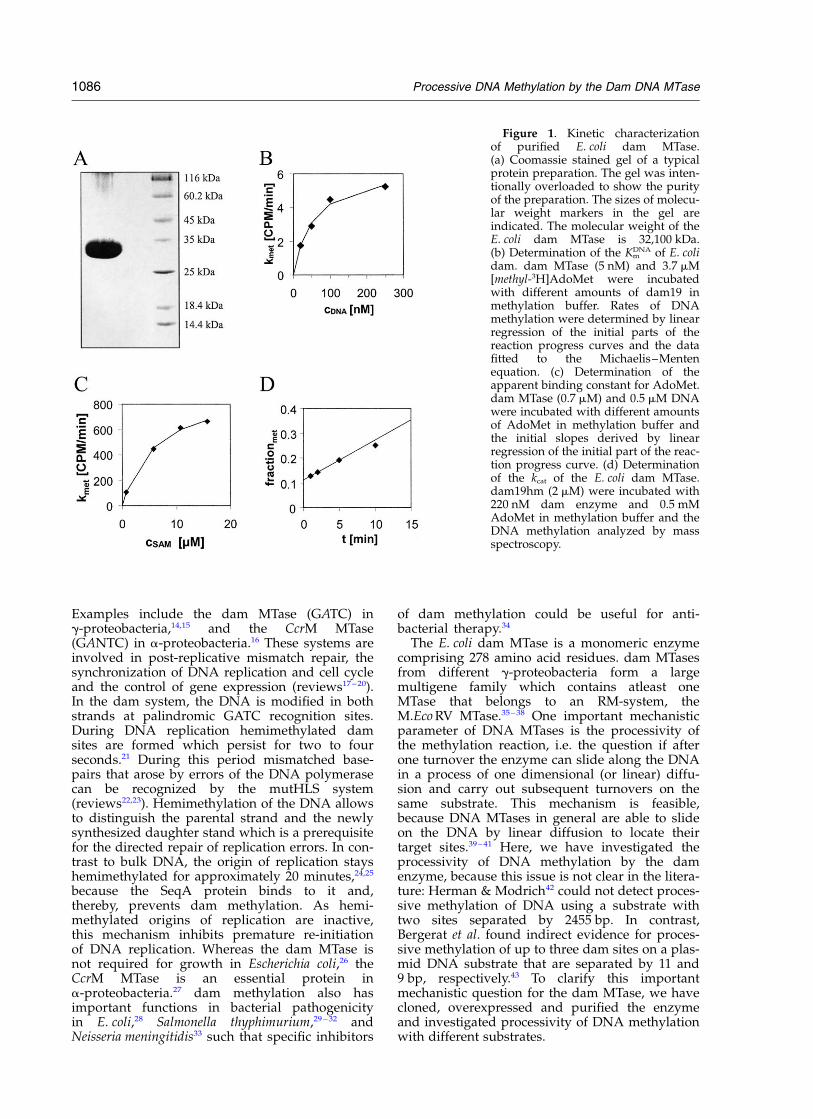
Figure 1. Kinetic characterizationof purified E. coli dam MTase.(a) Coomassie stained gel of a typicalprotein preparation. The gel was inten-tionally overloaded to show the purityof the preparation. The sizes of molecu-lar weight markers in the gel areindicated. The molecular weight of theE. coli dam MTase is 32,100 kDa.(b) Determination of the Km
DNA of E. colidam. dam MTase (5 nM) and 3.7 mM[methyl-3H]AdoMet were incubatedwith different amounts of dam19 inmethylation buffer. Rates of DNAmethylation were determined by linearregression of the initial parts of thereaction progress curves and the datafitted to the Michaelis–Mentenequation. (c) Determination of theapparent binding constant for AdoMet.dam MTase (0.7 mM) and 0.5 mM DNAwere incubated with different amountsof AdoMet in methylation buffer andthe initial slopes derived by linearregression of the initial part of the reac-tion progress curve. (d) Determinationof the kcat of the E. coli dam MTase.dam19hm (2 mM) were incubated with220 nM dam enzyme and 0.5 mMAdoMet in methylation buffer and theDNA methylation analyzed by massspectroscopy.
1086 Processive DNA Methylation by the Dam DNA MTase
Results
Cloning, protein purification and initialcharacterization of the E. coli dam MTase
The E. coli dam gene was cloned into pET28a,overexpressed in E. coli and purified over Ni-NTAagarose and phosphocellulose. Proteinpreparations were pure .98% as estimated fromoverloaded Coomassie blue stained SDS-gels(Figure 1(a)). Different preparations containedbetween 70 and 200 mM dam MTase. Thespecific activity of our preparations is8.9(^0.5) £ 105 units/mg† which is identical withvalues in the literature (8 £ 105 units/mg,44
1 £ 106 units/mg45). The enzyme preparations donot carry significant amounts of endogenousAdoMet, because 1 mg l-DNA (containing3.6 pmol dam sites) incubated with 100 pmol damMTase in the absence of external AdoMet is notcleaved by DpnI (which cuts DNA methylated atGATC sites) and fully digested by DpnII (which isinhibited by dam methylation) (data not shown).Using labeled [methyl-3H]AdoMet a Km
DNA of55(^15) nM was determined with dam19, a 19meroligonucleotide that contains one dam site(Figure 1(b)) which is identical with a valueobtained with a 14mer substrate.46 The rate ofDNA methylation was determined at different con-centrations of AdoMet. Under the assumption thatAdoMet binding is a fast equilibrium, the initialrate of DNA methylation reflects the saturation ofthe dam enzyme with the cofactor. Then, theresults can be analyzed with respect to theapparent dissociation constant for AdoMet whichwas determined to be 6(^1) mM (Figure 1(c)) close
to KmAdoMet values determined by others with oligo-
nucleotide substrates (3 mM,47 3.5 mM48). The kcat ofthe enzyme was determined using 2 mMdam19hm, 220 nM dam MTase and 1 mM AdoMetby a mass spectrometric analysis of the methyl-ation of the DNA. As shown in Figure 1(d) weobserved a clear burst of methylation becauseduring the first minute of the reaction 11(^1)% ofthe substrate becomes methylated which corre-sponds to 220(^10) nM product formation. Thisresult indicates that the enzyme preparation is100% active within experimental error. After thisphase a linear reaction was observed which takesplace with a rate of 31(^4) nM product formationper minute yielding a kcat of 0.14(^0.02) min21.
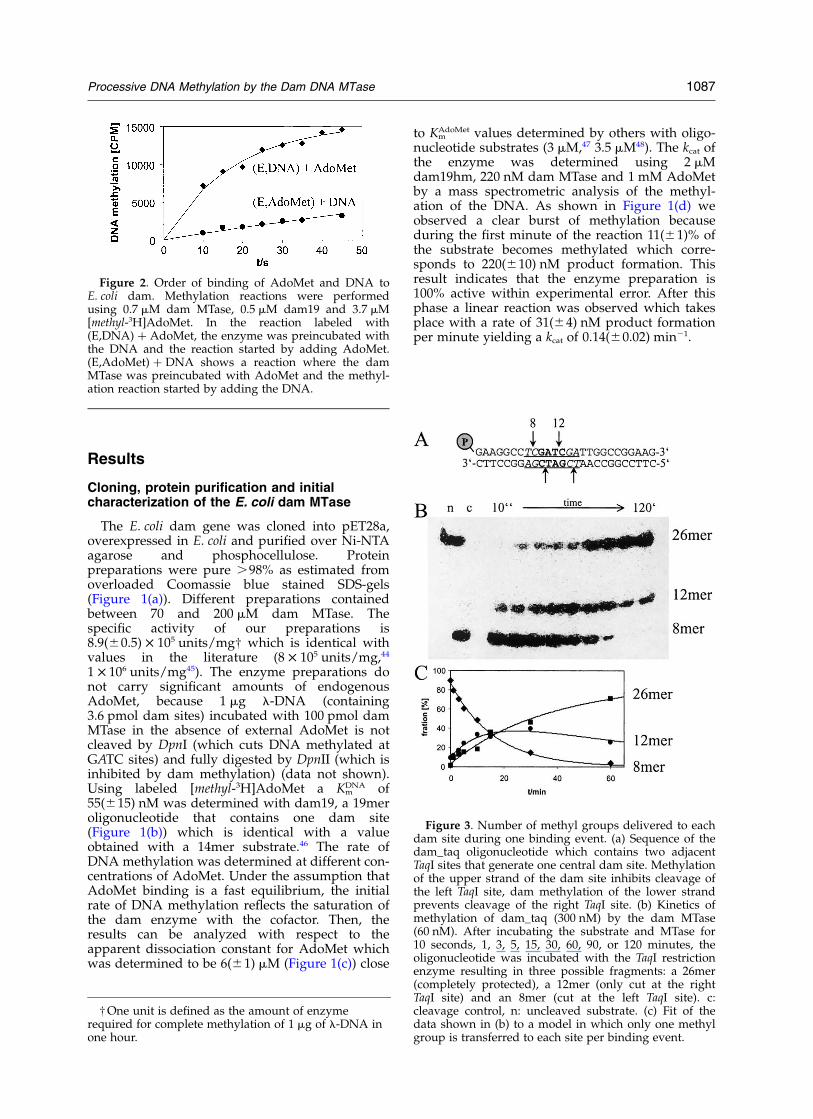
Figure 2. Order of binding of AdoMet and DNA toE. coli dam. Methylation reactions were performedusing 0.7 mM dam MTase, 0.5 mM dam19 and 3.7 mM[methyl-3H]AdoMet. In the reaction labeled with(E,DNA) þ AdoMet, the enzyme was preincubated withthe DNA and the reaction started by adding AdoMet.(E,AdoMet) þ DNA shows a reaction where the damMTase was preincubated with AdoMet and the methyl-ation reaction started by adding the DNA.
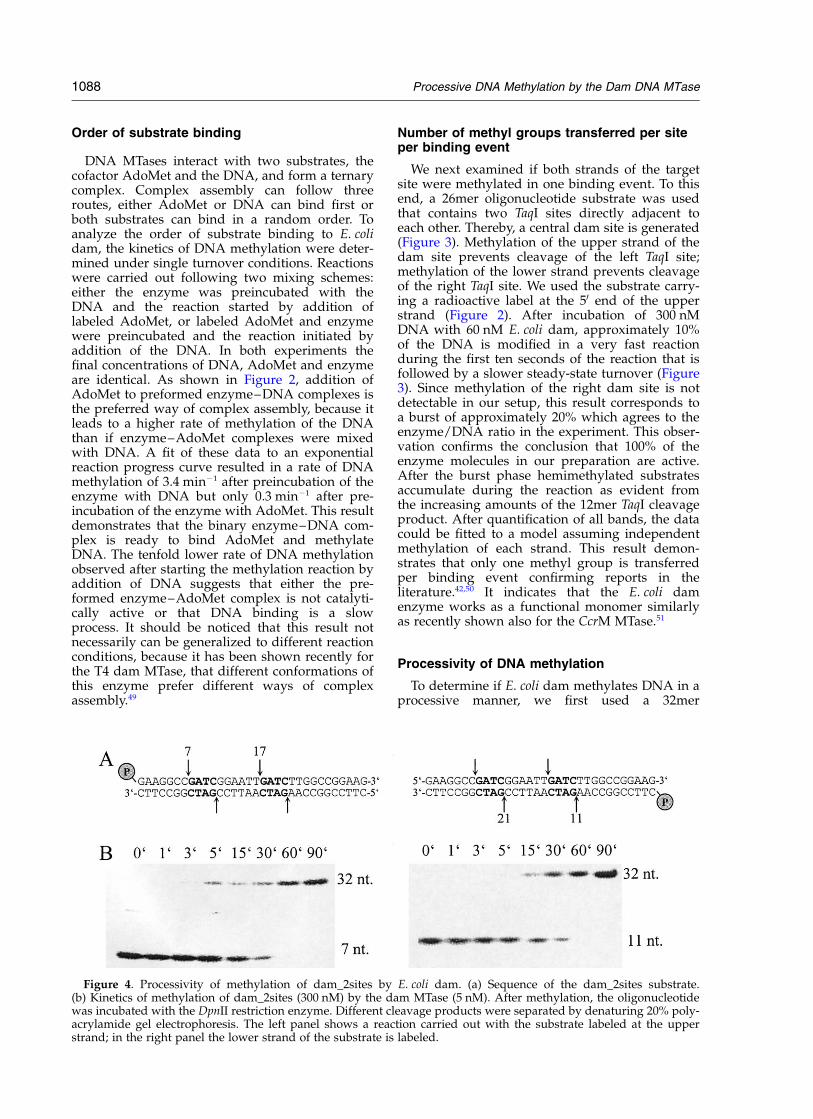
Figure 3. Number of methyl groups delivered to eachdam site during one binding event. (a) Sequence of thedam_taq oligonucleotide which contains two adjacentTaqI sites that generate one central dam site. Methylationof the upper strand of the dam site inhibits cleavage ofthe left TaqI site, dam methylation of the lower strandprevents cleavage of the right TaqI site. (b) Kinetics ofmethylation of dam_taq (300 nM) by the dam MTase(60 nM). After incubating the substrate and MTase for10 seconds, 1, 3, 5, 15, 30, 60, 90, or 120 minutes, theoligonucleotide was incubated with the TaqI restrictionenzyme resulting in three possible fragments: a 26mer(completely protected), a 12mer (only cut at the rightTaqI site) and an 8mer (cut at the left TaqI site). c:cleavage control, n: uncleaved substrate. (c) Fit of thedata shown in (b) to a model in which only one methylgroup is transferred to each site per binding event.
† One unit is defined as the amount of enzymerequired for complete methylation of 1 mg of l-DNA inone hour.
Processive DNA Methylation by the Dam DNA MTase 1087
Order of substrate binding
DNA MTases interact with two substrates, thecofactor AdoMet and the DNA, and form a ternarycomplex. Complex assembly can follow threeroutes, either AdoMet or DNA can bind first orboth substrates can bind in a random order. Toanalyze the order of substrate binding to E. colidam, the kinetics of DNA methylation were deter-mined under single turnover conditions. Reactionswere carried out following two mixing schemes:either the enzyme was preincubated with theDNA and the reaction started by addition oflabeled AdoMet, or labeled AdoMet and enzymewere preincubated and the reaction initiated byaddition of the DNA. In both experiments thefinal concentrations of DNA, AdoMet and enzymeare identical. As shown in Figure 2, addition ofAdoMet to preformed enzyme–DNA complexes isthe preferred way of complex assembly, because itleads to a higher rate of methylation of the DNAthan if enzyme–AdoMet complexes were mixedwith DNA. A fit of these data to an exponentialreaction progress curve resulted in a rate of DNAmethylation of 3.4 min21 after preincubation of theenzyme with DNA but only 0.3 min21 after pre-incubation of the enzyme with AdoMet. This resultdemonstrates that the binary enzyme–DNA com-plex is ready to bind AdoMet and methylateDNA. The tenfold lower rate of DNA methylationobserved after starting the methylation reaction byaddition of DNA suggests that either the pre-formed enzyme–AdoMet complex is not catalyti-cally active or that DNA binding is a slowprocess. It should be noticed that this result notnecessarily can be generalized to different reactionconditions, because it has been shown recently forthe T4 dam MTase, that different conformations ofthis enzyme prefer different ways of complexassembly.49
Number of methyl groups transferred per siteper binding event
We next examined if both strands of the targetsite were methylated in one binding event. To thisend, a 26mer oligonucleotide substrate was usedthat contains two TaqI sites directly adjacent toeach other. Thereby, a central dam site is generated(Figure 3). Methylation of the upper strand of thedam site prevents cleavage of the left TaqI site;methylation of the lower strand prevents cleavageof the right TaqI site. We used the substrate carry-ing a radioactive label at the 50 end of the upperstrand (Figure 2). After incubation of 300 nMDNA with 60 nM E. coli dam, approximately 10%of the DNA is modified in a very fast reactionduring the first ten seconds of the reaction that isfollowed by a slower steady-state turnover (Figure3). Since methylation of the right dam site is notdetectable in our setup, this result corresponds toa burst of approximately 20% which agrees to theenzyme/DNA ratio in the experiment. This obser-vation confirms the conclusion that 100% of theenzyme molecules in our preparation are active.After the burst phase hemimethylated substratesaccumulate during the reaction as evident fromthe increasing amounts of the 12mer TaqI cleavageproduct. After quantification of all bands, the datacould be fitted to a model assuming independentmethylation of each strand. This result demon-strates that only one methyl group is transferredper binding event confirming reports in theliterature.42,50 It indicates that the E. coli damenzyme works as a functional monomer similarlyas recently shown also for the CcrM MTase.51
Processivity of DNA methylation
To determine if E. coli dam methylates DNA in aprocessive manner, we first used a 32mer
Figure 4. Processivity of methylation of dam_2sites by E. coli dam. (a) Sequence of the dam_2sites substrate.(b) Kinetics of methylation of dam_2sites (300 nM) by the dam MTase (5 nM). After methylation, the oligonucleotidewas incubated with the DpnII restriction enzyme. Different cleavage products were separated by denaturing 20% poly-acrylamide gel electrophoresis. The left panel shows a reaction carried out with the substrate labeled at the upperstrand; in the right panel the lower strand of the substrate is labeled.
1088 Processive DNA Methylation by the Dam DNA MTase

oligonucleotide substrate that contains two damsites separated by 6 bp (Figure 4). The substratewas labeled on one strand, methylated by E. colidam for a defined period of time and cleaved withDpnII to determine which dam sites have beenmethylated at least in one strand of the DNA. Thelengths of all possible DpnII cleavage fragmentsare indicated in Figure 4(a). Most importantly, sub-strates that are methylated at only one dam sitewill produce characteristic DpnII cleavage pro-ducts: a substrate that is methylated on the leftdam site but unmethylated on the right one willproduce a 17mer cleavage product if it is labeledon the upper strand. Similarly, a substrate that isonly modified on the right dam site will producea 21mer cleavage product if it is labeled on thelower strand. As shown in Figure 4(b) such incom-pletely methylated DNA molecules are notdetected in our experiments regardless if theupper or lower strand of the DNA carries theradioactive label. This result can be explained onlyif both sites on each DNA molecule are modifiedin a processive reaction. Moreover, it means thatthe enzyme moves on the DNA in a randomfashion in both directions and that it has a highprobability of being reflected at the ends of linearDNA. This is a typical behavior for an enzymemoving on the DNA by linear diffusion withoutconsumption of energy (see41,52 – 54 and additionalreferences therein).
Methylation of oligonucleotide substratescontaining more than one GATC site
The observation of bursts of methylation inkinetics carried out under multiple turnover con-ditions (Figure 1(d) and Figure 3) suggests that thechemical step of DNA methylation is not rate limit-ing for E. coli dam. Most likely, for the dam MTaselike for many other DNA interacting enzymes pro-duct release is the rate limiting step. Then, onewould expect, that substrates containing morethan one GATC site which can be modified in aprocessive reaction should be methylated fasterthan substrates with only one site. To test this pre-sumption experimentally, we have determined therates of methylation of three different 30mer oligo-nucleotides that contain one (dam30), two(dam30_2sites) and three (dam30_3sites) damsites. In each case a time-course of methylationwas determined using 0.5 mM DNA, 0.1 mMenzyme and 3.7 mM [methyl-3H]AdoMet. The ratesof DNA methylation as determined by linearregression from the initial part of the reactionprogress curves were: dam30: 38(^5) CPM/minute, dam30_2sites: 71(^10) CPM/minute anddam30_3sites: 93(^5) CPM/minute. As expected,the substrate with three sites was modified thefastest, the substrates with two sites intermediateand the substrate with only one site was modifiedwith the slowest rate. Thus processive methylationof DNA leads to an increased overall rate ofmethylation.
Processive methylation of macromolecularDNA substrates
To test for processive methylation of longer DNAsubstrates, we first used an 879mer PCR productthat contains four dam sites. The substrate wasprepared after 32P-labeling of one of the PCRprimers on its 50 end such that one end of theDNA carries a radioactive label (Figure 5). Methyl-ation with E. coli dam and cleavage with DpnIIagain did not show any incompletely modifiedDNA molecules (Figure 5). Therefore, theunmethylated 879mer is directly converted intothe product that is modified at all four dam sites.This result demonstrates that the E. coli damenzyme methylates four sites on DNA substratescomprising 879 bp in a completely processive man-ner. As a control the methylation reaction was alsoanalyzed by DpnI digestion. Then, methylatedDNA will be degraded and unmethylated willremain uncleaved. However, no methylation inter-mediates were observed after the DpnI digest of879mer incubated with dam MTase in methylationbuffer for different periods of time (data notshown). This finding confirms our conclusion thatE. coli dam modifies the 879mer in a fully proces-sive reaction.
These experiments were carried out in a methyl-ation buffer containing 100 mM Hepes (pH 8.0),1 mM EDTA, 0.5 mM DTT that has been used inall other enzymological studies with the E. coli
Figure 5. Processivity of methylation of an 879merPCR product by E. coli dam. (a) Methylation of the PCRproduct that carries a radioactive label at the 50 end ofthe upper DNA strand. The position of the first nucleo-tide of each of the four dam sites on the 879mer is indi-cated. After incubation of the DNA (90 nM) and thedam MTase (5 nM) for 0, 1, 2, 3, 5, 10, 15, 30 or 60minutes, the DNA was incubated with the DpnII restric-tion enzyme. The cleavage products were separated ona native 6% polyacrylamide gel. (b) Same as in (a) exceptthat the PCR product is 50 labeled in the lower strand.
Processive DNA Methylation by the Dam DNA MTase 1089
dam enzyme.42 –45 The salt concentration of thisbuffer clearly is lower than physiological levels.We therefore carried out additional methylationexperiments with the 879mer in buffers containing50 and 100 mM NaCl, as well as 50 mM NaCl and5 mM MgCl2. The methylation reactions were fullyprocessive under all the conditions (data notshown) demonstrating that the E. coli dam enzymemodifies DNA in a highly processive reactionunder physiological conditions.
In our experiments we used an AdoMet concen-tration of 1 mM that is slightly higher than AdoMetconcentrations observed in vivo in bacterial cells(300–500 mM,55 30–300 mM,56 100 mM57). It wasused because AdoMet is known to be unstableand our in vitro experiments do not provide anAdoMet regenerating system. Therefore AdoHcywill accumulate which is known to be a strongcompetitive inhibitor for AdoMet binding to DNAMTases. In order to find out if the high concen-tration of AdoMet is critical for processivity, wehave carried out methylation experiments with the879mer at an AdoMet concentration of 100 mMand also observed full processivity (data notshown).
Processivity of methylation of l-DNA
To find out if more than four sites can be modi-fied in a processive reaction, we analyzed the
DpnII cleavage pattern of l-DNA during the time-course of dam methylation (Figure 6(a)). Clearly,l-DNA is not modified in a fully processive reac-tion at least in vitro because methylation inter-mediates are easily visible after DpnII digest.However, fully protected DNA and final cleavagefragments are observed simultaneously during themethylation reaction. Simulations of the methyl-ation reaction demonstrate that this result cannotbe obtained in the case of a non-processive reaction(see Figure 7). For a more quantitative analysis ofprocessivity, the fractions of full length l-DNAand final cleavage fragments with respect to whatis observed in uncleaved or fully cleaved DNAwere determined at each time point of the reactionand plotted against time (Figure 6(b)). In this plota characteristic crossover point is obtained wheresame fractions of final cleavage fragments and fulllength DNA are present. The position of the cross-over point in this plot is characteristic for the pro-cessivity of the reaction, the higher theprocessivity the higher the crossover point. In afully processive reaction this point is at 0.5 becauseno incompletely modified DNA molecules areformed. In a distributive reaction the crossoverpoint is close to zero, because for statistical reasonsonly very few DNA molecules carry two unmodi-fied sites next to each other at the same time whenother DNA molecules are already modified at all116 dam sites (Figure 7). We experimentally found
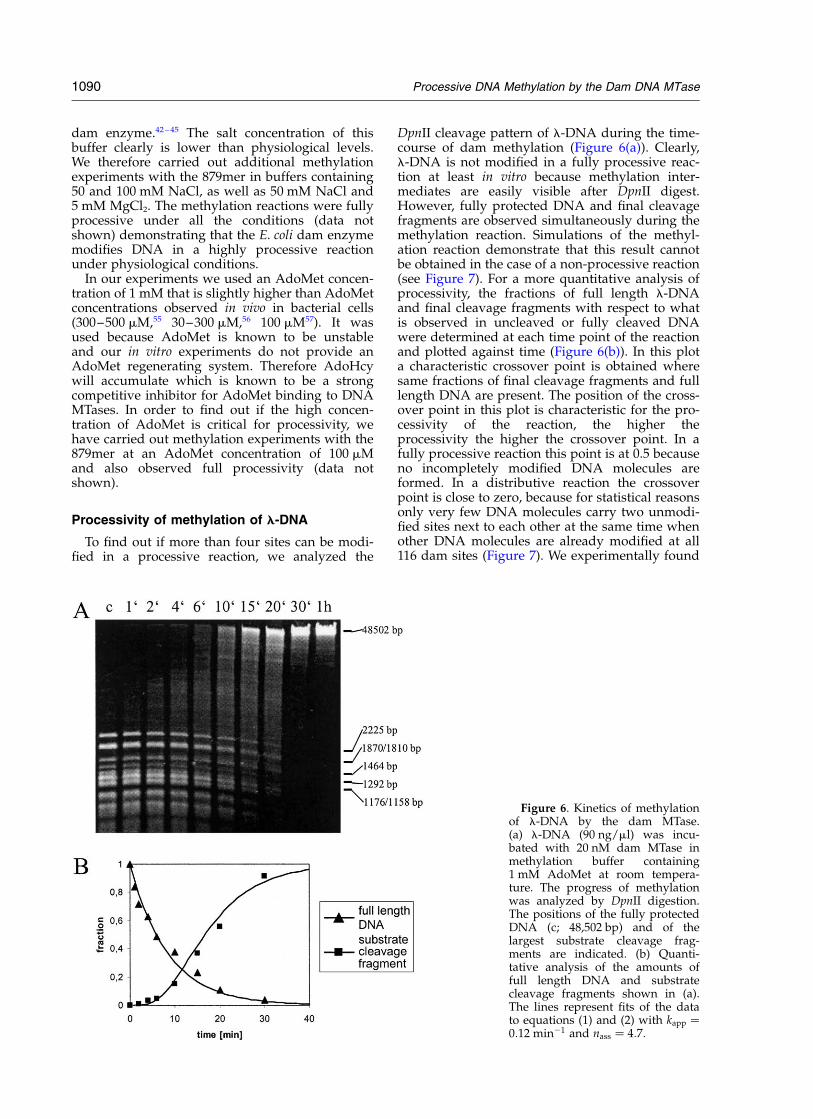
Figure 6. Kinetics of methylationof l-DNA by the dam MTase.(a) l-DNA (90 ng/ml) was incu-bated with 20 nM dam MTase inmethylation buffer containing1 mM AdoMet at room tempera-ture. The progress of methylationwas analyzed by DpnII digestion.The positions of the fully protectedDNA (c; 48,502 bp) and of thelargest substrate cleavage frag-ments are indicated. (b) Quanti-tative analysis of the amounts offull length DNA and substratecleavage fragments shown in (a).The lines represent fits of the datato equations (1) and (2) with kapp ¼0:12 min21 and nass ¼ 4:7:
1090 Processive DNA Methylation by the Dam DNA MTase
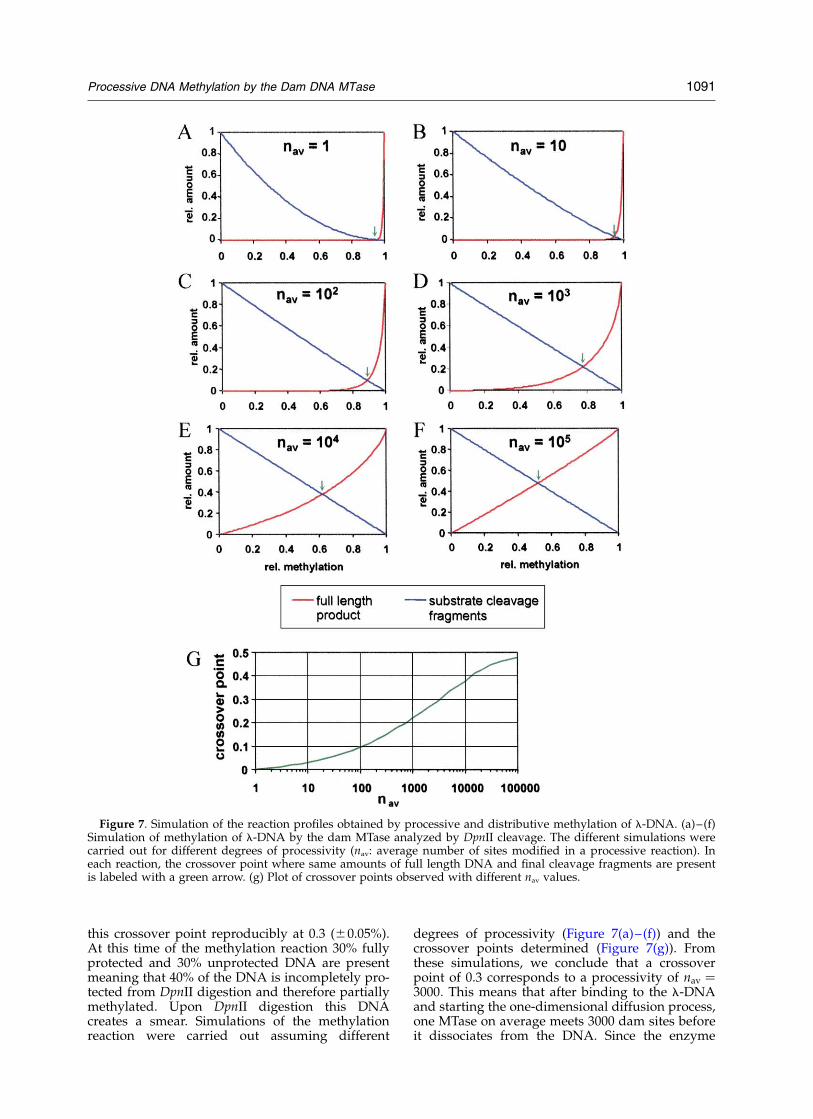
this crossover point reproducibly at 0.3 (^0.05%).At this time of the methylation reaction 30% fullyprotected and 30% unprotected DNA are presentmeaning that 40% of the DNA is incompletely pro-tected from DpnII digestion and therefore partiallymethylated. Upon DpnII digestion this DNAcreates a smear. Simulations of the methylationreaction were carried out assuming different
degrees of processivity (Figure 7(a)–(f)) and thecrossover points determined (Figure 7(g)). Fromthese simulations, we conclude that a crossoverpoint of 0.3 corresponds to a processivity of nav ¼3000: This means that after binding to the l-DNAand starting the one-dimensional diffusion process,one MTase on average meets 3000 dam sites beforeit dissociates from the DNA. Since the enzyme
Figure 7. Simulation of the reaction profiles obtained by processive and distributive methylation of l-DNA. (a)–(f)Simulation of methylation of l-DNA by the dam MTase analyzed by DpnII cleavage. The different simulations werecarried out for different degrees of processivity (nav: average number of sites modified in a processive reaction). Ineach reaction, the crossover point where same amounts of full length DNA and final cleavage fragments are presentis labeled with a green arrow. (g) Plot of crossover points observed with different nav values.
Processive DNA Methylation by the Dam DNA MTase 1091

performs a random walk, it will hit many sitesmore often than only once and the effective rangeof processivity corresponds to the square root ofnav. Therefore, the dam MTase is able to methylateapproximately 55 GATC sites on l-DNA in aprocessive reaction in vitro. In this simulation, 4.6associations of an MTase molecule to the samemolecule of l-DNA are required to obtain fullymethylated DNA. Similar results were obtained ifthe progress of DNA methylation was analyzedby DpnI cleavage (data not shown).
As shown in Figure 6(b) the disappearance of thefinal cleavage fragments is described by a hyper-bolic curve whereas the increase in full lengthDNA shows a sigmoidal curve shape. This differ-ence is caused by the fact that every binding eventof the dam MTase to the DNA leads to a reductionof the amount of substrate cleavage fragments. Incontrast, since the dam enzyme does not methylatethe l-DNA in a fully processive way, on averagemore than one enzyme molecule must bind eachsubstrate molecule before the first fully protectedDNA is generated. To analyze these data morequantitatively, all microscopic events occurringduring one binding event (association of the damenzyme to the DNA, linear diffusion, methylationof all sites on the way and dissociation of theenzyme from the DNA) were combined into oneapparent rate constant (kapp). Then, the increase inthe concentration of the full length DNA (cP for“product”) is given by equation (1):
cP ¼ ð1 2 e2kapptÞnass ð1Þ
where nass represents the number of enzymes thatmust bind to the l-DNA before fully protectedDNA is generated. The reduction in the concen-tration of the substrate cleavage fragments (cS) isgiven by equation (2):
cS ¼ e2kappt ð2Þ
A global fit of the data shown in Figure 6(b) tothese equations resulted in an apparent rate con-stant of 0.12 min21 and an nass number of 4.7.Given the accuracy of this semi-quantitativeanalysis and the assumptions inherent in thenumerical simulation, there is an excellent agree-ment between the values obtained for nass from theexperimental data (4.7) and in the numerical simu-lations of the methylation reaction (4.6, see above).It is also impressive to note the similarity of therate of 0.12 min21 and the kcat value of the enzymeon an oligonucleotide substrate that contains onlyone dam site (0.14 min21, see above). This resultsuggests that multiple turnover kinetics are almostcompletely dominated by the rate of productrelease, because methylation of several sites takesas long as methylation of just one single site.
Discussion
In this study we have investigated the kineticmechanism of the E. coli dam MTase. We have
shown that binding of the AdoMet after DNAbinding is the preferred way of ternary enzyme–DNA–AdoMet complex assembly under ourexperimental conditions. This allows for a proces-sive methylation of DNA. We have shown herethat E. coli dam methylates four dam sites onDNA molecules comprising 879 bp in a fully pro-cessive reaction. On l-DNA, the enzyme performsa one-dimensional random walk and encountersapproximately 3000 dam sites before it dissociatesform the DNA. Since it moves in a random fashion,this on average leads to a processive methylationof 55 dam site. Processive methylation of DNAmethylation has two important consequences: firstit considerably accelerates the rate of DNA methyl-ation. Second, it leads to an all or none behavior ofthe methylation pattern where fully methylatedstretches of the DNA alternate with completelyunmethylated.
The E. coli dam enzyme is closely related to theM.Eco RV adenine-N6 MTase that modifies DNA atGATATC sites.35 – 38 Both enzymes share 27% aminoacid identity and another 40% similarity and mostlikely are the outcome of divergent evolution. Thesimilarity between dam MTases and M.EcoRV isemphasized by the finding that M.EcoRV alsomodifies DNA at dam sites.38,58 However, we haveshown recently, that the kinetic mechanism ofM.EcoRV is dramatically different from that ofE. coli dam, because M.EcoRV binds the AdoMetfirst.41 M.EcoRV–DNA and also M.EcoRV–DNA–AdoHcy complexes are catalytically not competentand, therefore, the enzyme does not methylateDNA processively,41 which is in sharp contrast tothe results obtained here with E. coli dam. In fact,the E. coli dam/M.Eco RV pair is just one exampleof a general rule, because all MTases that areaccompanied by a restriction enzyme show a dis-tributive mechanism of DNA methylation(M.HhaI, M.HpaII, M.EcoRI and M.EcoRV)40,41,59
and all solitary MTases methylate DNA in a pro-cessive reaction (CcrM, M.SssI and E. coli dam)(this paper).59,60 The question appears if theremight be a biological reason why these relatedenzymes behave such differently.
As first discussed by Surby & Reich,40 the distri-butive mechanism of DNA methylation is animportant adaptation of DNA MTases in RM-systems to the biological function of these systemswhich is to cleave phage DNA invading the cellby the restriction enzyme. Since methylation pro-tects the DNA from cleavage, it is important thatthe endonuclease reaches a restriction site on thephage DNA, before it is modified by the corre-sponding MTase. As the phage DNA usually con-tains more than one recognition site for the RM-system and the endonuclease is able to scan theDNA by linear diffusion, methylation of somerecognition sites does not prevent cleavage of thephage DNA as long as not all the recognition sitesare modified before an endonuclease has bound tothe substrate. RM-systems evolved to avoid thissituation by making the MTases distributive which
1092 Processive DNA Methylation by the Dam DNA MTase

considerably slows down the rate of DNAmethylation.
The functional constraints for a solitary bacterialmaintenance MTase (not involved in an RM-system) are different. The E. coli genome comprises4,639,221 bp and contains 19,123 GATC sites thatare subject to methylation by the E. coli damenzyme.61 Therefore, after DNA replication 38,246dam sites (19,123 on each chromosome) must bemethylated in each cell cycle. Given the amount ofdam MTase (20–130 molecules per cell62,63), eachenzyme has to modify 382 sites in 20 minutes ifone assumes an amount of 100 dam MTasemolecules per cell. This corresponds to a kcat ofabout 20 turnovers/minute, which is approxi-mately 100-fold faster than observed with an oligo-nucleotide substrate with one methylation site.Processive methylation of the DNA can help toresolve this discrepancy. However, even under theconditions of processive DNA methylation it isstill difficult to rationalize, how the dam MTasecan modify all dam sites in one generation time,because in our l-DNA methylation experiments,2.8 nM l-DNA (325 nM dam sites) and 20 nMdam MTase are present (1 enzyme per 16 sites),but complete methylation takes 30 minutes invitro†, which altogether is 30–40 times slower thanthe required rate of 382 GATC sites in 20 minutes.However, there is one fundamental differencebetween the situation in vitro and in vivo: it isknown that the DNA polymerase “factory” isstationary located in the mid-cell region and expelsboth daughter DNA strands towards the two cell-poles.64,65 Therefore, it is possible that the damenzymes are also located at fixed positions in thecell and that the DNA is pushed through them.Then, the MTase would scan the DNA in a directedfashion which would mean that one dam MTasemolecule could modify 3000 dam sites processivelycorresponding to 750,000 bp or about 1/6 of theE. coli genome. Since scanning and methylation of3000 dam sites on l-DNA occurs with a rate of0.12 min21, the rate of DNA methylation by E. colidam could be as high as 6 s21. This rate would besufficient to follow a replication fork which moveswith approximately 1000 bp/second66 and onaverage produces four dam sites per second. Insummary, while the MTase must search for its rec-ognition sites in a random walk in vitro, the energydriven directional movement of the DNA in vivocould lead to a directed movement of the MTaseon the DNA. This mechanical coupling of DNApolymerase and MTase would accelerate the rateof DNA methylation by orders of magnitudes byconverting a random walk into a directed move-ment. In this model, in principle, one dam enzymeon the leading and lagging strand would be suffi-cient for complete re-methylation of the whole
E. coli genome after DNA replication. Future workwill show if this hypothetical model of a stationarydam enzyme and a mechanical coupling of thedam MTase to DNA replication is right.
Materials and Methods
Oligodeoxynucleotides
The following double-stranded oligodeoxynucleotidesubstrates obtained by annealing of complementarysingle-stranded oligonucleotides were used in thisstudy: dam_2sites (GAAGGCCGATCGGAATTGAT-CTTGGCCGGAAG), dam_taq (GAAGGCCTCGATCGA-TTGGCCGGAAG), dam19 (GCCCGGGGATCCGGC-CGCG), dam19hm (dam19 methylated in the lowerstrand), dam30 (GAAGGCCCTAGGTGATCTTCTAG-TGGCCGA), dam30_2sites (GAAGGCCGATCGTGATC-TTCTAGTGGCCGA), dam30_3sites (GAAGGCCGA-TCGTGATCTTGATCTGGCCGA). dam19, dam19hm,dam30, dam30_2sites and dam30_3sites carry a biotin atthe 50 end of the upper strand.
Cloning, expression and purification of the E. colidam MTase
The gene of the E. coli dam MTase (Entrez entry code:AAC76412) was amplified by PCR from E. coli JM109and cloned into pET28a þ (Novagen). The His6-damMTase fusion protein was expressed in BL21(DE3,pLysS). The cell pellet was resuspended in buffer A(20 mM KPi (pH 7.5), 0.1 mM DTT, 0.5 M NaCl, 10%(v/v) glycerol, 20 mM imidazole) and the cells weredisrupted by ultrasound. Cell debris was removed bycentrifugation. The supernatant was applied onto a Ni-NTA (Qiagen) column equilibrated with buffer A. Afterwashing with 150 ml buffer A, the dam MTase waseluted with 5 ml of buffer A containing 200 mM imida-zole. The eluate was diluted 1:5 with buffer B (20 mMKPi (pH 7.5), 1 mM EDTA, 0.1 mM DTT, 10% glycerol)and applied to a phosphocellulose column (Whatman).The column was washed with 100 ml buffer B containing100 mM NaCl, eluted with buffer B containing 1000 mMNaCl and collected in fractions. The fractions containingthe dam MTase were pooled, dialyzed overnight againststorage buffer (20 mM Hepes (pH 7.5), 40 mM NaCl,1 mM EDTA, 0.2 mM DTT, 50% glycerol) and stored at220 8C. The concentration of the dam MTase was deter-mined using an extinction coefficient of e280 nm ¼39; 935 M21 cm21 calculated from the contents of Trpand Tyr.67
Methylation kinetics with labeled [methyl-3H]-AdoMet
The methylation assays using labeled [methyl-3H]-AdoMet and biotinylated oligonucleotide substrateswere carried out as described.68 In most assays, DNAand labeled S-[methyl-3H]AdoMet (555 GBq/mmol;Hartmann Analytics, Braunschweig, Germany) wereused at concentrations of 0.5 mM and 3.7 mM, respect-ively, at ambient temperature in methylation buffer(100 mM Hepes (pH 8.0), 1 mM EDTA, 0.5 mM DTT).Usually the amount of radioactivity incorporated into2 pmol substrate DNA was analyzed. All in vitro methyl-ation experiments were carried out at least in duplicate.
† Similar results were obtained by others, since thespecific activity of our preparations is identical with thatof other groups (see above).
Processive DNA Methylation by the Dam DNA MTase 1093

Determination of kcat
The kcat of the E. coli dam MTase was determinedusing the dam19hm oligonucleotide. Oligonucleotide(2 mM), 20 nM dam MTase and 1 mM unlabeled AdoMet(Sigma) were incubated in methylation buffer at ambienttemperature. After defined times 10 ml aliquots werewithdrawn, stopped by heating to 95 8C and the amountof methyl groups incorporated into the DNA analyzedby MALDI-TOF mass spectrometry (A. H. A. Beck,C.-M. Becker, & A. J., unpublished results).
In vitro methylation of l-DNA
For methylation analyses, 0.45 mg l-DNA were incu-bated with the dam MTase in 5 ml of methylation buffercontaining 1 mM AdoMet (Sigma) at 37 8C. After definedtimes, the reaction was stopped by heating to 65 8C. DpnI(NEB) or DpnII (NEB) cleavage was carried out usingten units (as defined by the supplier) of the enzymes ina total volume of 15 ml for one hour at 37 8C andanalyzed on 0.8% agarose gels. DpnII cleaves DNA atunmethylated GATC sites whereas DpnI specificallycleaves DNA at methylated GATC sites.
Methylation analyis of an 879mer PCR product byDpnII restriction protection
An 879mer PCR product containing four GATC siteswas amplified from the m.ecorv gene. An end-labeledPCR fragment was generated using one labeled primerin the PCR reaction. Methylation reactions were carriedout at ambient temperature in methylation buffer con-taining 1 mM AdoMet. The reactions were started byaddition of the enzyme. At defined times, aliquots wereremoved and methylation reactions were stopped byaddition of nine volumes of ethanol. After precipitation,the DNA was dissolved in DpnII cleavage buffer (NEB).The samples were incubated with ten units of DpnII forone hour at 37 8C. Cleavage products were separated ona 6% (w/v) polyacrylamide gel and analyzed using anInstant Imager (Canberra Packard).
Restriction protection analysis of methylation ofoligonucleotide substrates
Methylation dependent restriction protection experi-ments with different oligonucleotide substrates (dam_2-sites, dam_taq) were carried out essentially as describedfor the PCR product. The oligonucleotides were end-labeled with T4-polynucleotide kinase (NEB) using[g-32P]ATP (Hartmann, Analytic). The TaqI or DpnIIdigested samples were analyzed on 20% polyacrylamidegels containing 7 M urea.
Computer simulation of methylation of l-DNA by thedam MTase
The DpnI and DpnII cleavage patterns of l-DNAduring methylation by the dam MTase were simulatedwith a program written in Delphie 3.0 (Borland). In thisnumerical simulation, an MTase binds randomly to adam site on 10,000 molecules of l-DNA. If an unmodi-fied site is bound, the MTase methylates it. The MTasethen searches by linear diffusion for the next dam site.The reach of linear diffusion is given by the probabilityof the enzyme to dissociate from the DNA (Poff) whichis the variable parameter in the simulation. Then, the
average number of dam sites scanned (nav) is given bynav ¼ 1=Poff: It is assumed that the MTase modifies allsites on its way and that it does not dissociate at theend of the linear DNA. During the methylation reaction,DpnI and DpnII digests were simulated. Completelymodified l-DNA is resistant against DpnII digest (“fulllength DNA”). DpnII cleavage of DNA molecules con-taining two consecutive unmethylated dam sites resultsin fragments that correspond to one of the fragmentsthat one would obtain after DpnII digest of completelyunmethylated DNA (“substrate cleavage fragments”). Itshould be noticed that substrate cleavage fragments canbe obtained from completely unmethylated but alsofrom partially methylated l-DNA molecules. DpnIIdigest of partially methylated DNA molecules producesadditional cleavage bands that do not correspond tocleavage fragments of unmodified DNA. Since l-DNAhas 116 dam sites, there are 6612 possible intermediatefragments which precludes any quantitative treatmentof experimental data with respect to methylation inter-mediates. Therefore, in this simulation we also did notconsider the intermediate bands but focus on theamounts of full length l-DNA and of the substratecleavage fragments. The processivity of the methylationreaction is described by the amounts of fully methylatedDNA and unmethylated DNA that are present at thesame time in the reaction mixture. Upon DpnII digest acharacteristic crossover point of the decreasing amountof substrate cleavage fragments and the increasingamount of full length DNA is observed; the higher thecrossover point the higher the processivity. Obviously,in a fully processive reaction a crossover point of 0.5 isobtained, because no intermediates are generated. DpnIdigests were analyzed in an analogous manner.
Acknowledgments
Thanks are due to B. Kleiber for technical assistance.Support and discussions by A. Pingoud and C.-M.Becker are gratefully acknowledged. This work hasbeen supported by the Deutsche Forschungs-gemeinschaft (JE 252/2-3).
References
1. Bestor, T. H. & Verdine, G. L. (1994). DNA methyl-transferases. Curr. Opin. Cell Biol. 6, 380–389.
2. Cheng, X. (1995). DNA modification by methyl-transferases. Curr. Opin. Struct. Biol. 5, 4–10.
3. Cheng, X. (1995). Structure and function of DNAmethyltransferases. Annu. Rev. Biophys. Biomol.Struct. 24, 293–318.
4. Robertson, K. D. & Wolffe, A. P. (2000). DNA methyl-ation in health and disease. Nature Rev. Genet. 1,11–19.
5. Jones, P. A. & Takai, D. (2001). The role of DNAmethylation in mammalian epigenetics. Science, 293,1068–1070.
6. Cheng, X. & Roberts, R. J. (2001). AdoMet-dependentmethylation, DNA methyltransferases and baseflipping. Nucl. Acids Res. 29, 3784–3795.
7. Jeltsch, A. (2002). Beyond Watson and Crick: DNAmethylation and molecular enzymology of DNAmethyltransferases. ChemBioChem. 3, 274–293.
1094 Processive DNA Methylation by the Dam DNA MTase

8. Warnecke, P. M. & Bestor, T. H. (2000). Cytosinemethylation and human cancer. Curr. Opin. Oncol.12, 68–73.
9. Baylin, S. B. & Herman, J. G. (2000). DNA hyper-methylation in tumorigenesis: epigenetics joinsgenetics. Trends Genet. 16, 168–174.
10. Wilson, G. G. & Murray, N. E. (1991). Restriction andmodification systems. Annu. Rev. Genet. 25, 585–627.
11. Heitman, J. (1993). On the origins, structures andfunctions of restriction-modification enzymes. InGenetic Engineering (Setlow, J. K., ed.), vol. 15, pp.57–108, Plenum Press, New York.
12. Pingoud, A. & Jeltsch, A. (1997). Recognition andcleavage of DNA by type-II restriction endo-nucleases. Eur. J. Biochem. 246, 1–22.
13. Pingoud, A. & Jeltsch, A. (2001). Structure and func-tion of type II restriction endonucleases. Nucl. AcidsRes. 29, 3705–3727.
14. Lacks, S. & Greenberg, B. (1977). Complementaryspecificity of restriction endonucleases of Diplococcuspneumoniae with respect to DNA methylation. J. Mol.Biol. 114, 153–168.
15. Hattman, S., Brooks, J. E. & Masurekar, M. (1978).Sequence specificity of the P1 modificationmethylase (M.EcoP1) and the DNA methylase(M.Ecodam) controlled by the Escherichia coli damgene. J. Mol. Biol. 126, 367–380.
16. Zweiger, G., Marczynski, G. & Shapiro, L. (1994). ACaulobacter DNA methyltransferase that functionsonly in the predivisional cell. J. Mol. Biol. 235,472–474.
17. Marinus, M. G. (1987). DNA methylation inEscherichia coli. Annu. Rev. Genet. 21, 113–131.
18. Messer, W. & Noyer-Weidner, M. (1988). Timing andtargeting: the biological functions of Dam methyl-ation in E. coli. Cell, 54, 735–737.
19. Barras, F. & Marinus, M. G. (1989). The great GATC:DNA methylation in E. coli. Trends Genet. 5, 139–143.
20. Reisenauer, A., Kahng, L. S., McCollum, S. &Shapiro, L. (1999). Bacterial DNA methylation: a cellcycle regulator? J. Bacteriol. 181, 5135–5139.
21. Stancheva, I., Koller, T. & Sogo, J. M. (1999). Asym-metry of Dam remethylation on the leading andlagging arms of plasmid replicative intermediates.EMBO J. 18, 6542–6551.
22. Modrich, P. (1994). Mismatch repair, genetic stability,and cancer. Science, 266, 1959–1960.
23. Modrich, P. & Lahue, R. (1996). Mismatch repair inreplication fidelity, genetic recombination, andcancer biology. Annu. Rev. Biochem. 65, 101–133.
24. Campbell, J. L. & Kleckner, N. (1990). E. coli oriC andthe dnaA gene promoter are sequestered from dammethyltransferase following the passage of thechromosomal replication fork. Cell, 62, 967–979.
25. Lu, M., Campbell, J. L., Boye, E. & Kleckner, N.(1994). SeqA: a negative modulator of replicationinitiation in E. coli. Cell, 77, 413–426.
26. Palmer, B. R. & Marinus, M. G. (1994). The dam anddcm strains of Escherichia coli—a review. Gene, 143,1–12.
27. Stephens, C., Reisenauer, A., Wright, R. & Shapiro, L.(1996). A cell cycle-regulated bacterial DNA methyl-transferase is essential for viability. Proc. Natl Acad.Sci. USA, 93, 1210–1214.
28. Hale, W. B., van der Woude, M. W., Braaten, B. A. &Low, D. A. (1998). Regulation of uropathogenicEscherichia coli adhesin expression by DNA methyl-ation. Mol. Genet. Metab. 65, 191–196.
29. Heithoff, D. M., Sinsheimer, R. L., Low, D. A. &Mahan, M. J. (1999). An essential role for DNAadenine methylation in bacterial virulence. Science,967–970.
30. Enserink, M. (1999). Gene may promise new route topotent vaccines. Science, 284, 883.
31. Garcia-Del Portillo, F., Pucciarelli, M. G. &Casadesus, J. (1999). DNA adenine methylasemutants of Salmonella typhimurium show defects inprotein secretion, cell invasion, and M cell cyto-toxicity. Proc. Natl Acad. Sci. USA, 96, 11578–11583.
32. Heithoff, D. M., Sinsheimer, R. L., Low, D. A. &Mahan, M. J. (2000). In vivo gene expression and theadaptive response: from pathogenesis to vaccinesand antimicrobials. Phil. Trans. Roy. Soc. ser. B, Biol.Sci. 355, 633–642.
33. Bucci, C., Lavitola, A., Salvatore, P., Del Giudice, L.,Massardo, D. R., Bruni, C. B. & Alifano, P. (1999).Hypermutation in pathogenic bacteria: frequentphase variation in meningococci is a phenotypictrait of a specialized mutator biotype. Mol. Cell, 3,435–445.
34. Low, D. A., Weyand, N. J. & Mahan, M. J. (2001).Roles of DNA adenine methylation in regulating bac-terial gene expression and virulence. Infect. Immun.69, 7197–7204.
35. Lauster, R., Kriebardis, A. & Guschlbauer, W. (1987).The GATATC-modification enzyme EcoRV is closelyrelated to the GATC-recognizing methyltransferasesDpn II and dam from E. coli and phage T4. FEBSLetters, 220, 167–176.
36. Malone, T., Blumenthal, R. M. & Cheng, X. (1995).Structure-guided analysis reveals nine sequencemotifs conserved among DNA amino-methyl-transferases, and suggests a catalytic mechanism forthese enzymes. J. Mol. Biol. 253, 618–632.
37. Tran, P. H., Korszun, Z. R., Cerritelli, S., Springhorn,S. S. & Lacks, S. A. (1998). Crystal structure of theDpnM DNA adenine methyltransferase from theDpnII restriction system of Streptococcus pneumoniaebound to S-adenosylmethionine. Structure, 6,1563–1575.
38. Beck, C., Cranz, S., Solmaz, M., Roth, M. & Jeltsch, A.(2001). How does a DNA interacting enzyme changeits specificity during molecular evolution? A site-directed mutagenesis study at the DNA binding siteof the DNA-(adenine-N6)-methyltransferase EcoRV.Biochemistry, 40, 10956–10965.
39. Nardone, G., George, J. & Chirikjian, J. G. (1986).Differences in the kinetic properties of BamHI endo-nuclease and methylase with linear DNA substrates.J. Biol. Chem. 261, 12128–12133.
40. Surby, M. A. & Reich, N. O. (1996). Contribution offacilitated diffusion and processive catalysis toenzyme efficiency: implications for the EcoRIrestriction-modification system. Biochemistry, 35,2201–2208.
41. Gowher, H. & Jeltsch, A. (2000). Molecular enzym-ology of the Eco RV DNA-(adenine-N6)-methyltrans-ferase: kinetics of DNA binding and bending,kinetic mechanism and linear diffusion of theenzyme on DNA. J. Mol. Biol. 303, 93–110.
42. Herman, G. E. & Modrich, P. (1982). Escherichia colidam methylase. Physical and catalytic properties ofthe homogeneous enzyme. J. Biol. Chem. 257,2605–2612.
43. Bergerat, A., Kriebardis, A. & Guschlbauer, W.(1989). Preferential site-specific hemimethylation of
Processive DNA Methylation by the Dam DNA MTase 1095

GATC sites in pBR322 DNA by dam methyltransfer-ase from Escherichia coli. J. Biol. Chem. 264, 4064–4070.
44. Geier, G. E. & Modrich, P. (1979). Recognitionsequence of the dam methylase of Escherichia coliK12 and mode of cleavage of DpnI endonuclease.J. Biol. Chem. 254, 1408–1413.
45. Bergerat, A., Guschlbauer, W. & Fazakerley, G. V.(1991). Allosteric and catalytic binding of S-adeno-sylmethionine to Escherichia coli DNA adeninemethyltransferase monitored by 3H NMR. Proc. NatlAcad. Sci. USA, 88, 6394–6397.
46. Thielking, V., Du Bois, S., Eritja, R. & Guschlbauer,W. (1997). Dam methyltransferase from Escherichiacoli: kinetic studies using modified DNA oligomers:nonmethylated substrates. Biol. Chem. 378, 407–415.
47. Bergerat, A. & Guschlbauer, W. (1990). The doublerole of methyl donor and allosteric effector ofS-adenosyl-methionine for Dam methylase of E. coli.Nucl. Acids Res. 18, 4369–4375.
48. Marzabal, S., DuBois, S., Thielking, V., Cano, A.,Eritja, R. & Guschlbauer, W. (1995). Dam methylasefrom Escherichia coli: kinetic studies using modifiedDNA oligomers: hemimethylated substrates. Nucl.Acids Res. 23, 3648–3655.
49. Evdokimov, A. A., Zinoviev, V. V., Malygin, E. G.,Schlagman, S. L. & Hattman, S. (2002). BacteriophageT4 dam DNA-[N6-adenine]methyltransferase.Kinetic evidence for a catalytically essential confor-mational change in the ternary complex. J. Biol.Chem. 277, 279–286.
50. Urieli-Shoval, S., Gruenbaum, Y. & Razin, A. (1983).Sequence and substrate specificity of isolated DNAmethylases from Escherichia coli C. J. Bacteriol. 153,274–280.
51. Shier, V. K., Hancey, C. J. & Benkovic, S. J. (2001).Identification of the active oligomeric state of anessential adenine DNA methyltransferase fromCaulobacter crescentus. J. Biol. Chem. 276, 14744–14751.
52. Jeltsch, A., Alves, J., Wolfes, H., Maass, G. &Pingoud, A. (1994). Pausing of the restriction endo-nuclease EcoRI during linear diffusion on DNA.Biochemistry, 33, 10215–10219.
53. Jeltsch, A., Wenz, C., Stahl, F. & Pingoud, A. (1996).Linear diffusion of the restriction endonucleaseEco RV on DNA is essential for the in vivo functionof the enzyme. EMBO J. 15, 5104–5111.
54. Jeltsch, A. & Pingoud, A. (1998). Kinetic character-ization of linear diffusion of the restrictionendonuclease EcoRV on DNA. Biochemistry, 37,2160–2169.
55. Javor, G. T. (1983). Depression of adenosylmethio-nine content of Escherichia coli by thioglycerol.Antimicrob. Agents Chemother. 24, 860–867.
56. Posnick, L. M. & Samson, L. D. (1999). Influence ofS-adenosylmethionine pool size on spontaneousmutation, dam methylation, and cell growth ofEscherichia coli. J. Bacteriol. 181, 6756–6762.
57. Borczuk, A., Stock, A. & Stock, J. (1987). S-Adenosyl-methionine may not be essential for signal trans-duction during bacteril chemotaxis. J. Bacteriol. 169,3295–3300.
58. Taylor, J. D., Goodall, A. J., Vermote, C. L. & Halford,S. E. (1990). Fidelity of DNA recognition by theEcoRV restriction/modification system in vivo.Biochemistry, 29, 10727–10733.
59. Renbaum, P. & Razin, A. (1992). Mode of action ofthe Spiroplasma CpG methylase M.Sss I. FEBS Letters,313, 243–247.
60. Berdis, A. J., Lee, I., Coward, J. K., Stephens, C.,Wright, R., Shapiro, L. & Benkovic, S. J. (1998). Acell cycle-regulated adenine DNA methyltransferasefrom Caulobacter crescentus processively methylatesGANTC sites on hemimethylated DNA. Proc. NatlAcad. Sci. USA, 95, 2874–2879.
61. Blattner, F. R., Plunkett, G., 3rd, Bloch, C. A., Perna,N. T., Burland, V., Riley, M. et al. (1997). The completegenome sequence of Escherichia coli K-12. Science, 277,1453–1474.
62. Szyf, M., Avraham-Haetzni, K., Reifman, A.,Shlomai, J., Kaplan, F., Oppenheim, A. & Razin, A.(1984). DNA methylation pattern is determined bythe intracellular level of the methylase. Proc. NatlAcad. Sci. USA, 81, 3278–3282.
63. Boye, E., Marinus, M. G. & Lobner-Olesen, A. (1992).Quantitation of Dam methyltransferase in Escherichiacoli. J. Bacteriol. 174, 1682–1685.
64. Lemon, K. P. & Grossman, A. D. (1998). Localizationof bacterial DNA polymerase: evidence for a factorymodel of replication. Science, 282, 1516–1519.
65. Cook, P. R. (1999). The organization of replicationand transcription. Science, 284, 1790–1795.
66. Marians, K. J. (1992). Prokaryotic DNA replication.Annu. Rev. Biochem. 61, 673–719.
67. Pace, C. N., Vajdos, F., Fee, L., Grimsley, G. & Gray, T.(1995). How to measure and predict the molarabsorption coefficient of a protein. Protein Sci. 4,2411–2423.
68. Roth, M. & Jeltsch, A. (2000). Biotin–Avidin micro-plate assay for the quantitative analysis of enzymaticmethylation of DNA by DNA methyltransferases.Biol. Chem. 381, 269–272.
Edited by J. Karn
(Received 17 January 2002; received in revised form 15 April 2002; accepted 16 April 2002)
1096 Processive DNA Methylation by the Dam DNA MTase