the behavioural responses of juvenile signal crayfish pacifastacus leniusculus to stimuli from perch...
TRANSCRIPT

Freshwater Biology (1993) 29, 89-97
The behavioural responses of juvenile signal crayfishPacifastacus leniusculus to stimuli from perchand eels
M.A. BLAKE AND P.J.B. HARTDepartment of Zoology, Adrian Building, University of Leicester, Leicester LEI 7RH, U.K.
SUMMARY
1. Experiments were designed to determine the relative importance of chemical andvisual stimuli in eliciting predator avoidance behaviour in juvenile freshwater crayfishPacifastacus leniusculus (Dana).2. Crayfish placed in visual and/or chemical contact v̂ nth one of two predators exhibitedmarked avoidance behaviour, spending less time walking and climbing and more timewithin shelters.3. The combined effects of both visual and chemical stimuli increased crayfish shelteruse and reduced walking and climbing activity to a greater degree than either stimuluswhen presented alone.4. Crayfish exhibited avoidance behaviour in response to chemical stimuli duringperiods of light and darkness. Visual detecfion of predators elicited avoidance behaviourduring the day.5. It is suggested that the behavioural response of P. leniusculus to chemical stimulireduces the likelihood of being detected by visual predators, and that chemical stimulilower the response threshold for avoidance behaviour in crayfish reacfing to visualstimuli. The adaptivity of using chemical cues to detect predators is emphasized.
Introduction
Behaviour against predation can be categorized aseither avoidance or escape behaviour (Sih, 1987).Escape behaviour occurs after prey have encounteredpredators. Avoidance behaviour by prey occursbefore an encounter with a predator, and reducesthe rate of encounters between predators and prey.
Avoidance behaviours that bring about changesin microhabitat selection and temporal shifts inactivity have been demonstrated in crayfish whenin the presence of crepuscular predators (Stein &Magnuson, 1976; Stein, 1977; Hamrin, 1987; Appelberg& Odelstrom, 1988). In freshwater systems, chemicalstimuli associated v̂ fith predators have been shownto cause avoidance behaviour in prey (Petranka,Kats & Sih, 1987; Alexander & Covich, 1991). Visual,chemical and tacfile stimuli elicit aspects of de-fensive behaviour in crayfish (Wine & Krasne, 1982;Hazlett, 1985).
Perch Perca fluviatilis (L.) and eels Anguilla anguilla(L.) are known to prey on crayfish (Svardson, 1972;Dehli, 1981; Hogger, 1988 for review). Each specieshas a different foraging strategy. AnguilHd eels arenocturnal and forage using chemoreception (Edel,1975; Tesch, 1977). The mean stomach fullness ofAnguilla australis (Richardson) has been reported toincrease throughout the night (Ryan, 1984). Perch arediurnal or crepuscular predators and forage visually(Disler & Simimov, 1977). It is possible that crayfishrespond to different predatory stimuli associatedwith the separate foraging techniques of perch andeels. Visual stimuli should be important for thedetection of diurnal predators such as perch. A re-liance on chemoreception or mechanoreceptionmight be expected for the detection of nocturnalpredators such as eels. This would be a likely productof the levels of illumination present when each pred-ator forages. Crayfish of the genus Orconectes respondto the loss of visual stimuli at night with compensatory
89
90 M.A. Blake and P.].B. Hart
increases in the use of mechanorecepfive organssuch as the antennae and chelae (Bruski & Dunham,1987; Smith & Dunham, 1990). No similar increasewas found in the use of the antennules, a major siteof chemoreception. The purpose of the followingexperiments was to determine the importance ofvisual and chemical stimuli, characterisfic of eelsand perch, in initiating defensive behaviour in thesignal crayfish Pacifastacus leniusculus (Dana).
The predators were presented to the prey undercondifions simulafing nocturnal and diumal lightlevels. Predators and prey were physically isolatedso as to mininuze the chances of the crayfish detecfingthe predators by means of mechanoreception. Underthese experimental condifions it was hypothesizedthat if chemical sfimuli were important in deter-mining defensive behaviour in crayfish, such be-haviour should occur under conditions of both lightand darkness. If visual sfimuli were important, tworesults would be expected. Firstly, defensive behav-iour should only be observed in the light. Secondly,defensive behaviour should be more marked inresponse to an increase in the frequency of visualdisturbances if the latter is a measure of potentialdanger (Sih, 1987).
The following experiments describe the changes inbehaviour observed in crayfish in response to perchand eels during four time periods in the diel cycle.
Materials and Methods
Experimental animals
Juvenile crayfish were obtained from a crayfish farm
near Gillingham, Dorset, U.K. and were stored inaquaria and fed part-boiled potato. The crayfishranged between 16.3 and 23.5 mm carapace length.Eels and perch were obtained by electofishing in theRiver Welland, Leicestershire and in a pond nearWalUngford, Oxfordshire. All fish were stored in a4 m'' arena tank. The total lengths of eels rangedbetween 40 and 60 cm. The perch ranged between15 and 20 cm total length.
Experimental design
Crayfish were placed individually into 12-1 aquariaand were subjected to one of four treatments arrangedin a 2 X 2 factorial design, based on the presence orabsence of visual and chemical contact betweenpredator and prey (Table 1).
Two of the 12-1 aquaria, filled to a depth of 18 cm,were placed adjacent to each other inside each offour 250-1 tanks. These tanks were filled to a depthof 15 cm. For Experiment 1, four eels were rotatedbetween the test tanks. In Experiment 2, the eelswere replaced by four perch. A single tank, containingtwo aquaria, was observed on each day of an ex-periment. Observations were made on each tank inrotafion. In the tanks the aquaria were subjectedto pairs of treatments, as set out in Table 1. Eightreplicates of each of the four treatments were per-formed. Inifially, each crayfish was used twice, oncein each of two different treatments. In some casesreplicates had to be repeated. In Experiment 1, twenty-two crayfish were used in thirty-two replicates ofthe four treatments. In Experiment 2, twenty-onecrayfish were used.
Table 1 Description of the experimental design for Experiments 1 and 2 (X, no stimulus; ^ , stimulus)
Presentation of stimuli
Eel (Experiment 1) Perch (Experiment 2)
Tank Aquarium Treatment Description Visual Chenucal Visual Chemical
A and C a
b
B and D a
b
Control.No stimuliVisual andchemicalstimuliChemicalstimuli onlyVisualstimuli only

Predator avoidance by juvenile crayfish 91
In treatments where the predator and the preywere to be in visual contact, the aquaria were trans-parent. The aquaria sides were covered with blackplastic sheeting in treatments with no visual contactbetween predator and prey. Water was circulatedbetween the aquaria and the tanks in the treatmentswhere predator and prey were to be in chemicalcontact. Where no chemical contact was requiredwater was circulated within each aquarium. AUtank and aquarium bottoms were covered withsand. Each aquarium was fitted with two pieces ofplastic tubing (2 cm diameter x 4 cm long) which thecrayfish could enter for shelter. To provide food forthe crayfish each aquarium was also supplied withpart-boiled potato in excess.
The water temperature in the tanks during theexperiments was 7-9°C. The tank room operated onan 8.5:15.5 L:D regime. The lights came on at 09.30hours and were turned off at 18.00 hours. The lightsdid not fade in or out.
Experimental procedure
The following procedure was used in the preparationand observation of each tank. The water in eachaquarium was replaced and aerated for 24 h. Thecrayfish were then placed in the aquaria 48 h beforeobservations began. The water circulation pumpswere started 24 h before observations began. Onthe day of each experimental trial the crayfish wereobserved for 30 min over four time periods: pre-dawn{08.45h), post-dawn (09.45h), pre-dusk (17.15h) andpost-dusk (18.15h). In the first experiment, the eelswere placed in the tanks 15 min before the start of thepre-dawn and pre-dusk observation periods so asto ensure that they were active throughout all fourperiods of observation. In trials conducted prior tothis experiment eels became inactive when left in thetanks for long periods of time. In Experiment 2, perchwere also placed into the tanks 15 min prior to thepre-dawn and pre-dusk observation periods.
Tank water was not changed between replicates.Water that was occupied by a predator 48 h pre-viously was pumped into aquaria that were to betreated with predator scent 24 h prior to the newpredators being placed in the tanks. As a result, thecrayfish in these aquaria were in contact with oldpredator water for 24 h prior to the onset of theexperiment. The introduction of the predator into
the tanks thus constituted a fresh input of predatorscent.
Crayfish behaviour
Observations of crayfish activity were made usingblack-and-white video recording equipment sensi-tive to infra-red light. Crayfish were filmed for fourperiods of 30 min as indicated above. These timeswere chosen for two reasons. Firstly, to ensure thatthe predators were active whilst filming. Secondly,crayfish change their activity in response to thechanges in light intensity associated with dawn anddusk (Hamrin, 1987). Therefore, any behaviouralchanges associated with predator avoidance shouldhave been discernible within these fime periods.The videos were analysed and crayfish behaviourwas noted every 30 s for each 30-min filming period.Categories of crayfish behaviour were determined inpreliminary trials. Only a small number of behavioursoccurred with regularity during the periods of obser-vation. As a result some categories were combinedfor the purpose of the analyses. The categories arelisted below.
1. Defensive shelter use —crayfish were withdrawninside a shelter and were either not visible or the tipof their rostrum and chelae were visible within theshelter.2. Active shelter use —crayfish were partly exposedoutside a shelter with either their carapace or carapaceand tail visible.3. Total shelter use—includes both defensive andacfive shelter use.4. Walking — forward walking in the open tank.5. Climbing — climbing the tank sides or water cir-culation tubes.
Analysis of behaviour
Counts were made of the number of 30-s observationsin which each behaviour occurred. These countswere expressed as a percentage of the total numberof 30-s observations made during the 30-min offilming. Within each time period the influences ofvisual and chemical stimuli on crayfish behaviourwere analysed using a 2 X 2 factorial, non-parametricanalysis of variance by ranks (Meddis, 1984). TheWilcoxon signed-rank test was used to determinethe effect of changes in light intensity on crayfish

92 M.A. Blake and PJ.B. Hart
behaviour (Siegel & Castellan, 1988). The reults foreach crayfish were paired within treatments andacross the following time periods: (i) pre-dawn vpost-dawn and (ii) pre-dusk v post-dusk. The nullhypothesis employed for these analyses was thatneither treatment nor light condifions affected crayfishbehaviour. Non-specific (two-tailed) alternativehypotheses were employed in all tests.
Results
Crayfish behaviour in response to predatory stimuli
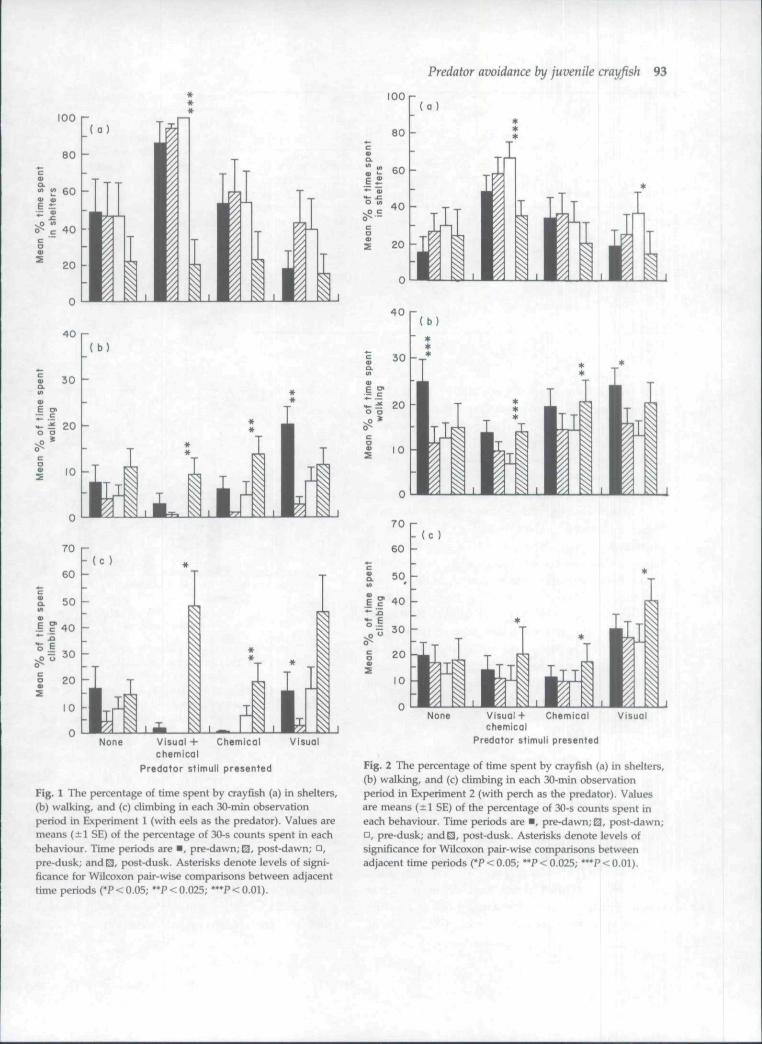
During the pre-dawn, post-dawn and pre-duskperiods, crayfish spent more time in shelters in re-sponse to the scent of both eels and perch (Table 2).Often the increase in shelter use was accompanied.by a reduction in walking and climbing activity,although this was not always the case. Behaviouralresponses of crayfish to fish scent were most markedwhere the fish could also be seen (Figs 1 and 2).Visual stimuli influenced crayfish behaviour, most
strongly during the pre-dusk period. Crayfish spentmore time within shelters and walked less in responseto seeing perch (H = 4.8 and 4.2, respectively, df = 1,m = 16, n = 16, P < 0.05). There was a similar trend inresponse to eels but this was not significant (P > 0.05).
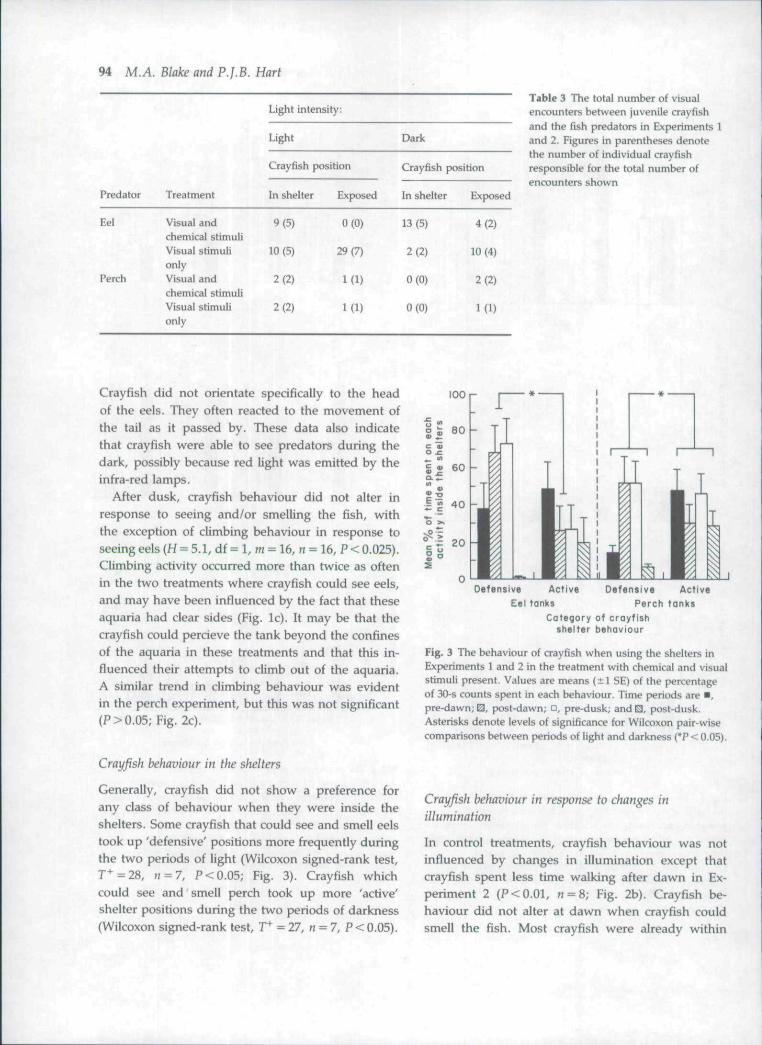
Crayfish behaviour appeared to be influencedmore by seeing eels than by seeing perch. Althoughit was not possible to test this, it may have been aconsequence of the greater activity of the eels. Thisis indicated by the predator-prey encounter data(Table 3). Encounters were only detected in thosetreatments in which fish could be seen by the crayfish.Fifteen of sixteen crayfish encountered eels a totalof seventy-seven times. Eight of sixteen crayfishencountered perch a total of nine times. There wasa clear difference in crayfish behaviour betweentreatments. Where the crayfish could see and smellthe eels, the majority of the encounters occurredwhilst the crayfish were in the shelters. Where the eelscould only be seen, the crayfish were most oftenexposed. This was true in the light (x̂ = 16.9, n = 48,P< 0.001) and the dark (x^=10.1, n = 29, P<0.01).
Time period
Pre-dawn
Post-dawn
Pre-dusk
Post-dusk
Behaviour
In shelter
Walking
Climbing
In shelter
Walking
Climbing
In shelter
Walking
Climbing
In shelter
Walking
Climbing
H-valuePH-valuePH-valuePH-valuePH-valueprH-valuePH-valuePH-valuePH-valuePH-valuenrH-valuepI
H-valueP
Predator
Eel (Experiment 1)
4.39 { + )<0.05
5.49 (-)<0.025
7.34 (-)<0.01
4.46 (+)<0.05
N S • •
3.19 (-)<0.07
4.13 (+)<0.05
3.02 (-)<0.08
2.92 (-)<0.09
NS
NS
NS
Perch (Experiment 2)
6.31 (+)<0.025
5.32 (-)<0.025
5.86 (-)<0.025
5.52 (+)<0.025
NS
4.49 (-)<0.05
3.74 (+)<0.05
NS
3.61 (-)<0.06
NS
NS
NS -
Table 2 Results of part of a 2 x 2 factorialanalysis showing the effect of chemicalstimuli on crayfish behaviour for eachobservation period in Experiment 1 and2. The analyses compared crayfishactivity between treatments with scent(2 + 3) and treatments without scent(1 + 4). Values of H and associatedprobability values (P<0.1) are given inthe body of the table. Figures inparentheses denote the direction of thebehavioural change. In all tests df = 1,ni = 16 and « = 16
Predator avoidance by juvenile crayfish 93
100 r
4 0
30
^ 20t
10
0
70
60
. 50
1*40
I 30
20
10
0
( b )
- (c )
None Visual + Chemicalchemical
Predator stimuli presented
Visual
Fig. 1 The percentage of time spent by crayfish (a) in shelters,(b) walking, and (c) climbing in each 30-min observationperiod in Experiment 1 (with eels as the predator). Values aremeans (±1 SE) of the percentage of 30-s counts spent in eachbehaviour. Time periods are • , pre-dawn;0, post-dawn; • ,pre-dusk; and^, post-dusk. Asterisks denote levels of signi-ficance for Wilcoxon pair-wise comparisons between adjacenttime periods (*P < 0.05; **P < 0.025; *'*P < 0.01).
100
80
60
4 0
20
( a )
1
1
None Visual + Chemicalchemical
Predator stimuli presented
Visual
Fig. 2 The percentage of time spent by crayfish (a) in shelters,(b) walking, and (c) climbing in each 30-min observationperiod in Experiment 2 (with perch as the predator). Valuesare means (±1 SE) of the percentage of 30-s counts spent ineach behaviour. Time periods are • , pre-dawn;0, post-dawn;• , pre-dusk; and^, post-dusk. Asterisks denote levels ofsignificance for Wilcoxon pair-wise comparisons betweenadjacent Hme periods ('P < 0.05; *'P < 0.025; "*P < 0.01).
94 M.A. Blake and P.J.B. Hart
Light intensity:
Light Dark
Predator
Eel
Perch
Treatment
Visual andchemical stimuliVisual stimulionlyVisual andchemical stimuliVisual stimulionly
Crayfish position
In shelter
9(5)
10(5)
2(2)
2(2)
Exposed
0(0)
29(7)
1(1)
1(1)
Crayfish position
In shelter
13(5)
2(2)
0(0)
0(0)
Exposed
4(2)
10(4)
2(2)
1(1)
Table 3 The total number of visualencounters behveen juvenile crayfishand the fish predators in Experiments 1and 2. Figures in parentheses denotethe number of individual crayfishresponsible for the total number ofencounters shown
Crayfish did not orientate specifically to the headof the eels. They often reacted to the movement ofthe tail as it passed by. These data also indicatethat crayfish were able to see predators during thedark, possibly because red light was emitted by theinfra-red lamps.
After dusk, crayfish behaviour did not alter inresponse to seeing and/or smelling the fish, withthe exception of climbing behaviour in response toseeing eels (H = 5.1, df = 1, m = 16, « = 16, P < 0.025).Climbing activity occurred more than twice as oftenin the two treatments where crayfish could see eels,and may have been influenced by the fact that theseaquaria had clear sides (Fig. lc). It may be that thecrayfish could percieve the tank beyond the confinesof the aquaria in these treatments and that this in-fluenced their attempts to climb out of the aquaria.A similar trend in climbing behaviour was evidentin the perch experiment, but this was not significant{P>0.05;Fig. 2c).
Crayfish behaviour in the shelters
Generally, crayfish did not show a preference forany class of behaviour when they were inside theshelters. Some crayfish that could see and smell eelstook up 'defensive' positions more frequently duringthe two periods of light (Wilcoxon signed-rank test,T+=28, n=7, P<0.05; Fig. 3). Crayfish whichcould see and smell perch took up more 'active'shelter positions during the two periods of darkness(Wilcoxon signed-rank test, T̂ = 27, « = 7, P < 0.05).
100 r
5S>
Defensive Active Defensive ActiveEel tanks Perch tanks
Category of crayfishsheiter bflhoviour
Fig. 3 The behaviour of crayfish when using the shelters inExperiments 1 and 2 in the treatment with chemical and visualstimuli present. Values are means ( i l SE) of the percentageof 30-s counts spent in each behaviour. Time periods are • ,pre-dawn; E, post-dawn; •, pre-dusk; and^, post-dusk.Asterisks denote levels of significance for Wilcoxon pair-wisecomparisons between periods of light and darkness {*P < 0.05).
Crayfish behaviour in response to changes inillumination
In control treatments, crayfish behaviour was notinfluenced by changes in illumination except thatcrayfish spent less time walking after dawn in Ex-periment 2 (P<0.01, n = 8; Fig. 2b). Crayfish be-haviour did not alter at dawn when crayfish couldsmell the fish, Most crayfish were already within

shelters prior to dawn and remained so after dawn.Again, there was one exception. Crayfish that couldsmell but not see perch spent less time walking afterdawn, although this was not significant (P>0.05).After dusk, crayfish that could smell fish becamemuch more active, spending less time in shelters andmore time walking and climbing (Figs 1 and 2).
Crayfish that could only see eels spent less timewalking and more time under shelter after dawn(Fig. 1). Crayfish also spent less time walking inresponse to seeing perch (Fig. 2b). After dusk, crayfishthai could see perch became more active (Fig. 2). Nosimilar change in behaviour occurred in responseto seeing eels.
Discussion
Predators behaved in a similar manner in the twoexperiments and crayfish responded similarly toboth eels and perch. The time spent in the sheltersappears to be the best indicator of the defensivestate of the crayfish, as they reduced the chance ofvisual detection by a predator. The use of cover bycrayfish to minimize detection has been shown byStein & Magnuson (1976) and Appelberg & Odelstrom(1988). The use of shelter by P. leniusculus in theseexperiments is thus interpreted to be an avoidanceresponse. Walking and climbing are most likely tooccur in the absence of a predatory threat.
Crayfish {Astacus astacus L.) tend to be more activeat night, particularly in the presence of a crepuscularpredator (Hamrin, 1987). In this study, changes inlight intensity exerted only a weak influence oncrayfish behaviour in the control treatments. Overthe first three time periods, crayfish behaviourchanged in response to predator scent, confirmingprevious studies (Hazlett, 1985; M. Appelberg, pers.comm.). Crayfish showed their most marked changesin behaviour in response to both seeing and smellingthe fish. These crayfish spent more time in sheltersbefore dawn and during the day. After dusk therewas a switch to locomotory activity. This behaviourcorresponded to the loss of the dawn peak and theenhancement of the dusk peak in locomotory activityfound in A. astacus (L.) in response to the presenceof perch (Hamrin, 1987).
It was hypothesized that visual stimuli wouldonly cause crayfish to show defensive behaviour inthe light. This proved to be the case in treatments
Predator avoidance by iuvenile crayfish 95
where only visual stimuli were presented. Due tothe experimental design, there was little differencein the behaviour of the two predator species in thetanks. Perch were expected to be more active thaneeis, thus providing a stronger visual stimulus tothe crayfish. The encounter data show the oppositewas true, however, and we observed no conclusiveevidence to support the hypothesis that an increasedfrequency of visual disturbance would produce astronger avoidance response in crayfish.
If crayfish detect predators chemically they shouldshow behavioural responses in both the light anddark. This proved to be the case. Before dawn, crayfishincreased their use of the shelters in response topredator scent. This behaviour persisted duringthe post-dawn and pre-dusk periods. As a result,crayfish that could smell the predators showed nochange in shelter use in response to dawn. Wherecrayfish occur in sympatry with crepuscular pred-ators, reducing exposure prior to dawn should beselected for as this interrupts the chain of predator-prey interactions before a dangerous visual encountercan occur (Endler, 1991). it also enables crayfishto monitor the habitat from within a shelter thusreducing exposure to predators. Such an effect wasdemonstrated by the behavioural differences be-tween the crayfish that could both see and smell eelsand those that could only see eels. The crayfish inchemical contact with eels were most often withintheir shelters when a visual encounter took place.
Remaining stationary in a confined space may notbe a successful strategy for avoiding predation frompredators such as eels that feed by chemoreception.Capture might even be made easier. Thus the increasein walking and climbing by crayfish in response tothe reduction in light levels after dusk may reducethe chances of being eaten by nocturnal predators.A further result of the increased movement is thatit may increase the chances of crayfish moving tohabitats where there is a smaller risk of predation.Crayfish might be expected to commence feedingimmediately during the dusk activity peak after aperiod of reduced diurnal activity. The crayfish,however, spent the majority of the post-dusk periodeither climbing or walking.
Crustaceans are sensitive to their chemical en-vironment (Ache, 1982). The source of chemicalinformation about the proximity of predators maybe the predators themselves or their prey. Prey may

96 M.A. Blake and P.J.B. Hart
release alarm chemicals as a result of physical damageinflicted by a predator, or disturbance chemicals asa result of being disturbed by a predator {Hazlett,1985; Petranka et al., 1987; Alexander & Covich, 1991).Alarm chemicals are often low-molecular-weighthydrocarbons which differ between species (Carr,1986 for review). Fish of the superorder Ostariophysipossess an alarm chemical system and may learn torespond to other chemicals associated with the releaseof alarm chemicals. One such chemical stimulus isthe scent of the predator (Smith, 1992 for review).Hazlett (1990) suggested that crayfish (Orconectesvirilis Hagen) respond to conspecific disturbancesignals comprised of ammonia released from the gillsand pheromones released from the green gland inthe excretory system. Whilst their behavioural re-sponses to conspecific disturbance chemicals weregreater in magnitude, crayfish also responded todisturbance chemicals produced by different taxa,including the leech Macrobdella decora and the darterEtheostoma exile. This suggests that some componentof the disturbance chemicals, probably ammonia,was common to the different taxa. In this studycrayfish responded to the predators' scent, althoughit was not possible to determine the nature of thechemicals which produced the response.
Crayfish must obtain chemical information fromwater which is by nature a turbulent medium. Hencechemical concentrations will be patchy. By extension,information transfer will be discontinuous andadditional sensory information is often required toimprove search efficiency (Atema, 1988). This is likelyto be the case for predator detection by crayfish.In this study an interaction of visual and chemicalstimuli produced a greater behavioural reaction incrayfish than did either stimulus alone. Such aneffect has also been shown in the cyprinid fish,Leucaspius delineatus (Heckel) (RCippell & Gosswein,1979) and in other crayfish, whose latency of responseto visual stimuli declined after being exposed todisturbance chemicals (Hazlett, 1990). We suggestthat chemical perception of predators lowered thethreshold of the crayfish avoidance behaviour inresponse to visual stimuli in this study. This wouldmake evolutionary sense. Behavioural responsesshould be related to the degree of threat (Stein, 1979).If it is assumed that chemical stimuli act over tem-poral and spatial ranges greater than those of visualstimuli, then chemical detection of a predator should
predispose prey to adopt defensive behaviour beforea visual encounter. The distance between predatorand prey must be reduced for a visual encounter tooccur. This will represent a more dangerous situationfor many potential prey species and so the behav-ioural response should be greater.
Acknowledgments
Thanks to Ian Henderson and Andy Gill for theiradvice and help, and to David Holdich for his com-ments on the manuscipt. This research was fundedby a NERC case award (GR4/89/ALS/18), and furthersupported by Simontorp Aquaculture AB, Blentarp,Sweden.
References
Ache B.W. (1982) Chemoreception and thermoreception.The Biology of Crustacea Volume 3. Neurobiology: Struc-ture and Function (Ed. D.E. Bliss), pp. 369-398. Aca-demic Press, New York.
Alexander J.E. Jr. & Covich A.P. (1991) Predator avoid-ance by the freshwater snail Physella virgata in re-sponse to the crayfish Procambarus siniulans. Oecologia,87, 435-442.
Appelberg M. & Odelstrom T. (1988) Interaction be-tween European perch {Perca fluviatilis) and juvenilePadfastacus leniusculus (Dana) in a pond experiment.Freshwater Crayfish VII (Ed. P. Goeldlin de Tiefenau),pp. 37-45. Papers from the Seventh InternationalSymposium of Astacoiogy, Lausanne, Switzerland,3-5 August 1987.
Atema J. (1988) Distribution of chemical stimuli. SensoryBiology of Aquatic Animals (Eds J. Atema, R.R. Fay,A.N. Popperand W.N.Tavolga), pp. 29-58. Springer-Verlag, Berlin.
Bruski C.A. & Dunham D.W. (1987) The importance ofvision in agonistic communication of the crayfishOrconectes rusticus. I: an analysis of bout dynamics.Behaviour, 103 (1-3), 83-107.
Carr W.E.S. (1988) The molecular nature of chemicalstimuli in the aquatic environment. Sensory Biology ofAquatic Animals (Eds J. Atema, R.R. Fay, A.N. Popperand W.N. Tavoiga), pp. 3-28. Springer-Verlag, Berlin.
Dehli E. (1981) Abor og ferskvannskreps. Fauna, 34,64-67.
Disler N.N. & Smimov S.A. (1977) Sensory organs ofthe lateral line canal system in two percids and theirimportance in behaviour. Journal of the FisheriesResearch Board of Canada, 34, 1492-1503.

Edel R.K. (1975) The effect of shelter availability onthe activity of male silver eels. Helgolaender Wissen-schafttiche Meeresunters uchuttgen, 27, 167-174.
Endler I.A. (1991) Interactions between predators andprey. Behavioural Ecology. An Evolutionary Approach(EdsI.R. Krebs and N.B. Davis), pp. 169-196. BlackwellScientific Publications, Oxford.
Hamrin S.F. (1987) Seasonal crayfish activity as in-fluenced by fluctuating water levels and presence of afish predator. Holarctic Ecology, 10, 45-51.
Hazlett B.A. (1985) Disturbance pheromones in thecrayfish Orconectes virilis. Journal of Chemical Ecology,11(12), 1695-1711.
Hazlett B.A. (1990) Source and nature of disturbance-chemical system in crayfish, jounial of Chemical Ecology,16 (7), 2263-2275.
Hogger J.B. (1988) Ecology, population biology andbehaviour. Freshioater Crayfish. Biology, Managementand Exploitation (Eds D.M. Holdich and R.S. Lowery),pp. 114-144. Croom Helm. London.
Meddis R. (1984) Statistics Using Ranks: A UnifiedApproach. Basil Blackwell, Oxford.
Petranka J.W., Kats L.B. & Sih A. (1987) Predator-prey interactions among fish and larval amphibians:use of chemical cues to detect predatory fish. AnimalBehaviour, 35, 420-425.
Ruppell G. & Gosswein E. (1979) Die schwarme vonLeucaspius delineatus (Cyprinidae, Teleosteii) beigefahr in hellen und im dunkeln. Zeitschrift fuerVergleichende Physiologic, 76, 333-340.
Ryan P.A. (1984) Diel and seasonal feeding activity ofthe short-finned eel, Anguilla australis schmidtii, inLake EUesmere, Canterbury, New Zealand. Environ-mental Biotogy of Fishes, 11 (3), 229-234.
Siegel S. & Castellan N.J. Jr. (1988) Nonparametric Stat-istics for the Behavioural Sciences. McGraw-Hill,Maidenhead.
Predator avoidance by juvenile crayfish 97
Sih A. (1987) Predators and prey lifestyles: an evol-utionary and ecological overview. Predation. Directand Indirect Impacts on Aquatic communities (Eds. W.C.Kerfoot and A. Sih), pp. 201-224. University Pressof New England, Hanover.
Smith M.R. & Dunham D.W. (1990) Chela postureand vision: compensation for sensory deficit in thecrayfish Orconectes propinquus (Girard) (Decapoda,Cambaridae). Crustaceana, 59 (3), 309-313.
Smith R.J.F. (1992) Alarm signals in fishes. Reviews inFish Biology and Fisheries, 2, 33-63.
Stein R.A. (1977) Selective predation, optimal foraging,and the predator-prey interaction between fish andcrayfish. Ecology, 58, 1237-1253.
Stein R.A. (1979) Behavioural response of prey to fishpredators. Predator-Prey Systems in Fisheries Manage-ment (Eds R.H. Stroud and H. Clepper), pp. 343-353.International Symposium on Predator-Prey Systemsin Fish Communities and their Role in FisheriesManagement. Aflanta, Georgia, 24-27July 1978. SportFishing Institute, Washington, D.C.
Stein R.A. & Magnuson J.J. (1976) Behavioural responseof crayfish to a fish predator. Ecology, 57, 751-761.
Svardsson G. (1972) The predatory impact of eel (Anguillaanguilla L.) on populations of crayfish {Astacus atacusL.). Report of the Institute of Freshwater Research.Drottningholm Nr. 52, 149-191.
Tesch F.-W. (1977) Biology and Management of AnguillidEels. Chapman and Hall Ltd, London.
Wine J.J. & Krasne F.B. (1982) The cellular organisationof crayfish escape behaviour. The Biology of CrustaceaVolume 4. Neural Integration and Behaviour (Ed. D.E.Bliss), pp. 242-292. Academic Press, New York.
{Manuscript accepted 8 September 1992)
