the alpha chains of goat hemoglobins: old and new variants in native apulian breeds
TRANSCRIPT

www.elsevier.com/locate/cbpb
Comparative Biochemistry and Physio
The alpha chains of goat hemoglobins: Old and
new variants in native Apulian breeds
Elisa Pieragostini a,*, Rosario Rullo b, Andrea Scaloni b, Grazia Bramante a, Aldo Di Luccia c
aDepartment of Engineering and Management of the Agricultural, Livestock and Forest Systems�University of Bari, 70100 Bari, ItalybISPAAM�National Research Council, 80147 Napoli, Italy
cDepartment of Animal Production�University of Bari, 70100 Bari, Italy
Received 18 January 2005; received in revised form 29 April 2005; accepted 5 May 2005
Available online 11 July 2005
Abstract
Blood samples were collected from 324 goats belonging to the native Apulian breeds Garganica and Jonica; 60 Alpine goats were also
sampled to serve as a comparison. Hemoglobin phenotypes were analyzed with isoelectric focusing in a pH range of 6.7–7.7. Heterogeneity
of globin chains was evidenced both by AUT-PAGE and RP-HPLC. The primary structure of four alpha globins was analyzed by combined
mass spectrometry approaches. Two of these globins had never been sequenced before. One was a new alpha variant, an allele of the HBA1A
gene from which it differed for the mutation A26T and has been registered with a low frequency only in Apulian breeds; the other was a
globin encoded by the HBA2 locus, whose primary structure was previously derived from the corresponding gene. The two alleles recorded
at the HBA2 locus presented a different frequency in the three breeds but may be considered to be generally rather common. Notwithstanding
the sample size no goat was found to exhibit HbA1B. The Authors discuss their findings in the light of the results reported by other
researchers and argue that, in spite of what had been inferred in pioneer works on goat hemoglobins, HBA1B is not a common allele.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Goat hemoglobin; a-chain; Primary structure; Non allelic variants; Gene expression; Gene frequency; Apulian native breeds; Supernumerary
haplotypes; CpG site; Hot spot
1. Introduction
In Apulia, the region covering the heel of the boot-
shaped Italian peninsula, there are 8 recognized native
breeds of farm animals. The rather harsh pedoclimatic
conditions as well as the selective pressure of endemic
hemotropic parasites have yielded genetic pools that are
generally rustic and tolerant to the diseases caused by these
parasites. Phenotypic variability induced by parasites is a
matter of fact, as impressively exemplified by the high
number of hemoglobinopathies in human populations living
in malaria-endemic areas (Evans and Wellems, 2002). The
past decade has registered remarkable advances in the study
1096-4959/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.cbpc.2005.05.005
* Corresponding author. Dipartimento PROGESA-Facolta di Agraria, via
Amendola 165/A, 70126 Bari, Italy. Tel.: +39 080 5442842; fax: +39 080
5442504.
E-mail address: [email protected] (E. Pieragostini).
of hemoglobin polymorphism in Apulian livestock (Pier-
agostini et al., 1994; Scaloni et al., 1998; Pieragostini et al.,
2000) and a special focus has been the peculiar hemato-
logical features of native breeds which seem to be related to
their general ability to thrive in endemic TBD areas
(Pieragostini and Petazzi, 2000). In particular, extra alpha
genes were found both in sheep and horses (Pieragostini et
al., 2002, 2003).
The alpha globin genes are a relevant example of gene
families which have arisen from a single gene by gene
duplication events (Hardison, 1998). In mammals, non-
allelic loci produce unequal amounts of alpha globin which
may give rise to quantitative polymorphism with the
upstream gene being the most efficient; in particular, as
observed in sheep, the percent gene efficiencies in the aa
and aaa gene haplotypes, from the 5Vto the 3Vend, are about32–18 and 30–14–6, respectively (Vestri et al., 1991). A
recent investigation on Apulian native sheep compared the
logy, Part B 142 (2005) 18 – 27

Hemoglobin Tetramers Globin chain
responsible
for the Hb
name
References
Protein sequence Nucleotide
sequence
HbA a2Ah2
A IaA Huisman et al.,
1968
Schon et al.,
1982HbB a2Bh2
A IaB
HbC a2Ah2
C Ch Huisman et al.,
1967
Haynes et al.,
1980a2Bh2
C
HbD a2Ah2
D hD Adams et al.,
1968
Pirastru et al.,
2000
HbDMalta a2Ah2
D Malta hD Malta Bannister et al.,
1979
Pirastru et al.,
2000
HbE a2Ah2
E hE Wrightstone
et al., 1970
Pirastru et al.,
2000
HbF a2Ag2 g Kleinschmidt and
Braunitzer, 1982
Haynes et al.,
1980a2Bg2
E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–27 19
total amount of a�globin chains with the total h�chains and
found that the a /h ratio appears to be linearly related to the
number of a�globin genes; specifically, the more genes an
individual possesses, the greater the imbalance. The related
hematological pattern, which mimics a thalassemia-like
syndrome, was suggested to impart some protection against
endemic TBD parasites. This is analogous to what happens
in thalassemic red blood cells where malaria parasite growth
is unimpaired (Nagel, 1990). In Apulia, the relatively high
frequency of individuals characterized by unusual extra
a�globin genes may then be taken as a telltale signature of a
positive selection (Pieragostini et al., 2003).
Based on these considerations, we have focussed our
attention on the alpha globin genetic system in Apulian
native goat breeds. Unfortunately since it was first reported,
hemoglobin polymorphism in goat populations appeared to
be a complex issue.
The first biochemical studies were performed in the late
sixties and seventies, directly on the primary structure of
five h-and three a-globin chains (as reviewed by Garrick
and Garrick, 1983). Investigations into fetal hemoglobin
highlighted the existence of monomorphism at the gamma
gene level while heterogeneity was related to the presence of
polymorphic alpha globins (Wilson et al., 1969); analo-
gously, hemoglobin switching to HbC in adults during
anemic episodes is known to conceal polymorphism at the
beta globin level because of the substitution of all beta
variants by the beta C globin chain encoded by the dormant
gene Ch. In the eighties research by the Cincinnati Group
thoroughly explored the structure and evolution of the h-globin gene clusters and of the relative developmentally
regulated genes, hF, Ch and hA (Lingrel et al., 1985). The
primary structure of a fourth a-globin was deduced from the
nucleotide sequence (Schon et al., 1982) followed by the
nucleotide sequence of the embryonic alpha globin gene
(zeta) and by the evolutionary analysis of the complete
alpha globin cluster (Wernke and Lingrel, 1986).
Parallel investigations undertaken by European research-
ers with high resolution techniques provided new insights
into hemoglobin polymorphism in goat populations (Tucker
et al., 1983; Braend et al., 1987a,b). Using IPG electro-
phoresis, which is a very sensitive technique, the HbA band
was found to be a mixture of tetramers, differing for the beta
chains produced by the hA2, hA4, hA6, hA8 beta alleles
(Braend et al., 1987a) and simply defined on the basis of
their different electrophoretic mobility. In the nineties no
steps were taken to identify the electrophoretic variants
found so far but in 2003 Pirastru et al. analyzed the DNA
sequences of four new beta genes (hL125E, hK104R, L125V,
hQ87H, K104R, hK104R), possibly encoding for the beta globin
allele described by Braend et al. (1987a).
Thus, present knowledge of goat hemoglobin may be
summarized into two essential points. 1) As to the alpha
globin system, the protein sequence of four non-allelic
alpha-chains has been described in the literature; two of the
chains are encoded by the 5Vupstream gene-IaA and IaB-and
the other two by the 3Vdownstream gene (Huisman et al.,
1967; Schon et al., 1982). 2) For the beta globin system, the
protein sequence of four allelic (hA, hE, hD and hD Malta)
and two non-allelic globin genes (hF and Ch) can be
numbered. Only seven of the possible tetramers resulting
from the combination between such a heterogeneity of alpha
and beta globins are identified by a name, i.e., HbA, HbB,
HbE, HbD, HbDMalta, HbF and HbC; the former two
derived their name from the respective alpha globin, all the
others from the respective beta globin. These pieces of
information are catalogued according to the nomenclature
for polypeptide chains as described by Huisman and
Schroeder (1971) and listed in the following scheme.
In particular, the findings concerning HbB appear to be
controversial because HbDMalta and HbB exhibit a similar
electrophoretic behavior and, in the absence of other
analytical investigations or biological trials (such as
response to anemic stress or developmental investigations),
one cannot exclude that HbDMalta (Bannister et al., 1979)
may have been misidentified as HbB. In other instances,
details from quantitative evaluations offer indications as to
the nature of the recorded polymorphism; in fact, due to the
already mentioned characteristic differences in the average
levels of globin production in a�loci, the relative densities
of the electrophoretic bands can deduce many genotypes.
Hence, even supernumerary haplotypes, where present, may
be detected. The aim of this work, therefore, is to expand the
evidence provided so far by thoroughly investigating alpha
globin polymorphism in two native Apulian goat breeds, the
Garganica and the Jonica, and by evaluating the results in
the light of literature.
2. Materials and methods
2.1. Animals
Blood samples were collected from 324 individual goats
(Capra hircus) belonging to the Garganica and Jonica

E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–2720
breeds and reared in pure breeding on different farms. A
total of 60 individual samples from a flock of over 1000
pure Alpine goats were also analyzed to serve as a
comparison since this breed is present throughout the world.
Blood samples were processed through standard procedures
as described in Di Luccia et al. (1991).
2.2. Isoelectric focusing
Lysates were analyzed by isoelectric focusing (IEF) in a
narrow pH range (6.7–7.7) as already described by Di
Luccia et al. (1991) to assess the Hb phenotypes. Gel slabs
were scanned with a computerized Ultroscan XL enhanced
laser densitometer equipped with gelscan 2.0 software from
Pharmacia-LKB (Uppsala-Sweden).
2.3. Reverse phase HPLC
Globin chain separation was performed by a reverse
phase HPLC procedure described by Manca et al. (1991).
Hemoglobin solutions were brought to a final concentration
of 70–100 Ag and filtered through 0.45 Am HV filters
(Millipore, Bredford, MA). Fifteen AL of globin solution
were then applied onto a large pore Vydac C4 column
250�4.60 mm, (300A) (The Separation Group, Hesperia,
USA). The chromatogram was developed with a linear
gradient between solvent B (60% acetonitrile containing
0.1% trifluoroacetic acid ) and solvent A (20% acetonitrile
containing 0.1% trifluoroacetic acid ) at a flow rate of 1 mL/
min. The gradient was 48–65% developer B in 85 min. The
HPLC equipment was a Beckman Gold System.
2.4. Acid-Urea Triton X-100 polyacrylamide gel electro-
phoresis (AUT-PAGE)
Acid-Urea Triton X-100 polyacrylamide gel electro-
phoresis (AUT-PAGE) of the globin chains was carried
out according to Di Luccia et al. (1991).
2.5. Structural analysis
Electrospray mass spectra of intact globins were
recorded by using an API-100 single quadrupole mass
spectrometer (Applied Biosystems, USA) equipped with an
atmospheric pressure ionization source, as reported else-
where (Scaloni et al., 1998). Mass calibration was
performed by means of the multiply charged ions from a
separate injection of horse heart myoglobin (molecular
mass 16,951.5 Da). All masses have been reported as
average values.
Trypsin and endoproteinase Asp-N digestions of indi-
vidual globin chains were performed in 0.4% ammonium
bicarbonate, pH 8, at 37 -C, overnight, using an enzyme to
substrate ratio of 1 :100 (w/w).
Peptide mixtures were loaded onto the MALDI target
using the dried droplet technique and a-cyano-4-hydroxy-
cinnamic as a matrix and analysed with a Voyager-DE PRO
mass spectrometer (Applied Biosystems, Framingham,
USA). Internal mass calibration was performed with
peptides deriving from protease autoproteolysis. The mass
spectra were acquired in either reflectron or linear mode
with delayed extraction. Post-source decay fragment ion
spectra were acquired for intense signals after isolation of
the appropriate precursor by using timed ion selection, as
already reported (Scaloni et al., 1998).
Edman degradation was performed using a Procise 491
protein sequencer (Applied Biosystems, Foster City, USA)
equipped with a 140 C microgradient apparatus and a 785A
UV detector for the automated identification of phenyl-
thiohydantoin amino acids.
2.6. Statistics
To reduce bias in the quantitative evaluation of gene
products and consequent differential gene expressions, only
homozygous phenotypes at the HbA2 locus were selected
from the data set obtained by densitometric evaluation of the
Hb bands, and least square means with their standard
deviations were calculated.
The alpha gene frequencies were estimated assuming
equilibrium conditions and statistically assessed with the
v2 test.
3. Results
3.1. Isoelectric focusing of hemoglobins
Polyacrylamide gel isoelectric focusing in a narrow pH
range (6.7–7.7) of goat hemoglobins carrying different a
and h globin chains showed various hemoglobin band
patterns (Fig. 1). Two main electrophoretic zones were
recognized in the gel; Fig. 1 clearly shows that each major A
and/or B zone consists of stronger forward bands and one or
two weaker backward bands.
A more complex hemoglobin pattern was observed in
lanes 4 and 5 where the minimal difference of isoelectric
points observed (0.04 pH units) suggested the presence of
globin variants bearing non-charged amino acid substitu-
tions. Resolving power reduction towards the cathode was
due to gradient drifting for an electro-osmotic effect.
3.2. Densitometric evaluation of Hb bands
Table 1 shows the output of the HBA1 and HBA2 genes
as estimated from the concentrations of different alpha
globin chains on the basis of the densitometric evaluation of
their relevant Hb PAGIF bands (Fig. 1). The values
observed have been compared to the values expected on
the basis of those reported in the literature and those of
previous experiences with sheep and buffalo (Pieragostini et
al., 2003; Iorio et al., 2004).

Table 1
Output of alpha I and alpha II genes in goat as compared with buffalo and
sheep
Specie N Ia IIa Ia / IIa Reference
Goat 120a 32.02T2.59 18.19T1.44 Present paper
21 36.32T2.73b – Deduced from
Braend et al., 1987b
Buffalo 362 32.56T3.12 17.50T2.6 1.86 Iorio et al., 2004
Sheep 30 32.20T2.68 17.60T1.91 1.83 Pieragostini et al.,
2003
a To reduce bias, only densitometric data of homozygotes at alpha 2 locus
were considered.b This finding refers to the so-called reverse type that supposedly takes
into account the expression level of a single Ia gene.
Fig. 1. Polyacrylamide gel isoelectric focusing in narrow pH range, 6.7–
7.7, of goat hemoglobins carrying different a and h globin chains. Sample
1: two non-allelic alpha globins (IaA and IIaT) and one beta globin (hA )
result in two hemoglobin bands, Ia2Ah2
A and IIa2Th2
A; sample 2: two non-
allelic alpha globins (IaA and IIaT) and two beta globins (hA and hDMalta)
result in a twin couple of hemoglobin bands, the former couple migrating
in the zone A (Ia2Ah2
A and IIa2Th2
A) and the latter one in zone B (Ia2Ah2
DMalta
and IIa2Th2
DMalta); sample 3: one non-allelic alpha globins (IaA), two allelic
alpha globins (IIaA and IIaT) and two beta globins (hA and hD) result in a
twin triplet of hemoglobin bands, the former triplet migrating in the zone
A (Ia2Ah2
A, IIa2Th2
A and IIa2Ah2
A) and the latter one in zone B (Ia2Ah2
D, IIa2Th2
A
and IIa2Ah2
D); sample 4: two allelic alpha globins (the wild type IaA and the
new variant IaT), a non-allelic alpha (IIaA) and two beta (hA and hE) result
in six hemoglobins all migrating in the zone A; sample 5: the same alpha
globins as in sample 4 (IaA, IaT and IIaA) but different beta globins (hE
and hD) result in six hemoglobins among which the three tetramers
containing the hE globin migrate in the zone A, the others containing the
hD globin in the zone B.
E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–27 21
No patterns showing unusual quantitative polymorphism
were found, suggesting that only common duplicate alpha
arrangements were present in the sampled populations.
3.3. Electrophoretic and chromatographic analyses of
globins
The results of the AUT-PAGE and RP-HPLC analyses of
two hemoglobin phenotypes relevant for the purposes of our
interests are shown in Fig. 2. Both separation techniques are
primarily based on hydrophobicity and well define the
tetramer composition, displaying the differences between
the a and h globins. Fig. 2A shows the AUT-PAGE and RP-
HPLC globin pattern of one of the phenotypes exhibiting
the new alpha variant. As to the AUT-PAGE, it can be
observed that, at the alpha globin zone, two major bands, as
well as a minor one are detectable. The IaA globin chain is
slower than the new alpha variant, attesting a primary
structure change with a decrease in hydrophobicity. The IEF
results excluded the substitution of a charged amino acid so
the effect recorded can be ascribed to a neutral to polar
amino acid substitution. RP-HPLC analysis of the above
samples confirmed the results obtained by AUT-PAGE.
In the beta globin zone, a co-migration of hA ed hDmalta
can be observed since in an acid environment the Asp
residue at position 69 in the hDmalta is not charged and
behaves like a neutral or polar amino acid. Conversely, these
two beta chains were well resolved by RP-HPLC (Fig. 2B).
RP-HPLC analysis of the above samples confirmed the
results obtained by AUT-PAGE. Furthermore, direct molec-
ular mass measurement by ESI-MS unambiguously ascer-
tained the nature of each globin component. Globin identity,
defined by this combined approach, definitively demon-
strated the occurrence of a new a-globin variant in goat
hemolysates, together with the ubiquitous absence of IaB.
As to the genes responsible for different products, the
evaluation of the relative peak areas and retention times
confirmed that the slower-migrating proteins are encoded by
the upstream a-globin gene while the faster-migrating
proteins are encoded by the downstream a-globin gene.
3.4. Structural analysis
AMALDI-TOF peptide mass mapping approachwas used
to determine the primary structure of this new variant and
confirmed the nature of the other a-globins observed.
Analysis of tryptic digests highlighted the strict structural
homology between IaA and the new variant, suggesting that
the amino acid differences were confined to peptide 17–31,
as shown in Fig. 3A and B. These results were confirmed by
independent mass mapping analysis on endoprotease Asp-N
digests (data not shown). Post-source decay MALDI-TOF
analysis of the mutated peptide at m/z 1465.6 definitively
demonstrated the occurrence of the A26T substitution in this
new a-globin variant (Fig. 3C). These results were confirmed
by Edman degradation analysis on electroblotted protein
samples. The nature of this amino acid replacement was in
perfect accordance with the relative migration observed in
AUT-PAGE and RP-HPLC. Table 2 summarizes the data
obtained for the different goat a-globins reported in this study
and compares them with those reported in the literature.
3.5. Nomenclature
On the basis of the structure analyses and in agreement
with what had been suggested in previous papers on
sheep and buffalo (Pieragostini et al., 2003; Iorio et al.,
2004) and in the guidelines for gene nomenclature in
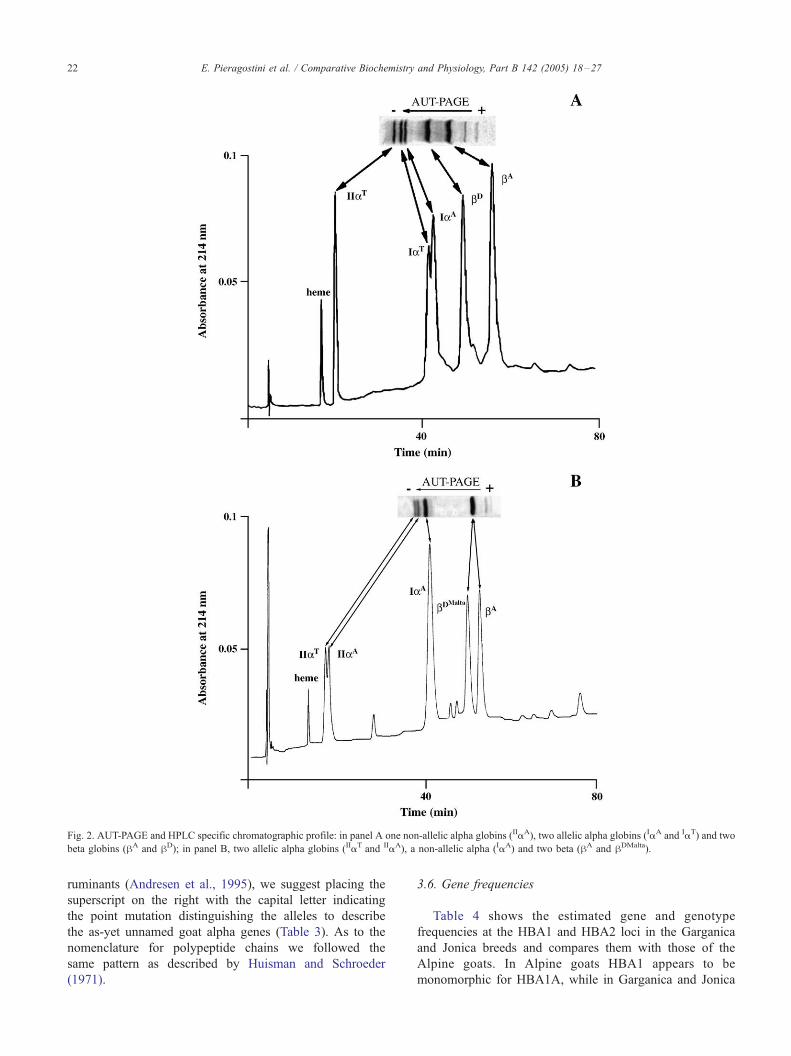
Fig. 2. AUT-PAGE and HPLC specific chromatographic profile: in panel A one non-allelic alpha globins (IIaA), two allelic alpha globins (IaA and IaT) and two
beta globins (hA and hD); in panel B, two allelic alpha globins (IIaT and IIaA), a non-allelic alpha (IaA) and two beta (hA and hDMalta).
E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–2722
ruminants (Andresen et al., 1995), we suggest placing the
superscript on the right with the capital letter indicating
the point mutation distinguishing the alleles to describe
the as-yet unnamed goat alpha genes (Table 3). As to the
nomenclature for polypeptide chains we followed the
same pattern as described by Huisman and Schroeder
(1971).
3.6. Gene frequencies
Table 4 shows the estimated gene and genotype
frequencies at the HBA1 and HBA2 loci in the Garganica
and Jonica breeds and compares them with those of the
Alpine goats. In Alpine goats HBA1 appears to be
monomorphic for HBA1A, while in Garganica and Jonica
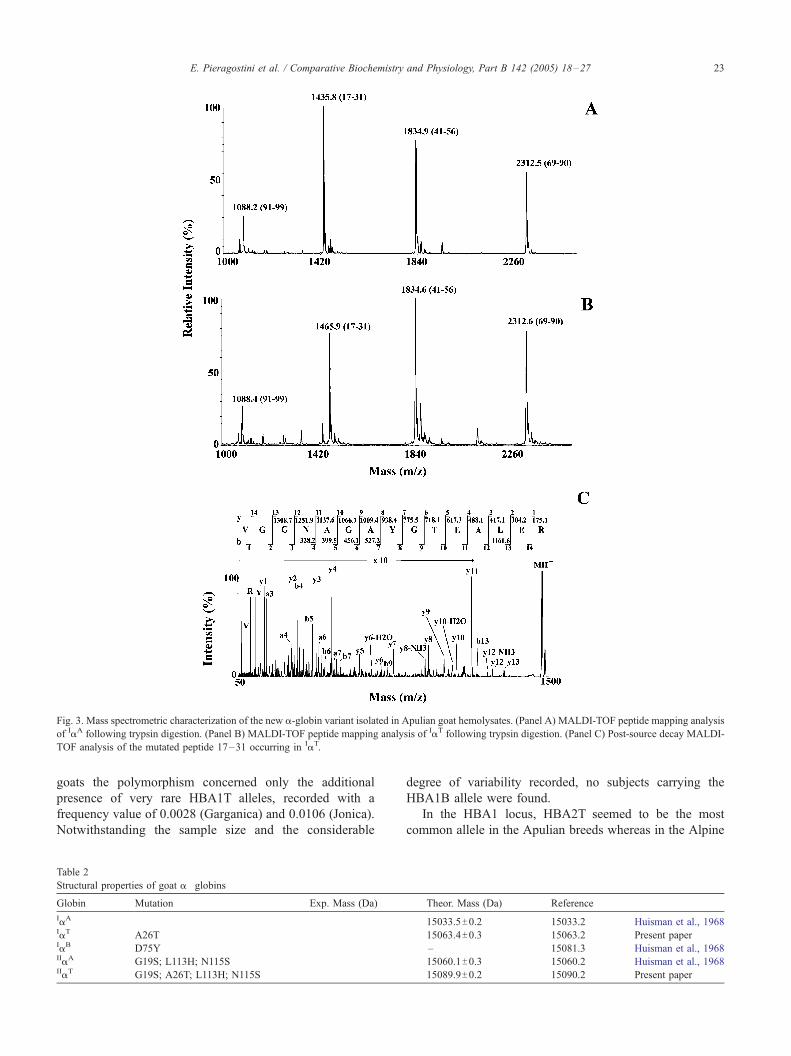
Fig. 3. Mass spectrometric characterization of the new a-globin variant isolated in Apulian goat hemolysates. (Panel A) MALDI-TOF peptide mapping analysis
of IaA following trypsin digestion. (Panel B) MALDI-TOF peptide mapping analysis of IaT following trypsin digestion. (Panel C) Post-source decay MALDI-
TOF analysis of the mutated peptide 17–31 occurring in IaT.
E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–27 23
goats the polymorphism concerned only the additional
presence of very rare HBA1T alleles, recorded with a
frequency value of 0.0028 (Garganica) and 0.0106 (Jonica).
Notwithstanding the sample size and the considerable
Table 2
Structural properties of goat a�globins
Globin Mutation Exp. Mass (Da)
IaA
IaT A26TIaB D75YIIaA G19S; L113H; N115SIIaT G19S; A26T; L113H; N115S
degree of variability recorded, no subjects carrying the
HBA1B allele were found.
In the HBA1 locus, HBA2T seemed to be the most
common allele in the Apulian breeds whereas in the Alpine
Theor. Mass (Da) Reference
15033.5T0.2 15033.2 Huisman et al., 1968
15063.4T0.3 15063.2 Present paper
– 15081.3 Huisman et al., 1968
15060.1T0.3 15060.2 Huisman et al., 1968
15089.9T0.2 15090.2 Present paper

Table 3
Nomenclature of alpha globin genes and products
HBA1 Locus HBA1 Locus
Gene Symbol HBA1A HBA1B HBA1T HBA2A HBA2T
Globin symbol IaA IaB IaT IIaA IIaT
E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–2724
goats both HBA2T and HBA2A were present with almost
the same frequency.
The alleles and genotypes in each breed sample were in
HW equilibrium.
4. Discussion
The main information provided by our results concern: a)the lack of evidence that alpha globin extra genes are
present; b) the identification of a new alpha globin variant
and the primary structure of the globin encoded by the alpha
globin gene sequenced by Schon et al. (1982); c) the
quantitative data to deduce the output of alpha genes and to
assign the new variant to the appropriate gene; and d) the
allele frequencies at the alpha loci.
4.1. Alpha globin genes and chains
In Table 2, the values recorded in goats have been
compared with the other species for the HbA1 and HbA2
gene outputs. Based on the size of the data set and the fact
that the range of variability records is comparable to that
reported previously for sheep and buffalo with the same
duplicated gene arrangements (Pieragostini et al., 2003;
Iorio et al., 2004) the values in Table 2 may confidently be
defined as appropriate levels of gene expression for goats,
and more generally, the alpha gene expression gradient for
goats is confirmed to be close to a 38% output reduction of
the 5Vupstream gene.
On the basis of the quantitative evaluation of the PAGIF
bands and the RP-HPLC peaks, it can be inferred that the
new alpha variant found is encoded by an allele of the
HBA1A gene from whose product it differs for the
substitution of alanine with threonine at position 26. This
result is consistent with the results obtained through previous
analytical approaches with the side chain hydrophobicity
values for alanine and threonine amounting to 0.616 and
0.359, respectively (Black and Mould, 1991). The difference
of 0.257 between the two justifies the electrophoretic and
chromatographic behavior of the corresponding proteins.
Table 4
Estimated gene and genotype frequencies both HB1A and HB2A loci in Gargani
Breed Herds Genotypes at HBA1 locus Genotypes at H
AA AT AA AT
Garganica 5 182 1 13 63
Ionica 7 141 3 8 19
Alpine 1 60 0 17 25
The substitution of alanine with threonine at position 26
is present not only in the newly detected Ia variant but also
in the sequence of the globin chain encoded by one of the
alleles at the IIa locus. Yet, considering the differences in the
amino acid sequences of the two chains, this is not the case
of a tandem repetition of individual gene copies. Thus the
phenomenon cannot be included amongst the genomic
mechanisms of turnover capable of achieving sequence
homogeneity, such as gene conversion or unequal crossing-
over between individual copies of the gene family. The
presence of the same mutation in the two non-allelic genes
in goats is more likely to be associated with independent
events since the change of alanine to threonine at position
26 involves a CpG dinucleotide which is a mutation hot-
spot. According to Perutz (1990), however, the presence of
the highly mutagenic CpG dinucleotide is a necessary but
not a sufficient condition for a mutation to occur. Only
CpGs methylated in the presence of a specific methylase
(Roy and Weissbach, 1975) act as hot spots because
deamination converts 5-methylcytosine to thyamine which
then pairs with adenine.
Position 26, as may be observed by analyzing the amino
acid sequences of mammalian a-globin chains present in the
data bank (ExPASy Proteomics Server: http://www.expasy.
org/), is constantly occupied by an alanine residue. The
evidence reported above suggests that only goat Ia-and Ia-
globin genes contain methylated CpG, thus producing the
Ala26Thr mutation. However, one cannot exclude that this
conservative substitution exists in other species because it
may have escaped detection since it does not interfere with
structure and is not electrophoretically visible by conven-
tional methods.
4.2. Frequency data
The focal point of a discussion on frequency data is the
comparison between populations to measure genetic dis-
tances or check latitudinal or altitudinal trends so that the
adaptive significance of genes, such as beta B in sheep
(Agar et al., 1972; Pieragostini et al., 1994), may be
accounted for. Unfortunately, there are not many data in the
literature to compare with the variability recorded in this
investigation on goats.
As to frequency values at the HBA1 and HBA2 loci, the
results concerning the HBA2 alleles in native Apulian goats,
where IIaT is the most frequent form, are very similar to
those reported by Pirastru et al. (2003) in Sardinian breeds
ca and Jonica breeds compared to Alpine one
BA2 locus Alleles at HBA1 locus Alleles at HBA2 locus
TT HBA1A HBA1T HBA2A HBA2T
107 0.997 0.0028 0.2432 0.7568
90 0.989 0.0106 0.1736 0.8264
18 1 0 0.4917 0.5083

E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–27 25
and by Nyamsamba et al. (2003) in Mongolian goats whereIIaT practically seems to be the only form present. By
contrast both alleles are equally present in the Alpine goat
population-at least as far as our sample is concerned. As to
the HBA1 alleles, the IaT variant seems to be very similar
both for analytical behavior and frequency to that found in
Sardinian goats (Pirastru et al., 2003) but the most intriguing
point is the absence of HbA1B (Table 4). Since the findings
on HbA1B in various populations, as already pointed out,
seem to be rather controversial, we reconsidered our results
in the light of those found in the relevant literature. It is
worth remembering that in 1979 Bannister et al. highlighted
that the Ia chain variant HbB was absent in Maltese goats.
Unfortunately, several of the investigations inferring the
occurrence of HbB on the basis of electrophoretic evidence
did not confirm the identification with structural techniques
or biological trials (Nguyen and Bunch, 1980; Tunon et al.,
1987; Igarashi et al., 2000). In other cases, such as in Sartore
et al. (1981) or Fesus et al. (1983), the conclusions drawn by
the Authors are not entirely corroborated by well-grounded
arguments. Sartore et al. (1981) analysed 398 adults and 568
kids of the Saanen breed by starch gel electrophoresis and
found both HbA and HbB in the adults while the kids were
monomorphic for HbA; considering that the frequency
value assumed for HBA1B was 0.11 in the adults, it is rather
surprising that no HBA1B genes were found in such a large
sample of kids. Analogously, in their evaluation of
hemoglobin polymorphism in native Hungarian breeds
Fesus et al. (1983) took into account the presence of a
certain number of HbB goats but, in the mean time,
concluded that all the newborn kids exhibited a single type
of hemoglobin. In the erythrocytes of newborn kids only
foetal hemoglobin can be found, thus polymorphic alpha
genes, where present, should be evident while polymor-
phism at the beta locus is known to become apparent only
after the start of adult beta chain synthesis. Hence, whether
the lack of HbB in the kids studied was a sampling effect or
the HbB in the adults was an effect of misidentification
based on the analytical system remains a moot question.
In other cases an initial misidentification of HbB was
recognized and corrected by the biological trials. Tucker et
al. (1983) bled their supposed AB and BB goats but, rather
surprisingly, they found them to produce an HbC identical
to that produced by AA goats. Real biallelism at HBA1
would have produced two different tetramers, namely
a2ACh2 and a2
BCh2. The presence of the same band proved
that the polymorphism was due to allelism at the h locus
and particularly that the HbB band (a2Bh2
A) was mistaken for
that of the HbD (a2Ah2
D). In particular, IaB differing from
IaA for the amino acid substitution D75Y and hD and hD
differing from hA, respectively for a D21H and D69G, it
implies that the corresponding hemoglobins exhibit almost
the same electrophoretic mobility at alkaline pH. Thus, it is
possible that Hb D and DMalta may be mistaken for HbB
when there is no unambiguous identification. In the case of
Hb D the confusion can be generated only at pH above 7
because titration and the loss of charge of the His residue—
whose pKa is 6-can be observed in an alkaline environment.
The existence of the HbB seems to be unquestionable.
Huisman et al., 1967, 1968 extensively documented
biallelism at the Hb1A locus. Electrophoretic, chromato-
graphic and structural analyses and biological trials pro-
vided evidence for the existence of the A and B alpha
variants. On the basis of these results Huisman et al. (1967)
suggested that goat hemoglobin polymorphism was similar
to that of the buffalo in that it was due to variations in the
alpha globin genetic system. The only weak point in this
argument is that structure analyses of the alpha chain variant
HbB were all performed sampling the blood of a single goat,
namely Huisman’s goat n-5 of an unidentified breed, while
the analytical procedures and subsequent results seem to be
indisputable.
Subsequently John and Barnabas (1978) sampled goats
from a local slaughterhouse in India and found goats that
were phenotypically HbA, HbAB and HbB. The identi-
fication was based on starch gel electrophoresis of
hemoglobins and DEAE-Sephadex chromatographic sepa-
ration of globins.
Actually, Braend might have found an HbB goat when
analyzing an experimental herd in Norway. In their paper of
1987b, Braend et al. described the presence of ‘‘hemoglobin
reverse phenotypes’’ detected by IEF with Immobiline gels
at a pH gradient of 6.9–7.5. The crucial point is that this pH
range reveals only tetramers which normally migrate into
the A zone, while potential Ia2Bh2
A (amounting to 30% of the
total Ia2h2 concentration) may remain undetected. In this
case, the zone A may be characterized by weaker forward
bands (30% of the Ia2Ah2 tetramers) and stronger backward
bands (40% of the IIa2h2 tetramers). This view seems to be
supported precisely by the findings reported in Braend et al.
(1987a). In this paper, Braend, together with Tucker and
Clarke, examined 150 Norwegian goats by PAGIF in a pH
range of 6–8 and found 5 HbAD individuals; in order to
resolve the variability recorded in the A zone, the remaining
HbA goats were analyzed only by Immobiline gels and
curiously no reverse phenotypes were found! It is our
opinion that Braend et al. (1987b) simply did not consider
the possibility of a IaB allele, chose a system a priori which
did not permit the detection of HbB tetramers and were then
biased in evaluating the results of the screening.
Finally, Johnson et al. (2002) found HbB in Omani goats.
Although there are no structural analyses or biological trials,
the densitometric data of the Hb bands suggested that in one
of the three breeds the IaB gene was rather frequent.
Evidence of this conclusion was provided by the AB
phenotypes which exhibited two-band patterns differing in
the relative band intensities (one with 67% A and 33% B
and the other with 67% A and 33% B). The analytical
procedure they used does not permit separation of different
Ia and IIa in the A zone. Thus, considering the data in Table
2, the A band is supposedly composed by the tetramers
encoded both by the Ia genes and IIa genes, with relative

E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–2726
efficiencies of about 32 and 18, respectively. This entails
that the truly AB Omani goats are only those exhibiting the
67:33 band pattern; 67 being equal to the percent gene
efficiencies of both IIa genes plus that of the single IaA
genes (�36%+�32%), while 33% is equal to the percent
gene efficiencies of the single IaB gene. The goats
exhibiting the 33:67 band pattern reasonably seem to be
BB where 33 is equal to the percent gene efficiencies of twoIIa genes, while 67 is equal to the percent gene efficiencies
of the two IaB genes. The same type of misinterpretation the
results may have occurred in Canatan and Boztepe (2000)
who did not find BB goats in their populations, though a
relatively high number of heterozygotes was recorded in
both cases.
The above considerations suggest that the IaB is not a
common allele. Thus, in contrast with the conclusions
drawn by Huisman et al. (1967), hemoglobin polymorphism
in goats is not similar to that of the buffalo but to that of the
sheep where the rare allele, IaD, has been found only in
some European breeds (Tucker, 1981) and particularly in
Apulian native sheep (Pieragostini et al., 1994, 2003)
although the suggested similarities with sheep do not
include the presence of supernumerary haplotypes for the
time being.
Acknowledgements
We are indebted to Dr Athina Papa for her accuracy in
revising the English of the manuscript. This research has
been funded by the Italian Ministry for University.
References
Adams, H.R., Boyd, E.M., Wilson, J.B., Miller, A., Huisman, T.H., 1968.
The structure of goat hemoglobins. 3. Hemoglobin D, a beta chain
variant with one apparent amino acid substitution 21 AspYHis. Arch.
Biochem. Biophys. 127, 398–405.
Agar, N.S., Evans, J.V., Roberts, J., 1972. Red blood cell potassium and
haemoglobin polymorphism in sheep. A review. Anim. Breed. Abstr.
40, 407–436.
Andresen, E., Broad, T.E., Brown, S., Cooper, D.W., Di Stasio, L., Dolling,
C.H.S., Fleet, M., Hill, D.F., Lauvergne, J.J., Lundie, R.S., Maddox, J.,
Nicholas, F.W., Rae, A.L., Renieri, C., Sponenberg, D.P., Tucker, E.M.,
1995. Revised guideline for gene nomenclature in ruminants, 1993.
Genet. Sel. Evol. 27, 1 c.
Bannister, J.V., Bannister, W.H., Wilson, J.B., Lam, H., Miller, A.,
Huisman, T.H., 1979. The structure of goat hemoglobins. V. A fourth
beta chain variant beta-D-Malta; 69 Asp is replaced by Gly with
decreased oxygen affinity and occurring at a high frequency in Malta.
Hemoglobin 3, 57–75.
Black, S.D., Mould, D.R., 1991. Development of hydrophobicity param-
eters to analyze proteins which bear post or cotranslational modifica-
tions. Anal. Biochem. 193, 72–82.
Braend, M., Tucker, E.M., Clarke, S.W., 1987a. Search for genetic variation
in the blood of Norwegian dairy goats reveals a new polymorphism at
the Hb beta A locus. Anim. Genet. 18, 75–79.
Braend, M., Nesse, L.L., Efremov, G.D., 1987b. Expression and genetics of
caprine haemoglobins. Anim. Genet. 18, 223–231.
Canatan, T., Boztepe, S., 2000. The polymorphisms of hemoglobin and
transferrin in Turkish hair goat. Indian Vet. J. 77, 966–968.
Di Luccia, A., Iannibelli, L., Addato, E., Masala, B., Manca, L., Ferrara, L.,
1991. Evidence for the presence of two different beta-globin chains in
the hemoglobin of the river buffalo (Bubalus bubalis L.). Comp.
Biochem. Physiol., B 99, 887–892.
Evans, A.G., Wellems, T.E., 2002. Coevoultionary genetics of Plasmodium
malaria parasites and their human hosts. Int. Comp. Biol. 42, 401–407.
Fesus, L., Varkonyi, J., Ats, A., 1983. Biochemical polymorphisms in goats
with special reference to the Hungarian Native breed. Anim. Blood
Groups Biochem. Genet. 14, 1–6.
Garrick, M.D., Garrick, L.M., 1983. Hemoglobin and globin genes. In:
Agar, N.S., Board, P.G. (Eds.), Red Blood Cells of Domestic Mammals.
Elsevier, Amsterdam, pp. 165–207.
Hardison, R., 1998. Hemoglobins from bacteria to man: evolution of
different patterns of gene expression. J. Exp. Biol. 201, 1099–1117.
Haynes, J.R., Rosteck, P. Jr., Schon, E.A., Gallagher, P.M., Burks, D.J.,
Smith, K., Lingrel, J.B., 1980. The isolation of the beta A-, beta C-, and
gamma-globin genes and a presumptive embryonic globin gene from a
goat DNA recombinant library. J. Biol. Chem. 255, 6355–6367.
Huisman, T.H.J., Schroeder, W.A., 1971. New Aspects of the Structure,
Function and Synthesis of Hemoglobins. CRC Press, Cleveland, Ohio.
Huisman, T.H., Adams, H.R., Dimmock, M.O., Edwards, W.E., Wilson,
J.B., 1967. The structure of goat hemoglobins. I. Structural studies of
the beta chains of the hemoglobins of normal and anemic goats. J. Biol.
Chem. 242, 2534–2541.
Huisman, T.H.J., Brandt, G., Wilson, J.B., 1968. The structure of goat
hemoglobins. II. Structural studies of the alpha chains of the
hemoglobins A and B. J. Biol. Chem. 243, 3675–3686.
Igarashi, M.L., Machado, T.M., Ferro, J.A., Contel, E.P., 2000. Structure
and genetic relationship among Brazilian naturalized and imported goat
breeds. Biochem. Genet. 38, 353–365.
Iorio, M., Vincenti, D., Annunziata, M., Rullo, R., Bonamassa, R., Di
Luccia, A., Pieragostini, E., 2004. Biochemical and molecular inves-
tigations on Hb qualitative and quantitative polymorphism in the river
buffalo (Bubalus bubalis L.) population reared in southern Italy. Genet.
Mol. Biol. 27, 167–173.
John, M.E., Barnabas, J., 1978. Gene diversity of bovid hemoglobins.
Biochem. Genet. 16, 787–798.
Johnson, E.H., Nam, D., Al-Busaidy, R., 2002. Observations on haemo-
globin types in three breeds of Omani goats. Vet. Res. Commun. 26,
353–359.
Kleinschmidt, T., Braunitzer, G., 1982. The primary structure of the
hemoglobin gamma-chains of fetal sheep (Ovis ammon) and goat
(Capra aegagrus). Hoppe-Seyler Z. Physiol. Chem. 363, 789–796.
Lingrel, J.B., Townes, T.M., Shapiro, S.G., Wernke, S.M., Liberator, P.A.,
Menon, A.G., 1985. Structural organization of the alpha and beta globin
loci of the goat. Prog. Clin. Biol. Res. 191, 67–79.
Manca, L., Masala, B., Ledda, S., Naitana, S., 1991. Separation of caprine
globin chains by reversed-phase high-performance liquid chromatog-
raphy: evidence for the presence of a silent beta B-globin allele in
Sardinian sheep. J. Chromatogr. 563, 158–165.
Nagel, R.L., 1990. Innate resistance to malaria: the intraerythrocytic cycle.
Blood Cells 16, 321–339.
Nguyen, T.C., Bunch, T.D., 1980. Blood groups and evolutionary relation-
ships among domestic sheep (Ovis Aries), domestic goat (Capra
hircus), audad (Ammotragus lervia) and European mouflon (Ovis
musimon). Ann. Genet. Sel. Anim. 12, 169–180.
Nyamsamba, D., Nomura, K., Nozawa,, Yokohama, M., Zagdsuren, K.Yo.,
Amano, T., 2003. Genetic relationship among Mongolian native goat
populations estimated by blood protein polymorphism. Small Rumin.
Res. 47, 171–181.
Perutz, M.F., 1990. Frequency of abnormal human haemoglobins caused by
C–T transitions in CpG dinucleotides. Biophys. Chemist. 37, 25–29.
Pieragostini, E., Petazzi, F., 2000. Genetics and tolerance to tick-borne
diseases in South Italy: experience in studying native Apulian and
exotic sheep breed. Parassitologia 41 (Suppl. 1), 89–94.

E. Pieragostini et al. / Comparative Biochemistry and Physiology, Part B 142 (2005) 18–27 27
Pieragostini, E., Dario, C., Bufano, G., 1994. Hemoglobin phenotypes
and hematological factors in Leccese sheep. Small Rumin. Res. 13,
177–185.
Pieragostini, E., Scaloni, A., Rullo, R., Di Luccia, A., 2000. Identical
marker alleles in Podolic cattle (Bos taurus) and zebu (Bos indicus).
Comp. Biochem. Physiol., B 127, 1–9.
Pieragostini, E., Di Luccia, A., Rullo, R., Bottiglieri, C., 2002. Hemoglobin
phenotyes in Murgese horse. Ital. J. Anim. Sci. 1, 159–163.
Pieragostini, E., Petazzi, F., Di Luccia, A., 2003. The relationship between
the presence of extra alpha-globin genes and blood cell traits in
Altamurana sheep. Genet. Sel. Evol. 35 (Suppl. 1), S121–S133.
Pirastru, M., Palici di Suni, M., Vacca, M., Masala, B., Manca, L., 2000.
The hemoglobin polymorphism in sardinian goats: nucleotide sequence
and frequency of hA, hD, hD-Malta, and hE globin genes. In: Di Prisco,
G., Giardina, B., Weber, R. (Eds.), Hemoglobin Function in Vertebrates.
Molecular Adaptations in Extreme and Non-Extreme Environments.
Springer Verlag, Berlin, pp. 97–108.
Pirastru, M., Manca, L., Masala, B., 2003. Characterization of four novel
variants of goat beta(A)-globin gene. Biochem. Genet. 41, 209–217.
Roy, P.H., Weissbach, A., 1975. DNA methylase from HeLa cell nuclei.
Nucleic Acids Res. 2, 1669–1684.
Sartore, G., Facello, C., Bianchi, M., 1981. Ricerche sui polimorfismi
biochimici nella capra: tipi di emoglobina. Ann. Fac. Med. Vet. Torino
28, 3–8.
Scaloni, A., Pieragostini, E., Malori, A., Ferrara, L., Di Luccia, A., 1998.
Bovine hemoglobin alpha-globin chain polymorphism: primary struc-
ture determination of two new genetic variants by mass spectrometry
and amino acid sequencing. Biochimie 80, 333–338.
Schon, E.A., Wernke, S.M., Lingrel, J.B., 1982. Gene conversion of two
functional goat alpha-globin genes preserves only minimal flanking
sequences. J. Biol. Chem. 257, 6825–6835.
Tucker, E.M., 1981. Haemoglobin D in three rare Dutch breeds of sheep.
Anim. Blood Groups Biochem. Genet. 12, 107–112.
Tucker, E.M., Clarke, S.W., Osterhoff, D.R., Groenewald, J., 1983. An
investigation of five genetic loci controlling polymorphic variants in the
red cells of goats. Anim. Blood Groups Biochem. Genet. 14, 269–277.
Tunon, M.J., Gonzalez, P., Vallejo, M., 1987. Blood biochemical poly-
morphism in Spanish goat breeds. Comp. Biochem. Physiol., B 88,
513–517.
Vestri, R., Pieragostini, E., Yang, F., Di Gregorio, P., Rando, A., Masina, P.,
1991. Expression of triplicated and quadruplicated alfa globin genes in
sheep. Br. J. Haematol. 77, 110–116.
Wernke, S.M., Lingrel, J.B., 1986. Nucleotide sequence of the goat
embryonic alpha globin gene zeta and linkage and evolutionary
analysis of the complete alpha globin cluster. J. Mol. Biol. 192,
457–471.
Wilson, J.B., Adams, H.R., Huisman, T.H., 1969. The heterogeneity of the
fetal hemoglobin of the goat. Biochim. Biophys. Acta 181, 367–372.
Wrightstone, R.N., Wilson, J.B., Miller, A., Huisman, T.H., 1970. The
structure of goat hemoglobins. IV. A third beta chain variant beta E with
three apparent amino acid substitutions. Arch. Biochem. Biophys. 138,
451–456.