temporal dynamics of masked word reading
TRANSCRIPT

Consciousness
Consciousness and Cognition 16 (2007) 112–123
andCognition
www.elsevier.com/locate/concog
Temporal dynamics of masked word reading
Scott L. Fairhall *, Jeff P. Hamm, Ian J. Kirk
Research Centre for Cognitive Neuroscience, Department of Psychology, The University of Auckland, Auckland, New Zealand
Received 24 July 2005Available online 20 February 2006
Abstract
The repercussions of unconscious priming on the neural correlates subsequent cognition have been explored previously.However, the neural dynamics during the unconscious processing remains largely uncharted. To assess both the complexityand temporal dynamics of unconscious cognition the present study contrasts the evoked response from classes of maskedstimuli with three different levels of complexity; words, consonant strings, and blanks. The evoked response to maskedword stimuli differed from both consonant strings and blanks, which did not differ from each other. This response wasqualitatively different to any evoked potential observed when stimuli were consciously visible and peaked at 140 ms, earlierthan is usually associated with differences between words and strings and 100 ms earlier than word-consonant string dif-ferences in the visible condition. The evoked response demonstrates a qualitatively distinct signature of unconscious cog-nition and directly demonstrates the extraction of abstract information under subliminal conditions.� 2006 Elsevier Inc. All rights reserved.
Keywords: Priming; Subliminal priming; Unconscious perception; EEG; ERP; Reading; Visual word form area
1. Introduction
Central to many current models of the neural substrates of conscious awareness is the idea that phenom-enological awareness results from the formation of Hebbian reverberant, re-entrant neural assemblies into adynamic core (Crick & Koch, 1998; Dehaene & Naccache, 2001; Tononi & Edelman, 1998). Under such mod-els the backwards masking of a stimulus, known to interrupt the neural activity elicited by the stimulus (Rolls& Tovee, 1994), is believed to prevent the closure of re-entrant connections, sustained activity, and the con-comitant access to the dynamic core (Dehaene & Naccache, 2001; Tononi & Edelman, 1998). The result beingthat the stimulus is not consciously perceived, though paradoxically, an influence on later behaviour, and neu-ral activity has often been documented (Cheesman & Merikle, 1986; Debner & Jacoby, 1994; Dehaene et al.,1998, 2001; Draine & Greenwald, 1998; Greenwald, Draine, & Abrams, 1996).
A recent model has proposed that backwards masking results from the inconsistency between higher andlower order representation of the stimulus resulting in a failure to establish a reentrant representation of themasked stimulus (Enns & Di Lollo, 2000). A similar failure to establish reentrancy may underlie the occasional
1053-8100/$ - see front matter � 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.concog.2006.01.002
* Corresponding author. Fax: +64 9 373 7450.E-mail address: [email protected] (S.L. Fairhall).

S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 113
failure to perceive non-masked stimuli where an absence of late components of the visual response is observed(Super, Spekreijse, & Lamme, 2001).
Backwards masking has been shown to infere with the durations of neural activity evoked in the superiortemporal sulcus (STS) of the macaque. Following presentation of face stimuli, neurons in the STS respond75 ms after the presentation of the stimulus. Activity then persists under normal viewing conditions for some200–300 ms after a 16 ms stimulus presentation. If a masking pattern is introduced in immediate succession ofthe face stimuli, neural firing is extinguished 20–30 ms after its initiation (Rolls & Tovee, 1994). It has beenestimated that �65% of the information present in the firing of a single neuron over the 400 ms period is pres-ent within this 20–30 ms period (Tovee, Rolls, Treves, & Bellis, 1993). It should be noted that these findingsmay not reflect true unconscious processing, as under similar stimulus conditions humans correctly performed50% over and above chance performance (Rolls, Tovee, Purcell, Stewart, & Azzopardi, 1994). However, thesestudies show that masking exerts its influence by suppressing the sustained tail of neural discharge, not theinitial burst.
During backwards masking, although according to Tovee et al. (1993) much of the information remainspresent in the first initial burst of neural activity, the stimulus does not enter conscious awareness. Suggestingperhaps that it is not the amount of information that determines stimuli awareness; rather it is the form ofneural activity. During binocular rivalry, neurons in the visual cortex have been shown to continue to dis-charge even when their output is not part of the conscious percept (Fries, Roelfsema, Engel, Konig, & Singer,1997; Leopold & Logothetis, 1996). Although cells continue to response to the suppressed image, recent fMRIwork has suggested that neural activity in V1 is modulated during rivalry (Lee & Blake, 2002; Polonsky, Blake,Braun, & Heeger, 2000) and further that the spread of BOLD signal is consistent with the spread of the per-ceptual transition during image-dominance transfer (Lee, Blake, & Heeger, 2005). The discord between singlecell and haemodynamic measure of activity leaves the exact nature of V1 activity ambiguous. It has been sug-gested that the increase in bold response may reflect the result of reentrant feedback from extrastriate areasrather than output from this region (Logothetis, 2002; Logothetis, Pauls, Augath, Trinath, & Oeltermann,2001). Single cell recordings have shown that although the rate of firing in V1 does not change, an increasein the gamma range synchrony occurs in neurons receiving input from the dominant eye with the introductionof a rivalrous stimulus in strabismic cats (Fries et al., 1997). Likewise, increased gamma synchrony emergeswith the conscious representation of near threshold tactile stimulation (Meador, Ray, Echauz, Loring, &Vachtsevanos, 2002) and upon the subjective separation of temporally proximate tones (Joliot, Ribary, & Lli-nas, 1994). These findings suggest that it is synchrony of firing, not the rate of firing, that is important in deter-mining what stimuli will be available for conscious processing.
Evidence from the effects of backwards masking in the monkey suggests an interesting possibility. Theneural activity conveying stimulus driven cognition sweeps through a cell assembly, reaching the Hebbianthreshold of ignition, spreading unconscious automatic cognisance and gaining access to all elements ofthe assembly. Then, if the stimulus is not eclipsed by masking stimuli, the network reaches a thresholdfor reverberation and sustained activity persists across the same network, in a style amenable to consciousrepresentation. Under such a sweep and sustain model, complex complete processing of the stimuli occursduring the rapid sweep phase. This is followed by a sustained phase that contains relatively less information,across which the conscious representation of the cognition is contained. Rolls et al. (1994) speculate that thehigh information capacity of the initial burst leaves little need for the encoding of information through syn-chrony. It may be that reverberant activity is not required for its capacity for information processing, but tocreate a sustainable representation. The complex stage of processing has already been performed uncon-sciously. The information poor but long lived stage may allow the representation of the result of this cog-nition through the closure of reentrant loops and possibly the establishment of synchrony. This theoryprovides a working model for distinctions between conscious and unconscious processing in relation tomasked stimuli, explaining both the sustained sensory stimulation required for the representation of cogni-tion in consciousness and the wealth of information associated with unconscious processing that leads tothis representation.
Binocular fusion of two color-inverted stimuli prevents a stimulus from forming a conscious representation.Despite the perceptual absence of the stimuli an extensive, content-specific BOLD response is observed overmany of the regions activated during the visible perception of the stimulus (Moutoussis & Zeki, 2002). The

114 S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123
activation of visual association areas within the temporal cortex had previously been shown to covary with thecontents of consciousness (Tong, 2001). Moutoussis and Zeki (2002) demonstrated that conscious and uncon-scious neural mechanisms cannot be distinguished on the basis of anatomical representation. Rather, theseresearchers showed that it was a diminished of activation within these regions that varied from consciousto unconscious perception. Because of the temporal constraints of fMRI it is not possible to determinewhether this decrease result from an overall decrement in neural activity or to a decrease in the duration ofneural discharge.
This sweep and sustain model predicts that, during masked word priming, activity will initially sweepthrough the network of neurons associated with reading, and then as reverberant assemblies build, will travelthrough consciously in a manner consistent with wave transfer. Such a model predicts an early, punctuatedactivation of a widely distributed neural population during unconscious word processing and a slower multi-stage progression during conscious word processing.
Functional magnetic resonance imaging (fMRI) and electroencephalographic (EEG) imaging of the pro-cesses that accompany unconscious word perception have suggested a diminished and delayed form of theneural activity associated with conscious word perception (Dehaene et al., 2001). These findings are consistentwith a model of unconscious processing where neural activity forms segregated self sustaining local reentrantassemblies successively along the neural pathways associated with reading. If unconscious neural activity pro-ceeds in an isolated but sustained fashion then the unconscious neural events would be occurring in the tailend of this distribution of neural firing observed in the monkey cortex during backward masking (Rolls &Tovee, 1994), after, not before, neural activity has been interrupted by masking. The work of Rolls and Toveefurther suggests that the neural activity underlying unconscious cognition should be more rapid in nature.
Traditional approaches to imaging sub-threshold priming have examined the effect of a masked primethrough an indirect measure, the neural response to a subsequently presented target stimulus. However, neu-roimaging removes the need for this indirect measure as it provides direct insight into the processing occurringduring masked stimulus presentation. It is possible to compare the neural response elicited by two classes ofstimuli and to determine if differing forms of cognition, evidenced by different neural responses, are occurring.A benefit of this method is that tightly controlled comparisons between stimulus classes are possible allowinginferences to be made about the type of cognition contributing to observed neural responses. The greatestadvantage of this method is that, unlike the traditional method, it affords the opportunity to examine theevents occurring during unconscious word processing and not solely the repercussions.
The goal of the present experiment is to investigate the temporal dynamics associated with masked uncon-scious word reading. The sweep hypothesis of unconscious processing postulates that close temporal proxim-ity should exist between the neural events underlying the wide range of processing associated with reading.Further it should have a brief time course and information extraction should occur earlier than is usually asso-ciated with conscious processing of the same stimuli.
To test this hypothesis, a comparison will be made between the evoked response to the absences of stimuliand the masked presentation of semantic free orthographically illegal stimuli (unpronounceable non-words)and words. In an attempt to isolate abstract processes specifically related to reading, words will be comparedto word-like stimuli. Previous techniques have measured the level of processing achieved unconsciouslythrough performance on an indirect measure, such as reaction time or repetition suppression. This study shalldirectly compare two masked stimuli of differing complexities. If a difference is observed in the evokedresponse then this must be due to the processing of the differing complexity.
2. Methods
2.1. Participants
Eighteen right handed subjects (9 female) participated. The mean age was 27, ages ranged from 19to 45 years. All subjects had normal or corrected to normal vision and no history of neurologicalimpairment. All procedures were approved by the University of Auckland Human Subjects EthicsCommittee.

S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 115
2.2. Materials
Three classes of stimuli were utilized: words, orthographically-illegal non-words, and ‘blanks’ (whereno stimulus was presented). Three hundred and twenty word stimuli, 160 nouns, 160 verbs between threeand eight letters in length (mean length 5.53) were presented. The 320 orthographically illegal (unpro-nounceable) non-words were matched for string length (mean 5.54). Stimuli presentation was controlledby a PC compatible computer. Stimuli were presented on a LCD screen. A masking pattern of overlap-ping triangles and squares was presented so that it subtended 3� by 1.5� degrees of visual angle, fullycovering the letter stimuli (2 · 0.5�). Black (10 Lumen) stimuli were presented on a white (135 Lumen)background.
2.3. Procedure
Subjects were presented with a continuous stream of masking patterns interspersed with the experimentalstimuli. Individual masks were presented for 67 ms, experimental stimuli for 27 ms. In the visible condition,the experimental stimuli were flanked by a 67 ms blank. In the masked condition stimuli were flanked bymasks but each flanking mask was preceded or succeeded by a 67 ms blanks (see Fig. 1) as per the methodof Dehaene et al. (2001). Between 11 and 15 masking patterns separated each stimulus presentation, each triallasting between 764 and 1032 ms. The veracity of internal time audits was verified via an external photosen-sitive diode. To ensure the subjects attended to the stimuli they were instructed to press the space key whenthey viewed a series of seven uppercase ‘x’s. A total of 40 such targets were present.
The experiment consisted of 1000 trials, 320 in each experimental condition and 40 target stimuli. Stimulipresentation was ordered randomly. Subjects received a break period every 2 min.
Fig. 1. Experimental paradigm. Masking stimuli present for 67 ms flank the experimental stimuli. Visible stimuli were flanked by a 67 msblank which was the same color and contrast as the background. Shown is the Word condition. The same sequence was presented in thenon-word and blank conditions.
116 S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123
2.4. Behavioural measures
Following the main experiment subjects participated in two forced choice tests and an identification test.Each test consisted of 40 trials. In the first test, subjects indicated whether the word stimuli occurred beforeor after a briefly presented tone. In the second test subjects discriminated between words and illegal non-words. For the third test, subjects reported the presented word. Words were selected randomly from the320 words used in the imaging study.
2.5. Electrophysiological recordings
Recordings were made in an electrically shielded, sound-attenuated room, using a 128 channel ElectricalGeodesic amplifier (200 MX input impedance) with Ag/AgCl electrodes. EEG was recorded continuously witha sampling rate of 500 Hz and referenced to the vertex channel. Impedances were set below 40 kX. Duringrecording the data were band-passed filtered between 0.1 and 100 Hz.
Following data acquisition EEG recordings were averaged across trials, within conditions. Trialepochs consisted of a 250 ms pre-stimulus baseline and a 700 ms post stimulus time period. The resultingevent related potentials (ERPs) were baselined over the 250 ms pre-stimulus interval, re-referenced to thealgebraic average of all the channels (the common average reference) and band-pass filtered (0.1–20 Hz).
Automated artefact rejection algorithms were employed to remove trials contaminated by eye blinks orother artefacts.
3. Results
3.1. Behavioural results
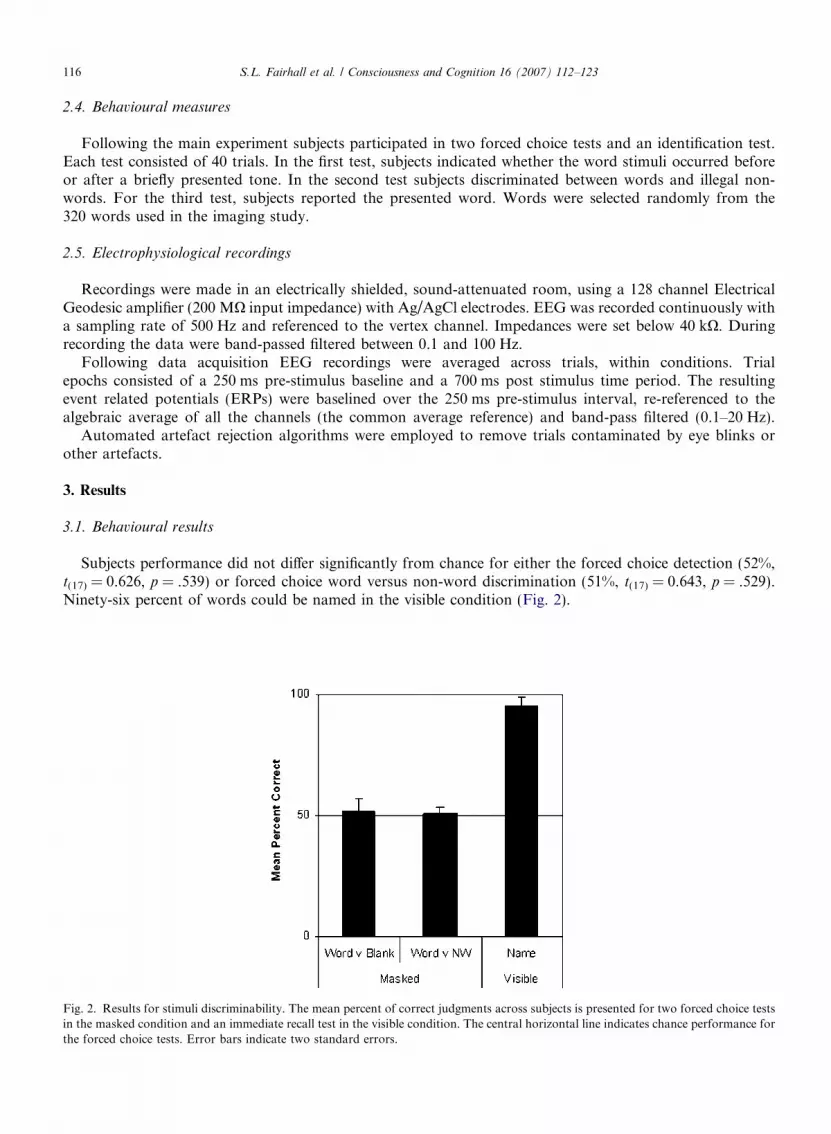
Subjects performance did not differ significantly from chance for either the forced choice detection (52%,t(17) = 0.626, p = .539) or forced choice word versus non-word discrimination (51%, t(17) = 0.643, p = .529).Ninety-six percent of words could be named in the visible condition (Fig. 2).
Fig. 2. Results for stimuli discriminability. The mean percent of correct judgments across subjects is presented for two forced choice testsin the masked condition and an immediate recall test in the visible condition. The central horizontal line indicates chance performance forthe forced choice tests. Error bars indicate two standard errors.

S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 117
3.2. Time window selection
To prevent the inflation of type 1 error associated with large sets of EEG data it is important to define apriori the focus of analysis. A previously employed technique for identifying time windows where evokedresponses to masked words are present is to identify time windows through peaks in differential evokedresponse to visible words (Dehaene et al., 2001). To remove the effects of the masking pattern alone the‘blanks’ condition was subtracted from both the word, and non-word, conditions. The resulting differencewaves were then averaged and the Global Field Power (GFP) calculated. The GFP time series reflected acti-vation predominantly due to the presence of the stimuli (word or non-word) and not masking stimuli. Impor-tantly, this time series contained no information about differences between words and non-words allowing forthe blind selection of time windows with respect to word/non-word differences. Further, it contained no infor-mation of differences within the masked condition, allowing blind time window selection for comparisons ofall three levels of stimuli. This process will contain information about the difference between visible blanks andletter string stimuli but allows the blind selection of time windows with respect to the remaining contrasts.Four time windows with distinct topographical distributions were observed. Time windows were extended20 ms before and after the corresponding peaks. These were: 122–162, 226–266, 306–346, and 422–462 ms.These time windows and their respective topographic distributions were similar, though not identical, to thoseobserved in Dehaene et al. (2001).
3.3. Analysis of time windows
To test for significant difference over the whole scalp the mean voltage for each electrode was calculated ineach time window a repeated measures ANOVA was performed with the within subject factors of conditionand electrode. This analysis was repeated for all four time windows. Alpha was adjusted to compensate for thefour comparisons (acorrected = 0.0125). To compensate for the lack of independence between the electrodes theGreenhouse–Geiser adjusted degrees of freedom were used. A significant condition by electrode interactionindicates a different pattern of activations across conditions. Such an analysis is sensitive to differences in bothtopography and amplitude.
3.4. Masked condition
Analysis revealed a significant condition by electrode interaction in the first time window (F(9.7,14.7) = 2.390,p = .012). No significant difference was found in the three later time windows.
The significant effect of condition occurring in the first time window appears to coincide with a positiveoccipital activation typical of the p100 visual evoked potential. The presence of this p100-type componentin the blank condition suggests that this component was the result of changes in the series of masking patternsnot the experimental stimuli.
To investigate which conditions were contributing to the interaction a, further post hoc pair wiseANOVA was conducted comparing each combination of conditions. The introduction of non-words intothe mask stream did not produce a discernable change in the evoked potential (F(7,118.9) = 1.076,p = .383). However, the inclusion of words in the masked stream produced an evoked response signifi-cantly different to that produced by either non-words (F(7.2,122.2) = 3.743, p = .001) or blanks(F(7.2,123) = 2.517, p = .018).
Words elicited a greater parietal negativity and bilateral fronto-temporal positivity than did non-words. Toassess whether this reflected a change in topography, and the recruitment of different neural populations, rath-er than a change merely in amplitude, individual subject data was normalized to a GFP of 1 and the dissim-ilarity in potential maps between conditions was compared to a distribution generated through 5000permutation randomisation test (Strik, Fallgatter, Brandeis, & Pascual-Marqui, 1998). This yielded a signif-icantly different topography between word and non-word conditions (p = .001) (Fig. 3).
The differences between words and blanks were smaller than between words and non-words. Comparisonsof the topographical distribution using the above method revealed that the words and blanks produce differingscalp distributions (p = .03). The parietal negativity was less wide-spread and stronger over the right hemi-
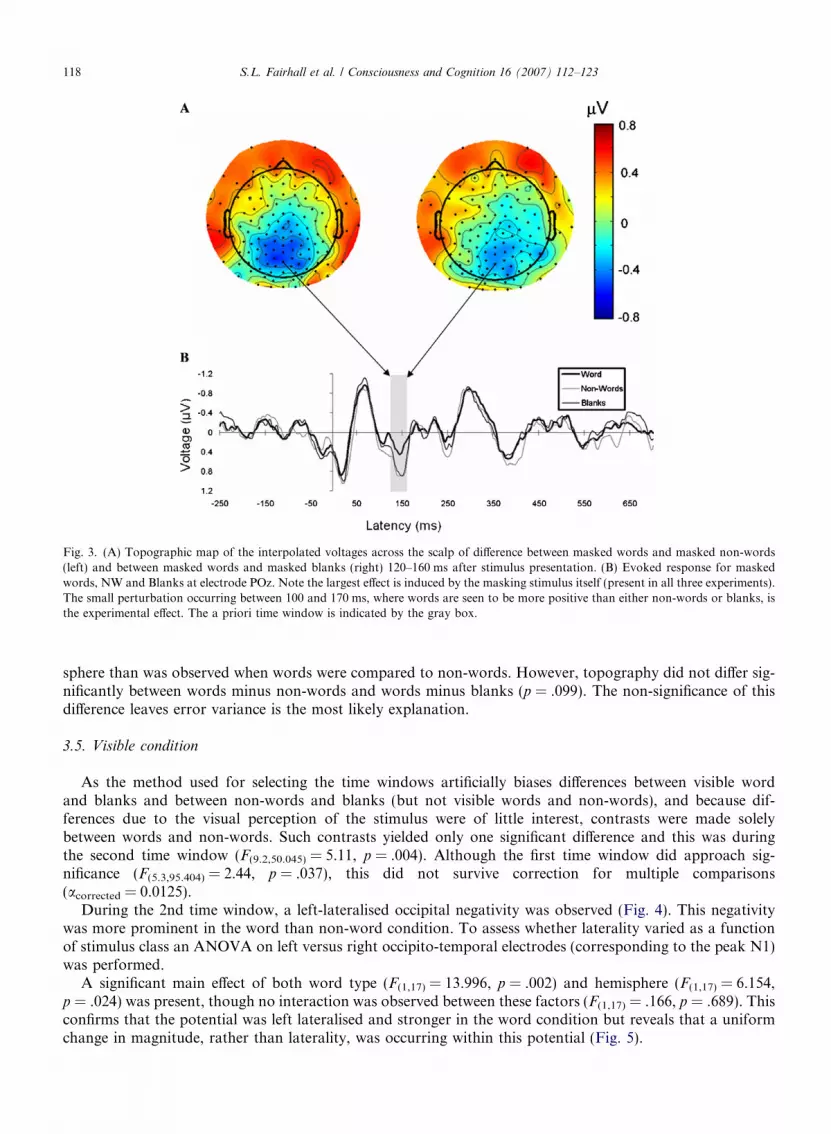
Fig. 3. (A) Topographic map of the interpolated voltages across the scalp of difference between masked words and masked non-words(left) and between masked words and masked blanks (right) 120–160 ms after stimulus presentation. (B) Evoked response for maskedwords, NW and Blanks at electrode POz. Note the largest effect is induced by the masking stimulus itself (present in all three experiments).The small perturbation occurring between 100 and 170 ms, where words are seen to be more positive than either non-words or blanks, isthe experimental effect. The a priori time window is indicated by the gray box.
118 S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123
sphere than was observed when words were compared to non-words. However, topography did not differ sig-nificantly between words minus non-words and words minus blanks (p = .099). The non-significance of thisdifference leaves error variance is the most likely explanation.
3.5. Visible condition
As the method used for selecting the time windows artificially biases differences between visible wordand blanks and between non-words and blanks (but not visible words and non-words), and because dif-ferences due to the visual perception of the stimulus were of little interest, contrasts were made solelybetween words and non-words. Such contrasts yielded only one significant difference and this was duringthe second time window (F(9.2,50.045) = 5.11, p = .004). Although the first time window did approach sig-nificance (F(5.3,95.404) = 2.44, p = .037), this did not survive correction for multiple comparisons(acorrected = 0.0125).
During the 2nd time window, a left-lateralised occipital negativity was observed (Fig. 4). This negativitywas more prominent in the word than non-word condition. To assess whether laterality varied as a functionof stimulus class an ANOVA on left versus right occipito-temporal electrodes (corresponding to the peak N1)was performed.
A significant main effect of both word type (F(1,17) = 13.996, p = .002) and hemisphere (F(1,17) = 6.154,p = .024) was present, though no interaction was observed between these factors (F(1,17) = .166, p = .689). Thisconfirms that the potential was left lateralised and stronger in the word condition but reveals that a uniformchange in magnitude, rather than laterality, was occurring within this potential (Fig. 5).
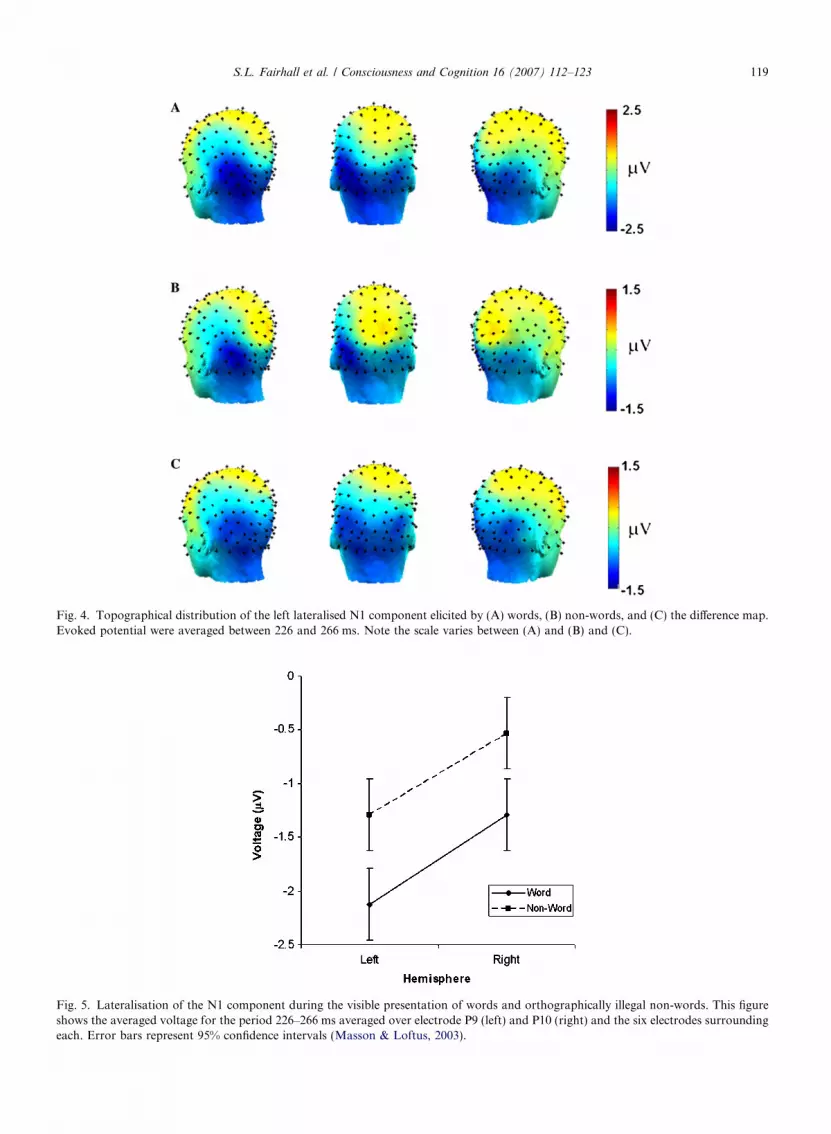
Fig. 4. Topographical distribution of the left lateralised N1 component elicited by (A) words, (B) non-words, and (C) the difference map.Evoked potential were averaged between 226 and 266 ms. Note the scale varies between (A) and (B) and (C).
Fig. 5. Lateralisation of the N1 component during the visible presentation of words and orthographically illegal non-words. This figureshows the averaged voltage for the period 226–266 ms averaged over electrode P9 (left) and P10 (right) and the six electrodes surroundingeach. Error bars represent 95% confidence intervals (Masson & Loftus, 2003).
S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 119

120 S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123
4. Discussion
The behavioural studies show that subjects were unaware of the stimuli in the masked condition and thatstimuli were easily observable in the visible condition. The masked condition can therefore be consideredunconsciously processed as assessed by the objective threshold criterion (Merikle, Smilek, & Eastwood, 2001).
Dehaene and colleagues (2001) demonstrated that perturbations could be detected in the ERP evoked byunconsciously perceived stimuli. Further, they showed in a supplementary experiment that, under such presen-tation conditions, abstract information pertaining to the characteristics of the letter characters was extracted.Left ambiguous was whether the activation observed in response to masked words was a reflection of suchabstract information processing, or merely due to the presence of the naked stimulus.
In this study, the differences observed between words and both non-words and blanks demonstrate that therecorded response was not due to physical characteristics of the stimulus. The significantly different evokedresponse between words and non-words provides direct evidence of abstract information extraction abovethe level of letter character information, demonstrating that during unconscious word reading words are pro-cessed to at least pre-lexical level in the absence of conscious awareness.
The evoked response observed in this study provides a qualitative distinction between conscious and uncon-scious conditions. A classic criticism of unconscious perception paradigms is that the direct measure of aware-ness may not be sensitive to all aspects of conscious processing (Holender, 1986). Undetected consciouscognition may seep into trials in the unconscious condition and produce an increase in performance that iserroneously attributed to non-existent unconscious processing. The observation of a temporally and topo-graphically distinct pattern of activation in the masked condition amounts to a qualitative distinction. Suchqualitative distinctions offer rare evidence of dissociable unconscious processes. Such dissociations have beenfound in the process dissociation paradigm where explicit instruction is placed in opposition to automatic pro-cessing (Cheesman & Merikle, 1986; Debner & Jacoby, 1994) and have previously been observed via modifi-cations of the time course of the N400 evoked by masked and visibly primed target stimuli (Kiefer & Spitzer,2000).
The transient pattern of the evoked response is consistent with the transient pattern of the behaviouralfacilitation. Priming that requires extraction of the symbolic meaning of the word is maximal 60 ms after stim-ulus presentation and disappears after 100 ms (Greenwald et al., 1996). The single fleeting burst of activationrelated to the abstract nature of the word is consistent with the evanescent nature of masked semantic priming.
The brief, punctuated, activation observed in the masked conditions of the present study, supports thehypothesised model of unconscious processing, where neural activity is transmitted rapidly across a neuralnetwork in the absence of reverberant activity. However, the failure to find evidence of prolonged activity doesnot preclude its occurrence. In a similar study, masked words were found to produce evoked responses in threetime windows spanning a period from 150 to 500 ms after stimulus presentation (Dehaene et al., 2001). Theseresults were not, however, replicated in the present study suggesting that further research will be required toresolve this issue.
The posited model requires that all neural activity, visual, pre-lexical, and semantic, is occurring within theevoked potential recorded in this study. In a single subject (12 recording sessions) MEG study, differences inthe evoked response were found over parieto-occipital sites to words of differing semantic strengths beginning100 ms post stimulus (Pulvermuller, Assadollahi, & Elbert, 2001). This response occurred to highly practisedwords (16 stimuli each presented 240 times) which may have attenuated conscious processing. The authorsspeculate that, due to frequent repetition, the relevant memory traces may have been more active, facilitatingthe response to semantic word properties. The similarity of the distribution and time course (both peaked at140) of the activation of both this study and that of Pulvermuller and colleagues (2001) allows the possibilitythat the differences observed during this study may reflect the differences between the semantic content in theunconsciously processed non-words and words.
The non-lateralised topographical distribution of the difference in the evoked response is consistent with aprevious study that has shown the absence of a visual field advantage during unconscious word processing
In the current study, the failure of masked orthographically illegal non-words to evoke a discernable neuralresponse may reflect the absence of a pre-existing neural assembly responsive to these novel stimuli. Thehypothesised model of the transmission of neural activity related to unconscious cognition relies on the rapid

S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 121
transmission of neural signals across existing pathways. If no such assembly exists then the model predicts thatthe stimuli will not produce a discernable evoked response.
The left lateralised negative occipito-temporal component of the current study is identical to that observedduring similar stimuli presentation conditions (Dehaene et al., 2001). This waveform has the distinctive topog-raphy of the late N1, thought to reflect activity of the VWFA (visual word form area Cohen et al., 2000) anddemonstrated a delayed onset consistent with other masking studies (Dehaene et al., 2001). During the visiblepresentation of words, differences between orthographic and non-orthographic stimuli have not been observedprior to the N1 (Simon, Bernard, Largy, Lalonde, & Rebai, 2004). If the VWFA is the site of the pre-lexicalprocessing then there is a disparity between the difference observed between words and non-words at 140 ms inthe masked condition and the difference observed at 240 ms in the visible condition. The earlier activation inthe masked condition suggests that visual word form has already been extracted 100ms prior to the potentialusually attributed the initial extraction of the word form. It is possible that this disparity is a reflection of thedifferent time courses associated with an initial burst of high information firing.
The amplitude of the late N1 component related to visual word form showed a monotonic increase acrosshemispheres from letter strings to words. This suggests that activation of the putative VWFA may reflect bilat-eral generators and trans-hemispheric cell assemblies. The possibility of the presence of bilateral generators issupported by the finding of an increase in the BOLD response to words over non-words not only in the leftVWFA of all subjects, but in the right fusiform area, observed in six out of seven subjects (Dehaene, LeClec’H, Poline, Le Bihan, & Cohen, 2002).
The difference between words and consonant strings observed for masked stimuli was not found for visiblestimuli. This may have been due to the increase in neural activity associated with the visible condition. Thiswould increase intra-subject variability in the evoked response (the error term in the statistical test used) andthe probability of a neural generator which cancels out the masked condition potential before it travels to thescalp.
An alternate explanation has further reaching theoretical implications. There exists the possibility that themasked pattern of activation was not observed in the visible condition because as the stimuli enters conscious-ness the normal route of unconscious processing is reduced or removed. This supports a model where uncon-scious processing is actively suppressed during conscious processing leaving the elements of unconsciousprocessing more active in the absence of awareness of the stimuli.
Although this remains speculative, such a finding would have repercussions on existing theories of con-scious and unconscious processing where unconscious processing is conceptualized as a constant that canbe linearly added to the level of conscious processing, allowing the estimation of the degree of unconsciousprocessing (Debner & Jacoby, 1994). However, if unconscious processing varies with stimulus perceptibilitythen such theories become untenable.
The evoked potential reported in this study was not reported by Dehaene and colleagues (2001) despite analmost identical experimental paradigm and examination of the same time window. This disparity may havearisen due to the type of analysis used. Dehaene and colleagues focussed on a pair of occipital electrode (O1and O2) where no significant difference was observed in this study. Another possibility is that the repetition ofstimuli used during the study of Dehaene and colleagues may have attenuated the neural response throughrepetition suppression (Naccache & Dehaene, 2001)
In summary, this study provides evidence for qualitatively different neural response during unconscious,relative to conscious, word processing. This response proceeded with a time course more rapid than that asso-ciated with visual word form processing during conscious cognition. The brief duration of observed activationmay reflect a single rapid processing period underlying unconscious processing, although further unconsciouscognition may be below the resolution of EEG techniques. The comparison of the evoked response to wordsand to letter strings demonstrates that abstract information has been extracted despite subjects being unable toreport the difference between words and non-words. The time course and distribution of this response is con-sistent with a component previously observed to be modulated by semantic processing (Pulvermuller et al.,2001). The earlier processing of information without awareness is compatible with an initial sweep of activa-tions through neural circuits which is distinguished from conscious processing by its failure to develop reen-trant stability, although it still remains to be shown that the recorded evoked response was the sum of allneural processing occurring in response to the masked stimuli.

122 S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123
Acknowledgments
This research was supported by the University of Auckland Emerging Research Excellence Award (23137),awarded to Jeff Hamm.
References
Cheesman, J., & Merikle, P. M. (1986). Distinguishing conscious from unconscious perceptual processes. Canadian Journal of Psychology-
Revue Canadienne De Psychologie, 40(4), 343–367.Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-Lambertz, G., Henaff, M. A., et al. (2000). The visual word form area:
Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain, 123(2),291–307.
Crick, F., & Koch, C. (1998). Consciousness and neuroscience. Cerebral Cortex, 8(2), 97–107.Debner, J. A., & Jacoby, L. L. (1994). Unconscious perception: Attention, awareness, and control. Journal of Experimental Psychology.
Learning, Memory, and Cognition, 20(2), 304–317.Dehaene, S., Le Clec’H, G., Poline, J. B., Le Bihan, D., & Cohen, L. (2002). The visual word form area: a prelexical representation of
visual words in the fusiform gyrus. Neuroreport, 13(3), 321–325.Dehaene, S., & Naccache, L. (2001). Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework.
Cognition, 79(1–2), 1–37.Dehaene, S., Naccache, L., Cohen, L., Le Bihan, D., Mangin, J. F., Poline, J. B., et al. (2001). Cerebral mechanisms of word masking and
unconscious repetition priming. Nature Neuroscience, 4(7), 752–758.Dehaene, S., Naccache, L., Le Clec’H, G., Koechlin, E., Mueller, M., Dehaene-Lambertz, G., et al. (1998). Imaging unconscious semantic
priming. Nature, 395(6702), 597–600.Draine, S. C., & Greenwald, A. G. (1998). Replicable unconscious semantic priming. Journal of Experimental Psychology-General, 127(3),
286–303.Enns, J. T., & Di Lollo, V. (2000). What’s new in visual masking? Trends in Cognitive Sciences, 4(9), 345–352.Fries, P., Roelfsema, P. R., Engel, A. K., Konig, P., & Singer, W. (1997). Synchronization of oscillatory responses in visual cortex
correlates with perception in interocular rivalry. Proceedings of the National Academy of Sciences of the United States of America,
94(23), 12699–12704.Greenwald, A. G., Draine, S. C., & Abrams, R. L. (1996). Three cognitive markers of unconscious semantic activation. Science, 273(5282),
1699–1702.Holender, D. (1986). Semantic activation without conscious identification in dichotic-listening, parafoveal vision, and visual masking - a
Survey and Appraisal. Behavioral and Brain Sciences, 9(1), 1–23.Joliot, M., Ribary, U., & Llinas, R. (1994). Human oscillatory brain activity near 40 Hz coexists with cognitive temporal binding.
Proceedings of the National Academy of Sciences of the United States of America, 91(24), 11748–11751.Kiefer, M., & Spitzer, M. (2000). Time course of conscious and unconscious semantic brain activation. Neuroreport, 11(11), 2401–2407.Lee, S. H., & Blake, R. (2002). V1 activity is reduced during binocular rivalry. Journal of Vision, 2(9), 618–626.Lee, S. H., Blake, R., & Heeger, D. J. (2005). Traveling waves of activity in primary visual cortex during binocular rivalry. Nature
Neuroscience, 8(1), 22–23.Leopold, D. A., & Logothetis, N. K. (1996). Activity changes in early visual cortex reflect monkeys’ percepts during binocular rivalry.
Nature, 379(6565), 549–553.Logothetis, N. K. (2002). The neural basis of the blood-oxygen-level-dependent functional magnetic resonance imaging signal.
Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 357(1424), 1003–1037.Logothetis, N. K., Pauls, J., Augath, M., Trinath, T., & Oeltermann, A. (2001). Neurophysiological investigation of the basis of the fMRI
signal. Nature, 412(6843), 150–157.Masson, M. E. J., & Loftus, G. R. (2003). Using confidence intervals for graphically based data interpretation. Canadian Journal of
Experimental Psychology-Revue Canadienne De Psychologie Experimentale, 57(3), 203–220.Meador, K. J., Ray, P. G., Echauz, J. R., Loring, D. W., & Vachtsevanos, G. J. (2002). Gamma coherence and conscious perception.
Neurology, 59(6), 847–854.Merikle, P. M., Smilek, D., & Eastwood, J. D. (2001). Perception without awareness: Perspectives from cognitive psychology. Cognition,
79(1–2), 115–134.Moutoussis, K., & Zeki, S. (2002). The relationship between cortical activation and perception investigated with invisible stimuli.
Proceedings of the National Academy of Sciences of the United States of America, 99(14), 9527–9532.Naccache, L., & Dehaene, S. (2001). The priming method: Imaging unconscious repetition priming reveals an abstract representation of
number in the parietal lobes. Cerebral Cortex, 11(10), 966–974.Polonsky, A., Blake, R., Braun, T., & Heeger, D. J. (2000). Neuronal activity in human primary visual cortex correlates with perception
during binocular rivalry. Nature Neuroscience, 3(11), 1153–1159.Pulvermuller, F., Assadollahi, R., & Elbert, T. (2001). Neuromagnetic evidence for early semantic access in word recognition. European
Journal of Neuroscience, 13(1), 201–205.Rolls, E. T., & Tovee, M. J. (1994). Processing speed in the cerebral-cortex and the neurophysiology of visual masking. Proceedings of the
Royal Society of London Series B-Biological Sciences, 257(1348), 9–15.

S.L. Fairhall et al. / Consciousness and Cognition 16 (2007) 112–123 123
Rolls, E. T., Tovee, M. J., Purcell, D. G., Stewart, A. L., & Azzopardi, P. (1994). The responses of neurons in the temporal cortex ofprimates, and face identification and detection. Experimental Brain Research, 101(3), 473–484.
Simon, G. G., Bernard, C., Largy, P., Lalonde, R., & Rebai, M. (2004). Chronometry of visual word recognition during passive and lexicaldecision tasks: An ERP investigation. International Journal of Neuroscience, 114(11), 1401–1432.
Strik, W. K., Fallgatter, A. J., Brandeis, D., & Pascual-Marqui, R. D. (1998). Three-dimensional tomography of event-related potentialsduring response inhibition: evidence for phasic frontal lobe activation. Electroencephalography and Clinical Neurophysiology, 108(4),406–413.
Super, H., Spekreijse, H., & Lamme, V. A. (2001). Two distinct modes of sensory processing observed in monkey primary visual cortex(V1). Nature Neuroscience, 4(3), 304–310.
Tong, F. (2001). Competing theories of binocular rivalry: A possible resolution. Brain and Mind, 2(1), 55–83.Tononi, G., & Edelman, G. M. (1998). Neuroscience—consciousness and complexity. Science, 282(5395), 1846–1851.Tovee, M. J., Rolls, E. T., Treves, A., & Bellis, R. P. (1993). Information encoding and the responses of single neurons in the primate
temporal visual-cortex. Journal of Neurophysiology, 70(2), 640–654.