switching the polarity of a bacteriophage integration system
TRANSCRIPT

Molecular Microbiology (2004)
51
(6), 1719–1728 doi:10.1111/j.1365-2958.2003.03942.x
© 2004 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004
? 2004
51
617191728
Original Article
Polarity of phage integrationM. C. A. Smith, R. Till and M. C. M. Smith
Accepted 17 November, 2003. *For correspondence. E [email protected]; Tel. (+1) 01224 555739; Fax (+1) 01224555844.
†
Present address: Institute of Medical Sciences, Universityof Aberdeen, Foresterhill, Aberdeen AB25 2ZD UK.
Switching the polarity of a bacteriophage integration system
Matthew C. A. Smith, Rob Till and Margaret C. M. Smith
†
*
Institute of Genetics, University of Nottingham, Queens Medical Centre, Nottingham NG7 2UH, UK.
Summary
During lysogenic growth many temperate bacte-riophage genomes are integrated into the host’schromosome and efficient integration and excisionare therefore an essential part of the phage lifecycle. The
Streptomyces
phage
ffff
C31 encodes anintegrase related to the resolvase/invertases and isevolutionarily and mechanistically distinct from theintegrase of phage
llll
. We show that during
ffff
C31integration the polarity of the recombination sites,
attB
and
attP
, is dependent on the sequences ofthe two base pairs (bp) where crossover occurs. Aloss or switch in polarity of the recombination sitescan occur by mutation of this dinucleotide, leadingto incorrectly joined products. The properties of themutant sites implies that
ffff
C31 integrase interactssymmetrically with the substrates, which duringsynapsis can align apparently freely in either oftwo alternative forms that lead to correct or incor-rect joining of products. Analysis of the topologiesof the reaction products provided evidence thatintegrase can synapse and activate strandexchange even when recombinant products cannotform due to mismatches at the crossover site. Thetopologies of the recombination products are com-plex and indicative of multiple pathways to productformation. The efficiency of integration of a
ffff
C31derivative, KC859, into an
attB
site with switchedpolarity was assayed
in vivo
and shown to be nodifferent from integration into a wild-type
attB
. Thusneither the host nor KC859 express a factor thatinfluences the alignment of the recombination sitesat synapsis.
Introduction
Site-specific recombination systems are frequentlyresponsible for movement of genetic elements from oneDNA molecule to another. In temperate bacteriophagesthe recombination system is tightly regulated to ensureintegration and excision at the appropriate stages in thelife cycle. Moreover the mechanism of integration mustensure that the excision pathway, once it is activated,occurs efficiently. The preservation of the polarity of therecombination sites is a crucial factor in phage-encodedand many other site-specific recombination system (Nash,1996). This means that the left side of one substrate sitebecomes joined to the right side of the other substrate siteand
vice versa
. Incorrect joining, i.e. left side to left sideor right to right may result in inactive recombination sitesand, in some elements, deleterious DNA rearrangements.
In bacteria the serine recombinase family of site-specific recombinases includes the resolvases, inver-tases, phage integrases and transposases (Smith andThorpe, 2002). The best-studied serine recombinases arethe resolvase/invertases, which contain an N-terminal cat-alytic domain and a small C-terminal domain (Grindley,2002; Johnson, 2002). Others such as the phage inte-grases contain the resolvase/invertase-like N-terminalcatalytic domains but as their C-terminal regions are muchlarger and very diverse, they have been called the largeserine recombinases (Smith and Thorpe, 2002). Somegenetic elements that encode clinically important antibi-otic resistance determinants are dependent on membersof the large serine recombinases for movement. Theseinclude Tn
4451
and Tn
5397
in
Clostridia
encodingchloramphenicol or tetracycline resistance, respectively,and the SCC
mec
, a novel genetic element from strains of
Staphylococcus aureus
containing methicillin resistance(Crellin and Rood, 1997; Katayama
et al
., 2000; Wangand Mullany, 2000; Smith and Thorpe, 2002). Little isknown about the mechanism and control of the largeserine recombinases.
The integrase encoded by the
Streptomyces
phage
f
C31 is a large serine recombinase. Mutation of the pro-posed catalytic serine residue in
f
C31 integrase abolishesrecombination function suggesting that this protein catal-yses recombination using a similar mechanism to
gd
andTn
3
resolvases (Thorpe and Smith, 1998). The first step

1720
M. C. A. Smith, R. Till and M. C. M. Smith
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
51
, 1719–1728
in
gd
and Tn
3
resolution is the formation of a productivesynapse in which resolvase binds to sequences in twoidentical
res
sites, bringing the two DNA strands contain-ing the crossover sites together in a specific complex(Bednarz
et al
., 1990; Sarkis
et al
., 2001; Grindley, 2002).A considerable body of evidence suggests that strandexchange occurs via the double-strand cut rotation model(Grindley, 2002). Staggered breaks are generated at the3
¢
ends of a 2-bp sequence and transient phospho-serinelinkages are formed between the active site serine in eachof four resolvase monomers to the four recessed 5
¢
ends.The DNA strands are positioned into the recombinantconfiguration and the DNA backbone is then ligated toform the products. The invertases function by a similarmechanism (Johnson, 2002). The details of howresolvases/invertases manipulate the DNA strands duringrecombination are not clear but measurements of thechanges in DNA topology have provided key insights. Thetopology studies indicate that a 180
∞
right-handed rotationof one pair of half sites about the axis of alignment occurs(Stark
et al
., 1989; Heichman
et al
., 1991;Grindley, 2002).Iteration of this rotation has been observed through mul-tiples of 180
∞
to form complex knots and catenanes (Kras-now and Cozzarelli, 1983; Wasserman
et al
., 1985;Kanaar
et al
., 1990; Heichman
et al
., 1991; Stark
et al
.,1991).
Unlike resolvase the pairs of sites that take part inintegration of
f
C31 are different (Kuhstoss and Rao,1991). As in
l
, integration occurs between an
attP
and
attB
site, recombining to form
attL
and
attR
. However, in
f
C31 both
attP
and
attB
are small, the minimal sites being39 and 34 bp for
attP
and
attB,
respectively, and recom-bination occurs at the identical core sequence, 5
¢
TT(Fig. 1) (Kuhstoss and Rao, 1991; Groth
et al
., 2000;Combes
et al
., 2002). Integrative recombination by
f
C31integrase occurs
in vitro
but not excision, i.e.
attL
and
attR
recombination. Indeed recombination
in vitro
between anyother pair of
att
sites other than
attP
and
attB
has neverbeen observed (Thorpe and Smith, 1998; Thorpe
et al
.,2000). Thus integrase on its own is directional and it isproposed that an accessory factor is required for phageexcision. It is the combination of the irreversible nature ofthe integrative reaction
in vitro
and the small recombina-tion sites that has attracted many researchers to investi-gate the potential of
f
C31 integrase in genomemanipulation and in gene therapy (Olivares
et al
., 2001;2002; Andreas
et al
., 2002; Ortiz-Urda
et al
., 2002; Stoll
et al
., 2002; Belteki
et al
., 2003).Our aim was to determine how the polarity of recombi-
nation sites is maintained during phage integration by alarge serine recombinase, such as
f
C31 integrase. Weshow that during
f
C31 integration the polarity of therecombination sites,
attB
and
attP
, is wholly dependenton the sequences of the two base pairs (bp) where cross-
over occurs. A loss or switch in polarity of the recombina-tion sites can occur by mutation of this dinucleotide,leading to incorrectly joined products.
Results
Switching the polarity of integration
in vitro
In order to investigate the role of the core sequence indetermining the polarity of the attachment sites, singlebase pair changes were introduced into
attB
and
attP
togenerate palindromic core sequences, i.e.
attP
TA
and
att-B
TA
(Fig. 1A).
In vitro
recombination assays were per-formed using two supercoiled plasmids as substrates, oneencoding
attB
or
attB
TA
and the other
attP
or
attP
TA
. Theproducts of recombination are co-integrates or productcircles (Fig. 1). The reactions were analysed by restrictionto detect the recombined molecules and by nicking withDNaseI, which provides information on the topology of theproducts (Fig. 2). Restriction analysis of the recombina-tion products showed that the majority of the wild-type
attB
and
attP
sites could be converted to
attL
and
attR
after 30 min (Fig. 2A). The product of this reaction ispRT602700 (Fig. 1B) and this plasmid can be propagatedin
E. coli
and used as a marker for recombinant products(Fig. 2, PC). When mismatched core sequences wereused, i.e.
attB
TA
¥
attP
(Fig. 2A) or with
attB
¥
attP
TA
(notshown), no
attL
or
attR
containing fragments wereobserved. However when the
attP
TA
¥
attB
TA
substrateswere used similar overall levels of recombinant productswere made compared to the reactions with the wild-typesites but only about half of the products were correctlyjoined to form
attR
and
attL
. The remaining half wasincorrectly joined with the left side of
attP
joining to theleft side to
attB
(L
P
-L
B
joining) and the reciprocal, right sideto right side joint (R
B
-R
P
joining) (Figs 1 and 2A). Theproducts of the incorrect joining were confirmed by DNAsequencing. Thus the sites with matched core sequences,
attB
TA
¥
attP
TA had lost polarity completely.To investigate the polarity further we constructed two
more mutant sites, attBAA and attPAA both containing 5¢AAat the core sequence (Fig. 1A). All the products betweenattP ¥ attBAA or attPAA ¥attB were incorrectly joined indi-cating that the polarities of the attBAA or attPAA sites hadswitched compared to their wild-type counterparts(Fig. 2C). The sequences of the incorrectly joined productsfrom the attP ¥ attBAA reaction are shown in Fig. 1C. Onlycorrectly joined products were obtained with the attBAA
¥attPAA, i.e. when the polarities of both sites were switched(Fig. 2C). Thus the 2 bp core sequence where crossoveroccurs determines the polarities of the attB and attP sitesand correct formation of the products, attL and attR.
This switching of polarity of the sites has implicationsfor synapsis, which is the initial bringing together of the

Polarity of phage integration 1721
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
attB and attP sites through their interactions with inte-grase. In order to generate the correct left-to-right joinedproducts, as observed with the wild-type sites, we canregard the alignment of the att sites at synapsis as ‘par-allel’ (shown schematically in Fig. 3A). On the other hand,left-to-left and right-to-right products such as those gen-erated by recombination between attP ¥ attBAA and attPAA
¥attB must occur via an alternative alignment of sub-strates, which we term ‘antiparallel’ (Fig. 3A). The terms‘parallel’ and ‘antiparallel’ have been used previously to
describe alternative versions of the synapse (Bednarzet al., 1990; Moskowitz et al., 1991). It should be notedthat we do not use these terms to infer the actual pathsof the DNA within the synaptic complex. As the sites withthe symmetrical core sequences can form both the correctand incorrectly joined products with equal frequency, theyappear to be able to form both the ‘parallel’ and the ‘anti-parallel’ synaptic complexes with apparently no prefer-ence between them. We therefore propose that integraseis free to bring recombination sites in either the ‘parallel’
Fig. 1. fC31 attachment sites, outline of the in vitro recombination assay and the sequences of the products.A. The sequences of the attP, attB and the mutant sites used in this study. A dot indicates the sequence is identical to the sequence on the line above. The boxed bases are the identical core sequences where crossover occurs.B. Inter-plasmid recombination and detection of the products by restriction analysis. A schematic is shown of the reaction between substrates containing attP (black solid arrow) and those containing attB (dotted black arrow) producing the product circles containing either the correctly joined products attL (grey dotted arrow) and attR (grey solid arrow) or the incorrectly joined products where the left site of attB has joined to the left side of attP (LP-LB, shown by the grey dotted line) and vice versa (RB-RP, shown the by grey double headed arrow).C. The sequences of the correctly joined products, attL and attR and the incorrectly joined products, LP-LB (and its inverted sequence, LB-LP, shown in grey) and RB-RP (and its inverted sequence, RP-RB, also shown in grey) derived from recombination between substrates containing attP and attBAA. The sequences have been aligned to either attLTA or attRTA to show where the bases are identical (shown by a dot) between the incorrectly and the correctly joined products.
AAAG AC C CAC
CGC A AG TCAA
LB–Lp
Rp–RB
1722 M. C. A. Smith, R. Till and M. C. M. Smith
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
or ‘antiparallel’ versions of the synapse but that the forma-tion of the recombinant products is dependent on compat-ibility of the sequences at the crossover point.
Integrase leaves a ‘topological footprint’ on substrates that cannot recombine
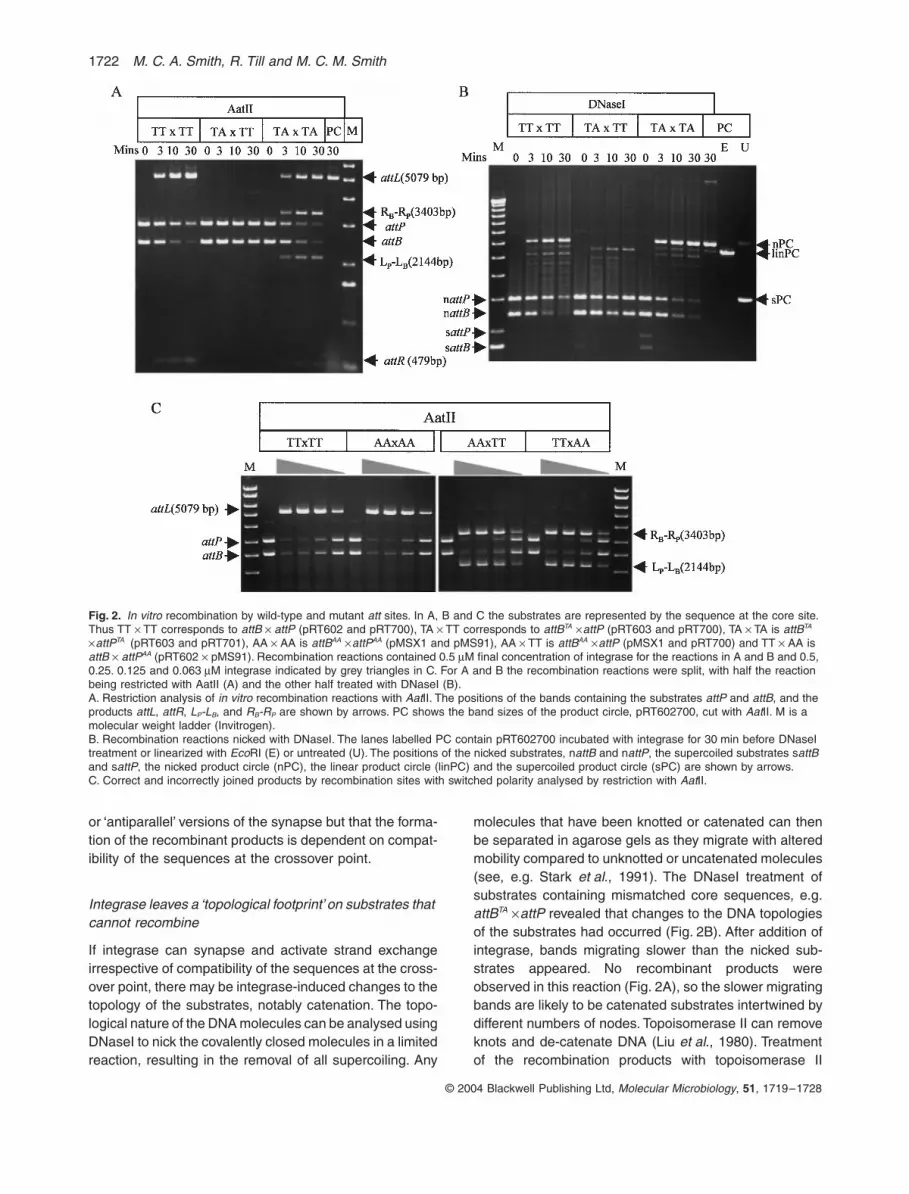
If integrase can synapse and activate strand exchangeirrespective of compatibility of the sequences at the cross-over point, there may be integrase-induced changes to thetopology of the substrates, notably catenation. The topo-logical nature of the DNA molecules can be analysed usingDNaseI to nick the covalently closed molecules in a limitedreaction, resulting in the removal of all supercoiling. Any
molecules that have been knotted or catenated can thenbe separated in agarose gels as they migrate with alteredmobility compared to unknotted or uncatenated molecules(see, e.g. Stark et al., 1991). The DNaseI treatment ofsubstrates containing mismatched core sequences, e.g.attBTA ¥attP revealed that changes to the DNA topologiesof the substrates had occurred (Fig. 2B). After addition ofintegrase, bands migrating slower than the nicked sub-strates appeared. No recombinant products wereobserved in this reaction (Fig. 2A), so the slower migratingbands are likely to be catenated substrates intertwined bydifferent numbers of nodes. Topoisomerase II can removeknots and de-catenate DNA (Liu et al., 1980). Treatmentof the recombination products with topoisomerase II
Fig. 2. In vitro recombination by wild-type and mutant att sites. In A, B and C the substrates are represented by the sequence at the core site. Thus TT ¥ TT corresponds to attB ¥ attP (pRT602 and pRT700), TA ¥ TT corresponds to attBTA ¥attP (pRT603 and pRT700), TA ¥ TA is attBTA ¥attPTA (pRT603 and pRT701), AA ¥ AA is attBAA ¥attPAA (pMSX1 and pMS91), AA ¥ TT is attBAA ¥attP (pMSX1 and pRT700) and TT ¥ AA is attB ¥ attPAA (pRT602 ¥ pMS91). Recombination reactions contained 0.5 mM final concentration of integrase for the reactions in A and B and 0.5, 0.25. 0.125 and 0.063 mM integrase indicated by grey triangles in C. For A and B the recombination reactions were split, with half the reaction being restricted with AatII (A) and the other half treated with DNaseI (B).A. Restriction analysis of in vitro recombination reactions with AatII. The positions of the bands containing the substrates attP and attB, and the products attL, attR, LP-LB, and RB-RP are shown by arrows. PC shows the band sizes of the product circle, pRT602700, cut with AatII. M is a molecular weight ladder (Invitrogen).B. Recombination reactions nicked with DNaseI. The lanes labelled PC contain pRT602700 incubated with integrase for 30 min before DNaseI treatment or linearized with EcoRI (E) or untreated (U). The positions of the nicked substrates, nattB and nattP, the supercoiled substrates sattB and sattP, the nicked product circle (nPC), the linear product circle (linPC) and the supercoiled product circle (sPC) are shown by arrows.C. Correct and incorrectly joined products by recombination sites with switched polarity analysed by restriction with AatII.

Polarity of phage integration 1723
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
Fig. 3. Nature of the synapse and its effect on recombination and the topology of the substrates and products.A. Putative events at the crossover sites with wild-type sites, attB ¥ attP, and with attBAA (with switched polarity) ¥ attP. The attB and attBAA sites are in grey and the attP sites are shown in black. A putative intermediate is shown in all four reactions where integrase has cleaved the DNA (lines indicate the proposed staggered breaks), and initiated strand exchange but is yet to religate the products. In the reaction with attB ¥ attP where the ‘antiparallel’ synapse has formed and in attBAA ¥attP with the ‘parallel’ synapse, there are non-complementary base pairs at the crossover sequence and it is proposed that integrase iterates rotation of two half sites to reform the substrates.B. Some of the proposed topologies generated by integrase-catalysed reactions. The substrates are shown in black and the two parental attachment sites are shown by the small black solid and dashed arrows. Large black arrows indicate iteration of rotation of the half sites during strand exchange and grey arrows indicate recombination. The products are shown in grey. Recombination between wild type sites in an ‘antiparallel’ synapse and iteration of strand exchange leads to catenated parental plasmids (a). Catenation is also predicted between sites with mismatched core sequences irrespective of the type of synapse formed. The putative catenane can then re-synapse in either form to yield one of three possible outcomes; a recombinant product (b), two separated parental plasmids [(c); after formation of a ‘parallel’ synapse and iteration of strand exchange] or a 4-noded catenane [(d); formed for example via further iteration of reaction (a)]. The three versions of the 4-noded catenane, where the att sites are positioned at different relative locations with respect to nodes, is intended to indicate that synapsis of the recombination sites can randomly trap nodes leading to topological complexity in subsequent reactions. Integrase continues to synapse the parental att sites whenever present. Thus the separated parental plasmids from (c) could form a ‘parallel’ synapse and form an unknotted product circle (e) or, after iterative strand exchange (f) a 3-noded product knot (g).

1724 M. C. A. Smith, R. Till and M. C. M. Smith
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
removed most of the bands migrating slower than thesubstrates leaving only some substrate topoisomers, andthese could be completely removed by further treatmentwith DNaseI (Fig. 4). Thus although recombination withthese substrates containing mismatched core sequencesdid not occur, integrase left a ‘topological footprint’ con-sisting of catenated substrate molecules.
The catenation of the substrates containing the mis-matched core sequences may have occurred if strandexchange is initiated and substrates become covalentlyattached to integrase via phospho-serine linkages butrejoining of the DNA backbone to form the recombinantsfails due to non-complementary base pairing. Integrasecan then either reverse the reaction and go back to reformthe original substrates or, by analogy with resolvase/inver-tases, iterate rotation of the cleaved substrate half-sitesto form a catenane (Fig. 3B; Kanaar et al., 1990; Heich-man et al., 1991; Stark et al., 1991). These catenatedsubstrates still contain attBTA and attP and could thereforeact as substrates for repeated rounds of recombination toform more complex catenanes and this would account forthe ladder of different topological forms observed inFig. 2B. These data provide direct evidence that synapsisoccurs between the mismatched sites and integrase isactivated irrespective of whether the products can form.
Integrase forms topologically complex recombination products
In reactions with the wild type sites attB ¥ attP, a ‘parallel’synapse is expected to lead to recombinant productsafter a single strand exchange reaction leading to anunknotted circle (Fig. 3B). Conversely if an ‘antiparallel’synapse forms, the sequences at the core will be non-complementary after strand exchange and integrase isunlikely to form recombinant products (Fig. 3A). Asobserved with the mismatched core sequences above,we expect integrase to iterate rotation of the half sitesleading to the formation of catenated substrates(Fig. 3B). Integrase will continue to act on these sub-strates until a ‘parallel’ synapse is formed and strandexchange leads to recombinant products (Fig. 3B).DNaseI treatment of the attB ¥ attP reactions showedthat unknotted circles are indeed the major topologicalform generated amongst the recombination products, butthere are clearly many other topological forms produced,seen as a ladder of bands migrating faster than themarker plasmid pRT602700 or its linear form (Fig. 2B).As the restriction digest on the same reaction productsindicated that the majority of substrate had recombined(Fig. 2A), we propose that this ladder of bands is a mix-ture of topologically different, knotted recombinants(Fig. 2B). Treatment of the recombination products withtopoisomerase II removed most of the bands migrating
faster than the unknotted relaxed product leaving onlysome topoisomers, which were removed by DNaseI(Fig. 4). Thus integrase acting on the wild-type attP andattB sites produced unknotted circles and a variety ofknotted products.
In reactions with the symmetrical core sites, attPTA ¥attBTA, we expected to observed only unknotted productcircles as, irrespective of the type of synapse formedrecombinant products are not prevented by mismatchesat the crossover sequences. However although the major-ity of the products are unknotted circles, a ladder of topo-logically different products was observed that was asubset of those seen with the wild-type sites and of differ-ent mobilities to the putative catenanes obtained with themismatched sites (Fig. 2B). Thus the symmetrical coresubstrates still apparently generated knotted products.These knots can only arise if integrase fails to rejoinrecombinant products even when there is no mismatchand instead iterates the rotation of the half sites, leadingfirst to catenated substrates but then to recombinant prod-ucts (Fig. 3B). The catenated parental substrates areunlikely to persist as the att sites are in close proximity,favouring the formation of a synapse and further strandexchanges until recombinant products are formed.
It should be noted that once attL and attR are formedthere is no further initiation of strand exchange. Integrasehad no detectable activity on the substrate, pRT602700(Fig. 2) implying that there is no initiation of strandexchange with this combination of substrates. Moreoverassays designed to detect any integrase activity on other
Fig. 4. Topoisomerase II can remove the putative knots and cat-enanes introduced by integrase. Recombination reactions with either wild type (TT ¥ TT using plasmids pRT602 and pRT700) or mis-matched (TA ¥ TT with pRT603 and pRT700) att sites were performed then treated with DNaseI, topoII or DNaseI and topoII. As a control pRT602700 was also incubated with integrase, topoisomerase II and DNaseI. The arrows indicate the position of the nicked product circle (nPC), the linear product (linPC), the supercoiled product (sPC), the nicked substrates (nattB and nattP) and partially relaxed substrates (tattB and tattP).

Polarity of phage integration 1725
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
combinations of att sites, e.g. attL ¥ attR, attP ¥ attL,attB ¥ attB, etc. also failed to show any recombinationproducts or changes in the topology of the substrates(data not shown).
The polarity of phage integration in vivo can be switched using a mutant attB site
We have shown that the sole determinant of the polarityof the recombination sites in fC31 integration in vitro isthe core sequence. We sought to determine whether thisis true in vivo. The fragments encoding attB, attBTA andattBAA were cloned into the vector pRT801 (Gregory
et al., 2003) to form pMX3, pMX4 and pMX5, respec-tively, which when introduced into S. coelicolor, integrateinto the chromosome at a site distal to the fC31 attB site.pMX3, pMX4 and pMX5 were inserted into S. coelicolorJ1929 (attB+) to make JX3, JX4 and JX5 containing thewild-type fC31 attB site in its natural position as well asthe ectopically introduced attB derivates. The same plas-mids, pMX3, pMX4 and pMX5 were also introduced intoPCS41(DattB), an isogenic derivative of J1929 containinga deletion of the natural attB (Combes et al., 2002), tomake DX3, DX4 and DX5, which only have the ectopicallyinserted attB sites. After infection of all six host strainswith a derivative of fC31 carrying a thiostrepton resis-tance determinant, KC859 (Kieser et al., 2000), lysogenswere selected and PCR was then used to determine theorientation of insertion of the phage genome (Fig. 5 andTable 1). The results indicated first, that insertion of anadditional attB site did not prevent occupancy of the nat-ural attB site; all of the JX3, JX4 and JX5 lysogens exam-ined contained KC859 in the natural attB site and theJX3 and JX5 isolates also contained KC859 in theectopic attB sites. Second, when this ectopic site wasattBAA (JX5 and DX5), KC859 was integrated in the incor-rect orientation whereas when it was the wild-type attB(JX3 and DX3), KC859 was in the correct orientation.There was no difference in the relative frequency ofthiostrepton resistant colonies with these strains, sug-gesting there was no inhibition of KC859 integration inattBAA (Table 1). Third, this experiment showed that thereis no context effect on the use of attB for phage integra-tion in vivo. Finally, the poor frequency of thiostreptonresistant colonies in DX4 (DattB, attBTA) demonstratedthat the mismatched core sequence is a powerful barrierto integration in vivo (Table 1). These data show that theorientation of integration of the fC31 derivative KC859can be manipulated by changing the sequence of the2 bp core within attB. Moreover neither the host norKC859 express a factor that influences the alignment ofthe recombination sites at synapsis.
Fig. 5. Switching the polarity of phage integration in vivo. The host strains used for the selection of KC859 lysogens were JX3, JX4 and JX5, derivatives of S. coelicolor J1929 (attB+) (Bedford et al., 1995) or DX3, DX4 and DX5, derivatives of PCS41 (J1929DattB) (Combes et al., 2002) and contain attB (JX3, DX3) attBTA (JX4, DX4) or attBAA (JX5, DX5) introduced ectopically.A. Location of the primer binding sites used to determine the orien-tation of insertion of KC859. The S. coelicolor chromosome is shown as a grey line containing the fC31 attB site (white box) and the attB sites integrated ectopically via the plasmids pMX3, pMX4 and pMX5 (grey box). The integrated KC859 is shown as a line flanked by black circles and containing a central arrowhead that indicates the orientation.B. Shows the PCR products obtained from representative KC859 lysogens. Primer pairs were used to amplify recombination joints at the genomic attB (lanes marked G; RMS73 and RMS69), in the correct orientation at the ectopic attB (lanes marked C; RMS68 and RMS62) and in the incorrect orientation at the ectopic attB (lanes marked I; RMS69 and RMS71).
Table 1. The attB core sequence determines the orientation of inte-gration of KC859.
HostaEctopicattB site
Relative numberof thiostreptonresistant clones
Orientation ofKC859 in the ectopicattB site (% of 20 clones tested):
Correct IncorrectJX3 attB 0.41 100 0JX4 attBTA 0.24 0 0JX5 attBAA 0.42 0 100DX3 attB 1 100 0DX4 attBTA 1.55 ¥ 10-4 0 0DX5 attBAA. 0.362 0 100
a. For host strains see legend to Fig. 4.

1726 M. C. A. Smith, R. Till and M. C. M. Smith
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
Discussion
The data presented here provide an insight into the mech-anism of serine integrases. The effects of changes in thecore sequence within the recombination sites and theproduction of knots and catenanes that are observed withfC31 integrase can all be interpreted using the double-strand cut rotation model as described for the resolvase/invertases (Grindley, 2002; Johnson, 2002). fC31 inte-grase interacts with the attP and attB sites bringing themtogether in either of two alternative forms of synapse, the‘parallel’ and ‘antiparallel’ forms, that lead to either cor-rectly and incorrectly joined molecules depending on the2 bp sequence at the crossover point (Fig. 3A). The 2 bpcore sequence determines the polarity of the sites and theoutcome of recombination.
The ability of fC31 integrase to synapse attB and attPsites freely in either of the two forms is reminiscent of themutant activated resolvases and of Fis-independentmutants of Gin invertase that form correctly and incor-rectly joined products via inter and intramolecular reac-tions (Klippel et al., 1988; Klippel et al., 1993; Arnoldet al., 1999). As with the resolvase/invertases once a syn-apse is formed, irrespective of the compatibility of thecrossover sequences, strand exchange is activated. Onlyif the crossover sequences are complementary willrecombinant products form. If there is a mismatch in thecrossover sequences, generated in our experiments eitherby a single base change or if the wild-type sites form an‘antiparallel’ synapse, integrase can iterate rotation of thehalf-sites to reform the parental substrates, which undergofurther rounds of synapsis and initiation of strandexchange leaving a complex ‘topological footprint’(Figs 2B and 3B). The ability to perform multiple strandexchanges by integrase is also reminiscent of the proper-ties of the resolvase/invertase (Wasserman et al., 1985;Kanaar et al., 1990; Heichman et al., 1991; Stark et al.,1991). We conclude that there are close similaritiesbetween the resolvase/invertases and fC31 integrase inall the steps involved in synapsis and strand exchange.
Unlike the resolvases/invertases, fC31 integrase gen-erates a variety of topologically different recombinationproducts (Fig. 2B). We observed that the range of differentproduct topologies obtained with the wild type sites wasmore complex than those generated with the symmetricalsites (Fig. 2B) and we propose that this is a consequenceof the non-palindromic nature of the core sequence andthe ability by integrase to form both the ‘parallel’ and‘antiparallel’ synapse. In theory the symmetrical sites canproduce either a correct or an incorrectly joined productfrom every synapse. Despite this, evidence was presentedfor iteration of the strand exchange in the form of differentproduct topologies. With the wild-type sites only a ‘parallel’synapse can give rise to recombinants, but data pre-
sented here indicate that an ‘antiparallel’ synapse canoccur with equal frequency, increasing the overall iterativeactivity in the reaction. As the level of recombination is thesame with the symmetrical and the wild-type sites, wepropose that integrase will continue to synapse attP andattB sites until they have recombined to form attL andattR. If in the process nodes, where one DNA strandcrosses another, are trapped randomly at every synapsis,this will lead to variable topologies of the substrate cat-enanes and products. In summary we propose thatrecombination between the wild type sites, attP ¥ attBoften takes a complex pathway through multiple synapsesand iterations of strand exchange until the products attLand attR are formed, after which there is no further activityby integrase.
Changes in the recombination sites that caused aswitch in polarity had no great effect on the efficiency ofintegration in vitro or in vivo. This experiment confirmedthat neither the phage nor the host produced a factor thatinfluenced the structure of the synapse to favour correctjoining. Given that phage-encoded integration/excisionsystems catalysed by tyrosine recombinases have redun-dant mechanisms to maintain the recombination sitepolarity, the work presented here underlines the mecha-nistic and evolutionary differences between these twofamilies of phage integrases (Nash, 1996).
Experimental procedures
Strains and plasmids
Escherichia coli strain DS941 (Summers and Sherratt, 1988)was used to prepare plasmid DNA according to standardmethods (Sambrook and Russell, 2001). pRT700, pRT701and pMS91 contain the attP, attPTA and attPAA , respectively,cloned into pGEM7 (Promega) cut with EcoRI and BstBI byinsertion of annealed oligonucleotides, 5¢-AAT TAG TAG TGCCCC AAC TGG GGT AAC CTT/A t/AGA GTT CTC TCA GTTGGG GGC GTA-3¢ and 5¢-CGT ACG CCC CCA ACT GAGAGA ACT CA/TA/T AGG TTA CCC CAG TTG GGG CAC TACT-3¢. The attB sites were cloned using annealed oligonucle-otides, 5¢-AAT TCC GCG GTG CGG GTG CCA GGG CGTGCC CT/At/A GGG CTC CCC GGG CGC GTA CTC C-3¢and 5¢-CGG GAG TAC GCG CCC GGG GAG CCC a/TA/TGGGC ACG CCC TGG CAC CCG CAC CGC GG-3¢ intopGEM7 to form pRT600, pRT601 and pMS91 encoding attB,attBTA and attBAA and then transferred into pSP72 via thecommon BamHI and EcoRI sites to form pRT602, pRT603and pMSX1 respectively. The inserts from pRT600, pRT601and pMS91 were transferred into the fBT1-derived integrat-ing vector pRT801 (Gregory et al., 2003) via the XbaI/SpeIand BamHI sites to generate pMSX3, pMSX4 and pMSX5respectively. These plasmids were introduced into the non-methylating E. coli strain ET12567(pUZ8001) (MacNeil,1988) for conjugation into the S. coelicolor strains J1929(DpglY) (Bedford et al., 1995) or PCS41 (DpglY, DattB)(Combes et al., 2002) as described by Kieser et al. (2000).The plasmid, pRT602700, was prepared by introduction of

Polarity of phage integration 1727
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
the products of a recombination reaction between pRT602and pRT700 into E. coli DH5a and screening the transfor-mants for the presence of the recombinant plasmid byrestriction analysis.
Recombination assays
One hundred and fifty nanograms of each plasmid substratewere mixed in R buffer (10 mM Tris pH 7.5, 1 mM EDTA pH 8,100 mM NaCl, 5 mM DTT, 5 mM spermidine, 4.5% glyceroland 0.5 mg ml-1 bovine serum albumin). For a time-course,reactions were initiated by adding 0.5 mM of purified integraseand incubating at 30∞C and then stopped at intervals by heat(70∞C, 10 min). For a dilution series, twofold dilutions of inte-grase were performed in R buffer and reactions were incu-bated at 30∞C for 30 min before heating. Treatment afterrecombination may involve splitting the reactions, with halfthe reaction being restricted with AatII and the other halftreated with DNaseI as described previously (Stark et al.,1991). Digestions were halted by the addition of4 ¥ proteinase K mix (50% glycerol, 0.1%SDS, 10 mM TrispH 8.2, 100 mg ml-1 proteinase K, 1 mg ml-1 bromophenolblue), the DNaseI samples were extracted with phenol/chlo-roform and the aqueous layer loaded onto a 0.7% Tris-ace-tate-EDTA buffered agarose gel and electrophoresed atapproximately 3 V cm-1 for about 16 h. Bands were visualisedby staining with ethidium bromide.
Topoisomerase II assays
The products of the recombination reactions were precipi-tated with ethanol and resuspended gently in topoisomeraseII (USB) buffer. Selected samples were then treated with40 ml topoisomerase II (USB) at 30∞C for 1 h, and whereindicated some of these samples were also treated withDNaseI as described above. As a control pRT602700 wasalso incubated with integrase, topoisomerase II and DNaseI.
Preparation and screening of lysogens
Lysogens were prepared as described previously (Kieseret al., 2000). KC859 was mixed with S. coelicolor spore sus-pensions, which were then diluted, plated on R1M mediumand after 8 h overlayed with thiostrepton to select forlysogens. After amplification of spores from 20 colonies fromeach infected strain, PCR analysis was performed usingstandard techniques and the primers RMS62 (5¢-CCCTGAAGGACCGTGTGACG-3¢), RMS68 (5¢-GACCCTACGCCCCCAACTGAG-3¢) RMS69 (5¢-CCCGCGTACGTGTCCACCCCG-3¢), RMS71 (5¢-GCTTCCGGCTCGTATGTTGTG-3¢)and RMS73 (5¢-ACGATGTAGGTCACGGTCTCGAA-3¢).Spore and phage titres of the spore suspensions were per-formed as described (Kieser et al., 2000).
Acknowledgements
We thank Dr William Brown and Dr Peter McGlynn for criticalreading of this manuscript and to Dr Graham Hatfull, DrMarshall Stark and Dr Martin Boocock for stimulating discus-
sion. Funding for this work was provided by the UK Biotech-nology and Biological Sciences Research Council.
References
Andreas, S., Schwenk, F., Kuter-Luks, B., Faust, N., andKuhn, R. (2002) Enhanced efficiency through nuclear local-ization signal fusion on phage fC31-integrase: activitycomparison with Cre and FLPe recombinase in mamma-lian cells. Nucleic Acids Res 30: 2299–2306.
Arnold, P.H., Blake, D.G., Grindley, N.D.F., Boocock, M.R.,and Stark, W.M. (1999) Mutants of Tn3 resolvase whichdo not require accessory binding sites for recombinationactivity. EMBO J 18: 1407–1414.
Bedford, D.J., Laity, C., and Buttner, M.J. (1995) Two genesinvolved in the phase-variable fC31 resistance mechanismof Streptomyces coelicolor A3 (2). J Bacteriol 177: 4681–4689.
Bednarz, A.L., Boocock, M.R., and Sherratt, D.J. (1990)Determinants of correct res site alignment in site-specificrecombination by Tn3 resolvase. Genes Dev 4: 2366–2375.
Belteki, G., Gertsenstein, M., Ow, D.W., and Nagy, A. (2003)Site-specific cassette exchange and germline transmissionwith mouse ES cells expressing fC31 integrase. Nat Bio-technol 21: 321–324.
Combes, P., Till, R., Bee, S., and Smith, M.C. (2002) TheStreptomyces genome contains multiple pseudo-attB sitesfor the fC31-encoded site-specific recombination system.J Bacteriol 184: 5746–5752.
Crellin, P.K., and Rood, J.I. (1997) The resolvase/invertasedomain of the site-specific recombinase TnpX is functionaland recognizes a target sequence that resembles the junc-tion of the circular form of the Clostridium perfringenstransposon Tn4451. J Bacteriol 179: 5148–5156.
Gregory, M.A., Till, R., and Smith, M.C.M. (2003) Integrationsite for Streptomyces phage fBT1 and the development ofnovel site-specific integrating vectors. J Bacteriol 185:5320–5323.
Grindley, N.D.F. (2002) The movement of Tn3-like elements.Transposition and cointegrate resolution. In Mobile DNA II.Craig, N.L., Craigie, R., Gellert, M., and Lambowitz, A.M.,(eds). Washington, DC: American Society for MicrobiologyPress.
Groth, A.C., Olivares, E.C., Thyagarajan, B., and Calos, M.P.(2000) A phage integrase directs efficient site-specific inte-gration in human cells. Proc Natl Acad Sci USA 97: 5995–6000.
Heichman, K.A., Moskowitz, I.P., and Johnson, R.C. (1991)Configuration of DNA strands and mechanism of strandexchange in the Hin invertasome as revealed by analysisof recombinant knots. Genes Dev 5: 1622–1634.
Johnson, R.C. (2002) Bacterial site-specific DNA inversionsystems. In Mobile DNA II. Craig, N.L., Craigie, R., Gellert,M., and Lambowitz, A.M., (eds). Washington DC: AmericanSociety for Microbiology Press, pp. 230–271.
Kanaar, R., Klippel, A., Shekhtman, E., Dungan, J.M., Kah-mann, R., and Cozzarelli, N.R. (1990) Processive recom-bination by the phage Mu Gin system: implications for themechanisms of DNA strand exchange, DNA site alignment,and enhancer action. Cell 62: 353–366.

1728 M. C. A. Smith, R. Till and M. C. M. Smith
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 51, 1719–1728
Katayama, Y., Ito, T., and Hiramatsu, K. (2000) A newclass of genetic element, staphylococcus cassettechromosome mec, encodes methicillin resistance in Sta-phylococcus aureus. Antimicrob Agents Chemother 44:1549–1555.
Kieser, T., Bibb, M.J., Buttner, M.J., Chater, K.F., and Hop-wood, D.A. (2000) Practical Streptomyces Genetics. Nor-wich: The John Innes Foundation.
Klippel, A., Cloppenborg, K., and Kahmann, R. (1988) Isola-tion and characterization of unusual Gin mutants. EMBO J7: 3983–3989.
Klippel, A., Kanaar, R., Kahmann, R., and Cozzarelli, N.R.(1993) Analysis of strand exchange and DNA binding ofenhancer-independent Gin recombinase mutants. EMBOJ 12: 1047–1057.
Krasnow, M.A., and Cozzarelli, N.R. (1983) Site-specificrelaxation and recombination by the Tn3 resolvase: recog-nition of the DNA path between oriented res sites. Cell 32:1313–1324.
Kuhstoss, S., and Rao, R.N. (1991) Analysis of the integra-tion function of the Streptomycete bacteriophage fC31. JMol Biol 222: 897–908.
Liu, L.F., Liu, C.C., and Alberts, B.M. (1980) Type II DNAtopoisomerases: enzymes that can unknot a topologicallyknotted DNA molecule via a reversible double-strandbreak. Cell 19: 697–707.
MacNeil, D.J. (1988) Characterization of a unique methyl-specific restriction system in Streptomyces avermitilis. JBacteriol 170: 5607–5612.
Moskowitz, I.P., Heichman, K.A., and Johnson, R.C.(1991) Alignment of recombination sites in Hin-mediatedsite-specific DNA recombination. Genes Dev 5: 1635–1645.
Nash, H. (1996) Site-specific recombination: Integration,excision, resolution, and inversion of defined DNA seg-ments. In Escherichia Coli and Salmonella, Vol. I. Curtiss,R., III, Ingraham, J., Lin, E., Low, K., Magasanik, B., Rezni-koff, W., et al., (eds). Washington, DC: American Societyfor Microbiology Press, pp. 2363–2376.
Olivares, E.C., Hollis, R.P., and Calos, M.P. (2001) PhageR4 integrase mediates site-specific integration in humancells. Gene 278: 167–176.
Olivares, E.C., Hollis, R.P., Chalberg, T.W., Meuse, L., Kay,M.A., and Calos, M.P. (2002) Site-specific genomic inte-
gration produces therapeutic Factor IX levels in mice. NatBiotechnol 20: 1124–1128.
Ortiz-Urda, S., Thyagarajan, B., Keene, D.R., Lin, Q., Fang,M., Calos, M.P., and Khavari, P.A. (2002) Stable nonviralgenetic correction of inherited human skin disease. NatMed 8: 1166–1170.
Sambrook, J., and Russell, D.W. (2001) Molecular Cloning:A Laboratory Manual. Cold Spring Harbor, New York: ColdSpring Harbor Laboratory Press.
Sarkis, G.J., Murley, L.L., Leschziner, A.E., Boocock, M.R.,Stark, W.M., and Grindley, N.D. (2001) A model for thegamma delta resolvase synaptic complex. Mol Cell 8: 623–631.
Smith, M.C., and Thorpe, H.M. (2002) Diversity in the serinerecombinases. Mol Microbiol 44: 299–307.
Stark, W.M., Boocock, M.R., and Sherratt, D.J. (1989) Site-specific recombination by Tn3 resolvase. Trends Genet 5:304–309.
Stark, W.M., Grindley, N.D.F., Hatfull, G.F., and Boocock,M.R. (1991) Resolvase-catalyzed reactions between sitesdiffering in the central dinucleotide of subsite-I. EMBO J10: 3541–3548.
Stoll, S.M., Ginsburg, D.S., and Calos, M.P. (2002) PhageTP901-1 site-specific integrase functions in human cells. JBacteriol 184: 3657–3663.
Summers, D.K., and Sherratt, D.J. (1988) Resolution ofColE1 dimers requires a DNA sequence implicated in thethree-dimensional organization of the cer site. EMBO J 7:851–858.
Thorpe, H.M., and Smith, M.C.M. (1998) In vitro site-specificintegration of bacteriophage DNA catalyzed by a recombi-nase of the resolvase/invertase family. Proc Natl Acad SciUSA 95: 5505–5510.
Thorpe, H.M., Wilson, S.E., and Smith, M.C.M. (2000) Con-trol of directionality in the site-specific recombination sys-tem of the Streptomyces phage fC31. Mol Microbiol 38:232–241.
Wang, H., and Mullany, P. (2000) The large resolvase TndXis required and sufficient for integration and excision ofderivatives of the novel conjugative transposon Tn5397. JBacteriol 182: 6577–6583.
Wasserman, S.A., Dungan, J.M., and Cozzarelli, N.R. (1985)Discovery of a predicted DNA knot substantiates a modelfor site-specific recombination. Science 229: 171–174.