survey of pyrethroids resistance in indian isolates of rhipicephalus (boophilus) microplus:...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Acta Tropica 125 (2013) 237– 245
Contents lists available at SciVerse ScienceDirect
Acta Tropica
journa l h o me pa g e: www.elsev ier .com/ locate /ac ta t ropica
Survey of pyrethroids resistance in Indian isolates of Rhipicephalus (Boophilus)microplus: Identification of C190A mutation in the domain II of thepara-sodium channel gene
Rinesh Kumar, Gaurav Nagar, Anil Kumar Sharma, Sachin Kumar, D.D. Ray, Pallab Chaudhuri,Srikanta Ghosh ∗
Entomology Laboratory, Parasitology Division, Indian Veterinary Research Institute, Izatnagar 243122, Uttar Pradesh, India
a r t i c l e i n f o
Article history:Received 17 July 2012Received in revised form 3 October 2012Accepted 14 October 2012Available online 22 October 2012
Keywords:Rhipicephalus (Boophilus) microplusDeltamethrin resistanceEsteraseMutationPara-sodium channel gene
a b s t r a c t
Monitoring acaricide resistance and understanding the underlying mechanisms are critically importantin developing strategies for resistance management and tick control. Eighteen isolates of Rhipicephalus(Boophilus) microplus collected from four agro-climatic regions of India were characterized and the resis-tant data were correlated with bioassay results, esterase enzyme activities and with the presence/absenceof point mutation in the para-sodium channel gene. The adult immersion test was standardized to assessthe level of resistance and resistant factors (RF) in the range of 1.2–95.7 were detected. Out of eighteenisolates, three were categorized as susceptible (RF < 1.4), five isolates at level I (RF = 1.5–<5), eight at levelII (RF = 5.1–<25), and one isolate each at level III (RF = 26–<40) and level IV (RF = >41). The esterase enzymeratio and survival% of tick isolates was observed significantly (p < 0.001) correlated with correlation coef-ficient (r) in �- and �-esterase activity. The correlation of determination (R2) for �- and �-esterase activityindicated that 73.3% and 55.3% data points of field isolates were very close to the correlation lines. Fordetection of point mutation, three sites (mutation in domain IIS6, T2134A mutation in domain IIIS6 andC190A mutation in domain IIS4-5 linker) of sodium channel gene were amplified and sequenced. Com-parative sequence analysis identified a cytosine (C) to adenine (A) nucleotide substitution (CTC to ATC) atposition 190 in domain II S4-5 linker region of para-sodium channel gene in six isolates and in referencedeltamethrin resistant IVRI-IV line. The occurrence of mutation in the tick isolates having high resistancefactor suggested that target site insensitivity and enhanced esterase activity is the possible mechanismof resistance to deltamethrin in the Indian isolates of R. (B.) microplus. These results also concluded thatthe mutation site in Indian tick isolates is similar to Australian and Brazilian tick isolates while it is dif-ferent in tick isolates from Mexico and North America. This is the first report of occurrence of mutationin para-sodium channel gene of deltamethrin resistant Indian isolates of R. (B.) microplus.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The cattle tick, Rhipicephalus (Boophilus) microplus (Canestrini)pose serious economic threat to cattle producers in many partsof the world, both directly from physical effects upon infestedanimals and indirectly through the diseases caused by protozoanparasites transmitted by this tick species. The global economiclosses due to tick infestations have been roughly estimated as14,000–18,000 million US$ annually (De Castro and Newson, 1993).In India, the annual control cost of ticks and tick borne diseases(TTBDs) has been estimated at 498.7 million US$ (Minjauw and McLeod, 2003). Besides, a 20–30% reduction in the value of hides
∗ Corresponding author. Tel.: +91 941 0261029; fax: +91 581 2303284.E-mail addresses: [email protected], [email protected] (S. Ghosh).
and skin has been determined due to tick bite marks (Biswas,2003). Control of the tick species along with other medically impor-tant vectors and agricultural pests is largely focused on repeateduse of chemical acaricides viz., synthetic pyrethroids (SPs) andorganophosphates (OPs). The rampant and indiscriminate use ofacaricides by livestock farmers has culminated into developmentof resistance in ticks, which is considered as the main technicalproblem for tick control programme in livestock (Shidrawi, 1990;Ghosh et al., 2006). A survey based on questionnaires in a sampledpopulation of manufacturers and farmers reported the presence ofa wide spread acaricidal resistance in India (FAO, 2004). However,no comprehensive study was undertaken to study the mecha-nism of resistance to SPs operating in Indian cattle tick, R. (B.)microplus.
Insensitive target site SP resistance mechanism has been closelyexamined in many arthropods. The sodium channels are the
0001-706X/$ – see front matter © 2012 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.actatropica.2012.10.006

Author's personal copy
238 R. Kumar et al. / Acta Tropica 125 (2013) 237– 245
primary targets of SPs, which are structural derivatives of the nat-urally occurring pyrethrins (Narahashi, 1988). Intensive use of SPsin arthropod control has led to a worldwide emergence of resistantpopulations. Many of the resistant arthropods carry specific pointmutations in the sodium channel gene and structural alterationsin the sodium channel protein may diminish the interaction ofSPs with sodium channel, reducing the sensitivity to pyrethroids(Dong, 2007; Soderlund, 2008). Most of the mutations in sodiumchannel gene have been reported in domain II S6 transmembranesegment and domain II S4-5 linker region. A small number ofmutations have also been found outside of domain II usuallyin domain I or III. Insensitive target site resistance mechanism,which is now quite ubiquitous among disease vectors and otherarthropod species is also reported in R. (B.) microplus. A mutationin the sodium channel gene was found in Corrales and San Felipeisolates of R. (B.) microplus from Mexico that were extremelyresistant to the acaricide permethrin. This mutation involved asingle substitution of an adenosine (A) for thymidine (T) at theposition 2134 (T2134A) in the sodium channel gene sequence(GenBank accession no. AF134216), resulting in the replacementof a phenylalanine by an isoleucine in transmembrane segmentsix of domain III of the sodium channel gene (He et al., 1999).Studies have shown that this mutation correlates with flumethrin,deltamethrin and cypermethrin resistance in Mexican tick popula-tions (Jamroz et al., 2000; Rosario-Cruz et al., 2005). More recently,another mutation in the domain II S4-5 linker region of the sodiumchannel gene has been reported in cattle tick R. (B.) microplus fromAustralia (Morgan et al., 2009) and Brazil (Nogueira Domingueset al., 2012). The cytosine (C) to adenine (A) mutation at position190 (C190A) in the para-sodium channel gene sequence results inan amino acid substitution from leucine in the susceptible isolate toisoleucine in the resistant isolate. The mutation found in Australianand Brazilian ticks has not been detected in ticks from Mexico(Chen et al., 2009; Rosario-Cruz et al., 2009). The second andless understood mechanism involves esterase enzyme mediatedmetabolic detoxification. A number of assays have been developedto detect elevated expression of esterase via gene amplification(Field et al., 1988) and over-transcription (Fournier et al., 1992).However, In India there is a paucity of information on the statusand mechanisms of development of acaricide resistance in R. (B.)microplus, the most economically important tick infesting Indianlivestock. There is a need to closely monitor acaricide resistanceproblem in India as there is diversity reported in the mechanismof resistance to SPs in R. (B.) microplus from different regions ofthe world (He et al., 1999; Chen et al., 2009). Hence, the aim ofthe present study was to determine the mechanism of resistancein eighteen field isolates of R. (B.) microplus collected from highlytick infested areas of India through correlation of discriminat-ing dose (DD) bioassay results with esterase activity and thepresence/absence of mutation in the para-sodium channel gene.
2. Materials and methods
2.1. Reference susceptible tick line (IVRI-I)
The colony of acaricides susceptible reference IVRI-I line of R.(B.) microplus (NBAII-IVRI-BM-1-1998) was used as the standard toassess susceptibility/resistance status in tick isolates collected fromthe study area. The colony is maintained in the Entomology Labora-tory of Indian Veterinary Research Institute for the last 15 years andhas not been exposed to any acaricides. The susceptibility statusof the colony was established by periodical testing against sev-eral organo-phosphates, organo-chlorines, synthetic pyrethroidsand formamidine compounds in independent bioassays. Differentdevelopmental stages of ticks were reared in glass tubes covered
with cotton cloths and kept in BOD incubator maintained at 28 ◦Cwith 85 ± 2% RH. A group of 10–14 days old larvae were released onthe ears of disease free cross bred calves using ear bag method andthe bags were checked regularly. After 16–18 days, the engorgedfemales dropped in the ear bags were collected for in vitro bioas-says. After 2–3 feeding cycles, calves were set free for a month.The homogeneity amongst different generations of IVRI-I line hasbeen established by uniform entomological data and by analyzingthe sequences of 16s rRNA gene of the tick species (accession nos.GU222462, GU323287, and GU323288) (Kumar et al., 2011).
2.2. Reference deltamethrin resistant tick line (IVRI-IV)
The deltamethrin resistant IVRI-IV line of R. (B.) microplus wasoriginally collected from cattle shed located at Danapur village ofPatna, Bihar, India. The cattle owners of the village reported lowefficacy of deltamethrin used for the control of ticks. As the col-lected samples were not sufficient for effective AIT, adults werereared in the laboratory at 28 ◦C with 85 ± 2% RH for oviposi-tion and hatching of larvae. As mentioned above, larvae werereleased on separate batch of calves and significant number ofadult females was obtained. To determine the acaricide resistancestatus, previously determined discriminating dose (DD) of tech-nical grade deltamethrin (99.9%) (AccuStandard® Inc., USA) wasused in AIT and the resistance factor (RF) was determined (Sharmaet al., 2012). Initially, the ticks were selected from the treatmentof 6X (X = 30 ppm) concentration of deltamethrin. The adult femalethat survived was allowed to lay eggs and the developed larvaewere released on calves for feeding. After completing the cycle theengorged females of the next generation were collected and againtreated with higher concentration of deltamethrin to get the LC50values. The experiment was continued for several generations withthe increasing acaricide pressure. The resistance increase in subse-quent generations was calculated by the method of Gopalan et al.(1996) using the following formula:
Resistance fold increased = LC50 values of the resistant ticksLC50 values of the susceptible ticks
.
The RF of reference IVRI-IV line was calculated as 42.5.
2.3. Sampling
Two stage stratified sampling method was adopted to collectlive engorged females of R. (B.) microplus from animals and fromthe cracks and crevices of organized and unorganized farms. Theareas of collection were selected from four agro-climatic regionsof India (Fig. 1) where tick infestation level is normally very highand SPs and OP insecticides are intensively used for animal hus-bandry and agricultural activities. The isolates (BEG, DNP, DRB,and SUL) were collected from middle gangetic region located at24◦20′10′′–27◦31′15′′N, 82◦19′50′′–88◦17′40′′E with annual tem-perature and rainfall in the range of 4–40 ◦C and 1000–1200 mm,respectively. The isolates (N-24P, S-24P) were collected from areaslocated at 22.56◦N–88.36◦E receiving annual rainfall from 1250to 2500 mm with an average annual temperature in the rangefrom 15 to 35.5 ◦C. The isolates (PAT, BTH, and LDH) were col-lected from trans-gangetic plain region located at 29.30◦–32.32◦N,73.55◦–76.50◦E with very cold winter (−2 ◦C) and hot summer(40 ◦C) and receiving 460–960 mm annual rainfall. The other iso-lates (COR, PRT, UDP, BLW, BSW, JPR, BHT, ALW, and SKR) ofwestern region located at 23.30◦–30.11◦N, 69.29◦–78.17◦E havingvery low annual rainfall of 200–400 mm and average temperatureof 8–48 ◦C.
The female ticks were collected in separate vials, covered withcotton cloths to allow air and moisture exchange, and were trans-ported to the local processing centers. The samples collected from a
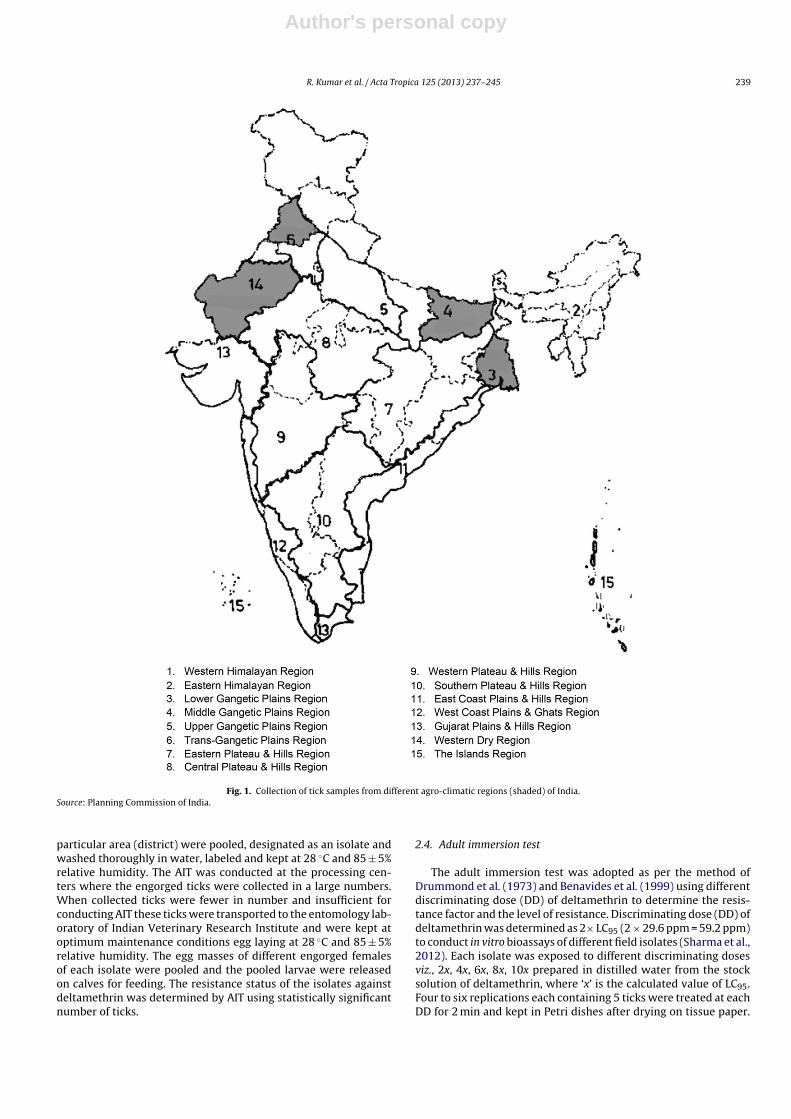
Author's personal copy
R. Kumar et al. / Acta Tropica 125 (2013) 237– 245 239
Fig. 1. Collection of tick samples from different agro-climatic regions (shaded) of India.Source: Planning Commission of India.
particular area (district) were pooled, designated as an isolate andwashed thoroughly in water, labeled and kept at 28 ◦C and 85 ± 5%relative humidity. The AIT was conducted at the processing cen-ters where the engorged ticks were collected in a large numbers.When collected ticks were fewer in number and insufficient forconducting AIT these ticks were transported to the entomology lab-oratory of Indian Veterinary Research Institute and were kept atoptimum maintenance conditions egg laying at 28 ◦C and 85 ± 5%relative humidity. The egg masses of different engorged femalesof each isolate were pooled and the pooled larvae were releasedon calves for feeding. The resistance status of the isolates againstdeltamethrin was determined by AIT using statistically significantnumber of ticks.
2.4. Adult immersion test
The adult immersion test was adopted as per the method ofDrummond et al. (1973) and Benavides et al. (1999) using differentdiscriminating dose (DD) of deltamethrin to determine the resis-tance factor and the level of resistance. Discriminating dose (DD) ofdeltamethrin was determined as 2× LC95 (2 × 29.6 ppm = 59.2 ppm)to conduct in vitro bioassays of different field isolates (Sharma et al.,2012). Each isolate was exposed to different discriminating dosesviz., 2x, 4x, 6x, 8x, 10x prepared in distilled water from the stocksolution of deltamethrin, where ‘x’ is the calculated value of LC95.Four to six replications each containing 5 ticks were treated at eachDD for 2 min and kept in Petri dishes after drying on tissue paper.

Author's personal copy
240 R. Kumar et al. / Acta Tropica 125 (2013) 237– 245
Table 1Primers used to amplify the targeted regions of sodium channel gene.
Name Primer sequence 5′-3′ Region amplified Product size Ref.
D2FD2R
ACGTTCGTTTCGTCTGCTAGATTCGCTTGGGACAGATT
Domain II-S6 (kdr) 434 bp Jamroz et al. (2000)
D3FD3R
CTGGTTACATCATATCTAATTGCCACCCAGCCTTCTTCTTTTGTTCATTG
Domain III-S6 193 bp Chen et al. (2009)
L2FL2R
TACGTGTGTTCAAGCCTAACTTTCTTCGTAGTTCTTGC
Domain II S4-5 linker (Superkdr) 167 bp Morgan et al. (2009)
After 24 h, the treated ticks were transferred to 10 ml tick-rearingtubes covered with cotton cloths and were placed in incubatormaintained at 28 ◦C and 85 ± 5% relative humidity. The entomo-logical data was recorded regularly. The LC50 value of differentisolates was determined by applying regression equation analysisto the probit transformed data of mortality using GraphPad Prismversion 4.0, San Diego, CA, USA. Resistance factors (RFs) for dif-ferent isolates were worked out by the quiescent between LC50 offield isolates and LC50 of reference susceptible IVRI-I line of R. (B.)microplus (Castro-Janer et al., 2009). As per the calculated value ofresistance factor (RF), the resistance status in field isolates of R. (B.)microplus was categorized as susceptible (RF ≤ 1.4), resistance levelI (RF = 1.5–5.0), level II (RF = 5.1–25.0), level III (RF = 26–40) and levelIV (RF ≥ 41) (Kumar Sachin et al., 2011).
2.5. Esterase assay
Esterase activities with the substrates �-and �-naphthyl acetatewere determined in the ticks according to the method ofHemingway (1998) with some modifications. Twenty deep frozenlarvae were homogenized in a precooled glass pestle in 200 �lof distilled water. The homogenates were spun at 1100 × g ina refrigerated centrifuge at 4 ◦C for 15 min and resulting super-natant was used for assay. Reaction mixtures contained 20 �l ofthe homogenate in quadruplicate adjacent wells (two wells eachfor �-and �-naphthyl acetate) of microtitre plate and 200 �l of �-and �-naphthyl acetate solution (250 �l of 30 mM stock in 25 mlof phosphate buffer 0.02 M, pH 7.2.), respectively. The reactionmixtures were incubated at room temperatures for 30 min beforeaddition of 50 �l of fast blue solution (0.023 g fast blue salt dissolvedin 2.25 ml distilled water and 5.25 ml of 5% SDS in 0.1 M sodiumphosphate buffer, pH 7.2) to each well. The plates were incubatedfor 5 min at room temperature and absorbance was measured at570 nm in a microtitre plate reader (Tecan, Austria) operated by apersonal computer using Magellan 6 software. The resulting opti-cal densities (ODs) were compared with standard curves of ODs forknown concentrations of the products �-and �-naphthyl acetate,respectively. The esterase activities were expressed as enzymeratio (mean activity of enzyme in resistant isolate/mean activityof enzyme in reference susceptible IVRI-I line).
2.6. Extraction of genomic DNA, RNA, amplification andsequencing
Ten to fifteen days old, unfed larvae emanating from referenceIVRI-I, IVRI-IV lines and field isolates, whose resistance status wascharacterized were used to isolate total RNA and genomic DNA.The total RNA was extracted from about 100 mg tick larvae usingTrizol reagent (Sigma, USA) following the manufacturer’s protocol.The integrity of RNA was checked by gel electrophoresis and con-centration was determined in Nanodrop 3300C spectrophotometer(Thermo scientific, USA). The cDNA was synthesized from 3 �g oftotal RNA using the RevertAidTM H minus Reverse Transcription Kitusing OligodT primer (Fermentas, Germany). The cDNA was storedat −20 ◦C until use. The genomic DNA was extracted from 400 mg
tick larvae by phenol–chloroform extraction as per the standardtechnique (Sambrook et al., 2001). Genomic DNA was preserved in200 �l of TE buffer.
The PCR primers to amplify the fragments of the sodium chan-nel gene flanking the mutation sites were designed from the partialsodium channel R. (B.) microplus gene sequence (Mexican strain,GenBank accession no. AF134216). Nucleotide sequences of theprimer pairs, the product sizes and the regions amplified are indi-cated in Table 1. Domain IIS6 was amplified by primer pair D2F andD2R; domain IIIS6 was amplified by primer pair D3F and D3R andS4-5 linker region in domain-II was amplified by primer pair L2Fand L2R. The lyophilized primers were resuspended in TE bufferand the stock solution was further diluted in nuclease free water toobtain a working solution of 10 pmol/�l.
First strand cDNA generated from the larvae of IVRI-I, IVRI-IVlines and from 18 field isolates were used as a template for PCRamplification of knockdown resistance (kdr) region (domain IIS6)of the sodium channel gene. PCR reaction was carried out in a 25 �lreaction volume containing 2.5 �l of 10× AccuPrime PCR buffer I,5.0 �l cDNA (1:5 dilution, 100 ng/�l); 1.0 �l of each primer, D2Fand D2R, 0.3 �l of AccuPrime Taq DNA polymerase (5 IU/�l) (Invi-trogen, USA). The PCR conditions optimized as one cycle of initialdenaturation at 94 ◦C for 2 min followed by 35 cycles of 94 ◦C for1 min, 50 ◦C for 1 min, 68 ◦C for 1 min and a final extension at 68 ◦Cfor 10 min.
Genomic DNA was isolated from larvae of IVRI-I, IVRI-IV linesand eighteen field isolates and was used as a template for PCRamplification of domain IIIS6 region (encompassing the T2134Amutation site) and domain II S4-5 (encompassing the C190Amutation site) in the voltage-gated sodium channel gene. Foramplification of T2134A mutation site, a 25 �l PCR reaction was setup using 2.5 �l of 10× PCR buffer; 5.0 �l genomic DNA (1:5 dilu-tion, 50 ng/�l); 0.5 �l dNTP (10 mM), 0.75 �l of each primer, D3Fand D3R, 0.3 �l of DreamTaq DNA polymerase (5 IU/�l) (Fermentas,Germany). The PCR conditions optimized as an initial denaturationstep at 95 ◦C for 2 min, followed by 34 cycles of 95 ◦C for 1 min,55 ◦C for 30 s and 72 ◦C for 30 s with a final extension step at 72 ◦Cfor 10 min.
The C190A mutation site was amplified by PCR using a set ofprimers, L2F and L2R. A 25 �l PCR reaction was performed using2.5 �l of 10× AccuPrime PCR buffer II, 5.0 �l genomic DNA (1:5dilution, 50 ng/�l), 1.0 �l of each primer, 0.3 �l of AccuPrime TaqDNA polymerase (5 IU/�l) (Invitrogen, USA). Thermal cycling con-ditions were: initial denaturation of 94 ◦C for 2 min and followedby 40 cycles each consisting of successive incubations at 94 ◦C for1 min, 50 ◦C for 30 s, 68 ◦C for 30 s with a final extension step at68 ◦C for 10 min. All amplifications were carried out in a Veriti Ther-mal Cycler (Applied Biosystems, USA). The positive amplificationof genes was visualized by electrophoresis of the product in ethid-ium bromide stained 1.5% and 3% metaphor agarose gel. The PCRproducts were purified using QIAquick gel extraction kit (Qiagen,Germany).
The purified PCR product of T2134A mutation site (193 bp) andC190A mutation site (167 bp) were subjected to double strandedcustom DNA sequencing. The purified PCR product of kdr region
Author's personal copy
R. Kumar et al. / Acta Tropica 125 (2013) 237– 245 241
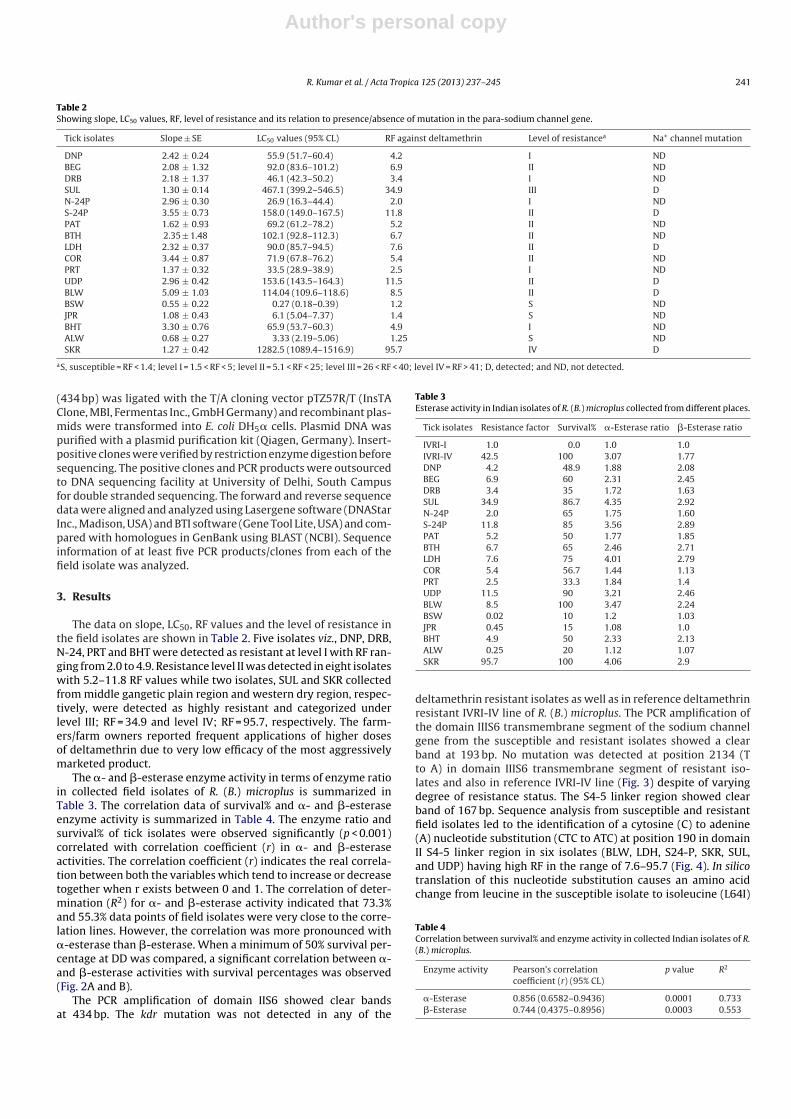
Table 2Showing slope, LC50 values, RF, level of resistance and its relation to presence/absence of mutation in the para-sodium channel gene.
Tick isolates Slope ± SE LC50 values (95% CL) RF against deltamethrin Level of resistancea Na+ channel mutation
DNP 2.42 ± 0.24 55.9 (51.7–60.4) 4.2 I NDBEG 2.08 ± 1.32 92.0 (83.6–101.2) 6.9 II NDDRB 2.18 ± 1.37 46.1 (42.3–50.2) 3.4 I NDSUL 1.30 ± 0.14 467.1 (399.2–546.5) 34.9 III DN-24P 2.96 ± 0.30 26.9 (16.3–44.4) 2.0 I NDS-24P 3.55 ± 0.73 158.0 (149.0–167.5) 11.8 II DPAT 1.62 ± 0.93 69.2 (61.2–78.2) 5.2 II NDBTH 2.35 ± 1.48 102.1 (92.8–112.3) 6.7 II NDLDH 2.32 ± 0.37 90.0 (85.7–94.5) 7.6 II DCOR 3.44 ± 0.87 71.9 (67.8–76.2) 5.4 II NDPRT 1.37 ± 0.32 33.5 (28.9–38.9) 2.5 I NDUDP 2.96 ± 0.42 153.6 (143.5–164.3) 11.5 II DBLW 5.09 ± 1.03 114.04 (109.6–118.6) 8.5 II DBSW 0.55 ± 0.22 0.27 (0.18–0.39) 1.2 S NDJPR 1.08 ± 0.43 6.1 (5.04–7.37) 1.4 S NDBHT 3.30 ± 0.76 65.9 (53.7–60.3) 4.9 I NDALW 0.68 ± 0.27 3.33 (2.19–5.06) 1.25 S NDSKR 1.27 ± 0.42 1282.5 (1089.4–1516.9) 95.7 IV D
aS, susceptible = RF < 1.4; level I = 1.5 < RF < 5; level II = 5.1 < RF < 25; level III = 26 < RF < 40; level IV = RF > 41; D, detected; and ND, not detected.
(434 bp) was ligated with the T/A cloning vector pTZ57R/T (InsTAClone, MBI, Fermentas Inc., GmbH Germany) and recombinant plas-mids were transformed into E. coli DH5� cells. Plasmid DNA waspurified with a plasmid purification kit (Qiagen, Germany). Insert-positive clones were verified by restriction enzyme digestion beforesequencing. The positive clones and PCR products were outsourcedto DNA sequencing facility at University of Delhi, South Campusfor double stranded sequencing. The forward and reverse sequencedata were aligned and analyzed using Lasergene software (DNAStarInc., Madison, USA) and BTI software (Gene Tool Lite, USA) and com-pared with homologues in GenBank using BLAST (NCBI). Sequenceinformation of at least five PCR products/clones from each of thefield isolate was analyzed.
3. Results
The data on slope, LC50, RF values and the level of resistance inthe field isolates are shown in Table 2. Five isolates viz., DNP, DRB,N-24, PRT and BHT were detected as resistant at level I with RF ran-ging from 2.0 to 4.9. Resistance level II was detected in eight isolateswith 5.2–11.8 RF values while two isolates, SUL and SKR collectedfrom middle gangetic plain region and western dry region, respec-tively, were detected as highly resistant and categorized underlevel III; RF = 34.9 and level IV; RF = 95.7, respectively. The farm-ers/farm owners reported frequent applications of higher dosesof deltamethrin due to very low efficacy of the most aggressivelymarketed product.
The �- and �-esterase enzyme activity in terms of enzyme ratioin collected field isolates of R. (B.) microplus is summarized inTable 3. The correlation data of survival% and �- and �-esteraseenzyme activity is summarized in Table 4. The enzyme ratio andsurvival% of tick isolates were observed significantly (p < 0.001)correlated with correlation coefficient (r) in �- and �-esteraseactivities. The correlation coefficient (r) indicates the real correla-tion between both the variables which tend to increase or decreasetogether when r exists between 0 and 1. The correlation of deter-mination (R2) for �- and �-esterase activity indicated that 73.3%and 55.3% data points of field isolates were very close to the corre-lation lines. However, the correlation was more pronounced with�-esterase than �-esterase. When a minimum of 50% survival per-centage at DD was compared, a significant correlation between �-and �-esterase activities with survival percentages was observed(Fig. 2A and B).
The PCR amplification of domain IIS6 showed clear bandsat 434 bp. The kdr mutation was not detected in any of the
Table 3Esterase activity in Indian isolates of R. (B.) microplus collected from different places.
Tick isolates Resistance factor Survival% �-Esterase ratio �-Esterase ratio
IVRI-I 1.0 0.0 1.0 1.0IVRI-IV 42.5 100 3.07 1.77DNP 4.2 48.9 1.88 2.08BEG 6.9 60 2.31 2.45DRB 3.4 35 1.72 1.63SUL 34.9 86.7 4.35 2.92N-24P 2.0 65 1.75 1.60S-24P 11.8 85 3.56 2.89PAT 5.2 50 1.77 1.85BTH 6.7 65 2.46 2.71LDH 7.6 75 4.01 2.79COR 5.4 56.7 1.44 1.13PRT 2.5 33.3 1.84 1.4UDP 11.5 90 3.21 2.46BLW 8.5 100 3.47 2.24BSW 0.02 10 1.2 1.03JPR 0.45 15 1.08 1.0BHT 4.9 50 2.33 2.13ALW 0.25 20 1.12 1.07SKR 95.7 100 4.06 2.9
deltamethrin resistant isolates as well as in reference deltamethrinresistant IVRI-IV line of R. (B.) microplus. The PCR amplification ofthe domain IIIS6 transmembrane segment of the sodium channelgene from the susceptible and resistant isolates showed a clearband at 193 bp. No mutation was detected at position 2134 (Tto A) in domain IIIS6 transmembrane segment of resistant iso-lates and also in reference IVRI-IV line (Fig. 3) despite of varyingdegree of resistance status. The S4-5 linker region showed clearband of 167 bp. Sequence analysis from susceptible and resistantfield isolates led to the identification of a cytosine (C) to adenine(A) nucleotide substitution (CTC to ATC) at position 190 in domainII S4-5 linker region in six isolates (BLW, LDH, S24-P, SKR, SUL,and UDP) having high RF in the range of 7.6–95.7 (Fig. 4). In silicotranslation of this nucleotide substitution causes an amino acidchange from leucine in the susceptible isolate to isoleucine (L64I)
Table 4Correlation between survival% and enzyme activity in collected Indian isolates of R.(B.) microplus.
Enzyme activity Pearson’s correlationcoefficient (r) (95% CL)
p value R2
�-Esterase 0.856 (0.6582–0.9436) 0.0001 0.733�-Esterase 0.744 (0.4375–0.8956) 0.0003 0.553
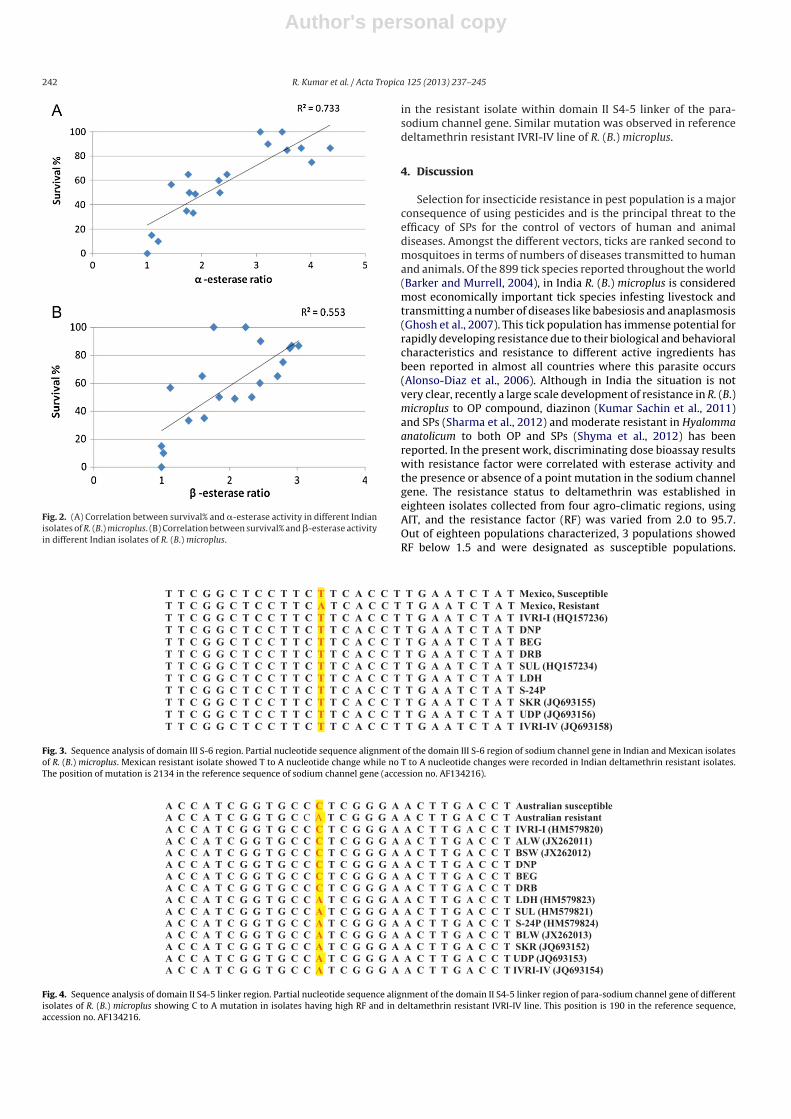
Author's personal copy
242 R. Kumar et al. / Acta Tropica 125 (2013) 237– 245
Fig. 2. (A) Correlation between survival% and �-esterase activity in different Indianisolates of R. (B.) microplus. (B) Correlation between survival% and �-esterase activityin different Indian isolates of R. (B.) microplus.
in the resistant isolate within domain II S4-5 linker of the para-sodium channel gene. Similar mutation was observed in referencedeltamethrin resistant IVRI-IV line of R. (B.) microplus.
4. Discussion
Selection for insecticide resistance in pest population is a majorconsequence of using pesticides and is the principal threat to theefficacy of SPs for the control of vectors of human and animaldiseases. Amongst the different vectors, ticks are ranked second tomosquitoes in terms of numbers of diseases transmitted to humanand animals. Of the 899 tick species reported throughout the world(Barker and Murrell, 2004), in India R. (B.) microplus is consideredmost economically important tick species infesting livestock andtransmitting a number of diseases like babesiosis and anaplasmosis(Ghosh et al., 2007). This tick population has immense potential forrapidly developing resistance due to their biological and behavioralcharacteristics and resistance to different active ingredients hasbeen reported in almost all countries where this parasite occurs(Alonso-Diaz et al., 2006). Although in India the situation is notvery clear, recently a large scale development of resistance in R. (B.)microplus to OP compound, diazinon (Kumar Sachin et al., 2011)and SPs (Sharma et al., 2012) and moderate resistant in Hyalommaanatolicum to both OP and SPs (Shyma et al., 2012) has beenreported. In the present work, discriminating dose bioassay resultswith resistance factor were correlated with esterase activity andthe presence or absence of a point mutation in the sodium channelgene. The resistance status to deltamethrin was established ineighteen isolates collected from four agro-climatic regions, usingAIT, and the resistance factor (RF) was varied from 2.0 to 95.7.Out of eighteen populations characterized, 3 populations showedRF below 1.5 and were designated as susceptible populations.
T T C G G C T C C T T C T T C A C C T T G A A T C T A T Mexico, Susceptible
T T C G G C T C C T T C A T C A C C T T G A A T C T A T Mexico, Resistant
T T C G G C T C C T T C T T C A C C T T G A A T C T A T IVRI-I (HQ157236 )
T T C G G C T C C T T C T T C A C C T T G A A T C T A T DNP
T T C G G C T C C T T C T T C A C C T T G A A T C T A T BEG
T T C G G C T C C T T C T T C A C C T T G A A T C T A T DRB
T T C G G C T C C T T C T T C A C C T T G A A T C T A T SUL (HQ157234 )
T T C G G C T C C T T C T T C A C C T T G A A T C T A T LDH
T T C G G C T C C T T C T T C A C C T T G A A T C T A T S-24P
T T C G G C T C C T T C T T C A C C T T G A A T C T A T SKR (JQ693155)
T T C G G C T C C T T C T T C A C C T T G A A T C T A T UDP (JQ693156)
T T C G G C T C C T T C T T C A C C T T G A A T C T A T IV RI-IV (JQ693158)
Fig. 3. Sequence analysis of domain III S-6 region. Partial nucleotide sequence alignment of the domain III S-6 region of sodium channel gene in Indian and Mexican isolatesof R. (B.) microplus. Mexican resistant isolate showed T to A nucleotide change while no T to A nucleotide changes were recorded in Indian deltamethrin resistant isolates.The position of mutation is 2134 in the reference sequence of sodium channel gene (accession no. AF134216).
A C C A T C G G T G C C C T C G G G A A C T T G A C C T Australian susceptible
A C C A T C G G T G C C A T C G G G A A C T T G A C C T Aus tralian resistant
A C C A T C G G T G C C C T C G G G A A C T T G A C C T IVRI-I (HM579820 )
A C C A T C G G T G C C C T C G G G A A C T T G A C C T ALW (JX262011 )
A C C A T C G G T G C C C T C G G G A A C T T G A C C T BSW (JX262012 )
A C C A T C G G T G C C C T C G G G A A C T T G A C C T DNP
A C C A T C G G T G C C C T C G G G A A C T T G A C C T BEG
A C C A T C G G T G C C C T C G G G A A C T T G A C C T DRB
A C C A T C G G T G C C A T C G G G A A C T T G A C C T LDH (HM579823 )
A C C A T C G G T G C C A T C G G G A A C T T G A C C T SUL (HM579821)
A C C A T C G G T G C C A T C G G G A A C T T G A C C T S-24P (HM579824 )
A C C A T C G G T G C C A T C G G G A A C T T G A C C T BLW (JX262013 )
A C C A T C G G T G C C A T C G G G A A C T T G A C C T SKR (JQ693152)
A C C A T C G G T G C C A T C G G G A A C T T G A C C T UDP (JQ693153)
A C C A T C G G T G C C A T C G G G A A C T T G A C C T IVRI -IV (JQ693154 )
Fig. 4. Sequence analysis of domain II S4-5 linker region. Partial nucleotide sequence alignment of the domain II S4-5 linker region of para-sodium channel gene of differentisolates of R. (B.) microplus showing C to A mutation in isolates having high RF and in deltamethrin resistant IVRI-IV line. This position is 190 in the reference sequence,accession no. AF134216.

Author's personal copy
R. Kumar et al. / Acta Tropica 125 (2013) 237– 245 243
The other fifteen populations were considered resistant at I–IVlevel according to the classification of Kumar Sachin et al. (2011).From the data it is very clear that two isolates, SUL and SKRhaving RF of 34.9 and 95.7, respectively, were collected from theareas where SPs are probably not at all effective against the tickpopulations. In both the areas use of SPs has reached at alarminglyhigh level without maintaining any dose regime (Sharma et al.,2012). Besides, six samples (Table 2) were having RF more than6.0, a level considered enough to impair the use of deltamethrin inthe field (Patarroyo and Costa, 1980). The isolates characterized assusceptible were collected from the areas where OP compoundsare more frequently used than SP and thus high level of resistanceto diazinon was recorded in these areas (Kumar Sachin et al., 2011).
Esterase based resistance has been demonstrated to be one ofthe mechanisms for SPs and OP detoxification in insects and in R. (B.)microplus. However, the specific mechanism through which resis-tance is conferred has not been suitably elucidated (Hemingwayet al., 1993; Rosario-Cruz et al., 1997; Jamroz et al., 2000; Zhu et al.,2004). In the present study, 73.3% and 55.3% data points for � and�-esterase, respectively, of field isolates were close to the correla-tion point with survival percentages confirming esterase mediatedresistant mechanism is operating in R. (B.) microplus population inIndia (Table 4).
The in silico analysis was performed to detect point mutationsin three specific regions of the sodium channel gene of the fieldisolated, R. (B.) microplus. One major mechanism of resistance tar-geting the sodium channel gene is known as knock down resistance(kdr) in which there is reduced target site sensitivity for pyrethroidsresulting from one or more point mutations in domain IIS6 ofsodium channel gene. The most frequently encountered mutationof kdr found in the house fly include a substitution of leucine byphenylalanine (L1014F) and a variety of this mutations (L1014Sor L1014H) are found in a range of important agricultural anddisease-transmitting arthropods including tobacco budworm (Parkand Taylor, 1997), horn fly, Haematobia irritans (Guerrero et al.,1997), diamondback moth, Plutella xylostella (Schuler et al., 1998),peach-potato aphid, Myzus persicae (Martinez-Torres et al., 1999a),mosquitoes, Anopheles gambiae and Culex pipiens (Martinez-Torreset al., 1999b) and Colorado potato beetle, Leptinotarsa decemlineata(Lee et al., 1999). Although kdr mutation is the most widely foundmutation associated with pyrethroid resistance, it is not detectedin any of the pyrethroid resistant Mexican isolates of southern cat-tle tick, R. (B.) microplus (He et al., 1999; Jamroz et al., 2000). Thepresent investigation also failed to detect mutation in this regionof sodium channel gene (domain IIS6) in the resistant isolates of R.(B.) microplus from India (Fig. 3).
Another nucleotide substitution at position 2134 (T2134A) indomain IIIS6 transmembrane segment of the sodium channel genewas detected in San Felipe and Corrales isolates of R. (B.) microplusin Mexico that were extremely resistant to pyrethroid permethrin(He et al., 1999). To date this mutation in domain IIIS6 has beendetected in many tick isolates from North America (Guerrero et al.,2002; Rosario-Cruz et al., 2005; Miller et al., 2007; Chen et al.,2009; Aguirre et al., 2010; Rodriguez-Vivas et al., 2012). Resis-tance conferring mutations in the domain IIIS6 transmembranesegment of the sodium channel gene have also been identified inseveral pyrethroid resistant arthropods such as fruitfly, Dorsophilamelanogaster (Pittendrigh et al., 1997; Martin et al., 2000), two-spotted spider mites, Tetranychus urticae (Tsagkarakou et al., 2009),itch mites, Sarcoptes scabiei (Pasay et al., 2008) and mosquito, Aedesaegypti (Yanola et al., 2010). The double stranded sequence analysisfrom eighteen pyrethroid resistant tick isolates and from laboratoryestablished deltamethrin resistant IVRI-IV line did not detect muta-tions in the domain IIIS6 region of sodium channel gene of Indianisolates of R. (B.) microplus. The absence of T2134A mutation hasalso been reported in various pyrethroid resistant tick isolates from
Australia and Brazil (Li et al., 2007; Chen et al., 2009; Rosario-Cruzet al., 2009; Andreotti et al., 2011).
A mutation which included substitution of adenine (A) by cyto-sine (C) (CTC to ATC) was reported at position 190 in the domainII S4-5 linker of the sodium channel gene of Parkhurst isolate of R.(B.) microplus from Australia, which was resistant to all pyrethroidsincluding flumethrin, cyhalothrin and deltamethrin (Morgan et al.,2009). A similar mutation has been discovered in whitefly, B. tabaci(Morin et al., 2002) and head lice, Pediculus capatis (Lee et al., 2000)in which it confers resistance to SPs. In the present investigations, amutation in the domain II S4-5 linker region of the sodium channelgene has been detected in six populations having high resistancefactors (level II–level IV). This is the first report from India detec-ting a point mutation in the para-sodium channel gene possiblyresponsible for conferring high level of resistance against SP in R.(B.) microplus.
In the present study, a direct correlation between RF, esteraseactivity and mutation (C190A) in the domain II S4-5 linker of para-sodium channel gene was observed when RF is reached more than7.6. The results gives a significant clue to develop a monitoring andwarning system to restrict the use of SPs in area (s) where RF hasreached above the threshold level of 7.6.
The analysis of mutation in the sodium channel gene of R.(B.) microplus from Australia, Brazil, Mexico and India leads tothe conclusion that different resistance mechanism have appar-ently developed between these isolates of R. (B.) microplus. Theseresults suggest that distinct sodium channel gene mutations maybe selected in different arthropod species in response to pyrethroiddrug pressure and due to geographical isolation (He et al., 1999;Pasay et al., 2006). In a recent review Guerrero et al. (2012) reportedthat Domain III mutation seems to be localized to North America,the Morgan et al. (2009) mutation was discovered in Australia butalso reported in Brazil (Nogueira Domingues et al., 2012) while theJonsson et al. (2010) mutation is only reported in Australia. Thepresent information added new dimension to the distribution ofdomain II mutation in the cattle tick, R. (B.) microplus.
5. Conclusions
In India, the R. (B.) microplus populations have developedresistance to deltamethrin and mechanism of development of resis-tance has possibly been mediated by over-expression of esteraseenzymes and mutation in domain II S4-5 linker region of para-sodium channel gene.
Acknowledgements
The authors are grateful to Indian Council of Agricul-tural Research, New Delhi for funding through World Bankfunded National Agricultural Innovation Project No. NAIP/Comp-4/C2066/2008-09. Authors are also grateful to the Veterinaryofficers posted at different tick collection spots for their support.
References
Aguirre, M., Flores, E.A., Alvarez, G., Molina, A., Rodriguez, I., Ponce, G., 2010. Anovel amino acid substitution in the para-sodium channel gene in Rhipicephalusmicroplus (Acari, Ixodidae) associated with knockdown resistance. Experimentaland Applied Acarology 52, 377–382.
Alonso-Diaz, M.A., Rodriguez-Vivas, R.I., Fragoso-Sanchez, H., Rosario-Cruz, R., 2006.Ixodicide resistance of the Boophilus microplus tick to ixodicides. Archivos deMedicina Veterinaria 38, 105–114.
Andreotti, R., Guerrero, F.D., Soares, M.A., Barros, J.C., Miller, R., Perez De Leon, A.A.,2011. Acaricide resistance of Rhipicephalus (Boophilus) microplus in State of MatoGrosso do Sul, Brazil. Revista Brasileira de Parasitologia Veterinária 20, 127–133.
Barker, S.C., Murrell, A., 2004. Systematics and evolution of ticks with a list of validgenus and species names. Parasitology 129, S15–S36.
Benavides, O.E., Romero, N.A., Rodriguez, J.L., Silva, Z.J., 1999. Evidencia preliminarde la aparicion de resistencia a lactonas macrociclicas en cepas de garrapata

Author's personal copy
244 R. Kumar et al. / Acta Tropica 125 (2013) 237– 245
Boophilus microplus en Colombia. In: Memorias IV Seminarios Internacional deParasitologia, Puerto Vallarta, Jalisco, Mexico, October 20–22, pp. 260–264.
Biswas, S., 2003. Effects of ticks on animal production system. In: Proceedings ofthe National Seminar on Leather Industry in Today’s Perspective, Kolkata, India,November 14–15.
Castro-Janer, E., Rifran, L., Piaggio, J., Gil, A., Miller, R.J., Schumaker, T.T.S., 2009.In vitro tests to establish LC50 and discriminating concentrations for fipronilagainst Rhipicephalus (Boophilus) microplus (Acari, Ixodidae) and their standard-ization. Veterinary Parasitology 162, 120–128.
Chen, A.C., He, H., Temeyer, K.B., Jones, S., Green, P., Barker, S.C., 2009. A surveyof Rhipicephalus microplus populations for mutation associated with pyrethroidresistance. Journal of Economic Entomology 102, 373–380.
De Castro, J.J., Newson, R.M., 1993. Host resistance in cattle tick control. ParasitologyToday 9, 13–17.
Dong, K., 2007. Insect sodium channels and insecticide resistance. Invertebrate Neu-roscience 7, 17–30.
Drummond, R.O., Crust, S.E., Trevino, J.L., Gladney, W.J., Grahan, O.H., 1973. Boophilusannulatus and laboratory tests of insecticides. Journal of Economic Entomology66, 130–133.
FAO, 2004. Guidelines Resistance Management and Integrated Parasite Control inRuminants. Module 1: Ticks: Acaricide Resistance: Diagnosis, Management andPrevention. Food and Agriculture Organization, Animal Production and HealthDivision, Rome, pp. 1–53.
Field, L.M., Devonshire, A.L., Forde, B.G., 1988. Molecular evidence that insecticideresistance in peach-potato aphids (Myzus persicae Sulz,) results from amplifica-tion of an esterase gene. Biochemical Journal 251, 309–315.
Fournier, D., Bride, M., Hoffman, F., Karch, F., 1992. Acetylcholinesterase: two typesof modifications confer resistance to insecticide. Journal of Biological Chemistry15, 14270–14274.
Ghosh, S., Bansal, G.C., Gupta, S.C., Ray, D., Khan, M.Q., Irshad, H., Shahiduzzaman,M., Seitzer, U., Ahmed, J.S., 2007. Status of tick distribution in Bangladesh, Indiaand Pakistan. Parasitology Research 101 (Suppl. 2), S207–S216.
Ghosh, S., Azhahianambi, P., de la Fuente, J., 2006. Control of ticks of ruminants, withspecial emphasis on livestock farming systems in India: present and future pos-sibilities for integrated control – a review. Experimental and Applied Acarology40, 49–66.
Gopalan, N., Prakash, S., Bhattacharya, B.K., Anand, O.P., Rao, K.M., 1996. Develop-ment of malathion resistance in Culex quinquefasciatus Say (Diptera, Culicidae).Indian Journal of Medical Research 103, 84–90.
Guerrero, F.D., Leonore, L., Martins, J.R., 2012. Acaricide resistance mechanisms inRhipicephalus (Boophilus) microplus. Revista Brasileira de Parasitologia Veter-inária 21, 1–6.
Guerrero, F.D., Li, A.Y., Hernandez, R., 2002. Molecular diagnosis of pyrethroid resis-tance in Mexican strains of Boophilus microplus (Acari: Ixodidae). Journal ofMedical Entomology 39, 770–776.
Guerrero, F.D., Jamroz, R.C., Kammlah, D., Kunz, S.E., 1997. Toxicological and molec-ular characterization of pyrethroid-resistant horn flies, Haematobia irritans:identification of kdr and super-kdr point mutations. Insect Biochemistry andMolecular Biology 27, 745–755.
He, H., Chen, A.C., Davey, R.B., Ivie, G.W., George, J.E., 1999. Identification of a pointmutation in the para-type sodium channel gene from a pyrethroid-resistantcattle tick. Biochemical and Biophysical Research Communications 261,558–561.
Hemingway, J., 1998. Techniques to Detect Insecticide Resistance Mechanisms (Fieldand Laboratory Manual). Document WHO/CDS/CPC/MAL/98.6, World HealthOrganization, Geneva.
Hemingway, J., Dunbar, S.J., Monro, A.G., Small, G.J., 1993. Pyrethroid resistance inGerman cockroaches (Dictyoptera: Blattelidae): resistance levels and underly-ing mechanisms. Journal of Economic Entomology 86, 1631–1638.
Jamroz, R.C., Guerrero, F.D., Pruett, J.H., Oehler, D.D., Miller, R.J., 2000. Molecular andbiochemical survey of acaricide resistance mechanisms in larvae from Mex-ican strains of the southern cattle tick, Boophilus microplus. Journal of InsectPhysiology 46, 685–695.
Jonsson, N.N., Cutullè, C., Corley, S.W., Seddon, J.M., 2010. Identification of a muta-tion in the para-sodium channel gene of the cattle tick Rhipicephalus (Boophilus)microplus associated with resistance to flumethrin but not to cypermethrin.International Journal for Parasitology 40, 1659–1664.
Kumar, R., Paul, S., Kumar, S., Sharma, A.K., Gupta, S., Rawat, A.K.S., Chaudhuri, P., Ray,D.D., Ghosh, S., 2011. Nucleotide specific changes in the hypervariable region of16S rDNA gene as possible marker to differentiate the tick genera. Indian Journalof Animal Sciences 81, 1204–1207.
Kumar, S., Paul, S., Sharma, A.K., Kumar, R., Tewari, S.S., Chaudhuri, P., Ray,D.D., Rawat, A.K.S., Ghosh, S., 2011. Diazinon resistant status in Rhipicephalus(Boophilus) microplus collected from different agro-climatic zones of India. Vet-erinary Parasitology 181, 274–281.
Lee, S.H., Dunn, J.B., Soderlund, J.M., Clark, D.M., 1999. Molecular analysis of kdr-likeresistance in a permethrin-resistant strain of Colorado potato beetle. PesticideBiochemistry and Physiology 63, 63–75.
Lee, S.H., Yoon, K.S., Williamson, M.S., Goodson, S.J., Takano-Lee, M., Devonshire,A.L., Clark, J.M., 2000. Molecular analysis of kdr-like resistance in permethrin-resistant strains of head lice Pediculus capitis. Pesticide Biochemistry andPhysiology 66, 130–143.
Li, A.Y., Chen, A.C., Miller, R.J., Davey, R.B., George, J.E., 2007. Acaricide resistance andsynergism between permethrin and amitraz against susceptible and resistantstrains of Boophilus microplus (Acari, Ixodidae). Pest Management Science 63,882–889.
Martin, R.L., Pittendrigh, B., Liu, J., Reenan, R., ffrench-Constant, R., Hanck, D.A., 2000.Point mutations in domain III of a Drosophila neuronal Na channel confer resis-tance to allethrin. Insect Biochemistry and Molecular Biology 30, 1051–1059.
Martinez-Torres, D., Foster, S.P., Field, L.M., Devonshire, A.L., Williamson, M.S.,1999a. A sodium channel point mutation is associated with resistance to DDTand pyrethroid insecticides in the peach-potato aphid, Myzus persicae (Sulzer)(Hemiptera: Aphididae). Insect Molecular Biology 8, 339–346.
Martinez-Torres, D., Chevillon, C., Brun-Barale, A., Bergé, J.B., Pasteur,.N., Pauron, D.,1999b. Voltage-dependent Na+ channels in pyrethroid-resistant Culex pipiensmosquitoes. Pesticide Science 55, 1012–1020.
Miller, R.J., Davey, R.B., George, J.E., 2007. First report of Permethrin resistantBoophilus microplus (Acari: Ixodidae) collected within the United States. Journalof Medical Entomology 44, 308–315.
Minjauw, B., Mc Leod, A., 2003. Research Report, DFID Animal Health Programme.Centre for Tropical Veterinary Medicine, University of Edinburgh, UK.
Morgan, J.A.T., Corley, S.W., Jackson, L.A., Lew-Tabor, A.E., Moolhuijzen, P.M., Jon-sson, N.N., 2009. Identification of point mutation in the para-sodium channelgene of the cattle Rhipicephalus (Boophilus) microplus associated with resistanceto synthetic pyrethroids acaricides. International Journal for Parasitology 39,775–779.
Morin, S., Williamson, M.S., Goodson, S.J., Brown, J.K., Tabashnik, B.E., Dennehy, T.J.,2002. Mutations in the Bemisia tabaci para sodium channel gene associated withresistance to a pyrethroid plus organophosphate mixture. Insect Biochemistryand Molecular Biology 32, 1781–1789.
Narahashi, T., 1988. Chemical modulation of sodium channels. In: Soria, B., Cena, V.(Eds.), Ion Channel Pharmacology. Oxford University Press, Oxford, pp. 23–73.
Nogueira Domingues, L., Dos Santos Alves Figueiredo Brasil, B., Passos de PaivaBello, A.C., Pinto da Cunha, A., Thadeu Medeiros de Barros, A., CerqueiraLeite, R., Silaghi, C., Pfister, K., Friche Passos, L.M., 2012. Survey of pyrethroidand organophosphate resistance in Brazilian field populations of Rhipicephalus(Boophilus) microplus: Detection of C190A mutation in domain II of the para-type sodium channel gene. Veterinary Parasitology http://dx.doi.org/10.1016/j.vetpar.2012.04.031
Park, Y., Taylor, M.F.J., 1997. A novel mutation L1029H in sodium channel genehscp associated with pyrethroid resistance for Heliothis virescens (Lepidoptera,Noctuidae). Insect Biochemistry and Molecular Biology 27, 9–13.
Pasay, C., Arlian, A., Morgam, M., Moher-Vyszenski, D., Rose, A., Holt, D., Walton, S.,McCarthy, J., 2008. High-resolution melt analysis for the detection of a mutationassociated with permethrin resistance in a population of scabies mites. Medicaland Veterinary Entomology 22, 82–88.
Pasay, C., Walton, S., Fischer, K., Holt, D., McCarthy, J., 2006. PCR-based assay tosurvey for knockdown resistance to pyrethroid acaricides in human scabiesmites (Sarcoptes scabiei var hominis). American Journal of Tropical Medicine andHygiene 74, 649–657.
Patarroyo, J.H., Costa, J.O., 1980. Susceptibility of Brazilian samples of Boophilusmicroplus to organophosphorus acaricides. Tropical Animal Health and Produc-tion 12, 6–10.
Pittendrigh, B., Reenan, R., Ffrench-Constant, R.R., Ganetzky, B., 1997. Point muta-tions in the Drosophila para sodium channel gene associated with resistanceto DDT and pyrethroid insecticides. Molecular and General Genetics 256,602–610.
Rodriguez-Vivas, R.I., Hodgkinson, J.E., Rosado-Aguilar, J.A., Villegas-Perez, S.L.,Trees, A.J., 2012. The prevalence of pyrethroid resistance phenotype and geno-type in Rhipicephalus (Boophilus) microplus in Yucatan, Mexico. VeterinaryParasitology 184, 221–229.
Rosario-Cruz, R., Almazan, C., Miller, R.J., Dominguez-Gracia, D.I., Hernandez-Ortiz,R., De la Fuente, J., 2009. Genetic basis and impact of tick acaricide resistance.Frontiers in Bioscience 14, 2657–2665.
Rosario-Cruz, R., Guerrero, F.D., Miller, R.J., Rodriguez-Vivas, R.I., Dominguez-Garcia,D.I., Cornel, A.J., Hernandez-Ortiz, R., George, J.E., 2005. Roles played by esteraseactivity and by a sodium channel mutation involved in pyrethroid resistancein populations of Boophilus microplus (Acari: Ixodidae) collected from Yucatan,Mexico. Journal of Medical Entomology 42, 1020–1025.
Rosario-Cruz, R., Miranda, M.E., Garcia, V.Z., Ortiz, E.M., 1997. Detection of esteraseactivity in susceptible and resistant Boophilus microplus tick strains. Bulletin ofEntomological Research 87, 197–202.
Sambrook, J., Russell, D.W., Irwin, N., Janseen, K.A., 2001. Molecular cloning-A labo-ratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NewYork, pp. 6.1–6.62.
Schuler, T.H., Martinez-Torres, D., Thompson, A.J., Devonshire, A.L., Duce, I.R.,Williamson, M.S., 1998. Toxicological, electrophysiological and molecular char-acterization of knockdown resistance to pyrethroid insecticides in the diamondblack moth, Plutella xylostella (L.). Pesticide Biochemistry and Physiology 59,169–182.
Sharma, A.K., Kumar, R., Kumar, S., Nagar, G., Singh, N.K., Rawat, S.S.,Dhakadd, M.L., Rawat, A.K.S., Ray, D.D., Ghosh, S., 2012. Deltamethrinand cypermethrin resistance status of Rhipicephalus (Boophilus) micropluscollected from six agro-climatic regions of India. Veterinary Parasitologyhttp://dx.doi.org/10.1016/j.vetpar.2012.03.050
Shidrawi, G.R., 1990. A WHO global programme for monitoring vector resistance topesticide. Bulletin of the World Health Organization 68, 403–408.
Shyma, K.P., Kumar, S., Sharma, A.K., Ray, D.D., Ghosh, S., 2012. Acaricide resistancestatus in Indian isolates of Hyalomma anatolicum. Experimental and AppliedAcarology, http://dx.doi.org/10.1007/s10493-012-9592-3.
Soderlund, D.M., 2008. Pyrethroids, knockdown resistance and sodium channels.Pest Management Science 64, 610–616.

Author's personal copy
R. Kumar et al. / Acta Tropica 125 (2013) 237– 245 245
Tsagkarakou, A., Van Leeuwen, T., Khajehali, J., Ilias, A., Grispou, M., Williamson, M.S.,Tirry, L., Vontas, J., 2009. Identification of pyrethroid resistance associated muta-tions in the para sodium channel of the two-spotted spider mite Tetranychusurticae (Acari, Trtranychidae). Insect Molecular Biology 18, 583–593.
Yanola, J., Somboon, P., Walton, C., Nachaiwieng, W., Prapanthadara, L., 2010. Anovel F1552/C1552 point mutation in the Aedes aegypti voltage-gated sodium
channel gene associated with permethrin resistance. Pesticide Biochemistry andPhysiology 96, 127–131.
Zhu, Y.C., Snodgrass, G.L., Chen, M.S., 2004. Enhanced esterase gene expres-sion and activity in a malathion-resistant strain of the tarnished plantbug, Lygus lineolaris. Insect Biochemistry and Molecular Biology 34,1175–1186.