successive sheep grazing reduces population density of
TRANSCRIPT

1 3
Oecologia (2016) 180:149–159DOI 10.1007/s00442-015-3455-7
POPULATION ECOLOGY - ORIGINAL RESEARCH
Successive sheep grazing reduces population density of Brandt’s voles in steppe grassland by altering food resources: a large manipulative experiment
Guoliang Li1,2 · Baofa Yin1,3 · Xinrong Wan1 · Wanhong Wei3 · Guiming Wang4 · Charles J. Krebs5 · Zhibin Zhang1
Received: 2 April 2015 / Accepted: 11 September 2015 / Published online: 7 October 2015 © Springer-Verlag Berlin Heidelberg 2015
Brandt’s voles along a gradient of three grazing intensities by using 12 large-scale enclosures. Responses of Brandt’s voles to livestock grazing varied with grazing intensity and year. As compared to the control group, sheep graz-ing had no effect on vole abundance in the first year but an overall negative effect on vole abundance in the following 3 years. Successive grazing caused decreases in survival and male body mass of voles, but had no significant effect on fecundity. Negative effects of grazing were associated with a grazing-induced deterioration in both food quantity (reflected by biomass and cover of less-preferred plants), and food quality (measured by tannin and total phenol con-tent). Our findings highlight the urgent need for more flex-ible management of yearly rotational grazing to optimize livestock production while maintaining species diversity and ecosystem health.
Keywords Biodiversity loss · Food quality · Food quantity · Pest management · Rotational grazing
Introduction
As human populations grow, anthropogenic disturbances are imposing increasing threats to biodiversity and eco-system functioning (Dirzo et al. 2014; Zhang et al. 2014). Livestock grazing has been recognized as one of the old-est and most geographically expansive forms of terrestrial anthropogenic activities (Diaz et al. 2007; Pielke et al. 2011), and has significantly shaped grassland ecosystems through both direct and indirect effects (Golluscio et al. 2009). Understanding of the effects of grazing on key spe-cies is urgently needed to address and thereby provide guidelines for managing grassland ecosystems under accel-erated human disturbances (Lindsey et al. 2013).
Abstract Livestock grazing has shaped grassland ecosys-tems around the world. Previous studies indicated grazing showed various impacts on small rodents; however, most studies were conducted over 1–2 years without controlling for confounding factors such as immigration/emigration and predation in rodents. Brandt’s voles (Lasiopodomys brandtii) are generally recognized as pests because of food overlap with domestic herbivores, but are also important for biodiversity conservation because they provide nests or food to many birds. Fully understanding the ecological relationship between domestic herbivores and small mam-mals is essential to making ecosystem management deci-sions. To address these needs, we carried out a field experi-ment during the period 2010–2013 to assess the effects of sheep grazing on vegetation and the population density of
Communicated by Janne Sundell.
Electronic supplementary material The online version of this article (doi:10.1007/s00442-015-3455-7) contains supplementary material, which is available to authorized users.
* Zhibin Zhang [email protected]
1 State Key Laboratory of Integrated Management of Pest Insects and Rodents in Agriculture, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
2 College of Life Science, University of Chinese Academy of Sciences, Beijing 100049, China
3 College of Bioscience and Biotechnology, Yangzhou University, Yangzhou 225009, China
4 Department of Wildlife, Fisheries and Aquaculture, Mississippi State University, Starkville, MS 39762, USA
5 Department of Zoology, University of British Columbia, Vancouver, BC V6T 1Z4, Canada

150 Oecologia (2016) 180:149–159
1 3
Small rodents play important roles in helping to main-tain biodiversity and functioning of grassland ecosystems (Davidson et al. 2012). They also play a significant role in promoting nutrient cycling of grassland ecosystems as con-sumers. However, they occasionally cause damage to grass-lands as pests when their populations become dense (Zhong et al. 1999). There is a need to find a trade-off between con-servation and control in managing small rodents in grass-land ecosystems.
Grazing has been recognized to influence populations of small rodents in grasslands, but responses of small rodents to grazing may vary significantly due to differences in spe-cies or systems. Some studies have demonstrated that graz-ing has negative effects on small rodents due to the deterio-ration of food quantity [i.e., a shortage of food resources caused by livestock as competitors (e.g., Milchunas et al. 1998; Steen et al. 2005)] or food quality [a grazing-induced increase of secondary chemicals (e.g., Bryant et al. 1985; Lindroth and Batzli 1984)]. Some studies indicated that grazing benefited small rodents from a reduction of vegeta-tion and the creation of preferred open habitats (Davidson et al. 2010) or stimulation of grass regrowth during the growing season (Arsenault and Owen-Smith 2002). Several other studies did not reveal any significant grazing effects (Saetnan et al. 2012).
Previous studies on this subject have three limitations. First, most of the previous studies were conducted in open grasslands, which allowed the confounding influence of immigration/emigration and predation in rodents. Second, the grazing experiments lasted only for 1–2 years (Foster et al. 2014); in such studies it is impossible to detect the differences between short-duration and extended-dura-tion effects of grazing. Finally, the underlying mecha-nisms which cause change are not fully understood due to a lack of manipulative experiments with information on both the demography of the rodents and on plant-animal interactions.
Brandt’s vole is a small herbivore and the predominant rodent species in the steppe grassland of Inner Mongolia. They are social animals, and their breeding season mainly lasts from May to August. Females give birth from one to three times each year, with each litter consisting of usu-ally two to 13 pups. Its population oscillates greatly among years (Jiang et al. 2011; Zhang et al. 2003b; Zhong et al. 1999), but the underlying mechanisms have not been tested with manipulative experiments. In this study, by the use of large replicated enclosures which excluded the confound-ing influences of immigration/emigration and predation, we investigated the effects of sheep grazing on the popu-lation dynamics and the underlying response mechanisms of Brandt’s vole populations in Inner Mongolia grasslands during the period 2010–2013. This is the first study which addresses the underlying demographic mechanisms for the
impacts of grazing on small mammal populations in the context of the plant-vole-sheep system.
Materials and methods
Study area
The study was conducted in the Maodeng pasture (44°11′N, 116°27′E; 1100 m in elevation; Fig. 1), 38 km northeast of Xilinhot, Inner Mongolia, China. Meteorological data gathered from 1999 to 2009 showed that the study site has a very cold winter (average daily maximum tempera-ture approximately −10 °C) and a warm summer (average daily maximum temperature approximately 26 °C). Mean annual precipitation at the study site was 241 mm, with 75 % occurring during the growing season (May–August). The study site is located in a semiarid typical steppe, domi-nated by Krylov’s needle grass (Stipa krylovii), Chinese lyme grass (Leymus chinensis), and scabrous Cleistogenes (Cleistogenes squarrosa). Brandt’s voles are the most abun-dant small herbivores in the grassland. The experimen-tal area had been grazed mainly by sheep until the start of 2008, and subsequently rested for 2 years prior to this study. The local stock rate at the study site was about six sheep ha−1 during the growing season.
Experimental design
We constructed a set of twenty-four 0.48-ha enclosures of 80 × 60 m with galvanized iron sheets extending 1 m underground and 1.4 m above ground to prevent move-ments of burrowing rodents, sheep, and carnivores into or out of enclosure (see Li et al. 2015). Each enclosures was covered completely by 50-cm-high wire netting (1-cm mesh size) on all four sides and by nylon netting (10-cm mesh size) on the top to exclude avian predators. These enclosures eliminated all emigration and immigration of rodents and excluded all their predators.
Twelve enclosures were randomly assigned into three treatments: control, light grazing (LG), moderate grazing (MG), with four replicates for each treatment (the other 12 enclosures were used for other study purposes; Fig. 1c). According to the local standard of sheep grazing intensity (Schonbach et al. 2011), the grazing intensity for the three treatments C, LG and MG was set at 0, 2, and 4 sheep ha−1, respectively. We did not set up a heavy grazing treatment to avoid serious vegetation deterioration. We grazed 40 adult sheep for a half day or a whole day biweekly during the growing season, which was equivalent to the grazing inten-sity of the LG group or the MG group, respectively.
Experimental voles were trapped in Abaga Qi (43°31′30″N, 116°04′50″E) or Dongwu Qi (45°33′39″N,

151Oecologia (2016) 180:149–159
1 3
116°37′48″E) of Xilingol League in the early spring (April) of the year of experiment and in autumn (September and October) of the previous year using live traps made of wire mesh [trap size, height × width × length = 10 × 10 × 24 cm; mesh size, 1 cm; Table S1, Electronic supplemen-tary material (ESM)]. Voles captured in autumn were raised in an open enclosure (480 × 300 m; near to experimental enclosures) as supplemental source populations. Founder individuals of experimental populations were mostly from spring-captured individuals; if the number of voles was not sufficient, individuals from the supplemental pool popu-lation were also used. In 2010, due to a shortage of field voles, we introduced five or six non-sibling laboratory-bred voles to each enclosure. During the period 2010–2013, thirteen to 15 pairs of adult voles were randomly assigned to each of the 12 enclosures in early spring, and allowed to adapt to the enclosure environments for 10–15 days before the grazing was initiated. We added new voles into enclosures to compensate for losses of voles due to mor-talities during the adaptation period and to have an aver-age founder population size of 13–15 pairs at the begin-ning for each year (see Table S1 of the ESM). The grazing began when the 10- to 15-day adaptation period finished. Voles in enclosures were allowed to survive to the follow-ing spring to determine impacts of grazing during the non-breeding season. All voles were removed from enclosures at the beginning of the ensuing spring so as to repeat the experiments the following year to evaluate the impacts of
successive grazing. This resetting of the experimental sys-tem was designed to minimize the differences of founder populations, and to test the successive effects of grazing on voles in multiple years. Studies on the carry-over effects of grazing on voles in 2 successive years started in 2014.
Animal trapping
Vole populations were monitored biweekly from May to October using mark-capture-recapture methods and each trapping session lasted for 2 successive days. We checked the traps three times during each trapping session. About 180 traps baited with peanuts were set near burrows in each enclosure. The voles were marked with a numbered metal ear tag at their first capture. The tag number, body mass, sex and reproductive condition of voles were recorded. After measurements, voles were immediately released at the capture site. In order to minimize the effects of the bait on the chemical composition of fecal samples, fresh sam-ples of feces were collected within 1 h after the traps were set. Diet compositions of voles and relative frequency (RF) indices of each plant species were estimated by identify-ing food items based on counting of 100 microscope fields of feces sampled in each enclosure. The RF index for each plant species was calculated following Gill et al. (1983):
RFi =Number of fields where species i is identified
Number of fields with fragments of any species× 100.
Fig. 1 Location of the study site in China (a), Brandt’s vole Lasiopodomys brandtii (b), enclosure layout from Google Earth in 2011 (c), a grazing enclosure (d)

152 Oecologia (2016) 180:149–159
1 3
Vegetation survey
Vegetation was sampled in the first week following a trap-ping session once each month. We evaluated changes in food quantity by surveying plant cover, biomass and den-sity. The cover (%), density (the number of plants rooted within each quadrat) and biomass (grams per square meter) of each plant species were measured in five randomly selected quadrats (1 × 1 m) within each enclosure. Subse-quently, each 1 × 1-m quadrat was further subdivided into 16 subquadrats (0.25 × 0.25 m), and aboveground mate-rial of each plant species in one of these subquadrats was clipped at ground level. Locations of clipped quadrats dif-fered in each sampling session to minimize the impact of clipping. Clipped plants were separated by species, stored temporarily in paper envelopes, and then dried in an oven at 65 °C to a constant mass. Plant samples were also ana-lyzed for food quality using the chemical and nutritional properties including crude protein, hydrolysable tannins (hereafter “tannins”) and total phenols. Crude protein con-tent was determined using the Kjeldahl method. Total phe-nols and tannins were measured by using a modified Folin-Ciocalteu method (Makkar et al. 1993). Plant samples were not taken within about 8 m of the enclosure wall to avoid the extra heat effect of galvanized iron sheet flashing on the vegetation.
The preference for different plant species by voles was evaluated by the relative preference index (RPI) (Vandyne and Heady 1965). RPI was calculated as the ratio of the percent of a plant species in the diet of voles from fecal analysis to the percent of the plant species present in the enclosure. If the mean RPI was greater than 1.0 and the 95 % confidence interval (CI) did not include 1.0, the par-ticular plant species was defined as a “preferred plant” for voles; otherwise, it was defined as a “less-preferred plant.”
Data analysis
The population density was first estimated by the minimum number of animals known alive (MNA) method (Krebs 1966). It is a reasonable index for estimating population size when the recapture rate is high (Hilborn et al. 1976; Ozgul et al. 2004). In our study, the average probability of recapture during these 4 years was 86.8 % (95 % CI 86–87.6 %). Thus, MNA was used as an index of popula-tion size of Brandt’s voles in the enclosures. The popula-tion density for each enclosure was also estimated by the jackknife estimator for model Mh (where h = heterogeneity in capture probabilities among animals) with the program CAPTURE (Otis et al. 1978; available at http://www.mbr-pwrc.usgs.gov/software.html). The population density of voles was defined as the number of voles per hectare. As suggested by Redfield et al. (1978), recruits were defined
simply as the number of any previously unmarked voles, regardless of age or body size of animals.
Significant differences in population parameters of voles (i.e., density, recruits, body mass) and parameters of plants (i.e., cover, biomass, density, crude protein content, tannin content and total phenol content) between control and graz-ing groups were tested by means of linear mixed models (LMM) and package nlme (Pinheiro et al. 2014) in R (R Core Team 2013). In the LMM analysis, a repeated covari-ance structure was used to take account of variables meas-ured over time. Enclosure and year were both specified as random effects, which allowed for correlated responses within years and enclosures. Factor enclosure was nested within factor year. Trapping session and treatment as well as their interaction were used as fixed effects in all anal-yses. Firstly, we used the data of the first year to detect the short-duration grazing effects; secondly, we used the pooled data from the 4 years to detect the extended-dura-tion grazing effects. We applied the function glht of the multcomp package to perform the multiple comparisons. Three enclosures [one for the control (B3), one for LG (B1) and one for MG (A1); Fig. 1] in 2010 were not included in the analysis due to their invasion by domestic cats. Data of male body mass were analyzed to avoid the effects of preg-nancy in females. In order to identify differences of total tannin or phenol content of the three preferred plant spe-cies in the diets of voles between the control and grazing groups, we weighted tannin or phenol content by the RF for each plant species in the fecal analysis of voles.
We estimated apparent survival (hereafter “survival”) and recapture probabilities with the classic Cormack-Jolly-Seber (CJS) models implemented in program MARK 7.1 (Lebreton et al. 1992; White and Burnham 1999). Instead of the “all possible models” approach (Anderson et al. 2001), we constructed a set of candidate models a priori, of which each represented survival as a function of the combination of covariates, including treatment group, time and sex. The CJS models assume statistical homoge-neity both on survival rates from time i to time i + 1 and on recapture rates at time i among different individuals. We estimated overdispersion in our trapping data using the median c (an estimate of the variance inflation factor) to scale the variance estimate. If the assumptions of the CJS model were met, then c = 1; otherwise c > 1. We used quasi Akaike’s information criterion corrected for small sample size (QAICc) and adjusted for overdispersion when c > 1 for model selection and statistical inference (Burnham and Anderson 2002).
Overwinter survival and density changes of voles from autumn (October) to the spring (April) were also estimated. Statistical differences in overwinter survivals and density changes between control and grazing groups were analyzed by one-way ANOVA followed by the post hoc multiple

153Oecologia (2016) 180:149–159
1 3
comparisons by Tukey’s honest significant difference with a significance level of P < 0.05. The CJS model could not estimate the overwinter survival since it is the last capture period, so overwinter survival per year was calculated as the proportion of animals captured during the last trap-ping session of the previous October that were recaptured in April of the current year, as suggested by Karels et al. (2000). October vole densities were also used as covari-ates (density dependence) when overwinter survival was analyzed. Due to an insufficient sample size of overwinter populations in 2011, the overwinter survival and density changes from autumn 2010 to spring 2011 could not be estimated.
The current study emphasizes comparisons between grazing and control groups in 4 years. Because of a limited sampling number of years, the yearly variations will be not analyzed and discussed.
Results
Population density
In the breeding season, data pooled from the 4 years showed that vole density was significantly affected by livestock grazing (F2, 39 = 4.2, P = 0.023) and trapping sessions (F10,
420 = 51.3, P < 0.001), with no interaction between trap-ping sessions × grazing (F20, 420 = 0.74, P = 0.79). Tuk-ey’s tests revealed that MG lowered vole density by 23 % on average compared to control vole density (t = −2.67, P = 0.02; Fig. 2); LG also marginally lowered vole den-sity by 21 % relative to the control group (t = −2.28, P = 0.058; Fig. 2). A separate analysis of the first-year data showed that grazing has no effect on vole density in 2010 (F2, 6 = 0.31, P = 0.75). Results using population density
estimated by using the jackknife estimator were similar to those determined by MNA (see Table S3 of the ESM). In non-breeding seasons, ANOVA results showed significant differences in overwinter density among control and graz-ing groups in 2013–2014 (F2,9 = 4.9, P = 0.036); the MG group had a 60 % lower overwinter density of voles than the control group (t = −2.97, P = 0.038; Fig. 3).
Survival
The median c-test significantly indicated a lack of fit or overdispersion in the general model (in 2010, c = 1.4; in 2011, c = 1.6; in 2012, c = 1.47; in 2013, c = 1.5); there-fore, the variance inflation factor was applied for all subse-quent model fitting.
In breeding seasons, model selection (Table 1) showed that the effect of sex on estimates of survival differed among years. Males had greater survival rates than females in 2010 (
∧
β = 0.21, 95 % CI 0.03–0.45) and in 2011 (∧
β = 0.12, 95 % CI 0.01–0.26). In 2012, the best-approx-imating model indicated there was no difference in survival between males and females (Table 1). In 2013, males had lower survival than females (
∧
β = −0.22, 95 % CI −0.41 to −0.03). Both light and MG significantly reduced the survival of voles in the breeding season in both 2012 (LG, ∧
β = −1.78, 95 % CI −2.8 to −0.76; MG, ∧
β = −2.93, 95 % CI −3.89 to −1.97) and 2013 (LG:
∧
β = −8.22, 95 % CI −12.62 to −3.82; MG,
∧
β = −9.84, 95 % CI −14.92 to −4.76), but no significant effects of grazing on survival
Fig. 2 Changes in population densities (mean ± SE, n = 4) of Brandt’s vole of control, light gazing (LG) and moderate grazing (MG) treatments. Note that the density scale varies in the different years
Fig. 3 Changes in overwinter densities in April and survival rates (nApril/nOctober) of control, LG and MG treatments. Data represent mean ± SE. Different letters indicate significant treatment effects. For abbreviations, see Fig. 2
154 Oecologia (2016) 180:149–159
1 3
rates were detected in 2010 and 2011. These results indi-cated that overall successive grazing decreased the survival rate of voles.
In the non-breeding season, ANOVA results showed no significant differences in overwinter survival among the control vs. grazing groups for any year (Fig. 3).
Recruitment
Pooled data showed that there were no significant effects of grazing (F2, 39 = 0.31, P = 0.74) and interaction between grazing and trapping session (F20, 420 = 0.59, P = 0.92) on recruits. Trapping session had significant effects on recruits
(F10, 420 = 50.2, P < 0.001). Separate analysis of the first year also showed that grazing had no effect on the number of recruits (F2, 6 = 0.26, P = 0.78).
Body mass
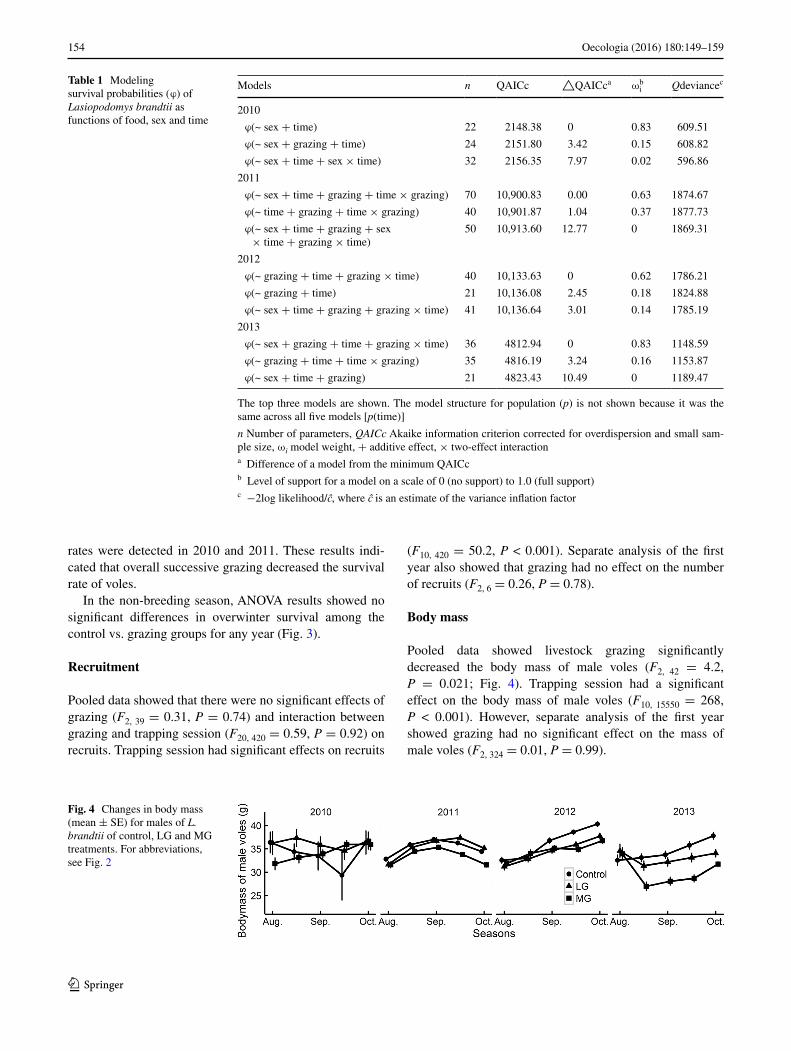
Pooled data showed livestock grazing significantly decreased the body mass of male voles (F2, 42 = 4.2, P = 0.021; Fig. 4). Trapping session had a significant effect on the body mass of male voles (F10, 15550 = 268, P < 0.001). However, separate analysis of the first year showed grazing had no significant effect on the mass of male voles (F2, 324 = 0.01, P = 0.99).
Table 1 Modeling survival probabilities (ϕ) of Lasiopodomys brandtii as functions of food, sex and time
The top three models are shown. The model structure for population (p) is not shown because it was the same across all five models [p(time)]
n Number of parameters, QAICc Akaike information criterion corrected for overdispersion and small sam-ple size, ωi model weight, + additive effect, × two-effect interaction a Difference of a model from the minimum QAICcb Level of support for a model on a scale of 0 (no support) to 1.0 (full support)c −2log likelihood/c, where c is an estimate of the variance inflation factor
Models n QAICc △QAICca ωib Qdeviancec
2010
ϕ(~ sex + time) 22 2148.38 0 0.83 609.51
ϕ(~ sex + grazing + time) 24 2151.80 3.42 0.15 608.82
ϕ(~ sex + time + sex × time) 32 2156.35 7.97 0.02 596.86
2011
ϕ(~ sex + time + grazing + time × grazing) 70 10,900.83 0.00 0.63 1874.67
ϕ(~ time + grazing + time × grazing) 40 10,901.87 1.04 0.37 1877.73
ϕ(~ sex + time + grazing + sex × time + grazing × time)
50 10,913.60 12.77 0 1869.31
2012
ϕ(~ grazing + time + grazing × time) 40 10,133.63 0 0.62 1786.21
ϕ(~ grazing + time) 21 10,136.08 2.45 0.18 1824.88
ϕ(~ sex + time + grazing + grazing × time) 41 10,136.64 3.01 0.14 1785.19
2013
ϕ(~ sex + grazing + time + grazing × time) 36 4812.94 0 0.83 1148.59
ϕ(~ grazing + time + time × grazing) 35 4816.19 3.24 0.16 1153.87
ϕ(~ sex + time + grazing) 21 4823.43 10.49 0 1189.47
Fig. 4 Changes in body mass (mean ± SE) for males of L. brandtii of control, LG and MG treatments. For abbreviations, see Fig. 2

155Oecologia (2016) 180:149–159
1 3
Food quantity
Over the 4 years of study the most preferred plant species for Brandt’s voles in our enclosures were Leymus chinensis, Setaria viridis and Medicago sativa, and the less-preferred plant species were most commonly Stipa krylovii, Cleis-togenes squarrosa, and Phlomis dentosa (Table S2, ESM).
For preferred plant species, pooled data showed that there was no significant differences in plant cover (F2, 42 = 0.28, P = 0.76), plant biomass (F2, 42 = 0.56,
P = 0.57; Fig. 5) or plant density (F2, 42 = 1.48, P = 0.24) between control and grazing groups. Separate analysis of the first year also showed that grazing exerted no signifi-cant effect on plant cover (F2, 9 = 0.3, P = 0.75), plant bio-mass (F2, 9 = 0.29, P = 0.75) or plant density (F2, 9 = 0.33, P = 0.73).
However, for less-preferred species, pooled data showed that grazing significantly decreased plant cover (F2,
42 = 20.6, P < 0.001) and plant biomass (F2, 42 = 8.98, P < 0.001; Fig. 6). Tukey’s test showed that plant cover and/
Fig. 5 Changes in vegetation quantity (cover, biomass and density) of plants preferred by L. brandtii in control, LG and MG treatments. Data represent mean ± SE. For abbreviations, see Fig. 2
Fig. 6 Changes in vegetation quantity (cover, biomass and density) of plants less preferred by L. brandtii in control, LG and MG treatments. Data repre-sent mean ± SE. For abbrevia-tions, see Fig. 2
156 Oecologia (2016) 180:149–159
1 3
or plant biomass were lower in LG (13 and 12 % decreases, respectively) and MG (26 and 28 % decreases, respectively) than those in the control group. Grazing showed no sig-nificant effects on plant density (F2, 42 = 0.72, P = 0.49). Separate analysis of the first year showed grazing exerted no significant effect on plant cover (F2, 9 = 2.75, P = 0.12), plant biomass (F2, 9 = 1.7, P = 0.24) or plant density (F2,
9 = 0.77, P = 0.5) for less-preferred species.
Food quality
The grass species L. chinensis, S. krylovii and C. squarrosa were common food for Brandt’s vole. Based on pooled data from 4 years of study, their proportions in the dietary composition of Brandt’s vole were 42.5, 36.8, and 16.6 %, respectively.
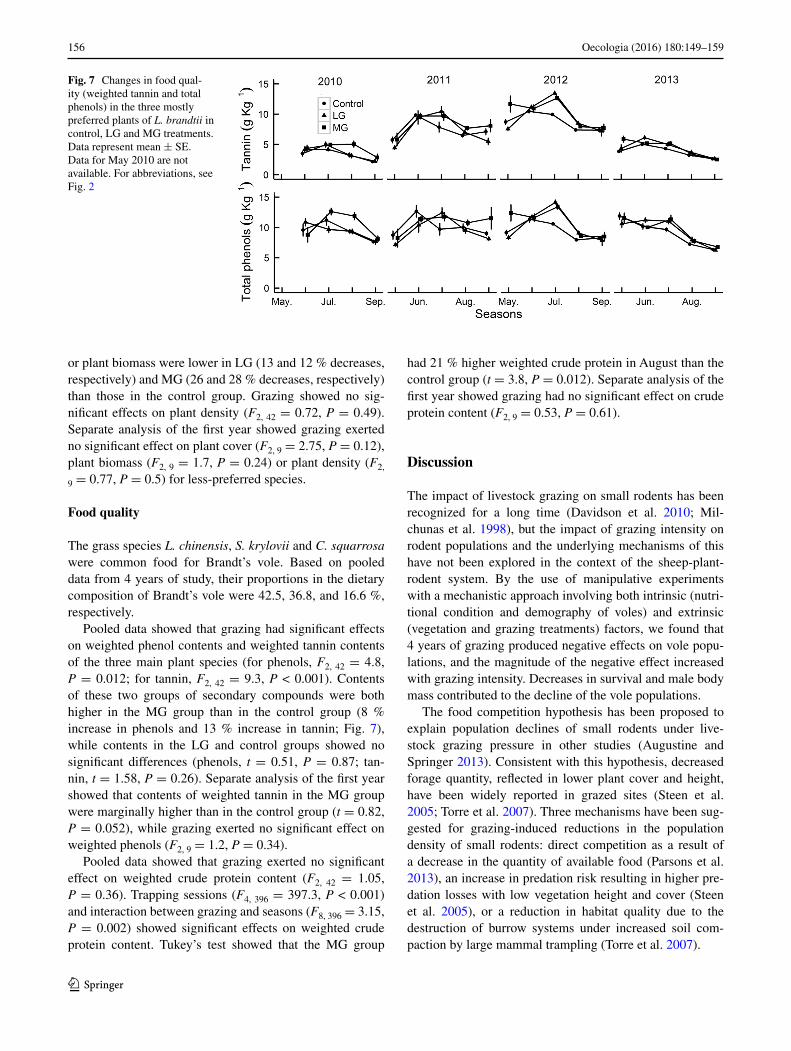
Pooled data showed that grazing had significant effects on weighted phenol contents and weighted tannin contents of the three main plant species (for phenols, F2, 42 = 4.8, P = 0.012; for tannin, F2, 42 = 9.3, P < 0.001). Contents of these two groups of secondary compounds were both higher in the MG group than in the control group (8 % increase in phenols and 13 % increase in tannin; Fig. 7), while contents in the LG and control groups showed no significant differences (phenols, t = 0.51, P = 0.87; tan-nin, t = 1.58, P = 0.26). Separate analysis of the first year showed that contents of weighted tannin in the MG group were marginally higher than in the control group (t = 0.82, P = 0.052), while grazing exerted no significant effect on weighted phenols (F2, 9 = 1.2, P = 0.34).
Pooled data showed that grazing exerted no significant effect on weighted crude protein content (F2, 42 = 1.05, P = 0.36). Trapping sessions (F4, 396 = 397.3, P < 0.001) and interaction between grazing and seasons (F8, 396 = 3.15, P = 0.002) showed significant effects on weighted crude protein content. Tukey’s test showed that the MG group
had 21 % higher weighted crude protein in August than the control group (t = 3.8, P = 0.012). Separate analysis of the first year showed grazing had no significant effect on crude protein content (F2, 9 = 0.53, P = 0.61).
Discussion
The impact of livestock grazing on small rodents has been recognized for a long time (Davidson et al. 2010; Mil-chunas et al. 1998), but the impact of grazing intensity on rodent populations and the underlying mechanisms of this have not been explored in the context of the sheep-plant-rodent system. By the use of manipulative experiments with a mechanistic approach involving both intrinsic (nutri-tional condition and demography of voles) and extrinsic (vegetation and grazing treatments) factors, we found that 4 years of grazing produced negative effects on vole popu-lations, and the magnitude of the negative effect increased with grazing intensity. Decreases in survival and male body mass contributed to the decline of the vole populations.
The food competition hypothesis has been proposed to explain population declines of small rodents under live-stock grazing pressure in other studies (Augustine and Springer 2013). Consistent with this hypothesis, decreased forage quantity, reflected in lower plant cover and height, have been widely reported in grazed sites (Steen et al. 2005; Torre et al. 2007). Three mechanisms have been sug-gested for grazing-induced reductions in the population density of small rodents: direct competition as a result of a decrease in the quantity of available food (Parsons et al. 2013), an increase in predation risk resulting in higher pre-dation losses with low vegetation height and cover (Steen et al. 2005), or a reduction in habitat quality due to the destruction of burrow systems under increased soil com-paction by large mammal trampling (Torre et al. 2007).
Fig. 7 Changes in food qual-ity (weighted tannin and total phenols) in the three mostly preferred plants of L. brandtii in control, LG and MG treatments. Data represent mean ± SE. Data for May 2010 are not available. For abbreviations, see Fig. 2

157Oecologia (2016) 180:149–159
1 3
In this study, continuous herbivory by voles sharply decreased the total biomass and cover of preferred plants in the enclosure during the 4-year study. The average ratio of non-preferred plants to preferred plants increased year by year (e.g., for biomass, 2.8 in 2010, 13.5 in 2013; for cover, 3 in 2010, 13.9 in 2013), which resulted in the less-preferred plants becoming the main supply of food for voles (Fig. S1, ESM). We found the negative effects of suc-cessive grazing were closely linked to a significant reduc-tion in cover and biomass of less-preferred plant species in grazing enclosures, which resulted in a shortage of food for Brandt’s voles. Dietary overlap was great between Brandt’s vole and sheep, the grass species L. chinensis the main food for voles in summer was also preferred by sheep. There-fore, the disappearance of preferred plants enhanced com-petition between voles and sheep. This explains why the grazing effect in the first year was not significant because voles had an excess of preferred plants to eat. This observa-tion supports the food competition hypothesis (Steen et al. 2005; Torre et al. 2007).
Grazing may change plant quality (crude protein, sec-ondary compounds), and subsequently may affect popula-tion abundance of other herbivores in grasslands (Augus-tine and Springer 2013; Lindroth and Batzli 1986). Grazing-induced secondary compounds, including tannins and proteinase inhibitors, have been shown to have nega-tive effects on small rodents in laboratory conditions (Lin et al. 2012; Lindroth and Batzli 1984), but there are few field studies of such effects (but see Saetnan et al. 2012). Repeated grazing is often necessary for maintaining high levels of secondary compounds (Gustafson and Ryan 1976), because the responses of food quality to grazing may be delayed (Saetnan et al. 2012). The response of for-age nitrogen content and digestibility in plants to grazing may be positive (Augustine and Springer 2013) or neutral (Saetnan et al. 2012). In our study, MG in August showed a positive effect on crude protein. There is strong evidence in our study that grazing induced higher secondary compound contents (e.g., tannin and total phenols). Thus, grazing-induced deterioration in food quality may be treated as an additional explanation for the observation of the reduced abundance of voles.
In our study, we confirmed that grazing intensity plays a key role in explaining the mechanisms of the response of voles to livestock grazing. LG with an extended duration (4 years) would impose a negative effect only by decreas-ing the food quantity (e.g., cover and biomass), while MG with an extended duration would reduce the vole popula-tion both by decreasing the food quantity and by decreas-ing the food quality (e.g., secondary compounds). The reduction in vole abundance caused by LG was slightly lower than that of MG. Long-time high grazing intensity could lead to a decline in the population of native keystone
species by modifying plants both in their community struc-ture and nutritional characteristics; this would cause a loss in the critical ecosystem functioning in which animals play a central role.
Overwinter survival is a critical demographic parameter for voles living in the northern hemisphere and has attracted extensive attention by field ecologists (e.g., Haapakoski et al. 2012; Karels et al. 2000). In our study, we found evi-dence of a lower overwinter density in years of successive grazing, which may have affected the population growth rate in the following years. In this study, we only focused on testing the effects of successive grazing on voles within four breeding seasons. Future studies should focus on test-ing the impact of grazing on small rodents across years.
The yearly variation in our vole populations might be caused by variations in weather in different years. As com-pared to 2011 and 2012, the low density of vole popula-tions in 2010 and 2013 was linked to low values of spring precipitation and temperature, which do not favor vole breeding (Xie et al. 2012). However, due to the limited data of 4 years, we are not able to draw reliable conclusions about the effects of climate on vole populations in the long term. Long-term experiments are needed to reveal the tem-poral effects of climate on vole populations in our enclo-sures (Evans et al. 2015; Krebs 2015).
Some small rodents play a significant role in perform-ing essential ecosystem functions and have been perceived as keystone species or ecosystem engineers in their native ranges (Davidson et al. 2012; Krebs 2014). Local extinc-tion of these native species (e.g., pika on the Qinghai-Tibet plateau) under accelerated human disturbance or poison-ing may damage ecosystem functions (Zhang et al. 2003a). Brandt’s vole is an important prey for some native apex predators (e.g., mustelids, canids and raptors). Its burrows are also ideal habitat for other rodents, herpetofauna and arthropods. Thus this species is very important in maintain-ing biodiversity and ecosystem functioning. Voles could occasionally damage pastures during their outbreak years (Zhong et al. 1999). Our study suggests that the short dura-tion of the LG treatment had little effect on voles, but the long duration of the MG caused population declines. Tra-ditionally, grazing has been sustained without or with an infrequent short-duration rotation by local herders, which leads to biodiversity loss, and thereby threatens ecosystem services. To achieve an ecological balance for managing Brandt’s vole, it is necessary to adjust the traditional graz-ing strategy. At present, moderate or heavy grazing without yearly rotation is widely adopted by local herdsmen. Based on our study, we recommend that in years without obvious vole damage to grassland, low-intensity and long-dura-tion grazing rotations should be adopted within the next 2–3 years to conserve the native species, while in years or regions with obvious vole damages, high-intensity and

158 Oecologia (2016) 180:149–159
1 3
short-duration grazing rotations within the next 3–4 years are recommended to reduce vole populations.
Acknowledgments We thank the Maodeng Pasture of Xilinhot City and the Grassland Station of Xilingol League for their kind help in our field experiments. We thank all the students and volunteers involved in the field work. We are grateful to Prof. Marcel Holyoak of the University of California Davis for his valuable assistance and comments on this manuscript. This study was supported by the State Basic Research Program of the Ministry of Science and Technology (2007CB109100) and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB11050000).
Author contribution statement Z. Z. designed the experiments. G. L., B. Y., X. W. and W. W. performed the experiments. G. L. analyzed the data. G. L. and Z. Z. wrote the manuscript; G. W. and C. J. K. contributed to the data analysis and manuscript writing.
References
Anderson DR, Burnham KP, Gould WR, Cherry S (2001) Concerns about finding effects that are actually spurious. Wildlife Soc B 29:311–316
Arsenault R, Owen-Smith N (2002) Facilitation versus competi-tion in grazing herbivore assemblages. Oikos 97:313–318. doi:10.1034/j.1600-0706.2002.970301.x
Augustine DJ, Springer TL (2013) Competition and facilitation between a native and a domestic herbivore: trade-offs between forage quantity and quality. Ecol Appl 23:850–863
Bryant JP, Wieland GD, Clausen T, Kuropat P (1985) Interactions of snowshoe hare and feltleaf willow in Alaska. Ecology 66:1564–1573. doi:10.2307/1938018
Burnham KP, Anderson DR (2002) Model selection and multimodal inference: a practical information theoretic approach. Springer, New York
Davidson AD et al (2010) Rapid response of a grassland ecosystem to an experimental manipulation of a keystone rodent and domestic livestock. Ecology 91:3189–3200. doi:10.1890/09-1277.1
Davidson AD, Detling JK, Brown JH (2012) Ecological roles and conservation challenges of social, burrowing, herbivorous mam-mals in the world’s grasslands. Front Ecol Environ 10:477–486. doi:10.1890/110054
Diaz S et al (2007) Plant trait responses to grazing—a global synthesis. Global Change Biol 13:313–341. doi:10.1111/j.1365-2486.2006.01288.x
Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B (2014) Defaunation in the Anthropocene. Science 345:401–406. doi:10.1126/science.1251817
Evans DM, Villar N, Littlewood N, Pakeman R, Evans S, Dennis P, Skartveit J, Redpath S (2015) The cascading impacts of livestock grazing in upland ecosystems: a 10 year experiment. Ecosphere. doi:10.1890/ES14-00316.1
Foster CN, Barton PS, Lindenmayer DB (2014) Effects of large native herbivores on other animals. J Appl Ecol 51:929–938. doi:10.1111/1365-2664.12268
Gill RB, Carpenter LH, Bartmann RM, Baker DL, Schoonveld GG (1983) Fecal analysis to estimate mule deer diets. J Wildlife Manage 47:902–915. doi:10.2307/3808149
Golluscio RA, Austin AT, Martinez GCG, Gonzalez-Polo M, Sala OE, Jackson RB (2009) Sheep grazing decreases organic car-bon and nitrogen pools in the Patagonian Steppe: combination of direct and indirect effects. Ecosystems 12:686–697. doi:10.1007/s10021-009-9252-6
Gustafson G, Ryan CA (1976) Specificity of protein turnover in tomato leaves: accumulation of proteinase-inhibitors, induced with wound hormone, Piif. J Biol Chem 251:7004–7010
Haapakoski M, Sundell J, Ylonen H (2012) Predation risk and food: opposite effects on overwintering survival and onset of breeding in a boreal rodent. J Anim Ecol 81:1183–1192. doi:10.1111/j.1365-2656.2012.02005.x
Hilborn R, Redfield JA, Krebs CJ (1976) Reliability of enumeration for mark and recapture census of voles. Can J Zool 54:1019–1024. doi:10.1139/Z76-114
Jiang G et al (2011) Effects of ENSO-linked clilmate and vegetation on population dynamics of sympatric rodent species in semiarid grasslands of Inner Mongolia, China. Can J Zool 89:678–691. doi:10.1139/Z11-048
Karels TJ, Byrom AE, Boonstra R, Krebs CJ (2000) The interactive effects of food and predators on reproduction and overwinter survival of arctic ground squirrels. J Anim Ecol 69:235–247. doi:10.1046/j.1365-2656.2000.00387.x
Krebs CJ (1966) Demographic changes in fluctuating populations of Microtus californicus. Ecol Monogr 36:240–273
Krebs CJ (2014) Rodent biology and management. Integr Zool 9:229–230. doi:10.1111/1749-4877.12090
Krebs CJ (2015) One hundred years of population ecology: suc-cesses, failures and the road ahead. Integr Zool 10:233–240. doi:10.1111/1749-4877.12130
Lebreton JD, Burnham KP, Clobert J, Anderson DR (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecol Monogr 62:67–118. doi:10.2307/2937171
Li GL, Hou XL, Wan XR, Zhang ZB (2015) Sheep grazing causes shift in sex ratio and cohort structure of Brandt’s vole: impli-cation of their adaptation to food shortage. Integr Zool. doi:10.1111/1749-4877.12163
Lin LJ et al (2012) Growth of sheep as affected by grazing system and grazing intensity in the steppe of Inner Mongolia, China. Livest Sci 144:140–147. doi:10.1016/j.livsci.2011.11.008
Lindroth RL, Batzli GO (1984) Plant phenolics as chemical defenses: effects of natural phenolics on survival and growth of prai-rie voles (Microtus Ochrogaster). J Chem Ecol 10:229–244. doi:10.1007/Bf00987851
Lindroth RL, Batzli GO (1986) Inducible plant-chemical defenses: a cause of vole population cycles. J Anim Ecol 55:431–449. doi:10.2307/4729
Lindsey HA, Gallie J, Taylor S, Kerr B (2013) Evolutionary rescue from extinction is contingent on a lower rate of environmental change. Nature 494:463–467. doi:10.1038/Nature11879
Makkar HPS, Blummel M, Borowy NK, Becker K (1993) Gravimet-ric determination of tannins and their correlations with chemical and protein precipitation methods. J Sci Food Agr 61:161–165. doi:10.1002/jsfa.2740610205
Milchunas DG, Lauenroth WK, Burke IC (1998) Livestock grazing: animal and plant biodiversity of shortgrass steppe and the relation-ship to ecosystem function. Oikos 83:65–74. doi:10.2307/3546547
Otis DL, Burnham KP, White GC, Anderson DR (1978) Statistical inference from capture data on closed animal populations. Wild-life Monogr 62:7–135
Ozgul A, Getz LL, Oli MK (2004) Demography of fluctuat-ing populations: temporal and phase-related changes in vital rates of Microtus ochrogaster. J Anim Ecol 73:201–215. doi:10.1111/j.0021-8790.2004.00797.x
Parsons EWR, Maron JL, Martin TE (2013) Elk herbivory alters small mammal assemblages in high-elevation drainages. J Anim Ecol 82:459–467. doi:10.1111/1365-2656.12009
Pielke RA et al (2011) Land use/land cover changes and climate: modeling analysis and observational evidence. Wires Clim Change 2:828–850. doi:10.1002/Wcc.144

159Oecologia (2016) 180:149–159
1 3
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2014) nlme: linear and nonlinear mixed effects models. R package version 3. 1–117. http://CRAN.R-project.org/package=nlme
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0, URL http://www.R-project.org/
Redfield JA, Taitt MJ, Krebs CJ (1978) Experimental alteration of sex ratios in populations of Microtus townsendii, a field vole. Can J Zool 56:17–27. doi:10.1139/Z78-003
Saetnan ER, Skarpe C, Batzli GO (2012) Do sheep affect vole populations in alpine meadows of central Norway? J Mammal 93:1283–1291. doi:10.1644/11-Mamm-a-226.1
Schonbach P et al (2011) Grassland responses to grazing: effects of grazing intensity and management system in an Inner Mongo-lian steppe ecosystem. Plant Soil 340:103–115. doi:10.1007/s11104-010-0366-6
Steen H, Mysterud A, Austrheim G (2005) Sheep grazing and rodent populations: evidence of negative interactions from a land-scape scale experiment. Oecologia 143:357–364. doi:10.1007/s00442-004-1792-z
Torre I, Diaz M, Martinez-Padilla J, Bonal R, Vinuela J, Fargallo JA (2007) Cattle grazing, raptor abundance and small mam-mal communities in Mediterranean grasslands. Basic Appl Ecol 8:565–575. doi:10.1016/j.baae.2006.09.016
Vandyne GM, Heady HF (1965) Botanical composition of sheep and cattle diets on a mature annual range. Hilgardia 36:465–492
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:120–139
Xie XH, Wen YL, Niu HX, Shi DZ, Zhang ZB (2012) Re-feed-ing evokes reproductive overcompensation of food-restricted Brandt’s voles. Physiol Behav 105:653–660. doi:10.1016/j.physbeh.2011.09.026
Zhang YM, Zhang ZB, Liu JK (2003a) Burrowing rodents as ecosystem engineers: the ecology and management of pla-teau zokors Myospalax fontanierii in alpine meadow eco-systems on the Tibetan Plateau. Mammal Rev 33:284–294. doi:10.1046/j.1365-2907.2003.00020.x
Zhang ZB, Pech R, Davis S, Shi DZ, Wan XR, Zhong WQ (2003b) Extrinsic and intrinsic factors determine the eruptive dynamics of Brandt’s voles Microtus brandti in Inner Mongolia, China. Oikos 100:299–310. doi:10.1034/j.1600-0706.2003.11810.x
Zhang MW et al (2014) Small mammal community succession on the beach of Dongting Lake, China after the Three Gorges Project. Integr Zool 9:294–308. doi:10.1111/1749-4877.12073
Zhong WQ, Wang MJ, Wan XR (1999) Ecological management of Brandt’s vole (Microtus brandti) in Inner Mongolia, China. In: Singleton GRHL, Leirs H, Zhang Z (eds) Ecologically-based management of rodent pests. ACIAR, Canberra, pp 199–214