subchronic oral toxicity of microcystin in common carp ( cyprinus carpio l.) exposed to microcystis...
TRANSCRIPT

Subchronic oral toxicity of microcystin in common carp (Cyprinus
carpio L.) exposed to Microcystis under laboratory conditions
Xiao-Yu Lib, Ik-Kyo Chungc, Jung-In Kimd, Jin-Ae Leea,*
aSchool of Environmental Science and Engineering, Inje University, Kimhae 621-749, South KoreabSchool of Life Science, Henan Normal University, XinXiang 453002, People’s Republic of China
cDepartment of Marine Science, Pusan National University, Busan 609-735, South KoreadSchool of Food and Life Science, Biohealth Products Research Center, Inje Univeristy, Kimhae 621-749, South Korea
Received 25 March 2004; accepted 17 June 2004
Abstract
The subchronic oral toxicity of microcystin in common carp (Cyprinus carpio L.) was investigated in this study. The fish
(mean body weight of 322G36 g, nZ10) were orally exposed to Microcystis by feeding with bloom scum at a dose of 50 mg
microcystins/kg body weight under laboratory conditions for 28 days. Growth assay results showed that microcystin could
completely inhibit the growth of carp, but failed to change the fish hepatosomatic index. Ultrastructural examination by electron
microscope revealed severe damage in hepatocytes derived from the treated fish. Serum biochemical assays with commercial
kits indicated that alanine aminotransferase and aspartate aminotransferase activities were significantly increased as compared
to control levels, but g-glutamyl transferase, alkaline phosphatase and lactate dehydrogenase activities remained unchanged.
Protein phosphatase inhibition assay revealed that the microcystin concentrations were 261.0G108.3 ng microcystin-LR
equivalent/g fresh weight in hepatopancreas and 38.3G12.3 ng microcystin-LR equivalent/g fresh weight in muscle. The latter
is above the limit recommended by the World Health Organization for human consumption. Therefore, we recommend that a
warning system be instituted for announcing the occurrence of microcystin-producing water bloom and the possible risk of
human intoxication.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: Microcystis bloom; Common carp; Subchronic toxicity of microcystin; Serum enzyme; Bioaccumulation
1. Introduction
Microcystis bloom occurs frequently in bodies of
freshwater all over the world (Codd et al., 1999). The
bloom not only deceases water quality, but also increases
the risk of toxicity to both animals and humans, because
many Microcystis species (mainly Microcystis aeruginosa)
can produce highly toxic microcystin (MCYST) (Toranzo
et al., 1990; Jochimsen et al., 1998; Falconer, 1999;
0041-0101/$ - see front matter q 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.toxicon.2004.06.010
* Corresponding author. Tel.: C82-55-320-3248; fax: C82-55-
334-7902.
E-mail address: [email protected] (J.-A. Lee).
Zimba et al., 2001). MCYST is a cyclic peptide hepatotoxin,
and acute dosage by intraperitoneal (i.p.) injection can kill
animals (Kotak et al., 1996; Gupta et al., 2003). Toxicolo-
gically, it appears that MCYST causes damage when it is
taken up by hepatocytes and inhibits serine/threonine
protein phosphatases 1 and 2A (Eriksson et al., 1990;
Yoshizawa et al., 1990; Runnegar et al., 1993). The resulting
imbalance in protein phosphorylation disrupts liver cytos-
keleton, which leads to massive hepatic haemorrhages that
cause death in rodents (Dawson, 1998).
MCYST-induced liver damage has been correlated with
the activity of certain plasma enzymes, such as g-glutamyl
transferase (GGT), alanine aminotransferase (ALT), aspar-
tate aminotransferase (AST), alkaline phosphatase (ALP)
Toxicon 44 (2004) 821–827
www.elsevier.com/locate/toxicon

X.-Y. Li et al. / Toxicon 44 (2004) 821–827822
and lactate dehydrogenase (LDH) (Falconer et al., 1994;
Fischer and Dietrich, 2000; Malbrouck et al., 2003). The
bioaccumulation of MCYST in fish tissues has been
reported (Eriksson et al., 1986; Magalhaes et al., 2001,
2003; Mohamed et al., 2003). This could lead to the transfer
of the toxin to human upon consumption of contaminated
fish, thereby affecting human health. Until now, there have
been no studies on subchronic oral exposure of carp to
microcystin under laboratory conditions.
The Naktong river is the second largest river in South
Korea, and it is an important resource for agriculture,
industrial development, and drinking water. Since 1993,
heavy cyanobacterial blooms have been observed in this
river, and a predominant species in the bloom material was
identified as toxic Microcystis aeruginosa (Lee et al., 1997).
Thus, there is much concern about using the Naktong river
as a source of drinking water and the possible effects of this
water on human health.
Common carp is a dominant species in the Naktong river
and often captured by the local fishermen for food. Because
the carp is omnivore, Microcystis can comprise a significant
portion of its diet, which might lead to MCYST accumu-
lation in its tissues during the bloom periods. Therefore, it is
important to evaluate MCYST toxicity to fish as well as
the toxic impact of Microcystis bloom on both fish and
human health.
This study aimed to determine the subchronic toxicity of
MCYST in common carp by examining ultrastructural
alternations in hepatocytes and changes in the activity of
various plasma enzymes. We also verified the presence of
MCYST in carp tissues, to evaluate the potential risks for
human health if MCYST-contaminated carp from the
Naktong river are consumed.
2. Materials and methods
2.1. Chemicals
MC-LR, -RR and -YR (purityO95%) were purchased
from Wako (Wako Chemical Co., Japan). Protein phospha-
tase 2A (PP 2A) and a commercial PP assay kit was obtained
from Promega (Promega, USA). Bovine serum albumin
(BSA), glutaraldehyde, EGTA, EDTA, and all other
reagents used were of the highest purity available and
were purchased from Sigma (Sigma, USA). Serum enzyme
assay kits were obtained commercially from ASAN (ASAN,
Korea).
2.2. Bloom scum collection and HPLC analysis
Microcystis bloom scum was collected from the Naktong
river using a phytoplankton net (25-mm diameter mesh) and
stored at K70 8C until used for fish feeding. Microcystis and
other associated algal species were identified as described
by Lee et al. (1997) and counted using a hemocytometer.
Extraction of MCYST from bloom scum and quantitative
HPLC analysis of MCYST levels within the scum was
performed according to method of Harada et al. (1988), with
minor modifications.
2.3. Fish
Common carp (mean body weight 322G36 g) was
kindly provided by Jinhae Inland Fisheries Research
Institute, National Fishery Research and Development
Institute, Korea. The fish were divided into two 400-L
tanks containing drinking water (one tank received treat-
ment and one tank served as a control). The fish were fed
adlibitum with commercial carp food at a rate of 1.5% of
body weight. Water temperature was maintained at 20G1 8C and dissolved oxygen values were maintained between
7.1 and 7.6 mg lK1. Fish were exposed to a 12-h light/12-h
dark photoperiod, and the tank water was changed weekly.
2.4. Exposure and sampling
After acclimatization for 2 weeks, fish in the treatment
tank (nZ10) were exposed orally to Microcystis by feeding
with bloom scum. The scum was mixed in the commercial
carp food powder and made into small sticky pellets. The
pellets were put into tank, and they fell down on the bottom
of tank for fish to take. It was ensured that fish should eat up
all pellets within 1 h. A low dose of 50 mg toxin/kg body wt,
about one tenth of the oral LD50 of MCYST in carp, was
adopted for subchronic exposure. According to this
exposure dose and the toxin concentration of bloom scum
(determined from HPLC analysis), we determined the
quantity of bloom scum that was necessary to add to carp
food. Control fish (nZ10) was fed only the commercial
food. The body length and weight of fish were measured
prior to exposure and fish were tagged for individual
recognition. Sampling was conducted after 4 weeks of
exposure. At sampling, the body length and weight of fish
were measured and blood was collected. Fish were then
killed by a blow to the head, the body cavity was opened,
and the liver weight was determined. The hepatosomatic
index (HSI) was obtained by expressing the liver weight as a
percentage of body weight. Liver and muscle samples from
all treated fish and several controls were taken for MYCST
analysis. Other liver samples from treatment and control fish
were obtained for transmission electron microscope (TEM)
analysis.
2.5. TEM
Liver samples were prefixed in a 2.5% glutaraldehyde
solution (in 0.1 M Na-cacodylate buffer, pH 7.4) for 2 h at
4 8C and postfixed in cold 1% osmium tetroxide (in 0.1 M
Na-cacodylate buffer, pH 7.2) for 2 h at 4 8C. Ultra-thin
sections were sliced with glass knives (LKB, ultra-
microtome, Nova, Sweden). The sections were examined

Table 1
Methods used for plasma enzyme assays performed with kits (ASAN, Korea)
Enzyme Method Reference
Alanine aminotransferase (ALT) Reitman–Frankel method Reitman and Frankel (1957)
Aspartate aminotransferase (AST) Reitman–Frankel method Reitman and Frankel (1957)
g-glutamyl transferase (GGT) 5-Aminosalicylic acid method Persijn and van der Slik (1976)
Lactate dehydrogenase (LDH) Wroblewski–LaDue method Wroblewski and LaDue (1955)
Alkaline phosphatase (ALP) Kind–King method Kind and King (1954)
X.-Y. Li et al. / Toxicon 44 (2004) 821–827 823
with an electron microscope (JEOL, JEM 1200EX-II,
Japan).
2.6. Blood collection and serum enzyme assay
At sampling, blood was collected from fish heart with a
syringe. Serum was recovered by centrifugation at 3000g for
10 min at 4 8C, and stored at K70 8C until needed. All
analyses were conducted using commercially available
serum enzyme assay kits (ASAN, Korea) based on
colorimetric reaction (Table 1).
2.7. Determination of MCYST concentration in liver
and muscle
MCYST was extracted from fish liver and muscle using
the method described by Malbrouck et al. (2003). MCYST
concentrations in fish tissues were analyzed by a highly
sensitive radioisotope protein phosphatase assay (Jones and
Orr, 1994). [32P]-phosphorylase-A was prepared by the
method of Cohen et al. (1988). Inhibition of protein
phosphatase was expressed as MCYST concentration
(MC-LR equivalent).
2.8. Statistical analysis
Results were expressed as the meanGSD. The two-tailed
Student’s t-test for sample equal variance was used to
compare the growth rate, hepatosomatic index and serum
enzyme activity changes between the treated and control
samples. Differences were considered to be statistically
significant at P!0.05.
Fig. 1. Growth rate of common carp during a period of 4 weeks of
oral exposure to Microcystis bloom scum (P!0.01). The exposure
dosage was 50 mg microcystin/kg body weight. Data are expressed
as the meanGSD (nZ10).
3. Results
3.1. Characteristics of Microcystis bloom scum
Microscopic examination revealed that cyanobacterial
bloom in the Naktong river was dominated by Microcystis
aeruginosa (Relative abundance, 91.3%). HPLC analysis
showed that three kinds of microcystins (MC-RR, -LR and
-YR) were identified in bloom material, of which MC-RR
was the dominant variant (Relative concentration, 73.7%)
and the total concentration of toxin in bloom scum was
357.3G26.8 mg/g dry weight.
3.2. Effect of MCYST on fish growth
Although the body color of treated fish became darker
than that of control fish, no obvious differences were
observed in appetite, swimming activity or other behavior of
control versus treated fish. No fish died during the course of
the experiment. Examination of the intestinal contents
revealed that most bloom scum could be digested by carp
(data not shown). Growth assay results showed that
the growth rate of the treated fish was significantly lower
than that of control fish (Fig. 1), while no significant
difference in the hepatosomatic index of fish was found in
the two groups of carp (Fig. 2).
3.3. Ultrastructural observations
TEM analysis revealed a number of ultrastructural
alterations in hepatocytes from MCYST-treated fish.
When hepatocytes from control and treated fish were
examined by electron microscope under low magnification
(2000!), sections derived from control fish had intact
plasmalemma and distinct cell junction (Fig. 3A). However,
hepatocytes from the treated fish had lost the intact
plasmalemma as well as the cell junction between
cells (Fig. 3B). Under higher magnification (5000!),
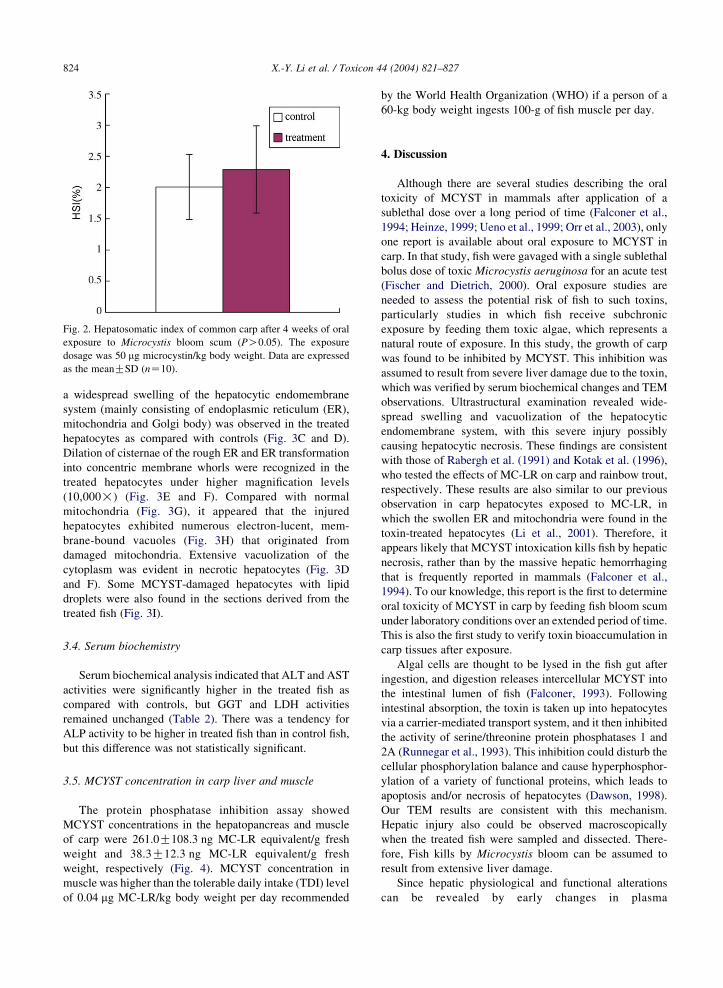
Fig. 2. Hepatosomatic index of common carp after 4 weeks of oral
exposure to Microcystis bloom scum (PO0.05). The exposure
dosage was 50 mg microcystin/kg body weight. Data are expressed
as the meanGSD (nZ10).
X.-Y. Li et al. / Toxicon 44 (2004) 821–827824
a widespread swelling of the hepatocytic endomembrane
system (mainly consisting of endoplasmic reticulum (ER),
mitochondria and Golgi body) was observed in the treated
hepatocytes as compared with controls (Fig. 3C and D).
Dilation of cisternae of the rough ER and ER transformation
into concentric membrane whorls were recognized in the
treated hepatocytes under higher magnification levels
(10,000!) (Fig. 3E and F). Compared with normal
mitochondria (Fig. 3G), it appeared that the injured
hepatocytes exhibited numerous electron-lucent, mem-
brane-bound vacuoles (Fig. 3H) that originated from
damaged mitochondria. Extensive vacuolization of the
cytoplasm was evident in necrotic hepatocytes (Fig. 3D
and F). Some MCYST-damaged hepatocytes with lipid
droplets were also found in the sections derived from the
treated fish (Fig. 3I).
3.4. Serum biochemistry
Serum biochemical analysis indicated that ALT and AST
activities were significantly higher in the treated fish as
compared with controls, but GGT and LDH activities
remained unchanged (Table 2). There was a tendency for
ALP activity to be higher in treated fish than in control fish,
but this difference was not statistically significant.
3.5. MCYST concentration in carp liver and muscle
The protein phosphatase inhibition assay showed
MCYST concentrations in the hepatopancreas and muscle
of carp were 261.0G108.3 ng MC-LR equivalent/g fresh
weight and 38.3G12.3 ng MC-LR equivalent/g fresh
weight, respectively (Fig. 4). MCYST concentration in
muscle was higher than the tolerable daily intake (TDI) level
of 0.04 mg MC-LR/kg body weight per day recommended
by the World Health Organization (WHO) if a person of a
60-kg body weight ingests 100-g of fish muscle per day.
4. Discussion
Although there are several studies describing the oral
toxicity of MCYST in mammals after application of a
sublethal dose over a long period of time (Falconer et al.,
1994; Heinze, 1999; Ueno et al., 1999; Orr et al., 2003), only
one report is available about oral exposure to MCYST in
carp. In that study, fish were gavaged with a single sublethal
bolus dose of toxic Microcystis aeruginosa for an acute test
(Fischer and Dietrich, 2000). Oral exposure studies are
needed to assess the potential risk of fish to such toxins,
particularly studies in which fish receive subchronic
exposure by feeding them toxic algae, which represents a
natural route of exposure. In this study, the growth of carp
was found to be inhibited by MCYST. This inhibition was
assumed to result from severe liver damage due to the toxin,
which was verified by serum biochemical changes and TEM
observations. Ultrastructural examination revealed wide-
spread swelling and vacuolization of the hepatocytic
endomembrane system, with this severe injury possibly
causing hepatocytic necrosis. These findings are consistent
with those of Rabergh et al. (1991) and Kotak et al. (1996),
who tested the effects of MC-LR on carp and rainbow trout,
respectively. These results are also similar to our previous
observation in carp hepatocytes exposed to MC-LR, in
which the swollen ER and mitochondria were found in the
toxin-treated hepatocytes (Li et al., 2001). Therefore, it
appears likely that MCYST intoxication kills fish by hepatic
necrosis, rather than by the massive hepatic hemorrhaging
that is frequently reported in mammals (Falconer et al.,
1994). To our knowledge, this report is the first to determine
oral toxicity of MCYST in carp by feeding fish bloom scum
under laboratory conditions over an extended period of time.
This is also the first study to verify toxin bioaccumulation in
carp tissues after exposure.
Algal cells are thought to be lysed in the fish gut after
ingestion, and digestion releases intercellular MCYST into
the intestinal lumen of fish (Falconer, 1993). Following
intestinal absorption, the toxin is taken up into hepatocytes
via a carrier-mediated transport system, and it then inhibited
the activity of serine/threonine protein phosphatases 1 and
2A (Runnegar et al., 1993). This inhibition could disturb the
cellular phosphorylation balance and cause hyperphosphor-
ylation of a variety of functional proteins, which leads to
apoptosis and/or necrosis of hepatocytes (Dawson, 1998).
Our TEM results are consistent with this mechanism.
Hepatic injury also could be observed macroscopically
when the treated fish were sampled and dissected. There-
fore, Fish kills by Microcystis bloom can be assumed to
result from extensive liver damage.
Since hepatic physiological and functional alterations
can be revealed by early changes in plasma

Fig. 3. Ultrastructural effects of MCYST on hepatocytes from carp exposed orally to Microcystis at a dose of 50 mg microcystin/kg body weight
for 28 days. (A) Control hepatocytes with intact plasmalemma and cell junction. (B) Hepatocytes from MCYST-treated carp. Note that these
hepatocytes have no visible plasmalemma and cell junction. (C) Ultrastructure of normal hepatocytes with stacks of rough ER. (D) Hepatocytes
from the treated fish showing a swollen endomembrane system. (E) The swollen RER in hepatocyte from toxin-treated fish. (F) Concentric
membrane whorls originating from the toxin-damaged ER (arrow). (G) Mitochondria of hepatocyte from control fish. (H) Vacuoles originating
from the toxin-damaged mitochondria. (I) MCYST-damaged hepatocytes with lipid droplets (arrow). RER, rough endoplasmic reticulum;
M, mitochondria; G, Golgi body; V, vacuole; N, nuclei.
X.-Y. Li et al. / Toxicon 44 (2004) 821–827 825
aminotransferase activity when an animal receives an
external shock, serum enzymes such as ALT, AST, and
GGT have been used as pathological and toxicological
indicators in clinical examination and toxicity research.
Table 2
Serum enzyme activities of common carp after 4 weeks of oral exposure
weight
ALT AST
Control 12.8G6.6 65.4G16.8
Treatment 37.4G27.3 91.1G26.3
Significance P!0.05 P!0.05
All analyses were conducted using commercially available serum enzyme
The International Unit (U lK1) was adopted as the enzyme activity value, e
meanGSD (nZ10).
Rabergh et al. (1991) reported that statistically significant
increases in activity of three liver enzymes, ALT, AST, and
LDH, were found in the plasma of common carp injected
with MC-LR at a dose of 150 mg/kg body weight.
to Microcystis bloom scum at a dose of 50 mg microcystin/kg body
GGT LDH (W-U) ALP
42.5G21.7 1305.5G706.1 12.1G7.3
33.1G30.3 1383.1G647.8 18.5G16.9
PO0.05 PO0.05 PO0.05
assay kits (ASAN, Korea) based on colorimetric reaction (Table 1).
xcept for LDH (W-U, Wroblewski Unit). Data are expressed as the
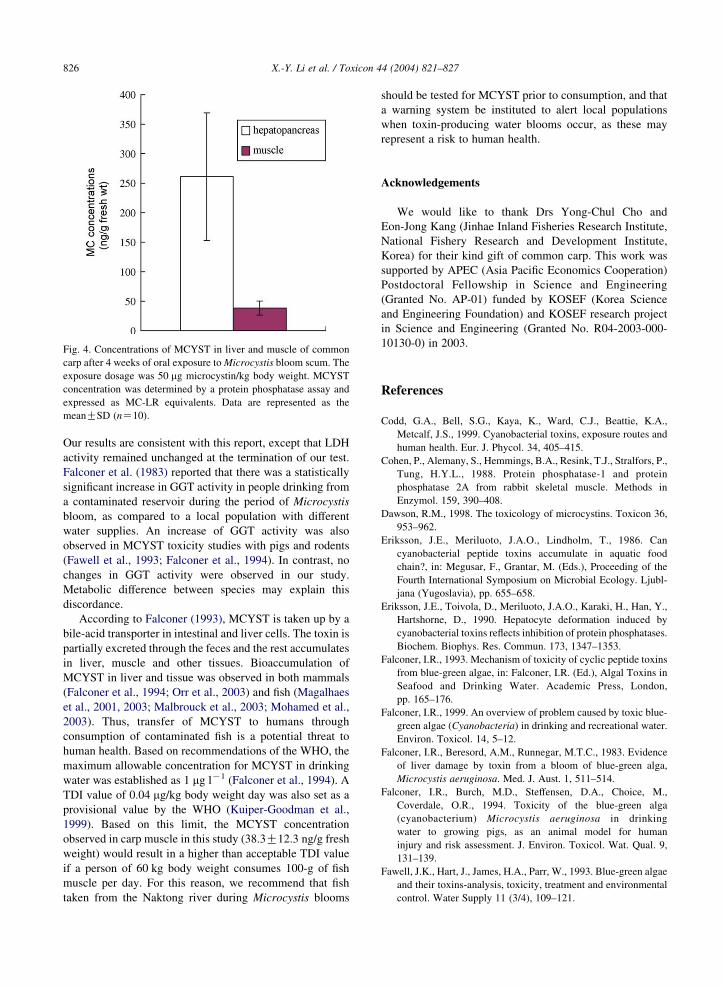
Fig. 4. Concentrations of MCYST in liver and muscle of common
carp after 4 weeks of oral exposure to Microcystis bloom scum. The
exposure dosage was 50 mg microcystin/kg body weight. MCYST
concentration was determined by a protein phosphatase assay and
expressed as MC-LR equivalents. Data are represented as the
meanGSD (nZ10).
X.-Y. Li et al. / Toxicon 44 (2004) 821–827826
Our results are consistent with this report, except that LDH
activity remained unchanged at the termination of our test.
Falconer et al. (1983) reported that there was a statistically
significant increase in GGT activity in people drinking from
a contaminated reservoir during the period of Microcystis
bloom, as compared to a local population with different
water supplies. An increase of GGT activity was also
observed in MCYST toxicity studies with pigs and rodents
(Fawell et al., 1993; Falconer et al., 1994). In contrast, no
changes in GGT activity were observed in our study.
Metabolic difference between species may explain this
discordance.
According to Falconer (1993), MCYST is taken up by a
bile-acid transporter in intestinal and liver cells. The toxin is
partially excreted through the feces and the rest accumulates
in liver, muscle and other tissues. Bioaccumulation of
MCYST in liver and tissue was observed in both mammals
(Falconer et al., 1994; Orr et al., 2003) and fish (Magalhaes
et al., 2001, 2003; Malbrouck et al., 2003; Mohamed et al.,
2003). Thus, transfer of MCYST to humans through
consumption of contaminated fish is a potential threat to
human health. Based on recommendations of the WHO, the
maximum allowable concentration for MCYST in drinking
water was established as 1 mg lK1 (Falconer et al., 1994). A
TDI value of 0.04 mg/kg body weight day was also set as a
provisional value by the WHO (Kuiper-Goodman et al.,
1999). Based on this limit, the MCYST concentration
observed in carp muscle in this study (38.3G12.3 ng/g fresh
weight) would result in a higher than acceptable TDI value
if a person of 60 kg body weight consumes 100-g of fish
muscle per day. For this reason, we recommend that fish
taken from the Naktong river during Microcystis blooms
should be tested for MCYST prior to consumption, and that
a warning system be instituted to alert local populations
when toxin-producing water blooms occur, as these may
represent a risk to human health.
Acknowledgements
We would like to thank Drs Yong-Chul Cho and
Eon-Jong Kang (Jinhae Inland Fisheries Research Institute,
National Fishery Research and Development Institute,
Korea) for their kind gift of common carp. This work was
supported by APEC (Asia Pacific Economics Cooperation)
Postdoctoral Fellowship in Science and Engineering
(Granted No. AP-01) funded by KOSEF (Korea Science
and Engineering Foundation) and KOSEF research project
in Science and Engineering (Granted No. R04-2003-000-
10130-0) in 2003.
References
Codd, G.A., Bell, S.G., Kaya, K., Ward, C.J., Beattie, K.A.,
Metcalf, J.S., 1999. Cyanobacterial toxins, exposure routes and
human health. Eur. J. Phycol. 34, 405–415.
Cohen, P., Alemany, S., Hemmings, B.A., Resink, T.J., Stralfors, P.,
Tung, H.Y.L., 1988. Protein phosphatase-1 and protein
phosphatase 2A from rabbit skeletal muscle. Methods in
Enzymol. 159, 390–408.
Dawson, R.M., 1998. The toxicology of microcystins. Toxicon 36,
953–962.
Eriksson, J.E., Meriluoto, J.A.O., Lindholm, T., 1986. Can
cyanobacterial peptide toxins accumulate in aquatic food
chain?, in: Megusar, F., Grantar, M. (Eds.), Proceeding of the
Fourth International Symposium on Microbial Ecology. Ljubl-
jana (Yugoslavia), pp. 655–658.
Eriksson, J.E., Toivola, D., Meriluoto, J.A.O., Karaki, H., Han, Y.,
Hartshorne, D., 1990. Hepatocyte deformation induced by
cyanobacterial toxins reflects inhibition of protein phosphatases.
Biochem. Biophys. Res. Commun. 173, 1347–1353.
Falconer, I.R., 1993. Mechanism of toxicity of cyclic peptide toxins
from blue-green algae, in: Falconer, I.R. (Ed.), Algal Toxins in
Seafood and Drinking Water. Academic Press, London,
pp. 165–176.
Falconer, I.R., 1999. An overview of problem caused by toxic blue-
green algae (Cyanobacteria) in drinking and recreational water.
Environ. Toxicol. 14, 5–12.
Falconer, I.R., Beresord, A.M., Runnegar, M.T.C., 1983. Evidence
of liver damage by toxin from a bloom of blue-green alga,
Microcystis aeruginosa. Med. J. Aust. 1, 511–514.
Falconer, I.R., Burch, M.D., Steffensen, D.A., Choice, M.,
Coverdale, O.R., 1994. Toxicity of the blue-green alga
(cyanobacterium) Microcystis aeruginosa in drinking
water to growing pigs, as an animal model for human
injury and risk assessment. J. Environ. Toxicol. Wat. Qual. 9,
131–139.
Fawell, J.K., Hart, J., James, H.A., Parr, W., 1993. Blue-green algae
and their toxins-analysis, toxicity, treatment and environmental
control. Water Supply 11 (3/4), 109–121.

X.-Y. Li et al. / Toxicon 44 (2004) 821–827 827
Fischer, W.J., Dietrich, D.R., 2000. Pathological and biochemical
characterization of microcystin-induced hepatopancreas and
kidney damage in carp (Cyprinus carpio). Toxicol. Appl.
Pharmacol. 164, 73–81.
Gupta, N., Pant, S.C., Vijayaraghavan, R., Lakshmana Rao, P.V.,
2003. Comparative toxicity of cyanobacterial cyclic peptide
toxin microcystin variants (LR, RR, YR) in mice. Toxicology
188, 285–296.
Harada, K.-I., Suzuki, M., Dahlem, A.M., Beasley, V.R.,
Carmichael, W.W., Rinehart, K.L., 1988. Improved method
for purification of toxic peptide produced by cyanobacteria.
Toxicon 26, 433–439.
Heinze, R., 1999. Toxicity of the cyanobacterial toxin microcystin-
LR to rate after 28 days intake with the drinking water. Environ.
Toxicol. 14, 57–60.
Jochimsen, E.M., Carmichael, W.W., An, J., Cardo, D.M.,
Cookson, S.T., Holmes, C.E.M., Antunes, M.B.C.,
Filho, D.A.M., Lyra, T.M., Barreto, V.S.T., Azevado, S.M.F.O.,
Jarvis, W.R., 1998. Liver failure and death after exposure
to microcystins at a hemodialysis center in Brazil. New Engl.
J. Med. 338, 873–878.
Jones, G.J., Orr, P.T., 1994. Release and degradation of microcystin
following algicide treatment of a Microcystis aeruginosa bloom
in a recreational lake, as determined by HPLC and protein
phosphatase inhibition assay. Water Res. 28, 871–876.
Kind, P.R.N., King, A.J., 1954. Elimination of plasma phosphate by
determination of hydrolysed phenol with aminoantipyrine.
J. Clin. Pathol. 7, 322–326.
Kotak, B.G., Semalulu, S., Fritz, D.L., Prepas, E.E., Hrudey, S.E.,
Coppock, R.W., 1996. Hepatic and renal pathology of
intraperitoneally administered microcystin-LR in rainbow
trout (Oncorhynchus mykiss). Toxicon 34, 517–525.
Kuiper-Goodman, T., Falconer, I.R., Fitzgerald, J., 1999. Human
health aspects, in: Chorus, I., Bartram, J. (Eds.), Toxic
Cyanobacteria in Water: A Guide to Their Public Health
Consequences, Monitoring and Management. E&FN Spon,
London, pp. 113–134.
Lee, J.A., Choi, A.R., Watanabe, M., 1997. Taxonomic implication
of the genus Microcystis (Cyanophyceae) from the Naktong
river. Algae 12, 167–176.
Li, X.Y., Liu, Y.D., Song, L.R., 2001. Cytological alterations in
isolated hepatocytes from common carp (Cyprinus carpio L.)
exposed to microcystin-LR. Environ. Toxicol. 16, 517–522.
Magalhaes, V.F., Soares, R.M., Azevedo, S.M.F.O., 2001. Micro-
cystin contamination in fish from the Jacarepagua Lagoon (Rio
de Janeiro, Brazil): ecological implication and human health
risk. Toxicon 39, 1077–1085.
Magalhaes, V.F., Marinhao, M.M., Domingos, P., Oliverira, A.C.,
Costa, S.M., Azevedo, L.O., Azevedo, S.M.F.O., 2003. Micro-
cystins (cyanobacterial hepatotoxins) bioaccumulation in fish
and crustaceans from Sepetiba Bay (Brasil, RJ). Toxicon 42,
289–295.
Malbrouck, C., Trausch, G., Devos, P., Kestemont, P., 2003.
Hepatic accumulation and effects of microcystin-LR on juvenile
goldfish Carassius auratus L.. Comp. Biochem. Physiol. Part C
135, 39–48.
Mohamed, Z.A., Carmichael, W.W., Hussein, A.A., 2003. Esti-
mation of microcystins in the freshwater fish Oreochromis
niloticus in an Egyptian fish farm containing a Microcystis
bloom. Environ. Toxicol. 18, 137–141.
Orr, P.T., Jones, G.J., Hunter, R.A., Berger, K., 2003. Exposure of
beef cattle to sub-clinical doses of Microcystis aeruginosa: toxin
bioaccumulation, physiological effects and human health risk
assessment. Toxicon 41, 613–620.
Persijn, J.P., van der Slik, W., 1976. A new method for the
determination of gamma-glutamyltransferase in serum. J. Clin.
Chem. Clin. Biochem. 14, 421–427.
Rabergh, C.M.I., Bylund, G., Eriksson, J.E., 1991. Histopathological
effects of microcystin-LR, a cyclic peptide toxin from the
cyanobacterium (blue-green alga) Microcystis aeruginosa, on
common carp (Cyprinus carpio L.). Aquat. Toxicol. 20, 131–146.
Reitman, S., Frankel, S., 1957. A colorimetric method for the
determination of serum glutamic oxalacetic and glutamic
pyruvic transaminases. Am. J. Clin. Pathol. 28, 56–63.
Runnegar, M.T., Kong, S., Berndt, N., 1993. Protein phosphatase
inhibition and in vivo hepatotoxicity of microcystins. Am.
J. Physiol. 265, G224–G230.
Toranzo, A.E., Nieto, F., Barja, J.L., 1990. Mortality associated with a
cyanobacterial bloom in farmed rainbow trout in Galicia (North-
western Spain). Bull. Eur. Assoc. Fish Pathol. 10, 106–107.
Ueno, Y., Makita, Y., Nagata, S., Tsutsumi, T., Yoshida, F.,
Tamura, S-I., Sekijima, M., Tashiro, F., Harada, T., Yoshida, T.,
1999. No chronic oral toxicity of a low dose of microcystin-LR,
a cyanobacterial hepatotoxin, in female BALB/c mice. Environ.
Toxicol. 14, 45–55.
Wroblewski, F., LaDue, J.S., 1955. Lactic dehydrogenase activities
in blood. Proc. Soc. Exp. Biol. Med. 90, 210–213.
Yoshizawa, S., Matsushima, R., Watanabe, M.F., Harada, K.I.,
Ichihara, A., Carmichael, W.W., Fujiki, H., 1990. Inhibition of
protein phosphatases by microcystins and nodularin associated
with hepatotoxicity. J. Cancer Res. Clin. Oncol. 116, 609–614.
Zimba, P.V., Khoo, L., Gaunt, P., Carmichael, W.W., Brittain, S.,
2001. Confirmation of catfish mortality from Microcystis toxins.
J. Fish Dis. 24, 41–47.