structural changes in the hippocampal formation after long-term alcohol consumption and withdrawal...
TRANSCRIPT

Addiction (1993) 88, 237-247
RESEARCH REPORT
Structural changes in the hippocampalformation after long-term alcoholconsumption and withdrawal in the rat
M. M. PAULA-BARBOSA, F. B R A N D A O , M . D . MADEIRA &A. CADETE-LEITE
Department of Anatomy, Porto Medical School, Alameda Hemdni Monteiro,4200 Porto, Portugal
AbstractThe effects of long-term alcohol consumption and withdrawal upon the structure of the rat hippocampal
formation were studied by applying morphometric methods to material processed for light and electronmicroscopy. The somatostatinergic neurons of the hilus were also studied. Groups of 6 rats were treated asfollows: (a) given alcohol for 6, 12 and 18 months; (b) paired controls; and (c) rats switched to a normaldiet in the 6 months after 6 and 12 months of alcohol intake. A progressive loss of hippocampal neurons afterchronic alcohol consumption was found. The loss was aggravated during withdrawal from alcohol, with theexception of the hilar cells. The dendrites of granule cells from the alcohol-treated rats displayed signs ofregrowing, but they did not do so in rats withdrawn from alcohol. The synapses between mossy fibre terminalsand CA3 dendrites appear to be rather resistant to alcohol insult, and evidence of morphological plasticity wasfound in withdrawn rats. If an homology can be established between humans and rodents then the changesobserved in alcohol-fed rats can be regarded as underpinning some of the functional and behaviouralalterations depicted under these circumstances. The peculiar changes found in some nerve cell populations afterwithdrawal of alcohol could be related to the deficient or incomplete functional recovery often seen afterabstinence from alcohol.
Introduction malnutrition,''' commonly accompany chronicAlcoholism continues to be a major health and alcohol consumption and are known also to af-social problem of most societies. The alcohol- feet the neuronal structure. In 1987 Riley &induced neurocytological alterations that under- Walker described marked neuronal loss in thelie brain dysfunction were first reponed in rodent hippocampal formation after 20 weeks ofhumans by Courville in 1955.' These structural alcohol intake using, for the first time in brainchanges needed to be carefully interpreted be- studies, a nutritional controlled model.cause other factors, vitamin deficiency" and Over the past 12 years, using an animal model
of alcohol consumption with periods of ethanolintake as long as 18 months, we have contributed
Correspondence should be addressed to: M. M, Paula- •_ i • • r L 'i • lBarbosa, Department of Anatomy, Porto Medical School, O the comprehensive picture of the Stnking al-Al. Hemaani Monteiro, 4200 Poito, Portugal. terations of the Structural organization of the
237

238 M. M. Paula-Barbosa et al.
Figure 1. Lateral view qfche right hippocampal formation (HF) exposed by removal of the neoconex. The line was tracedin its midseplotemporal pan, where the semithin section shown in Fig, 2A was obtained, x 4.5.
CNS after chronic alcohol consumption. We re-poned massive degenerative changes, such asneuronal death/'^ impoverishment of dendritictrees,** loss of synaptic contaets,'' formation ofneuritic plaques'" and a precocious increasedaccumulation of lipofuscin."''^ Notwithstandingthe massive deteriorative changes, there was evi-dence of remodeling activity, which includedsynaptic plasticity"'' '' and increase in size of thedendritic spine.'"
The morphological effects of chronic alcoholconsumption on the cerebellum were the first tobe studied because of the cerebellum's simplestructural arrangement." Subsequently, we ex-tended the investigations to the hippocampalformation,"^ ' taking into account its role incognitive deficits, • '' its relatively simple andhighly characteristic structure,^^ and its vulnera-bility to ethanol aggression.""'"^ More recently,we have paid attention to the effects of with-drawal after chronic alcohol consumption '' ' ^in order to gain an insight on the effects ofprolonged alcohol abstinence and to shed lighton the controversy concerning the reversibility ofthe alcohol-induced deteriorative changes.-'' ''''
Material and methodsTwo-month-old male Sprague-Dawley rats wereseparated into groups of 6 animals and weretreated as follows: (a) animals fed a 20%aqueous ethanol solution as the only availableliquid source for 6, 12 and 18 months; in theseanimals food and fluid intake were measured
every other day and the amounts consumed werecalculated; (b) paired controls; these animalswere given the same amount of food as theirmatched experimental animals, and water in asolution where sucrose replaced the ethanolisocalorically; (c) animals alcohol-fed for 6 and12 months and then switched to water for 6months; these groups will be called withdrawalgroups. All animals were given vitamins and saltsupplements.
At the end of each experimental period theanimals were perfused with 1% paraformalde-hyde and 1% glutaraldehyde. The righthippocampal formations (Fig. 1) were processedas for electron microscopy,'' and the left oneswere post-fixed in the solution used for the per-fusion and Golgi-impregnated.'' TTieobservations were carried out in horizontal sec-tions from the midseptotemporal part of thehippocampal formations (Fig. 2A). The CAlsubdivision was quantified only in 18—monthold animals, and the somatostatin-immunoreac-tive (SS-I) hilar cells"* were studied only in6-month and 12-month old rats.
Different sets of animals were used for im-munostaining; they were perfijsed with 4°/<>paraformaldehyde. The brains were sliced at 50Jim thick on a Vibratome. The antibody wassupplied by DAKO (Denmark) and the PAPmethod was used. **
After structural hippocampal componentsvisualized in semithin sections were drawn withthe aid of a camera lucida, the thickness of thegranular and CAl and CA3 pyramidal celt layers
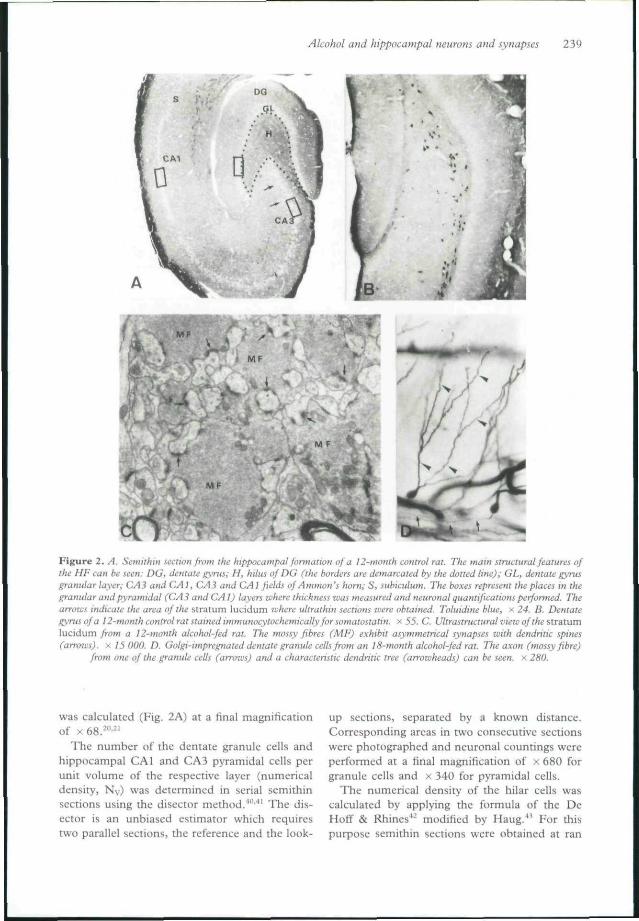
Alcohol and hippocampal neurons and synapses 239
MF
Figure 2. A. Semithin section from the hippocampal formation of a 12-month control rat. The main structural features ofihe HF can be seen: DO, dentate gyrus; H, hilus of DG (ifie borders are demarcated by the doited line); GL, dentate gyrusgranular layer; CA3 and CAl, CAJi and CAl fields of Amman's horn; S, subiculum. The boxes represent the places in thegranular and pyramidal (CA3 andCAl) layers where thickness was measured and neuronal quantifications performed. Thearrows indicate the area of the stratum lucidum luhere ultrathin sections were obtained. Toluidine blue, x 24. B. Dentategyrusofa 12-month control rat stained immunocytochemically for somatostatin. x 55. C. Ultrastructurat view of the stratumlucidum from a 12-month alcohol-fed rat. The mossy fibres (MF) exhibit asymmetrical synapses with dendritic spines(arrows), x 75 000. D. Golgi-impregnaied dentate granule cells from an 18-month alcohol-fed rat. The axon (mossy fibre)
from one of the granule cells (arrows) and a characteristic dendritic tree (arrowheads) can he seen, x: 280.
was calculated (Fig. 2A) at a final magnificationof x68.^"'^'
The number of the dentate granule cells andhippocampal CAl and CA3 pyramidal cells perunit volume of the respective layer (numericaldensity, Ny) was determined in serial semithinsections using the disector method.•"'•"" The dis-ector is an unbiased estimator which requirestwo parallel sections, the reference and the look-
up sections, separated by a known distance.Corresponding areas in two consecutive sectionswere photographed and neuronal countings wereperformed at a final magnification of x 680 forgranule cells and x 340 for pyramidal cells.
The numerical density of the hilar cells wascalculated by applying the formula of the DeHoff & Rhincs''' modified by Haug.*' For thispurpose semithin sections were obtained at ran

240 M. M. Paula-Barbosa et al.
dom from three blocks of tissue of each animaland the number of cells per surface area andneuronal diameters were evaluated.*^
The disector method is known to give the bestestimate of particle density since it is unbiased bypanicle size and shape and eliminates the prob-lem of lost caps, which must be taken intoconsideration when the classic stereologicalmethods are used." With the disector methodpanicles must be evenly distributed throughoutthe reference area for representative samples ofthe neuronal populations to be obtained. Forhilar cells this would require the entire hilus tobe photographed, which is difficult to perform.
The immunostained hilar cells (Fig. 2B) werecounted per section.
The disector method was also used to calcu-late the numerical density of MF-CA3 synapsesin the stratum luadum (Fig. 2C), those estab-lished between the mossy fibre terminals and theexcrescences of the proximal pan of the CA3pyramidal dendrites. Serial electron micrographswere analyzed at a final magnification ofX 18 000.^' The percentage of plasmalemmafrom the mossy fibre terminals occupied bysynaptic contacts was calculated from the samephotographs by measuring the perimeter of theterminals and the length of the postsynaptic den-sities with the aid of a Mop-Videoplan.^'
Fifteen Golgi-impregnated granule cells (Fig.2D) per animal, on average, were chosen forquantitative evaluation of the three-dimensionalbranching pattern and of granule cell dendriticspines.^"'" These evaluations were carried outusing a semi-automatic dendrite-measuring sys-tem developed at The Netherlands Institute forBrain Research. "'" '"'•'•'' Cells were selectedaccordingly to previously defined criteria^"' ' andfi-om each cell the following parameters werestudied: the projected area of the cell soma (so-matic surface), total number of dendrites, totalnumber of segments, total dendritic length, andradial distance from the cell soma to terminal
The non-parametric two-tailed Mann-Whit-ney U test was used to analyze differencesbetween groups. A value of p < 0.05 was consid-ered statistically significant.
ResultsQualitatively, in some of the alcohol-treated ani-mals, a loss of granule, pyramidal and hilar
neurons could be seen. The morphology of thegranule cells' dendritic trees and their spines didnot differ among the groups studied. At theultrastructural level, degenerated neurites wereabundant in alcohol-fed rats and withdrawn ratsand occurred in both axons and dendrites.
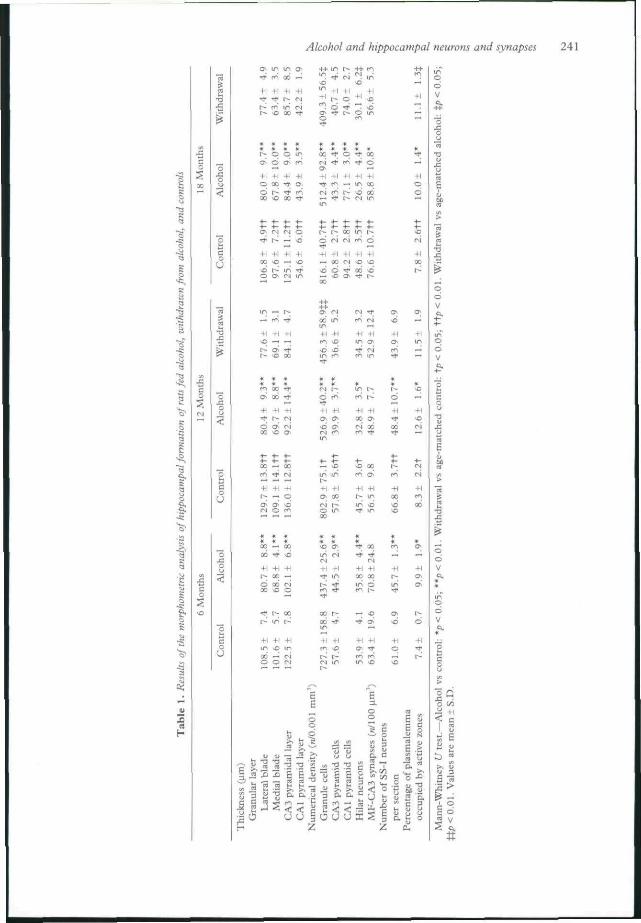
Results of the quantitative analysis are pre-sented in Table 1. In the dentate gyrus thegranule cell layer was significantly thinner inalcohol-fed rats and withdrawn rats than in therespective age-matched controls. The valuesfound in withdrawal groups matched those ob-tained in alcohol-fed groups. "' ** The number ofgranule cells in alcohol-fed rats was significantlylower than in their controls. Also, the numericaldensity of granule cells was significantly lower inthe withdrawn rats than in age-matched alcohol-fed animals.'' '-"
The thickness of CAl and CA3 pyramidallayers and the numerical density of their neuronswere significantly lower in alcohol-fed groupsthan in age-matched controls. The numericaldensity of the CA3 and CAl pyramidal cells waslower in withdrawal groups than in age-matchedalcohol-treated groups, although the differenceswere not significant.
The numerical density of hilar cells insemithin sections was significantly lower in alco-hol-treated and withdrawn animals than incontrols at all ages.'' In 18-month old animalsthe numerical density was significantly greater inthe withdrawal group than in the alcohol-fedone. The number of SS-I neurons per sectionwas significantly lower in both the alcohol-treated and withdrawal groups than in controls.''"
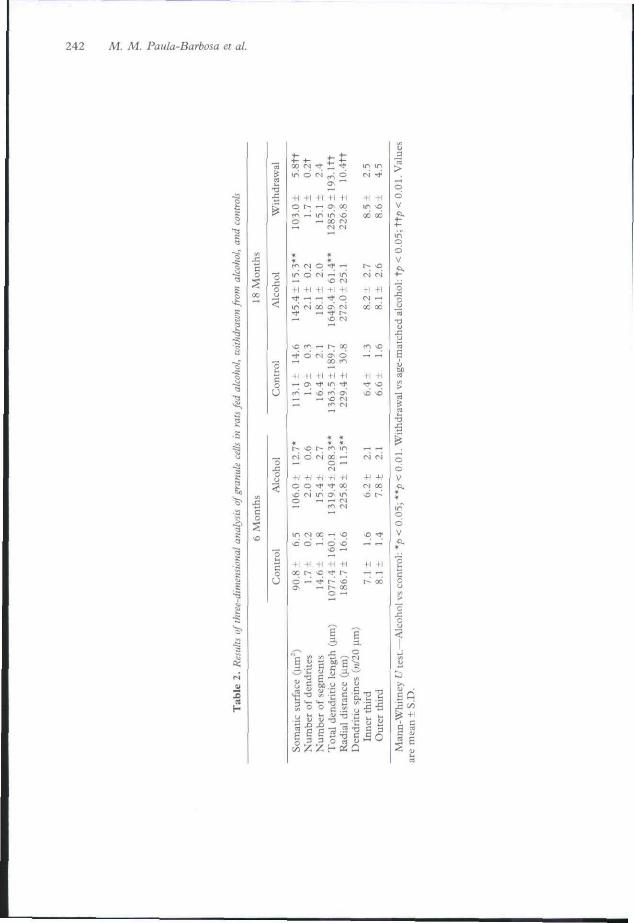
In Golgi-impregnated material (Table 2) wefound that the somatic surface was significantlygreater in 6-month and 18-month alcohol-fedanimals than in controls. There was no signifi-cant differences in the number of dendrites percell in alcohol-fed groups, but the withdrawalgroup had significantly fewer dendrites per cellthan did the controls. Total dendritic length wassignificantly greater in alcohol-fed animals andless in the withdrawal group. No differenceswere found in the number of dendritic segmentsper neuron. The radial distance of the terminalsegments was significantly greater after 6 tnonthsof alcohol intake than in controls. The density ofdendritic spines was not significantly differentbetween alcohol-fed or withdrawn animals and
In the ncuropil of the stratium lucidum
Alcohol and hippocampal neurons and synapses 241
^ f<^ 00 —
+1 +1 +1 +1
oo0(1
+10 0
^
4-4-
-HI f -
OTi
O-
IT't
—
f ^
i n
( N
4-4-
•0
i n
3^
-fla>
•d
1--*
+1•or ^
IN
+ 1—li-i
o
4-1
d-t-i—.
+100
d
00
M
+1I N
- t
i n
ci+1
00
d
•d
-n
6
«00OO
-H
a-"
*
-H
^^+1r-
— 00Tf- fvi
+1 +1— o
-.-i nr--fi
in
4-1ao
+1 4-1 4-1r^ 00 —d 00 (N
^ O —' *
^ 3 3 E E
r-1 m •—
"a c
a cta ,_,
[ J
U U a u u I 5 E
J^i -H
I u5 E
242 M. M. Paula-Barbosa et al.
00 <N "? _ • *in d i>J P-; d
41 +1 +1 +1 41q r- —• o; 00r — Lo Ln >co — ao f-J
+1 +1 +1 +1 +1^ OS -J- lO ^
+1 +1 +1 +1 41o q ^ rf CO\D oi m o- in
>D o ^ o ^
(I +1 -H 4-1 -H
o —• •* r~ ^a, — t-- X
o —
if
^ O O g w
p E fc 3 ^S 3 3 O rt
in in<N -*
-H 4-1in \qX 00
i n.—4-1
o
4-1 +1 +1— Tl-
•—•
i n(N-l-l
o
fN
41IN
fN
f l—
+1 4-1fN CO
+1 +1
t-- 00
E Q

Alcohol and hippocampal neurons and synapses 243
(Table 1), no significant differences were foundin the numerical density of the MF-CA3synapses when 6-month and 12-month alcohol-fed groups were compared with respectivecontrols, but after 18 months of experimentthere was significant reduction in the number ofsynapses, in both the alcohol-fed and withdrawalgroups.^' The percentage of mossy fibre plas-malemma occupied by postsynaptic densitieswas greater in the alcohol-fed and withdrawalgroups than in controls.^'
DiscussionOf the changes found in the brains of alcohol-treated animals, cell loss is one of the mostsevere. In the hippocampal formation of rodentsfed alcohol for 20 weeks. Walker et al?"-^'^showed that the number of dentate granule cellsper unit section area was reduced by approxi-mately 20%, despite the short period of alcoholfeeding. In a previous work** we showed that thedegree of cerebellar granule cell loss dependedon the duration of exposure to alcohol. Thepresent data demonstrate that the dentate gyrusis reduced by 40% of its granule cells after 6months of alcohol feeding.
As well, the numerical density of the granulecells was lower in the withdrawal groups than inthe alcohol-fed groups, which indicates thatwithdrawal from ethanol does not impede theongoing cell death. This observation is consistentwith the numerical reduction observed byPhillips & Cragg in CAl pyramidal cells andPurkinje cells after shoner periods of alcoholfeeding and withdrawal."'^^ This is an unex-pected finding and difficult to explain. Recentstudies by Begleiter et al.'"'^'' indicate that with-drawal from alcohol could involve a complexsequence of modifications which cause furtherneuronal changes, leading to alterations in thefunctioning of the CNS.
Walker & coworkers showed that the numberof the CAl and CA3 pyramidal cells was re-duced;'"'^' Lescaudron & Vema, " in a similarexperimental model, found a numerical reduc-tion of CAl neurons. Phillips & Cragg" foundno change in the number of CAl pyramidal cellsafter 4 months of alcohol feeding. We found asignificant loss of both CAl and CA3 pyramidalcells, reaching 20% and 30% respectively after18 months of alcohol feeding, together with athinning of the pyramidal cell layer. Contrary to
the findings with dentate granule cells, no differ-ences were observed between alcohol-fed andwithdrawn animals.
The maintenance of the altered hippocampalstructure after withdrawal from alcohol, or, evenworse, the increased neuronal degenerationfound under these circumstances, apparentlydoes not fit with the anatomical amelioration inhumans described in numerous scanner stud-ies.^"'"' However, it must be kept in mind thatchronic alcohol consumption induces markedand widespread metabolic changes, namely inthe hydric balance; for instance, dehydration iscommonly seen after prolonged alcohol inges-tion. As a consequence the shrunken appearanceof the brain of human alcoholics might resultfrom the altered water metabolism,"''''" and theestablishment of a normal milieu in ratswithdrawn from alcohol might contribute to thestructural improvement of the brain'sappearance.
The neuronal loss observed in the hippocam-pal formation might have electrophysiologicaland behavioral correlates. It seems obvious tospeculate that the hippocampal trisynaptic cir-cuitry will be severely damaged as the granuleand pyramidal neurons are important links inthis circuitry. Furthermore, the loss of SS-I hilarcells might inflict severe functional changes. It iswell known that these hilar cells colocalize theneuropeptide Y (NPY) and GABA. These cellsform 20-25% of the neurons of this area.GABAergic functions are deeply affected bychronic alcohol consumption, as Hunt &Majchrowicz, ' Leonard, * and Lescaudron etaf'' have shown. The SS-I neurons appear pri-marily to extend their neurites to the outer partsof the dentate molecular layer of both ipsilateraland contralateral dentata gyrus. It seems thatSS-I neurons modulate the entorhinal input tothe fascia dentata through inhibitory synapsesestablished over the dcndrites of granule cells,dendrites that receive the entorhinal perforantpath. A loss of inhibition may explain the in-creased excitability of the neurons found inexperimental epilepsy in the rat '' and in humantemporal lobe epilepsy,"" situations in whichthere is loss of SS-I neurons. Such loss may alsoexplain the increased susceptibility to convul-sions verified in the animals withdrawn fromethanol.-' The selective loss of SS-I and NPY-1neurons in the human hippocampal formation

244 M. M. Paula-Barbosa el al.
has also been observed in Alzheimer's disease byChan-Palay;'^ it may be consistent with a role forthese cerebral peptides in cognitive functions.
The increase in the granule cells' dendriticlength observed in alcohol-treated rats mighttherefore indicate that the surviving neurons takeover additional areas for new synaptic connec-tions with the input fibres. This means that,as previously reported in the hippocampalformation and cerebellum under similar circum-stances,"''"''^" the reduction of the dendriticnetwork is partially compensated by the prolifer-ation of the remaining granule cell dendrites.Dendritic plasticity was quantitatively shown byDurand et al.^'' in granule cell dendrites after 5months of alcohol feeding, using a shifted linearanalysis, but Riley & Walker and McMullen elal?^ were unable to describe it.
Withdrawn animals display a global reductionof their dendritic arborizations when comparedwith the group alcohol-fed for 18 months, as canbe seen from the somatic surface, total numberof dendrites, total dendritic length and the radialdistance, which arc significantly reduced. Tlienon-existence of plastic compensatory mecha-nisms in withdrawn animals shows that thesubstrate available for synaptic connections withthe afferents and for interaction with the surviv-ing close neurons is reduced after withdrawal.
Plastic alterations of dendritic spines followingchronic alcohol consumption have already beendescribed. In our laboratory an elongation inPurkinge cell spines was reponed,"' whereasKing et al. '' found that spine density decreasedin CAl pyramidal cells and increased in granulecells after 5 months of alcohol ingestion, reach-ing control levels during abstinence. Lescaudronei al.''" reported that CAl pyramidal cell den-dritic spines were reduced after 6 and 9 monthsof alcohol intake, returning to normal valuesafter withdrawal from ethanol. In granule cellswe were unable to show significant differences inthe density of granule cell spines in both thealcohol-treated and withdrawal groups.
A significant reduction (23Vo) of the MF-CA3synapses was found after 18 months of alcoholconsumption. It probably arises from the degen-eration of granule cell mossy fibres and CA3dendrites, screened throughout the straiiumlucidum of alcohol-fed animals. A synaptic de-crease would be expected to occur precociouslyas marked granule and pyramidal cells reduc-tions are already present after 6 and 12 months
of alcohol exposure; therefore fewer synapseswould be found in all alcohol-fed groups than incontrols. The maintenance of the number ofsynapses at control levels in 6-month and 12-month alcohol-fed groups suggest that newsynapses must have been formed in order tocompensate for the synapse loss due to neuronaldegeneration. At 18 months, due to the in-creased cell loss, it is likely that this plasticprocess collapses and leads to a decline in thedensity of synapses. '
Compensating for the synaptic reduction is theincrease in the synaptic extension, which inphysiological terms is known to cause a netincrease in the primary region of synaptic activa-tion.**' It provides an additional means ofpreserving the total amount of postsynaptic con-tact area.''
In summary, the structural organization of thehippocampal formation was found to bemarkedly affected by chronic alcohol consump-tion, and the magnitude of the effect can berelated to the length of the alcohol treatment.There are marked plastic potentialities in thehippocampal formation during alcohol feeding,but their functional implications are not clear.Withdrawal following alcohol consumption doesnot interfere with the structural changes occur-ring during alcohol consumption. On thecontrary, in some hippocampal subdivisions thedegenerative activity observed during withdrawalis more prominent than that seen after alcoholconsumption.
AcknowledgementsThe supervision of Prof Jens Zimmer in theimmunostaining of hilar cells is acknowledged.This study was supported by grants fromStiftung Volkswagenwerk (1-67432), InstitutoNacional de Investigagao Cientifica (CMEUP)and Junta Nacional de Investigagao Cientifica eTecnologica (Project PMCT/C/SAU/32/90).
References1. COURVILLE, C. B. (1955) Ejfecls of Alcohol on ike
Nervous System of Man (Los Angeles, San LucasPress).
2. YKW, M . L . S., MOORK, S, & BiKSELE, M. M.(1981) Effects of chronic 'moderate' alcohol con-sumption on vitamins A and C status of maleSprague-Dawley rats, Nutrition Reports Interna-tional, 23, pp. 427-436.
3. VICTOR, M . & ADAMS, R. D . & (1961) On the

Alcohol and hippocampal neurons and synapses 245
etiology of the alcoholic neurologic diseases withspecial reference to the role of nutrition, AmericanJournal of Climcal Nutrition, 9, pp, 379-397,
4. PAUIJ\-BARBOSA, M . M . , ANURADE, J . P.,
CASTKDO, J- L. , AZKVEDO, F . P., CAIMOHS, I., VOI.K,
B. & TAVARI-S, M . A. (1989) Cell loss in thecerebellum and hippocampal formation of adultrats after long-term low-protein diet. ExperimentalNeurology, 103, pp. 186-193.
5. RlUiYj. N. & W A I J < : E R , D . W . (1978) Morpholog-ical alterations in hippocampus after long-termalcohol consumption in mice, Science, 201, pp.646-648,
6. TAVARHS, M. A, & PAUIJ\-BARBOSA, M. M. (1982)
Alcohol-induced granule cell loss in the cerebellarcortex of the adult rat, Experimental Neurology, 78,pp. 574-582,
7. TAVARES, M , A., PAULA-BAKB{ISA, M . M , &
CADHTL-LEITH, A . (1987) Chronic alcohol con-sumption reduces the cortical layer volumes andthe number of neurons of the rat cerebellar cortex,Alcohottsrn: Clinical and Experime?ital Research, 11,pp. 315-319.
8. TAVARHS, M . A. , PAULA-BARBOSA, M , M . & GRAY,
E. G. (1983) A morphometric Golgi analysis of thePurkinge cell dendritic tree after long-term alcoholconsumption in the adult rai. Journal of Neurocycol-ogy, 12, pp. 939-948,
9. TAVARI-:S, M . A,, PALIA-BARBOSA, M . M , &
VERWER, R. W, H. (1987) Synapses of the cerebel-lar cortex molecular layer after chronic alcoholconsumption. Alcohol, 4, pp. 109-116.
10. PAULA-BARBOSA, M . M . & TAVERES, M . A. (1984)
Neuritic plaque-like structures in the rat cerebel-lum following prolonged alcohol consumption,Experientia, 40, pp, 110-112,
11. TAVARES, M . A. & PAL-IA-BARBOSA, M . M . (1983)
Lipofuscin granules in Purkinje cells after long-term alcohol consumption in rats, Aleoholism:(.'Mnical and Experimental Research, 7, pp. 302 -306.
12. TAVARES, M . A., pAutA-BARBosA, M, M., BAR-ROCA, H. & V01.K, B. (1985) Lipofuscin granulesin cerebellar intemeurons after long-term alcoholconsumption in the adult rat, Anatomy and Embry-ology, 171, pp. 61-69.
13, TAVARES, M. A. & PAULA-BARBOS.^ M. M. (1984)
Remodeling of the cerebellar glomeruli after long-term alcohol consumption in the adult rat. BrainResearch, 309, pp. 217-226.
14, TAVARES, M . A,, PAUIA-BARBDSA, M . M , &
CADE rii-LElTE, A. (1986) Morphological evidenceof climbing fiber plasticity after long-term alcoholintake, Neurobehavioral Toxicology and Teratology,8, pp. 481-485.
15. TAVARHS, M , A, , PAUIA-BARBOSA, M . M . & VOLK,
B. (1986) Chronic alcohol consumption inducesplastic changes in granule cell synaptic bouions ofthe rat cerebellar cortex. Journal of SubmicroscopicCytology, 18, pp. 725-730.
16, TAVARES, M , A., PAUIA-BARBOSA, M . M . & GRAY,
E. G. (1983) Dendritic spine plasticity and chronicalcoholism in rats, Neuroscience letters, 42, pp.235-238,
17, PAIAY, S, L, & CHAN-PAIAY, V. (1974) Cerebellar
Cortex. Cytology and Organisation (Berlin,Springer),
18. BoRGES, M. M., PAUL/\-BARBOSA, M . M , & Votjc,B. (1986) Chronic alcohol consumption induceslipofuscin deposition in the rat hippocampus, Neu-robiology of Aging, 7, pp. 347-355.
19. PAUIA-BARBOSA, M . M . , BORGES, M . M , , CADETE-
L E I I E , A, & TAVARES, M . A. (1986) Giantmultevisicular bodies in the rat hippocampal pyra-midal cells after chronic alcohol consumption,Neuroscience Letters, 64, pp. 345-349.
20. CADETE-LHITK, A,, TAVARES, M . A., UYIJNGS,
H, B. M. & PAUtA-BARBOSA, M. M. (1988)Granule cell loss and dendritic regrowing in thehippocampal dentate gyrus of the rat after chronicalcohol consumption. Brain Research, 473, pp.1-14,
21. CADETE-LEITE, A. , TAVARES, M . A,, P.-VCHECO,
M. M,, VOLK, B. & PAULA-BARBOS.A, M. M. (1989)
Hippocampal mossy fiber-CA3 synapses afterchronic alcohol consumption and withdrawal.Alcohol, 6, pp, 303-310,
22. CADETH-LHITE, A., T.AVERES, M . A,, ALVES, M , C ,
UYLINGS, H . B, M , & PAUIA-BARBOSA, M . M ,
(1989) Metric analysis of hippocampal granule celldendritic trees after alcohol withdrawal in rats.Alcoholism: Clinical and Experimental Research, 13,pp. 837-840.
23. BuTiERs, N,, CKR.VL^K, L, S., MONTGOMERY, K, &
AniNOt^Ei, A. (1977) Some comparisons of thememory and visuoperceptive deficits of chronicalcoholics, and patients with KorsakofTs disease,Alcoholism: Clinical and Experimental Research, 1,pp. 73-80.
24. WALKER, D . W . & HUNTER, B . E . (1978) Short-term memory impairment following chronicalcohol consumption in rats, Neuropsychologia, 16,pp. 545-553.
25. ANDERSEN, P. (1975) Organization of hippocam-pal neurons and their interconnections, in:ISAACSON, R. L . & PRIBRAM, K, H . (Eds) The
Hippocampus—Structure and Development, vol. 1,pp, 155-175 (New York, London, Plenum Press).
26. WALKER, D . W . , BARNES, D . E. , ZORNETZER, S . F . ,
HUNTER, B . E, & KUBANIS, P. (1980) Neuronalloss in hippocampus induced by prolonged ethanolconsumption in rats. Science, 209, pp. 711-713,
27. WALKER, D . W . , HUNiiiR, B, E, & ABRAMA-M,
W. C. (1981) Neuroanatomical and functionaldeficits subsequent to chronic ethanol administra-tion in animals. Alcoholism: Clinical andExperimental Research, 5, pp. 267-282.
28. CADETF-LI-ITE, A,, TAVARES, M . A. & PAUtA-
BARBOSA, M . M , (1983) Alcohol withdrawal doesnot impede hippocampal granule cell progressiveloss in chronic alcohol-fed rats, NeuroscienceLetters, 86, pp. 45-50,
29. CARLEN, P . L, , WoRizMAN, G., H01.GATE, R. C ,WILKINSON, D . A. & RANKIN, J. G. (1978) Re-
versible cerebral atropby in recently abstinentchronic alcoholics measured by computed tomo-graphy scans. Science, 200, pp, 1076--1078.
30. CARLEN, P. L., PEEN, R. D . , EORNAZZARI, L. ,
BENNETT, J., Wit.KiNSON, D. A, & W()RTz\iAN, G.

246 M. M. Paula-Barbosa el al
(1986) Computerized tomographic scan assess-ment of alcoholic brain damage and its potentialreversibility, Alcoholism: Clinical and ExperimentalResearch, 10, pp. 226-232.
31. PHILLIPS, S. C . & CRAGG, B . G . (1983) Chronicconsumption of alcohol hy adult mice: effect onhippocampal cells and synapses, Experimental Neu-rology, 80, pp. 218-226.
32. PHII.I.I|'S, S . C . & CRAGG, B . G . (1984) Alcoholwithdrawal causes a toss of cerebellar Purkinjecells in mice, Journal of Studies on Alcohol, 45, pp.475-480.
33. McMuLLKN, P. A., SAINT-CYR, J. A. & CARLI^N,
P. L. (1984) Morphological alterations in rat CAlhippocampal pyramidal cell dendriles resultingfrom chronic ethanol consumption and with-drawal. Journal of Comparative Neurology, 225, pp.111-118.
34. PHILLIPS, S . C . (1985) Qualitative and quantitativechanges of mouse cerebellar synapses after chronicalcohol consumption and withdrawal. ExperimentalNeurology, 88, pp. 748 756.
35. KING, M . A., HUNTER, B . E . & WALKHR, D . W .
(1988) Alterations and recovery of dendritic spinedensity in rat hippocampus following long-termethanol ingestion. Brain Research, 459, pp. 381-385.
36. DuRAND, D,, SAIN"I-CVR, J . A. , GURKVICH, N . &
CARLKN, P . L . (1989) Ethanol-induced dendriticalterations in hippocampal granule cells, BrainResearch, 477, pp. 373-377.
37. EcKENllOhH, M. F. & RAKIC, P. (1984) Radialorganization of the hippocampal dentate gyrus: aGolgi, ultrastructural, and immunocytochemicalanalysis in the developing rhesus Monkey, Journalof Comparative Neurology, 223, pp. 1--21.
38. ZiMMKR, J. & StiNDK, N. (1984) Neuropeptidesand astroglia intracerebral hippocampal trans-plants: an immunohistochemical study in the rat.Journal of Comparative Neurology, 227, pp. 331-347.
39. STERNBERGKR, L . A. (1979) Immunocytochemistry,2nd edn (New York, John Wiley and Sans).
40. STERIO, D . C . (1984) The unbiased estimation ofnumber and sizes of arbitrary particles using thedisscloi. Journal of Microscopy, 134, pp. 127-136.
41. BRAENDGAARD, H. & GuNDERSEN, H. J. G. (1986)TTie impact of recent stereological advances onquantitative studies of the nervous system, Journalof Neuroscience Methods, 18, pp. 39-78.
42. D E HOHH, R. T . & RHINES, E . N . (1961) Cited byE. R. Wl-.lBEt., R. P, BoLHNDKR (1973) Stereo-logical techniques for electron microscopy mor-phometry, in: HAYAT, M . A. (Ed.) Principles andTechniques of Electron Microscopy, vol. 3, pp. 239-296 (New York, Van Nostrand-ReinholdCompany).
43. HAUG, H . (1967) Cited by E. R. Wi-itiHi., R. P.BOLENDER (1973) Stereological techniques forelectron microscopy morphometry, in: HAYAT,M. A. (Ed.) Principles and Techniques of ElectronMicroscopy, vol. 3, pp. 239-296 (New York, VanNostrand-Reinhold Company).
44. OvHRDijK, J., UYLINGS, H . B . M . , KL-YPERS, K . &
KA,MS TRA, A. W. (1978) An economical semi-auto-matic system for measuring cellular tree structuresin three dimensions, with special emphasis onGolgi-impregnated sections, Journal of Microscopy,114, pp. 271-284.
45. UYLINGS, H . B . M . , RUIZ-MARCOS, A. & VAN
PELT, J. (1986) The metric analysis of three-dimensional dendritic tree patterns: a methodolog-ical review, youmal of Neuroscience Methods, 18, pp.127-151.
46. D E RuriT.R, J. P. & UYLINGS, H . B . M . (1987)Morphometric and dendritic analysis of fascia den-tata granule cells in human aging and seniledementia. Brain Research, 402, pp. 217-229.
47. ANDRADE, J. P., FKRNANDO, P. M., MADEIRA,
M. D., PAULA-BARBOSA, M . M . , CADETii-LEiTE, A.& ZiM.MER, J. (1992) Effects of chronic alcoholconsumption and withdraw] on the somatostatin-immunoreactive neurons of the rat hippocampaldentate hilus. Hippocampus, 2, pp. 65-72.
48. BI;GI.I-:ITER, H . & PORJKSZ, B . (1977) Persistence ofbrain hyperexcitatability following chronic alcoholexposure in rats. Advances in Experimental Medicineand Biology, 85B, pp. 209-222.
49. BEGt.[-rrER, H., D E NOBLH, V. & PORJRSZ, B .
(1980) Protracted brain dysfunction after alcoholwithdrawal in monkey, Advances in ExperimentalMedicine and Biology, 126, pp. 237-250.
50. LESCAUDRON, L . & VERNA, A. (1985) Effects ofchronic ethanol consumption on pyramidal neu-rons of the mouse dorsal and ventralhippocampus: a quantitative histological analysis.Experimental Brain Research, 58, pp. 362-367.
51. BESSON, J. A . O . , GLEN, A. I. M., FOREMAN, E . I.,
MACDONALD, A. , S.MITH, E . W . , HUTCHINSON,
J. M. C , MALIARI:), J. R. & AsHCRorr, G. M.(1981) Nuclear magnetic resonance observationsin alcoholic cerebral disorder and the role of vaso-pressin. Lancet, 4, pp. 923 924.
52. SMITH, M . A., CHICK, J., KEAN, D . M . , DOUGIAS,
R. H. B., SINGER, A., KENDALL, R. E . & BEST,
J. J. K. (1985) Brain water in chronic alcoholicpatients measured by magnetic resonance imaging.Lancet, 1, pp. 1273-1274.
53. HL-N-r, W. A. & MAJCHROWICZ, E . (1979) Alter-ations in neurotransmitter function after acute andchronic treatment with ethanol, in: MAJCHROWICZ,E. & NOBLE, E . P. (Eds) Biochemistry and Pharma-cology of Ethanol, vol. 2, pp. 167-185 (Nevf York,Plenum Press).
54. LEONARD, B . E . (1986) Is ethanol a neurotoxin?Effects of ethanol on neuronal structure and func-tion. Alcohol and Alcoholism, 21, pp. 325-338.
55. LESCAUDRON, L . , SEGUKIA, P., GI;I-HARI>, M . &
VERNA, A. (1986) Effects of long-term ethanolconsumption on GABAergic neurons in the mousehippocampus: a quantitative immunocytochemicaistudy. Drug and Alcohol Dependence, 18, pp. 377-384.
56. Sl.ovlTER, R. S. (1987) Decreased hippocampalinhibition and a selective loss of interneurons inexperimental epilepsy. Science, 235, pp. 73-76.
57. D E LANEROLLE, N. C , KiM,J. H., ROBBINS, R. J. &
SPENCER, D . D . (1989) Hippocampal intemeuron

Alcohol and hippocampal neurons and synapses 247
loss and plasticity in human temporal lobeepilepsy, Brain Research, 495, pp. 387-395.
58. CHAN-PAIJ\Y, V. (1987) Snmatostatin immuno-reactive neurons in tho human hippocampus andcortex shown by immunogold/silver intensificationon vibratome sections: coexistence with neuropep-tidc Y neurons, and efFects in Alzheimer-typedementia, Journal of Comparaiive Neurologyt 260,pp. 201 223.
59. PliN-l-NHV, R. J. & QUACKJ-NBUSH, L. J, (1990)Dendritic hypenrophy in Purkinje neurons of oldFisher 344 rats after long-term ethanol treatment,Alcoholism: Clinical and Experimental Research, 14,pp. 878-886.
60. LESCAUDRON, I.., JAFFARD, R. & VI;RNA, A.
(1989) Modficaiions in number and morphologyof dendritic spines resulting from chronicethanol consumption and withdrawal: a Golgistudy in the mouse anterior and posteriorhippocampus. Experimental Neurology, 106, pp.156-163.
61. DhSMOND.N. L. & LEVY, W. B. (1986) Changes inthe numerical density of synaptic contacts withlong-term potentiation in the hippocampal dentategyrus, Journal of Comparative Neurology, 253, pp.466-475.
