effects of chronic alcohol consumption and withdrawal on the somatostatin-immunoreactive neurons of...
TRANSCRIPT

HIPPOCAMPUS, VOL. 2, NO. 1, PAGES 65-72, JANUARY 1992
Effects of Chronic Alcohol Consumption and Withdrawal on the Somatostatin-immunoreactive Neurons of the Rat
Hippocampal Dentate Hilus Jos6 P. Andrade," Paulo M. Fernando,"
Maria D. Madeira," Manuel M. Paula-Barbosa,* Antonio Cadete-Leite," and Jens Zimmer-i-
*Department of Anatomy, Porto Medical School, 4200 Porto, Portugal, tlnstitute of Neurobiology , University of Aarhus, DK-8000 Aarhus, Denmark
Abstract Previous studies have demonstrated that the dentate granule and the CA3 pyramidal cells of
the rat hippocampal formation are neuronal populations vulnerable to the toxic effects of ethanol. It also has been shown that the resulting alterations do not end after withdrawal from ethanol. As the neurons in the dentate hilus are heavily interconnected with the dentate granule cells, the authors decided to examine the fate of the hilar neurons after chronic alcohol consumption and withdrawal, inasmuch as the hilar somatostatin-immunoreactive (SS-I) neurons were found to be sensitive to cerebral ischemia and to seizures.
The following groups of adult rats were studied: (1) alcohol-fed for 6 and 12 months; ( 2 ) alcohol- fed for 6 months and then switched to water for a further 6 months: (3) pair-fed controls; and (4) controls fed ad libitum. The authors determined the numerical density of hilar neurons and the number of its SS-I subpopulation. These were found to be significantly reduced in both the alcohol-fed and withdrawal groups when compared with the respective age-matched controls. The consequent loss of the integrative action of the hilar neurons, including the SS-Is, could explain some of the alcohol-related functional deficits as well as their persistence after with- drawal.
Key words: fascia dentata, ethanol, imrnunocytochernistry, morphometry
It has become evident over the past few years that pro- longed alcohol intake leads to permanent morphological al- terations in the central nervous system (CNS). The hippo- campal formation of rodents has been found to be particularly sensitive to this neurotoxic agent, as numerous morpholog- ical (Walker et al., 1980; Phillips and Cragg, 1983; Lescaudron and Verna, 1985; Lescaudron et al., 1986; Andrade et al., 1988; Cadete-Leite et al., 1988a; 1988b; 1989b) and functional (Walker et al., 1981; 1982; Arendt et al., 1989) studies have shown. In rats submitted to long-term alcohol consumption we observed a loss of hippocampal dentate granule and CA3 pyramidal cells that was related to the length of the exposure to alcohol (Cadete-Leite et al., 1988b; 1989b). In parallel to the neuronal cell loss, a reduction of mossy fiber-CA3 syn- apses was observed (Cadete-Leite et al., 1989b). Rather surprisingly, we also noted that withdrawal from alcohol, instead of stopping these alterations, induced an increased
Correspondence and reprint requests to J. P. Andrade, Department of Anatomy, Porto Medical School, Alameda Prof. Hemhi Mon- teiro, 4200 Porto, Portugal.
loss of neurons and synapses (Cadete-Leite et al., 1988a; 1989b).
Because both dentate granule and hippocampal CA3 py- ramidal cells were affected, we found it worthwhile to extend our studies to the closely related neurons in the dentate hilus (Amaral, 1978), particularly because these cells are consid- ered to play a major role in the regulation of the dentate gran- ule cell activity (Amaral and Witter, 1989). Although hilar neurons are morphologically heterogeneous (Amaral, 1978), it is possible, by immunocytochemical staining, to define among them a somatostatin-immunoreactive (SS-I) subpo- pulation (Zimmer et al., 1983; Zimmer and Sunde, 1984; Bakst et al., 1986; L&rAnth et al., 1990) that is highly susceptible to transient cerebral ischemia (Johansen et al., 1987; Tonder et al., 1990) and seizures (Sloviter, 1987). Because seizures (Sloviter, 1987) are commonly observed during withdrawal after long periods of ethanol intake (Walker et al., 1981; Maier and Pohorecky, 1989), we thought it would be of interest to quantify the dentate hilar cells and their SS-I subpopulation, using the model we developed (Cadete-Leite et al., 1988b; 1989a; 1989b) for evaluating the effects of long-term alcohol consumption and withdrawal on rodents.
65

66 HZPPOCAMPUS VOL. 2, NO. 1, JANUARY 1992
MATERIALS AND METHODS
Animals, diets, and treatments
Two-month-old male Sprague-Dawley rats, weighing 200 * 20 g, were obtained from the colony of the Gulbenkian Institute of Science (Oeiras, Portugal). The animals were in- dividually housed, and seven groups of four to six rats each were treated as follows.
Alcohol-fed rats had unrestricted access to a 20% (vol/vol) aqueous ethanol solution for periods of 6 and 12 months; food and fluid intake were measured every other day, and the amounts consumed were calculated. Control rats had free access to food and water through the entire experimental period. Pair-fed control rats were given the same amount of food consumed by alcohol-fed animals. Sucrose was added to the water to replace the caloric intake provided by ethanol; the percentage of sucrose in the fluid was calculated taking into account the caloric values of both alcohol (1 g-7 kcal) and sucrose (1 8-4 kcal) as well as the volume of ethanol consumed by alcohol-fed animals (10 mL of ethanol; 7,9 9). Withdrawal rats were alcohol-fed for 6 months and then switched to standard chow and water for another period of 6 months (Cadete-Leite et al., 1990a). The withdrawal from alcohol was performed progressively by reducing the ethanol concentration in the drinking solution gradually over a 2-week period.
A graded withdrawal was performed to minimize the symp- toms~ and lesions that might occur following an acute absti- nence from alcohol (Walker et al., 1981; Maier and Poho- recky, 1989), allowing the rats an adjustment to their new metabolic conditions (Eckardt et al., 1986).
In all groups, the drinking fluid supplied was supplemented with vitamins and salts.
Tissue preparation
For the estimation of the entire number of the hilar neu- rons, the animals were anesthetized with ether and perfused with a mixture of 1% glutaraldehyde and 1% paraformalde- hyde in 0.12 M phosphate buffer at pH 7.2 (Palay and Chan- Palay, 1974). The brains were removed, coded to allow sub- sequent analysis without knowledge of the group designation, and immersed for 2 hours in the fixative solution. The hip- pocampal formations were then separated from the overlying cortex and cut perpendicular to the septotemporal axis. Blocks from the midtemporal part of each hippocampal for- mation were processed as though being prepared for electron microscopy and embedded in Epon. From each block, sem- ithin sections of nominal thickness (2 Fm) were stained with toluidine blue.
For immunocytochemistry, the animals were perfused with approximately 300 mL of 4% paraformaldehyde in 0.15 M phosphate buffer at pH 7.4. The brains were then removed in toto and placed in the fixative for 2 hours (Zimmer and Sunde, 1984). The brains were then cut in 50-pm thick coronal sections on a Vibratome. Sections containing the midtem- poral level of the hippocampus and fascia dentata were then
selected under the stereomicroscope and processed immu- nocytochemically according to the peroxidase-antiperoxi- dase method (Sternberger, 1979). A rabbit antiserum against somatostatin (DAKO, Copenhagen, Denmark) was used as a primary antibody at a dilution of 1 : 2,500 in Tris-NaCI buffer with 0.5% Triton X-100. Free-floating sections were incu- bated overnight at 4°C. The peroxidase activity was visual- ized by incubation of the sections in 0.025% diaminobenzi- dine and 0.05% hydrogen peroxidase for 15 minutes. Some of the sections were counterstained with toluidine blue. All sections were finally mounted on glass slides, dehydrated, and coverslipped in a synthetic resin (Eukitt).
For control of the specificity of the immunostaining, sec- tions were incubated in the solutions of the primary antibody to which somatostatin (Clin-Midi) had been added 24 hours earlier in amounts of 100 pg/mL. None of the sections treated with this solution showed any neuronal or neuropil staining, leaving only staining of erythrocytes and macrophage-like cells.
In two different sets of animals, the morning and evening blood ethanol concentrations were enzymatically determined using a Boehringer-Mannheim kit.
Quantitative procedures
The number of hilar cells per unit volume (Nv , numerical density) was calculated by applying the formula of De Hoff and Rhines (1961) modified by Haug (1967):
"4 D + T - 2 h ' Nv =
in which NA is the number of hilar cells per unit area of section (areal density), D is the mean diameter of these cells, T is the section thickness, and h is the smallest nuclear diameter measured. The quantifications were performed in the 2-pm thick toluidine blue-stained sections obtained from three dif- ferent blocks per animal. The number of cells per unit area of the dentate hilus was estimated by counting the nuclear profiles of the hilar cells observed within the boundaries of an adequate plastic replica, using a camera lucida at a mag- nification of x 100. For the evaluation of the mean nuclear diameters, a minimum of 100 hilar cell nuclei was drawn from the same semithin sections, using a camera lucida at a mag- nification of x 340. The mean nuclear diameters were mea- sured from the drawings with the aid of a MOP-Videoplan.
The number of SS-I cell bodies was quantified in the 50- km thick Vibratome sections using a x 10 plan objective and a camera lucida. The boundaries of the hilus of the fascia dentata and the enclosed part of CA3 pyramidal cell layer were drawn. The positions of the SS-I cell bodies were plotted and later counted from the drawings. The area of the hilus in which the neurons were counted was measured with a MOP- Videoplan, and the results expressed as the mean number of SS-I cells observed in 7-10 slices per animal. Comparisons of the number of SS-I cells were made between sections at the midtemporal level with similar areas.
As brain tissue undergoes volumetric changes during his- tological processing, the tissue shrinkage factor (SFv) of the brains processed for immunocytochemistry was calculated in

HIPPOCAMPAL HILAR NEURONS AFTER ALCOHOL INTAKE AND WITHDRAWAL / Andrade et al. 67
separate groups of alcohol-fed and control rats. The SFV was calculated following a previously described methodology (Uylings et al., 1986; Madeira et al., 1990; 1991). The SFV of Epon-embedded material was not calculated because it is al- most nonexistent (West et al., 1988; Schiiz and Palm, 1989).
Statistical analysis
The quantitative results were expressed as mean 5 SD. The quantitative data were analyzed for statistical signifi- cance between groups using the two-tailed nonparametric Mann-Whitney U-test. Differences were considered signifi- cant if P < .05.
RESULTS
Animals and diets
Detailed data concerning animals and diets have been re- ported previously (Cadete-Leite et al., 1988b; 1990a). The average daily ethanol intake was approximately 9.0 glkg body weight. During the first 3 months, however, the mean intake was lower than 9.0 g/kg; it attained a mean of 13.0 g/kg after 12 months of ingestion, which corresponds to a mean value of 18 mL of daily aqueous ethanol intake (2.8 g of ethanol; 19.5 kcal) .
The water provided to pair-fed controls contained 11.7% (wthol) of sucrose (4.8 g; 19.5 kcal). The amount of sucrose was calculated with the average volume of water consumed by the pair-fed controls at 41 mL per day. We are aware that for correct pair-feeding the amount of fluid ingested by con- trols should be the same as that drunk by the respective al- cohol-treated rats. This can be achieved only using liquid diets, which, in long-term models like this one, induce marked collateral changes (A. Cadete-Leite, unpublished ob- servations).
The mean blood ethanol concentration was found to be 101.4 * 12.7 mg/dL. The values found ranged between 80 and 120 mg/dL. The highest concentrations were always ob- served in the blood samples collected in the morning (Cadete- Leite et al., 1990a).
During the first month of the experiment, the body weights of alcohol-fed animals remained unchanged, whereas the mean body weight increase of controls and pair-fed animals was 16.3 g and 12.2 g, respectively. Throughout the experi- ment, the body weights of alcohol-fed and withdrawn rats did not differ from those of age-matched pair-fed controls and were significantly smaller than those of controls. At the end of the experiment, the body weights of alcohol-fed animals were lower than those of withdrawal and pair-fed control rats, although the differences were not significant. Conversely, the mean body weights of the animals from the above-mentioned groups were significantly smaller than those found in con- trols.
Qualitative observations
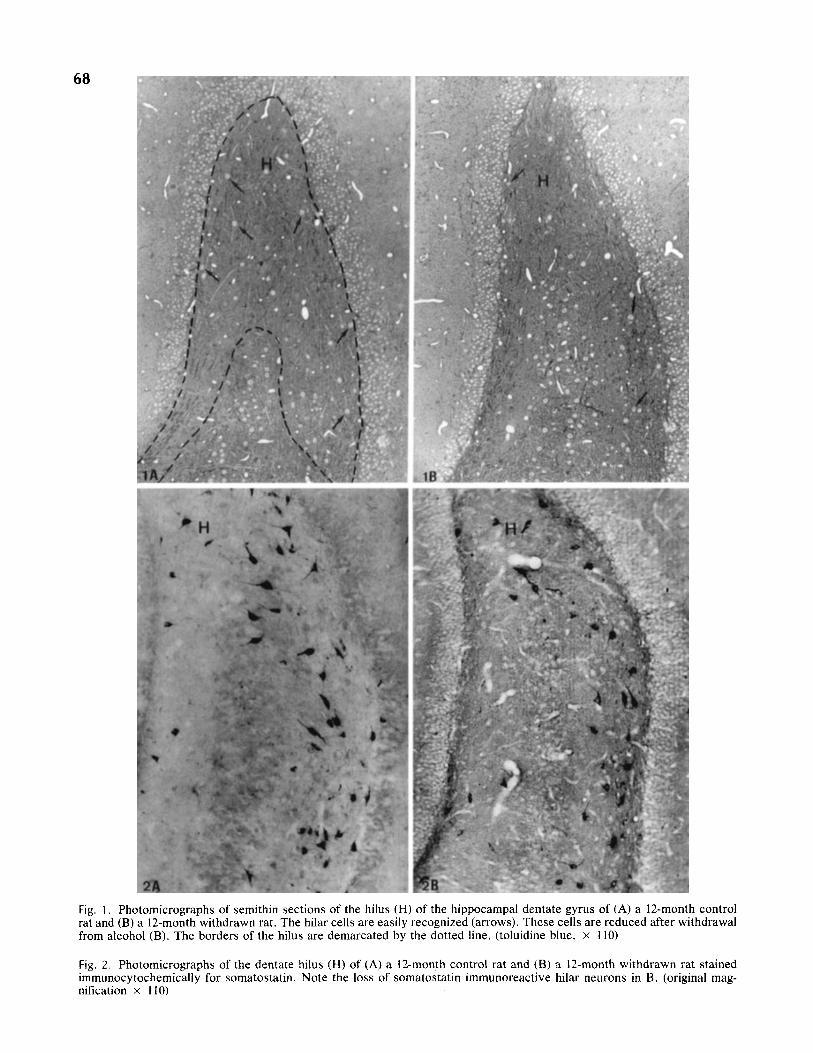
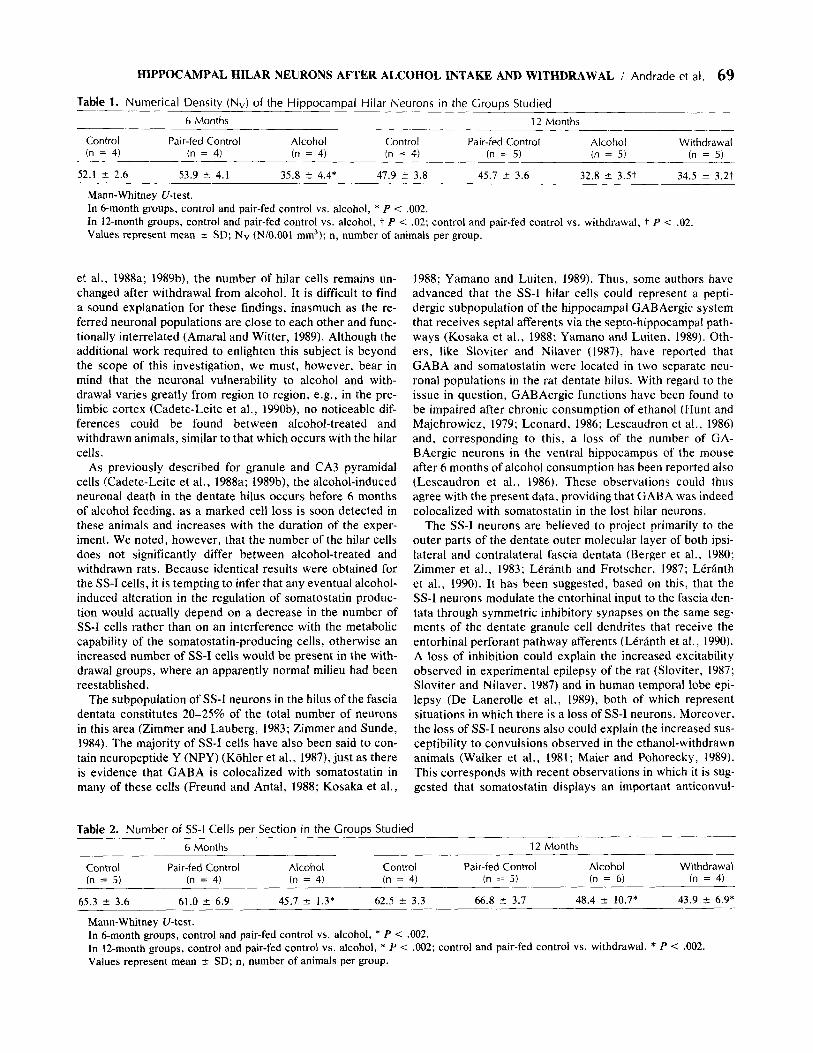
In most of the sernithin sections from 6- and 12-month al- cohol-fed and withdrawn rats, a loss of hilar cells could be recognized qualitatively (Fig. 1 A, B). In immunocytochem- ically stained material, a loss of SS-I neurons was also ap- parent in the referred groups (Fig. 2A, B). Since the soma-
tostatin-immunoreactivity is almost exclusively confined to the cell body and proximal parts of the dendrites, it was dif- ficult to classify the stained cells morphologically as belong- ing to one or the other of the known different types of hilar neurons (Amaral, 1978).
Quantitative results
The results of the quantitative analysis are shown in Tables 1 and 2.
The numerical densities of hilar cells from 6- and 12-month alcohol-treated rats were significantly reduced when com- pared with those from the respective control and pair-fed con- trol groups. A significant reduction in the numerical density of hilar cells was also found when the withdrawal group was compared with the 12-month control and pair-fed control groups. A nonsignificant reduction in the numerical density of hilar neurons was observed between 6- and 12-month al- cohol-fed rats. No significant differences were observed in the numerical density of hilar cells between 12-month alcohol- fed and withdrawn rats, as well as between controls and pair- fed controls (Table 1).
A significant reduction in the number of SS-I neurons was detected when 6- and 12-month alcohol-fed rats were com- pared with the respective controls. Furthermore, in the with- drawal group the number of SS-I hilar cells was significantly smaller than that of the 12-month control and pair-fed control groups. N o significant differences were observed in the num- ber of SS-I neurons between alcohol-fed and withdrawn rats. As mentioned above for the numerical densities of hilar cells (Table l), no significant differences were observed in the number of SS-I neurons between controls and pair-fed con- trols (Table 2).
The shrinkage factor calculated was 1.3 for both control and alcohol-fed rats.
DISCUSS I ON
In previous studies of the hippocampal formation of the rat, we reported a loss of dentate granule and CA3 pyramidal cells following prolonged alcohol consumption (Cadete-Leite et al., 1988b; 1989b). Also, the inclusion of withdrawal groups in our experiments enabled us to demonstrate that the re- ferred cell loss does not stop after withdrawal from alcohol (Cadete-Leite et al., 1988a; 1989b). Indeed, instead of evi- dence of any kind of regenerative activity that could be re- garded as a compensatory mechanism for the structural dam- age inflicted by the alcohol aggression, we found an increased neuronal degeneration (Cadete-Leite et al., 1989a; 1989b). Hence, our results do not provide a morphological basis for the reported partial functional recovery observed following abstinence from alcohol (Carlen et al., 1978; 1986).
We now demonstrate that neurons from the dentate hilus are also vulnerable to alcohol administration, which indicates that ethanol has a widespread neurotoxic action upon the dif- ferent cell types of the hippocampal formation. The SS-I neu- rons that constitute a subpopulation of the hilar cells are clearly affected, although it was not possible in the present experiment to determine whether the loss of SS-I neurons covered the entire hilar cell loss. Still, and as opposed to that which occurs with granule and pyramidal cells (Cadete-Leite
68
Fig. 1. Photomicrographs of semithin sections of the hilus (H) of the hippocampal dentate gyrus of (A) a 12-month control rat and (B) a 12-month withdrawn rat. The hilar cells are easily recognized (arrows). These cells are reduced after withdrawal from alcohol (B). The borders of the hilus are demarcated by the dotted line. (toluidine blue, x 110)
Fig. 2. Photomicrographs of the dentate hilus (H) of (A) a 12-month control rat and (B) a IZmonth withdrawn rat stained immunocytochemically for somatostatin. Note the loss of somatostatin immunoreactive hilar neurons in B. (original mag- nification x 110)
HIPPOCAMPAL HILAR NEURONS AFTER ALCOHOL INTAKE AND WITHDRAWAL / Andrade et al. 69
Table 1. Numerical Density (Nv) of the Hippocampal Hilar Neurons in the Groups Studied 6 Months 12 Months
Control Pair-fed Control Alcohol Control Pair-fed Control Alcohol Withdrawal (n = 4) (n = 4) (n = 4) (n = 4) (n = 5) (n = 5) (n = 5)
52.1 2 2.6 53.9 2 4.1 35.8 i 4.4* 47.9 f 3.8 45.7 f 3.6 32.8 f 3.5t 34.5 ? 3.2t
Mann-Whitney U-test. In 6-month groups, control and pair-fed control vs. alcohol, * P < ,002. In 12-month groups, control and pair-fed control vs. alcohol, t P i .02; control and pair-fed control vs. withdrawal, t P < .02. Values represent mean k SD; Nv (N/0.001 mm3); n , number of animals per group.
et al., 1988a; 1989b), the number of hilar cells remains un- changed after withdrawal from alcohol. It is difficult to find a sound explanation for these findings, inasmuch as the re- ferred neuronal populations are close to each other and func- tionally interrelated (Amaral and Witter, 1989). Although the additional work required to enlighten this subject is beyond the scope of this investigation, we must, however, bear in mind that the neuronal vulnerability to alcohol and with- drawal varies greatly from region to region, e.g., in the pre- limbic cortex (Cadete-Leite et al., 1990b), no noticeable dif- ferences could be found between alcohol-treated and withdrawn animals, similar to that which occurs with the hilar cells.
As previously described for granule and CA3 pyramidal cells (Cadete-Leite et al., 1988a; 1989b), the alcohol-induced neuronal death in the dentate hilus occurs before 6 months of alcohol feeding, as a marked cell loss is soon detected in these animals and increases with the duration of the exper- iment. We noted, however, that the number of the hilar cells does not significantly differ between alcohol-treated and withdrawn rats. Because identical results were obtained for the SS-I cells, it is tempting to infer that any eventual alcohol- induced alteration in the regulation of somatostatin produc- tion would actually depend on a decrease in the number of SS-I cells rather than on an interference with the metabolic capability of the somatostatin-producing cells, otherwise an increased number of SS-I cells would be present in the with- drawal groups, where an apparently normal milieu had been reestablished.
The subpopulation of SS-I neurons in the hilus of the fascia dentata constitutes 20-25% of the total number of neurons in this area (Zimmer and Lauberg, 1983; Zimmer and Sunde, 1984). The majority of SS-I cells have also been said to con- tain neuropeptide Y (NPY) (Kohler et al., 19871, just as there is evidence that GABA is colocalized with somatostatin in many of these cells (Freund and Antal, 1988; Kosaka et al.,
Table 2. Number of SS-I Cells per Section in the Groups Studied
1988; Yamano and Luiten, 1989). Thus, some authors have advanced that the SS-I hilar cells could represent a pepti- dergic subpopulation of the hippocampal GABAergic system that receives septa1 afferents via the septo-hippocampal path- ways (Kosaka et al., 1988; Yamano and Luiten, 1989). Oth- ers, like Sloviter and Nilaver (1987), have reported that GABA and somatostatin were located in two separate neu- ronal populations in the rat dentate hilus. With regard to the issue in question, GABAergic functions have been found to be impaired after chronic consumption of ethanol (Hunt and Majchrowicz, 1979; Leonard, 1986; Lescaudron et al., 1986) and, corresponding to this, a loss of the number of GA- BAergic neurons in the ventral hippocampus of the mouse after 6 months of alcohol consumption has been reported also (Lescaudron et al., 1986). These observations could thus agree with the present data, providing that GABA was indeed colocalized with somatostatin in the lost hilar neurons.
The SS-I neurons are believed to project primarily to the outer parts of the dentate outer molecular layer of both ipsi- lateral and contralateral fascia dentata (Berger et al., 1980; Zimmer et al., 1983; Leranth and Frotscher, 1987; Leranth et al., 1990). It has been suggested, based on this, that the SS-I neurons modulate the entorhinal input to the fascia den- tata through symmetric inhibitory synapses on the same seg- ments of the dentate granule cell dendrites that receive the entorhinal perforant pathway afferents (Leranth et al. , 1990). A loss of inhibition could explain the increased excitability observed in experimental epilepsy of the rat (Sloviter, 1987; Sloviter and Nilaver, 1987) and in human temporal lobe epi- lepsy (De Lanerolle et al., 1989), both of which represent situations in which there is a loss of SS-I neurons. Moreover, the loss of SS-I neurons also could explain the increased sus- ceptibility to convulsions observed in the ethanol-withdrawn animals (Walker et al., 1981; Maier and Pohorecky, 1989). This corresponds with recent observations in which it is sug- gested that somatostatin displays an important anticonvul-
6 Months 12 Months
Control Fair-fed Control Alcohol Control Pair-fed Control Alcohol Withdrawal (n = 5) (n = 4) (n = 4) (n = 4) (n = 5) (n = 6 ) (n = 4)
65.3 f 3.6 61.0 f 6.9 45.7 2 1.3* 62.5 t 3.3 66.8 .f 3.7 48.4 -t 10.7* 43.9 ir 6.9*
Mann-Whitney U-test. In 6-month groups, control and pair-fed control vs. alcohol, * P < ,002. In 12-month groups, control and pair-fed control vs. alcohol, * P < ,002; control and pair-fed control vs. withdrawal, * P < .002. Values represent mean f SD; n, number of animals per group.

70 HZPPOCAMPUS VOL. 2, NO. 1, JANUARY 1992
sivant role and that a decrease in somatostatin would lead to increased neuronal excitability (Manfridi et al., 1991). A se- lective loss of SS-I and NPY-I hippocampal neurons has been observed also in brains from patients with Alzheimer’s dis- ease (Chan-Palay, 1987), a finding that may be consistent with a role played by these cerebral peptides in cognitive functions (Chan-Palay, 1987; Haroutunian et al., 1987). These assump- tions are in keeping with the recent report of a reduction of cerebrospinal fluid somatostatin-like immunoreactivity in both Alzheimer’s disease and dementia associated with al- coholism (Koponen et al., 1990).
Bearing these observations in mind, it is likely that the loss of SS-I neurons, added to the loss of other hippocampal neu- rons, could contribute to the neurophysiological and behav- ioral deficits associated with alcohol abuse that are main- tained even after withdrawal from ethanol (Walker et al., 1981; 1982; Maier and Pohorecky, 1989).
ACKNOWLEDGMENTS
The authors are indebted to one anonymous referee whose criticisms and linguistic corrections very much improved the manuscript. This work was granted by Junta Nacional de In- vestiga@o Cientifica e Tecnologica (JNICT) Project PMCTI C/SAU/32/90, NATO Grant 91/89, and Instituto Nacional de InvestigaGBo Cientifica (INIC).
References
Amaral, D. G. (1978) A Golgi study of cell types in the hilar region of the hippocampus in the rat. J . Comp. Neurol. 182:851-914.
Amaral, D. G . , and M. P. Witter (1989) The three-dimensional or- ganization of the hippocampal formation: A review of anatomical data. Neuroscience 31571-591.
Andrade, J . P., A. Cadete-Leite, M. M. Paula-Barbosa, B. Volk, and M. A. Tavares (1988) Long-term alcohol consumption reduces the number of neuronal nuclear pores: A morphometric study under- taken in CA3 hippocampal pyramids of rats. Alcohol. Clin. Exp. Res. 12:286-289.
Arendt, T., Y. Allen, R. M. Marchbanks, M. M. Schugens, J . Sinden, P. L . Lantos, and J . A. Gray (1989) Cholinergic system and memory in the rat: Effects of chronic ethanol, embryonic basal forebrain brain transplants and excitotoxic lesions of cholinergic basal fore- brain projection system. Neuroscience 33:435-462.
Bakst, I., C. AvendBno, J . H. Morrison, and D. G . Amaral (1986) An experimental analysis of the origins of the somatostatin im- munoreactive fibers in the dentate gyrus of the rat. J. Neurosci. 6: 1452-1462.
Berger, T. W., S. Semple-Rowland, and J. L . Basset (1980) Hippo- campal polymorph neurons are the cells of origin for ipsilateral association and commissural afferents to the dentate gyrus. Brain Res. 215:329-336.
Cadete-Leite, A., M. A. Tavares, and M. M. Paula-Barbosa (1988a) Alcohol withdrawal does not impede hippocampal granule cell pro- gressive loss in chronic alcohol-fed rats. Neurosci. Lett. 86:45-50.
Cadete-Leite, A., M. A. Tavares, H. B. M. Uylings, and M. M. Paula- Barbosa (1988b) Granule cell loss and dendritic regrowth in the hippocampal dentate gyrus of the rat after chronic alcohol con- sumption. Brain Res. 473:l-14.
Cadete-Leite, A., M. A. Tavares, M. C. Alves, H. B. M. Uylings, and M. M. Paula-Barbosa (1989a) Metric analysis of hippocampal granule cell dendritic trees after alcohol withdrawal in rats. Al- cohol. Clin. Exp. Res. 133374340.
Cadete-Leite, A., M. A. Tavares, M. M. Pacheco, B. Volk, and M.
M. Paula-Barbosa (1989b) Hippocampal mossy fiber-CA3 syn- apses after chronic alcohol consumption and withdrawal. Alcohol
Cadete-Leite, A,, M. C. Alves, M. M. Paula-Barbosa, H. B. M. Uyl- ings, and M. A. Tavares (1990a) Quantitative analysis of basal den- drites of prefrontal pyramidal cells after chronic alcohol consump- tion and withdrawal in the adult rat. Alcohol. 25:467-475.
Cadete-Leite, A, , M. C. Alves, M. A. Tavares, and M. M. Paula- Barbosa (1990b) Effects of chronic alcohol intake and withdrawal on the prefrontal neurons and synapses. Alcohol 7: 145-152.
Carlen, P. L., R. C. Wortzman, D. A. Wilkinson, J. G . Rankin (1978) Reversible cerebral atrophy in recently abstinent chronic alcoholics measured by computed tomography scans. Science 200: 1076- 1078.
Carlen, P. L., R. D. Penn, L. Fornazzari, S . Bennett, D. A. Wilk- inson, and G. Wortzman (1986) Computerized tomographic scan assessment of alcoholic brain damage and its potential reversibility. Alcohol. Clin. Exp. Res. 10:226-232.
Chan-Palay, V. (1987) Somatostatin immunoreactive neurons in the human hippocampus and cortex shown by immunogold/silver in- tensification on vibratome sections: Coexistence with neuropeptide Y neurons, and effects in Alzheimer-type dementia. J . Comp. Neu- rol. 260: 201-223.
De Hoff, R. T., and E. N. Rhines (1961) Cited by E. R. Weibel, R. P. Bolender (1973) Stereological techniques for electron micros- copy morphometry. In Principles and Techniques of Electron Mi- croscopy, vol. 3, M. A. Hayat, ed., pp. 239-296, Van Nostrand- Reinhold Company, New York, NY.
De Lanerolle, N. C., J . H . Kim, R. J. Robbins, and D. D. Spencer (1989) Hippocampal interneuron loss and plasticity in human tem- poral lobe epilepsy. Brain Res. 495:387-395.
Eckardt, M. J . , G . A. Campbell, C. A. Marietta, E. Majchrowick, H. N. Wixon, and F. F. Weight (1986) Cerebral 2-deoxyglpcose uptake in rats during ethanol withdrawal and postwithdrawal. Brain Res. 366:l-9.
Freund, T. F., and M. Antal(1988) GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature
Haroutunian, V., R. Mantin, G . A. Campbell, G . K. Tsuboyama, and K. L. Davis (1987) Cysteamine-induced depletion of central so- matostatin-like immunoactivity: Effects on behavior, learning, memory and brain neurochemistry. Brain Res. 403:234-242.
Haug, H. (1967) Cited by E. R. Weibel, R. P. Bolender (1973) Ster- ological techniques for electron microscopy morphometry. In Prin- ciples and Techniques ofElectron Microscopy, vol. 3, M. A. Hayat, ed., pp. 239-296, Van Nostrand-Reinhold Company, New York, NY.
Hunt, W. A,, and E. Majchrowicz (1979) Alterations in neurotrans- mitter function after acute and chronic treatment with ethanol. In Biochemistry and Pharmacology of Ethanol, vol. 2 , E. Majchrow- icz and E. P. Noble, eds., pp. 167-185, Plenum Press, New York, NY.
Johansen, F. F., J . Zimmer, and N. H. Diemer (1987) Early loss of somatostatin neurons in dentate hilus after cerebral ischemia in the rat precedes CAI pyramidal cell loss. Acta Neuropathol. (Berl.) 73:llO-114.
Kohler, C., L. G . Eriksson, S. Davis, and V. Chan-Palay (1987) Co- localization of neuropeptide tyrosine and somatostatin immuno- reactivity in neurons of individual subfields of the rat hippocampal region. Neurosci. Lett. 78: 1-6.
Koponen H., K. J . Reinikainen, J . T. Jolkkonen, and P. J. Riekkinen (1990) Somatostatin-like immunoreactivity in the CSF of patients with dementia associated with alcoholism. Acta Neurol. Scand.
Kosaka, T., J.-Y. Wu, and R.‘Benoit (1988) GABAergic neurons containing somatostatin-like immunoreactivity in the rat hippocam- pus and dentate gyrus. Exp. Brain Res. 71:388-398.
6~303-310.
336: 170-173.
82:289-291.

HIPPOCAMPAL HILAR NEURONS AFTER ALCOHOL INTAKE AND WITHDRAWAL / Andrade et al. 71
Leonard, B. E. (1986) Is ethanol a neurotoxin? Effects of ethanol on neuronal structure and function. Alcohol Alcohol. 21 :325-338.
LCrhnth, C., and M. Frotscher (1987) Cholinergic innervation of hip- pocampal GAD- and somatostatin-immunoreactive commissural neurons. J. Comp. Neurol. 261:33-47.
LCrhnth, C., A. J. Malcolm, and M. Frotscher (1990) Afferent and efferent synaptic connections of somatostatin-inmunoreactive neurons in the rat fascia dentata. J. Comp. Neurol. 295:lll-122.
Lescaudron, L., and A. Verna (1985) Effects of chronic ethanol con- sumption on pyramidal neurons of the mouse dorsal and ventral hippocampus: A quantitative histological analysis. Exp. Brain Res.
Lescaudron, L., P. Seguela, M. Geffard, and A. Verna (1986) Effects of long-term ethanol consumption on GABAergic neurons in the mouse hippocampus: A quantitative immunocytochemical study. Drug Alcohol Depend. 18:377-384.
Madeira, M. D., A. Pereira, A. Cadete-Leite, and M. M. Paula-Bar- bosa (1990) Estimates of volumes and pyramidal cell numbers in the prelimbic subarea of the prefrontal cortex in experimental hypo- thyroid rats. J . Anat. 171:41-56.
Madeira, M. D., A. Cadete-Leite, N. Sousa, and M. M. Paula-Bar- bosa (1991) The supraoptic nucleus in hypothyroid and under- nourished rats: An experimental morphometric study. Neurosci- ence 41:827-839.
Maier, D. M., and L. A. Pohorecky (1989) The effect of repeated withdrawal episodes on subsequent withdrawal severity in ethanol- treated rats. Drug Alcohol Depend. 23:103-110.
Manfridi, A., G. L. Forloni, A. Vezzani, F. Fodritto, and M. G. de Simoni (1991) Functional and histological consequences of quin- olinic and kainic acid-induced seizures on hippocampal somatos- tatin neurons. Neuroscience 41:127-135.
Palay, S. L., and V. Chan-Palay (1974) Cerebellar Cortex. Cytology and Organization. Springer-Verlag, Berlin.
Phillips, S . C., and B. G. Cragg (1983) Chronic consumption of al- cohol by adult mice: Effect on hippocampal cells and synapses. Exp. Neurol. 80:218-226.
Schiiz, A., and G. Palm (1989) Density of neurons and synapses in the cerebral cortex of the mouse. J. Comp. Neurol. 286:442-455.
Sloviter, R. S. (1987) Decreased hippocampal inhibition and a selec- tive loss of interneurons in experimental epilepsy. Science 235:73- 76.
Sloviter, R. S., and G. Nilaver (1987) Immunocytochemical locali-
58~362-367.
zation of GABA- cholecystokinin-, vasoactive intestinal polypep- tide-, and somatostatin-like immunoreactivity in the area dentata and hippocampus of the rat. J. Comp. Neurol. 256:42-60.
Sternberger, L. A. (1979) frnmunocytochemistiy, 2nd edition, John Wiley, New York, NY.
TZnder, N., F. F. Johansen, C. J. Frederickson, J. Zimmer, and N. H. Diemer (1990) Possible role of zinc in the selective degeneration of dentate hilar neurons after cerebral ischemia in the adult rat. Neurosci. Lett. 109:247-252.
Uylings, H. B. M., C. G . Van Eden, and M. A. Hofman (1986) Mor- phometry of sizeholume variables and comparison of their bivar- iate relations in the nervous system under different conditions. J. Neurosci. Meth. 18:19-37.
Walker, D. W., D. E. Barnes, S. F. Zornetzer, B. E. Hunter, and P. Kubanis (1980) Neuronal loss in hippocampus induced by pro- longed ethanol consumption in rats. Science 209:711-713.
Walker, D. W., B. E. Hunter, and W. C. Abraham (1981) Neuroan- atomical and functional deficits subsequent to chronic ethanol ad- ministration in animals. Alcohol. Clin. Exp. Res. 5:267-282.
Walker, D. W., B. E. Hunter, D. E. Barnes, and J. N. Riley (1982) An animal model of alcohol-induced brain damage: A behavioral and anatomical analysis. In Cerebrul Deficits in Alcoholism, D. A. Wilkinson, ed., pp. 123-147, Addiction Research Foundation, To- ronto, Ontario.
West, M. J., P. D. Coleman, and D. G. Flood (1988) Estimating the number of granule cells in the dentate gyrus with the disector. Brain Res. 448:167-172.
Yamano, M., and P. G. M. Luiten (1989) Direct synaptic contacts of medial septa1 efferents with somatostatin immunoreactive neurons in the rat hippocampus. Brain Res. Bull. 22:993-1001.
Zimmer, J., and S. Lauberg (1983) Neuronal subpopulations in the hilus of fascia dentata (CA4) revealed by combinations of retro- grade tracing, AChE-histochemistry and somatostatin-like immu- noreactivity. Neurosci. Lett. Suppl. 14:S416.
Zimmer, J., and N. Sunde (1984) Neuropeptides and astroglia in in- tracerebral hippocampal transplants: An immunohistochemical study in the rat. J. Comp. Neurol. 227:331-347.
Zimmer, J., S. Lauberg, and N. Sunde (1983) Neuroanatomical as- pects of normal and transplanted hippocampal tissue. In Neuro- biology of the Hippocampus, W. Seifert, ed., pp. 39-64, Academic Press, London.