stage-specific gene expression in early differentiating oligodendrocytes
TRANSCRIPT

Stage-Specific Gene Expression inEarly Differentiating
OligodendrocytesFRANCESCA BLASI,1 ALESSIA CIARROCCHI,1 ALICE LUDDI,2
MICHELINA STRAZZA,2 MASSIMO RICCIO,3 SPARTACO SANTI,3ROSARIA ARCONE,4 CONCETTA PIETROPAOLO,5 ROMINA D’ANGELO,1
ELVIRA COSTANTINO-CECCARINI,2 AND MARIALUISA MELLI1*1Department of Biology, Bologna University, Bologna, Italy2Centro Studio Cellule Germinali, CNR Siena, Siena, Italy
3Istituto di Citomorfologia NP, CNR, Bologna, Italy4Dipartimento di Scienze Farmaco-biologiche, Universita di Catanzaro, Magna Graecia, Catanzaro, Italy5Dipartimento di Biochimica e Biotecnologie Mediche, Universita di Napoli Federico II, Naples, Italy
KEY WORDS cell-specific mRNA; CNS; brain development; oligodendrocyte precur-sor cells; differential library
ABSTRACT The screening of a differential library from precursor and differentiatedoligodendrocytes, obtained through the representational difference analysis (RDA) tech-nique, has generated a number of cDNA recombinants corresponding to mRNA coding forknown and unknown proteins: (1) mRNA coding for proteins involved in protein synthesis,(2) mRNA coding for proteins involved in the organization of the cytoskeleton, and (3) mRNAcoding for proteins of unknown function. The expression profile of the mRNA was studied byNorthern blot hybridization to the poly-A� mRNA from primary rat progenitor and differ-entiated oligodendrocytes. In most cases, hybridization to the precursor was higher thanhybridization to the differentiated mRNA, supporting the validity of the differential screen-ing. Hybridization of the cDNA to rat cerebral hemisphere and brain stem poly-A� mRNA,isolated from 1- to 90-day-old rats, confirms the results obtained with the mRNA fromdifferentiating oligodendrocytes. The intensity of the hybridization bands decreases asdifferentiation proceeds. The pattern of expression observed in oligodendrocytes is differentfrom that found in the brain only in the case of the nexin-1 mRNA, the level of whichremains essentially constant throughout differentiation both in the brain stem and in thecerebral hemispheres, in agreement with the published data. In contrast, the intensity ofhybridization to the oligodendrocyte mRNA is dramatically lower in the differentiated cellscompared with the progenitor oligodendrocyte cells. Some of the recombinant cDNA repre-sent mRNA sequences present at high frequency distribution in the cells, while othersbelong to the rare sequences group. Six recombinants code for proteins of the ribosomalfamily, suggesting that of approximately 70 known ribosomal proteins, only a few areupregulated during oligodendrocyte differentiation. The third category of open readingframe (ORF) is represented by rare messengers coding for proteins of unknown functionsand includes six clones: RDA 279, 11, 95, 96, 254, and 288. GLIA 39:114–123, 2002.© 2002 Wiley-Liss, Inc.
INTRODUCTION
The isolation of primary cultures of oligodendrocyteprecursor cells from newborn rats or mice representsan important contribution to the study of oligodendro-cyte differentiation (Raff et al., 1983; Gard and Pfeiffer,1989). In fact, in primary cell cultures, the discretestages of oligodendrocyte differentiation from neuralstem cells to mature myelin-forming cells can be repro-duced quite faithfully. Under appropriate conditions,
the oligodendrocyte precursor cells can also revert tomultipotent stem cells (Kondo and Raff, 2000).
Grant sponsor: Istituto Superiore di Sanita; Grant sponsor: Multiple SclerosisProject; Grant sponsor: Ministero dell’Universita e della Ricerca Scientifica eTecnologica (PRIN).
*Correspondence to: Marialuisa Melli, Department of Biology, Bologna Uni-versity, I-40126 Bologna, Italy. E-mail: [email protected]
Received 18 January 2002; Accepted 22 March 2002
DOI 10.1002/glia.10092Published online 00 Month 2002. in Wiley InterScience (www.interscience.
wiley.com).
GLIA 39:114–123 (2002)
© 2002 Wiley-Liss, Inc.

The generation of the correct number of oligodendro-cytes in the brain involves several steps. Oligodendro-cyte precursors must be induced from cells of the neu-roepithelium. The precursors migrate through thecentral nervous system (CNS), driven by definedgrowth factors and signaling molecules, and undergoproliferation in the developing white matter. Once asufficient number of cells are generated, proliferation isdownregulated and oligodendrocyte precursors differ-entiate into mature oligodendrocytes (Orentas andMiller, 1998). Understanding the regulation of the in-duction, migration, and maturation of oligodendrocyteprecursors has important implications in both CNSdevelopment and functional recovery after focal demy-elinating pathologies, such as multiple sclerosis andinjury.
Although antigens specific for the various stages ofdifferentiation have been identified, little is knownabout variations in gene expression throughout oligo-dendrocyte differentiation. Changes in the transcrip-tional program of cells at different stages of differenti-ation are the basis of the modifications in cell growth,physiology, and environment. The identification ofgenes whose expression depends on the state of the cellmay bring new insights into, and help define, the com-plex networks and regulatory mechanisms underpin-ning the processes of differentiation and development.The elucidation of genome sequences specifically in-volved in a differentiation stage of a given cell wouldprovide basic information on gene expression, that canbe further studied with genetic and biochemical meth-ods. This work describes the isolation of recombinantcDNA specific to primary rat oligodendrocyte precur-sors by means of the representational difference anal-ysis (RDA) technique initially described by Hubankand Schatz (1994). Primary oligodendrocyte progeni-tors were isolated according to standard techniques, asdescribed in the Material and Methods. The purity ofthe progenitor cell population was analyzed by inter-action with the specific A2B5 antibodies. On average,92–96% of the cells were A2B5 positive. The screeningof the differential library has generated cDNA recom-binants coding for proteins involved in the process ofprotein synthesis in the organization of the cytoskele-ton, as well as a number of recombinants coding forproteins of unknown function. Northern blot hybridiza-tion of the cDNA clones to the poly-A� mRNA fromprimary rat progenitor and differentiated oligodendro-cytes confirms the differential expression of the corre-sponding messenger molecules. Data are also pre-sented on the hybridization of cDNA to cerebralhemisphere and brain stem poly-A� mRNA obtainedfrom brains of 1- to 90-day-old rats.
MATERIALS AND METHODS
All common techniques and routine DNA manipula-tions, including transformation, plasmid preparation,
and gel electrophoresis were carried out according tostandard procedures (Sambrook et al., 1989). Restric-tion and modification enzymes (New England Biolabs,Stratagene, Roche Molecular Biochemicals) were usedfollowing the manufacturers’ instructions. DNA wasextracted from agarose gel using the QIAquick SpinGel Extraction kit (QIAgen).
Cell Culture
Oligodendrocyte primary cultures
Cerebra from 3- to 4-day-old Sprague-Dawley ratsafter removal of the meninges were minced and passedsequentially through nylon meshes of 210-, 130-, and35-�m pore size (McMorris, 1983). The dissociated cellsuspension was plated on 75-cm2 flasks in OM-5 con-taining 10% fetal bovine serum (FBS; Hyclone, Logan,UT); after 7–8 days, the oligodendrocytes and oligoden-drocyte precursors were separated from the astrocytemonolayer by shaking overnight at 200 rpm on a rotaryshaker. Oligodendrocytes were purified from contami-nating astrocytes by immunopanning on plates coatedwith Ran2 antibodies. Oligoprogenitors were plated onpoly-D-lysine-coated dishes and maintained in OM-6medium containing 30% conditioned medium fromB104 neuroblastoma cells; they were grown in the pres-ence of 10 ng/ml platelet-derived growth factor-AA(PDGF-AA) and of 10 ng/ml basic fibroblast growthfactor (bFGF) (Peprotech). Oligoprogenitors were dif-ferentiated in mature oligodendrocytes by culturingthem for 5 days on poly-D-lysine-coated dishes in OM-6medium containing 0.5% serum (Brogi et al., 1997).Oligoprogenitors were differentiated into type 2 astro-cytes, as previously described (Raible and McMorris,1993).
RNA Preparation and cDNA Synthesis
Total RNA was prepared from cells and tissues ac-cording to Chomczynski and Sacchi (1987), and poly-A�
RNA was isolated by double selection over an oligo-dTcellulose column (Ausubel et al., 1989). Double-stranded cDNA was prepared using oligo-dT primers(Ausubel et al., 1989). To assess the size of cDNA, on analiquot from the second-strand reaction, the synthesiswas placed in the presence of �-32P dATP. The reactionproduct was electrophoresed on a denaturing agarosegel together with a 32P terminally labeled DNA molec-ular weight marker, and exposed to an X-ray film(Kodak X-Omat AR).
Generation of the Subtraction cDNA Library byRepresentational Difference Analysis
After polymerase chain reaction (PCR) amplification,we subtracted the differentiated oligodendrocyte cDNA
115CELL-SPECIFIC mRNA IN CNS DURING BRAIN DEVELOPMENT
(driver) from oligodendrocyte precursors cDNA (tester)according to the representational difference analysis(RDA) technique described by Hubank and Schatz(1994). We introduced the following modifications.First, to avoid the loss of sequences that may containonly one DpnII site or none, the blunt-ended cDNA wasligated to a DNA adaptor containing a DpnII site. Sec-ond, as the ideal conditions for the amplification of eachmRNA may not be the same for all sequences, wecarried out the PCR amplifications under four differentconcentrations of Mg2�: 1.5, 2, 2.5, 3.5 mM Mg2�. Aftereach PCR reaction, the four products were pooled anddigested with the DpnII restriction endonuclease, ac-cording to the protocol. Third, the subtractive hybrid-ization was carried out in 3 �l in 0.3 M Na�, at 67°C,for 70 h. After three rounds of subtractive hybridiza-tion, the RDA products were DpnII-digested and clonedinto the BamHI site of the pBluescript SK II� (Strat-agene).
We checked the validity of the technique by adding atrace amount of the adenovirus E1A gene, to the initialdouble-stranded cDNA from progenitor cells; 10 pg of a500-bp DNA fragment of the adenovirus type 5 cDNAE1A12S (Chroboczek et al., 1992) was added to 1 �g ofoligodendrocyte precursor cDNA, before the ligation tothe DpnII adaptors.
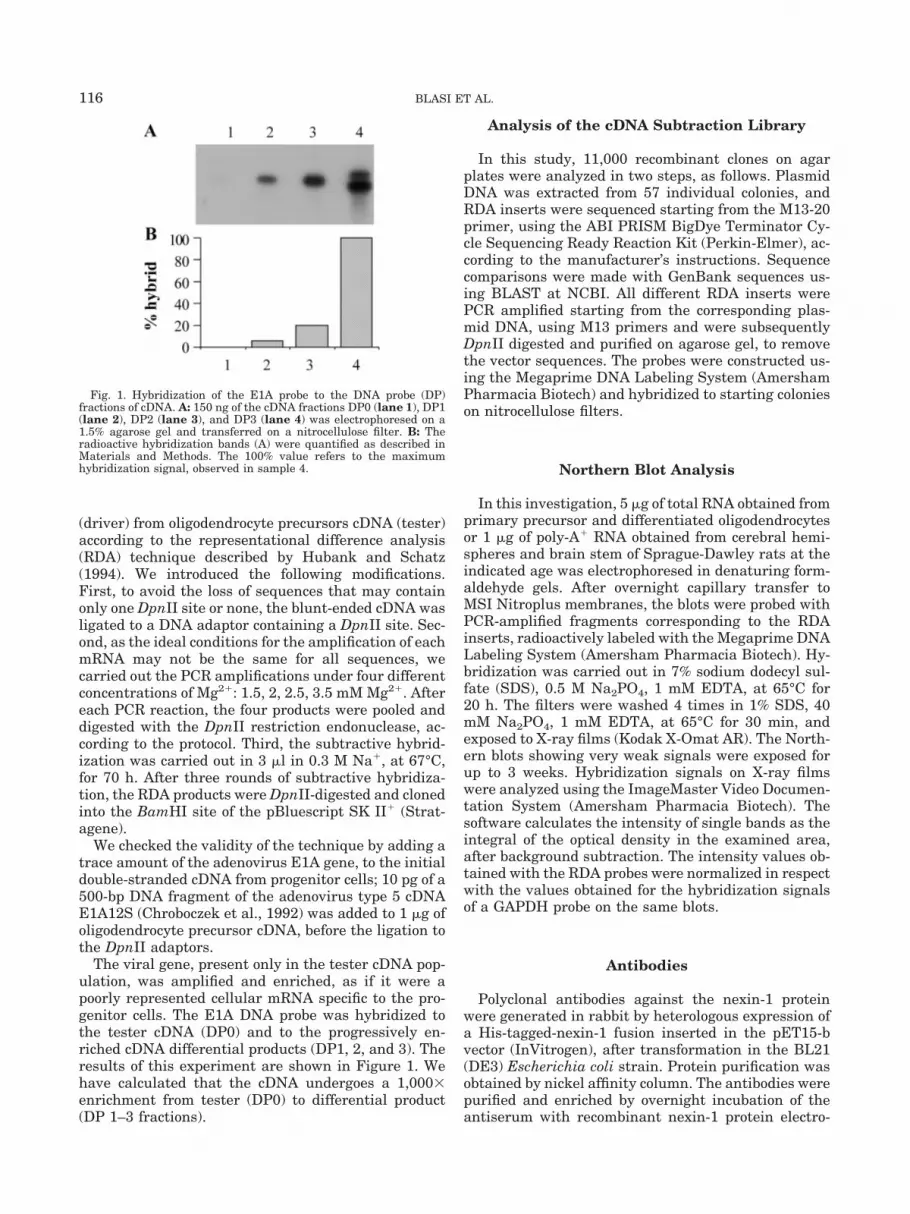
The viral gene, present only in the tester cDNA pop-ulation, was amplified and enriched, as if it were apoorly represented cellular mRNA specific to the pro-genitor cells. The E1A DNA probe was hybridized tothe tester cDNA (DP0) and to the progressively en-riched cDNA differential products (DP1, 2, and 3). Theresults of this experiment are shown in Figure 1. Wehave calculated that the cDNA undergoes a 1,000�enrichment from tester (DP0) to differential product(DP 1–3 fractions).
Analysis of the cDNA Subtraction Library
In this study, 11,000 recombinant clones on agarplates were analyzed in two steps, as follows. PlasmidDNA was extracted from 57 individual colonies, andRDA inserts were sequenced starting from the M13-20primer, using the ABI PRISM BigDye Terminator Cy-cle Sequencing Ready Reaction Kit (Perkin-Elmer), ac-cording to the manufacturer’s instructions. Sequencecomparisons were made with GenBank sequences us-ing BLAST at NCBI. All different RDA inserts werePCR amplified starting from the corresponding plas-mid DNA, using M13 primers and were subsequentlyDpnII digested and purified on agarose gel, to removethe vector sequences. The probes were constructed us-ing the Megaprime DNA Labeling System (AmershamPharmacia Biotech) and hybridized to starting colonieson nitrocellulose filters.
Northern Blot Analysis
In this investigation, 5 �g of total RNA obtained fromprimary precursor and differentiated oligodendrocytesor 1 �g of poly-A� RNA obtained from cerebral hemi-spheres and brain stem of Sprague-Dawley rats at theindicated age was electrophoresed in denaturing form-aldehyde gels. After overnight capillary transfer toMSI Nitroplus membranes, the blots were probed withPCR-amplified fragments corresponding to the RDAinserts, radioactively labeled with the Megaprime DNALabeling System (Amersham Pharmacia Biotech). Hy-bridization was carried out in 7% sodium dodecyl sul-fate (SDS), 0.5 M Na2PO4, 1 mM EDTA, at 65°C for20 h. The filters were washed 4 times in 1% SDS, 40mM Na2PO4, 1 mM EDTA, at 65°C for 30 min, andexposed to X-ray films (Kodak X-Omat AR). The North-ern blots showing very weak signals were exposed forup to 3 weeks. Hybridization signals on X-ray filmswere analyzed using the ImageMaster Video Documen-tation System (Amersham Pharmacia Biotech). Thesoftware calculates the intensity of single bands as theintegral of the optical density in the examined area,after background subtraction. The intensity values ob-tained with the RDA probes were normalized in respectwith the values obtained for the hybridization signalsof a GAPDH probe on the same blots.
Antibodies
Polyclonal antibodies against the nexin-1 proteinwere generated in rabbit by heterologous expression ofa His-tagged-nexin-1 fusion inserted in the pET15-bvector (InVitrogen), after transformation in the BL21(DE3) Escherichia coli strain. Protein purification wasobtained by nickel affinity column. The antibodies werepurified and enriched by overnight incubation of theantiserum with recombinant nexin-1 protein electro-
Fig. 1. Hybridization of the E1A probe to the DNA probe (DP)fractions of cDNA. A: 150 ng of the cDNA fractions DP0 (lane 1), DP1(lane 2), DP2 (lane 3), and DP3 (lane 4) was electrophoresed on a1.5% agarose gel and transferred on a nitrocellulose filter. B: Theradioactive hybridization bands (A) were quantified as described inMaterials and Methods. The 100% value refers to the maximumhybridization signal, observed in sample 4.
116 BLASI ET AL.

blotted on a nitrocellulose membrane. The antibodieswere eluted with 0.2 M glycine-HCl, pH 2.8.
Immunofluorescence Microscopy Analysis
Immunofluorescence studies and the confocal mi-croscopy were carried out using a laser scanner (Radi-ance 2000, Bio-Rad) attached to a Nikon microscope(Eclipse TE300) as previously described (Riccio et al.,2001).
Semiquantitative RT-PCR Analysis of mRNA
In this study, 10 �g of total RNA from primary ratoligodendrocyte precursor or differentiated oligoden-drocytes was incubated with 50 U/50 �l RNase freeDNAase (Promega), for 30 min; 2 �g RNA was reverse-transcribed with 200 U/20 �l reverse transcriptase(RT) (Moloney-murine leukemia virus, Genenco), witholigo-dT as the primer. RT-PCR was carried out in a50-�l final volume, containing the cDNA template (1 �lof the 20-�l mix described), 1.5 U Taq gold DNA poly-merase (Perkin-Elmer), and 0.2 �M-specific oligonucle-otide primers.
The following oligonucleotide primers were con-structed:
RDA279 (melting point [Tm] 55°); left primer, 5�CTC-CGCAATGAAATCAGAG3�, right primer, 5�GGCT-TCACAGGAAATTTGA3�
HIP-1 (Tm 55°); left primer, 5�GAAAACGACCTGCA-GAAGGA3�, right primer, 5�GTGGGGAGTGTCA-CAGAAGG3�
Dynein heavy chain (Tm 50°); left primer, 5�CAGA-CAGTATGTGGCACAAG3�, right primer 5�AGGTA-AAGTCACCACACTGG3�
Nexin-1 (Tm 55°); left primer, 5�GAGACGAGGGGCAT-GATTGAC3�, right primer 5�GCCACAGCTGT-GAACTTGGG3�
�-Actin (Tm 50°); left primer, 5�TGGAATCCTGTG-GCATCC3�, right primer, 5�TCGTACTCCTGCTT-GCTG3�
The sequences were amplified as follows: nexin-1, 20cycles; HIP-1 and �-actin, 25 cycles; and RDA279 anddynein h.c., 27 cycles. The intensities of the amplifiedbands from the agarose gel were quantified as de-scribed above.
RESULTS AND DISCUSSIONDifferentially Expressed Genes
As mentioned earlier, only cultures containing 92–96% A2B5� cells (progenitors) and 92–96% O1�, O4�
cells (oligodendrocytes) were used for the RNA
extraction and construction of the cDNA subtractionlibrary. Using the RDA technique, we have identified anumber of genes that may be specific to rat oligoden-drocyte progenitors. The screening of 11,000 clones hasgenerated cDNA recombinants classified into three cat-egories, two of which represent sequences of knownbiological significance. The recombinants are listed inTable 1 according to their function and frequency dis-tribution in the subtraction library. A category in-cludes sequences coding for proteins of the cytoskele-ton, a second category contains cDNA coding forribosomal proteins, and a third category contains se-quences coding for other proteins, mostly of unknownfunction. Most 3� fragments of the vimentin and L3ribosomal protein cDNA are present in the library atvery high frequencies, totaling almost 70% of the re-combinants. The remaining 30% of the library containsrecombinant cDNA present at lower frequency. Thesecode for the ribosomal proteins S3a, L23a, for the heavychain of dynein, for the RACK1, EF2, and the most 5�fragment of the vimentin and L3 proteins. Most of therecombinant cDNA isolated are present in the librarywith a low frequency, within a range of 0.01–0.2%. Onthe basis of these results, the screening of 11,000 clonesis probably not exhaustive, although from a furtherscreening we should yield almost exclusively rare se-quences. In the following discussion, we show the ex-pression pattern in the CNS, of some of the mRNAcorresponding to the isolated cDNA recombinants.
TABLE 1. Distribution of cloned DP3 fragments in the library
Proteins involved in protein synthesis %L3 rp (1) 33.2S3a rp 3.5L23a rp 3.5Elongation factor 2 3.5L3 rp (2) 1.7PO rp 0.08L19 rp 0.02L15 rp 0.01
Proteins involved in cytoskeletal functionsVimentin (1) 35Dynein heavy chain 8.7Receptor for activated C kinase 1 (RACK1) 5.2Vimentin (2) 1.7Huntingtin interacting protein 1 (HIP1) 0.01GTP binding protein G8 �-subunit 0.01
Other functionsHeat shock protein 90 (hsp90) 3.5Nexin-1 0.2Ferritin-H 0.08Adenine-phosphoribosyl-transferase(APTR)
0.02
Proteasome subunit p45 0.01RDA279 0.02RDA11 0.01RDA95 0.01RDA96 0.01RDA254 0.01RDA288 0.01
L3 cDNA fragments: (1) spanning nt 923–1212 and (2) spanning nt 575–866(accession X62166). Vimentin cDNA fragments: (1) spanning nt 844–1183 and(2) spanning nt 563–802 (accession X62952). The Data Bank accession numberfor RDA11 is bankit 452928; for RDA95, bankit 452938; for RDA96, bankit452942; and for RDA 254, bankit 452936.
117CELL-SPECIFIC mRNA IN CNS DURING BRAIN DEVELOPMENT

Expression Profile of the RDA Clones
cDNA coding for proteins involved in proteinsynthesis
We have isolated six cDNA coding for different ribo-somal proteins. One of these cDNA, L3, is abundant inthe cell and represents 33% of the recombinant cDNAof the library. S3a and L23a represent approximately3% of the library, while P0, L19, and L15 are amongthe rare clones. All these mRNA are downregulatedmore or less drastically during oligodendrocyte differ-entiation (Fig. 2A). Hybridization of L23a, P0, and S3ato the poly-A� RNA from brain hemispheres at progres-sively later stages after birth is similar to that ob-served in the oligodendrocyte mRNA (Fig. 2B–D). TheL23a and P0 probes, hybridized to the brain stemmRNA, are also downregulated (Fig. 2E,F). Becausevery few of the known ribosomal proteins were isolatedby the differential screening, it seemed interesting toanalyze the expression profile of L32, a well-studied,
abundant, and constitutive ribosomal protein that canvary its concentration in cells kept under conditions ofdifferential growth (Curcio et al., 1997; Sienna et al.,2000). The intensity of hybridization of the L32 cDNAprobe to the oligodendrocyte RNA is very similar inboth progenitor and differentiated cells, confirmingthat not all ribosomal protein mRNA are differentiallyexpressed during glial differentiation.
More than 70 proteins are necessary for the assem-bly of ribosomes, and their synthesis is such thatequimolar amounts of each protein are available forpackaging with stoichiometric amounts of rRNA. How-ever, the assembly of the ribosomal subunits is regu-lated only by a few ribosomal proteins, and the evi-dence for extraribosomal functions of the ribosomalproteins is substantial. In this context, the presence ofzinc-finger, bZIP, and helix-loop-helix motifs in thestructure of the ribosomal proteins is suggestive (Wool,1996). The ribosomal proteins have been involved insuch processes as replication, transcription, DNA re-pair, RNA processing, regulation of development, andmalignant transformation, some of which could be crit-ical during cell differentiation (Yacoub et al., 1996;Naora et al., 1998; Koyama et al., 1999). The evidence,in oligodendrocyte progenitor cells, of an upregulationinvolving only a few ribosomal proteins may reflect aregulatory role of these proteins during the process ofoligodendrocyte differentiation.
cDNA coding for proteins involved incytoskeletal functions
mRNA coding for vimentin, dynein heavy chain,Huntingtin interacting protein-1 (HIP1), and theRACK1 receptor of the activated PKC are all involvedin the organization and regulation of cytoskeletal func-tions. During the differentiation of glial and neuronalcells, the transition of each cell type to the successivestage is associated with variations of the intermediatefilaments, implicating these proteins in the process ofdevelopment. Vimentin, a protein belonging to the fam-ily of intermediate filaments, is expressed only in im-mature cells of the neuronal and glial lineage (Bignamiet al., 1982). During the transition from astroblasts toastrocytes, which occurs in rodents at 12–14 days afterbirth, glial fibrillar acidic protein (GFAP) replaces vi-mentin, while vimentin is not expressed in differenti-ated oligodendrocytes (Tardy et al., 1989; Meyer et al.,1989). Hybridization of a cDNA probe from the vimen-tin mRNA sequence to the RNA of progenitor and dif-ferentiated oligodendrocytes confirms that this mRNAis present mainly in progenitor cells and decreasesstrongly in differentiated cells (Fig. 3A). Hybridizationof the same probe to the poly-A� RNA of rat brainhemispheres shows a maximum signal up to 4 daysafter birth and a drop by the 6th day, remaining fairlyconstant thereafter also in the adult animal (Fig. 3B).Hybridization to brain stem mRNA, which containsmainly myelin-producing cells and astrocytes, gener-
Fig. 2. Northern blot hybridization of the cDNA probes coding forproteins involved in protein synthesis. A: Hybridization of total RNAfrom primary progenitor oligodendrocytes (white bars) and differen-tiated oligodendrocytes (black bars). The probes are indicated underthe histogram. The L23a (B,E), P0 (C,F), and S3a (D) probes werehybridized to rat cerebral hemispheres (B,C,D) and brain stem (E,F).The hybridization signals were quantified as described in Materialsand Methods. The vertical axis indicates the percentage hybridiza-tion. The 100% value represents the maximum hybridization observedin each experiment. Numbers on the horizontal axis indicate the ageof the animals in days after birth.
118 BLASI ET AL.

ates a signal whose intensity decreases noticeablymore than 8 days after birth (Fig. 3C). The differencebetween the drastic decrease of the hybridization sig-nal in primary oligodendrocytes and its milder declinein the cerebral hemispheres may be due to the com-plexity of the brain cell population, which contains avariety of cell types, i.e., glial, neuronal, ependymal,and stem cells. During the cell cycle, vimentin under-goes a series of phosphorylation/dephosphorylationevents with the consequent modification of the organi-zation of its filaments (Inada et al., 1999; Janosch etal., 2000). Its presence in growing cells is not thereforesurprising. However, the precise function of vimentinremains unknown, and a mouse knockout for this pro-tein is fertile, with a phenotype characterized by ne-crosis of some of the Purkinje cells and decrease of theBergmann glial cells in the cerebellum (Colucci-Guyonet al., 1999).
Northern blot analysis of oligodendrocyte progenitorand differentiated mRNA using the dynein heavychain/MAP1C probe (Mikami et al., 1993) producedalmost undetectable signals, even after very long expo-sure of the hybridization filter. We therefore decided touse a semiquantitative RT-PCR technique; the resultsare shown in Figure 4. �-Actin represents the controlsample for a stable mRNA expressed constitutively,while nexin-1 is the control sample for downregulatedmRNA. The results indicate a decrease of the dynein
Fig. 3. Northern blot hybridization of the cDNA probes coding forthe remaining proteins. The total RNA from primary progenitor (p)and differentiated (d) oligodendrocytes (A,H,M,P,S), and the poly-A�
RNA from cerebral hemispheres (B,D,F,I,N,Q,T,V) and from brainstem (C,E,G,L,O,R,U,Z) of rats was hybridized to the following RDAprobes: (A–C) vimentin, (D,E) dynein heavy chain, (F,G) HIP1, (H–L)RACK1, (M–O) nexin-1, (P–U) RDA288, (V,Z) RDA279. The probeRDA288 hybridized to two messenger bands migrating in the gel atpositions corresponding to 1,200 and 1,600 bp, respectively. P–R:Quantitation of the 1,600-bp hybridization band. S–O: Quantificationof the 1,200-base band. The RDA288 probe was hybridized to thepoly-A� RNA of oligodendrocytes because the signal was not de-tactable on total RNA. No hybridization signal of the RDA279 probewas detectable to either the poly-A� or the total oligodendrocyte RNA.The values on the horizontal and vertical axes are the same as in Fig.2. The data bank accession number for RDA 288 is AF388527; for RDA279 is AF388528; for rat HIP1 is AF388529.
Fig. 4. Semiquantitative polymerase chain reaction (PCR) analysisof oligodendrocyte RNA from primary progenitor and differentiatedcells. White bars, quantification of the mRNA in oligodendrocyteprogenitor cells; black bars, quantification of mRNA in differentiatedoligodendrocyte cells. The 100% value represents the maximum am-plification observed in each experiment. The relative percentage of thetwo samples (from progenitor and differentiated cells) is indicated onthe vertical bar.
119CELL-SPECIFIC mRNA IN CNS DURING BRAIN DEVELOPMENT

heavy chain mRNA in differentiated primary oligoden-drocytes. The mRNA is upregulated during early de-velopment in the brain hemispheres and in the brainstem (Fig. 3D,E). The intensity of hybridization tomRNA from the cerebral hemispheres starts falling in4-day-old rats, while hybridization to RNA from brainstem is already weakened in 2-day-old rats.
The dynein heavy chain is part of a molecular com-plex responsible for movements associated with micro-tubules. The complex is made of two heavy chainsbound to intermediate and light chains. The ATPaseactivity responsible for the motor function is part of theheavy chain (Vallee, 1993; Barton and Goldstein,1996). Dynein is responsible for the transport of endo-somes, lysosomes, and elements of the Golgi apparatusand is essential for the mitotic spindle formation andpositioning. Through its interaction with dynactin, dy-nein assembles the spindle and organizes the spindlepole (Gaglio et al., 1996; Merdes et al., 2000). Upregu-lation of this mRNA in oligodendrocyte progenitors isin good agreement with the essential role of dynein inproliferating cells.
Another protein upregulated in oligodendrocyte pro-genitors is the Huntingtin interacting protein-1(HIP1), first isolated for its interaction with the proteinproduct of the Huntington disease gene (Kalchman etal., 1997). This mRNA is rare and poorly represented inthe cDNA library. The RT-PCR analysis shown in Fig-ure 4 indicates downregulation of the HIP1 RNA indifferentiated oligodendrocytes. The quantification ofthe hybridization signals of the HIP1 probe to the RNAof cerebral hemispheres and brain stem, isolated from1- to 90-day-old rats is shown in Figure 3F,G. Theintensity of the hybridization to the poly-A� RNA frombrain hemisphere is already decreasing in 4-day-oldrats, reaching a minimum in the adult animals. Theintensity of the signal in the brain stem RNA decreasesmore gradually.
The function of HIP1 is unknown, but the proteincontains evolutionarily conserved sequences. By anal-ogy with its yeast homologue, the SLA2 protein, in-volved in assembly of cortical cytoskeleton, endocyto-sis, and vesicle trafficking, HIP1 may have acytoskeletal function (Holtzman et al., 1993). HIP1 be-longs to a family of proteins that includes SLA2, talin,a cytoskeletal protein involved in cell–cell interaction,and HIP-1R, the human and mouse isoform of HIP1with ubiquitous tissue distribution. The SLA2 family ofproteins is characterized by the presence of a conservedN-terminal domain, a talin-like domain at the C-termi-nus, and three coiled-coil forming domains (Engqvist-Goldstein et al., 1999). Since both HIP-1R and theHuntingtin protein are found to colocalize and associ-ate with clathrin, and SLA2p has been implicated inendocytosis and vesicle trafficking, it has been pro-posed that this family of proteins is involved in func-tions linking the actin cytoskeleton to the sites of en-docytosis (Engqvist-Goldstein et al., 1999; Waelter etal., 2001).
Figure 3H shows the results of the hybridization ofthe RACK1 cDNA to oligodendrocyte RNA. RACK1mRNA is more abundant in oligodendrocyte progeni-tors than in differentiated cells. Hybridization to thepoly-A� RNA from rat cerebral hemispheres and brainstem decreases steadily from the newborn to the adultanimals (Fig. 3I,L). RACK 1 is a receptor protein thatbinds the active conformation of some of the isoforms ofprotein kinase C (PKC) and mediates the interactionbetween the activated kinase and the cell membranes(Smith and Mochly-Rosen, 1992; Newton, 1997). Thehighest affinity is for the �-isoform of PKC (Ron et al.,1995). In addition, RACK 1 interacts with the cytoplas-mic domain of the integrin �-subunit, indicating aninvolvement in the interaction between plasma mem-brane and cytoskeleton (Liliental and Chang, 1998).The decreased PKC� activity with aging correlateswith the decrease of membrane-bound RACK1 (Pascaleet al., 1996; Battaini et al., 1997). Baron et al. (1998)showed that the activation of PKC in oligodendroblastcultures (O-2A cells) inhibits the transition to imma-ture oligodendrocytes and the expression of the genescoding for the myelin proteins. These investigators sug-gest that PKC activation inhibits the changes of thecytoskeletal organization necessary to the extension ofoligodendrocyte processes during differentiation. A dif-ferential expression of the RACK1 mRNA in progenitoroligodendrocytes is in agreement with a role of RACK1in the early differentiation of oligodendrocytes.
cDNA coding for proteins with different orunknown functions
Nexin-1 is one of the mRNA drastically downregu-lated in differentiated oligodendrocytes. Figure 3Mshows the hybridization of the cDNA to the RNA ofprogenitor and differentiated oligodendrocytes. In con-trast, the hybridization of the same probe to the mRNAfrom rat brain hemispheres: as shown in Figure 3N,there is no difference between the levels of hybridiza-tion of the same probe to mRNA from brain hemi-spheres of rats at any of the stages examined.
The result is the same when nexin-1 cDNA is hybrid-ized to brain stem mRNA (Fig. 3O). Our results clearlyshow that the downregulation of nexin-1 mRNA duringdifferentiation, is a unique feature of the oligodendro-cyte lineage. The brain hemispheres and the brainstem, which, in addition to oligodendrocytes, containneurons and astrocytes, maintain constant levels ofnexin-1 mRNA.
Figure 5 displays immunofluorescence microscopy ofCG4 (Fig. 5A,C,E) and primary (Fig. 5B,D,F) tissueculture cells stained with polyclonal antibodies againstthe nexin-1 protein. As expected, the progenitor cells(Fig. 5A,B) and the type 2 astrocytes (Fig. 5E,F) displaya strong fluorescent signal, while the oligodendrocytes(Fig. 5C,D) display very weak fluorescence. We con-clude that the differentiated oligodendrocytes show a
120 BLASI ET AL.

decrease of the nexin-1 protein which is consistent withthe fall of the corresponding mRNA.
Nexin-1, an inhibitor of serine proteases, belongs tothe serpin superfamily of proteins and is specific forthrombin and thrombin-like proteases (Guenther et al.,1985; Stone et al., 1987; Sommer et al., 1987). It isexpressed in neurons and astrocytes (Simpson et al,1994; Reinhard et al., 1994; Sommer et al., 1987), inwhich it promotes neurite outgrowth, the formation ofsynapses, cell migration, and astrocyte differentiation
and stellation (Canavaugh et al., 1990; Beecher et al.,1994). Kury et al. (1997) showed that the nexin-1 geneis activated after administration of bFGF to the cells.bFGF blocks the differentiation of preoligodendrocytes(Gard and Pfeiffer, 1993; Mayer et al., 1993). Further-more, Erno et al. (1996) showed that the nexin-1promoter contains a binding site for Krox-24, a tran-scription factor that is downregulated during oligoden-drocyte differentiation (Sock et al., 1997). Both obser-vations are in good agreement with our finding of areduction of the levels of nexin-1 mRNA and suggestthat the switching off of the nexin-1 gene is importantfor the correct differentiation of oligodendrocyte pro-genitors.
We have also examined some of the cDNA of un-known function. Those analyzed so far show weak hy-bridization signals to both the mRNA from oligoden-drocytes and from the two regions of the brain. Thisfinding is in good agreement with the rare nature ofthese mRNA both in the cDNA library and in themRNA population. Figure 3P–Z shows the results ofthese experiments. When RDA288 probe is used, thehybridization product yields two bands, a larger band,corresponding to �1,600 bases and a smaller band,corresponding to �1,200 bases (not shown).
The hybridization patterns for the two componentsare different and are shown in Figure 3:P–R show datarelating to the larger component, S–U to the smaller.
The process of differentiation appears to involve thedownregulation of RDA288 mRNA, irrespective of thematerial of origin: brain hemispheres, stem, or oligo-dendrocytes. However, the larger component ofRDA288 mRNA shows a decrease in the brain stem bythe time the rat is 2–4 days old (Fig. 3R), while in thebrain hemispheres, it shows a more gradual decrease,with maximal intensity in 4-day-old rats (Fig. 3Q). Thesmaller band exhibits a bell-shaped pattern in thebrain hemispheres (Fig. 3T), while in the brain stemhistogram, it decreases gradually from postnatal day 1to the adult animal (Fig. 3U). Both mRNAs are clearlydownregulated in differentiated oligodendrocytes, thesmaller band to a somewhat lower level than the largerband (Fig. 3P,S).
Figure 3V and Z shows the results of the hybridiza-tion of the RDA clone 279 to the cerebral hemispheresand the brain stem poly-A� RNA. This mRNA de-creases steadily from day 1 in both brain regions. Thehybridization of this probe to the mRNA of oligoden-drocytes was hardly detectable. We have thereforeused a semiquantitative PCR technique for the detec-tion of this mRNA too (Fig. 4). In contrast with theresults obtained with the brain tissues, little if anydifference can be observed between the amplified pro-genitor and differentiated oligodendrocyte RNA.
It is interesting to note that the abundance of themRNA coding for the dynein heavy chain, HIP1, andthe RDA279 protein is higher in the brain stem andhemispheres than in oligodendrocytes. In fact, in theNorthern blots of brain RNA, the hybridization signals,although weak, are detectable. This may be due to the
Fig. 5. Immunostaining of CG4 and primary rat cells of the oligo-dendrocyte lineage. Paraformaldehyde-fixed CG4 (A, progenitor cells;C, differentiated oligodendrocytes, E, A2 astrocytes) and primary ratcells (B, progenitor; D, differentiated oligodendrocytes; F, astrocytes)were incubated with polyclonal antibodies against rat nexin-1. G:Results of incubation of oligodendrocyte progenitor cells with antibod-ies preabsorbed with the nexin-1 protein. H: Phase-contrast image ofthe cells in G.
121CELL-SPECIFIC mRNA IN CNS DURING BRAIN DEVELOPMENT

presence of cells, other than oligodendrocytes, that ex-press these mRNA in the brain stem and hemispheres.
CONCLUSIONS
The RDA technique has permitted isolation of anumber of cDNA clones that are differentially regu-lated in primary oligodendrocytes during differentia-tion and, in general, are downregulated, in the brainstem and in the brain hemispheres, during the devel-opment of the CNS. The change in the rate of messen-ger expression observed in the brain tissues representsan average of the specific expression in the differentcell types. In some cases, the downregulation is muchmore dramatic in the RNA from the oligodendrocytesthan in that from the CNS tissues and, in other cases,is the opposite. These differences are very clear for thenexin-1 mRNA, which remains constant throughoutCNS development in the brain stem and hemispheresand is strongly downregulated in oligodendrocytes. Thevariance between the RNA expression in differentiat-ing oligodendrocytes and that in brain tissues is a goodindicator of the purity of the primary oligodendrocytecells.
We are currently studying the role of the isolatedcDNA in relation to the differentiation program of theglial lineage in an effort to identify those that may havea regulatory function during development.
ACKNOWLEDGMENTS
The authors are very grateful to Dr. Cesira Galeotti,Dr. Maria Luisa Sartirana, and Dr. Davide Ambrosettifor helpful discussions and critical reading of themanuscript.
REFERENCES
Ausubel FM, Brent K, Kingston RE, Moore DD, Seidman JG, SmithJA, Struhl K. 1989. Current protocols in molecular biology. NewYork: John Wiley & Sons.
Baron W, De Jonge JC, De Vries H, Hoekstra D. 1998. Regulation ofoligodendrocyte differentiation: protein kinase C activation pre-vents differentiation of O-2A progenitor cells toward oligodendro-cytes. Glia 22:121–129.
Barton NR, Goldstein LS. 1996. Going mobile: microtubule motorsand chromosome segregation. Proc Natl Acad Sci U S A 93:1735–1742.
Battaini F, Pascale A, Paoletti R, Govoni S. 1997. The role of anchor-ing protein RACK1 in PKC activation in the ageing rat brain. TINS20:410–415.
Beecher TT, Anderson TT, Fenton JWI, Festoff BW. 1994. Thrombinreceptor peptides induce shape changes in neonatal murine astro-cytes in culture. J Neurosci Res 37:108–115.
Bignami A, Raju T, Dahl D. 1982. Localization of vimentin, the non-specific intermediate filament protein, in embryonal glia and inearly differentiating neurons. Dev Biol 91:286–295.
Brogi A, Strazza M, Melli M, Costantino-Ceccarini E. 1997. Inductionof intracellular ceramide by interleukin-1� in oligodendrocytes.J Cell Biochem 66:532–541.
Canavaugh K, Gurwitz D, Cunningham DD, Bradshaw R. 1990. Re-ciprocal modulation of astrocyte stellation by thrombin and pro-tease nexin-1. J Neurochem 54:1735–1743.
Chomczynski P, Sacchi N. 1987. Single-step method of RNA isolationby acid guanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem 162:156–159.
Chroboczek J, Bieber F, Jacrot B. 1992. The sequence of the genomeof adenovirus type 5 and its comparison with the genome of adeno-virus type 2. Virology 186:280–285.
Colucci-Guyon E, Gimenez Y, Ribotta M, Maurice T, Babinet C, PrivatA. 1999. Cerebellar defect and impaired motor coordination in micelacking vimentin. Glia 25:33–43.
Curcio D, Glibetic M, Larson DE, Sells BH. 1997. GA-binding proteinis involved in altered expression of ribosomal protein L32 gene.J Cell Biochem 65:287–307.
Engqvist-Goldstein AE, Kessels MM, Chopra VS, Hayden MR, DrubinDG. 1999. An actin-binding protein of the Sla2/Huntingtin inter-acting protein 1 family is a novel component of clathrin-coated pitsand vesicles. J Cell Biol 147:1503–1518.
Erno H, Kury P, Botteri FM, Monard D. 1996. A Krox binding siteregulates protease nexin-1 promoter activity in embryonic heart,cartilage and parts of the nervous system. Mech Dev 60:139–150.
Gaglio T, Saredi A, Bingham JB, Hasbani MJ, Gill SR, Schroer TA,Compton DA. 1996. Opposing motor activities are required for theorganization of the mammalian mitotic spindle pole. J Cell Biol135:399–414.
Gard AL, Pfeiffer SE. 1989. Oligodendrocyte progenitor cells isolateddirectly from telencephalon at a specific phenotypic stage: myelino-genic potential in a defined environment. Development 106:119–132.
Gard AL, Pfeiffer SE. 1993. Glial cell mitogens bFGF and PDGFdifferentially regulate development of O4� GalC- oligodendrocyteprogenitors. Dev Biol 159:618–630.
Guenther J, Nick H, Monard D. 1985. A glia-derived neurite-promot-ing factor with protease inhibitory activity. EMBO J 4:1963–1966.
Holtzman DA, Yang S, Drubin DG. 1993. Synthetic-lethal interac-tions identify two novel genes, SLA1 and SLA2, that control mem-brane cytoskeleton assembly in Saccharomyces cerevisiae. J CellBiol 122:635–644.
Hubank M, Schatz DG. 1994. Identifying differences in mRNA ex-pression by representational difference analysis of cDNA. NucleicAcids Res 22:5640–5648.
Inada H, Togashi H, Nakamura Y, Kaibuchi K, Nagata K, Inagaki M.1999. Balance between activities of Rho kinase and type 1 proteinphosphatase modulates turnover of phosphorylation and dynamicsof desmin/vimentin filaments. J Biol Chem 274:34932–34939.
Janosch P, Kieser A, Eulitz M, Lovric J, Sauer G, Reichert M, GounariF, Buscher D, Baccarini M, Mischak H, Kolch W. 2000. The Raf-1kinase associates with vimentin kinases and regulates the struc-ture of vimentin filaments. FASEB J 14:2008–2021.
Kalchman M, Koide BH, McCutcheon K, Graham RK, Nichol K,Nishyama K, Kazemi-Esfarjani P, Lynn FC, Wellington C, MetzlerM, Goldberg PY, Kanazawa I, Gietz RD, Hayden MR. 1997. HIP1, ahuman homologue of S. cerevisiae Sla2p, interacts with membrane-associates huntingtin in the brain. Nature Genet 16:44–53.
Kondo T, Raff M. 2000. Oligodendrocyte precursor cells repro-grammed to become multipotential CNS stem cells. Science 289:1754–1757.
Koyama Y, Katagiri S, Hanai S, Uchida K, Miwa M. 1999. Poly(ADP-ribose) polymerase interacts with novel Drosophila ribosomalproteins, L22 and L23a, with unique histone-like amino-terminalextensions. Gene 226:339–345.
Kury P, Schaeren-Wiemers N, Monard D. 1997. Protease nexin-1 isexpressed at the mouse met-/mesencephalic junction and FGF sig-naling regulates its promoter activity in primary met-/mesence-phalic cells. Development 124:1251–1262.
Liliental J, Chang DD. 1998. RACK1, a receptor for activated proteinkinase C, interacts with integrin � subunit. J Biol Chem 273:2379–2383.
Mayer M, Bogler O, Noble M. 1993. The inhibition of oligodendrocytedifferentiation of O-2A progenitors caused by bFGF is overridden byastrocytes. Glia 8:12–19.
Merdes A, Heald R, Samejima K, Earnshaw WC, Cleveland DW. 2000.Formation of spindle poles by dynein/dynactin-dependent transportof NuMA. J Cell Biol 149:851–862.
Meyer SA, Ingraham CA, McCarthy KD. 1989. Expression of vimentinby cultured astroglia and oligodendroglia. J Neurosci Res. 259:251–259.
Mikami A, Paschal BM, Mazumdar M, Vallee RB. 1993. Molecularcloning of the retrograde transport motor cytoplasmic dynein (MAP1C). Neuron 10:787–796.
Naora H, Takai I, Adachi M, Naora H. 1998. Altered cellular re-sponses by varying expression of a ribosomal protein gene: sequen-tial coordination of enhancement and suppression of ribosomal pro-
122 BLASI ET AL.

tein S3a gene expression induces apoptosis. J Cell Biol 141:741–753.
Newton AC. 1997. Regulation of protein kinase C. Curr Opin Cell Biol9:161–167.
Orentas DM, Miller RH. 1998. Regulation of oligodendrocyte develop-ment. Mol Neurobiol 18:247–259.
Pascale A, Fortino I, Govoni S, Trabucchi M, Wetsel WC, Battaini F.1996. Functional impairment in protein kinase C by RACK1 (recep-tor for activated C kinase 1) deficiency in aged rat brain cortex.J Neurochem 67:2471–2477.
Raff MC, Miller RH, Noble M. 1983. A glial progenitor cell thatdevelops in vitro into an astrocyte or an oligodendrocyte dependingon culture medium. Nature 303:390–396.
Raible DW, McMorris FA. 1993. Oligodendrocyte differentiation andprogenitor cell proliferation are independently regulated by cyclicAMP. J Neurosci Res 34:287–294.
Reinhard E, Suidan HS, Pavlik A, Monard D. 1994. Glia-derivednexin/protease nexin-1 is expressed by a subset of neurons in therat brain. J Neurosci Res 37:256–270.
Riccio M, Di Giaimo R, Pianetti S, Palmieri PP, Melli M, Santi S.2001. Nuclear localization of cystatin B, the cathepsin inhibitorimplicated in myoclonus epilepsy (EPM1). Exp Cell Res 262:89–94.
Ron D, Luo J, Mochly-Rosen D. 1995. C2 region-derived peptidesinhibit translocation and function of � protein kinase C in vivo.J Biol Chem 270:24180–24187.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: a lab-oratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Lab-oratory Press.
Sienna N, Larson DE, Sells BH. 2000. Dexamethasone stimulatesribosomal protein L32 gene transcription in rat myoblasts. Mol CellEndocrinol 167:127–37.
Simpson CS, Johnston HM, Morris BJ. 1994. Neuronal expression ofprotease nexin-1 mRNA in rat brain. Neurosci Lett 170:286–290.
Smith BL, Mochly-Rosen D. 1992. Inhibition of protein kinase Cfunction by injection of intracellular receptor for the enzyme. Bio-chem Biophys Res Commun 188:1235–1240.
Sock E, Leger H, Kuhlbrodt K, Schreiber J, Enderich J, Richter-Landsberg C, Wegner M. 1997. Expression of Krox proteins duringdifferentiation of the O-2A progenitor cell line CG4. J Neurochem68:1911–1919.
Sommer J, Gloor SM, Rovelli GF, Hofsteenge J, Nick H, Meier R,Monard D. 1987. cDNA sequence encoding for a rat glia-derivednexin and its homology to members of the serpin superfamily.Biochemistry 26:6407–6410.
Stone SR, Nick H, Hofsteenge J, Monard D. 1987. Glial-derived-neurite-promoting factor is a slow-binding inhibitor of trypsin,thrombin, and urokinase. Arch Biochem Biophys 252:237–244.
Tardy M, Fages C, Riol H, Le Prince G, Rataboul P, Charriere-Bertrand C, Nunez J. 1989. Developmental expression of the glialfibrillary acidic protein mRNA in the central nervous system and incultured astrocyte. J Neurochem 52:162–167.
Vallee R. 1993. Molecular analysis of the microtubule motor dynein.Proc Natl Acad Sci U S A 90:8769–8772.
Waelter S, Scherzinger E, Hasenbank R, Nordhoff E, Lurz R, GoehlerH, Gauss C, Sathasivam K, Bates GP, Lehrach H, Wanker EE.2001. The Huntingtin interacting protein HIP1 is a clathrin and�-adaptin-binding protein involved in receptor-mediated endocyto-sis. Hum Mol Genet 10:1807–1817.
Wool IG. 1996. Extraribosomal functions of ribosomal proteins.Trends Biochem Sci 21:164–165.
Yacoub A, Kelley MR, Deutsch WA. 1996. Drosophila ribosomal pro-tein P0 contains apurimic/apyrimidinic endonuclease activity. Nu-cleic Acids Res 24:4298–4303.
123CELL-SPECIFIC mRNA IN CNS DURING BRAIN DEVELOPMENT