src homology 3 domain of hepatitis c virus ns5a protein interacts with bin1 and is important for...
TRANSCRIPT

SII
SL
B(apHbiitittttfitmmecusNbtcttamo
Tsopptp
GASTROENTEROLOGY 2006;130:794–809
rc Homology 3 Domain of Hepatitis C Virus NS5A Proteinnteracts With Bin1 and Is Important for Apoptosis andnfectivity
ANTOSH K. NANDA, DAVID HERION, and T. JAKE LIANG
iver Diseases Branch, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, Marylandatepatitsottcs
Nmi3wvcpplcpcSHb
nhldce
See editorial on page 995.
ackground & Aims: HCV nonstructural protein 5ANS5A) has been implicated in regulating cell growthnd interferon response. The NS5A protein containsroline-rich regions that are highly conserved amongCV genotypes and match Src homology 3 (SH3)-inding motifs (PxxP) found in various cellular signal-
ng molecules. Methods: We screened for HCV NS5Anteracting proteins by using the yeast 2-hybrid sys-em and studied the functional consequence of thisnteraction. Results: Several independent clones con-aining SH3 domains were isolated along with Bin1, aumor suppressor with pro-apoptotic properties, beinghe most frequently identified clone. The protein–pro-ein interaction between NS5A and Bin1 was con-rmed by in vitro binding, in vivo co-immunoprecipi-ation, and confocal microscopy. Deletion andutation analyses indicated that the SH3 bindingotif of HCV NS5A and SH3 domain of Bin1 are
ssential for interaction. Human hepatoma (HepG2)ells lacking expression of Bin1 undergo apoptosispon infection with adeno-Bin1. Bin1-induced apopto-is was inhibited in HepG2 cells expressing wild-typeS5A but not NS5A mutant with mutations in the SH3inding motif. Infectious HCV genome containing mu-ations in the SH3 binding motif was not infectious inhimpanzees. Conclusions: Our results indicate thathis interaction is implicated in productive HCV infec-ion and may contribute to the pathogenesis of hep-tocellular carcinoma. In addition, the NS5A PxxPotif may represent a novel target for antiviral devel-
pment.
he HCV genome is a single-stranded positive-senseRNA molecule of �9.5 kilobases in length with a
ingle, large, open-reading frame encoding a polyproteinf �3000 amino acids. Processing of the precursorolyprotein requires both host and viral proteases toroduce the structural (core, E1, and E2) and nonstruc-ural (NS2, NS3, NS4A, NS4B, NS5A, and NS5B)
roteins. The mature NS5A protein is generated by thection of the NS3/4A serine protease. NS5A is localizedo the cytoplasmic and perinuclear regions of the cell andxists in a basal or hyperphosphorylated state (p56 or58) that regulates the HCV life cycle.1 NS5A protein isphosphoprotein with pleiotropic functions.2 It has at-
racted considerable interest because of its potential rolen modulating the interferon response.3 Adaptive muta-ions have been found in the NS5A in the context ofelectable subgenomic HCV replicons, suggesting a rolef NS5A in modulating viral expression and replica-ion.4,5 The exact function of NS5A is unknown, al-hough it is speculated to form a multiprotein replicationomplex with other nonstructural proteins on the cyto-olic side of the endoplasmic reticulum membrane.6
An increasing body of evidence has demonstrated thatS5A interacts with a number of cellular proteins anday also interfere with host cell signaling pathways
ncluding Grb2 and p85 subunits of phosphatidylinositol-kinase (PI3K).7 NS5A was shown to interact directlyith the interferon-induced double-stranded RNA-acti-ated protein kinase PKR, and this interaction seems toorrelate with inhibition of PKR function.8 Recent re-orts suggest that NS5A may also regulate cell-cyclerogression by modulating a number of cell-cycle regu-atory genes and can transform murine fibroblasts.9 Be-ause NS5A does not bind to the DNA directly, itrobably exerts its effect through interaction with otherellular protein(s) like cellular transcription factorRCAP, the human karyopherin 3 protein, or p53.2
owever, the physiologic significance of these NS5A-inding proteins in the HCV life cycle remains unclear.
Abbreviations used in this paper: AD, activation domain; 3AT, 3-ami-otriazole; DBD, DNA binding domain; 5FOA, 5-fluoroorotic acid; HCC,epatocellular carcinoma; MBD, myc-binding domain; NLS, nuclear
ocalization signal; NS5A, nonstructural protein 5A; PI3K, phosphati-ylinositol 3-kinase; SH3, Src homology 3; TUNEL, terminal deoxynu-leotidyl transferase-mediated deoxyribonucleotide triphosphate nick-nd labeling.© 2006 by the American Gastroenterological Association Institute
0016-5085/06/$32.00
doi:10.1053/j.gastro.2005.12.030
l(HbtpdmpcbsroSNpbtaaite
ptfctt1ciAoGTtN1eTNttN
wfWpsvu
hl2sbHMTT(bc2tsNtmlwtTc
ay1tmltil(mctmtpbAT
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 795
A potential clue that NS5A may perturb certain cel-ular signaling functions is the presence of a proline-richPxxP) sequence,10 which is conserved among variousCV genotypes 1 and matches Src homology 3 (SH3)-
inding motif. These motifs form extended helical struc-ures and are found in a number of viral and cellularroteins involved in signal transduction.11 The SH3omain found in a diverse group of signal-transducingolecules serves as the binding surface to proline-rich
eptides of polyproline type II helix with the minimalonsensus PxxP. Although isolated SH3 domains canind constitutively to their cognate ligands in vitro,ome SH3-binding reactions have been suggested to beegulatable in vivo.12,13 It was previously shown that onef these PxxP motifs in NS5A interacts directly with theH3 domains of Grb2.10 To elucidate the role of HCVS5A in viral pathogenesis, we searched for cellular
roteins interacting with the NS5A by the yeast 2-hy-rid screening of 2 human cDNA libraries. We iden-ified Bin1, a member of the BAR family of genes,14 asmajor cellular interactor of NS5A. In this report, we
lso demonstrated the functional importance of thisnteraction, which may be important for the produc-ive infection of HCV and contribute to the oncogenicffect of chronic HCV infection.
Materials and Methods
NS5A and Bin1 Constructs
The full-length NS5A gene (aa 1974-2419) was am-lified and cloned into pCRII (Invitrogen, Carlsbad, CA) fromhe HCV CG1b clone.15 All constructs were further derivedrom pCRIINS5A plasmid. The pCRIINS5A*P3 mutant (nu-leotide C7314G, C7317G, and C7320G) was generatedhrough site-directed mutagenesis (Stratagene, La Jolla, CA) ofhe pCRIINS5A. The pPC97 NS5A constructs listed in FigureA were generated by PCR cloning with the sense primersontaining a Bgl II site and the antisense primers containing ann-frame stop codon and an Spe I site. A Flag epitope with anTG start codon was introduced by cloning a hybridizedligonucleotide sequence (forward 5=-AGCTTCCACCATG-ACTACAAGGACGACGATGACAA-3=, reverse 5=-GATC-TGTCATCGTCGTCCTTGTAGTCCATGGTGGA-3=) be-
ween Hind III and Bgl II sites of pcDNA3. The wild-typeS5A was subcloned from pPC97NS5A (plasmid 1, Figure
A) as a Bgl II–Not I fragment into pcDNA3 vector with Flagpitope at Bam HI–Not I sites to generate pcDNA3FlagNS5A.he pcDNA3FlagNS5A*P3 was generated by PCR cloning ofS5A*P3 from pCRIINS5A*P3 with the sense primer con-
aining a Bgl II site in frame with the Flag epitope and withhe antisense primer containing an in-frame stop codon and a
ot I site. RThe Bin1 and its deletion constructs listed in Figure 1B,ere generated through ligation of PCR products amplified
rom the Bin1 constructs obtained from Dai Sakamuro (Theistar Institute, Philadelphia, PA), with sequence-specific
rimers containing a Sal I site in the sense primer and a Spe Iite in the antisense primers as described previously.16 Thearious Bin1 constructs were cloned into the pPC86 backbonesing the Sal I and Spe I sites.
Gal4-Based Yeast Two-Hybrid System
MaV103 (MATa, ura3-52, leu2-3, 112 trp1-901,is3�200, ade2-101, gal4�gal80�GAL1:LacZ, GAL1:HIS3,ys2 SPAL10; URA3)17 was used as the host strain for the-hybrid system based on the Gal4 protein. The MaV103train contains 3 reporter genes whose expression is regulatedy different Gal4-responsive promoters. In this study, we usedIS3 and URA3 reporter genes integrated in the genome ofaV103 to detect the HCV NS5A-host protein interaction.he pPC97 vector was used as the bait plasmid in this system.his vector contains the DNA binding domain (DBD) of Gal4
aa 1–147) and the LEU2 selectable marker. The HCV NS5Aait plasmid (pPC97NS5A, plasmid 6) was constructed byloning the C-terminally truncated HCV NS5A (aa 1974–396) gene in frame with the Gal4 DNA-binding domain inhe pPC97 vector,17 because the full-length HCV NS5A was aelf-activator in this 2-hybrid system. The Gal4 DBD-HCVS5A fusion protein (plasmid 6, Figure 1A) is expressed under
he control of constitutive Saccharomyces cerevisiae ADC1 pro-oter. For the library screen, an activated human T-cell cDNA
ibrary (Marc Vidal, MGH Cancer Center, Charlestown, MA)as cloned into the prey plasmid (pPC86),17 which contains
he activation domain (AD) of Gal4 (aa 768–881) and theRP1-selectable marker. Gal4 AD fusion host proteins areonstitutively expressed under the control of ADC1 promoter.
Growth and manipulation of the yeast strain was doneccording to standard procedures.18 For the library screen,east MaV103 carrying the pPC97NS5A (plasmid 6, FigureA) was transformed with the pPC86-cDNA library, andransformants were replica plated onto synthetic completeedium containing 3-aminotriazole (3AT, 20 mmol), lacking
eucine, tryptophan, and histidine. Interaction phenotypes ofhe MaV103 transformants were determined by the differencesn growth on dropout plates containing 20 mmol of 3ATacking histidine and on plates containing uracil and 0.1%wt/vol) 5-fluoroorotic acid (5FOA). The pPC86-cDNA plas-ids were recovered and reintroduced into yeast MaV103
ontaining pPC97NS5A (plasmid 6, Figure 1A) for confirma-ion by plating the transformants onto synthetic completeedium containing 20 mmol of 3AT but lacking leucine,
ryptophan, and histidine; medium lacking leucine, trypto-han, and uracil; and medium lacking leucine and tryptophanut containing X-gal, a substrate for the lacZ-encoded enzyme.
similar strategy was followed to screen a PROQUESTwo-Hybrid human liver cDNA library (Life Technologies,
ockville, MD).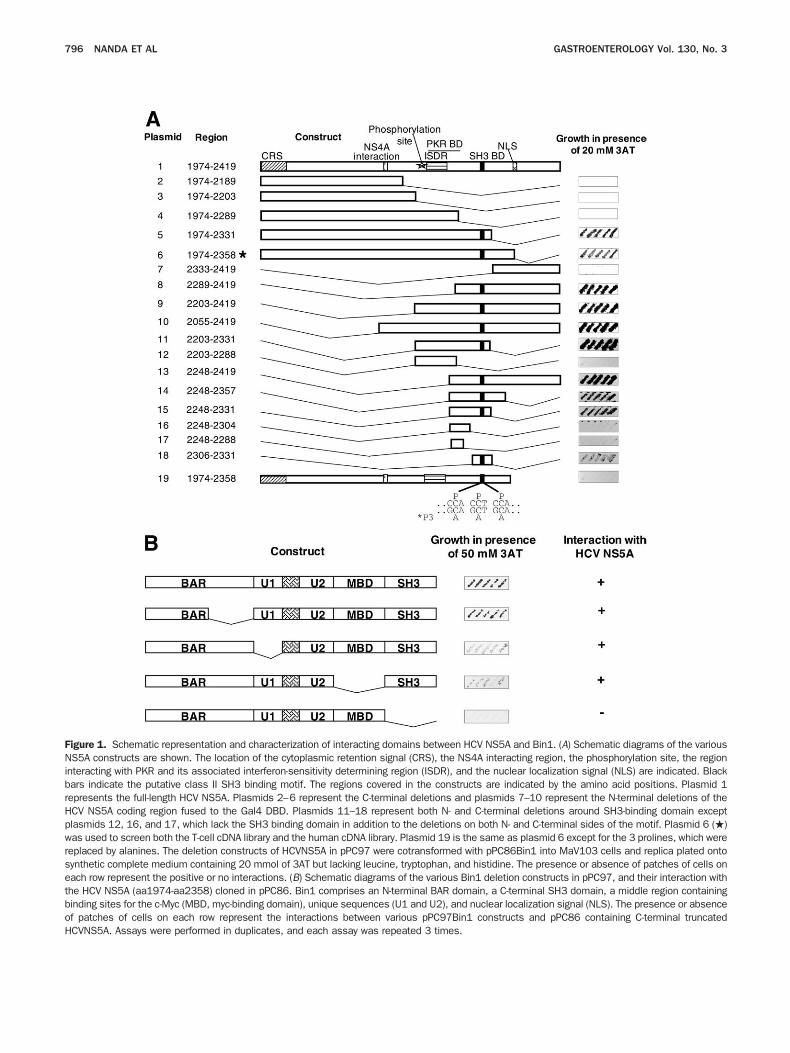
FNibrHpwrsetboH
796 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
igure 1. Schematic representation and characterization of interacting domains between HCV NS5A and Bin1. (A) Schematic diagrams of the variousS5A constructs are shown. The location of the cytoplasmic retention signal (CRS), the NS4A interacting region, the phosphorylation site, the region
nteracting with PKR and its associated interferon-sensitivity determining region (ISDR), and the nuclear localization signal (NLS) are indicated. Blackars indicate the putative class II SH3 binding motif. The regions covered in the constructs are indicated by the amino acid positions. Plasmid 1epresents the full-length HCV NS5A. Plasmids 2–6 represent the C-terminal deletions and plasmids 7–10 represent the N-terminal deletions of theCV NS5A coding region fused to the Gal4 DBD. Plasmids 11–18 represent both N- and C-terminal deletions around SH3-binding domain exceptlasmids 12, 16, and 17, which lack the SH3 binding domain in addition to the deletions on both N- and C-terminal sides of the motif. Plasmid 6 (�)as used to screen both the T-cell cDNA library and the human cDNA library. Plasmid 19 is the same as plasmid 6 except for the 3 prolines, which were
eplaced by alanines. The deletion constructs of HCVNS5A in pPC97 were cotransformed with pPC86Bin1 into MaV103 cells and replica plated ontoynthetic complete medium containing 20 mmol of 3AT but lacking leucine, tryptophan, and histidine. The presence or absence of patches of cells onach row represent the positive or no interactions. (B) Schematic diagrams of the various Bin1 deletion constructs in pPC97, and their interaction withhe HCV NS5A (aa1974-aa2358) cloned in pPC86. Bin1 comprises an N-terminal BAR domain, a C-terminal SH3 domain, a middle region containinginding sites for the c-Myc (MBD, myc-binding domain), unique sequences (U1 and U2), and nuclear localization signal (NLS). The presence or absencef patches of cells on each row represent the interactions between various pPC97Bin1 constructs and pPC86 containing C-terminal truncated
CVNS5A. Assays were performed in duplicates, and each assay was repeated 3 times.
gPwicwPitgwbZ0wabn
ouhmTliw(gSa
(Nflp1p(lCcAatmbMtb
b5ANTws(AlmcttP
51ttibEirc
tdRoipwPss�rdtrcat
R
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 797
In Vitro Binding and In VivoCoimmunoprecipitation
The full-length NS5A was cloned in frame with thelutathione S-transferase (GST) gene of pGEXKG (Amershamharmacia, Piscataway, NJ). The resulting GST fusion proteinas expressed in Escherichia coli, and solubilized from bacteria
n lysis buffer (1% Triton X-100 in PBS) by sonication. Afterentrifugation at 20,000 � g for 20 minutes, the supernatantas added to the glutathione Sepharose 4B beads (Amershamharmacia) and mixed gently at 4°C for 1–2 hours. Approx-
mately equal amounts of GST and GST-NS5A fusion pro-eins, as judged by Coomassie Blue staining, were bound tolutathione-Sepharose beads. The beads were then incubatedith in vitro translated [35S]Met-labeled Bin1 in bindinguffer (40 mmol HEPES, pH 7.4, 100 mmol KCl, 100 �molnCl2, 0.1% Nonidet P-40, 20 mmol �-mercaptoethanol, and.1 mmol phenylmethylsulfonyl fluoride [PMSF]). Beads wereashed thoroughly with the binding buffer, and proteins were
nalyzed by SDS-polyacrylamide gel electrophoresis, followedy autoradiography. [35S]Met-labeled luciferase was used as aegative control.Huh7 cells grown in 6-well plates were transfected with 1 �g
f pcDNA3FlagNS5A, the NS5A*P3 mutant, or the vector alonesing TransIT LT1 (Mirus Corporation, Madison, WI). After 48ours of transfection, cell lysates were prepared in lysis buffer (50mol Tris HCl, pH 7.4, 150 mmol NaCl, 1 mmol EDTA, 1%riton X-100) containing protease inhibitors (10 �g/mL each of
eupeptin and aprotinin, 1 mmol PMSF) and phosphatase inhib-tors (10 �mol NaVO4 and 1 mmol NaF). Each cell lysate dilutedith 1� lysis buffer was incubated with anti-Bin1 antibodies
Chemicon, Temecula, CA) bound to protein G beads and mixedently for 2 hours. The immunoprecipitates were separated byDS-PAGE and detected by Western blot using anti-Flag M2ntibody (Sigma, St. Louis, MO).
BB7 cell line containing the HCV subgenomic repliconBart79I),19 provided by Charles Rice (Rockefeller University,ew York, NY), was maintained in DMEM containing 10%
etal bovine serum and 500 �g/mL G418. The BB7 cells wereysed with the M-PER reagent (Seize X mammalian immuno-recipitation kit; Pierce, Rockford, IL). After centrifugation at3,000 � g for 5 minutes, the supernatant was analyzed forrotein concentration by Coomassie Plus Protein Assay reagentPierce). Anti-Bin1 antibody was cross-linked with immobi-ized Protein G beads (50% slurry) by disuccinimidyl suberate.ell extracts were added to antibody-coupled gel in the spinolumn and incubated with gentle mixing for at least 2 hours.n irrelevant antibody was also tested in the same manner asnegative control. Bound materials were washed thoroughly 4
imes and eluted with ImmunoPure elution buffer. The im-unoprecipitates were separated by SDS-PAGE and detected
y Western blot using anti-NS5A antibodies (Biodesign, Saco,E). For analysis of c-Myc expression, cell lysate was subjected
o SDS-PAGE and Western blotting with anti–c-Myc anti-
ody (Santa Cruz Biotech, Santa Cruz, CA). vConfocal Microscopy
HCV replicon cells were grown on Lab-Tek II cham-ered coverglass (Nalge Nunc International, Naperville, IL) to0% confluence and fixed with 3% paraformaldehyde in PBS.fter being washed with 10 mmol Tris-HCl and 50 mmolH4Cl, cells were permeabilized for 2 minutes with 1%riton-X-100 in PBS. Overnight blocking was carried outith PBS-Tween 20 (0.1%) containing 10% normal goat
erum. Both the anti-NS5A (Biodesign) and anti-Bin1Chemicon) monoclonal antibodies were labeled directly withlexa Fluor 488 and Alexa Fluor 555 Microscale Protein
abeling kit (Molecular Probes, Eugene, OR), respectively, peranufacturer’s guidelines. Antibodies were diluted with PBS
ontaining 0.1% Tween 20; all washes were performed withhe same solution. After overnight primary antibody incuba-ions, the cells were washed 4 times. Cells were embedded inroLong Antifade kit (Molecular Probes).Laser confocal microscopy was performed with Zeiss LSM
10 laser confocal microscope system at C-Apochromat 63x/.2W objective. Scanning parameters were adjusted to nega-ive control slides to eliminate nonspecific fluorescence collec-ion. Fixed parameters were used for data acquisition andmage analysis. The thickness of each digital section obtainedy the microscope was 1.2 �m at a pinhole size of 120 �m.ach experiment was performed at least 3 times, and approx-
mately 10 fields were examined to ensure that the results wereepresentative. Two or more individual cell images were re-orded for each experiment.
Screening of TranSignal SH3 DomainArrays
TranSignal SH3 domain arrays were generated usinghe recombinant conserved binding sites of individual SH3omain proteins (SH3 Domain Array I and II; Panomics,edwood City, CA). Proteins were purified and immobilizednto a membrane. Each SH3 domain on the arrays was spottedn duplicate with the top position containing 10 times morerotein than the lower position. The full-length HCV NS5Aas cloned into the ligand expression vector (pEXP vector,anomics) and transformed into DH5� cells. Protein expres-ion was induced by IPTG (isopropyl-�-D-thiogalactopyrano-ide) at 30°C, and bacterial lysate was prepared and stored at20°C until further use. Array membranes were blocked at
oom temperature for 1 hour and further incubated with theiluted bacterial extract at 4°C for 2 hours. After incubation,he membranes were washed 3 times for 10 minutes each atoom temperature. The membranes were developed withhemiluminescence reagent after incubation for 1 hour withntihistidine HRP conjugate. The spots were quantified withhe NIH Image software.
HCV Replicon Assays
The replicon plasmid Bart79I19 (from Charles Rice,ockefeller University) was digested with Sca I and used for in
itro transcription with a high-yield MEGAscript T7 tran-
selMe
h1i(trtmmf
pwfcotDf(t�aB
(eup(tHcpvlptcCceiwn2Dh
abflii
pvSTosatidispLsPt(
at
ftbiylt(ltoymtn
798 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
cription kit (Ambion, Austin, TX). RNA was purified byxtraction with TRIzol (Gibco BRL, Gaithersburg, MD),oaded onto an RNeasy minicolumn (Qiagen, Germantown,
D), and treated with DNase in the column. The RNA wasluted from the column in nuclease-free water.
For HCV replicon assay, Huh7 cells were seeded 16–20ours prior to transfection at a density of 2 � 106 cells per0-cm dish. The transfection reagent was prepared by vortex-ng vigorously 5 mL of OptiMEM I and 25 �L of DMRIE-CInvitrogen) with 10 �g of replicon RNA. Cells were washedwice with PBS and then incubated with the transfectioneagent for 6 hours at 37°C in a humidified incubator con-aining 5% CO2, followed by replacement with completeedium. For colony formation assay, the medium was supple-ented with 500 �g/mL of G418 24 hours following trans-
ection. Colonies were then counted at 4 weeks.5,20
For transient replication assay, Huh7 cells with varyingermissiveness for HCV replication20,21 (3 � 105 cells/well)ere transfected with 5 �g of the replicon RNA transcribed
rom HCV replicon carrying the luciferase gene22 and 3 cellulture-adaptive mutations that enhance RNA replication co-peratively (E1202G, T1280I, and S2197P).5 Four hours post-ransfection, cells were recovered with 2 mL of completeMEM without neutral red. At various time points posttrans-
ection, medium was removed and 100 �L of 1 X lysis bufferPromega) was added and incubated for 10 minutes at roomemperature. Twenty microliters of lysate was mixed with 100L of luciferase assay buffer (Promega), and the luciferase
ctivity was measured in a luminometer (Lumat LB9507;erthold, Pforzheim, Germany) for 15 seconds.
Apoptosis Assays
The Ad-Bin1 virus obtained from George PrendergastWistar Institute) contains Bin1 cDNA 99fE, which includesxons 1–11 and 13–16, but not brain-specific exons 12A–12D,nder the control a cytomegalovirus (CMV) enhancer-romoter.14,23 Adenovirus containing the �-galactosidase geneAd-LacZ) downstream of CMV promoter was used as a con-rol. Adenoviruses were amplified by standard methods.epG2 cells (ATCC, Rockville, MD) were grown in 6-well
ulture plates to about 50%– 60% confluence. ThecDNA3FlagNS5A, pcDNA3FlagNS5A*P3, and pCDNA3ectors were transfected separately into HepG2 cells usingipofectin (Invitrogen). After 16 hours, the medium was re-laced with fresh medium and cells were grown for an addi-ional 48 hours. The cells were then exposed to a selectiveoncentration of 800 �g/mL of geneticin (Clontech, Palo Alto,A) to isolate stably transfected cells. Clonal populations wereonfirmed by Western blot analysis for HCV NS5A proteinxpression. For the apoptosis assay, HepG2 cell lines express-ng either wild-type NS5A or NS5A*P3 mutant sequenceere plated at 2 � 106 cells per 10-cm culture dish. On theext day, virus was added to the cells at the indicated MOI for–3 hours at 37°C. Cells were then washed and fed withMEM � 10% FBS. At 48 hours postinfection, cells were
arvested by trypsinization and fixed in 1% paraformaldehyde mnd then 80% ethanol. TUNEL assay was performed by incu-ating samples for 1 hour at 37°C in TdT reaction buffer withuorescein-labeled nucleotide mix (Promega). Cells were thenncubated for 30 minutes at room temperature with propidiumodide and analyzed by flow cytometry.
Intrahepatic Inoculation of ChimpanzeesWith HCV RNA
The procedure has been described previously.15 TheHCVCG1b*P3 was linearized with BstZ17I and used for initro transcription of RNA. Chimpanzees were maintained atouthwest Foundation for Biomedical Research (San Antonio,X), an Association for Assessment and Accreditation of Lab-ratory Animal Care (ALAAC)-accredited facility, and thetudy protocol was approved by the Institutional Animal Carend Use Committee (IACUC) at the foundation. The in vitroranscribed HCV RNA was first purified and examined forntactness by denaturing formaldehyde agarose gel and thenissolved in 0.5 mL of phosphate-buffered saline (1 mg/mL) fornoculation intrahepatically into chimpanzee X205 at severalites with ultrasound guidance. After inoculation, serum sam-les were collected each week and tested for signs of infection.iver enzyme levels (ALT and AST) were determined bytandard assays. HCV RNA was assayed using the HCV RT-CR Cobas Amplicor kit (Roche Molecular Systems, Pleasan-on, CA). Anti-HCV seroconversion was monitored by EIA 2.0Abbott Laboratories, Abbott Park, IL).
Statistical Analysis
Overall significance was assessed by 1-way ANOVAnd significance between groups was performed by Student’s-test.
Results
Identification of Cellular ProteinsInteracting With HCV NS5A
On alignment of the proline-rich (PxxP) sequencerom HCV NS5A of all the genotypes of HCV, we foundhat it is conserved and belongs to the class II SH3inding motif (Figure 2). To identify cellular factorsnteracting with the HCV NS5A protein, we utilized theeast 2-hybrid system for screening of 2 human cDNAibraries. The amino-terminal 384 aa of the NS5A fusedo the Gal4 DNA binding domain was used as baitpPC97NS5A plasmid 6; Figure 1A) because the full-ength NS5A was a self-activator in this 2-hybrid sys-em. Increased HIS3 expression results as a consequencef an interacting clone. Because HIS3 expression renderseast resistant to 3AT, 3AT at a concentration of 20mol was used to optimize the growth difference be-
ween interaction-positive transformants and interaction-egative transformants for histidine auxotrophy deter-
ined on histidine dropout plates. A protein–protein
isaphlsnemcdatcoltfitBa
tm
fcuwtdtNNaomsBbwetpct
w2NacldStcCrisSB
BeaI
Fvtgg2c
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 799
nteraction was indicated by the viability of yeast cells onynthetic complete medium lacking leucine, tryptophan,nd histidine plates containing 20 mmol 3AT, and onlates containing uracil and 0.1% (wt/vol) 5FOA. Auman T-cell cDNA library and a human liver cDNAibrary were screened separately. From the T-cell librarycreen (1.5 � 106 transformants), 25 independent colo-ies were selected for growth. The corresponding cDNAncoding plasmids were isolated and retested for confir-ation of interaction on synthetic complete medium
ontaining 3AT, lacking leucine, tryptophan, and histi-ine; medium lacking leucine, tryptophan, and uracil;nd medium containing X-gal but lacking leucine andryptophan (data not shown). The cDNA clones withonfirmed interaction with NS5A were sequenced and 3f them were identified to be Bin1. From the humaniver cDNA library screen (1.65 � 106 independentransformants), 34 positive clones were selected and con-rmed. DNA sequence analysis of these clones showedhat 14 of the 34 interacting clones encode portions ofin1 cDNA (Table 1). The interaction between NS5A
igure 2. Alignment of amino acid sequences (aa 348–356) fromarious genotypes of HCV NS5A sequences. GenBank CDS transla-ion identification numbers (gi) of the aligned sequences are: 1a,i:2327074; 1b, gi:329770; 2a, gi:221650; 2b, gi:221608; 3a,i:514395; 3b, gi:676877; 4a, gi:2252489; 5a, gi:2462303; 6a, gi:326454. The alignment was generated by the CLUSTAL W and theonsensus SH3-binding motif is marked.
nd Bin1 was also confirmed by reciprocally exchanging w
he corresponding sequences in the bait and prey plas-ids of the yeast 2-hybrid system (data not shown).
The NS5A–Bin1 Interaction Is Mediated bythe SH3 Domain
To determine the region of HCV NS5A requiredor interaction with Bin1, yeast 2-hybrid analyses werearried out using various NS5A deletion constructs (Fig-re 1A). The yeast plasmids used for the 2-hybrid systemere co-introduced into MaV103 yeast strain, and the
ransformants were grown on synthetic complete me-ium containing 20 mmol of 3AT but lacking leucine,ryptophan, and histidine. As shown in Figure 1A, theS5A C-terminal deletion constructs (plasmid 2–6) and-terminal deletion constructs (plasmid 7–10) were re-
ctive. Additional truncation experiments showed thatnly constructs containing the SH3 binding motif (plas-ids 11, 13, 14, and 15) interacted with Bin1. The
mallest NS5A construct (plasmid 18) that interacts within1 contains aa 2306–2331, which includes the SH3inding motif, although the interaction appeared to beeaker than the other longer constructs. This could be
xplained by the possibility that some sequences outsidehis region are required either for Bin1 interaction orroper protein folding. These observations were alsoonfirmed by �-gal assay as an independent reporter ofhe yeast 2-hybrid interaction (data not shown).
To define the interacting domain of Bin1 with NS5A,e generated several deletion constructs of Bin1 for yeast-hybrid analysis. The Bin1 gene product comprises an-terminal BAR domain, a C-terminal SH3 domain,
nd a middle region containing binding sites for the-Myc (MBD, myc-binding domain) preceded by nuclearocalization signal (NLS) (Figure 1B). As expected, alleletion constructs of Bin1 except for the deletion ofH3 domain interacted with NS5A. To further evaluatehe SH3 binding motif of NS5A in this interaction, theonserved 3 proline residues (nucleotide C7314G,7317G, and C7320G) were mutated to alanine. The
esulting construct NS5A*P3 mutant was unable tonteract with Bin1 (plasmid 19, Figure 1A), furtherupporting the importance of the proline-rich region as aH3 binding motif interacting with the SH3 domain ofin1.
NS5A Binds to Bin1 In Vitro and In Vivo
To confirm an interaction between the NS5A andin1, we performed a GST pull-down assay. E coli-xpressed NS5A fused to GST (GST-NS5A) or GSTlone was purified with glutathione-Sepharose 4B beads.n vitro translated [35S]Met-labeled Bin1 or luciferase
as incubated with GST-NS5A or GST alone to assay for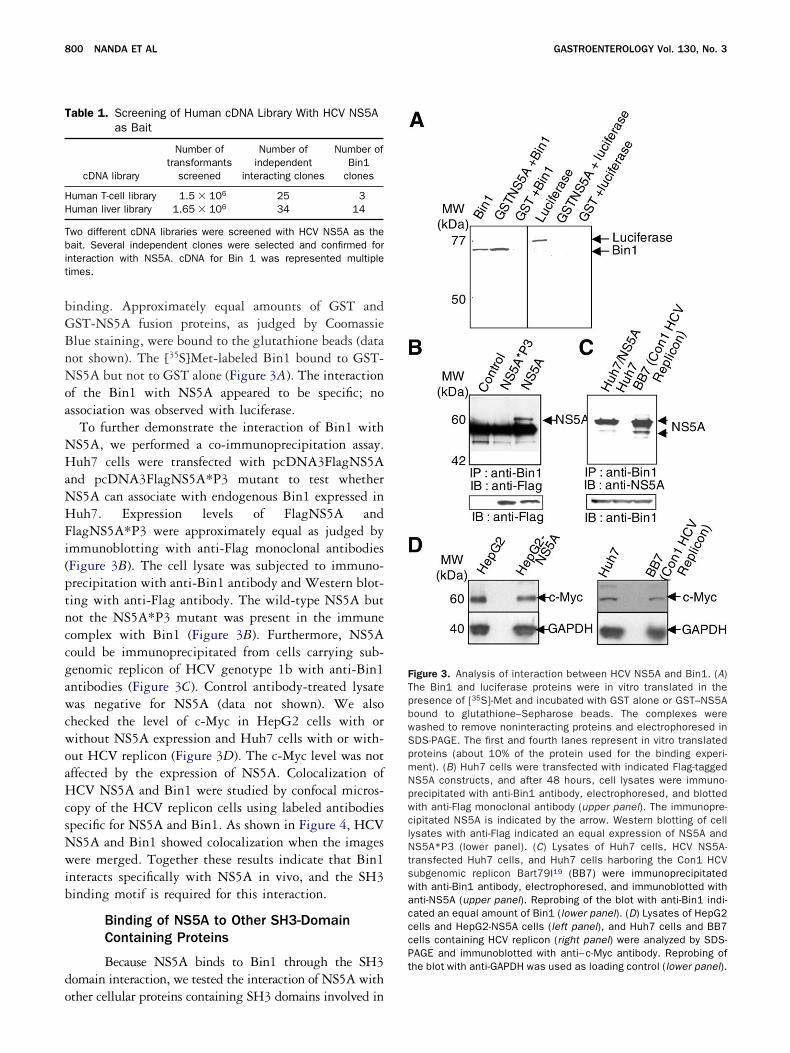
bGBnNoa
NHaNHFi(ptnccgawcwoaHcsNwib
do
FTpbwSpmNpwclNtswacccPthe blot with anti-GAPDH was used as loading control (lower panel).
T
HH
Tbit
800 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
inding. Approximately equal amounts of GST andST-NS5A fusion proteins, as judged by Coomassielue staining, were bound to the glutathione beads (dataot shown). The [35S]Met-labeled Bin1 bound to GST-S5A but not to GST alone (Figure 3A). The interaction
f the Bin1 with NS5A appeared to be specific; nossociation was observed with luciferase.
To further demonstrate the interaction of Bin1 withS5A, we performed a co-immunoprecipitation assay.uh7 cells were transfected with pcDNA3FlagNS5A
nd pcDNA3FlagNS5A*P3 mutant to test whetherS5A can associate with endogenous Bin1 expressed inuh7. Expression levels of FlagNS5A and
lagNS5A*P3 were approximately equal as judged bymmunoblotting with anti-Flag monoclonal antibodiesFigure 3B). The cell lysate was subjected to immuno-recipitation with anti-Bin1 antibody and Western blot-ing with anti-Flag antibody. The wild-type NS5A butot the NS5A*P3 mutant was present in the immuneomplex with Bin1 (Figure 3B). Furthermore, NS5Aould be immunoprecipitated from cells carrying sub-enomic replicon of HCV genotype 1b with anti-Bin1ntibodies (Figure 3C). Control antibody-treated lysateas negative for NS5A (data not shown). We also
hecked the level of c-Myc in HepG2 cells with orithout NS5A expression and Huh7 cells with or with-ut HCV replicon (Figure 3D). The c-Myc level was notffected by the expression of NS5A. Colocalization ofCV NS5A and Bin1 were studied by confocal micros-
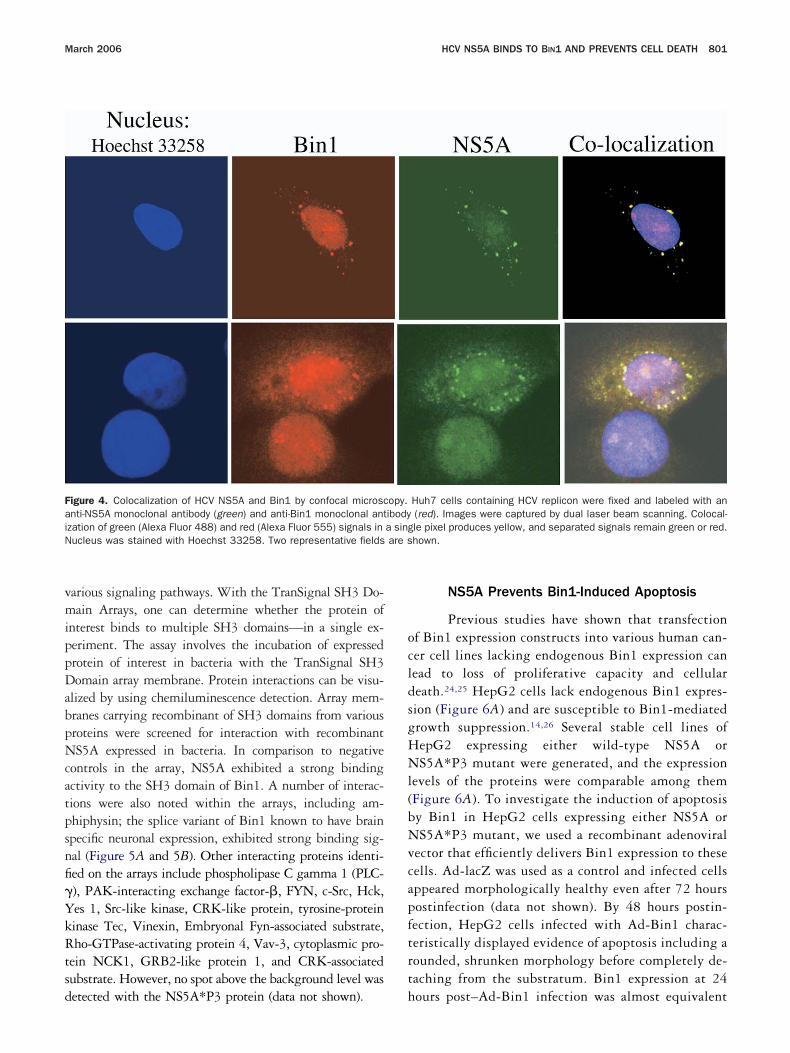
opy of the HCV replicon cells using labeled antibodiespecific for NS5A and Bin1. As shown in Figure 4, HCVS5A and Bin1 showed colocalization when the imagesere merged. Together these results indicate that Bin1
nteracts specifically with NS5A in vivo, and the SH3inding motif is required for this interaction.
Binding of NS5A to Other SH3-DomainContaining Proteins
Because NS5A binds to Bin1 through the SH3omain interaction, we tested the interaction of NS5A with
able 1. Screening of Human cDNA Library With HCV NS5Aas Bait
cDNA library
Number oftransformants
screened
Number ofindependent
interacting clones
Number ofBin1
clones
uman T-cell library 1.5 � 106 25 3uman liver library 1.65 � 106 34 14
wo different cDNA libraries were screened with HCV NS5A as theait. Several independent clones were selected and confirmed for
nteraction with NS5A. cDNA for Bin 1 was represented multipleimes.
ther cellular proteins containing SH3 domains involved in
igure 3. Analysis of interaction between HCV NS5A and Bin1. (A)he Bin1 and luciferase proteins were in vitro translated in theresence of [35S]-Met and incubated with GST alone or GST–NS5Aound to glutathione–Sepharose beads. The complexes wereashed to remove noninteracting proteins and electrophoresed inDS-PAGE. The first and fourth lanes represent in vitro translatedroteins (about 10% of the protein used for the binding experi-ent). (B) Huh7 cells were transfected with indicated Flag-taggedS5A constructs, and after 48 hours, cell lysates were immuno-recipitated with anti-Bin1 antibody, electrophoresed, and blottedith anti-Flag monoclonal antibody (upper panel). The immunopre-ipitated NS5A is indicated by the arrow. Western blotting of cellysates with anti-Flag indicated an equal expression of NS5A andS5A*P3 (lower panel). (C) Lysates of Huh7 cells, HCV NS5A-
ransfected Huh7 cells, and Huh7 cells harboring the Con1 HCVubgenomic replicon Bart79I19 (BB7) were immunoprecipitatedith anti-Bin1 antibody, electrophoresed, and immunoblotted withnti-NS5A (upper panel). Reprobing of the blot with anti-Bin1 indi-ated an equal amount of Bin1 (lower panel). (D) Lysates of HepG2ells and HepG2-NS5A cells (left panel), and Huh7 cells and BB7ells containing HCV replicon (right panel) were analyzed by SDS-AGE and immunoblotted with anti– c-Myc antibody. Reprobing of
vmippDabpNcatpsnfiYkRtsd
ocldsgHNl(bNvcapftrt
Fai a singN are s
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 801
arious signaling pathways. With the TranSignal SH3 Do-ain Arrays, one can determine whether the protein of
nterest binds to multiple SH3 domains—in a single ex-eriment. The assay involves the incubation of expressedrotein of interest in bacteria with the TranSignal SH3omain array membrane. Protein interactions can be visu-
lized by using chemiluminescence detection. Array mem-ranes carrying recombinant of SH3 domains from variousroteins were screened for interaction with recombinantS5A expressed in bacteria. In comparison to negative
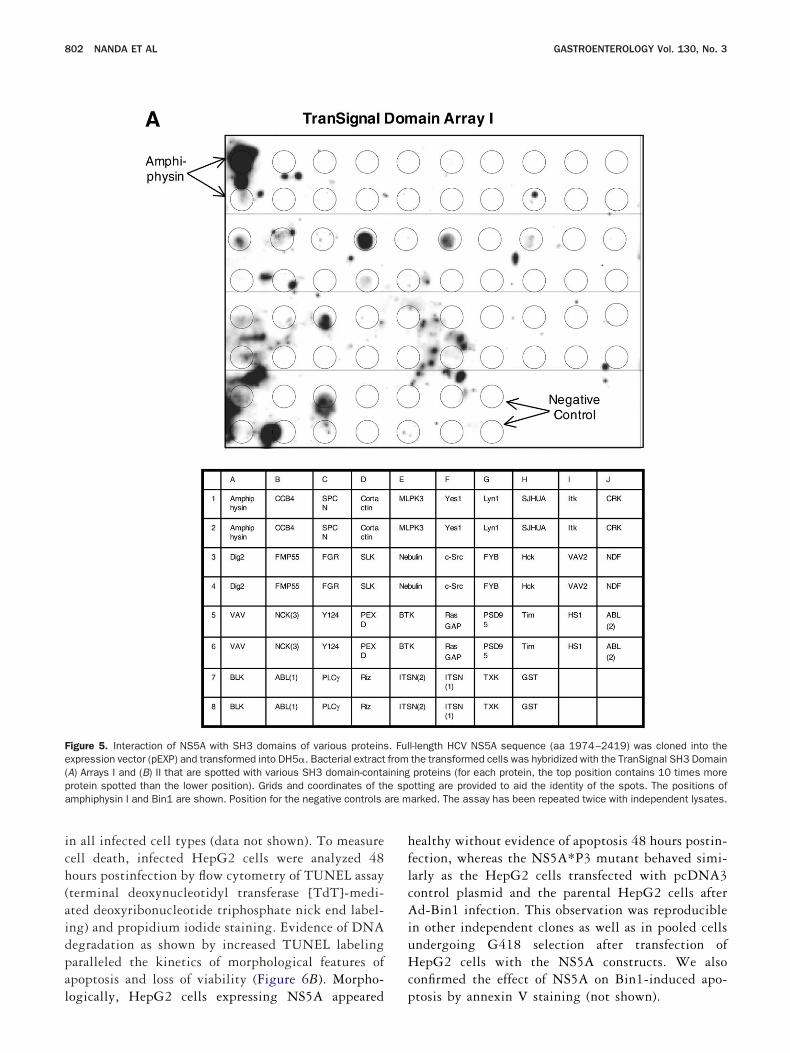
ontrols in the array, NS5A exhibited a strong bindingctivity to the SH3 domain of Bin1. A number of interac-ions were also noted within the arrays, including am-hiphysin; the splice variant of Bin1 known to have brainpecific neuronal expression, exhibited strong binding sig-al (Figure 5A and 5B). Other interacting proteins identi-ed on the arrays include phospholipase C gamma 1 (PLC-), PAK-interacting exchange factor-�, FYN, c-Src, Hck,es 1, Src-like kinase, CRK-like protein, tyrosine-proteininase Tec, Vinexin, Embryonal Fyn-associated substrate,ho-GTPase-activating protein 4, Vav-3, cytoplasmic pro-
ein NCK1, GRB2-like protein 1, and CRK-associatedubstrate. However, no spot above the background level was
igure 4. Colocalization of HCV NS5A and Bin1 by confocal microscnti-NS5A monoclonal antibody (green) and anti-Bin1 monoclonal antzation of green (Alexa Fluor 488) and red (Alexa Fluor 555) signals inucleus was stained with Hoechst 33258. Two representative fields
etected with the NS5A*P3 protein (data not shown). h
NS5A Prevents Bin1-Induced Apoptosis
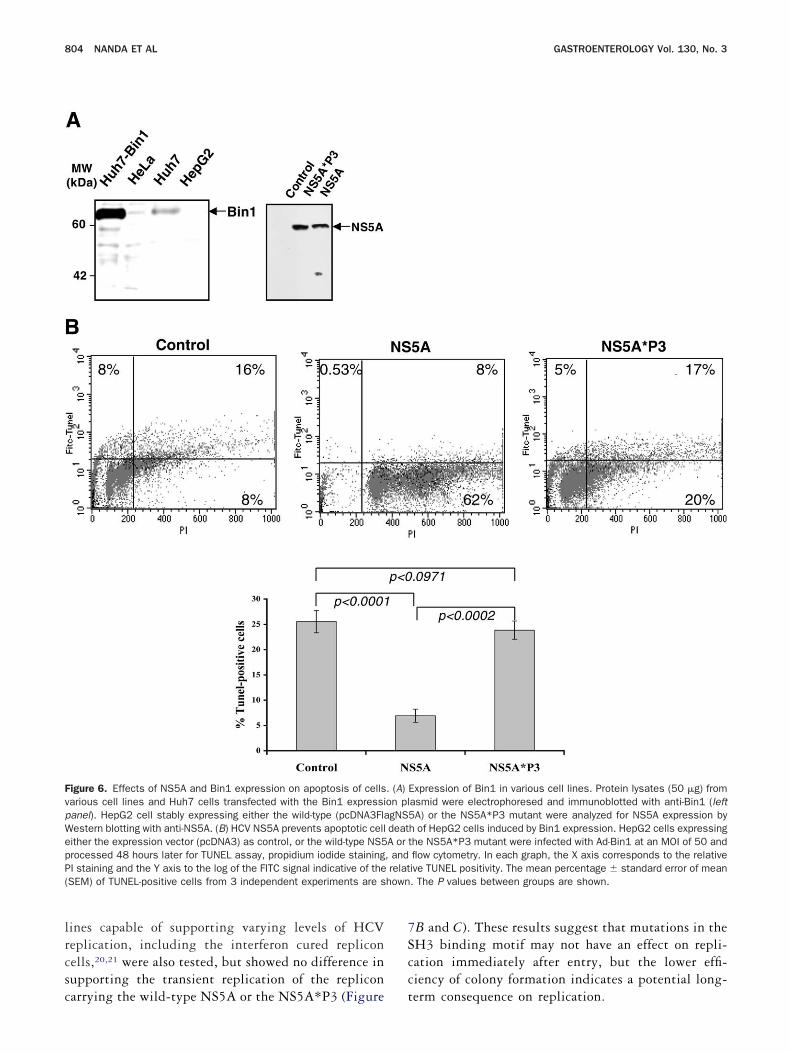
Previous studies have shown that transfectionf Bin1 expression constructs into various human can-er cell lines lacking endogenous Bin1 expression canead to loss of proliferative capacity and cellulareath.24,25 HepG2 cells lack endogenous Bin1 expres-ion (Figure 6A) and are susceptible to Bin1-mediatedrowth suppression.14,26 Several stable cell lines ofepG2 expressing either wild-type NS5A orS5A*P3 mutant were generated, and the expression
evels of the proteins were comparable among themFigure 6A). To investigate the induction of apoptosisy Bin1 in HepG2 cells expressing either NS5A orS5A*P3 mutant, we used a recombinant adenoviral
ector that efficiently delivers Bin1 expression to theseells. Ad-lacZ was used as a control and infected cellsppeared morphologically healthy even after 72 hoursostinfection (data not shown). By 48 hours postin-ection, HepG2 cells infected with Ad-Bin1 charac-eristically displayed evidence of apoptosis including aounded, shrunken morphology before completely de-aching from the substratum. Bin1 expression at 24
Huh7 cells containing HCV replicon were fixed and labeled with an(red). Images were captured by dual laser beam scanning. Colocal-le pixel produces yellow, and separated signals remain green or red.hown.
opy.ibody
ours post–Ad-Bin1 infection was almost equivalent
ich(aidpal
hflcAiuHc
Fe(p e spa are m
802 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
n all infected cell types (data not shown). To measureell death, infected HepG2 cells were analyzed 48ours postinfection by flow cytometry of TUNEL assayterminal deoxynucleotidyl transferase [TdT]-medi-ted deoxyribonucleotide triphosphate nick end label-ng) and propidium iodide staining. Evidence of DNAegradation as shown by increased TUNEL labelingaralleled the kinetics of morphological features ofpoptosis and loss of viability (Figure 6B). Morpho-
igure 5. Interaction of NS5A with SH3 domains of various proteinsxpression vector (pEXP) and transformed into DH5�. Bacterial extractA) Arrays I and (B) II that are spotted with various SH3 domain-contarotein spotted than the lower position). Grids and coordinates of thmphiphysin I and Bin1 are shown. Position for the negative controls
ogically, HepG2 cells expressing NS5A appeared p
ealthy without evidence of apoptosis 48 hours postin-ection, whereas the NS5A*P3 mutant behaved simi-arly as the HepG2 cells transfected with pcDNA3ontrol plasmid and the parental HepG2 cells afterd-Bin1 infection. This observation was reproducible
n other independent clones as well as in pooled cellsndergoing G418 selection after transfection ofepG2 cells with the NS5A constructs. We also
onfirmed the effect of NS5A on Bin1-induced apo-
l-length HCV NS5A sequence (aa 1974–2419) was cloned into thethe transformed cells was hybridized with the TranSignal SH3 Domainproteins (for each protein, the top position contains 10 times more
otting are provided to aid the identity of the spots. The positions ofarked. The assay has been repeated twice with independent lysates.
. Fulfromining
tosis by annexin V staining (not shown).

mtlpe(aac
wmrd
twippd
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 803
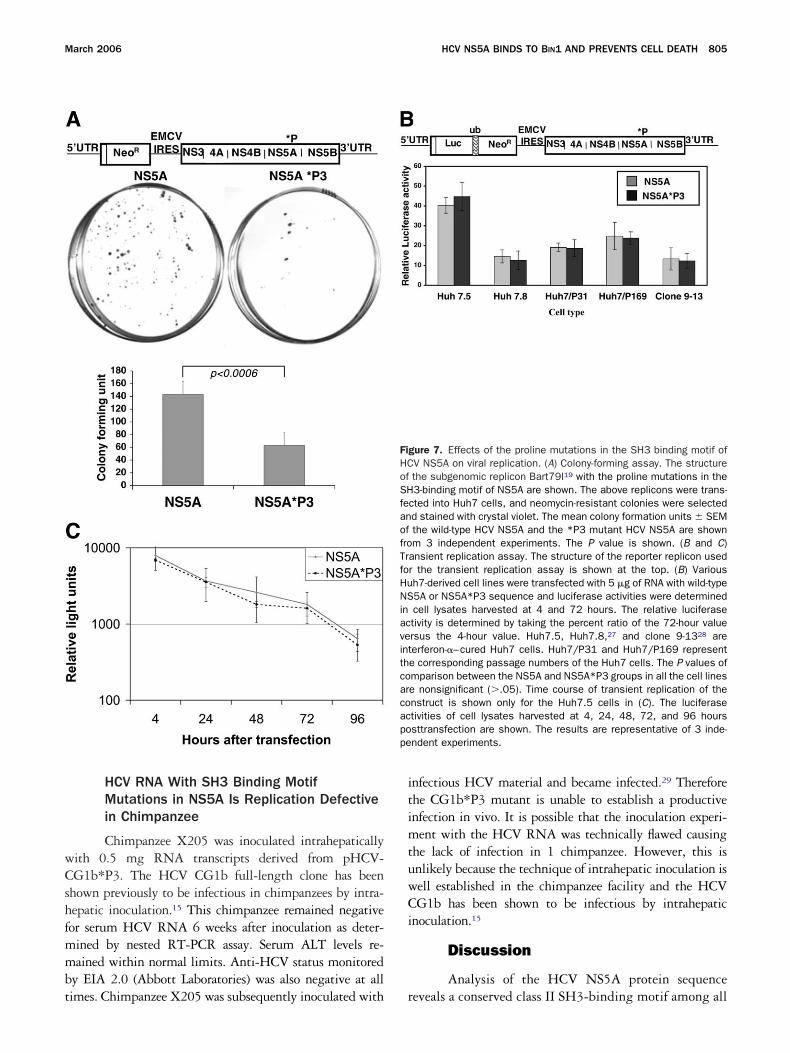
Effects of Mutations in the SH3 BindingMotif in NS5A on HCV RNA Replication
The conservation of this class II SH3 bindingotif in all known HCV isolates suggest that muta-
ion of this motif might be detrimental to the HCVife cycle. To test this hypothesis, we introduced theroline mutations into the NS5A gene of a high-fficiency bicistronic subgenomic HCV repliconBart79I).19 The replication efficiency of the wild-typend NS5A*P3 replicons were first tested for theirbility to establish G418-resistant colonies in Huh7
Fig. 5
ells. As shown in Figure 7A, although the replicon t
ith wild-type NS5A gave rise to numerous colonies,utations within the SH3-binding motif of NS5A
esulted in decreased colony formation (�3-fold re-uction) that was statistically significant (P .0006).To assess the effect of NS5A*P3 mutations on
ransient HCV replication, replicon RNA with orithout the NS5A*P3 mutations were transfected
nto Huh7 cells and harvested at 4 and 72 hoursosttransfection. The replication efficiency was ex-ressed as the relative luciferase activity calculated byividing the light units of the sample at 72 hours by
nt’d)
. (Cohat at 4 hours posttransfection. Different Huh7 cell
lrcsc
7Scc
FvpWepP( hown
804 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
ines capable of supporting varying levels of HCVeplication, including the interferon cured repliconells,20,21 were also tested, but showed no difference inupporting the transient replication of the replicon
igure 6. Effects of NS5A and Bin1 expression on apoptosis of cellsarious cell lines and Huh7 cells transfected with the Bin1 expressianel). HepG2 cell stably expressing either the wild-type (pcDNA3Flaestern blotting with anti-NS5A. (B) HCV NS5A prevents apoptotic cellither the expression vector (pcDNA3) as control, or the wild-type NS5rocessed 48 hours later for TUNEL assay, propidium iodide stainingI staining and the Y axis to the log of the FITC signal indicative of theSEM) of TUNEL-positive cells from 3 independent experiments are s
arrying the wild-type NS5A or the NS5A*P3 (Figure t
B and C). These results suggest that mutations in theH3 binding motif may not have an effect on repli-ation immediately after entry, but the lower effi-iency of colony formation indicates a potential long-
Expression of Bin1 in various cell lines. Protein lysates (50 �g) fromasmid were electrophoresed and immunoblotted with anti-Bin1 (left5A) or the NS5A*P3 mutant were analyzed for NS5A expression byh of HepG2 cells induced by Bin1 expression. HepG2 cells expressingthe NS5A*P3 mutant were infected with Ad-Bin1 at an MOI of 50 andflow cytometry. In each graph, the X axis corresponds to the relativeive TUNEL positivity. The mean percentage � standard error of mean. The P values between groups are shown.
. (A)on plgNSdeatA or, andrelat
erm consequence on replication.
wCshfmmbt
itimtuwCi
pe
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 805
HCV RNA With SH3 Binding MotifMutations in NS5A Is Replication Defectivein Chimpanzee
Chimpanzee X205 was inoculated intrahepaticallyith 0.5 mg RNA transcripts derived from pHCV-G1b*P3. The HCV CG1b full-length clone has been
hown previously to be infectious in chimpanzees by intra-epatic inoculation.15 This chimpanzee remained negativeor serum HCV RNA 6 weeks after inoculation as deter-ined by nested RT-PCR assay. Serum ALT levels re-ained within normal limits. Anti-HCV status monitored
y EIA 2.0 (Abbott Laboratories) was also negative at all
imes. Chimpanzee X205 was subsequently inoculated with rnfectious HCV material and became infected.29 Thereforehe CG1b*P3 mutant is unable to establish a productivenfection in vivo. It is possible that the inoculation experi-ent with the HCV RNA was technically flawed causing
he lack of infection in 1 chimpanzee. However, this isnlikely because the technique of intrahepatic inoculation isell established in the chimpanzee facility and the HCVG1b has been shown to be infectious by intrahepatic
noculation.15
Discussion
Analysis of the HCV NS5A protein sequence
gure 7. Effects of the proline mutations in the SH3 binding motif ofV NS5A on viral replication. (A) Colony-forming assay. The structurethe subgenomic replicon Bart79I19 with the proline mutations in the3-binding motif of NS5A are shown. The above replicons were trans-
cted into Huh7 cells, and neomycin-resistant colonies were selectedd stained with crystal violet. The mean colony formation units � SEMthe wild-type HCV NS5A and the *P3 mutant HCV NS5A are shownm 3 independent experiments. The P value is shown. (B and C)
ansient replication assay. The structure of the reporter replicon usedr the transient replication assay is shown at the top. (B) Varioush7-derived cell lines were transfected with 5 �g of RNA with wild-type5A or NS5A*P3 sequence and luciferase activities were determinedcell lysates harvested at 4 and 72 hours. The relative luciferasetivity is determined by taking the percent ratio of the 72-hour valuersus the 4-hour value. Huh7.5, Huh7.8,27 and clone 9-1328 areerferon-�–cured Huh7 cells. Huh7/P31 and Huh7/P169 represente corresponding passage numbers of the Huh7 cells. The P values ofmparison between the NS5A and NS5A*P3 groups in all the cell linese nonsignificant (�.05). Time course of transient replication of thenstruct is shown only for the Huh7.5 cells in (C). The luciferasetivities of cell lysates harvested at 4, 24, 48, 72, and 96 hourssttransfection are shown. The results are representative of 3 inde-ndent experiments.
FiHCofSHfeanoffroTrfoHuNSinacveintthcoarcoacpo
eveals a conserved class II SH3-binding motif among all

giNBpcivHsBdtDdettfiktkilbiwiipatstqwttp
tBtdBApntep
csttfmdBsiciidvtmaaoepp
dorsaeveHaaapipsaet
twNfpNq
806 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
enotypes. Using the yeast 2-hybrid system to screen fornteracting cellular proteins, we demonstrated thatS5A interacts with a newly identified tumor suppressorin1, which contains a SH3 domain. The C-terminalart of the NS5A containing the SH3-binding motif wasrucial for the interaction of Bin1. The protein–proteinnteraction between NS5A and Bin1 was confirmed by initro binding and in vivo coimmunoprecipitation inuh7 cells with endogenous Bin1 expression. NS5A was
hown to form an immunoprecipitable complex within1 in the HCV replicon cells. Confocal microscopyemonstrated that NS5A and Bin1 colocalize to a dis-inct membranous compartment in the replicon cells.eletion and mutation analyses indicated that the SH3omain of Bin1 and the SH3 binding motif of NS5A aressential for the interaction. Recently, through pro-eomic analysis, Zech et al30 demonstrated the associa-ion of NS5A and Amph I, a Bin1-related protein but nounctional analysis of this interaction was provided. Us-ng a protein array containing SH3 domains from manynown proteins, we identified several proteins that in-eract with NS5A. Members of the Src family of tyrosineinases Fyn and Hck had been shown previously tonteract with NS5A,31 whereas others like PLC-, CRK-ike protein, Tec, vinexin, and Vav-3 were not known toe involved in interaction with NS5A. Some of the newlydentified interactors in the protein array, like vinexin,as also tested in the yeast 2-hybrid system, but no
nteraction could be observed. The yeast 2-hybrid systems probably a more stringent system to assess protein–rotein interaction than the protein array. The function-lity of these additional interactions needs to be studiedo establish their biological significance. Although thepecificity of interaction depends on amino acids flankinghe core PxxP motif and the overall SH3 domain se-uences,32 the observation that NS5A appears to interactith a variety of SH3 domain-containing cellular pro-
eins involved in various signaling pathways may explainhe pleiotropic functions associated with the HCV NS5Arotein.2
Bin1 is a nucleocytoplasmic c-Myc–interacting pro-ein with tumor suppressor and apoptotic properties.in1 and its isoforms contain a highly conserved amino-
erminal region (BAR domain), a carboxy-terminal SH3omain, and a variable central region. Four mammalianAR protein-encoding genes have been reported so far:mphiphysin I (Amph I), Bin1 (also known as Am-hiphysin II), Bin2, and Bin3.33 Amphiphysin-I is aeuronal protein enriched in nerve terminals and is likelyo be involved in endocytosis. Amphiphysin II is widelyxpressed in different tissues, and is most highly ex-
ressed in the brain and striated muscle. Amphiphysin II tonsists of �6 isoforms generated by alternative mRNAplicing, all of which contain the common carboxy-erminal SH3 domain,34 by which Amphiphysin II bindso canonical PxxP sequences. Loss of Bin1 expression is arequently associated with malignancies.14,35,36 Theechanism responsible for loss of the Bin1 expression
uring malignant transformation is not well understood.in1 functionally associates with c-Myc in cells and
electively inhibits the oncogenic properties of c-Myc byts pro-apoptotic action.14,25 Epigenetic inactivation orhromosomal loss of the Bin1 gene that frequently occursn human tumor cells may enable these cells to escape thenduction of apoptosis associated with c-Myc activationuring transformation.25,37 Progressive inactivation ofarious suicide mechanisms are characteristic of neoplas-ic progression.38 The anti-neoplastic effects of Bin1 inalignant cells are a consequence of their ability to
ctivate programmed cell death. Bin1 alone is not pro-poptotic when overexpressed in many cell types, but itsverexpression induces apoptosis in cells lacking endog-nous Bin1 where c-Myc is deregulated. Therefore, Bin1rovides a conditional linkage between c-Myc, deathathway, and malignant transformation.39
Programmed cell death constitutes an important en-ogenous defense system against virus infection. Eukary-tic cells have the innate ability to undergo apoptosis inesponse to viral infection to limit viral replication andpread. In addition, cytotoxic T cells, the mainstay ofdaptive immunity against viral infection, function toliminate virus-infected cells by induction of apoptosisia several death mechanisms. However, many virusesncode anti-apoptotic genes to counter this defense.40
CV may also evolve a similar mechanism as in thenti-apoptotic function of NS5A to ensure a productivend persistent infection. In addition to the inhibition ofpoptosis induced by Bin1 shown here, NS5A also im-airs TNF-�–mediated apoptosis.41 Finally, NS5A alsonteracts and interferes with the activity of PKR, whichlays a major role in interferon-activated antiviral re-ponse and apoptosis.42 Therefore, NS5A appears to actt several levels of host antiviral response, resulting in annvironment that is favorable to productive viral infec-ion.2
It is known that NS5A interacts with other nonstruc-ural proteins in a membrane-bound replicase complexhere viral replication occurs.6 Several domains of theS5A have been shown to be required for this complex
ormation and viral replication. An N-terminal amphi-athic helix is important for membrane anchoring of theS5A in the replicase complex and several internal se-
uences of the NS5A are required for interaction with
he viral RNA-dependent RNA polymerase. Inactivation
oawrsome5HNSf
StfiiTitowctrntaitcHHtuacovttisrtairms
minhvtcpsdBfHliedcagbidcwc
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 807
f any of these domains resulted in defective replication,t least in the HCV replicon model.19 It is not knownhether the conserved SH3 binding motif of NS5A is
equired for viral replication. Using the HCV repliconystem, we could not demonstrate any significant effectf the NS5A*P3 mutations on replication, other than aodest reduction in the colony formation assay. How-
ver, Zech et al,30 in a recent publication, did show a0% reduction in the transient replication assay withCV mutant defective in the SH3 binding motif ofS5A. These effects notwithstanding, the function of the
H3 binding motif in NS5A appears to be dispensableor viral replication, at least in the replicon system.
HCV RNA harboring the proline mutations in theH3 binding motif of NS5A appeared not to be infec-ious in chimpanzees. Although it remains to be con-rmed in another animal, this observation raised annteresting point about the function of NS5A in vivo.he function of NS5A as mediated by the SH3 binding
s probably crucial for certain steps of productive infec-ion in vivo. This is consistent with the pleiotropic effectf NS5A in interacting with a variety of signaling path-ays in cells.2 The anti-apoptotic activity of NS5A dis-
ussed is consistent with this hypothesis. The findinghat the NS5A*P3 mutant was less efficient in theeplicon colony formation assay is also supportive of thisotion, because colony formation likely requires addi-ional events other than mere viral replication. There aremple examples in other viruses where viral genes aremportant for in vivo infection but not in vitro replica-ion, and many of these genes interact extensively withellular factors to affect profound cellular alterations.11
owever several caveats need to be considered. First, theCV replicon system does not completely reproduce all
he events in viral replication. The binding, entry, andncoating events are not included, and neither is thessembly process. It is possible that NS5A may play aritical role in any of these processes. The fact that somef the Bin1-related proteins are intimately involved inesicular transport suggests that NS5A may be impor-ant in viral assembly.33 Moreover, a novel adaptor pro-ein, amphiphysin IIm, is required for particle internal-zation during phagocytosis.43 Second, the repliconystem appeared to require different factors for efficienteplication from those in vivo infections, as evidenced byhe noninfectious nature of the HCV RNA harboring thedaptive mutations that confer a high level of replicationn vitro.44 NS5A could be one of the factors differentiallyequired for the 2 processes. It is interesting to note thatany of the highly adaptive mutations in the replicon
ystem occur in the NS5A region.1,5
It is not clear whether Bin1 plays a role in the develop-ent of liver cancer. In HepG2 cells where Bin1 expression
s absent, chromosomal loss or gross rearrangement appearsot to be the cause.26 Like other cancers, the development ofepatocellular carcinoma (HCC) is a multistep process in-olving multiple genetic and epigenetic alterations of hepa-ocytes.45 The long latency of HCV-associated HCC indi-ates that, in addition to a direct oncogenic and genotoxicotential of HCV proteins and replication, other eventsuch as interference of apoptosis likely occur during tumorevelopment. It is possible that NS5A, by interacting within1, interferes with the apoptosis pathway to set the stage
or additional genetic or epigenetic changes that lead toCC formation. As discussed, Bin1’s function is closely
inked to c-Myc induced apoptosis and c-Myc deregulations commonly observed in the pathogenesis of HCC.46 How-ver, transgenic mice expressing NS5A in the liver do notevelop HCC, suggesting that NS5A by itself is not suffi-ient to cause liver cancer.47 This is consistent with thencillary role of Bin1 as a Myc-dependent anti-apoptoticene in cellular transformation. The molecular mechanismsy which NS5A functions to contribute to HCC formationn the context of chronic HCV infection remains to beefined. However, the identification and characterization ofertain cellular pathways, such as the interaction of NS5Aith Bin1, underscores an important effort to elucidate this
omplex process.
References1. Evans MJ, Rice CM, Goff SP. Phosphorylation of hepatitis C virus
nonstructural protein 5A modulates its protein interactions andviral RNA replication. Proc Natl Acad Sci U S A 2004;101:13038–13043.
2. Macdonald A, Harris M. Hepatitis C virus NS5A: tales of a pro-miscuous protein. J Gen Virol 2004;85:2485–2502.
3. Brechot C. The direct interplay between HCV NS5A protein andinterferon transduction signal: from clinical to basic science.J Hepatol 1999;30:1152–1154.
4. Blight KJ, Kolykhalov AA, Rice CM. Efficient initiation of HCV RNAreplication in cell culture. Science 2000;290:1972–1974.
5. Krieger N, Lohmann V, Bartenschlager R. Enhancement of hepa-titis C virus RNA replication by cell culture-adaptive mutations.J Virol 2001;75:4614–4624.
6. Moradpour D, Evans MJ, Gosert R, Yuan Z, Blum HE, Goff SP,Lindenbach BD, Rice CM. Insertion of green fluorescent proteininto nonstructural protein 5A allows direct visualization of func-tional hepatitis C virus replication complexes. J Virol 2004;78:7400–7409.
7. He Y, Nakao H, Tan SL, Polyak SJ, Neddermann P, Vijaysri S,Jacobs BL, Katze MG. Subversion of cell signaling pathways byhepatitis C virus nonstructural 5A protein via interaction withGrb2 and P85 phosphatidylinositol 3-kinase. J Virol 2002;76:9207–9217.
8. Gale MJ Jr, Korth MJ, Tang NM, Tan SL, Hopkins DA, Dever TE,Polyak SJ, Gretch DR, Katze MG. Evidence that hepatitis C virusresistance to interferon is mediated through repression of thePKR protein kinase by the nonstructural 5A protein. Virology
1997;230:217–227.
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
808 NANDA ET AL GASTROENTEROLOGY Vol. 130, No. 3
9. Ghosh AK, Steele R, Meyer K, Ray R, Ray RB. Hepatitis C virusNS5A protein modulates cell cycle regulatory genes and pro-motes cell growth. J Gen Virol 1999;80:1179–1183.
0. Tan SL, Nakao H, He Y, Vijaysri S, Neddermann P, Jacobs BL,Mayer BJ, Katze MG. NS5A, a nonstructural protein of hepatitis Cvirus, binds growth factor receptor-bound protein 2 adaptor pro-tein in a Src homology 3 domain/ligand-dependent manner andperturbs mitogenic signaling. Proc Natl Acad Sci U S A 1999;96:5533–5538.
1. Krajcsi P, Wold WS. Viral proteins that regulate cellular signalling.J Gen Virol 1998;79:1323–1335.
2. Mayer BJ. SH3 domains: complexity in moderation. J Cell Sci2001;114:1253–1263.
3. Musacchio A. How SH3 domains recognize proline. Adv ProteinChem 2002;61:211–268.
4. Sakamuro D, Elliott KJ, Wechsler-Reya R, Prendergast GC. BIN1is a novel MYC-interacting protein with features of a tumoursuppressor. Nat Genet 1996;14:69–77.
5. Thomson M, Nascimbeni M, Gonzales S, Murthy KK, RehermannB, Liang TJ. Emergence of a distinct pattern of viral mutations inchimpanzees infected with a homogeneous inoculum of hepatitisC virus. Gastroenterology 2001;121:1226–1233.
6. Wechsler-Reya R, Elliott K, Herlyn M, Prendergast GC. The puta-tive tumor suppressor BIN1 is a short-lived nuclear phosphopro-tein, the localization of which is altered in malignant cells. CancerRes 1997;57:3258–3263.
7. Sardet C, Vidal M, Cobrinik D, Geng Y, Onufryk C, Chen A,Weinberg RA. E2F-4 and E2F-5, two members of the E2F family,are expressed in the early phases of the cell cycle. Proc Natl AcadSci U S A 1995;92:2403–2407.
8. Treco DA, Winston F. Growth and manipulation of yeast, unit13.2. In: Ausbel FM, Brent R, Kingston RE, et al, eds. Currentprotocols in molecular biopsy. New York, NY: John Wiley & Sons,Inc, 1995.
9. Elazar M, Cheong KH, Liu P, Greenberg HB, Rice CM, Glenn JS.Amphipathic helix-dependent localization of NS5A mediates hep-atitis C virus RNA replication. J Virol 2003;77:6055–6061.
0. Blight KJ, McKeating JA, Rice CM. Highly permissive cell lines forsubgenomic and genomic hepatitis C virus RNA replication. J Virol2002;76:13001–13014.
1. Lohmann V, Hoffmann S, Herian U, Penin F, Bartenschlager R.Viral and cellular determinants of hepatitis C virus RNA replica-tion in cell culture. J Virol 2003;77:3007–3019.
2. Vrolijk JM, Kaul A, Hansen BE, Lohmann V, Haagmans BL,Schalm SW, Bartenschlager R. A replicon-based bioassay for themeasurement of interferons in patients with chronic hepatitis C.J Virol Methods 2003;110:201–209.
3. Wechsler-Reya R, Sakamuro D, Zhang J, Duhadaway J, Prender-gast GC. Structural analysis of the human BIN1 gene. Evidencefor tissue-specific transcriptional regulation and alternate RNAsplicing. J Biol Chem 1997;272:31453–1458.
4. Elliott K, Ge K, Du W, Prendergast GC. The c-Myc-interactingadaptor protein Bin1 activates a caspase-independent cell deathprogram. Oncogene 2000;19:4669–4684.
5. Elliott K, Sakamuro D, Basu A, Du W, Wunner W, Staller P,Gaubatz S, Zhang H, Prochownik E, Eilers M, Prendergast GC.Bin1 functionally interacts with Myc and inhibits cell proliferationvia multiple mechanisms. Oncogene 1999;18:3564–3573.
6. Livezey KW, Negorev D, Simon D. Investigation of the expressionof Bin1, a putative suppressor, in human hepatoma cells. CancerGenet Cytogenet 2000;116:35–39.
7. Blight KJ, McKeating JA, Rice CM. Highly permissive cell lines forsubgenomic and genomic hepatitis C virus RNA replication. J Virol2002;76:13001–13014.
8. Lohmann V, Hoffmann S, Herian U, Penin F, Bartenschlager R.Viral and cellular determinants of hepatitis C virus RNA replica-
tion in cell culture. J Virol 2003;77:3007–3019.9. Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z,Murthy K, Habermann A, Krausslich HG, Mizokami M, Barten-schlager R, Liang TJ. Production of infectious hepatitis C virus intissue culture from a cloned viral genome. Nat Med 2005;11:791–796.
0. Zech B, Kurtenbach A, Krieger N, Strand D, Blencke S, MorbitzerM, Salassidis K, Cotten M, Wissing J, Obert S, Bartenschlager R,Herget T, Daub H. Identification and characterization of am-phiphysin II as a novel cellular interaction partner of the hepatitisC virus NS5A protein. J Gen Virol 2003;84:555–560.
1. Macdonald A, Crowder K, Street A, McCormick C, Harris M. Thehepatitis C virus NS5A protein binds to members of the Src familyof tyrosine kinases and regulates kinase activity. J Gen Virol2004;85:721–729.
2. Sparks AB, Rider JE, Hoffman NG, Fowlkes DM, Quillam LA, KayBK. Distinct ligand preferences of Src homology 3 domains fromSrc, Yes, Abl, Cortactin, p53bp2, PLCgamma, Crk, and Grb2.Proc Natl Acad Sci U S A 1996;93:1540–1544.
3. Zhang B, Zelhof AC. Amphiphysins: raising the BAR for synapticvesicle recycling and membrane dynamics. Bin-Amphiphysin-Rvsp. Traffic 2002;3:452–460.
4. Ramjaun AR, McPherson PS. Multiple amphiphysin II splice variantsdisplay differential clathrin binding: identification of two distinctclathrin-binding sites. J Neurochem 1998;70:2369–2376.
5. DuHadaway JB, Lynch FJ, Brisbay S, Bueso-Ramos C, Troncoso P,McDonnell T, Prendergast GC. Immunohistochemical analysis ofBin1/Amphiphysin II in human tissues: diverse sites of nuclearexpression and losses in prostate cancer. J Cell Biochem 2003;88:635–642.
6. Ge K, Duhadaway J, Sakamuro D, Wechsler-Reya R, Reynolds C,Prendergast GC. Losses of the tumor suppressor BIN1 in breastcarcinoma are frequent and reflect deficits in programmed celldeath capacity. Int J Cancer 2000;85:376–383.
7. Ge K, DuHadaway J, Du W, Herlyn M, Rodeck U, Prendergast GC.Mechanism for elimination of a tumor suppressor: aberrant splic-ing of a brain-specific exon causes loss of function of Bin1 inmelanoma. Proc Natl Acad Sci U S A 1999;96:9689–9694.
8. Williams GT. Programmed cell death: apoptosis and oncogene-sis. Cell 1991;65:1097–1098.
9. DuHadaway JB, Sakamuro D, Ewert DL, Prendergast GC. Bin1mediates apoptosis by c-Myc in transformed primary cells. Can-cer Res 2001;61:3151–3156.
0. Tschopp J, Thome M, Hofmann K, Meinl E. The fight of virusesagainst apoptosis. Curr Opin Genet Dev 1998;8:82–87.
1. Majumder M, Ghosh AK, Steele R, Zhou XY, Phillips NJ, Ray R,Ray RB. Hepatitis C virus NS5A protein impairs TNF-mediatedhepatic apoptosis, but not by an anti-FAS antibody, in transgenicmice. Virology 2002;294:94–105.
2. Gale M Jr, Kwieciszewski B, Dossett M, Nakao H, Katze MG.Antiapoptotic and oncogenic potentials of hepatitis C virus arelinked to interferon resistance by viral repression of the PKRprotein kinase. J Virol 1999;73:6506–6516.
3. Gold ES, Morrissette NS, Underhill DM, Guo J, Bassetti M, AderemA. Amphiphysin IIm, a novel amphiphysin II isoform, is required formacrophage phagocytosis. Immunity 2000;12:285–292.
4. Bukh J, Pietschmann T, Lohmann V, Krieger N, Faulk K, Engle RE,Govindarajan S, Shapiro M, St Claire M, Bartenschlager R. Mu-tations that permit efficient replication of hepatitis C virus RNA inHuh-7 cells prevent productive replication in chimpanzees. ProcNatl Acad Sci U S A 2002;99:14416–14421.
5. Thorgeirsson SS, Grisham JW. Molecular pathogenesis of humanhepatocellular carcinoma. Nat Genet 2002;31:339–346.
6. Shachaf CM, Kopelman AM, Arvanitis C, Karlsson A, Beer S,Mandl S, Bachmann MH, Borowsky AD, Ruebner B, Cardiff RD,Yang Q, Bishop JM, Contag CH, Felsher DW. MYC inactivationuncovers pluripotent differentiation and tumour dormancy in hep-
atocellular cancer. Nature 2004;431:1112–1117.
4
9J
N
fBtCsB
March 2006 HCV NS5A BINDS TO BIN1 AND PREVENTS CELL DEATH 809
7. Majumder M, Steele R, Ghosh AK, Zhou XY, Thornburg L, Ray R,Phillips NJ, Ray RB. Expression of hepatitis C virus non-structural5A protein in the liver of transgenic mice. FEBS Lett 2003;555:528–532.
Received April 12, 2005. Accepted September 20, 2005.Address requests for reprints to: T. Jake Liang, MD, Building 10-
B16, 10 Center Drive, Bethesda, Maryland 20892-1800. e-mail:
[email protected]; fax: (301) 402-0491. DSupported in part by the Intramural Research Program of the NIH,ational Institute of Diabetes and Digestive and Kidney Diseases.We thankfully acknowledge the reagents, constructs, and cell lines
rom Drs Marc Vidal, Dai Sakamuro, Charles Rice, Zhi Hong, Ralfartenschlager, and George Prendergast. We thank the members of
he Liver Diseases Branch for helpful discussions, the staff in the NIHlinical Center Department of Transfusion Medicine for virologic anderologic testing, Dr Krishna Murthy of Southwest Foundation foriomedical Research for performing the chimpanzee experiment, and
r Boris Baibakov of LCDB/NIDDK for confocal microscopy.