site-directed mutagenesis and deletion of three phosphorylation sites of calsequestrin of skeletal...
TRANSCRIPT

HIH
ttrbbdtrHecarre
Experimental Cell Research 260, 40–49 (2000)doi:10.1006/excr.2000.4989, available online at http://www.idealibrary.com on
Site-Directed Mutagenesis and Deletion of Three Phosphorylation Sitesof Calsequestrin of Skeletal Muscle Sarcoplasmic Reticulum
Effects on Intracellular Targeting
Alessandra Nori, Sandra Furlan, Fioroula Patiri, Marcello Cantini, and Pompeo Volpe1
Centro di Studio per la Biologia e la Fisiopatologia Muscolare del CNR, Dipartimento di Scienze Biomediche Sperimentali,
Universita di Padova, viale G. Colombo 3, 35121 Padova, Italyati
Calsequestrin (CS) is segregated to the junctionalsarcoplasmic reticulum (jSR) of skeletal muscle fibersand is responsible for intraluminal Ca21 binding. Achimeric CS-HA1, obtained by adding the nine-amino-acid viral epitope hemagglutinin (HA1) to the carboxy-terminal of CS and shown to be correctly segregated toskeletal muscle jSR in vivo (A. Nori, K. A. Nadalini, A.Martini, R. Rizzuto, A. Villa, and P. Volpe, 1997, Am. J.Physiol. 272, C1420–C1428), is mutagenized in order toidentify domains of CS involved in targeting. Since aputative targeting mechanism of CS implies phospho-rylation-dependent steps in the endoplasmic reticu-lum (ER) and/or Golgi complex, five CS-HA1 mutantsdisrupting the three phosphorylation sites of CS(Thr189, Thr229, and Thr353) were engineered by eithersite-directed mutagenesis or deletion: CS-HA1DP1(Thr189 3 Ile); CS-HA1DP2 (Thr229 3 Asn); CS-
A1DP1,2; in which Thr189 and Thr229 were changed tole and Asn, respectively; and CS-HA1D14COOH and CS-A1D49 COOH, in which 14 residues (Glu354-Asp367) and 49
residues (Asp319-Asp367), respectively, were deleted athe carboxy-terminal. Mutant cDNAs were transientlyransfected in either HeLa cells, cultured myoblasts ofat skeletal muscle, or regenerating soleus muscle fi-ers of adult rats. Each CS-HA1 mutant was identifiedy Western blot as a single polypeptide of the pre-icted molecular weight. The intracellular localiza-ion of CS-HA1 mutants was studied by immunofluo-escence using specific antibodies against either CS orA1. CS-HA1 mutants colocalized with ER markers,
.g., calreticulin, and partially overlapped with Golgiomplex markers, e.g., a-mannosidase II, in HeLa cellsnd myotubes. CS-HA1 mutants were expressed andetained in ER and ER/SR of HeLa cells and myotubes,espectively, and correctly segregated to jSR of regen-rating soleus muscle fibers. Thus, the targeting mech-
1 To whom correspondence and reprint requests should be ad-dressed at Dipartimento di Scienze Biomediche Sperimentali, Uni-versita degli Studi di Padova, viale G. Colombo 3, 35121 Padova,
Italy. Fax: 011-39-49-8276040. E-mail: [email protected].400014-4827/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
nism of CS in vivo is not affected by phosphoryla-ion(s); i.e., sorting and segregation of CS appear to bendependent of posttranslational phosphorylation(s).
© 2000 Academic Press
Key Words: skeletal muscle; sarcoplasmic reticulum;calsequestrin; site-directed mutagenesis; proteintargeting.
INTRODUCTION
The SR2 of skeletal muscle, a subcompartment of theER, is composed of two continuous and specializedmembrane compartments, devoted to intracellularCa21 homeostasis, the nonjunctional SR, enriched inCa21-pump molecules, and the jSR, enriched in Ca21
release channels (also known as RYRs) and CS [1–3].The molecular differentiation of SR appears to occurfrom an ER-derived reticular network, include at anearly stage the concentration of CS within membrane-bound structures, and progressively evolve into theestablishment of triads between TC and TT [1–3].
CS the acidic, low-affinity (K d ;1 mM), high-capac-ity (40–50 mol/mol), intralumenal Ca21-binding pro-tein [4, 5] segregates to the jSR lumen [6, 7, and Refs.therein], and plays a key role in the storage of Ca21
between uptake and release cycles. Moreover, bindingand unbinding of Ca21 to CS [8 and Refs. therein] aswell as changes of the phosphorylation state of CS [9,10] influence, directly and indirectly, the activity of theRYR-Ca21 release channel. CS has long been known tobe phosphorylated, in vitro at least [11].
2 The abbreviations used are SR, sarcoplasmic reticulum; jSR,junctional SR; TT, transverse tubules; RYR, ryanodine receptor; CS,calsequestrin; ER, endoplasmic reticulum; TC, terminal cisternae;TD, triadin; JC, junctin; PDI, protein disulfide isomerase; BiP, im-munoglobulin binding protein; FCS, fetal calf serum; HS, horse se-rum; DMEM, Dulbecco’s modified Eagle’s medium; PKA, proteinkinase A; CK II, casein kinase II; HA1, hemagglutinin 1; CR, calre-ticulin; PAGE, polyacrylamide gel electrophoresis; PDI, protein di-
sulfide isomerase.
“ccb
iw
41CALSEQUESTRIN PHOSPHORYLATION AND TARGETING TO SR
CS lacks the carboxy-terminal tetrapeptide KDEL[4] that ensures luminal retention, without segrega-tion, to several ER proteins (BiP, PDI, CR) throughcontinuous recycling from pre-Golgi and Golgi com-partments. A stage in the Golgi complex has beendescribed for CS of avian skeletal muscle [12], andmammalian skeletal muscle CS contains a complexcarbohydrate side-chain that would require processingby the Golgi complex [4, 6, 13]. However, no directevidence exists concerning the role of ER- and Golgi-dependent processes, e.g., phosphorylation(s), in sort-ing and segregation of CS.
CS segregation to the jSR can be accounted for byinteractions between (a) integral proteins of jSR able tobind CS with their luminal domains, e.g., TD, JC, RYR,[14–19 and Refs. therein] and (b) specific recognitionsites on CS. It is not known, however, which and wheresuch CS domains may be and whether posttransla-tional modifications of CS, not only glycosylation(s) butalso phosphorylation(s), determine and/or influence CStargeting. Interestingly, Cala and Jones [20] specu-lated that phosphorylation of cardiac CS could occur inthe Golgi complex and “affect how the protein is pack-aged or targeted to the SR.”
We have developed a chimeric, epitope-tagged cDNA,CS-HA1 cDNA whose expression allows studies on CStargeting mechanisms (i.e., sorting, routing, retention,multimerization, and segregation) and have shownthat CS-HA1 segregates to the jSR of regeneratingskeletal muscle fibers of adult rats, following in vivotransfection of CS-HA1 cDNA [21]. CS-HA1 cDNA,thus, is a powerful tool for the identification of specifictargeting sequences of CS by either site-directed mu-tagenesis or deletion.
In order to asses the role of posttranslational modi-fications of CS, we engineered one mutant in which theunique N-glycosylation site of CS (Asn316) was changedto Ile (Nori et al., submitted for publication)3 and fiveCS-HA1 mutants disrupting the three phosphorylationsites of CS (Thr189, Thr229 in Ref. 4, and Thr353 in Ref.20): CS-HA1DP1, in which Thr189 was changed to Ile;CS-HA1DP2, in which Thr229 was changed to Asn; CS-HA1DP1,2, in which both Thr189 and Thr229 werechanged to Ile and Asn, by site-directed mutagenesis;CS-HA1D14COOH, in which the carboxy-terminal of 14amino acid residues (Glu354-Asp367) was deleted; andCS-HA1D49COOH, in which 49 residues at the carboxy-terminal (Asp319-Asp367) were deleted. The five CS-HA1phosphorylation” mutants were transfected in HeLaells, rat myotubes, and rat regenerating skeletal mus-les, and their subcellular localization was monitoredy immunofluorescence.The present results show that phosphorylation of CS
3 A. Nori, G. Valle, M. L. Massimino, and P. Volpe, 2000, submitted
for publication.is not involved in sorting, retention, multimerization,and segregation of skeletal muscle CS to the jSR, i.e.,CS targeting proceeds despite lack of PKA- or CK II-dependent phosphorylation. The results are also inter-preted in the framework of knowledge derived from thecrystal structure of CS [22].
MATERIALS AND METHODS
Generation of CS-HA1DP1, CS-HA1DP2, CS-HA1DP1,2, CS-HA1D14COOH, and CS-HA1D49COOH cDNAs. The cDNA correspond-ng to rabbit skeletal muscle CS was used [4]. The CS-HA1 cDNAas generated as previously described [21].The CS-HA1D14COOH cDNA was generated as previously described
[23]. Modification of the 39 end with deletion of 42 bp and addition of27 bases coding for nine amino acids of HA1 was performed by PCR.The Glu354-Asp367 domain was deleted in CS-HA1D14COOH.
The CS-HA1D49COOH cDNA was developed using an identical clon-ing strategy with the same forward primer and a different reverseprimer, as previously described [23]. The final construct was devoidof 147 bp at the 39 end with the addition of 27 bases coding for HA1.The Asp319-Asp367 domain was deleted in CS-HA1D49COOH.
The CS-HA1DP1, CS-HA1DP2, and CS-HA1DP1,2 cDNAs weregenerated with the QuikChange site-directed mutagenesis kit(Stratagene) according to manufacturer’s specifications. The follow-ing mutagenic (bold letters) primers were used: DP1 sense primer, 59-GCGAAGAAGCTGATCCTGCAGCTGAACGAGATCG-39, antisenseprimer,59-CGA TCTCGTTCAGCTGCAGGATCAGCTTCTTCG-39; andDP2 sense primer, 59-GGAGCACAGGAGGGTTAACCTGAGGAAGC-TGAAGCC-39, antisense primer, 59-GGCTTCAGCTTCCTCAGGTTA-ACCCTCCTGTGCTCC-39. To generate CS-HA1DP1 and CS-HA1DP2the mutagenic primers were annealed to pBSK 1 CS-HA1 as a tem-plate; CS-HA1DP1,2 was obtained by a second round of mutagenesisusing CS-HA1DP1 as a template [24].
For expression in eukaryotic systems, all chimeric cDNAs wereisolated and inserted into NotI–XhoI sites of the expression vectorpcDNA3 (InVitrogen) downstream of the CMV promoter. Orientationand correct sequence of CS-HA1 mutants were checked by restrictionassays, and sequence of the synthetic region was obtained by thedideoxy chain termination method [25], using modified T7 DNApolymerase.
Cell cultures. HeLa cells were grown in DMEM containing 2 mMglutamine and 10% FCS. Primary myoblasts were isolated fromhindlimb skeletal muscles of 0- to 3-day-old rats. After severalwashes of the isolated muscles in 125 mM PBS, pH 7.4, and three20-min cycles of trypsinization (2.5% trypsin in PBS) at 37°C, super-natants were collected and trypsin inhibited by addition of 2% HS.Cells were then collected by centrifugation and preplated in 9-cm-diameter plates for 1 h at 37°C. The myoblast-enriched supernatantwas centrifuged and cells were finally resuspended in DMEM sup-plemented with 20% FCS, 20 mM glucose, counted, and plated.Differentiation to myotubes was obtained by changing the medium toDMEM with 10–20% HS and subsequently DMEM with 2% HS.
Bupivacaine-induced necrosis and regeneration of adult rat skele-tal muscle. Male adult Wistar rats (;250 g body wt) were anesthe-tized with ketamine (1.5 mg/100 g body wt). Soleus muscles wereexposed and injected with 0.5 ml of 0.5% bupivacaine, as described[21, 23]. In agreement with previous reports [26], bupivacaine in-duced almost complete necrosis of the whole soleus by day 3; regen-eration started by day 3 and was completed by day 10. Rat weresacrificed at day 10 or day 15. Muscles were removed and either fixedfor immunocytochemistry or frozen in liquid nitrogen for biochemicalanalysis.
Generation of transient transfectants. Twenty-four hours before
transfection, either HeLa cells or primary myoblasts were seeded
oawc
ipfm[ptc
p
p
t3d
42 NORI ET AL.
onto 25-mm-diameter wells of a 24-well Corning plate, containing a13-mm-diameter round coverslip with a cell density suitable to ob-tain 50% confluence at the moment of transfection. Plasmid DNAs orthe control pcDNA3 vector (4 mg/well) were transfected by the cal-cium phosphate precipitation method, as previously described [21,23]. Forty-eight hours after transfection, cells were fixed for immu-nofluorescence; incubation of myoblasts was prolonged and the me-dium was changed for differentiation.
Adult rat soleus muscles were exposed, 3 days after bupivacaineinjection under ketamine anesthesia, and injected with about 100 mgf plasmid DNA in 20% sucrose. Rats were sacrificed 7–12 days later,nd both transfected and mocked-transfected, contralateral musclesere excised and processed for either immunocytochemistry or bio-
hemical analysis.Immunofluorescence. Cells were fixed in 4% paraformaldehyde,
n PBS, for 15 min and permeabilized with 0.3% Triton X-100, 20 mMhosphate buffer, pH 7.4, 450 mM NaCl, 15% goat serum (buffer A)or 30 min. Longitudinal 6-mm sections were obtained from soleus
uscles, as described [1, 21, 23]. Incubation with polyclonal anti-CS1, 3] and monoclonal anti-CS (Affinity Bioreagents), monoclonal andolyclonal anti-HA1 antibodies (BabCo and Santa Cruz, respec-ively), polyclonal anti-CR antibodies (Affinity Bioreagents), poly-lonal antibodies for BiP (Stress Gene), or polyclonal antibodies for
a-mannosidase II [27] was performed at room temperature for 90min, in buffer A. After 60 min of washing, cells and sections wereincubated for 30 min with either rhodamine isothiocyanate- or fluo-rescein-conjugated anti-mouse or anti-rabbit antibodies (DAKO). Im-ages were obtained with a Leica DMRB microscope.
Preparation of homogenates from HeLa cells and rat soleus muscleand of SR vesicles from rabbit skeletal muscle. HeLa cells werecultured as described above, transiently transfected with plasmidDNAs for 2 days, harvested in PBS, rinsed, and lysed in 1 ml of 150mM NaCl, 15 mM MgCl2, 1 mM EGTA, 1 mM PMSF, 50 mM Hepes,
H 7.5, 10% glycerol, 1% Triton X-100, 2% SDS, 20% b-mercapto-ethanol (Buffer B), for 30 min at 4°C under shaking. Homogenates oftransfected, regenerating soleus muscle of the rat were obtained asfollows: frozen tissues were triturated in a mortar, resuspended in 10vol of Buffer B, and centrifuged at 900g in order to remove debris.Homogenates were kept at 220°C until use. Protein concentrationwas determined according to Lowry et al. [28].
Purified SR vesicles, referable to CS-enriched TC, were preparedfrom rabbit predominantly fast-twitch skeletal muscles, as described[1].
SDS–PAGE and Western blot. SDS–polyacrylamide linear gradi-ent (5–15%) gels, transferred to nitrocellulose sheets and immuno-blot, with either anti-CS or anti-HA1 antibodies, were carried outessentially as previously described [1, 29].
Materials. DNA modification and restriction enzymes were fromBoehringer Mannheim GmbH and Promega. All other chemicalswere from Sigma Chemical Co.
RESULTS
Strategy and Construction of CS-HA1 Mutants
Amino acid sequences around the three phosphory-lation sites of skeletal muscle CS indicate that Thr189
and Thr229 are substrates for PKA [4], whereas Thr353 issubstrate for CK II [20]. Site-directed mutagenesis wasimplemented to change Thr189 and Thr229 to Ile andAsn, respectively, whereas deletion mutants were de-veloped to either disrupt or remove the CK II phos-phorylation site on Thr353. Figure 1 schematically de-
icts the basic features of the five CS-HA1 mutants.
In order to obtain CS-HA1DP1 cDNA, a mutant chi-meric CS cDNA encoding for a CS immunologicallydistinguishable from endogenous CS and lacking one ofthe PKA phosphorylation sites (Thr189), the coding re-gion of rabbit skeletal muscle CS cDNA was modifiedby: (a) substitution of a single nucleotide (C567) so thatthe mutated triplet codes for Ile instead of Thr and (b)addition of a 27-bp fragment coding for nine aminoacids of the influenza virus HA1. A similar approachwas used for construction of CS-HA1DP2 cDNA andCS-HA1DP1,2 cDNA, as detailed under Materials andMethods. The strategy was devised so that conserva-tive amino acid substitutions preserved the configura-tion of the involved microdomains, as implied by Flie-gel et al. [4] and shown by the crystal structure ofWang et al. [22]. The tertiary structure of CS-HA1DP1,CS-HA1DP2, and CS-HA1DP1,2 was deemed to be onlyslightly modified in comparison to that of wild-type CS[22], based on computer simulations of protein model-ing provided by SWISS-MODEL protein modelingserver [30] and viewed with RasMol 2.6.
A deletion strategy was implemented based onknowledge about the minimal structural requirementfor substrate phosphorylation by CK II, i.e., a gluta-mate or aspartate at the n 1 3 position, where n is theposition of the phosphorylated serine or threonine [20].Since the CK II phosphorylation site of CS (Thr353) isfollowed by seven consecutive acidic amino acids, CS-HA1D14COOH should not be phosphorylated because thestructural requirements are lacking CS-HA1D49COOH
because the site itself is deleted.The five constructs were, thus, amenable for trans-
fection and expression of mutant chimeric CSs, immu-nologically distinguishable from endogenous CS, andsuitable to test the role of each phosphorylation site in
FIG. 1. Scheme of CS-HA1 “phosphorylation” mutants. From topto bottom, A, wild type CS; B, CS-HA1; C, CS-HA1DP1; D, CS-HA1DP2; E, CS-HA1DP1,2; F, CS-HA1D14COOH; and G, CS-HA1D49COOH. Phoshorylation sites on Thr are indicated by the posi-ion of relative amino acid residue in the CS sequence, 189, 229, and53, respectively. Cloning procedures for each CS-HA1 mutant areetailed under Materials and Methods.
the targeting mechanism of CS.

H
43CALSEQUESTRIN PHOSPHORYLATION AND TARGETING TO SR
Expression of CS-HA1 Mutants in TransientlyTransfected, Double-Labeled HeLa Cells:Recognition by Anti-CS and Anti-HA1 Antibodies
After transfection of HeLa cells, expression of CS-HA1 mutants was studied in immunofluorescenceexperiments with either anti-CS antibodies (Figs.2A–2D) or anti-HA1 antibodies (Figs. 2E–2H in thesecond row). About 20 –30% of transfected cells werestrongly CS positive and no differences were de-tected comparing the reactivity patterns with thetwo antibodies, as indicated by relative merge im-ages (Figs. 2I–2N).
The epifluorescence pattern obtained with both an-tibodies demonstrated that CS-HA1 mutants were re-tained to the endomembrane network of HeLa cells anddid not have a cytoplasmic distribution [21]. In con-trast, HeLa cells transfected with the empty pcDNA3vector (mocked-transfected cells) were CS negative, asexpected from the lack of expression of endogenous CSin HeLa cells, nor did they immunostain with anti-HA1antibodies (not shown).
Distribution of CS-HA1 Mutants and of ER andGolgi Complex Markers in HeLa Cells
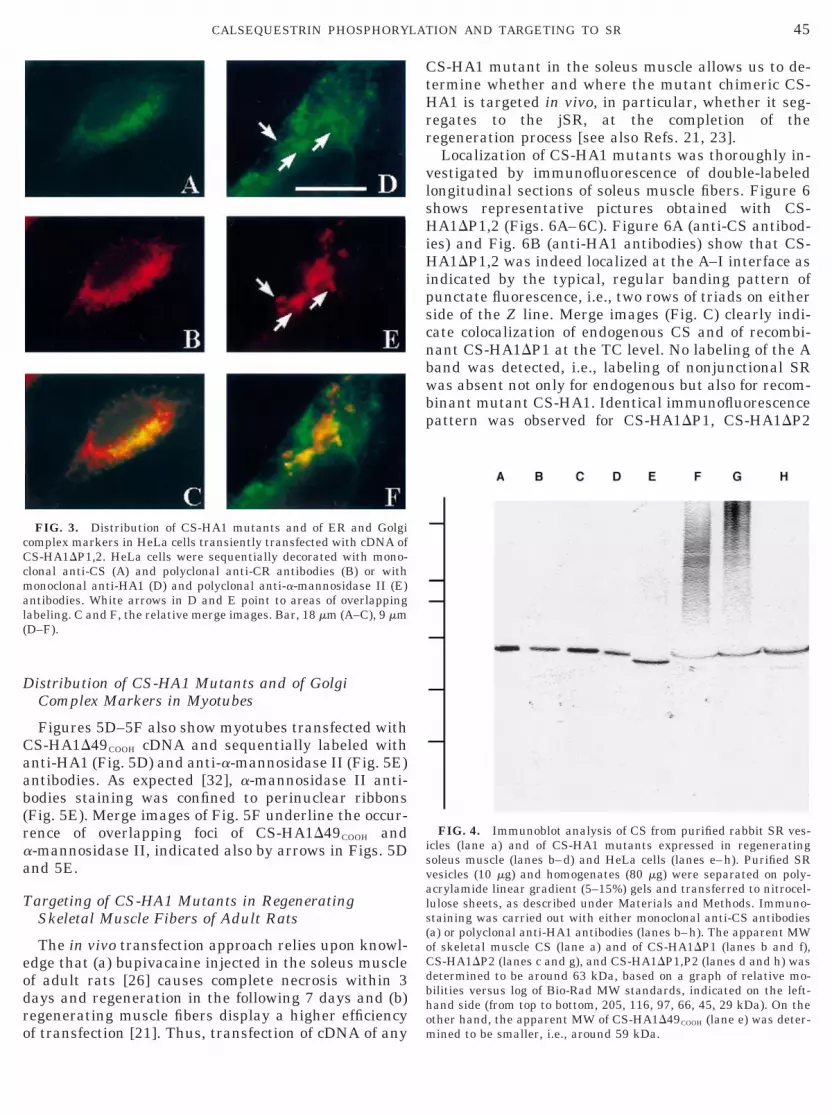
Double labeling with antibodies for CS (Fig. 3A) andfor CR (Fig. 3B), a specific ER marker and molecularchaperone [31], clearly shows that CS-HA1DP1,2 waswithin the ER lumen, as judged from the almost com-plete labeling overlap in the merge image (Fig. 3C);very few green spots, corresponding to areas of sole CSlabeling, are probably referable to medial/late Golgicomplex compartments that exclude CR [31]. Identicalresults were obtained with all other CS-HA1 mutants(not shown).
Since disruption of the phosphorylation site couldhave influenced sorting and routing of recombinantCS-HA1 through the Golgi complex, we investigatedthe subcellullar localization of CS-HA1 mutants rela-tive to that of known Golgi complex markers, e.g.,a-mannosidase II [27, 32]. In Figs. 3D–3F, the immu-nofluorescence pattern of HeLa cells transfected withCS-HA1DP1,P2 cDNA and double labeled with anti-HA1 (Fig. 3D) and anti-a-mannosidase II (Fig. 3E)antibodies, shows areas of overlap between CS anda-mannosidase II (Fig. 3F): the paranuclear domainsreferable to the Golgi complex (Fig. 3E) were also foundto be heavily labeled by CS-HA1DP1,2, as clearly in-ferred by the merge image of Fig. 3F. Thus, it appearsthat CS-HA1DP1,2 cycled through the Golgi complex.Similar pictures were obtained for all other CS-HA1mutants although the overlapping areas were less pro-
nounced (not shown).Detection of CS-HA1 Mutants in HomogenatesDerived from HeLa Cells and Soleus Muscle
The expression of recombinant CS-HA1 mutants wasalso assayed by SDS–PAGE and Western blots (Fig. 4).Homogenates from both regenerating soleus muscles(Fig. 4, lanes b–d) and HeLa cells (Fig. 4, lanes e–h)transfected with cDNAs of CS-HA1 mutants (Fig. 4,lanes b–h) were obtained as described in Materials andMethods. In Western blots, anti-HA1 antibodies recog-nized four CS-HA1 mutants (CS-HA1DP1, CS-
A1DP2, CS-HA1DP1,2) as a single protein band hav-ing an apparent MW of ;63 kDa (Fig. 4, lanes b–d andf–h), comparable to that of wild-type CS from rabbitskeletal muscle SR, detected by anti-CS antibodies(Fig. 4, lane a). As expected, the MW of CS-HA1D49COOH was smaller, around 59 kDa (Fig. 4, lanee).
The results also indicate that the epitope recognizedby anti-HA1 antibodies was within the recombinantprotein and that no proteolytic breakdown productscould be detected, ruling out the possibility that chi-meric proteins, as it may happen [33], undergo accel-erated or altered turnover which, in turn, may bring tomisleading interpretation of immunofluorescence data.
Compartmentalization of CS-HA1 Mutants in SR\ERof Rat Myotubes in Primary Culture
The consequences of disrupting the phosphorylationsite(s) of CS were further studied upon transfection ofCS-HA1 mutants in myoblasts. Myoblasts from 0- to3-day-old rat hindlimb skeletal muscles were culturedin vitro, induced to differentiate into myotubes (see fordetails Ref. 21 and Materials and Methods), and har-vested 4 days after induction. “Transfected myotubes,”thus, indicate myotubes obtained from transfectedmyoblasts by this procedure.
Expression of CS-HA1 mutants was detected inabout 10–20% of rat myotubes by anti-HA1 antibodies,whereas all myotubes are CS positive, as indicated byreactivity with anti-CS antibodies (cf. Ref. 23). Figure 5displays representative pictures for double-labeledmyotubes expressing CS-HA1D49COOH. In both Fig. 5Aand Fig. 5B (with anti-CS and anti-HA1 antibodies,respectively), fluorescent strands running parallel tothe longitudinal axis of the myotube and several dis-crete CS foci showed the prevalent longitudinal ar-rangement of ER/SR along with areas of CS clustering.Merge images (Fig. 5C) show complete overlap betweenendogenous CS and CS-HA1D49COOH; i.e., wild-type andrecombinant CS colocalize. The overall picture ob-tained with all other CS-HA1 mutants (not shown),including CS-HA1D14COOH [see Ref. 23], was virtually
similar to that reported in Figs. 5A–5C.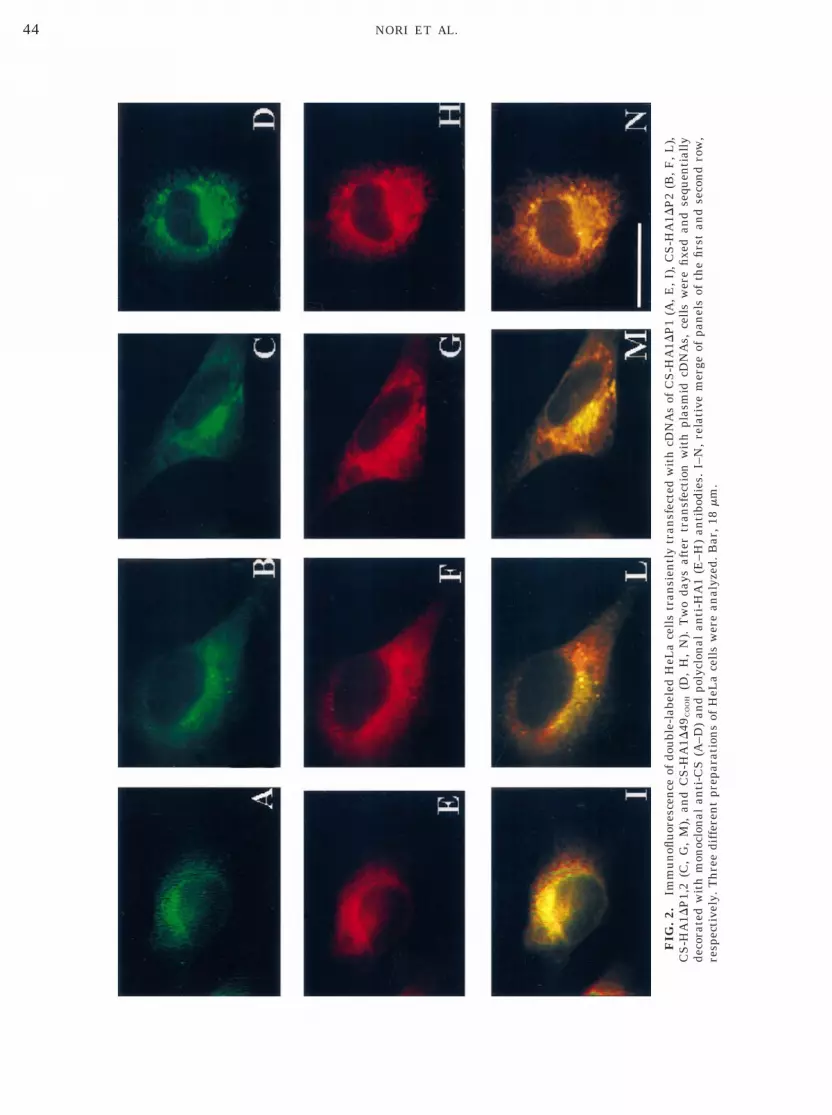
FIG
.2.
Imm
un
oflu
ores
cen
ceof
dou
ble-
labe
led
HeL
ace
lls
tran
sien
tly
tran
sfec
ted
wit
hcD
NA
sof
CS
-HA
1DP
1(A
,E
,I)
,C
S-H
A1D
P2
(B,
F,
L),
CS
-HA
1DP
1,2
(C,
G,
M),
and
CS
-HA
1D49
CO
OH
(D,
H,
N).
Tw
oda
ysaf
ter
tran
sfec
tion
wit
hpl
asm
idcD
NA
s,ce
lls
wer
efi
xed
and
sequ
enti
ally
deco
rate
dw
ith
mon
oclo
nal
anti
-CS
(A–D
)an
dpo
lycl
onal
anti
-HA
1(E
–H)
anti
bodi
es.
I–N
,re
lati
vem
erge
ofpa
nel
sof
the
firs
tan
dse
con
dro
w,
resp
ecti
vely
.T
hre
edi
ffer
ent
prep
arat
ion
sof
HeL
ace
lls
wer
ean
alyz
ed.
Bar
,18
mm
.
44 NORI ET AL.
eodro
CtHrr
vlsH
(
45CALSEQUESTRIN PHOSPHORYLATION AND TARGETING TO SR
Distribution of CS-HA1 Mutants and of GolgiComplex Markers in Myotubes
Figures 5D–5F also show myotubes transfected withCS-HA1D49COOH cDNA and sequentially labeled withanti-HA1 (Fig. 5D) and anti-a-mannosidase II (Fig. 5E)antibodies. As expected [32], a-mannosidase II anti-bodies staining was confined to perinuclear ribbons(Fig. 5E). Merge images of Fig. 5F underline the occur-rence of overlapping foci of CS-HA1D49COOH anda-mannosidase II, indicated also by arrows in Figs. 5Dand 5E.
Targeting of CS-HA1 Mutants in RegeneratingSkeletal Muscle Fibers of Adult Rats
The in vivo transfection approach relies upon knowl-dge that (a) bupivacaine injected in the soleus musclef adult rats [26] causes complete necrosis within 3ays and regeneration in the following 7 days and (b)egenerating muscle fibers display a higher efficiency
FIG. 3. Distribution of CS-HA1 mutants and of ER and Golgicomplex markers in HeLa cells transiently transfected with cDNA ofCS-HA1DP1,2. HeLa cells were sequentially decorated with mono-clonal anti-CS (A) and polyclonal anti-CR antibodies (B) or withmonoclonal anti-HA1 (D) and polyclonal anti-a-mannosidase II (E)antibodies. White arrows in D and E point to areas of overlappinglabeling. C and F, the relative merge images. Bar, 18 mm (A–C), 9 mmD–F).
f transfection [21]. Thus, transfection of cDNA of any m
S-HA1 mutant in the soleus muscle allows us to de-ermine whether and where the mutant chimeric CS-A1 is targeted in vivo, in particular, whether it seg-
egates to the jSR, at the completion of theegeneration process [see also Refs. 21, 23].Localization of CS-HA1 mutants was thoroughly in-
estigated by immunofluorescence of double-labeledongitudinal sections of soleus muscle fibers. Figure 6hows representative pictures obtained with CS-A1DP1,2 (Figs. 6A–6C). Figure 6A (anti-CS antibod-
ies) and Fig. 6B (anti-HA1 antibodies) show that CS-HA1DP1,2 was indeed localized at the A–I interface asindicated by the typical, regular banding pattern ofpunctate fluorescence, i.e., two rows of triads on eitherside of the Z line. Merge images (Fig. C) clearly indi-cate colocalization of endogenous CS and of recombi-nant CS-HA1DP1 at the TC level. No labeling of the Aband was detected, i.e., labeling of nonjunctional SRwas absent not only for endogenous but also for recom-binant mutant CS-HA1. Identical immunofluorescencepattern was observed for CS-HA1DP1, CS-HA1DP2
FIG. 4. Immunoblot analysis of CS from purified rabbit SR ves-icles (lane a) and of CS-HA1 mutants expressed in regeneratingsoleus muscle (lanes b–d) and HeLa cells (lanes e–h). Purified SRvesicles (10 mg) and homogenates (80 mg) were separated on poly-acrylamide linear gradient (5–15%) gels and transferred to nitrocel-lulose sheets, as described under Materials and Methods. Immuno-staining was carried out with either monoclonal anti-CS antibodies(a) or polyclonal anti-HA1 antibodies (lanes b–h). The apparent MWof skeletal muscle CS (lane a) and of CS-HA1DP1 (lanes b and f),CS-HA1DP2 (lanes c and g), and CS-HA1DP1,P2 (lanes d and h) wasdetermined to be around 63 kDa, based on a graph of relative mo-bilities versus log of Bio-Rad MW standards, indicated on the left-hand side (from top to bottom, 205, 116, 97, 66, 45, 29 kDa). On theother hand, the apparent MW of CS-HA1D49COOH (lane e) was deter-
ined to be smaller, i.e., around 59 kDa.

46 NORI ET AL.

risr
lsactMl
Haarmri
47CALSEQUESTRIN PHOSPHORYLATION AND TARGETING TO SR
(not shown), CS-HA1D14COOH [23], or CS-HA1D49COOH
[23].Double labeling with antibodies specific for either
HA1 or BiP, a soluble intraluminal protein and chap-erone [31] distributed to the entire SR lumen [1, 3],was carried out to unambiguously discriminate reten-tion to SR from segregation to jSR. Figure 6D is themerge image and shows that CS-HA1DP1,2 segregatedto junctional SR (green areas and yellow-orange dots),whereas BiP (red fluorescence) was also observed atthe A band around the H line, i.e., nonjunctional SRwas labeled (white arrows in Fig. 6D) [cf. also Refs.1, 3].
DISCUSSION
Targeting of CS to the jSR of skeletal muscle in-cludes several putative steps and phases, i.e., sorting,routing, retention, multimerization, and segregation.Any single step and phase, in possibly different subcel-lular compartments, e.g., ER, Golgi complex, and SR,may conceivably be controlled by posttranslationalphosphorylation(s), which may serve as routing sig-nal(s) and/or influence both homologous and heterolo-gous protein–protein interactions within the SR lu-men. The mutant CS-HA1 cDNAs were designed sothat either conservative substitutions or deletions in-volving three specific phosphorylation sites of CS(Thr189, Thr229, and Thr353) abolished posttranslationalphosphorylation(s) and substantially preserved thetertiary structure of relevant CS domains II and III[22].
The present results show that CS-HA1 mutants are:(a) expressed (Fig. 4) and retained to ER compartmentsin HeLa cells (Figs. 2 and 3), (b) sorted and retained toSR of differentiating rat myotubes (Fig. 5), and (c)expressed (Fig. 4) and segregated to jSR of skeletalmuscle fibers (Fig. 6), following in vivo transfection ofecombinant cDNAs. Conclusive data are derived fromn vivo transfection of mutant CS-HA1 cDNAs intoatellite cells of regenerating skeletal muscle of adultats: mutant CS-HA1s are sorted and retained to SR,
FIG. 5. Immunofluorescence of double-labeled rat skeletal musTransfection was carried out with plasmid cDNAs or pcDNA3 (not shanti-CS (A) and polyclonal anti-HA1 (B) or with monoclonal anti-HA1in D and E point to overlapping areas of perinuclear labeling. C anpreparations of myotubes were analyzed. Bar, 6 mm (A–C), 3 mm (D
FIG. 6. Immunofluorescence of double-labeled soleus muscle fiA1DP1,2. All sections were obtained 15 days after bupivacaine treafter transfection. Double-labeled, longitudinal sections were stainenti-HA1 antibodies (B). Note two rows of punctate labeling on eitheespectively. Three different muscle preparations were examined. Ionoclonal anti-HA1 antibodies (revealed by fluorescein-conjugated
hodamine isothiocyanate secondary antibodies). Only the merge ima
s observed at the A band around the H line. Bar, 5 mm.and, more importantly, segregate to TC. Thus, expres-sion of “phosphorylation” mutants during muscle re-generation, i.e., during SR biogenesis, TC develop-ment, and triad formation, indicates that substitution/deletion of any phosphorylation site does not interferewith CS targeting to jSR.
In vivo phosphorylations can occur both in the lumenof ER and in the Golgi complex [34] and could serve asspecific sorting signal(s) for ER export and/or vectorialsignal(s) for routing to Golgi complex and recyclingback to either ER or SR. Recombinant CS-HA1 and allCS-HA1 mutants partially overlap with Golgi complexmarkers in HeLa cells and myotubes (Figs. 3 and 5,respectively), as expected for a protein cycling throughthe Golgi complex. Our present data indicate that CSrouting through the Golgi complex proceeds unabatedwith “phosphorylation” mutants. Thus, phosphoryla-tion(s) is(are) not necessary for CS sorting and do nothamper subsequent targeting steps. These experimen-tal conclusions are discrepant with previous sugges-tions by Cala and Jones [20] who argued that phos-phorylation of cardiac CS could occur in the Golgicomplex and “affect how the protein is packaged ortargeted to the SR”.
We do not know whether CS-HA1 “phosphorylation”mutants are correctly folded following synthesis andtranslocation across ER in any of our experimentalsystems. Proper protein folding, assisted and directedby a variety of soluble and membrane-bound chaper-ones [30, 35], might be hampered in CS-HA1 mutants.Should CS-HA1 mutants be misfolded and thus di-verted to degradative pathways and compartments, wewould have to find some evidence that any CS-HA1mutant aggregated, coprecipitated with ER chaper-ones, and was en route to degradation. In contrast, noarge CS-HA1 mutant containing vacuoles were ob-erved in HeLa cells (Figs. 2 and 3) [cf. also Ref. 33],nd recombinant CS-HA1 mutants, from both HeLaells and soleus muscles, were detected as single pro-eins in the absence of proteolytic fragments (Fig. 4).echanisms for protein folding are so redundant that
ectin-based modes [36] might be replaced for by those
myotubes transiently transfected with cDNA of CS-HA1D49COOH.n), and fixed myotubes were sequentially decorated with monoclonal) and polyclonal anti-a-mannosidase II (E) antibodies. White arrows, the relative merge of A, B, D, and E, respectively. Three different.
s after bupivacaine treatment and transfection with cDNA of CS-ent, i.e., when muscle regeneration is deemed complete, and 12 daysequentially with monoclonal anti-CS antibodies (A) and polyclonalide of Z lines corresponding to TC. C ,the merge images of A and B,nother series of experiments, double labeling was carried out withondary antibodies) and polyclonal anti-BiP antibodies (revealed byis shown in D: the exclusive immunolabeling for BiP (white arrows)
cleow(D
d F–F)bertmd sr sn asecge

btbtmbtitaeC([
1
1
1
48 NORI ET AL.
exerted by BiP and similar chaperonines localized inthe SR lumen [1, 3]. In fact, based on both the primarysequence [4] and the crystal structure [22] of CS, it isentirely possible that folding is ensured by chaperoni-nes of the BiP family through binding to hydrophobicsequences exposed on the surface of incompletelyfolded CS. It is also known that some misfolded pro-teins are retained as aggregates that are too large orinsoluble to be exported from the ER [35]: if such werethe case for CS-HA1 mutants, we could have misinter-preted predegradative aggregation for ER retention, inboth HeLa cells and rat myotubes. Data on soleusmuscle fibers, however, provide evidence for segrega-tion to jSR, because hypothetical predegradative ag-gregates of CS-HA1 mutants would have been divertedand never reached jSR, their final location (Fig. 6).Moreover, separation from the BiP signal in regener-ating soleus fibers (Fig. 6D) unambiguously shows thatrecombinant CS-HA1 mutants are not simply retainedor aggregated, with or without BiP, but are specificallysegregated to jSR.
Segregation of CS to jSR is a process entailing mul-timerization of CS, i.e., homologous protein–proteininteractions, and docking, i.e., heterologous protein–protein interactions. CS-HA1 mutants segregate in jSRof regenerating soleus muscle fibers (Fig. 6), althoughour experimental approach cannot indicate whethermultimerization or docking to jSR are affected by themutagenized CSs. Multimerization of CS, a calcium-dependent phenomenon, entails initially two extensivedimerization contacts that require the juxtaposition ofmany negative groups at each CS monomer surface[22]. Since Thr189, Thr229, and Thr353 are located instretches of domains II and III of CS, regions not di-rectly involved in CS dimerization [22] multimeriza-tion of “phosphorylation” mutants is conceivably guar-anteed.
Since we have also shown that deletion of the uniqueN-glycosylation site on CS (Asn253) does not affect tar-geting to jSR in regenerating soleus muscle fibers (Noriet al., submitted for publication),3 it would appear thatoth homologous and heterologous protein–protein in-eractions involved in CS targeting are not regulatedy co- and/or posttranslational modifications. Segrega-ion may rely mainly on interactions between comple-entary charged amino acid residues on CS and CS-
inding proteins; although the mostly acidic carboxy-erminal of CS, irrespective of length of deletion, is notnvolved in targeting to jSR [23], it is entirely plausiblehat other CS domains are involved, and among themre to be listed acidic stretches located on the surface ofach of the three topological domains [22], relevant forS dimerization, the switch points of domain II and III
around residues 228–229), rich in acidic amino acids22], as well as the NH2-terminus [14, 17, 22].
Since mutagenized, dephosphorylated forms of CS
still segregate to jSR and changes of the phosphoryla-tion state of CS seem to influence, directly and indi-rectly, the activity of the RYR-Ca21 release channel [9,10], it appears that CS is a complex molecule withdistinct structural and functional domains, “more thanonly a luminal Ca21 buffer” [10]. Functional studies on“phosphorylation” mutants of CS may be very valuableto unravel additional mechanistic aspects of CS.
We thank Mr. G. A. Tobaldin for skillful assistance in surgicalprocedures. This work was supported by Telethon, Italy, Grant 669,and by funds from the Ministero dell’Universita e della RicercaScientifica e Tecnologica (1999–2001 Programma di ricerca di rile-vante interesse nazionale on “Biopatologia della fibra muscolarescheletrica”).
REFERENCES
1. Volpe, P., Villa, A., Podini, P., Martini, A., Nori, A., Panzeri,M. C., and Meldolesi, J. (1992). The endoplasmic reticulum-sarcoplasmic reticulum connection: Distribution of endoplasmicreticulum markers in the sarcoplasmic reticulum of skeletalmuscle fibers. Proc. Natl. Acad. Sci. USA 89, 6142–6146.
2. Flucher, B. E. (1992). Structural analysis of muscle develop-ment: Transverse tubules, sarcoplasmic reticulum and thetriad. Dev. Biol. 154, 245–260.
3. Villa, A., Podini, P., Nori, A., Panzeri, C., Martini, A., Meldolesi,J., and Volpe, P. (1993). The endoplasmic reticulum–sarcoplas-mic reticulum connection. II. Postnatal differentiation of thesarcoplasmic reticulum in skeletal muscle fibers. Exp. Cell Res.209, 140–148.
4. Fliegel, L., Ohnishi, M., Carpenter, M. R., Khanna, V. K., Re-ithmeier, R. A. F., and MacLennan, D. H. (1987). Amino acidsequence of rabbit fast-twitch skeletal muscle calsequestrindeduced from cDNA and peptide sequence. Proc. Natl. Acad.Sci. USA 84, 1167–1171.
5. Scott, B. J., Simmermann, H. K. B., Collins, J. H., Nadal-Ginard, B., and Jones, L. R. (1988). Complete amino acid se-quence of canine cardiac calsequestrin deduced by cDNA clon-ing. J. Biol. Chem. 263, 8958–8964.
6. Jorgensen, A. O., Kalnins, V. I., Zubrzycka, E., and MacLennan,D. H. (1977). Assembly of the sarcoplasmic reticulum proteinsin differentiating rat skeletal muscle cell cultures. J. Cell Biol.74, 287–298.
7. Franzini-Armstrong, C., Kenney, L. J., and Varriano-Marston,M. (1987). The structure of calsequestrin in triads of vertebrateskeletal muscle: A deep-etch study. J. Cell Biol. 105, 49–56.
8. Kawasaki, T., and Kasai, M. (1994). Regulation of calcium chan-nel of sarcoplasmic reticulum by calsequestrin. Biochem. Bio-phys. Res. Commun. 199, 1120–1127.
9. Herrmann-Frank, A., and Varsanyi, M. (1993). Enhancement ofCa21 release channel activity by phosphorylation of the skeletalmuscle ryanodine receptor. FEBS Lett. 332, 237–242.
0. Szegedi, C., Sarkozi, S., Herzog, A., Jona, I., and Varsanyi, M.(1999). Calsequestrin: More than ‘only’ a luminal Ca21 bufferinside the sarcoplasmic reticulum. Biochem. J. 337, 19–22.
1. Campbell,K. P., and Shamoo, A. E (1980). Phosphorylation ofheavy sarcoplasmic reticulum vesicles: Identification and char-acterization of three phosphorylated proteins. J. Membr. Biol.56, 241–248.
2. Thomas, K., Navarro, J., Benson, R. J. J., Campbell, K. P.,Rotundo, R. L., and Fine, R. E. (1989). Newly synthesized
calsequestrin, destined for the sarcoplasmic reticulum, is con-
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
RR
49CALSEQUESTRIN PHOSPHORYLATION AND TARGETING TO SR
tained in early/intermediate Golgi-derived clathrin-coated ves-icles. J. Biol. Chem. 264, 3140–3145.
3. Campbell, K. P., MacLennan, D. H., Jorgensen, A. O., andMintzer, M. C. (1983). Purification and characterization ofcalsequestrin from canine cardiac sarcoplasmic reticulum andidentification of the 53,000 dalton glycoprotein. J. Biol. Chem.258, 1197–1204.
4. Guo, W., and Campbell, K. P. (1995). Association of triadin withthe ryanodine receptor and calsequestrin in the lumen of thesarcoplasmic-reticulum. J. Biol. Chem. 270, 9027–9030.
5. Jones, L. R., Zhang, L., Sanborn, K., Jorgensen, A. O., andKelley, J. (1995). Purification, primary structure, and immuno-logical characterization of the 26-kDa calsequestrin bindingprotein (junctin) from cardiac junctional sarcoplasmic reticu-lum. J. Biol. Chem. 270, 30787–30796.
6. Knudson, C. M., Stang, K. J., Moomaw, C. R., Slaughter, C. A.,and Campbell, K. P. (1993). Primary structure and topologicalanalysis of a skeletal muscle-specific junctional sarcoplasmicreticulum glycoprotein (triadin). J. Biol. Chem. 268, 12646–12654.
7. Zhang, L., Kelley, J., Schmeisser, G., Kobayashi, Y. M., andJones, L. R. (1997). Complex formation between junctin, tria-din, calsequestrin, and the ryanodine receptor. J. Biol. Chem.272, 23389–23397
8. Kobayashi, Y. M., Alseikham, B. A., and Jones, L. R. (2000).Localization and characterization of the calsequestrin-bindingdomain of triadin 1: Evidence for a charged b-strand in medi-ating the protein–protein interaction. J. Biol. Chem. 275,17639–17641.
9. Murray, B. E., and Ohlendieck, K. (1998). Complex formationbetwen calsequestrin and the ryanodine receptor in fast- andslow-twitch rabbit skeletal muscle. FEBS Lett. 429, 317–322.
0. Cala, S. E., and Jones, L. R. (1991). Phosphorylation of cardiacand skeletal muscle calsequestrin isoforms by casein kinase. II.Demonstration of a cluster of unique rapidly phosphorylatedsites in cardiac calsequestrin. J. Biol. Chem. 266, 391–398.
1. Nori, A., Nadalini, K. A., Martini, A., Rizzuto, R., Villa, A., andVolpe, P. (1997). Chimeric calsequestrin and its targeting to thejunctional sarcoplasmic reticulum of skeletal muscle. Am. J.Physiol. 272, C1420–C1428
2. Wang S., Trumble, W. R., Liao, H., Wesson, C. R., Dunker,A. K., and Kang, C. H. (1998). Crystal structure of calsequestrinfrom rabbit skeletal muscle sarcoplasmic reticulum. NatureStruct. Biol. 5, 476–483.
3. Nori, A., Gola, E., Tosato, S., Cantini, M., and Volpe, P. (1999).
Targeting of calsequestrin to the sarcoplasmic reticulum follow-ing deletions of its acidic carboxy-terminal. Am. J. Physiol. 277,C974–C981.
4. Kunkel, T. A., Monaco, A. P., Middlesworth, W., Ochs, H. D.,and Latt, S. A. (1985). Specific cloning of DNA fragments absentfrom the DNA of a male patient with an X chromosome deletion.Proc. Natl. Acad. Sci. USA 82, 488–492.
5. Sanger, F., Nicklen, F., and Coulson, A. R. (1977). DNA se-quencing with chain-terminating inhibitors. Proc. Natl. Acad.Sci. USA 74, 5463–5467.
6. Hall-Graggs, E. C. B. (1974). Rapid degeneration and regener-ation of a whole skeletal muscle following treatment with bu-pivacaine (marcaine). Exp. Neurol. 43, 349–358.
7. Moremen, K. W., Touster, O., and Robbins, P. W. (1991). Novelpurification of the catalytic domain of Golgi a-mannosidasi II:Characterization and comparison with the intact enzyme.J. Biol. Chem. 266, 16876–16885.
28. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.(1951). Protein measurement with the Folin phenol reagent.J. Biol. Chem. 193, 9453–9459.
29. Nori, A., Villa, A., Podini, P., Witcher, D. R., and Volpe, P.(1993). Intracellular Ca21 stores of rat cerebellum: Heterogene-ity within and distinction from endoplasmic reticulum. Bio-chem. J. 291, 199–204.
0. Peitsch, M. C. (1996). ProMod and Swiss-Model: Internet-basedtools for automated comparative protein modeling. Biochem.Soc. Trans. 24, 274–279.
1. Krause, K. H., and Michalak, M. (1997). Calreticulin. Cell 88,439–443.
2. Ralston, E. (1993). Changes in architecture of the Golgi complexand other subcellular organelles during myogenesis. J. CellBiol. 120, 399–409.
3. Raichman, M., Panzeri, M. C., Clementi, E., Papazafiri, P.,Eckley, M., Clegg, D. O., Villa, A., and Meldolesi, J. (1995).Differential localization and functional role of calsequestrin ingrowing and differentiated myoblasts. J. Cell Biol. 128, 341–354.
4. Hirschberg, C. B., Robbins, P. W., and Abeijon, C. (1998).Transporters of nucleotide sugars, ATP, and nucleotide sulfatein the endoplasmic reticulum and Golgi apparatus. Annu. Rev.Biochem. 67, 49–69.
5. Helenius, A., Trombetta, S. E., Hebert, D. N., and Simons, J. F.(1997). Calnexin, calreticulin and the folding of glycoproteins.Trends Cell Biol. 7, 193–200.
6. Pelham, H. R. B. (1991). Recycling of proteins between theendoplasmic reticulum and Golgi complex. Curr. Opin. Cell
Biol. 3, 585–591.eceived May 10, 2000evised version received July 3, 2000
The sarcoplasmic reticulum of an ultrafast lobster muscle: First evidence of a tubular configuration
Mutagenesis, biochemical, and biophysical characterization of Mycoplasma arthritidis-derived mitogen