short interfering rna-mediated gene silencing in globodera pallida and meloidogyne incognita...
TRANSCRIPT

International Journal for Parasitology 40 (2010) 91–100
Contents lists available at ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Short interfering RNA-mediated gene silencing in Globodera pallidaand Meloidogyne incognita infective stage juveniles
Johnathan J. Dalzell a, Steven McMaster a, Colin C. Fleming b, Aaron G. Maule a,*
a Parasitology, School of Biological Sciences, Queen’s University Belfast, Belfast, UKb Agri-Food Biosciences Institute (AFBI), Belfast, UK
a r t i c l e i n f o a b s t r a c t
Article history:Received 12 June 2009Received in revised form 21 July 2009Accepted 22 July 2009
Keywords:RNA interference (RNAi)Short interfering RNA (siRNA)Globodera pallidaMeloidogyne incognitaFMRFamide-like peptidesQuantitative-PCRPlant parasitic nematodeThermodynamic manipulation
0020-7519/$36.00 � 2009 Australian Society for Paradoi:10.1016/j.ijpara.2009.07.003
* Corresponding author. Address: School of Biologisity Belfast, Medical Biology Centre, 97 Lisburn RoaIreland, UK. Tel.: +44 (0)28 9097 2059; fax: +44 (0)28
E-mail address: [email protected] (A.G. Maule).
The analysis of gene function through RNA interference (RNAi)-based reverse genetics in plant parasiticnematodes (PPNs) remains inexplicably reliant on the use of long double-stranded RNA (dsRNA) silencingtriggers; a practice inherently disadvantageous due to the introduction of superfluous dsRNA sequence,increasing chances of aberrant or off-target gene silencing through interactions between nascent shortinterfering RNAs (siRNAs) and non-cognate mRNA targets. Recently, we have shown that non-nematode,long dsRNAs have a propensity to elicit profound impacts on the phenotype and migrational abilities ofboth root knot and cyst nematodes. This study presents, to our knowledge for the first time, gene-specificknockdown of FMRFamide-like peptide (flp) transcripts, using discrete 21 bp siRNAs in potato cyst nem-atode Globodera pallida, and root knot nematode Meloidogyne incognita infective (J2) stage juveniles. Bothknockdown at the transcript level through quantitative (q)PCR analysis and functional data derived frommigration assay, indicate that siRNAs targeting certain areas of the FMRFamide-like peptide (FLP) tran-scripts are potent and specific in the silencing of gene function. In addition, we present a method ofmanipulating siRNA activity through the management of strand thermodynamics. Initial evaluation ofstrand thermodynamics as a determinant of RNA-Induced Silencing Complex (RISC) strand selection(inferred from knockdown efficacy) in the siRNAs presented here suggested that the purported influenceof 50 stand stability on guide incorporation may be somewhat promiscuous. However, we have found thaton strategically incorporating base mismatches in the sense strand of a G. pallida-specific siRNA, we couldspecifically increase or decrease the knockdown of its target (specific to the antisense strand), presum-ably through creating more favourable thermodynamic profiles for incorporation of either the sense(non-target-specific) or antisense (target-specific) strand into a cleavage-competent RISC. Whilst the effi-cacy of similar approaches to siRNA modification has been demonstrated in the context of Drosophilawhole-cell lysate preparations and in mammalian cell cultures, it remained to be seen how these sensestrand mismatches may impact on gene silencing in vivo, in relation to different targets and in differentsequence contexts. This work presents the first application of such an approach in a whole organism; ini-tial results show promise.
� 2009 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Plant parasitic nematodes (PPNs) impose a significant economicburden on plant cultivation efforts worldwide through reductionsin yield and quality; current best estimates predict losses acrossall sectors of approximately $125 billion annually (Bird andKaloshian, 2003; Chitwood, 2003). As a result of their obligate sed-entary parasitism in plant root vascular tissue, root knot (Meloido-gyne spp.) and cyst (Heterodera and Globodera spp.) nematodescomprise the most severe antagonists. The nematode feeding site
sitology Inc. Published by Elsevier
cal Sciences, Queen’s Univer-d, Belfast BT9 7BL, Northern
9097 5877.
within the root swells in response to parasitism factors secretedfrom the nematode pharyngeal glands and impedes normal vascu-lar activity (Gheysen and Fenoll, 2002; Bird, 2004; Caillaud et al.,2008), which results in systemic nutritional deficiency and reducedvigour of the plant host. Conventionally, an integrated approach toPPN management has relied heavily on the utility of nematicides,which although increasing the costs associated with plant cultiva-tion significantly, remain the only viable option for adequate con-trol. As environmental concerns rise over the systemic effects ofsustained nematicide use, withdrawal has left a significant short-coming in our ability to manage this problem and highlights theneed for novel and robust control methods.
The RNA interference (RNAi) pathway elicits sequence-specificsilencing of targeted mRNA transcripts through the introductionof homologous double-stranded (ds)RNA. Therefore, RNAi
Ltd. All rights reserved.

92 J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100
represents an attractive prospect as a reverse genetics tool, espe-cially in dealing with parasitic species difficult to culture andmanipulate at a genetic level. RNAi offers the ability to disruptthe expression of individual genes and consequently the expositionof novel molecular traits which could be targeted by therapeutics.Moreover, the application of RNAi in planta towards the control ofPPNs may prove a useful counterweight as part of an integrated ap-proach to management (Huang et al., 2006; Steeves et al., 2006;Yadav et al., 2006; Fairbairn et al., 2007; Sindhu et al., 2009),through stable transformation of the plant and production of nem-atode-specific dsRNAs in the saleable crop or trap crops.
The efficacy of gene silencing has been demonstrated previouslyin both Globodera pallida (Urwin et al., 2002; Kimber et al., 2007)and Meloidogyne incognita (Bakhetia et al., 2005; Rosso et al.,2005; Shingles et al., 2006) using long dsRNA silencing triggers.As yet there is no published account of short interfering (si)RNAutility for the study of gene function in these worms, or indeedany PPN. siRNA-mediated gene silencing has been attempted in afew animal parasitic nematode species, to varying effect. Issaet al. (2005) report that siRNAs targeted to a ubiquitin resultedin a significant and specific reduction in transcript abundance,and a pronounced developmental lag in Trichostrongylus colubrifor-mis L1s; these effects were considerably more profound on electro-poration (Issa et al., 2005). By contrast, the intestinal nematodeHeligmosomoides polygyrus did not present with a specific pheno-type on electroporation of tropomyosin-specific siRNAs, nor wastropomyosin mRNA abundance reduced.
Unfortunately, specific knockdown as a principle of RNAi beliesthe process in practice. Many now ascribe a cautionary approach tofunctional genetics conducted thus, due to the apparent relaxationof exact homology requirements for knockdown in many biologicalsystems, including parasitic nematodes (Jackson et al., 2006; Lend-ner et al., 2008). Furthermore, our recent demonstration of the im-pact of non-nematode derived, long dsRNAs on root knot and cystnematode phenotypes, calls into question our ability to rely onselective transcript knockdown by long dsRNAs as being theunequivocal trigger for observed phenotypic impacts on PPNs(Dalzell et al., 2009). This undermines the reliability of gene func-tion determinations in PPNs which are based on long dsRNAs.When considering these elements of uncertainty together, it seemslogical to minimise both the size and concentration of the dsRNAemployed in RNAi experiments.
To this end, we evaluated the performance of discrete 21 bpsiRNAs, spanning FMRFamide-like peptide (flp) transcripts; fivesiRNAs against Gp-flp-12 in G. pallida, and five siRNAs againstMi-flp-18 in M. incognita. In addition, we tested the efficacy of ran-domized and non-specific siRNAs as negative controls. FLPs com-prise the largest known family of neuropeptides in nematodes,and play crucial and diverse roles in the modulation of behaviour,motor and sensory functions, feeding and reproduction. In con-trast, vertebrate FLPs appear considerably less abundant and havea much more focused functional remit. Accordingly, these peptidesand their associated processes have considerable appeal as poten-tial control targets for parasitic helminths (Maule et al., 2002;Mousley et al., 2005; McVeigh et al., 2006).
Following simple soaking protocols we compared knockdown attranscript level by quantitative (q)PCR, and the physiological im-pact of knockdown through migration assay. Our results indicatethat with certain siRNAs we can easily achieve significant and spe-cific knockdown. However, we also present findings of a single siR-NA which elicits non-target gene knockdown and an aberrantphenotype in both G. pallida and M. incognita J2s.
siRNA strand selection and incorporation into a RNA-InducedSilencing Complex (RISC) is thought to be mediated by helical ther-modynamics (Schwarz et al., 2003), in that the strand with thelowest thermodynamic stability at its 50 is predominantly selected
as a guide for incorporation into RISC. We present a simple tech-nique which may allow the investigator to influence the degreeof knockdown achieved by a selected siRNA, through incorporatingbase mismatches into the sense (non-mRNA specific) strand of thesiRNA helix, consequently decreasing 50 or 30 thermodynamic sta-bility. Although the efficacy of similar strategies has been reportedpreviously in the context of Drosophila whole-cell lysates (Schwarzet al., 2003) and mammalian cell preparations (Ding et al., 2008;Sano et al., 2008), to our knowledge this is the first reported exam-ple of such an approach in a whole organism.
2. Materials and methods
2.1. Nematode collection
Globodera pallida (pathotype Pa2/3) were collected from cul-tures of the potato cultivar Cara maintained at the Agri-Food andBiosciences Institute (AFBI), Belfast, Northern Ireland. Infective J2stage worms were hatched from the cysts in potato root diffusateunder complete darkness at room temperature, washed in diethyl-pyrocarbonate (DEPC)-treated water and used immediately for siR-NA soaking.
Meloidogyne incognita were maintained on cultures of suscepti-ble tomato species under greenhouse conditions. Roots were har-vested from infected plants and washed rigorously in water.Subsequently, egg masses were removed by brief dissolution in so-dium hypochlorite (2.5% v/v), and the free eggs washed thoroughlyin water. Eggs were isolated by sequential washing through a ser-ies of nested sieves, and placed in tomato root diffusate undercomplete darkness at room temperature. Freshly hatched J2 stageworms were taken immediately for siRNA soaking.
2.2. siRNA synthesis
siRNAs 1–4 of each species set, together with Gp-flp-12 siRNA 1aand Gp-flp-12 siRNA 1b, were synthesised using the Silencer� siRNAConstruction Kit (Applied Biosystems), and aliquots were stored atboth 4–7 �C, and �80 �C until required. Freezing aliquots was ini-tially avoided in an effort to maintain the integrity of 30 2 nucleo-tide (nt) overhangs, however we did not find any obvious reductionin siRNA knockdown efficacy following up to three cycles offreeze–thaw under our standard experimental conditions (siRNAdilutions were not freeze–thawed). siRNA 5 from each set was cus-tom designed and synthesised by Ambion, together with each spe-cies-specific randomized control (siRNA C) (see Table 1 for siRNAsequences).
2.3. siRNA soaks for qPCR analysis
Approximately 300 worms each of G. pallida and M. incognitawere soaked in 0.1 mg/ml siRNA diluted with DEPC-treated springwater to 50 ll total volume for 24 h. Control worms were soaked in50 ll DEPC-treated spring water, again for 24 h. Soaks were carriedout in duplicate, at room temperature in hydrophobically-lined,1.5 ml microcentrifuge tubes (Anachem, Bedfordshire, UK). Follow-ing the 24 h soak period, worms were washed three times in DEPC-treated spring water and mRNA extracted with the DynabeadsmRNA DIRECTTM Kit (Invitrogen Dynal AS), and treated with DNaseI (Invitrogen). mRNA concentrations were equalised to a 20 ng totalbetween replicates following spectrophotometric quantification at260 nm using NanoDropTM 1000 (NanoDrop Technologies), and usedas template for cDNA synthesis by SuperScriptTM II RNase H-ReverseTranscriptase, with an Oligo (dT) 12–18 primer and 10 mM dNTPMix (Invitrogen). The resultant cDNAs was used in triplicate qPCRsto analyse selected transcript levels post-soak (see Table 2 for

Table 1Globodera pallida (Gp)-flp-12 and Meloidogyne incognita (Mi)-flp-18 short interfering (si)RNA sequences; mismatches are shown in lower case where appropriate.
Gene (GenBank Accession No.) Species siRNA strand sequence (S: 50–30; A: 30–50) Construct size (bp)
Gp-flp-12 siRNA 1 (AJ300488) G. pallida S: UGAACGGAGUUGAUGGGGAUU21A: UUACUUGCCUCAACUACCCCU
Gp-flp-12 siRNA 1a (derived from AJ300488) G. pallida S: aGAACGGAGUUGAUGGGGAUU21A: UUaCUUGCCUCAACUACCCCU
Gp-flp-12 siRNA 1b (derived from AJ300488) G. mllida S: UGAACGGAGUUGAUGcGGAUU21A: UUACUUGCCUCAACUACcCCU
Gp-flp-12 siRNA 2 (AJ300488) G. pallida S: CGCUUCUGAUGCUGUCGGGUU21A: UCGCGAAGACUACGACAGCCC
Gp-flp-12 siRNA 3 (AJ300488) G. pallida S: GGAAUCCCGACUUUUGGGCUU21A: UACCUUAGGGCUGAAAACCCG
Gp-flp-12 siRNA 4 (AJ300488) G. pallida S: AGAUGCUCCCAUCGUCCCCUU21A: UCUCUACGAGGGUAGCAGGGG
Gp-flp-12 siRNA 5 (AJ300488) G. pallida S: ACGACCGACCAAUGAACGGUU21A: UUUGCUGGCUGGUUACUUGCC
Gp-flp-12 siRNA C (AJ300488) G. pallida S: GUACAUUUAACGCCAGCCUUU21A: UUCAUGUAAAUUGCGGUCGGA
Mi-flp-18 siRNA 1 (AY729022) M. incognita S: GCCGAUUUAAUGAACAAUUUU21A: UACGGCUAAAUUACUUGUUAA
Mi-flp-18 siRNA 2 (AY729022) M. incognita S: UUGCAAAUGGAGAGGCGGGUU21A: UUAACGUUUACCUCUCCGCCC
Mi-flp-18 siRNA 3 (AY729022) M. incognita S: CAAUUUUGGAGGACCUAGUUU21A: UUGUUAAAACCUCCUGGAUCA
Mi-flp-18 siRNA 4 (AY729022) M. incognita S: GAAUGCCUCAAGUCCUCCGUU21A: UUCUUACGGAGUUCAGGAGGC
Mi-flp-18 siRNA 5 (AY729022) M. incognita S: AUGGAGAGGCGGGUCAUAAUU21A: UUUACCUCUCCGCCCAGUAUU
Mi-flp-18 siRNA C (AY729022) M. incognita S: GUACAUUUAACGCCAGCCUUU21A: UUCAUGUAAAUUGCGGUCGGA
flp, FMRFamide-like peptide; S, sense; A, antisense; C, control.
J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100 93
primer sequences). qPCR was conducted using the Platinum�
SYBR� Green qPCR SuperMix-UDG kit (Invitrogen). Primer sets tobe used for qPCR were optimised for working concentration,annealing temperature and analysed by dissociation curve for con-tamination or non-specific amplification by primer–dimer as stan-dard. The PCR efficiency of each specific amplicon was calculated(Schmittgen and Livak, 2008) and used in the relative quantifica-tion of targets by the augmented comparative Ct method of Pfaffl(2001). Fold-change in transcript abundance was calibrated againstwater soaked worms and data were analysed by ANOVA andTukey’s Honestly Significant Difference (HSD) test using SPSS16.0 package for Windows (SPSS Inc., IL, USA).
2.4. Analysis of siRNA impact on phenotype and motor function bymigration assay
Approximately 300 J2 worms of G. pallida, and M. incognita weresoaked in 0.1 mg/ml of siRNA diluted with DEPC-treated springwater to a final volume of 50 ll for 24 h. Control worms weresoaked in 50 ll DEPC-treated spring water for 24 h. Soaks werecarried out in triplicate, at room temperature in hydrophobically-lined, 1.5 ml microcentrifuge tubes (Anachem). Following the24 h soak, worms from each of the preparations were subjectedto a migration assay. The soaked worms were added by pipetteto a column of washed, moistened USGA (US Golf Association)specification (grain diameter 0.25–1.0 mm) sand, and held in aglass tube (sand column length 80 mm, diameter 5 mm) by washednylon muslin. The glass tube sand column was retained verticallyin a glass vial containing 1 ml potato (for G. pallida), or tomato(for M. incognita) root diffusate which covered the base of the glasstube and sand column. The number of worms able to completemigration of the sand column was counted every hour for a total
of 4 h. Following completion of the 4 h period, worms unable tocomplete the migration were washed out of the glass tube togetherwith the sand column, isolated by sucrose centrifugation andcounted. The results were analysed by one-way ANOVA, and Tu-key’s HSD tests using the SPSS 16.0 package for Windows (SPSSInc., Chicago, IL, USA); probabilities of less than 5% (P < 0.05) weredeemed statistically significant. Migration percentages of watersoaked J2s were standardised to 100% and, subsequently, migra-tion percentages of dsRNA-soaked J2s were expressed relative tothe water-soaked controls.
3. Results
3.1. Real-time qPCR analyses of target and selected non-target genetranscripts in G. pallida following 24 h soak in 0.1 mg/ml siRNA
Gp-flp-12 siRNAs 1 (q = 14.33, P < 0.001), 2 (q = 5.627, P < 0.01)and 4 (q = 20.49, P < 0.001) yielded a significant level of targetmRNA knockdown in G. pallida J2s (75.52 ± 5.91-, 29.57 ± 7.39-,and 107.98 ± 15.513-fold reductions in Gp-flp-12 abundance,respectively). Although Gp-flp-12 siRNA 5 delivered a mean19.35 ± 1.71-fold knockdown of Gp-flp-12 mRNA, this was not ad-judged statistically significant (q = 3.673, P > 0.05). Gp-flp-12 siR-NA 3 (q = 0.00246, P > 0.05) did not lead to any significantdecrease in target transcript. All Gp-flp-12 siRNAs, with the excep-tion of Gp-flp-12 siRNA 4 (and siRNA 3 which had no effect) weredeemed specific, as no significant deviation in non-target genetranscript abundance was observed following siRNA soak. Gp-flp-12 siRNA 4 induced reductions in all tested transcripts; a31.27 ± 1.99-fold reduction of Gp-b actin (q = 45.95, P < 0.001),16.12 ± 0.37-fold reduction of Gp-b tubulin (q = 54.32, P < 0.001),

Table 2Primer sequences used for quantitative (q)PCR analysis of selected Globodera pallida and Meloidogyne incognita transcripts. Forward (F) and reverse (R) oligonucleotide sequencesgiven 50–30 .
Gene (GenBank Accession No.) Species Oligonucleotide sequence Construct size (bp)
b-Tubulin (AY689396) G. pallida F: CTTACGGCGATCTCAACCA 257R: CTTGGTCAGTTCCGCTACAG
FMRFamide-like peptide 12 (AJ300488) G. pallida F: CACTCCTTCTGGTCACATTGG 254R: GCGGAAAAGACGATAGAACG
b-Actin (CV579373) G. pallida F: CCGAGAAAAGATGACCCAAA 253R: AAGGAGTAACCACGCTCAGTG
FMRFamide-like peptide 14 (AJ310668) G. pallida F: CATCTTCCTCCTCTGTTGACC 248R: CCTCAGGTATTCGTGCTTCC
b-Tubulin (CN443135) M. incognita F: TGGAAAGTATGTCCCAAGAGC 257R: CACCACCAAGCGAGTGAGT
FMRFamide-like peptide 18 (AY729022) M. incognita F: CTGGGGTACTTCGATTTGGT 254R: GCGGAAAAGACGATAGAACG
b-Actin (BE225475) M. incognita F: GACTTGGCAGGACGTGATTT 253R: GACTTGGCAGGACGTGATTT
FMRFamide-like peptide 12 (AY804187) M. incognita F: TCAACTTATGGGGGCAATG 248R: CAACATTTATTGAGTGAGTTCCAA
94 J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100
and an 8.67 ± 1.42-fold decrease in Gp-flp-14 transcript abundance(q = 17.63, P < 0.001) (Fig. 1).
3.2. Real-time qPCR analyses of target and selected non-target genetranscripts in M. incognita following 24 h soak in dilutions of siRNA
Mi-flp-18 siRNAs 1 (q = 44.56, P < 0.001), 3 (q = 20.55, P <0.001) and 5 (q = 21.03, P < 0.001) induced significant reductions
Fig. 1. Mean fold-change in selected Globodera pallida (Gp) gene transcripts; (A) Gp-flp-1interfering (si)RNA soak. flp, FMRFamide-like peptide. ***P < 0.005.
in target transcript over the 24 h soak period. Mi-flp-18 siRNA 1was the most active of the set (q = 24.01, P < 0.001), yielding a36.05 ± 1.2-fold decrease in Mi-flp-18 transcript relative to thoseworms soaked only in water. Mi-flp-18 siRNAs 2 (q = 0.076, P >0.05) and 4 (q = 0.95, P > 0.05) did not elicit any significant reduc-tion of target transcript. None of the Mi-flp-18-specific siRNAs in-duced knockdown of the endogenous control genes, nor did G.pallida specific Gp-flp-12 siRNAs 1, 2, 3, or 5 (P > 0.05). Surprisingly
2, (B) Gp-b actin, (C) Gp-b tubulin and (D) Gp-flp-14 following 24 h 0.1 mg/ml short

J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100 95
however, Gp-flp-12 siRNA 4 did elicit non-specific knockdown of alltested gene transcripts in M. incognita J2s, as was the case with G.pallida J2 soaks. Knockdown of the M. incognita genes varied withthe target, resulting in a 2.08 ± 0.39-fold decrease in Mi-flp-12(q = 7.026, P < 0.001), a 3.48 ± 0.41-fold decrease in Mi-b tubulin(q = 10.46, P < 0.001), a 3.95 ± 0.66-fold decrease of Mi-flp-18(q = 4.918, P < 0.05), and a 7.78 ± 1.45-fold decrease of Mi-b actin(q = 15.06, P < 0.001) (Fig. 2).
3.3. siRNA-mediated transcript knockdown in G. pallida J2s on serialdilution
The two most active Gp-flp-12 siRNAs were selected for appraisalof knockdown efficacy following a series of dilutions. Gp-flp-12 siR-NA 1 yielded a mean 84.46 ± 3.29-fold decrease of target transcriptfrom 0.1 mg/ml to 0.1 lg/ml siRNA. On further dilution to 10 ng/ml (10,000-fold), 1 ng/ml (100,000-fold) and 10 pg/ml (1,000,000-fold) Gp-flp-12 knockdown was no longer statistically significant(q = 0.764, P > 0.05; q = 0.760, P > 0.05; and q = 0.0045, P > 0.05,respectively). Gp-flp-12 siRNA 4-mediated knockdown followed adifferent dilution dynamic, yielding a mean 139.77 (±6.82)-fold de-crease in target mRNA from 0.1 mg/ml to 1 lg/ml siRNA; however,unlike Gp-flp-12 siRNA 1, Gp-flp-12 siRNA 4 became significantly lesspotent on 1,000-fold dilution to 0.1 lg/ml, resulting in a 71.77(±3.36)-fold decrease of Gp-flp-12 transcript (q = 8.352, P < 0.001).Further dilution to 10 ng/ml and 10 pg/ml resulted in a high degreeof variability between repetitions (46.8 ± 28.67-fold and 3.48 ±3.32-fold reductions in Gp-flp-12, respectively). None of the dilu-
Fig. 2. Mean fold-change in selected Meloidogyne incognita (Mi) gene transcripts; (A) Mshort interfering (si)RNA soak. flp, FMRFamide-like peptide. *P < 0.05 and ***P < 0.005.
tions below 0.1 lg/ml yielded a statistically significant knockdownof transcript (P > 0.05). However, significant inhibition of wormmotility was observed following soaks in both 0.1 lg/ml and10 ng/ml siRNA (Fig. 3A).
Subsequently, we sought to characterise the non-target genesilencing activity observed on soaking G. pallida J2s with Gp-flp-12 siRNA 4, following dilution (Fig. 3B). Gp-b actin knockdownaveraged 32.21 (±1.3)-fold from 0.1 mg/ml to 1 lg/ml (q = 18.34,P < 0.001). However, on soaking with 0.1 lg/ml Gp-flp-12 siRNA4, Gp-b actin knockdown dropped to 7.3 ± 0.86-fold, statisticallysignificant from those J2s soaked in 0.1 mg/ml–1 lg/ml(q = 15.06, P < 0.001). Subsequent dilutions did not yield any signif-icant knockdown of Gp-b actin.
3.4. siRNA-mediated transcript knockdown in M. incognita J2s onserial dilution
Mi-flp-18 siRNA 1, as the most severe protagonist of Mi-flp-18transcript knockdown, was tested following a series of dilutions.A consistent mean 33.07 ± 0.83-fold decrease of Mi-flp-18 tran-script was recorded at siRNA amounts ranging from 0.1 mg/ml to0.1 lg/ml (1,000-fold dilution) (q = 10.82, P < 0.001). On furtherdilution to 10 ng/ml Mi-flp-18 siRNA 1, knockdown of target tran-script began to decrease (25.74 ± 1.42-fold decrease; q = 7.870,P < 0.01) and was statistically abolished on dilution to 1 ng/ml(q = 3.539, P > 0.05) and 10 pg/ml (q = 0.002, P > 0.05) siRNA(Fig. 3C). We also looked to characterise the non-specific gene tran-script knockdown elicited in M. incognita J2s by Gp-flp-12 siRNA 4,
i-flp-18, (B) Mi-b actin, (C) Mi-b tubulin and (D) Mi-flp-12 following 24 h 0.1 mg/ml
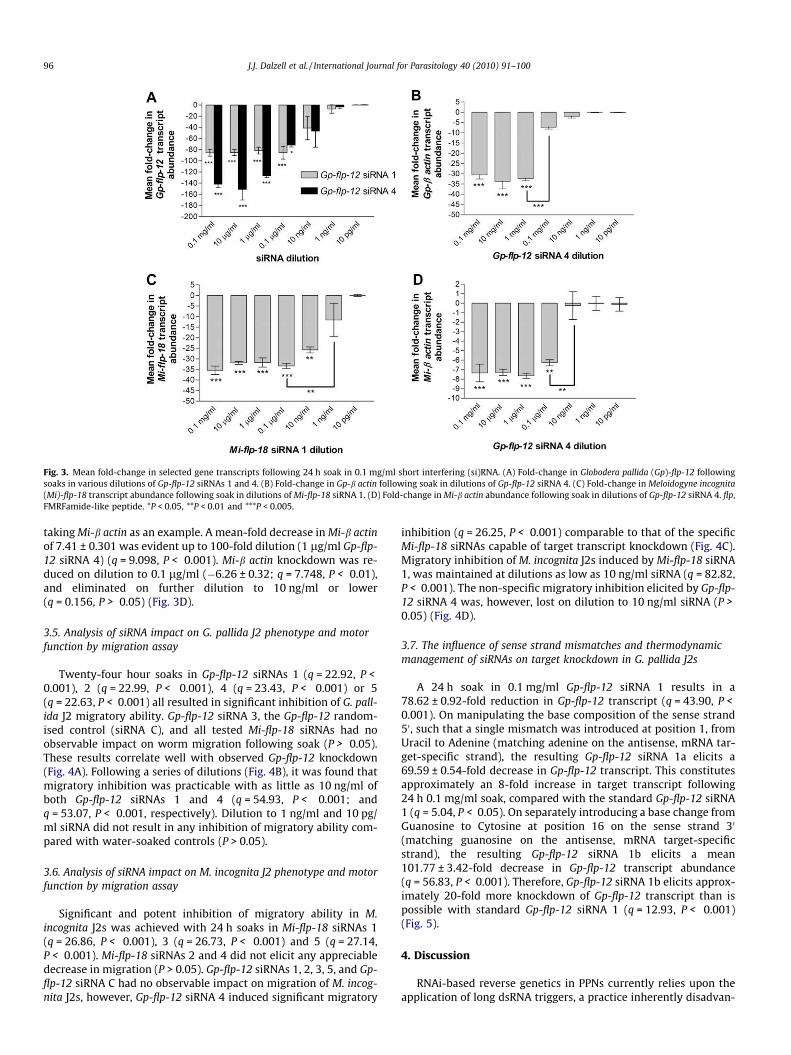
Fig. 3. Mean fold-change in selected gene transcripts following 24 h soak in 0.1 mg/ml short interfering (si)RNA. (A) Fold-change in Globodera pallida (Gp)-flp-12 followingsoaks in various dilutions of Gp-flp-12 siRNAs 1 and 4. (B) Fold-change in Gp-b actin following soak in dilutions of Gp-flp-12 siRNA 4. (C) Fold-change in Meloidogyne incognita(Mi)-flp-18 transcript abundance following soak in dilutions of Mi-flp-18 siRNA 1. (D) Fold-change in Mi-b actin abundance following soak in dilutions of Gp-flp-12 siRNA 4. flp,FMRFamide-like peptide. *P < 0.05, **P < 0.01 and ***P < 0.005.
96 J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100
taking Mi-b actin as an example. A mean-fold decrease in Mi-b actinof 7.41 ± 0.301 was evident up to 100-fold dilution (1 lg/ml Gp-flp-12 siRNA 4) (q = 9.098, P < 0.001). Mi-b actin knockdown was re-duced on dilution to 0.1 lg/ml (�6.26 ± 0.32; q = 7.748, P < 0.01),and eliminated on further dilution to 10 ng/ml or lower(q = 0.156, P > 0.05) (Fig. 3D).
3.5. Analysis of siRNA impact on G. pallida J2 phenotype and motorfunction by migration assay
Twenty-four hour soaks in Gp-flp-12 siRNAs 1 (q = 22.92, P <0.001), 2 (q = 22.99, P < 0.001), 4 (q = 23.43, P < 0.001) or 5(q = 22.63, P < 0.001) all resulted in significant inhibition of G. pall-ida J2 migratory ability. Gp-flp-12 siRNA 3, the Gp-flp-12 random-ised control (siRNA C), and all tested Mi-flp-18 siRNAs had noobservable impact on worm migration following soak (P > 0.05).These results correlate well with observed Gp-flp-12 knockdown(Fig. 4A). Following a series of dilutions (Fig. 4B), it was found thatmigratory inhibition was practicable with as little as 10 ng/ml ofboth Gp-flp-12 siRNAs 1 and 4 (q = 54.93, P < 0.001; andq = 53.07, P < 0.001, respectively). Dilution to 1 ng/ml and 10 pg/ml siRNA did not result in any inhibition of migratory ability com-pared with water-soaked controls (P > 0.05).
3.6. Analysis of siRNA impact on M. incognita J2 phenotype and motorfunction by migration assay
Significant and potent inhibition of migratory ability in M.incognita J2s was achieved with 24 h soaks in Mi-flp-18 siRNAs 1(q = 26.86, P < 0.001), 3 (q = 26.73, P < 0.001) and 5 (q = 27.14,P < 0.001). Mi-flp-18 siRNAs 2 and 4 did not elicit any appreciabledecrease in migration (P > 0.05). Gp-flp-12 siRNAs 1, 2, 3, 5, and Gp-flp-12 siRNA C had no observable impact on migration of M. incog-nita J2s, however, Gp-flp-12 siRNA 4 induced significant migratory
inhibition (q = 26.25, P < 0.001) comparable to that of the specificMi-flp-18 siRNAs capable of target transcript knockdown (Fig. 4C).Migratory inhibition of M. incognita J2s induced by Mi-flp-18 siRNA1, was maintained at dilutions as low as 10 ng/ml siRNA (q = 82.82,P < 0.001). The non-specific migratory inhibition elicited by Gp-flp-12 siRNA 4 was, however, lost on dilution to 10 ng/ml siRNA (P >0.05) (Fig. 4D).
3.7. The influence of sense strand mismatches and thermodynamicmanagement of siRNAs on target knockdown in G. pallida J2s
A 24 h soak in 0.1 mg/ml Gp-flp-12 siRNA 1 results in a78.62 ± 0.92-fold reduction in Gp-flp-12 transcript (q = 43.90, P <0.001). On manipulating the base composition of the sense strand50, such that a single mismatch was introduced at position 1, fromUracil to Adenine (matching adenine on the antisense, mRNA tar-get-specific strand), the resulting Gp-flp-12 siRNA 1a elicits a69.59 ± 0.54-fold decrease in Gp-flp-12 transcript. This constitutesapproximately an 8-fold increase in target transcript following24 h 0.1 mg/ml soak, compared with the standard Gp-flp-12 siRNA1 (q = 5.04, P < 0.05). On separately introducing a base change fromGuanosine to Cytosine at position 16 on the sense strand 30
(matching guanosine on the antisense, mRNA target-specificstrand), the resulting Gp-flp-12 siRNA 1b elicits a mean101.77 ± 3.42-fold decrease in Gp-flp-12 transcript abundance(q = 56.83, P < 0.001). Therefore, Gp-flp-12 siRNA 1b elicits approx-imately 20-fold more knockdown of Gp-flp-12 transcript than ispossible with standard Gp-flp-12 siRNA 1 (q = 12.93, P < 0.001)(Fig. 5).
4. Discussion
RNAi-based reverse genetics in PPNs currently relies upon theapplication of long dsRNA triggers, a practice inherently disadvan-
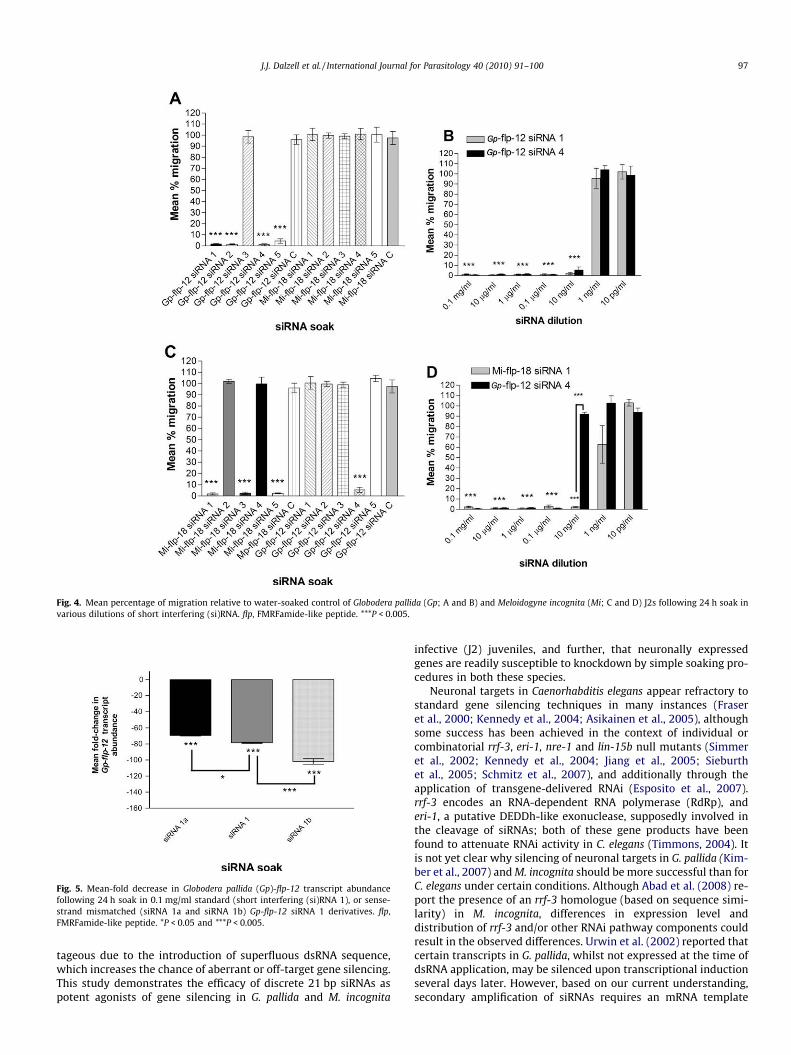
Fig. 4. Mean percentage of migration relative to water-soaked control of Globodera pallida (Gp; A and B) and Meloidogyne incognita (Mi; C and D) J2s following 24 h soak invarious dilutions of short interfering (si)RNA. flp, FMRFamide-like peptide. ***P < 0.005.
Fig. 5. Mean-fold decrease in Globodera pallida (Gp)-flp-12 transcript abundancefollowing 24 h soak in 0.1 mg/ml standard (short interfering (si)RNA 1), or sense-strand mismatched (siRNA 1a and siRNA 1b) Gp-flp-12 siRNA 1 derivatives. flp,FMRFamide-like peptide. *P < 0.05 and ***P < 0.005.
J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100 97
tageous due to the introduction of superfluous dsRNA sequence,which increases the chance of aberrant or off-target gene silencing.This study demonstrates the efficacy of discrete 21 bp siRNAs aspotent agonists of gene silencing in G. pallida and M. incognita
infective (J2) juveniles, and further, that neuronally expressedgenes are readily susceptible to knockdown by simple soaking pro-cedures in both these species.
Neuronal targets in Caenorhabditis elegans appear refractory tostandard gene silencing techniques in many instances (Fraseret al., 2000; Kennedy et al., 2004; Asikainen et al., 2005), althoughsome success has been achieved in the context of individual orcombinatorial rrf-3, eri-1, nre-1 and lin-15b null mutants (Simmeret al., 2002; Kennedy et al., 2004; Jiang et al., 2005; Sieburthet al., 2005; Schmitz et al., 2007), and additionally through theapplication of transgene-delivered RNAi (Esposito et al., 2007).rrf-3 encodes an RNA-dependent RNA polymerase (RdRp), anderi-1, a putative DEDDh-like exonuclease, supposedly involved inthe cleavage of siRNAs; both of these gene products have beenfound to attenuate RNAi activity in C. elegans (Timmons, 2004). Itis not yet clear why silencing of neuronal targets in G. pallida (Kim-ber et al., 2007) and M. incognita should be more successful than forC. elegans under certain conditions. Although Abad et al. (2008) re-port the presence of an rrf-3 homologue (based on sequence simi-larity) in M. incognita, differences in expression level anddistribution of rrf-3 and/or other RNAi pathway components couldresult in the observed differences. Urwin et al. (2002) reported thatcertain transcripts in G. pallida, whilst not expressed at the time ofdsRNA application, may be silenced upon transcriptional inductionseveral days later. However, based on our current understanding,secondary amplification of siRNAs requires an mRNA template

98 J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100
(Aoki et al., 2007). Perhaps this may be indicative of the reduced ornull influence of antagonistic factors such as eri-1, which may al-low for persistence of the primary siRNA population, or simplysome level of translational control applied to these transcriptsdownstream of RNAi.
Our results indicate that the efficacy of individual siRNAs, interms of transcript knockdown, varies significantly. Thus, rigorousinterpretation of post-silencing data may be necessary to delineatethe function of certain genes. Gp-flp-12 is predicted to encode a sin-gle mature peptide based on dibasic cleavage sites and sequencesimilarity with other characterised FLPs (KNKFEFIRFamide),whereas Mi-flp-18 encodes five predicted peptides (DDFIAPGVLRFamide, MPGVLRFamide, AVPGVLRFamide, QSQESGAVPGVLRFamide and MPQVLRFamide) (McVeigh et al., 2006). In the caseof the two flp genes studied here, it appears that a relatively lowlevel of transcript knockdown is necessary to elicit migratoryinhibition of the soaked J2s. This indicates that the neuropeptidesencoded by Gp-flp-12 and Mi-flp-18 are crucial for normal neuro-muscular function in G. pallida (Kimber et al., 2007) and M. incog-nita, respectively.
Whilst nearly all of the tested siRNAs seemed to function specif-ically, the finding that Gp-flp-12 siRNA 4 elicits non-target tran-script knockdown in both G. pallida and M. incognita wasunexpected. Based on sequence similarities, we had no obviousreason to expect serious interactions between this siRNA and othernematode transcripts in vivo. As expected, BLASTn analysis of bothGp-flp-12 siRNA 4 strands revealed many short contiguousstretches of base similarity to a variety of transcripts in both nem-atodes. None of the shared identities spanned more than 11 contig-uous base pairs, and few of these involved the seed-region of therelevant siRNA strand, which is believed to be heavily implicatedin limited homology interactions of small non-coding RNAs (Jinekand Doudna, 2009). Those sequence similarities which did spanthe guide RNA seed-region were invariably situated in predictedopen reading frames (ORFs), a situation shown to inhibit miRNA-like translational suppression and likely to reduce the influenceof off-target RNAi (Gu et al., 2009). Sequence similarity betweensiRNA and identifiable non-target nematode transcripts was asextensive in the entire group of siRNAs employed here (data notshown), and thus, at this stage we consider classical ‘‘off-target”RNAi, as we currently understand it, an insufficient basis to explainthe activity of Gp-flp-12 siRNA 4. It is possible that we observed aconserved toxicity event in response to Gp-flp-12 siRNA 4. ToxicRNA motifs have been reported previously in a range of biologicalcontexts (Fedorov et al., 2006; Ma et al., 2006; Li et al., 2008; Readet al., 2009) and may play a role in the observed phenomenon.
The ability to manipulate siRNA activity by influencing strandincorporation through the management of 50 strand thermody-namics represents an easy and achievable method of regulating,up or down, the silencing of a chosen mRNA. Schwarz et al.(2003) showed that sense strand mismatches could be used toinfluence guide strand selection and incorporation into RISC dur-ing Drosophila cell lysate assays, and Ding et al. (2008) demon-strate that a similar approach to nucleotide mismatch in thesense strand of an siRNA or hairpin (hp)RNA can be used to influ-ence strand selection in human embryonic kidney cells. Sanoet al. (2008) further demonstrate that on HeLa cell transfection,unilaterally blunted siRNAs which contain only one 30 2 nt over-hang on the antisense (guide) strand, consistently incorporate theantisense and thus reduce off-targets as a result of passenger(sense, in this context) strand incorporation into RISC; they addi-tionally showed that DNA mismatches in siRNAs can be used inisolation or in conjunction with unilaterally blunted siRNAs toimprove strand selection. Obvious applications of such a strategycould include the optimisation of siRNAs used to silence individ-ual members of highly conserved gene families, for example, in
isolated, yet divergent regions of sequence which may not natu-rally favour siRNA antisense (target-specific) strand incorporationinto RISC.
Mammalian and Drosophila cells do not encode currently recog-nisable RdRps, and thus may not possess the secondary amplifica-tion potential of C. elegans, a poorly understood mechanismwhereby RRF-1 (an RdRp) is recruited to the target transcriptwhere it initiates de novo synthesis of target-specific secondarysiRNAs, the origins of which propagate 50 and 30 along the relevantmRNA. These secondary siRNAs are single-stranded, exclusively ofantisense polarity, and characterised by a 50 triphosphate moiety.In C. elegans at least, these secondary siRNAs are then loaded intoa distinct set of Argonaute (AGO) proteins, primarily CSR-1 in C.elegans (Habig et al., 2008), which further elicits enzymatic cleav-age of cognate transcript (Aoki et al., 2007; Pak and Fire, 2007; Si-jen et al., 2007).
Caenorhabditis elegans encodes four distinct RdRps (Siomi andSiomi, 2009). Abad et al. (2008) report that based on sequence sim-ilarity, all four RdRp’s present in C. elegans have at least one puta-tive homologue in M. incognita. The impact of secondaryamplification on the RNAi response of M. incognita under experi-mentally relevant conditions is unknown. We would expect thatthe generation of secondary siRNAs would ablate modifications ap-plied to the primary pool of introduced siRNA; base compositionand mismatches would depend on the fidelity of RdRp amplifica-tion, and 30 overhangs are presumably no longer relevant in a pop-ulation of purportedly single-stranded secondary siRNAs.Therefore, we speculate that on exposure to low concentrationsof initial siRNA, the perceived benefit to reverse genetics applica-tions in M. incognita or any organism with active RdRps may be re-duced, or at best, manipulations of this type would result inquicker target knockdown up to a certain time point, followed bynormalisation of the RNAi response to that observed using unmod-ified siRNAs. The normalisation of response would correspond tothe point at which primary siRNA influence on target knockdownis surpassed by the RdRp-induced secondary population. We donot currently have the available sequence or annotation coverageto make informed predictions regarding the complement of RdRpsor RdRp-linked AGO proteins in G. pallida.
This study demonstrates modest but significant alterations ofGp-flp-12 transcript knockdown in response to sense strand mis-matches, which influence thermodynamic stability and asymmetryof the siRNA, as inferred by target knockdown. On introducing abase mismatch in the 50 of the sense strand, thermodynamic stabil-ity of the sense strand 50 is reduced, consequently increasing sus-ceptibility of the sense strand towards RISC incorporation. Onemay reverse the situation such that a base mismatch in the 30 ofthe sense strand (corresponding to the 50 of the antisense) will re-duce thermodynamic stability of the antisense 50, increasing thepropensity towards antisense incorporation as guide. The mis-matches applied in this study were selected at random within fournucleotides of the extreme 50 or 30 terminus of the sense strand, asappropriate. It is conceivable that either the resulting modificationof target knockdown presented in this study has been diluted bysecondary amplification of siRNAs in G. pallida, or that the time-frame of our experimental set-up is such that secondary amplifica-tion has not yet lead to appreciable synthesis of secondary siRNApools, and consequently the desired thermodynamic manipulationpersists in the dominant primary siRNA pool. Conversely, G. pallidamay not encode any functional RdRps. Regardless, we clearly dem-onstrated that under certain conditions, such an approach can beutilised to manipulate target knockdown in G. pallida J2s; a similarstrategy towards the manipulation of siRNA-mediated gene silenc-ing in M. incognita requires validation. We posit that the efficacy ofthermodynamic manipulation would invariably depend on the se-quence context of the passenger strand and gene silencing redun-

J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100 99
dancy of that strand on RISC incorporation. Unfortunately, it seemsthat the success of such modifications will follow a fairly ad hocdynamic as currently we are unable to predict the knockdown effi-cacy of any given siRNA a priori.
Little is known about how interactions of siRNA helical topologyand base mismatches may be tolerated by elements of the RNAipathway, as the mismatched stem-loop structures of miRNAs arethought to determine RISC loading (Siomi and Siomi, 2009).Although ‘‘RISC” is used as an encompassing term for any protein-aceous complex capable of eliciting RNAi effects, the activity of anygiven RISC is thought to be determined by the AGO protein con-tained within. It has been shown in a variety of different biologicalsystems, that only certain AGOs are cleavage-competent, acting bywhat we may consider ‘‘classical RNAi” and cleavage of cognatemRNAs, whilst others function purely through translational inhibi-tion (Tabara et al., 1999; Grishok et al., 2001; Tijsterman et al.,2002; Vastenhouw et al., 2003). However, in other organisms thesame AGO can function in both the enzymatic cleavage and trans-lational inhibition of targets in a context-dependent manner(Williams and Rubin, 2002; Meister et al., 2004; Okamura et al.,2004).
Current understanding dictates that central mismatches predis-pose guide incorporation into non-cleavage-competent AGOs, andfunctionally, towards translational inhibition – which may ormay not be relevant when attempting to target transcript ORFs(Gu et al., 2009). It is therefore possible that extensively mis-matched siRNAs may inadvertently load into AGOs which elicittranslational suppression rather than enzymatic cleavage.Although a useful discussion on the issues of control in siRNA-mediated RNAi is provided elsewhere (Nature Cell Biology, 2003),this has not yet been considered in the context of varying siRNAconformations.
We show that both randomized flp siRNAs and non-specific siR-NAs have (with exception) no global impact on mRNA abundance,phenotype or motile ability of the soaked J2s at 0.1 mg/ml siRNA,whereas dilution of the siRNAs to as little as 1 lg/ml still resultsin potent gene silencing and phenotype alteration. Importantly,this shows that siRNAs can be used to delineate specific gene func-tion following rigorous analysis, under these conditions. We havepreviously shown that at the same 0.1 mg/ml amount of longdsRNA, negative controls elicit an inhibitory phenotype in M. incog-nita J2s (Dalzell et al., 2009).
Whilst this study demonstrates successful application of siR-NAs towards the elucidation of specific gene function, it maybe pertinent to consider potential implications of such an ap-proach. Naturally, the biogenesis of small non-coding RNA spe-cies such as siRNAs and micro (mi)RNAs is tightly coupled withguide RISC incorporation, and in some cases, elements of the bio-synthetic pathway are themselves incorporated into the active or‘‘holo-RISC” (Jinek and Doudna, 2009). We are yet to consider theimplications of omitting the natural biosynthetic route to RISC onexperimentally introducing exogenous siRNA, and attempting todissect underlying mechanisms of transcript knockdown or toxic-ity may prove difficult to interpret in this context. The degree towhich eri-1 or related RNAi inhibitors attenuate the activity ofexperimentally administered siRNAs not coupled to biosyntheticRISC incorporation is unknown. Further, we cannot predict howthe thermodynamic asymmetry of administered siRNA correlatesto the loading mechanism of siRNAs evolved from dsRNAs or sec-ondary siRNA populations which may interact with additionalcofactors. Many unknowns remain with regard to gene silencingin parasitic nematodes, however we believe that reducing theconcentration and size of employed dsRNA may improve thereliability of RNAi screens in these species, and propose thatdiscrete 21 bp siRNAs provide a robust platform for furtherinvestigation.
Acknowledgements
This work was funded in part by both the European Social Fundand the Department of Agriculture and Rural Development forNorthern Ireland. The authors would like to thank Prof. HowardAtkinson, Dr. Wayne Charlton and Mandy Fearnehough of LeedsUniversity, UK, and Marie-Noelle Rosso of INRA, France, for kindlyproviding M. incognita-infected root material. Thanks also to Bren-dan Moreland and Maggie Hainon-McDowell for technicalassistance.
References
Abad, P., Gouzy, J., Aury, J.M., Castagnone-Sereno, P., Danchin, E.G., Deleury, E.,Perfus-Barbeoch, L., Anthouard, V., Artiguenave, F., Blok, V.C., Caillaud, M.C.,Coutinho, P.M., Dasilva, C., De Luca, F., Deau, F., Esquibet, M., Flutre, T.,Goldstone, J.V., Hamamouch, N., Hewezi, T., Jaillon, O., Jubin, C., Leonetti, P.,Magliano, M., Maier, T.R., Markov, G.V., McVeigh, P., Pesole, G., Poulain, J.,Robinson-Rechavi, M., Sallet, E., Segurens, B., Steinbach, D., Tytgat, T., Ugarte, E.,van Ghelder, C., Veronico, P., Baum, T.J., Blaxter, M., Bleve-Zacheo, T., Davis, E.L.,Ewbank, J.J., Favery, B., Grenier, E., Henrissat, B., Jones, J.T., Laudet, V., Maule,A.G., Quesneville, H., Rosso, M.N., Schiex, T., Smant, G., Weissenbach, J., Wincker,P., 2008. Genome sequence of the metazoan plant-parasitic nematodeMeloidogyne incognita. Nat. Biotechnol. 26, 909–915.
Aoki, K., Moriguchi, H., Yoshioka, T., Okawa, K., Tabara, H., 2007. In vitro analyses ofthe production and activity of secondary small interfering RNAs in C. elegans.EMBO J. 26, 5007–5019.
Asikainen, S., Vartiainen, S., Lakso, M., Nass, R., Wong, G., 2005. Selective sensitivity ofCaenorhabditis elegans neurons to RNA interference. Neuroreport 16, 1995–1999.
Bakhetia, M., Charlton, W., Atkinson, H.J., McPherson, M.J., 2005. RNA interference ofdual oxidase in the plant nematode Meloidogyne incognita. Mol. Plant MicrobeInteract. 18, 1099–1106.
Bird, D.M., Kaloshian, I., 2003. Are roots special? Nematodes have their say. Physiol.Mol. Plant Pathol. 62, 115–123.
Bird, D.M., 2004. Signaling between nematodes and plants. Curr. Opin. Plant Biol. 7,372–376.
Caillaud, M.C., Dubreuil, G., Quentin, M., Perfus-Barbeoch, L., Lecomte, P., deAlmeida Engler, J., Abad, P., Rosso, M.N., Favery, B., 2008. Root-knot nematodesmanipulate plant cell functions during a compatible interaction. J. Plant Physiol.165, 104–113.
Chitwood, D.J., 2003. Research on plant-parasitic nematode biology conducted bythe United States Department of Agriculture–Agricultural Research Service. PestManag. Sci. 59, 748–753.
Dalzell, J.D., McMaster, S., Johnston, M.J., Kerr, R., Fleming, C.C., Maule, A.G., 2009.Non-nematode-derived double stranded RNAs induce profound phenotypicchanges in Meloidogyne incognita and Globodera pallida infective juveniles. Int. J.Parasitol. doi:10.1016/j.ijpara.2009.05.006.
Ding, H., Liao, G., Wang, H., Zhou, Y., 2008. Asymmetrically designed siRNAs andshRNAs enhance the strand specificity and efficacy in RNAi. J. RNAi Gene Silenc.4, 269–280.
Esposito, G., Di Schiavi, E., Bergamasco, C., Bazzicalupo, P., 2007. Efficient and cellspecific knock-down of gene function in targeted C. elegans neurons. Gene 395,170–176.
Fairbairn, D.J., Cavallaro, A.S., Bernard, M., Mahalinga-Iyer, J., Graham, M.W., Botella,J.R., 2007. Host-delivered RNAi: an effective strategy to silence genes in plantparasitic nematodes. Planta 226, 1525–1533.
Fedorov, Y., Anderson, E.M., Birmingham, A., Reynolds, A., Karpilow, J., Robinson, K.,Leake, D., Marshall, W.S., Khvorova, A., 2006. Off-target effects by siRNA caninduce toxic phenotype. RNA 12, 1188–1196.
Fraser, A.G., Kamath, R.S., Zipperlen, P., Martinez-Campos, M., Sohrmann, M.,Ahringer, J., 2000. Functional genomic analysis of C. elegans chromosome 1 bysystematic RNA interference. Nature 408, 325–330.
Gheysen, G., Fenoll, C., 2002. Gene expression in nematode feeding sites. Annu. Rev.Phytopathol. 40, 191–219.
Grishok, A., Pasquinelli, A.E., Conte, D., Parrish, S., Ha, I., Baillie, D.L., Fire, A., Ruvkun,G., Mello, C.C., 2001. Genes and mechanisms related to RNA interferenceregulate expression of the small temporal RNAs that control C. elegansdevelopmental timing. Cell 106, 23–34.
Gu, S., Jin, L., Zhang, F., Sarnow, P., Kay, M.A., 2009. Biological basis for restriction ofmicroRNA targets to the 30 untranslated region in mammalian mRNAs. Nat.Struct. Mol. Biol. 16, 144–150.
Habig, J.W., Aruscavage, P.J., Bass, B.L., 2008. In C. elegans, high levels of dsRNA allowRNAi in the absence of RDE-4. PLoS One 3, e4052.
Huang, G., Allen, R., Davis, E.L., Baum, T.J., Hussey, R.S., 2006. Engineering broadroot-knot resistance in transgenic plants by RNAi silencing of a conserved andessential root-knot nematode parasitism gene. Proc. Natl. Acad. Sci. USA 103,14302–14306.
Issa, Z., Grant, W.N., Stasiuk, S., Shoemaker, C.B., 2005. Development of methods forRNA interference in the sheep gastrointestinal parasite, Trichostrongyluscolubriformis. Int. J. Parasitol. 35, 935–940.
Jackson, A.L., Burchard, J., Leake, D., Reynolds, A., Schelter, J., Guo, J., Johnson, J.M.,Lim, L., Karpilow, J., Nichols, K., Marshall, W., Khvorova, A., Linsley, P.S., 2006.

100 J.J. Dalzell et al. / International Journal for Parasitology 40 (2010) 91–100
Position-specific chemical modification of siRNAs reduces ‘‘off-target”transcript silencing. RNA 12, 1197–1205.
Jiang, G., Zhuang, L., Miyauchi, S., Miyake, K., Fei, Y., Ganapathy, V., 2005. A Na+/Cl�-coupled GABA transporter, GAT-1, from Caenorhabditis elegans. J. Biol. Chem.280, 2065–2077.
Jinek, M., Doudna, J.A., 2009. A three-dimensional view of the molecular machineryof RNA interference. Nature 457, 405–412.
Kennedy, S., Wang, D., Ruvkun, G., 2004. A conserved siRNA-degrading RNasenegatively regulates RNA interference in C. elegans. Nature 427, 645–649.
Kimber, M.J., McKinney, S., McMaster, S.A., Day, T.A., Fleming, C.C., Maule, A.G.,2007. Flp gene disruption in a parasitic nematode reveals motor dysfunctionand unusual neuronal sensitivity to RNA interference. FASEB J. 21, 1233–1243.
Lendner, M., Doligalska, M., Lucius, R., Hartmann, S., 2008. Attempts to establishRNA interference in the parasitic nematode Heligmosomoides polygyrus. Mol.Biochem. Parasitol. 161, 21–31.
Li, L.B., Yu, Z., Teng, X., Boninin, M., 2008. RNA toxicity is a component of ataxin-3degeneration in Drosophila. Nature 453, 1107–1111.
Ma, Y., Creanga, A., Lum, L., Beachy, P.A., 2006. Prevalence of off-target effects inDrosophila RNA interference screens. Nature 443, 359–363.
Maule, A.G., Mousley, A., Marks, N.J., Day, T.A., Thompson, D.P., Geary, T.G., Halton,D.W., 2002. Neuropeptide signalling systems – potential drug targets forparasite and pest control. Curr. Top. Med. Chem. 2, 733–758.
McVeigh, P., Geary, T.G., Marks, N.J., Maule, A.G., 2006. The FLP-side of nematodes.Trends Parasitol. 22, 385–396.
Meister, G., Landthaler, M., Dorsett, Y., Tuschl, T., 2004. Sequence-specific inhibitionof microRNA- and siRNA-induced RNA silencing. RNA 10, 544–550.
Mousley, A., Maule, A.G., Halton, D.W., Marks, N.J., 2005. Inter-phyla studies onneuropeptides: the potential for broad-spectrum anthelmintic and/orendectocide discovery. Parasitology 131, S143–S167.
Nature Cell Biology Editorial, 2003. Whither RNAi. Nat. Cell Biol. 4, 489–490.Okamura, K., Ishizuka, A., Siomi, H., Siomi, M.C., 2004. Distinct roles for Argonaute
proteins in small RNA-directed RNA cleavage pathways. Genes Dev. 18, 1655–1666.Pak, J., Fire, A., 2007. Distinct populations of primary and secondary effectors during
RNAi in C. elegans. Science 315, 241–244.Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-
time RT-PCR. Nucleic Acids Res. 29, e45.Read, M.L., Mir, S., Spice, R., Seabright, R.J., Suggate, E.L., Ahmed, Z., Berry, M., Logan,
A., 2009. Profiling RNA interference (RNAi)-mediated toxicity in neural culturesfor effective short interfering RNA design. J. Gene Med. doi:10.1002/jgm.1325.
Rosso, M.N., Dubrana, M.P., Cimbolini, N., Jaubert, S., Abad, P., 2005. Application ofRNA interference to root-knot nematode genes encoding esophageal glandproteins. Mol. Plant Microbe Interact. 18, 615–620.
Sano, M., Sierant, M., Miyagishi, M., Nakanishi, M., Takagi, Y., Sutou, S., 2008. Effectof asymmetric terminal structures of short RNA duplexes on the RNAinterference activity and strand selection. Nucleic Acids Res. 36, 5812–5821.
Schmittgen, T.D., Livak, K.J., 2008. Analyzing real-time PCR data by the comparativeC(T) method. Nat. Protoc. 3, 1101–1108.
Schmitz, C., Kinge, P., Hutter, H., 2007. Axon guidance genes identified in a largescale RNAi screen using the RNAi hypersensitive Caenorhabditis elegans strainnre-1(hd20) lin-15b(hd126). Proc. Natl. Acad. Sci. USA 104, 834–839.
Schwarz, D.S., Hutvagner, G., Du, T., Xu, Z., Aronin, N., Zamore, P.D., 2003.Asymmetry in the assembly of the RNAi enzyme complex. Cell 115, 199–208.
Shingles, J., Lilley, C.J., Atkinson, H.J., Urwin, P.E., 2006. Meloidogyne incognita:molecular and biochemical characterisation of a cathepsin L cysteine proteinaseand the effect on parasitism following RNAi. Exp. Parasitol. 115, 114–120.
Sieburth, D., Ch’ng, Q., Dybbs, M., Tavazoie, M., Kennedy, S., Wang, D., Dupuy, D.,Raul, J.F., Hill, D.E., Vidal, M., Ruvkun, G., Kaplan, J.M., 2005. Systematic analysisof genes required for synapse structure and function. Nature 436, 510–517.
Sijen, T., Steiner, F.A., Thijssen, K.L., Plasterk, R.H., 2007. Secondary siRNAs resultfrom unprimed RNA synthesis and form a distinct class. Science 315, 244–247.
Simmer, F., Tijsterman, M., Parrish, S., Koushika, S.P., Nonet, M.L., Fire, A., Ahringer,J., Plasterk, R.H., 2002. Loss of the putative RNA-directed RNA polymerase RRF-3makes C. elegans hypersensitive to RNAi. Curr. Biol. 12, 1317–1319.
Sindhu, A.S., Maier, T.R., Mitchum, M.G., Hussey, R.S., Davis, E.L., Baum, T.J., 2009.Effective and specific in planta RNAi in cyst nematodes: expression interferenceof four parasitism genes reduces parasitic success. J. Exp. Bot. 60, 315–324.
Siomi, H., Siomi, M., 2009. On the road to reading the RNA-interference code. Nature457, 396–404.
Steeves, R.M., Todd, T.C., Essig, J.S., Trick, H.N., 2006. Transgenic soybeansexpressing siRNAs specific to a major sperm protein gene suppress Heteroderaglycines reproduction. Funct. Plant Biol. 33, 991–999.
Tabara, H., Sarkissian, M., Kelly, W.G., Fleenor, J., Grishok, A., Timmons, L., Fire, A.,Mello, C.C., 1999. The rde-1 gene, RNA interference, and transposon silencing inC. elegans. Cell 99, 123–132.
Timmons, L., 2004. Endogenous inhibitors of RNA interference in Caenorhabditiselegans. Bioessays 26, 715–718.
Tijsterman, M., Ketting, R.F., Plasterk, R.H., 2002. The genetics of RNA silencing.Annu. Rev. Genet. 36, 489–519.
Urwin, P.E., Lilley, C.J., Atkinson, H.J., 2002. Ingestion of double-stranded RNA bypreparasitic cyst nematodes leads to RNA interference. Mol. Plant MicrobeInteract. 15, 747–752.
Vastenhouw, N.L., Fischer, S.E., Robert, V.J., Thijssen, K.L., Fraser, A.G., Kamath, R.S.,Ahringer, J., Plasterk, R.H., 2003. A genome-wide screen identifies 27 genesinvolved in transposon silencing in C. elegans. Curr. Biol. 13, 1311–1316.
Williams, R.W., Rubin, G.M., 2002. ARGONAUTE1 is required for efficient RNAinterference in Drosophila embryos. Proc. Natl. Acad. Sci. USA 99, 6889–6894.
Yadav, B.C., Veluthambi, K., Subramaniam, K., 2006. Host-generated doublestranded RNA induces RNAi in plant-parasitic nematodes and protects thehost from infection. Mol. Biochem. Parasitol. 148, 219–222.