serotonergic mechanisms in the basolateral amygdala differentially regulate the conditioned and...
TRANSCRIPT

www.e l sev i e r. com/ loca te /eu roneu ro
European Neuropsychopharmacology (2007) 17, 717–724
Serotonergic mechanisms in the basolateralamygdala differentially regulate the conditionedand unconditioned fear organizedin the periaqueductal grayRaquel Chacon Ruiz Martinez, Amanda Ribeiro de Oliveira,Marcus Lira Brandão ⁎
Instituto de Neurociências & Comportamento-INeC, Campus USP, 14040-901, Ribeirão Preto, SP, BrazilLaboratório de Psicobiologia, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto,Universidade de São Paulo (USP) 14040-901, Ribeirão Preto, SP, Brazil
Received 9 December 2006; received in revised form 14 February 2007; accepted 20 February 2007
⁎ Corresponding author. Tel.: +55 164830.
E-mail address: [email protected]
0924-977X/$ - see front matter © 200doi:10.1016/j.euroneuro.2007.02.015
Abstract The amygdala is an important filter for unconditioned and conditioned aversiveinformation. The amygdala synthesizes the stimuli input from the environment and then signalsthe degree of threat that they represent to the dorsal periaqueductal gray (dPAG), which wouldbe in charge of selecting, organizing and executing the appropriate defense reaction. In thisstudy, we examined the influence of fluoxetine microinjections (1.75 and 3.5 nmol/0.2 μL) intothe lateral (LaA) and basolateral (BLA) amygdaloid nuclei on the freezing and escape responsesinduced by electrical stimulation of the dPAG. Freezing behavior was also measured after theinterruption of the electrical stimulation of the dPAG. On the following day, these rats were alsosubmitted to a contextual fear paradigm to examine whether these microinjections would affectthe conditioned freezing to contextual cues previously associated with foot shocks. Fluoxetineinjections into both amygdaloid nuclei did not change the freezing and escape thresholds, butdisrupted the dPAG-post-stimulation freezing. Moreover, the conditioned freezing was enhancedby fluoxetine. Whereas 5-HT mechanisms in the amygdala facilitate the acquisition ofconditioned fear they inhibit the dPAG-post-stimulation freezing. However, the unconditionedfear triggered by activation of the dPAG is produced downstream of the amygdala. These findingshave important implications for the understanding of the neurochemical substrates that underliepanic and generalized anxiety disorders.© 2007 Elsevier B.V. and ECNP. All rights reserved.
KEYWORDSUnconditioned andconditioned freezing;dPAG;Lateral nucleus ofthe amygdala;Basolateral nucleus ofthe amygdala;Serotonin
3602 3638; fax: +55 16 3602
(M.L. Brandão).
7 Elsevier B.V. and ECNP. All right
1. Introduction
It has been proposed that the amygdala, the medialhypothalamus, and the dorsal periaqueductal gray (dPAG)
s reserved.

718 R.C.R. Martinez et al.
together constitute the brain aversion system, which hasbeen related to the organization of anxiety-like states inlaboratory animals (Graeff, 1990, 2004). Electrical stimula-tion of the dPAG induces defensive responses such as arousal,freezing and escape that mimic fearful behavior elicited byenvironmental challenges (Brandão et al., 1994, 1999, 2005).The amygdala synthesizes the stimulus input from theenvironment and then signals to the dorsal periaqueductalgray (dPAG) the degree of threat that they represent to theorganism (Fanselow, 1991).
A differential involvement of the amygdaloid nuclei in theregulation of unconditioned and conditioned fear is wellestablished. For instance, the basolateral complex is crucialfor the acquisition of conditioned fear whereas the centralnucleus of the amygdala is involved in the expression ofinnate and learned responses (LeDoux, 1994; LeDoux et al.,1990; Davis et al., 1994; Killcross et al., 1997). However, theregulatory mechanisms of conditioned and unconditionedfear may differ even within a given amygdaloid nucleus.Indeed, injections of ketanserin – a 5-HT2 receptor antag-onist – into the basolateral complex of the amygdala havebeen found to enhance the aversiveness of the chemicalstimulation of the inferior colliculus in rats placed in anopen-field test and to reduce the conditioned fear (Macedoet al., 2007). In view of these results, it has been proposedthat the filtering mechanisms in the basolateral (BLA) andlateral (LaA) nuclei of the amygdala have special character-istics depending on the nature of the aversive stimulus.Furthermore, there is a consensus on the need of studies thatextend our knowledge on the involvement of 5-HT regulationof the basolateral complex of the amygdala on conditionedand unconditioned fear.
The serotonin (5-HT) systemhas been extensively studied inthe neurobiology of fear and anxiety (Maisonnette et al., 1993;Deakin and Graeff, 1991; Gray and McNaughton, 2000; Graeff,2004). Microinjection of 5-HT receptor antagonists within theamygdala releases punished behavior, whereas 5-HT receptoractivation increases response suppression (Wise et al., 1972;Iversen, 1984). In contrastwith conflict tests, however,modelsin which animals actively escape or avoid aversive brainstimulation point to an anxiolytic role for 5-HT in the amygdala(Macedo et al., 2007). Both the amygdala and the dPAG receiveserotonergic input mainly from the dorsal raphe nucleus(Vertes, 1991). The axons that project onto the amygdalafollow the DRN-forebrain tract, while those that go to the PAGrun through the dorsal raphe nucleus-periventricular tract(Azmitia and Segal, 1978). It seems that these two aversivesystems are not entirely independent and some interactionbetween them may exist. For example, it has been proposedthat anxiety states generated at the amygdala level mayinhibit panic attacks elicited by activation of the neuralsubstrates of aversion in the dPAG (Graeff, 1990, 2004). Basedon these findings, we set out to determine whether injectionsof fluoxetine (1.75 and 3.5 nmol/0.2 μL) into the BLA or LAwould affect the conditioned fear or the unconditioned fearproduced by electrical stimulation of the dPAG. In fearconditioning, a neutral conditioned stimulus, such as a spatialcontext, acquires the capacity to elicit defensive responsesafter being associated with an aversive unconditioned stimu-lus, typically a foot shock. The freezing and escape behaviorinduced by gradual increases in the electrical stimulation ofthe dPAG were the unconditioned responses. Fluoxetine is a
selective serotonin reuptake inhibitor (SSRI) acting at thenerve terminals leading to an increase of the extracellular 5-HT levels around cell bodies (Goldstein and Goodnick, 1998;Blier, 2001; Burghardt et al., 2004). The dose of 3.5 nmol/0.2 μL was chosen based on previous work (Pasini et al., 1996).The lateral and basolateral nuclei have been conventionallyconsidered as part of the basolateral complex of the amygdala(Krettek and Price, 1978; Ottersen, 1981; McDonald, 1992).
2. Methods
2.1. Animals
Fifty-eight male Wistar rats weighing 250−300 g from the animalhouse of the Campus of Ribeirão Preto of the University of São Paulowere housed in a temperature-controlled (22±1 °C) room andmaintained on a 12-h light/12-h dark cycle (lights on: 07:00–19:00).These animals weremaintained in pairs in Plexiglas-walled cages andgiven free access to food and water throughout the experiment. Therats were randomly assigned to one of two surgery groups: lateraland basolateral nuclei of amygdala. The experiments were carriedout according to the Brazilian Society for Neuroscience and BehaviorGuidelines for Care and Use of Laboratory Animals.
2.2. Surgery
The animals were anesthetized with tribromoethanol (250 mg/kg,i.p.) and fixed in a stereotaxic frame (David Kopf, Tujunga, CA).The upper incisor bar was set at 3.3 mm below the interaural line,such that the skull was horizontal between bregma and lambda.Each animal was implanted with a unilateral bipolar electrodeaimed at the right dPAG. The electrode (14 mm) was made ofstainless steel wire, 160 μm in diameter and insulated except at thecross-section. It was introduced at a 16° angle with lambda servingas the reference for each plane: antero-posterior (AP)=0.0 mm;medio-lateral (ML)=1.9 mm; and dorso-ventral (DV)=5.1 mm.Next, a unilateral guide-cannula (through which an injectioncannula could be inserted for fluoxetine or saline applicationsone week later) was stereotaxically implanted over the right LaA orBLA. Taking bregma as reference point, the coordinates for bothnuclei were: lateral (AP=3.0 mm, ML=5.1 mm, DV=7.0 mm);basolateral (AP=2.5 mm; ML=5.1 mm; DV=7.7 mm). For bothgroups, the electrode and cannula were fixed to the skull bymeans of acrylic resin and two stainless steel screws. The electrodewire was connected to a male pin, so that it could be plugged intoan amphenol socket at the end of a flexible electrical cable andused for brain stimulation. At the end of the surgery, each guide-cannula was sealed with a stainless steel wire to protect it fromcongestion.
2.3. Microinjections
Fluoxetine (1.75 and 3.5 nmol/0.2 μL) was microinjected in LaA orBLA. Control animals received the same volume of saline, which alsoserved as vehicle for the injections. The injection needle was a thindental needle (0.3 mm, o.d.) connected to a 5-μL Hamilton syringeby means of a polyethylene tube. The injection needle wasintroduced through the guide cannula until its lower end was 1 mmbelow the guide cannula. The solutions were injected into the LaA orBLA (0.2 μL/min), with the help of an infusion pump (HarvardApparatus, South Natick, MA). The displacement of an air bubbleinside the polyethylene catheter (PE-10; Becton-Dickinson, FranklinLakes, NJ) connecting the syringe needle to the intracerebral needlewas used to monitor the microinjection. The needle was held inplace for an additional 1 min to maximize diffusion away from the tip
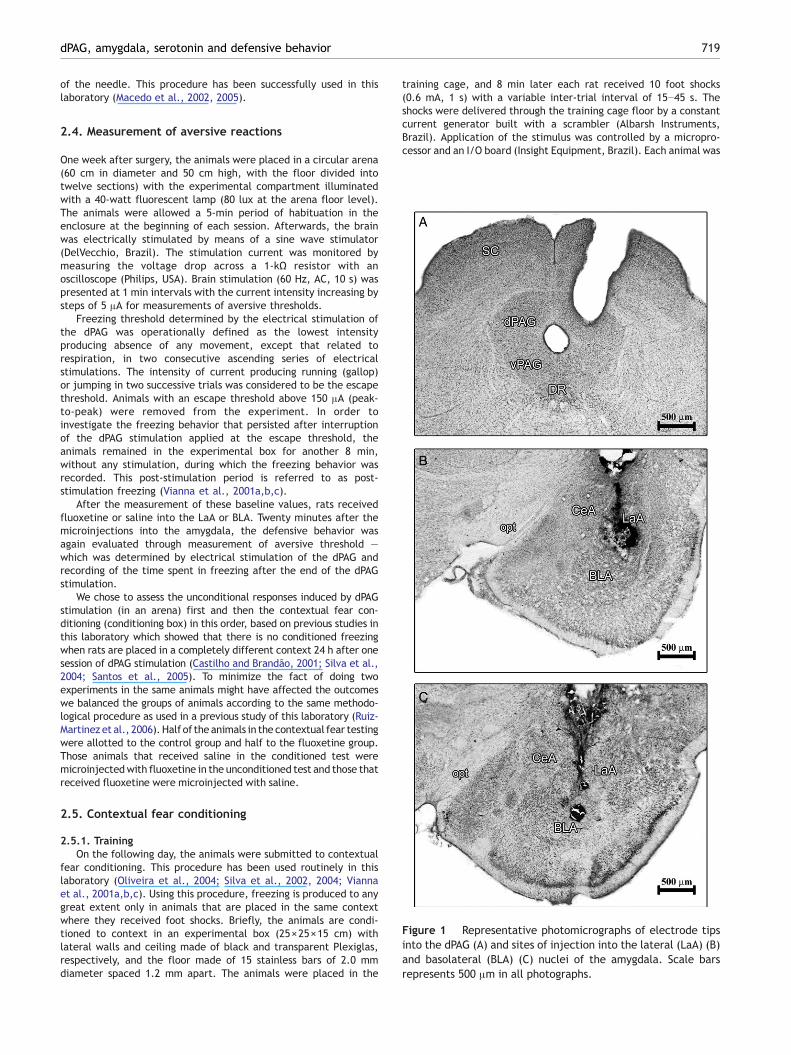
Figure 1 Representative photomicrographs of electrode tipsinto the dPAG (A) and sites of injection into the lateral (LaA) (B)and basolateral (BLA) (C) nuclei of the amygdala. Scale barsrepresents 500 μm in all photographs.
719dPAG, amygdala, serotonin and defensive behavior
of the needle. This procedure has been successfully used in thislaboratory (Macedo et al., 2002, 2005).
2.4. Measurement of aversive reactions
One week after surgery, the animals were placed in a circular arena(60 cm in diameter and 50 cm high, with the floor divided intotwelve sections) with the experimental compartment illuminatedwith a 40-watt fluorescent lamp (80 lux at the arena floor level).The animals were allowed a 5-min period of habituation in theenclosure at the beginning of each session. Afterwards, the brainwas electrically stimulated by means of a sine wave stimulator(DelVecchio, Brazil). The stimulation current was monitored bymeasuring the voltage drop across a 1-kΩ resistor with anoscilloscope (Philips, USA). Brain stimulation (60 Hz, AC, 10 s) waspresented at 1 min intervals with the current intensity increasing bysteps of 5 μA for measurements of aversive thresholds.
Freezing threshold determined by the electrical stimulation ofthe dPAG was operationally defined as the lowest intensityproducing absence of any movement, except that related torespiration, in two consecutive ascending series of electricalstimulations. The intensity of current producing running (gallop)or jumping in two successive trials was considered to be the escapethreshold. Animals with an escape threshold above 150 μA (peak-to-peak) were removed from the experiment. In order toinvestigate the freezing behavior that persisted after interruptionof the dPAG stimulation applied at the escape threshold, theanimals remained in the experimental box for another 8 min,without any stimulation, during which the freezing behavior wasrecorded. This post-stimulation period is referred to as post-stimulation freezing (Vianna et al., 2001a,b,c).
After the measurement of these baseline values, rats receivedfluoxetine or saline into the LaA or BLA. Twenty minutes after themicroinjections into the amygdala, the defensive behavior wasagain evaluated through measurement of aversive threshold —which was determined by electrical stimulation of the dPAG andrecording of the time spent in freezing after the end of the dPAGstimulation.
We chose to assess the unconditional responses induced by dPAGstimulation (in an arena) first and then the contextual fear con-ditioning (conditioning box) in this order, based on previous studies inthis laboratory which showed that there is no conditioned freezingwhen rats are placed in a completely different context 24 h after onesession of dPAG stimulation (Castilho and Brandão, 2001; Silva et al.,2004; Santos et al., 2005). To minimize the fact of doing twoexperiments in the same animals might have affected the outcomeswe balanced the groups of animals according to the same methodo-logical procedure as used in a previous study of this laboratory (Ruiz-Martinez et al., 2006). Half of the animals in the contextual fear testingwere allotted to the control group and half to the fluoxetine group.Those animals that received saline in the conditioned test weremicroinjectedwith fluoxetine in the unconditioned test and those thatreceived fluoxetine were microinjected with saline.
2.5. Contextual fear conditioning
2.5.1. TrainingOn the following day, the animals were submitted to contextual
fear conditioning. This procedure has been used routinely in thislaboratory (Oliveira et al., 2004; Silva et al., 2002, 2004; Viannaet al., 2001a,b,c). Using this procedure, freezing is produced to anygreat extent only in animals that are placed in the same contextwhere they received foot shocks. Briefly, the animals are condi-tioned to context in an experimental box (25×25×15 cm) withlateral walls and ceiling made of black and transparent Plexiglas,respectively, and the floor made of 15 stainless bars of 2.0 mmdiameter spaced 1.2 mm apart. The animals were placed in the
training cage, and 8 min later each rat received 10 foot shocks(0.6 mA, 1 s) with a variable inter-trial interval of 15−45 s. Theshocks were delivered through the training cage floor by a constantcurrent generator built with a scrambler (Albarsh Instruments,Brazil). Application of the stimulus was controlled by a micropro-cessor and an I/O board (Insight Equipment, Brazil). Each animal was
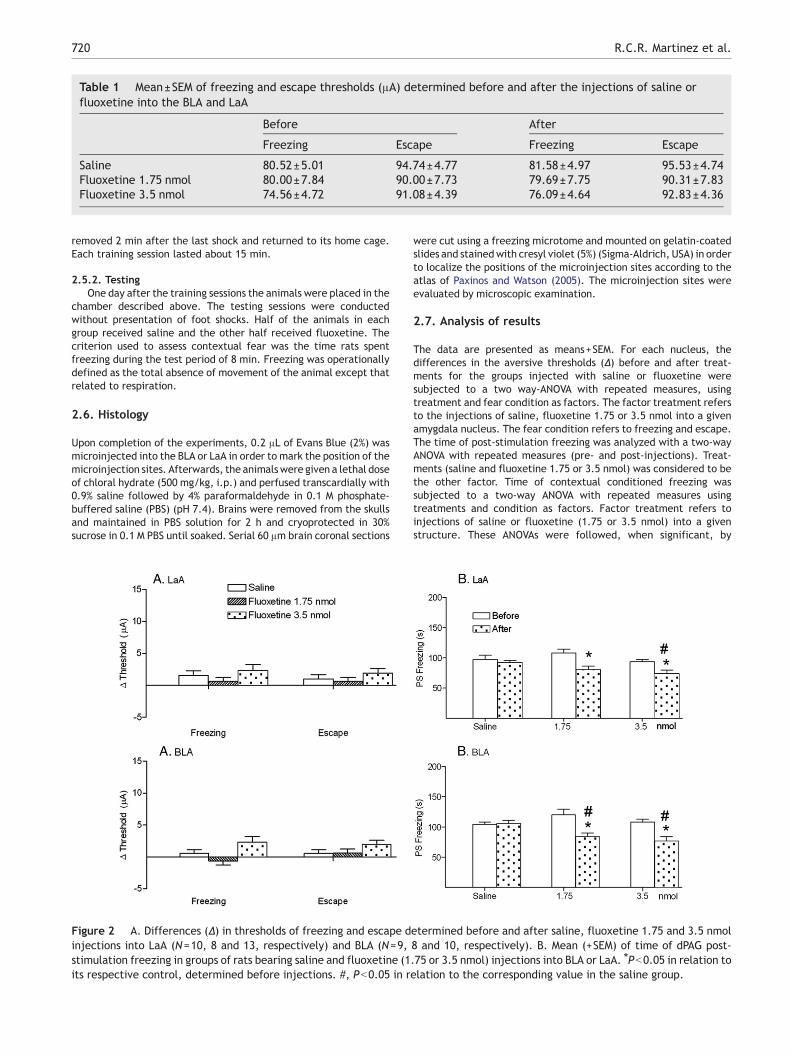
Table 1 Mean±SEM of freezing and escape thresholds (μA) determined before and after the injections of saline orfluoxetine into the BLA and LaA
Before After
Freezing Escape Freezing Escape
Saline 80.52±5.01 94.74±4.77 81.58±4.97 95.53±4.74Fluoxetine 1.75 nmol 80.00±7.84 90.00±7.73 79.69±7.75 90.31±7.83Fluoxetine 3.5 nmol 74.56±4.72 91.08±4.39 76.09±4.64 92.83±4.36
720 R.C.R. Martinez et al.
removed 2 min after the last shock and returned to its home cage.Each training session lasted about 15 min.
2.5.2. TestingOne day after the training sessions the animals were placed in the
chamber described above. The testing sessions were conductedwithout presentation of foot shocks. Half of the animals in eachgroup received saline and the other half received fluoxetine. Thecriterion used to assess contextual fear was the time rats spentfreezing during the test period of 8 min. Freezing was operationallydefined as the total absence of movement of the animal except thatrelated to respiration.
2.6. Histology
Upon completion of the experiments, 0.2 μL of Evans Blue (2%) wasmicroinjected into the BLA or LaA in order to mark the position of themicroinjection sites. Afterwards, the animalswere given a lethal doseof chloral hydrate (500 mg/kg, i.p.) and perfused transcardially with0.9% saline followed by 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) (pH 7.4). Brains were removed from the skullsand maintained in PBS solution for 2 h and cryoprotected in 30%sucrose in 0.1 M PBS until soaked. Serial 60 μm brain coronal sections
Figure 2 A. Differences (Δ) in thresholds of freezing and escape dinjections into LaA (N=10, 8 and 13, respectively) and BLA (N=9,stimulation freezing in groups of rats bearing saline and fluoxetine (1its respective control, determined before injections. #, Pb0.05 in r
were cut using a freezing microtome and mounted on gelatin-coatedslides and stainedwith cresyl violet (5%) (Sigma-Aldrich, USA) in orderto localize the positions of the microinjection sites according to theatlas of Paxinos and Watson (2005). The microinjection sites wereevaluated by microscopic examination.
2.7. Analysis of results
The data are presented as means+SEM. For each nucleus, thedifferences in the aversive thresholds (Δ) before and after treat-ments for the groups injected with saline or fluoxetine weresubjected to a two way-ANOVA with repeated measures, usingtreatment and fear condition as factors. The factor treatment refersto the injections of saline, fluoxetine 1.75 or 3.5 nmol into a givenamygdala nucleus. The fear condition refers to freezing and escape.The time of post-stimulation freezing was analyzed with a two-wayANOVA with repeated measures (pre- and post-injections). Treat-ments (saline and fluoxetine 1.75 or 3.5 nmol) was considered to bethe other factor. Time of contextual conditioned freezing wassubjected to a two-way ANOVA with repeated measures usingtreatments and condition as factors. Factor treatment refers toinjections of saline or fluoxetine (1.75 or 3.5 nmol) into a givenstructure. These ANOVAs were followed, when significant, by
etermined before and after saline, fluoxetine 1.75 and 3.5 nmol8 and 10, respectively). B. Mean (+SEM) of time of dPAG post-.75 or 3.5 nmol) injections into BLA or LaA. ⁎Pb0.05 in relation toelation to the corresponding value in the saline group.
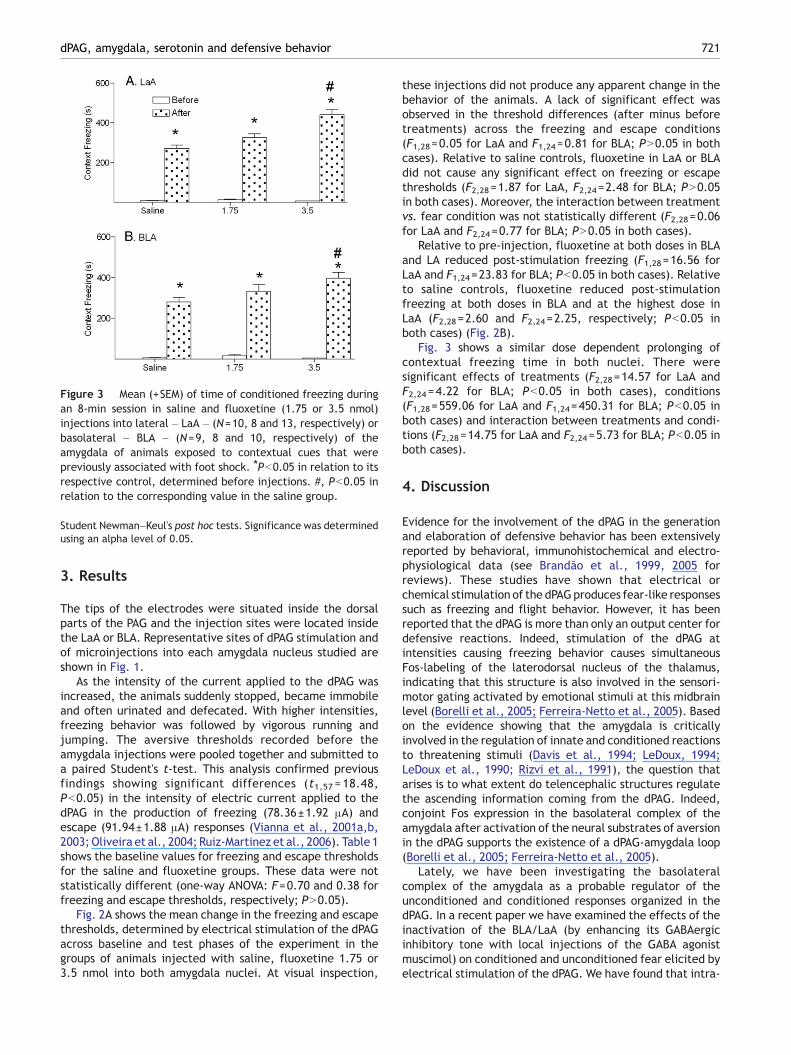
Figure 3 Mean (+SEM) of time of conditioned freezing duringan 8-min session in saline and fluoxetine (1.75 or 3.5 nmol)injections into lateral – LaA – (N=10, 8 and 13, respectively) orbasolateral – BLA – (N=9, 8 and 10, respectively) of theamygdala of animals exposed to contextual cues that werepreviously associated with foot shock. ⁎Pb0.05 in relation to itsrespective control, determined before injections. #, Pb0.05 inrelation to the corresponding value in the saline group.
721dPAG, amygdala, serotonin and defensive behavior
Student Newman–Keul's post hoc tests. Significance was determinedusing an alpha level of 0.05.
3. Results
The tips of the electrodes were situated inside the dorsalparts of the PAG and the injection sites were located insidethe LaA or BLA. Representative sites of dPAG stimulation andof microinjections into each amygdala nucleus studied areshown in Fig. 1.
As the intensity of the current applied to the dPAG wasincreased, the animals suddenly stopped, became immobileand often urinated and defecated. With higher intensities,freezing behavior was followed by vigorous running andjumping. The aversive thresholds recorded before theamygdala injections were pooled together and submitted toa paired Student's t-test. This analysis confirmed previousfindings showing significant differences (t1,57 = 18.48,Pb0.05) in the intensity of electric current applied to thedPAG in the production of freezing (78.36±1.92 μA) andescape (91.94±1.88 μA) responses (Vianna et al., 2001a,b,2003; Oliveira et al., 2004; Ruiz-Martinez et al., 2006). Table 1shows the baseline values for freezing and escape thresholdsfor the saline and fluoxetine groups. These data were notstatistically different (one-way ANOVA: F=0.70 and 0.38 forfreezing and escape thresholds, respectively; PN0.05).
Fig. 2A shows the mean change in the freezing and escapethresholds, determined by electrical stimulation of the dPAGacross baseline and test phases of the experiment in thegroups of animals injected with saline, fluoxetine 1.75 or3.5 nmol into both amygdala nuclei. At visual inspection,
these injections did not produce any apparent change in thebehavior of the animals. A lack of significant effect wasobserved in the threshold differences (after minus beforetreatments) across the freezing and escape conditions(F1,28=0.05 for LaA and F1,24=0.81 for BLA; PN0.05 in bothcases). Relative to saline controls, fluoxetine in LaA or BLAdid not cause any significant effect on freezing or escapethresholds (F2,28=1.87 for LaA, F2,24=2.48 for BLA; PN0.05in both cases). Moreover, the interaction between treatmentvs. fear condition was not statistically different (F2,28=0.06for LaA and F2,24=0.77 for BLA; PN0.05 in both cases).
Relative to pre-injection, fluoxetine at both doses in BLAand LA reduced post-stimulation freezing (F1,28=16.56 forLaA and F1,24=23.83 for BLA; Pb0.05 in both cases). Relativeto saline controls, fluoxetine reduced post-stimulationfreezing at both doses in BLA and at the highest dose inLaA (F2,28=2.60 and F2,24=2.25, respectively; Pb0.05 inboth cases) (Fig. 2B).
Fig. 3 shows a similar dose dependent prolonging ofcontextual freezing time in both nuclei. There weresignificant effects of treatments (F2,28=14.57 for LaA andF2,24 =4.22 for BLA; Pb0.05 in both cases), conditions(F1,28=559.06 for LaA and F1,24=450.31 for BLA; Pb0.05 inboth cases) and interaction between treatments and condi-tions (F2,28=14.75 for LaA and F2,24=5.73 for BLA; Pb0.05 inboth cases).
4. Discussion
Evidence for the involvement of the dPAG in the generationand elaboration of defensive behavior has been extensivelyreported by behavioral, immunohistochemical and electro-physiological data (see Brandão et al., 1999, 2005 forreviews). These studies have shown that electrical orchemical stimulation of the dPAGproduces fear-like responsessuch as freezing and flight behavior. However, it has beenreported that the dPAG is more than only an output center fordefensive reactions. Indeed, stimulation of the dPAG atintensities causing freezing behavior causes simultaneousFos-labeling of the laterodorsal nucleus of the thalamus,indicating that this structure is also involved in the sensori-motor gating activated by emotional stimuli at this midbrainlevel (Borelli et al., 2005; Ferreira-Netto et al., 2005). Basedon the evidence showing that the amygdala is criticallyinvolved in the regulation of innate and conditioned reactionsto threatening stimuli (Davis et al., 1994; LeDoux, 1994;LeDoux et al., 1990; Rizvi et al., 1991), the question thatarises is to what extent do telencephalic structures regulatethe ascending information coming from the dPAG. Indeed,conjoint Fos expression in the basolateral complex of theamygdala after activation of the neural substrates of aversionin the dPAG supports the existence of a dPAG-amygdala loop(Borelli et al., 2005; Ferreira-Netto et al., 2005).
Lately, we have been investigating the basolateralcomplex of the amygdala as a probable regulator of theunconditioned and conditioned responses organized in thedPAG. In a recent paper we have examined the effects of theinactivation of the BLA/LaA (by enhancing its GABAergicinhibitory tone with local injections of the GABA agonistmuscimol) on conditioned and unconditioned fear elicited byelectrical stimulation of the dPAG. We have found that intra-

722 R.C.R. Martinez et al.
BLA/LaA muscimol decreased the acquisition of fear condi-tioning and the dPAG-post-stimulation freezing (Ruiz-Marti-nez et al., 2006). Interestingly, electrolytic or localinjections of muscimol into the BLA/LaA did not change thefreezing and escape thresholds determined by stepwiseincreases in the current of the electrical stimulation of thedPAG (Oliveira et al., 2004; Ruiz-Martinez et al., 2006).Based on this evidence, we suggested that distinct modula-tory mechanisms in the basolateral complex are recruitedduring the conditioned and unconditioned fear triggered byactivation of the dPAG. Once established that the basolateralcomplex of the amygdala regulates a specific type ofunconditioned fear generated by stimulation of the dPAGwe were also interested to find out whether or not theproposed connection dPAG-amygdala would also be modu-lated by 5-HT-mediated mechanisms since this biogenicamine has also been extensively studied in the neurobiologyof fear and anxiety (Maisonnette et al., 1993; Deakin andGraeff, 1991; Gray and McNaughton, 2000; Graeff, 2004).The present study brings further evidence on this topic, in sofar as the freezing behavior that persists after theinterruption of the dPAG is sensitive to fluoxetine injectionsinto amygdaloid nuclei indicating that a functional dPAG-amygdala connection seems to exist, which is probablyactivated during the processing of aversive information. Itseems that the regulatory role of 5-HT mechanisms of theamygdala on the unconditioned fear interact with GABAergicmechanisms. It has been proposed that 5-HT2 mechanismsactivate GABA interneurons in structures of the brainaversion system (Brandão et al., 1991; Stutzmann andLeDoux, 1999; Graeff, 2004). Indeed, injections of the 5-HT2 receptor antagonist ketanserin into the BLA enhancedthe unconditioned aversive effects elicited by midbraintectum stimulation (Macedo et al., 2007). Moreover, theanxiolytic-like effects induced by 5-HT2A receptor agonist α-methyl-serotonin infused into the basolateral complex ontonic immobility of guinea pigs was reversed by pretreat-ment of ketanserin (Leite-Panissi et al., 2006). An oppositemediation by 5-HT mechanisms in learning process takingplace in the BLA/LaA measured by a context conditioningtest and on the expression of emotional behavior induced bystimulation of the dPAG will be discussed later on.
A number of studies have demonstrated that the 5-HT-mediated mechanisms of the basolateral complex areinvolved in the processing of information during conditionedfear (Davis et al., 1994; LeDoux et al., 1990). The presentdata lend further support to this assumption, in so far asfluoxetine injections within the BLA and LA clearly enhancedthe conditioned aversion to the context in which rats havereceived foot shocks. This finding adds to our currentknowledge that the 5-HT-mediated mechanisms of thebasolateral complex have a crucial role in the conditionedfear (LeDoux et al., 1990; Davis et al., 1994). In contrast withthe 5-HT-mediation of unconditioned fear, enhancement ofthe 5-HT signal in the BLA/LaA results in an increase ofconditioned fear. As intra-amygdala fluoxetine did not causeany significant change in the escape threshold determined byelectrical stimulation of the dPAG it is unlikely that theseeffects of fluoxetine on conditioned fear could be due to thehypolocomotion usually caused by intraperitoneal adminis-tration of this SSRI (Bagdy et al., 2001). Thus, the presentdata go one step further in the neurobiology of fear and
anxiety showing that 5-HT-mediated mechanisms of the BLA/LaA enhance the processing of information in this structureduring conditioned fear. This effect is in apparent conflictwith previous work of our group showing that systemicinjections of fluoxetine did not change the freezing of themoderate fear conditioning (Santos et al., 2006). However, incontrast with intra-amygdala fluoxetine, intraperitonealinjections allow the dug to act at multiple brain structures.Systemic fluoxetine increases extracellular 5-HT levelsaround cell bodies, which in turn activates somatodendritic5-HT1A autoreceptors, which are highly abundant in theraphe nuclei. This leads to an inhibition of the firing of 5-HTneurons and to a subsequent decrease in the 5-HT release atthe nerve terminals of forebrain regions. This effect iscounteracted by inhibition of 5-HT reuptake at the nerveterminals. Thus, the algebraic sum of these actions offluoxetine could have resulted in the reported absence ofeffect on the moderate conditioned fear of this SSRI, whensystemically injected (Santos et al., 2006). The present datacannot discard the possibility that fluoxetine could havefacilitated retrieval of the fear memory, without having aneffect on the fear response itself.
The proposal of an increased functioning of 5-HT networkswithin the basolateral complex may strengthen the acquisi-tion of conditioned aversive information is supported byresults of other studies (LeDoux et al., 1990; Davis et al.,1994; Charney and Deutch, 1996; Davis and Whalen, 2001;Macedo et al., 2007). The fact that the 5-HT2 antagonistketanserin injections within the basolateral complex re-versed the conditioned fear responses suggests that 5-HTacting on 5-HT2 receptors could facilitate the expression ofconditioned fear at this level. This assumption is inagreement with the general idea that 5-HTenhances anxietyin the amygdala by acting on 5-HT2 receptors. In this regard,intra-amygdala injection of ketanserin releases respondingsuppressed by punishment (Petersen and Scheel-Kruger,1984). Moreover, a more recent study using systemicinjections of serotonergic drugs in rats submitted to anelevated T-maze test showed that 5-HT2-mediated mechan-isms have a facilitatory role in the expression of learned fear(Mora et al., 1997).
To study the functional role of the basolateral complex inthe unconditioned fear generated by activation of the neuralsubstrates of aversion in the dPAG, similar injections offluoxetine into this region were also performed in ratssubmitted to the dPAG electrical stimulation procedure. Theinjections of this SSRI into the BLA/LaA did not change theaversive thresholds determined by dPAG stimulation. Thesefindings confirm previous reports using electrolytic lesions orchemical inactivation with muscimol showing that thebasolateral complex does not regulate the defensivebehavior generated at the dPAG level (Oliveira et al., 2004,Ruiz-Martinez et al., 2006). The lack of change in thethresholds for freezing and escape induced by electricalstimulation of the dPAG observed after the microinjection offluoxetine into amygdaloid nuclei is probably linked to thefact that these unconditioned behaviors were generated bydirect stimulation of the dPAG efferents downstream of theamygdala as already suggested by Ruiz-Martinez et al.(2006). Thus, stimulating a structure closer to the motoroutput, as is the case for the dPAG, overrides influences fromupstream structures.

723dPAG, amygdala, serotonin and defensive behavior
One possibility that has been put forward to explain thedifferential role of the basolateral complex of the amygdalain conditioned and unconditioned fear is that serotonergicsystems may be called into play in the setting up of adaptiveresponses aimed at coping with or signaling the presence ofthreatening stimuli. Thus, the signal of the modulatory 5-HTmechanism on defensive behavior will depend on the type ofemotional stimulus triggering the coping reaction. Consonantwith such view, excessive functioning of the basolateralcomplex has been related to general anxiety disorders andthe beneficial effects of SSRIs on panic attacks are thought tobe the result of the depressive action of 5-HTon the activityof this region (Charney and Deutch, 1996; Davis et al., 1994;De Oca et al., 1998; LeDoux et al., 1990).
Altogether, the present data indicate that the 5-HT-mediated mechanisms of the BLA/LaA appear to have oppositeinfluences on the conditioned and unconditioned fear. Whilethese mechanisms appear to facilitate the conditioned fear,they inhibit the unconditioned fear triggered by activation ofthe dPAG. These data support the two dimensional view ofdefense in that anxiety and fear are represented in parallelsystems in thebrain,which are probablymodulatedbyopposingneurochemical mechanisms (McNaughton and Corr, 2004).
Role of the funding source
Funding for this study was provided by FAPESP Grant 02/03705-0; theFAPESP had no further role in study design; in the collection, analysisand interpretation of data; in the writing of the report; and in thedecision to submit the paper for publication.
Contributors
Marcus Lira Brandão designed the study, wrote the protocol and thefirst draft of the manuscript. Raquel Chacon Ruiz-Martinez andAmanda Ribeiro deOliveira collected the data,managed the literaturesearches and analyses, and undertook the statistical analysis. Allauthors contributed to and have approved the final manuscript.
Conflict of interest
We hereby declare that there is no conflict of interest whatsoever forthe authors of thismanuscript (Marcus Lira Brandão, AmandaRibeiro deOliveira and Raquel Chacon Ruiz-Martinez) including any financial,personal or other relationships with other people or organizations thatcould inappropriately influence, or be perceived to influence thiswork.
Acknowledgements
This research was supported by FAPESP (Proc No. 02/03705-0). R.C.R.Martinez was the recipient of a doctor scholarship from CNPq andA.R. Oliveira holds a doctor scholarship from FAPESP.
References
Azmitia, E.C., Segal, M., 1978. An autoradiographic analysis of thedifferential ascending projections of the dorsal and medianraphe nuclei in the rat. J. Comp. Neurol. 179, 641–667.
Bagdy, G., Graf, M., Anheuer, Z.E., Modos, E.A., 2001. Anxiety-like effects induced by acute fluoxetine, sertraline or m-CPP
treatment are reversed by pretreatment with the 5-HT2Creceptor antagonist SB-242084 but not the 5-HT1A recep-tor antagonist WAY-100635. Int. J. Neuropsychopharmacol. 4,399–408.
Blier, P., 2001. Pharmacology of rapid-onset antidepressant treat-ment strategies. J. Clin. Psychiatry 62, 12–17.
Borelli, K.G., Ferreira-Netto, C., Coimbra, N.C., Brandão, M.L.,2005. Fos-like immunoreactivity in the brain associated withfreezing or escape induced by inhibition of either glutamic aciddecarboxylase or GABAA receptors in the dorsal periaqueductalgray. Brain Res. 105, 100–111.
Brandão, M.L., Lopez-Garcia, J.A., Roberts, M.H.T., 1991. Elec-trophysiological evidence for excitatory 5-HT2 and depressant5-HT1A receptors on neurons of the rat midbrain tectum. BrainRes. 556, 259–266.
Brandão, M.L., Cardoso, S.H., Melo, L.L., Motta, V., Coimbra, N.C.,1994. Neural substrate of defensive behavior in the midbraintectum. Neurosci. Biobehav. Rev. 18, 339–346.
Brandão, M.L., Anseloni, V.Z., Pandóssio, J.E., De Araújo, J.E.,Castilho, V.M., 1999. Neurochemical mechanisms of the defen-sive behavior in the dorsal midbrain. Neurosci. Biobehav. Rev. 23,863–875.
Brandão, M.L., Borelli, K.G., Nobre, M.J., Santos, J.M., Albrechet-Souza, L., Oliveira, A.R., Martinez, R.C., 2005. Gabaergicregulation of the neural organization of fear in the midbraintectum. Neurosci. Biobehav. Rev. 29, 1299–1311.
Burghardt, N.S., Sullivan, M., McEwen, B.S., Gorman, J.M., LeDoux,J.E., 2004. The selective serotonin reuptake inhibitor citalopramincreases fear after acute treatment but reduces fear withchronic treatment: a comparison with tianeptine. Biol. Psychi-atry 55, 1171–1178.
Castilho, V.M., Brandão, M.L., 2001. Conditioned antinociceptionand freezing using electrical stimulation of the dorsal periaque-ductal gray or inferior colliculus as unconditioned stimulus aredifferentially regulated by 5-HT2A receptors in rats. Psychophar-macology 155, 154–162.
Charney, D.S., Deutch, A., 1996. A functional neuroanatomy ofanxiety and fear: implications for the pathophysiology andtreatment of anxiety disorders. Crit. Rev. Neurobiol. 10, 419–446.
Davis, M., Whalen, P.J., 2001. The amygdala: vigilance and emotion.Mol. Psychiatry 6, 13–34.
Davis, M., Raiunnie, D., Cassell, M., 1994. Neurotransmission in therat amygdala related to fear and anxiety. TINS 17, 208–214.
De Oca, B.N., DeCola, J.P., Maren, S., Fanselow, M.S., 1998.Distinct regions of the periaqueductal gray are involved in theacquisition and expression of defensive responses. J. Neurosci.18, 3426–3432.
Deakin, J.F.W., Graeff, F.G., 1991. 5-HTand mechanisms of defense.Psychopharmacology 5, 305–315.
Fanselow, M.S., 1991. The midbrain periaqueductal gray as acoordinator of action in response to fear and anxiety. In:Depaulis, A., Blander, R. (Eds.), The Midbrain PeriaqueductalGray Matter: Functional, Anatomical and ImmunohistochemicalOrganization. Plenun, New York, pp. 151–173.
Ferreira-Netto, C., Borelli, KG., Brandão, M.L., 2005. Neuralsegregation of Fos-protein distribution in the brain followingfreezing and escape behaviors induced by injections of eitherglutamate or NMDA into the dorsal periaqueductal gray of rats.Brain Res. 1031, 151–163.
Goldstein, B.J., Goodnick, P.J., 1998. Selective serotonin reuptakeinhibitors in the treatment of affective disorders-III. Tolerability,safety and pharmacoeconomics. J. Psychopharmacol. 12, S55–S87.
Graeff, F.G., 1990. Brain defense systems and anxiety. In: Roth, M.,Burrow, G.D., Noyes, R. (Eds.), Handbook of Anxiety, vol. 3.Elsevier Science Publishers, New York, pp. 307–354.
Graeff, F.G., 2004. Serotonin, the periaqueductal gray and panic.Neurosci. Biobehav. Rev. 28, 239–259.

724 R.C.R. Martinez et al.
Gray, J.A., McNaughton, N., 2000. Fundamentals of the septo-hippocampal system, In: Gray, J.A., McNaughton, N. (Eds.),Neuropsychology of Anxiety, 2nd edn. Oxford University Press,Oxford, UK.
Iversen, S.D., 1984. 5-HT and anxiety. Neuropharmacology 23,1553–1560.
Killcross, S., Robbins, T.W., Everitt, B.J., 1997. Different types offear-conditioned behaviour mediated by separate nuclei withinamygdala. Nature 388, 377–380.
Krettek, J.E., Price, J.L., 1978. A description of the amygdaloidcomplex in the rat and cat with observations on intra-amygdaloidaxonal connections. J. Comp. Neurol. 178, 255–280.
LeDoux, J.E., 1994. The amygdala: contributions to fear and stress.Semin. Neurosci. 6, 231–237.
LeDoux, J.E., Cicchetti, P., Xagoraris, A., Romanski, L.M., 1990. Thelateral amygdaloid nucleus: sensory interface of the amygdala infear conditioning. J. Neurosci. 10, 1062–1069.
Leite-Panissi, C.R.A., Ferrarese, A.A., Terzian, A.L., Menescal-de-Oliveira, L., 2006. Serotoninergic activation of the basolateralamygdala and modulation of tonic immobility in guinea pig. BrainRes. Bull. 69, 356–364.
Macedo, C.E., Castilho, V.M., Souza Silva, M.A., Brandão, M.L., 2002.Dual 5-HT mechanisms in basolateral and central nuclei ofamygdala in the regulation of the defensive behavior induced byelectrical stimulation of the inferior colliculus. Brain Res. Bull.59, 189–195.
Macedo, C.E., Cuadra, G., Molina, V., Brandão, M.L., 2005. Aversivestimulation of the inferior colliculus changes dopamine andserotonin extracellular levels in the frontal cortex: modulationby the basolateral nucleus of amygdala. Synapse 55, 58–66.
Macedo, C.E., Martinez, R.C.R., Albrechet-Souza, L., Molina, V.A.,Brandão, M.L., 2007. 5-HT2 and D1 mechanisms of thebasolateral nucleus of the amygdala enhance conditioned fearand impair unconditioned fear. Behav. Brain Res. 177, 100–108.
Maisonnette, S.S., Morato, S., Brandão, M.L., 1993. Role ofresocialization and of 5-HT1A receptor activation on theanxiogenic effects induced by isolation in the elevated plusmaze test. Physiol. Behav. 54, 753–758.
McDonald, A.J., 1992. Projection neurons of the basolateral amygdala:a correlative Golgi and retrograde tract tracing study. Brain Res.Bull. 2, 179–185.
McNaughton, N., Corr, P.J., 2004. A two-dimensional neuropsychol-ogy of defense: fear/anxiety and defensive distance. Neurosci.Biobehav. Rev. 28, 285–305.
Mora, P.O., Netto, C.F., Graeff, F.G., 1997. Role of 5-HT2A and 5-HT2Creceptor subtypes in the two types of fear generated by theelevated T-maze. Pharmacol. Biochem. Behav. 58, 1051–1057.
Oliveira, L.C., Nobre, M.J., Brandão, M.L., Landeira-Fernandez, J.,2004. Role of amygdala in conditioned and unconditioned feargenerated in the periaqueductal gray. NeuroReport 15, 2281–2285.
Ottersen, O.P., 1981. Afferent connections to the amygdaloidcomplex of the rat with some observations in the cat. III.Afferents from the lower brain stem. J. Comp. Neurol. 202,335–356.
Pasini, A., Tortorella, A., Gale, K., 1996. The anticonvulsant actionof fluoxetine in substantia nigra is dependent upon endogenousserotonin. Brain Res. 724, 84–88.
Paxinos, G., Watson, C., 2005. The Rat Brain in StereotaxicCoordinates, 5th ed. Academic Press, San Diego, CA.
Petersen, E.N., Scheel-Kruger, J., 1984. Anticonflict effects of 5-HTantagonists by intraamygdaloid injection. Abstr 14th CINP Cong.,p. 654.
Rizvi, T.A., Ennis, M., Behbehani, M.M., Shipley, M.T., 1991.Connections between the central nucleus of the amygdala andthe midbrain periaqueductal gray: topography and reciprocity.J. Comp. Neurol. 303, 121–131.
Ruiz-Martinez, R.C., Oliveira, A.R., Brandão, M.L., 2006. Condi-tioned and unconditioned fear organized in the periaqueductalgray are differentially sensitive to injections of muscimol intoamygdaloid nuclei. Neurobiol. Learn. Mem. 85, 58–65.
Santos, J.M., Gárgaro, A.C., Oliveira, A.R., Masson, S., Brandão, M.L.,2005. Pharmacological dissociation of moderate and high contex-tual fear as assessed by freezing behavior and fear-potentiatedstartle. Eur. Neuropsychopharmacol. 15, 239–246.
Santos, J.M., Martinez, R.C., Brandão, M.L., 2006. Effects of acuteand subchronic treatments with fluoxetine and desipramine onthe memory of fear in moderate and high-intensity contextualconditioning. Eur. J. Pharmacol. 542, 121–128.
Silva, R.C., Cruz, A.P., Avanzi, V., Landeira-Fernandez, J., Brandão,M.L., 2002. Distinct contributions of median raphe nucleus tocontextual fear conditioning and fear-potentiated startle. NeuralPlast. 9, 233–247.
Silva, R.C.B., Gárgaro, A.C., Brandão, M.L., 2004. Differentialregulation of the expression of contextual freezing and fear-potentiated startle by 5-HT mechanisms of the median raphenucleus. Behav. Brain Res. 151, 93–101.
Stutzmann, G.E., LeDoux, J.E., 1999. GABAergic antagonists blockthe inhibitory effects of serotonin in the lateral amygdala: amechanism for modulation of sensory inputs related to fearconditioning. J. Neurosci. 19, 1–4.
Vertes, R.P., 1991. A PHA-L analysis of ascending projections of thedorsal raphe nucleus in the rat. J. Comp. Neurol. 313, 643–668.
Vianna, D.M.L., Graeff, F.G., Landeira-Fernandez, J., Brandão, M.L.,2001a. Lesion of the ventral periaqueductal gray reducesconditioned fear but does not change freezing induced bystimulation of the dorsal periaqueductal gray. Learn. Mem. 8,164–169.
Vianna, D.M.L., Graeff, F.G., Brandão, M.L., Landeira-Fernandez, J.,2001b. Defensive behavior evoked by electrical stimulation ofthe periaqueductal gray: comparison between dorsolateral andventrolateral region. NeuroReport 12, 4109–4113.
Vianna, D.M.L., Landeira-Fernandez, J., Brandão, M.L., 2001c.Dorsolateral and ventral regions of the periaqueductal graymatter are involved in distinct types of fear. Neurosci. Biobehav.Rev. 25, 711–719.
Vianna, D.M.L., Borelli, K.G., Ferreira-Netto, C., Macedo, C.E.,Brandão, M.L., 2003. Fos-like immunoreactive neurons followingelectrical stimulation of the dorsal periaqueductal gray atfreezing and escape thresholds. Brain Res. Bull. 62, 179–189.
Wise, C.D., Berger, B.D., Stein, L., 1972. Benzodiazepines: anxiety-reducing activity by reduction of serotonin turnover in the brain.Science 177, 180–183.