serology of orthopoxvirus cameli infection in dromedary camels: analysis by elisa and western...
TRANSCRIPT

~ Pergamon Comp. lmmun. Microbiol. infect. Dis. Vol. 19, No. 1, pp. 65-78, 1996 Copyright © 1996 Elsevier Science Ltd
0147-9571(95)00023-2 Printed in Great Britain. All rights reserved 0147-9571/96 $15.00 + 0.00
S E R O L O G Y O F O R T H O P O X V I R U S CAMEL1 I N F E C T I O N I N D R O M E D A R Y C A M E L S :
A N A L Y S I S B Y E L I S A A N D W E S T E R N B L O T T I N G
S. M. A Z W A I , I t S. D. C A R T E R , 1. Z. W O L D E H I W E T l and U. W E R N E R Y 2
~Department of Veterinary Pathology and Department of Veterinary Clinical Science and Animal Husbandry, University of Liverpool, P.O, Box 147, Liverpool, L69 3BX, U.K. and
2Central Veterinary Research Laboratory, P.O. Box 597, Dubai, U.A.E.
(Received for publication 24 April 1995)
Abstract--An enzyme-linked immunosorbent assay (ELISA) was developed, together with a Western blotting technique, for the detection of total and IgG and IgM antibodies to camelpox virus (Orthopoxvirus cameli) in camel (Camelus dromedarius) sera and for identifying the seroreactive antigens of the virus. A total of 520 camels from different regions in Libya were tested. The overall seropositivity rate in the examined herds was 9.8%, and varied between herds from 0 to 30%. Two viral antigenic determinants (31 and 35 kDa) were shared by the Western blotting patterns of all the positive camel sera tested. The developed ELISA assay showed ability to differentiate between orthopox and parapoxvirus infections in camels. It is considered that the ELISA technique is justified for serodiagnosis of camelpox in the camel and could be easily modified and usefully applied to other species at risk of poxvirus infection.
Key words: Camel, dromedary, Camelus dromedarius, Orthopoxvirus cameli, camelpox, contagious ecthyma, ELISA, monoclonal antibodies, Western blotting, Libya.
R6sum~-Un essai ELISA fut d6velopp6 ainsi q'une technique immunoblot, afin de mesurer les immunoglobulines totales et les anticorps IgG et IgM contre le virus variole (Orthopoxvirus cameli) des dromadaires (Camelus dromedarius), et d'identifier les antig6nes r6actifs du virus. 520 dromedaires en tout des regions diverses de la Libye furent examin6s. Le taux compl6t de la r6action positive dans les troupeaux etudi6s rut 9.8%, en variant entre les troupeaux de 0 fi 30%. Deux d6terminants antig6niques (31 et 35 kDa) furent partag6s par les profils immunoblots de tousles serums positifs examin6s. L'essai ELISA d6velopp6 se montra capable de distinguer entre les virus orthopox et parapox chez les dromadaires. On estime que la m6thode ELISA se justifie dans la diagnose du variole chez les dromadaires et pourrait s'adapter aux autres esp6ces pr6dispos6s l'infection.
Mots-clefs: Dromadaires, Camelus dromedarius, Orthopoxvirus cameli, Ecthyma contagiosum, ELISA, Immunoblot, Libye.
I N T R O D U C T I O N
As a result of frequent droughts and extending arid and semi-arid zones, there has been increased attention paid to camel health and welfare. Camels are raised for their milk, meat, fibre, hide and transport. The likely world population is about 20 million camels [1], mainly distributed in Africa, Asia and the Middle East. One of the most common important
*Author for correspondence. tPresent Address: Department of Veterinary Microbiology, E1-Fateh University, P.O. Box 13180, Tripoli, Libya.
65

66 S . M . A z w a i et al.
infectious and contagious diseases diagnosed in these old-world camelids is camelpox (caused by Orthopoxvirus cameli) which, from an economic point of view, is possibly the most important remaining orthopoxvirus disease. To herdsmen, camelpox is a well known and feared disease. Its considerable economic importance is due to the high morbidity, a relatively high mortality in younger animals, loss of condition and weight in all ages, as well as a fall in milk production. Camelpox may also be important because of potential zoonotic problems; camelpox virus is difficult to distinguish from smallpox virus and it occurs in areas where smallpox vaccination has now ceased. Although rare, pox-like lesions associated with camelpox have been described in man [2-4] and confirmed serologically [5]; it has also been suggested that camelpox virus could be as pathogenic for man as cow pox and monkey pox [6].
Camelpox virus (CPV) is one of the least studied members of orthopoxviruses, and the publications available are contradictory [7]. Although the causative virus was not isolated until 1970 [8], camelpox has long been recognized and is widespread in many camel-raising countries and infections are commonly encountered in the herds of the nomadic pastoralists in the semi-desert zones of Africa [5, 9-13], the Middle East [14-18], and the former USSR [6, 8] and although no exact data is available, some Asian countries (India, Pakistan, Afghanistan) are regarded as the main endemic areas of camelpox [2, 19].
Adequate surveillance for camelpox virus infection in migrating camels, particularly in large populations, is dependent on a sensitive and specific assay that is easy to perform and suited for mass screening. Poxvirus antibodies can be detected in animal sera much more frequently than poxviruses can be isolated from them [20].
Conventional serological tests (haemagglutination, haemagglutination inhibition, neutral- ization, complement fixation, fluorescent antibody) have been used to detect camelpox virus antibodies [6, 10, 15, 18, 21, 22], but have a number of restrictions. The simplicity of ELISA makes it suitable for retrospective studies and also for epidemiological investigations of animal sera, and even for field use. Marennikova et al. [23], showed that ELISA was more sensitive than virus neutralization, haemagglutination inhibition, passive haemagglutination, and indirect fluorescent antibody tests for detecting antibodies to vaccinia, whitepox and monkeypox viruses. Furthermore, the most commonly used test (virus neutralization) for diagnosis of orthopoxvirus infections, does not measure antibodies to envelope antigens, which are important in pathogenesis [24]. In any such study, it is necessary to consider the test specificity so as to differentiate between antibodies to different poxviruses (orthopox and parapoxviruses) which are known to cause similar pox-like lesions in camels [4, 25, 26].
We have previously described the structure of circulating IgM, IgG and IgG subclasses in the camel [27], have prepared monoclonal antibodies against both classes [28] and described ELISA assays for the detection of antibodies to parapoxvirus in camels [29]. The purpose of this study was to examine sera of Camelus drornedarius for antibodies to
Table 1. Serum samples from apparently normal camel herds in Libya
Herd Location No. of camels
A B C D E F
Total
Bir al Ghanam (100 km south-west of Tripoli) 103 Ghariyan (85 km south-west of Tripoli) 11 A1 Assah (150 km south-west of Tripoli) 171 Bin Ulid (160 km south-east of Tripoli) 20 Maknusah (1250 km south of Tripoli) 127 Wadi EI-Arial (1400 km south of Tripoli) 88
520

Serology of camelpox infection in dromedary camels 67
0. cameli. We demonstrated, for the first time, both evidence of camelpox infections in dromedaries in Libya and also the first analysis of camel antibodies to naturally acquired camelpox infection. This study describes a sensitive ELISA to detect the antibody response to the camelpox virus and demonstrates the specificity by Western blotting.
MATERIALS AND METHODS
Animals
A total of 520 camels (C. dromedarius) of mixed ages, both sexes and different localities from Libya were sampled (Table 1). No camels showed signs of any clinical disease at the time of blood sampling, except 39 camels at area (A) which showed pox-like lesions at the time of blood sampling and confirmed, serologically as contagious ecthyma [29]. The camel population of Libya has never been vaccinated against camelpox.
Blood samples
Blood was collected by jugular venepuncture from camels restrained in the sitting position. The samples were allowed to clot at room temperature, centrifuged at 3000 rpm for 15 min and the serum was stored in aliquots at -20°C until tested.
Antigen
Antigen source. A cell-culture adapted strain of O. cameli (Ducapox 298/89) is maintained by the Central Veterinary Research Laboratory (CVRL), Dubai, U.A.E. Originally the virus was isolated from severe pox-like lesions in a young camel and later identified as 0. cameli. The virus was initially cultured and then passaged 96 times in Vero cells as described earlier [I 7]. This isolate is currently being used for the production of a camelpox vaccine (Dubai camelpox vaccine; [17]).
Antigen preparation for ELISA. Virus for ELISA antigen was prepared as described by Thomas et aL [30] and Bell et al. [31] with some modifications [29]. Briefly, the virus suspension was sonicated (Rapidis 150, Ultrasonics Ltd. U.K.) intermittently on ice for 30 s with 15 s of rest, for a total sonication time of 2 min. After centrifugation at 1000g for l0 min, supernates were collected. The virus was pelleted by ultra-centrifugation at 45,000 g for 60 min at 4°C and the pellet resuspended in phosphate buffered saline (PBS). The viral suspension was stirred and NaC1 was added to 330 mM, followed by polyethylene glycol (PEG, mol. wt 6000 BDH) to a concentration of 7% and stirred at 4°C overnight. The suspension was centrifuged at 3000g for l0 min at 4°C and the pellet washed twice in 15 mM NaC1. Viral proteins were obtained by disruption with freezing and thawing in 15 mM NaCl and stripping of the surface proteins by 1% non-ionic detergent (Nonidet P40; Sigma) treatment at 37°C for 3 h, followed by freezing and thawing and centrifugation at 3000g for l0 min at 4°C. The supernatant containing viral antigens was collected and extensively dialysed against several changes of PBS. The protein concentration was determined as described by Lowry et al. [32]. Aliquots were stored at
_ 20oc. Antigen preparation for Western blotting. Purified virus particles were denatured, by a
modification of the method of Brik and Koepsell [33]. Briefly, antigens were solubilized by boiling for 5 min at 100°C in 62.5 mM Tris-HC1, pH 6.8, 2% (w/v) sodium dodecyl sulphate (SDS; BDH) and 5% (v/v) 2-mercaptoethanol (2-ME; Sigma). Aggregated material was removed by centrifugation.

68 S .M. Azwai et al.
Positive and negative control camel sera
Camel sera, which were collected from camels proven to be infected with camelpox (0. cameli was isolated from them) from, Dubai, were used as positive controls. Negative camel sera were obtained from camels which were born and raised in Chester Zoo, U.K.
Ant&era to camel immunoglobulins
Polyclonal antisera. Rabbit antiserum against camel IgG was generated and its specificity shown by Western blotting [27].
Monoclonal antibodies. Monoclonal antibodies reacting with camel IgG and IgM heavy chains were produced and their specificities were determined by ELISA and Western blotting [28]. The working dilutions of these reagents were obtained by titrating them against purified camel immunoglobulins in a direct ELISA.
Enzyme-linked immunosorbent assay (ELISA)
Non-activated, 96-weU, microtitre ELISA plates (Dynatech) were coated with the prepared camelpox virus antigens at 1 #g/ml in PBS pH 7.2, (the optimal antigen dilution was determined by titrating known positive and negative camel sera with serial antigen dilutions), the ELISA plates were incubated in a moist chamber (100% humidity) at 37°C for 1 h, then overnight at 4°C. Unbound antigen was washed off with PBS, three times, and camel sera to be tested were diluted 1/100 in 1% bovine serum albumin [BSA, Sigma/ PBS/Tween 20 (0.05%)] [The working serum dilution (1/100) was the highest dilution found to allow a clear distinction between positive and negative control sera] and added in duplicates. Positive and negative camel sera were treated similarly and included in duplicates in each test plate as controls. After incubation at 37°C for 1 h and three washes with PBS/Tween 20, antibodies to camel immunoglobulins were applied (rabbit anti-camel IgG at 1/10,000, biotinylated mouse monoclonal anti-camel IgG or IgM at 1/1000) and plates were incubated at 37°C for 1 h. Following washing with PBS/Tween, bound antibodies were detected with either alkaline phosphatase-conjugated goat anti-rabbit IgG (Sigma) at 1/1000 or extravidin alkaline phosphatase (Sigma) at 1/5000 as appropriate. Following incubation and washing as stated above, bound conjugate was detected with substrate (p-nitrophenyl phosphate; Sigma) diluted in glycine buffer pH 10.4 at 1 mg/ml. An ELISA reader (Titertek) was used to measure the absorbance at 405 nm. Results were expressed on a matrix 0-9 scale, setting the known positive sample as 9. Seropositivity was calculated as figures above the mean + 2 SD of the ELISA data from the whole population of camels.
SDS-PAGE
Polyacrylamide gel electrophoresis was carried out in a vertical slab mini-gel electro- phoresis system (2050 Midget; Pharmacia) with 0.75 mm thick, 8.5 × 6 cm gels using a trough comb, under reducing conditions as described by Laemmli [34]. Denatured camelpox virus preparation was loaded onto a 12% (w/v) acrylamide resolving gel via a 4% (w/v) stacking gel. Separation was carried out at a constant 30 mA/gel. Standard molecular weight markers, a mixture of proteins ranging from 6.9-202 kDa (Bio-Rad; Kaleidoscope prestained standards), were used to calculate the molecular weight of camelpox virus proteins. Gels were stained with silver stain (Pharmacia Biotech) for protein detection, or processed for Western blotting.

Serology of camelpox infection in dromedary camels 69
Western blotting
The electrophoretic transfer of proteins, after their separation in SDS-PAGE gels, to nitrocellulose membrane (0.20 pm) was performed in a semi-dry blotter (2117 Multiphor II, Pharmacia) with a discontinuous buffer, in a procedure similar to that described by Azwai et al. [29]. The transfer was performed at 18°C at a constant current (42 mA/gel)
Positivity (%) (11.7%) (0%) (9.9%) (30%) (7.1%) (8%)
i<
<
,,.]
O i i
O I O i
e l
O i O i
0 0 0 9 0
i l O O t
i
l i i lO
i O
i
t l l i
( l e
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ° . . . . . . . . . . . . . . . . . . . . . . .
l i i l •
O l l O i l l O l O i l OO
O i l i i
i l i l i l I l i i l i l i l l i i l i l i
l l l l l l l l i l l l i l l i l t
O O O l O O O O O i O O l i l i O O i O
0 0 0 0 0
O O * O i i i l O O O O i O i • O O l O O O O O O i
l i O l O l l
O O O I
l i i O
i l i tttthii O i l i l i l l O
i O O l i
!!!!!!It!!!
l l i l l l
0 0 0 0 0
OOilOi
O O i i O i i i O O e O O i O O i l i O O i
OI 000000000
. l l l l i l l l l .
0 0 0 0 9 0 0 0 0 • 0 0 0 9 0 0 0 0 0 0 0
O 0 0 O O 0 0 0 0 0 0 g O 0 0 0 0 0 i O 0 0 O 0 0 i O O O O 0 0 0
~ t g 34 t ~g~ g~ t 44 ~t: ~g 38 ~ t t O 0 0 e O O 0 0 8 0 O O0 O O O O O O O I O O O 0 0 0 0 t O O O O O 0 0 0 0 O
(A) (B) (C) (D) (E)
Camel herds
Fig. I. Total antibody ~sponse to camelpox virus in diffe~nt camel herdsin Libya.
l i
i
i i O
0 0 0 0 0
0OO0000
.°IIIII io°°
O066
0 0 0 0 0 0 0 * 0 0 0
I I I 30 IIII OOiiO000iOQ
(F)

70 s.M. Azwai et al.
for 1 h. To identify the protein bands on the nitrocellulose membrane, one strip was cut from the border including the molecular weight marker and was stained for 5 min in amido black stain. Unoccupied sites on nitrocellulose membranes were blocked by incubation in 5% BSA for 1 h at room temperature, followed by washing three times (10 min) in PBS/ Tween 20 (0.05%). The nitrocellulose sheet was cut into strips and each strip was incubated
Positivity (%) (10.7%) (0%) (9.4%) (30%) (7.1%) (8%)
5
E <
m 4
nl[t 0 O
OOO0
O0
0 0 9
*OOOO
0 0 9 0 9 9
9 0 O O 0 0 0 0 0 0 & O O
9 9 0
O O O * 9 * O O O 9 * O O O O
0 0 0 9 9
9 * 9 9 0 9 0 0 0 9 9
:11 46 1::1 OOOOOOOOOOO
qPo
QO
OOUO00
99
O0
O0
Oal)
O0
O(pq[a
0 0
OOql0
OOOO9
(B)
4 0 0 0 0 0 O
OQQO 9 0 O O O 9 0 0 0
, . , | | | | . , , .
, , , 1 5 , , , ,
. I I I I I I I I I .
OOOoooqNa
9 0 0 9 9 9 0 0 0 0 9
:1178 1111 O 0 0 0 0 0 0 0 1 0 0
(¢)
OOO
O 0 0 1
0 . 0 0 0 0 0 0 . 0 9 9 9 9 0 9 9 9 0 9
6 Q O 0 0 *
• , , , l l l l l l l l g ,
Q6 O O 0 1
0 D B 9 0 0 9 9 0 0 9
Ill: s7 :::: e o e e e o o o o o o o o e o o e
(D) (E)
Camel herds
00
o e
0 9
9 9 9 ~ o
o o 9 o 9 9 9 Q
, , | I | | I | ~ , ,
e g O O Q 0 9
O e 9 9 O O Q O 0 0 0
1:1 39 :::: D Q O * O * Q O 0 0 *
(F)
Fig. 2. Camelpox-specific IgG antibody response in different camel herds in Libya.
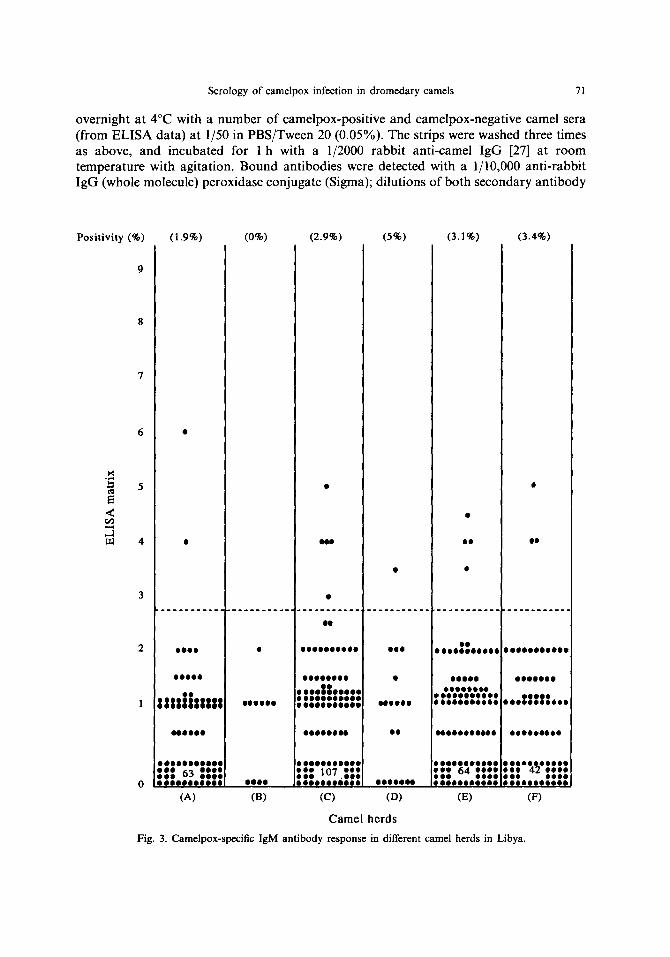
Serology of camelpox infection in dromedary camels 71
overnight at 4°C with a number of camelpox-positive and camelpox-negative camel sera (from ELISA data) at 1/50 in PBS/Tween 20 (0.05%). The strips were washed three times as above, and incubated for 1 h with a 1/2000 rabbit anti-camel IgG [27] at room temperature with agitation. Bound antibodies were detected with a 1/10,000 anti-rabbit IgG (whole molecule) peroxidase conjugate (Sigma); dilutions of both secondary antibody
Positivity (%) (1.9%) (0%) (2.9%) (5%) (3.1%) (3.4%)
5
<
4
2
000
O
O0 O( °
oq)
O00OO0
O 0 0 0 0 0 0 0 e O
0 9 0 0 0 0 0 0
o o o o
o o o o o
smilm, ° 0 ° O ° O
0 0 0 ~ 0 0 0 0 0 0 0
O 0 0 0 0 0 0 0 0 ~ O
(A)
o o o o ~ o o o o e o o o o 9 o o o o o o o o 0 4 o o 0 o 0 o o
OOO
e
o g o B o o
o o 40OOOOO0
O 0 0 0 0 0 0 0 O O 0 ::: 1o7:::
0 9 O 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 O 0 0 0 0 0 0 0
O 0 0 0 ° O O ° O O O O 0 0 0 0 0 0 0 0 0 0
O 0 0 0 0 0 0 0 0 0 0
OOOQ 0 6 0 0 0 0 Q O 0 0 0 O 0 0 0 0 e O
(B) (C) (D) (E)
O 0 0 0 0 0 0 0 0 0 0 : : : 64 : : : : O ° O 0 ° ° O 0 0 0 °
g O O d 0 0 0 0 0 0 0
0 0 0 0 0 0 0
, , ,II|SS,o,
QOOOOOe00
0 0 0 0 ° 9 0 * O O 0 0 ° O 0 0 0 0
(F)
Camel herds
Fig. 3. Camelpox-speeific IgM antibody response in different camel herds in Libya.

72 S.M. Azwai et al.
and conjugate were previously optimized by a dot blot technique. Development of the solid phase reactants was visualized with 3,3-diaminobenzidine tetrahydrochloride (DAB; Sigma).
Statistical analysis
Correlation coefficients were determined by linear regression analysis.
RESULTS
ELISA
The results of the ELISA assays are shown in Figs 1, 2 and 3. Camelpox infection appears to have occurred in all the herds examined except for a small herd (B). These findings show that there is a high prevalence of antibody to camelpox in herds examined regardless of their geographical location. This, together with the historical (none documented) and serological evidence, suggests that the disease is enzootic.
The overall antibody response to camelpox virus, measured by the rabbit anti-camel IgG, in all camels tested, gave a mean seropositivity rate of 9.8% (51/520), the camelpox- specific IgG and camelpox-specific IgM prevalence were 9.4 and 2.9% respectively. The frequency of raised antibody in camels in other herds varied from 0 (herd B) to 30% (herd D). In herd A, the seropositivity rate to O. cameli was 11.7% (12/103); among these, 3 camels were previously clinically diagnosed and serologically confirmed as being infected with parapox virus [29]. All 3 camels showed raised level of camelpox-specific IgG, but none had any detectable levels of camelpox-specific IgM. Linear regression analysis showed there was a good correlation between the ELISA using polyclonal antisera (rabbit anti- camel IgG) and that using monoclonal antibodies (mouse anti-camel IgG) (n = 520, r = 0.951, P < 0.001). There was a lesser correlation between the rabbit anti-camel IgG and mouse monoclonal anti-IgM (n--520, r--0.581, P <0.001); also the correlation was less between IgG and IgM responses, as detected by monoclonal antibodies (n --- 520, r = 0.566, P < 0.001).
S D S - P A G E
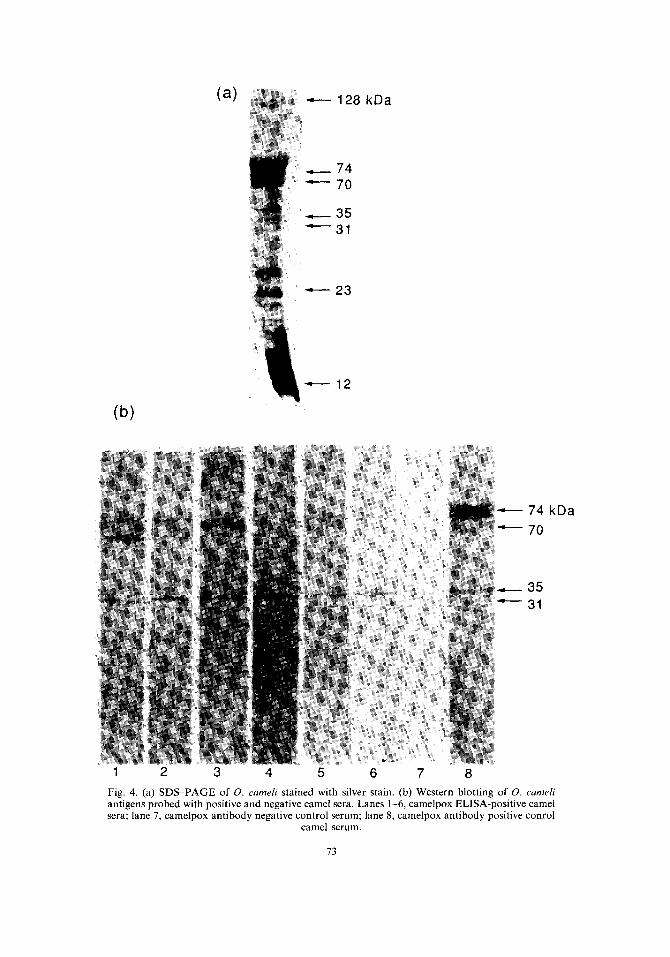
O. cameli gave a complex pattern of at least 20 polypeptides, when the gels were stained by silver stain, with a molecular weight of 12-128 kDa [Fig. 4(a)].
Western blotting
In order to identify the structural antigens which reacted with the ELISA-seropositive camel serum samples, immunoblotting experiments were performed and results are shown in Fig. 4(b).
Reactivity of serum antibodies from seropositive camel sera with virus polypeptides showed that, of the 20 virion polypeptides identified by SDS-PAGE, there were four major reacting viral antigens detected by most of the positive sera; these were 31, 35, 70 and 74 kDa. Some seropositive sera only bound two bands, at 31 and 35 kDa bands; these two bands have been only found in the electrophoretic profile of O. cameli and not in other orthopoxviruses [35]. These 31 and 35 kDa bands were bound by all ELISA-positive camel sera. Other bands were detected but were variable infrequency.
When a number of the ELISA-negative camel sera, chosen at random from different areas, were tested, their immunoblotting profiles revealed no binding of virus proteins.
(a) 128 kDa
74 70
35 31
23
(b)
12
74 kDa
70
35 31
1 2 3 4 5 6 7 8
Fig. 4. (a) SDS-PAGE of O. cameli stained with silver stain. (b) Western blotting of O. cameH antigens probed with positive and negative camel sera. Lanes 1-6, camelpox ELISA-positive camel sera; lane 7, camelpox antibody negative control serum; lane 8, camelpox antibody positive conrol
camel serum.
73

74 S.M. Azwai et al.
Area A was particularly interesting because contagious ecthyma had been diagnosed among its camel herd [29]. The sera of the three camels with dual infection with orthopox and parapox virus were Western blotted against O. cameli; all three gave a similar binding profile where they detected the four major polypeptides (31, 35, 70 and 74 kDa). These sera have been previously examined for parapox virus, where two of them detected three antigenic determinants of parapox virus of 22, 28 and 40 kDa and one detected 22 and 40 kDa bands [29]. When some of the orf-seropositive but pox-seronegative camel sera from area A were tested for binding to camelpox virus antigens, none of them detected any of the camelpox virus antigens, showing a lack of serological cross reaction between camelpox and orf infection.
DISCUSSION
The study of camelpox has been neglected because it presents no apparent risk to man or domestic animals in developed countries, but camelpox is a regular cause of camel morbidity under the traditional nomadic management systems and an incidence of 50% has been reported in camels in Somalia [5] and up to 100% in Kenya [10]. The mortality rate is usually lower (10-50%) and appears to be influenced by the presence ofintercurrent diseases (notably trypanosomiasis), stress, the age structure and the general nutritional status of the herd at the time of the outbreak as well as the virulence of the virus strain involved [4, 10, 18]. The requirements of the camel owners for the camel milk, which is an important component of their diet, frequently leave the camel calves undernourished and susceptible to diseases in the neonatal period [36] and the highest death rate is during the first year of life [5]. Outbreaks of camelpox have followed seasonal movements for grazing and watering purposes, which produce some mixing of the herds and the recent introductions of new camels into a herd; the role of a possible insect spread must also be considered because other poxviruses may be transmitted in this manner.
This work is the first comprehensive sero-epidemiological study to determine the extent of poxvirus infection in camels and provides data for prevalence of infection in Libya. ELISA have been developed for the serological detection of many infectious organisms, and this is the first description of an assay for O. cameli which detects camelpox-specific IgG and IgM. The Western blotting data described here confirms the specificity of the ELISA assay. The specificity of an ELISA is, to a large degree, determined by the quality of the antigen used in the solid phase; therefore, we chose to use purified camelpox virus antigens which gave exceptionally low background levels with the negative control camel sera. Furthermore, purified antigens lessen the likelihood of contaminating non-viral proteins.
Munz et al. [12] reported high O. cameli incidence rates (up to 95%) in camels from Kenya, Somalia and Sudan; in their ELISA assay they only used semi-purified CPV, which may explain their unexpected high seropositivity rate. The same limitation could be applied to the results from the ELISA assay developed by Pfahler et al. [37] who also used partially purified camelpox antigens.
The genome of an orthopoxvirus codes for a large number of different polypeptides, each of which is potentially antigenic; hence a large number of different antibodies are produced during orthopoxvirus infections [7]. These may be short-lived IgM or persistent IgG. These antibodies are considered to prevent generalization of the virus and the development of a cyclic disease and may persist for 2-3 years [38]. The current study has shown that many, but not all, of the viral antigenic determinants are detected by Western blotting, and hence could be used in differential diagnosis of orthopoxvirus infections.

Serology of camelpox infection in dromedary camels 75
The dual infection by camelpox and parapox viruses recorded in a number of camels in this study (herd A), is not an unusual observation and has been previously reported [25, 39]. Camels are not protected against camel contagious echyma after immunization with vaccinia virus, proving, for the camel, the lack of immunological cross reaction between orthopox and parapox viruses [40]. Tantawi [41] claimed that there was no cross-protection in camels or rabbits between strains of camelpox virus recovered from Egypt [9] and the Iran strain of camelpox virus or vaccinia virus. This finding suggests that the Egyptian strains may have been parapoxviruses rather than orthopoxviruses, since viruses of both genera may cause lesions described as "camelpox" [4, 42].
The results of the SDS-PAGE experiments confirm and extend the work of others which indicated that Orthopoxvirus species may be characterized by their protein electrophoretic patterns. Because different electrophoresis methods were used, direct comparison with the results of others is not always possible but the patterns obtained in this study were generally similar to those reported from some poxviruses and many components were shared [35, 43, 44]. However, careful examination of the camelpox virus pattern and its comparison with other orthopoxviruses (monkey, ectromelia, elephant, vaccinia, buffalopox and cowpox) reported by Arita and Tagaya [43], Esposito et al. [44] and November et al. [45] showed at least three main bands at mol. wt 23, 31 and 35 kDa which are characteristic for O. cameli and could be used to distinguish from the other orthopoxviruses. Similar observations were noted by other workers [35, 46] who high- lighted the importance of the 22, 23, 24, 34, 37, 53 and 54kDa bands in other orthopoxviruses. Camelpox virus is separated from the vaccinia group by the absence of a 53 kDa band and is distinguished from all others by possessing 31 and 35 kDa bands. Although the 23 kDa camelpox virus protein band has been found in other ortho- poxviruses (Ectromelia, elephant virus, vaccinia, buffalopox and cowpox), it is absent from monkeypox (human and monkey isolates).
Due to the lack of similar studies, we were not able to compare our Western blotting results with other workers. However, our findings from the Western blots confirmed the diagnostic value of the 31 and 35 kDa bands for camelpox infections, and their important role in differentiating O. cameli from other orthopoxviruses infections. Furthermore, as these two bands are the main antigens, this work shows the specificity, of the described ELISA, for O. cameli infection.
Increasing interest is being shown in the antigenic differences among Orthopoxvirus species. Well defined viral antigenic determinants will lead to production of monoclonal antibodies against them which could be applied in some assays for their detection [47, 48]. The production of monoclonal antibodies to the camelpox viral antigens (31 and 35 kDa) would enable development of assay systems to detect the virus in tissues and body fluids.
Cowpox-like virus strains have drawn extensive medical attention because they cause skin lesions in cats [49-51], a variety of zoo animals [52, 53] and also in man [54, 55]. These strains are characterized by a low contagiosity for healthy persons and are responsible for benign lesions. However, in predisposed and immunosuppressed persons who are not vaccinated against variola, they can also lead to severe cyclic diseases [56-59]. The permanent reservoir of such viruses is unknown, but several mouse species, rats, and other wildlife rodents have been considered [60-62]. The close relationship between strains of camelpox virus and variola virus [63] suggests that camelpox may need to be added to the growing list of poxviruses derived from animals whose possible importance for man is being thrown into prominence by the success of the WHO smallpox eradication programme.

76 s .M. Azwai et al,
It is poss ible tha t clinical or subcl inical man i fes t a t ion o f the camelpox infect ion in man a l r eady exists bu t no ep idemiologica l s tudies have been m a d e in man , p r imar i ly because o f the lack o f well descr ibed h u m a n cases. Immuno log i ca l surveys for specific camelpox an t ibod ies a m o n g unvacc ina ted h e r d s m a n at risk could , therefore, be helpful to de te rmine the poss ible t ransmiss ib i l i ty o f came lpox to man . This cou ld be pe r fo rmed by a m i n o r modi f i ca t ion o f the descr ibed E L I S A . In the future it is poss ible tha t h u m a n infections wi th came lpox could occur as the smal lpox immuni ty of the h u m a n popu l a t i on wanes; this wou ld only be detected by long- te rm serological moni tor ing . I f camelpox were to infect man , it wou ld p robab ly , as wi th o ther pox virus infections, be mos t i m p o r t a n t in i m m u n o s u p p r e s s e d indiv iduals [38, 56].
A p a r t f rom man , there is no in fo rma t ion a b o u t t ransmiss ion o f camelpox to o ther fa rm animals . However , Bor isovich and Orekhov [64] r epor ted tha t sheep pox hype r immune serum was no t effective aga ins t camelpox , and R a m y a r and Hessami [14] r epor ted that came lpox and sheep pox viruses are ant igenical ly different and that , to prevent camels f rom infect ion, immun iza t i on should be carr ied out by a vaccine p roduced with a h o m o l o g o u s virus. Recent ly , h o m o l o g o u s came lpox vaccines have been p roduced (Duba i camelpox vaccine) K a a d e n et al. [17] and Hafez et al. [65].
W e believe tha t the E L I S A technique descr ibed is a very useful m e t h o d for sero- ep idemio log ica l surveys for the de tec t ion o f an t ibodies agains t O. cameli. The test can also be app l i ed to differentiate, serological ly , between the cl inically s imilar infections o f o r t h o p o x and p a r a p o x viruses. As the app l i ca t ion o f e lectron microscopic invest igat ions to different iate between the two diseases based on the different m o r p h o l o g y o f o r t h o p o x and p a r a p o x viruses is no t poss ible everywhere, it is bel ieved tha t a relat ively quick d i f ferent ia t ion can be achieved using the E L I S A techniques descr ibed here, and could, if necessary, be conf i rmed by Wes te rn blot t ing. This should be a significant advance to con t ro l and assist in the prevent ion o f camelpox.
Acknowledgement--We would like to thank EI-Fateh University, Tripoli, Libya, for providing a project grant for this study.
REFERENCES
1. FAO-OIE-WHO. Animal Health Yearbook. Food and Agriculture Organisation of the United Nations, Rome (1990).
2. Leese A. S. Two diseases of young camels. J. Trop. Vet. Sci. 4, 1-7 (1909). 3. Amanschulow S. A., Samarzew A. A. and Arbusow L. N. Sur la variole du chameau de la r6gion de rOural.
Abstr. Bull. Inst. Pasteur 29, 96 (1930). 4. Jezek Z., Kriz B. and Rothbouer V. Camelpox and its risk to the human population. J. Hyg. Epidem.
Mierobiol. Immun. 27, 29-42 (1983). 5. Kriz B. A study of camelpox in Somalia. J. comp. Path. 92, 1-8 (1982). 6. Marrenikova S. S., Shenkman L. S., Shelukhina E. M. and Malsova N. N. Isolation of camelpox virus and
investigation of its properties. Acta Virol. 18, 423-428 (1974). 7. Fenner F., Wittek R. and Dumbell K. R. The orthopoxviruses. Academic Press Inc., London (1989). 8. Sadykov R. G. Cultivation ofcamelpox virus in chick embryos. In Virusnye Bolezni Selskokhozayaystvennykh
Zhivotnykh, Part I, p. 55, (in Russian) Moscow (1970). 9. Tantawi H. H., Saban M. S., Reda I. M. and Dahaby H. Camelpox virus in Egypt. l--Isolation and
characterization. Bull. Epizoot. Dis. Aft. 22, 315-319 (1974). 10. Davies F. G., Mungai J. N. and Shaw T. Characteristics of a Kenyan camelpox virus. J. Hyg. 75, 381-385
(1975). 11. Davies F. G., Mbugua H., Atema C. and Wilson A. The prevalence of antibody to camelpox virus in six
different herds in Kenya. J. comp. Path. 95, 633-635 (1985). 12. Munz E., Kropp E. and Reimann M. Demonstration of antibodies against Orthopoxvirus cameli in sera of
East African dromedaries by ELISA. J. Vet. Med. B. 33, 221-230 (1986). 13. Fassi-Fehri M. M. Diseases of camels. Rev. Sci. Tech. Off. Int. Epiz. 6, 337-354 (1987).

Serology of camelpox infection in dromedary camels 77
14. Ramyar H. and Hessami M. Isolation, cultivation and characterization of camelpox virus. Zbl. Vet. Med. B 19, 182-189 (1972).
15. AI-Falluji M. M., Tantawi H. H. and Shony M. O. Isolation, identification and characterization of camelpox virus in Iraq. J. Hyg. 83, 267-272 (1979).
16. Higgins A. J., Silvey R. E., Abdelghafir A. E. and Kitching R. P. The epidemiology and control of an outbreak of camelpox in Bahrain. Proc. 1st. Int. Camel Conf. 101-104 (1992).
17. Kaaden O. R., Walz A., Czerny C. P. and Wernery U. Progress in the development of a camelpox vaccine. Proc. 1st. Int. Camel Conf. 47-49 (1992).
18. Al Hendi A. B., Abuelzein E. M., Gameel A. A. and Hassanein M. M. A slow-spreading mild form of camelpox infection. J. Vet. Med. B. 41, 71-73 (1994).
19. Chauhan R. S. and Kanshik R. K. Isolation of camelpox virus in India. Br. Vet. J. 143, 581-582 (1987).
20. Marennikova S. S. The results of examinations of wildlife monkeys for the presence of antibodies and viruses of the pox group. Vopr. Virusol. 3, 321-326 (1975).
21. Schgal C. L. Camelpox. In Report of Summer Institute on Animal Poxviruses, pp. 161-163, G.B. Pant University of Agriculture and Technology, India (1977).
22. Tantawi H. H., EI-Dahaby H. and Fahmy L. S. Comparative studies on poxvirus strains isolated from camels. Acta ViroL 22, 451-457 (1978).
23. Marennikova S. S., Malceva N. N. and Habahpaseva N. A. ELISA--a simple test for detecting and differentiating antibodies to closely related orthopoxviruses. Bull. Wld Hlth. Org. 59, 365 369 (1981).
24. Boulther E. A., Zwartouw H. T., Titmuss D. H. and Maber H. B. The nature of the immune state produced by inactivated vaccinia virus in rabbits. Am. J. Epidem. 94, 612-618 (1971).
25. Munz E., Schillinger D., Reimann M. and Mahnel H. Electron microscopical diagnosis of Ecthyma contagiosum in camels (Camelus dromedarius): first report of the disease in Kenya. J. Vet. Med. B. 33, 73-77 (1986).
26. Munz E., Moallin A. S., Mahnel H. and Reimann M. Camel papiUomatosis in Somalia. Zbl. Vet. B 37, 191-196 (1990).
27. Azwai S. M., Carter S. D. and Woldehiwet Z. The isolation and characterization of camel (Camelus dromedarius) immunoglobulin classes and subclasses. J. comp. Path. 109, 187-195 (1993).
28. Azwai S. M., Carter S. D. and Woldehiwet Z. Monoclonal antibodies against camel (Camelus dromedarius) IgG, IgM and light chains. Vet. Immun. Immunopath. 45, 175-184 (1995).
29. Azwai S. M., Carter S. D. and Woldehiwet Z. Immune responses of camel (Camelus dromedarius) to contagious ecthyma (orf) virus infection. Vet. Microbiol. (In press) (1995).
30. Thomas V., Flores L. and Holowczak J. A. Biochemical and electron microscopic studies of the replication and composition of milker's node virus. J. Virol. 34, 244-255 (1980).
31. Bell S. C., Carter S. D. and Bennett D. Canine distemper viral antigens and antibodies in dogs with rheumatoid arthritis. Res. Vet. Sci. 50, 64-68 (1991).
32. Lowry O. H., Rosebrough N. J., Farr A. L. and Randall R. J. Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 265-275 (1951).
33. Birk H. W. and Koepsel H. Reaction of monoclonal antibodies with plasma membrane proteins after binding on nitrocellulose: Renaturation of antigenic sites and reduction of nonspecific antibody binding. Analyt. Biochem. 164, 12-22 (1987).
34. Laemmli U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-682 (1970).
35. Turner A. and Baxby D. Structural polypeptides of Orthopoxvirus: their distribution in various members and location within the virion. J. Gen. Virol. 45, 537-545 (1979).
36. Wilson A. J. and Dolan R. Important camel diseases in selected areas in Kenya. IPAL Technical report No. E 6, Veterinary Faculty, Nairobi (1978).
37. Pfahler W. H. E., Reimann M. and Munz E. A bitoin-avidin amplified enzyme immunoassay for detection and quantitation of orthopox virus camel antibodies in dromedaries. J. Vet. Med. B. 33, 477-484 (1986).
38. Mayr A. and Czerny C. P. Camelpox virus. In Virus Infections of Ruminants. Dinter, Z. and Morein, B. (Eds). pp. 19-22. Elsevier, Amsterdam (1990).
39. Wernary U. and Kaaden O. R. In Infectious Diseases ofCamelids. Blackwell, Wissenschafts-Verlag (1995). 40. Dashtseren T. S., Solovyev B. V., Varejka F. and Khokhoo A. Camel contagious ecthyma (Pastular
dermatitis). Acta Virol. 28, 12-127 (1984). 41. Tantawi H. H. Comparative studies on camelpox, sheeppox, and vaccinia viruses. Acta Virol. lg, 347-351
(1974). 42. Roslyakov A. A. Comparison of the ultrastructure of camel poxvirus, the virus of a pox-like disease of camels
and contagious ecthyma virus. Vopr. Virusol. 17, 26-30 [reprinted in Vet. Bull. 42, 519 (1972)]. 43. Arita M. and Tagaya I. Structural polypeptides of several strains of orthopoxvirus. Microbiol. Immun. 21,
343-346 (1977).

78 S.M. Azwai et al.
44. Esposito J. J., Obijeski J. F. and Nakano J. H. The virion and soluble antigen proteins of variola, monkeypox and vaccinia viruses. J. Med. Virol. 1, 95-110 (1977).
45. November F. J., Raska K. Jr and Holowczak J. A. The immune response to vaccinia virus infection in mice: analysis of the role of antibody. Arch. Virol. 107, 273-289 (1989).
46. Harper L., Bedson H. S. and Buchan A. Identification of orthopoxviruses by poiyacrylamide gel electro- phoresis of intracellular polypeptides. Virology 93, 435-444 (1979).
47. Meyer H., Osterrieder N. and Czerny C. P. Identification of binding-sites for neutralizing monoclonal- antibodies on the 14-kDa fusion protein of orthopox viruses. Virology 200, 778-783 (1994).
48. Czerny C. P., Johann S., Holzle L. and Meyer H. Epitope detection in the envelope of intracellular naked orthopox viruses and identifcation of encoding genes. Virology 200, 764-777 (1994),
49. Thomsett L. R., Baxby D. and Denham M. H. Cowpox in the domestic cat. Vet. Rec. 108, 567 (1978). 50. Bennett M., Gaskell R. M., Gaskell C. J., Baxby D. and Kelly D. F. Studies on poxvirus infection in cats.
Arch. Virol. 104, 19-33 (1989). 51. Mahnel H., Czerny C. P. and Mayr A. Nachweis und identifizierung yon pockenvirus bei hauskatzen.
J. Vet. Med. B. 36, 231-236 (1989). 52. Mahnel H. Identifizierung eines Kuhpockenvirus, isoliert von einem Kind. J. Vet. Med. B. 33, 362-370 (1986). 53. Nasemann T., Mayr A., Schrag G., Kimmig W. and Mahnel H. Infektion eines madchens mit kuhpockvirus.
Hautarzt 38, 414-418 (1987). 54. Klingebiel T., Vallbracht A., Doller G , Stierhof D., Gerth H. J., Glashauser E. and Herzau V. A severe
human cowpox infection in South Germany. Pediatr. Inf. Dis. J. 7, 883-885 (1988). 55. Marennikova S. S., Shelukhina E. M., Maltseva N. N., Efremova E. V. and Matsevich A. Data from
the serological examination of the population of the Republic of Congo for the presence of antibodies to orthopoxviruses: I. A comparative evaluation of different study methods and general results. J. Microbiol. Epidem. Immunobiol. 3, 95-100 (1984).
56. Redfield R. R., Wright D. C., James W. D., Jones T. S., Brown C. and Burkf D. S. Disseminated vaccinia in a military recruit with human immunodeficiency virus (HIV) disease. N. Engl. J. Med. 316, 673-676 (1987).
57. Eis-Hfibinger A. M., Gerritzen K. E., Schineweis K. E., Pfeiff B., Pullmann H., Mayr A. and Czerny C. P. Fatal cowpox-like infection transmitted by cat. Lancet 336, 880 (1990).
58. Czerny C. P., Eis-Hiibinger A. M., Mayr A., Schineweis K. E. and Pfeiff B. Animal poxviruses transmitted from cat to man: current event with lethal end. J. Vet. Med. B. 38, 421-431 (1991).
59. Pfeiff B., Pullmann H., Eis-Hiibinger A. M., Gerritzen K. E., Schineweis K. E. and Mayr A. Lethale tierpockeninfektion bei einem atopiker unter dem bild einer variola vera. Hautarzt 42, 293-297 (1991).
60. Baxby D., Shackleton W. B., Wheeler J. and Turner A. Comparison of cowpox-like viruses isolated from European zoos. Arch. Virol. 61, 337-340 (1979).
61. Marennikova S. S., Shelukhina E. M. and Efremova E. V. New outlook on the biology of cowpoxviruses. Acta Virol. 28, 437-444 (1984).
62. Munz E. and Mayr A. Problem der tierpocken nach aufhebung der pflichtimpfung gegen die menschenpocken. Tierarztl. Umsch. 39, 187-196 (1984).
63. Baxby D. Smallpox-like viruses from camels in Iran. Lancet 18, 1063-1065 (1972). 64. Borisovich Yu. F. and Orekhov M. D. Camelpox. Abstr. Vet. Bull. 36, 4739 (1966). 65. Hafez S. M., Alsukayran A., Delacruz D., Mazloum K. S., Albokmy A. M., Almukayel A. and Amj A. A.
Development of a live cell-culture camelpox vaccine. Vaccine 10, 533-539 (1992).