separation of a- versus c-nociceptive inputs into spinal–brainstem circuits
TRANSCRIPT

SS
DSa
Sb
q
AzcFhhotl(t
lCccnfifnvP
tc(w
iedtzttE
Kt
*EAd3DelP
Neuroscience 152 (2008) 1076–1085
0d
EPARATION OF A- VERSUS C-NOCICEPTIVE INPUTS INTO
PINAL– BRAINSTEM CIRCUITSAsittesMnBvtnMno
aidptseompc11Pohbrs(1
fecLhbcv(
. M. PARRY,a F. M. MacMILLAN,a S. KOUTSIKOU,a
. McMULLANb AND B. M. LUMBa*
Department of Physiology and Pharmacology, School of Medicalciences, University of Bristol, University Walk, Bristol BS8 1TD, UK
Australian School of Advanced Medicine, 3 Innovation Road, Mac-uarie University, Sydney 2109, Australia
bstract—This study was designed to determine the organi-ation of nociceptive inputs with different behavioral signifi-ance into spinal– brainstem circuits in the rat. Induction ofos protein was used to localize spinal dorsal horn andypothalamic neurons activated by noxious heating of theind paw dorsum at rates known to preferentially activate C-r A-heat nociceptors. This was combined with retrograderansport of cholera toxin subunit B from the dorsolateral/ateral- (DL/L-) or the ventrolateral- (VL-) periaqueductal grayPAG) in order to map the organization of A- and C-fiber inputo spinal– brainstem circuits.
The majority of dorsal horn heat-activated neurons wereocated in laminae I and II. A significantly larger proportion of-fiber-activated neurons projected to the VL-PAG (P<0.05)ompared with its DL/L-sector. In contrast, there was noolumnar separation in the projections of A-fiber-activatedeurons. However, a significantly greater proportion of A-ber-activated neurons (P<0.05) were retrogradely labeledrom the DL/L-PAG, when compared with C-fiber-activatedeurons. A large proportion (25–50%) of A- and C-fiber-acti-ated neurons in the lateral spinal nucleus projected to theAG.
A-fiber-activated neurons were found throughout the ros-ral hypothalamus but those projecting to the PAG were fo-used in the lateral area of the anterior hypothalamusLAAH), from where �20% projected to the VL-PAG, whichas significantly more than to the DL/L PAG (P<0.05).
We hypothesize that the organization of A- versus C-fibernputs to the PAG enables the coordination of coping strat-gies appropriate to meet the demands imposed by theseifferent noxious stimuli. Hypothalamic-PAG projections ac-ivated by A-fiber inputs did not reflect this level of organi-ation and we suggest that this may relate to their role inhermoregulation as opposed to autonomic responses to par-icular nociceptive inputs. © 2008 IBRO. Published bylsevier Ltd. All rights reserved.
ey words: Fos, nociception, pain, spinal cord, periaqueduc-al gray, hypothalamus.
Corresponding author. Tel: �44-117-33-12313; fax: �44-117-33-12288.-mail address: [email protected] (B. M. Lumb).bbreviations: AHAd, anterior hypothalamic area dorsal; CAAH, cau-al area anterior hypothalamus; CTb, cholera toxin subunit B; DAB,,3-diaminobenzidine tetrahydrochloride; DLH, DL-homocysteic acid;L/L, dorsolateral/lateral; FLI, Fos-like immunoreactivity; LAAH, lat-ral area anterior hypothalamus; LHA, lateral hypothalamic area; LSN,
Cateral spinal nucleus; PAG, periaqueductal gray; PB, phosphate buffer;BS-T, phosphate-buffered saline with Triton X-100; VL, ventrolateral.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2008.01.018
1076
lterations in cardiovascular function evoked by noxioustimuli contribute to protective strategies that enable an
ndividual to avoid, escape or cope with tissue damage inhe periphery and, it has long been predicted (Lewis, 1942)hat the precise patterns of autonomic change that arevoked by pain arising from cutaneous versus visceraltructures would be mediated by distinct central pathways.any autonomic control centers in the brain stem receiveociceptive inputs (Gauriau and Bernard, 2002; Keay andandler, 2002) and the induction of Fos protein has pro-ided detailed information about the functional organiza-ion of pathways within the brain that are activated byoxious stimuli arising from different peripheral organs.uch of this work has focused on the organization ofociceptive inputs to the periaqueductal gray (PAG) regionf the midbrain.
The PAG can be divided into functional columns thatre arranged longitudinally around the aqueduct. Neurons
n the different columns are thought to operate duringifferent environmental conditions to mediate either sym-athoexcitation and increased motor activity or sympa-hoinhibition and quiescence. The former pattern of re-ponse is coordinated by neurons in the dorsolateral/lat-ral (DL/L-) PAG, as part of an active coping strategy thatperates during states of arousal. In contrast, the latter isediated by neurons in the ventrolateral (VL-) sector, asart of passive coping strategies that operates during re-uperation or after intense exercise (Carrive, 1993; Lovick,993; Bandler and Shipley, 1994; Bernard and Bandler,998). Nociceptive inputs provide a powerful drive to theAG and recent work indicates a high degree of columnarrganization of these inputs, which is related to their be-avioral significance (Bandler et al., 2000); the DL/L-PAGeing more concerned with processing afferent informationelated to brief cutaneous insults and, in contrast, the VLector with processing deep somatic and visceral painKeay and Bandler, 1993; Keay et al., 1994; Clement et al.,996; Bandler et al., 2000).
The anterior hypothalamus, an important brain regionor the integration of autonomic functions, is thought toxert its effects, at least in part, after a relay in particularolumns of the PAG (Rizvi et al., 1996; Semenenko andumb, 1999). It is of considerable interest therefore, thatypothalamic-PAG projection neurons that are activatedy either noxious visceral or by noxious somatic stimulionstitute largely separate populations of cells that inner-ate the VL- or DL/L- columns of the PAG respectivelySnowball et al., 2000; Parry et al., 2002).
Visceral nerves contain a relatively high proportion of
-fiber nociceptors (Cervero and Tattersall, 1987) and, asved.

avtatcttlih
ttflndPac
Aa1iwo
I
EA(DdwmeD
mpaDrbp1dsPfis
wtt
P
Robc
atsnrttrta
tgpbp2
C
Ftag
T
Itmcabbw
D
SoFg
twa
FiTa1nsiEt(wv0owei(wp
D. M. Parry et al. / Neuroscience 152 (2008) 1076–1085 1077
consequence, visceral pain may be expected to be con-eyed principally by activity in C-fibers, in contrast to cu-aneous pain that may result from both C- and A-nociceptorctivation. As such, differential activation of, or organiza-ion of hypothalamic projections to, the DL/L- and VL-olumns of the PAG might be more directly related to theype of nociceptive afferent activated rather than the loca-ion of the particular end organ. The finding that hypotha-amic neurons driven by C-fiber activation project predom-nantly to the VL-PAG (Lumb et al., 2002) supports thisypothesis.
The hypothesis to be tested in the current study washat nociceptive inputs of different behavioral significancearget the PAG in a functionally relevant manner that re-ects their roles in triggering different patterns of auto-omic response. To test this hypothesis, experiments wereesigned to determine (i) the columnar organization of spino-AG projections activated by C- versus A-nociceptive inputsnd (ii) A-nociceptive input to hypothalamic neurons and theolumnar organization of their projections to the PAG.
EXPERIMENTAL PROCEDURES
ll procedures involving experimental animals were carried out inccordance with the U.K. Animals (Scientific Procedures) Act,986 and associated guidelines. All experiments conformed to
nternational guidelines on the ethical use of animals. Every effortas made to minimize animal suffering and to reduce the numberf animals used.
njection of retrograde tracer
xperiments were carried out on male Wistar rats (250–300 g).nesthesia was induced (60–70 mg kg�1, i.p.) and maintained
17–20 mg kg�1, i.p.) with sodium pentobarbitone (Sigma-Aldrich,orset, UK) at a level at which there was an absence of with-rawal reflexes to minor noxious stimuli. Arterial blood pressureas monitored via the carotid artery and body temperature wasaintained at approximately 37 °C. A small area of the cortex wasxposed just rostral to lambda. Under stereotaxic guidance, aL-homocysteic acid (DLH, 0.05 M; Sigma-Aldrich) –filled glassicropipette was attached to a pressure injection system andositioned in the PAG between 7.4 and 7.6 mm caudal to bregma,t a depth between 4.0 and 5.0 mm below the cortical surface.LH at 30–50 nl was injected and the evoked blood pressure
esponse recorded. Once a ‘depressor’ or a ‘pressor’ site hadeen located the coordinates of the site were noted and theipette was removed from the brain. Cholera toxin subunit B (CTb%; List Biological Laboratories Inc., Campbell, CA, USA) wasrawn up into the micropipette and the pipette repositioned at theame location. CTb (100–200 nl) was pressure-injected into theAG over approximately 10 min and the pipette left in place for a
urther 15 min before being withdrawn. A total of 14 animals werenjected with CTb at pressor sites and another 16 at depressorites.
The skin overlying the skull was sutured, the arterial cannulaas removed and the carotid artery ligated. The rats were allowed
o recover and left for 5–7 days to enable retrograde axonalransport of the cholera toxin.
eripheral stimulation
ats previously injected with retrograde tracer at pressor (n�14)r depressor (n�16) sites were re-anesthetized (sodium pento-arbitone, 60–70 mg kg�1 i.p.) before removal from their home
age. Anesthesia was monitored by assessing corneal reflexes gnd maintained with supplementary doses of sodium pentobarbi-one (17–20 mg kg�1, i.p.) as required. After 2 h, slow (2.5 °C s�1;tarting temperature 30 °C; cutoff 55 °C; ramp duration �60 s;�13) or fast (7.5 °C s�1; starting temperature 30 °C; cutoff 57 °C;amp duration �6 s; n�17) rates of skin heating were applied tohe dorsal surface of the hind paw ipsilateral to the injection site inhe PAG (six times per animal, with an 8 min gap between heatingamps), to preferentially activate C- or A-heat nociceptors respec-ively (Yeomans and Proudfit, 1996; McMullan et al., 2004; Leith etl., 2007).
The rats were maintained under anesthesia for a further 2 h,o allow time for the expression of Fos protein. They were theniven a lethal dose of pentobarbitone (100 mg kg�1 i.p.) anderfused transcardially with 0.1 M phosphate buffer (PB) followedy 4% paraformaldehyde in 0.1 M PB. The brains were removed,ost-fixed overnight in 4% paraformaldehyde then transferred to5% sucrose PB solution for 24–48 h.
ontrol animals
our rats were maintained, unstimulated, under anesthetic for aotal of 4 h and then perfused with 4% paraformaldehyde asbove. Of these, three had been injected previously with retro-rade tracer.
issue processing
n order to localize the CTb injection site, two series of 50 �mransverse sections were cut through the PAG using a freezingicrotome. One series was mounted onto gelatin/chrome alum–
oated microscope slides and counterstained with Neutral Red toid orientation and the other immunostained for CTb as describedy Clement et al. (2000), but using as secondary antibody aiotinylated anti-sheep immunoglobulin (Sigma-Aldrich). Sectionsere then washed in several changes of PB for at least 30 min.
ouble immunostaining
ections (40 �m) of spinal cord lumbar segments L3–L5 were cutn a freezing microtome and free-floating sections processed foros-like immunoreactivity (FLI) and for the presence of the retro-rade tracer CTb.
Three series of 40 �m transverse sections were cut throughhe hypothalamus. One series was mounted and counterstainedith Neutral Red, one series was immunostained for Fos and CTbnd the third was kept in buffer in reserve.
Free-floating hypothalamic sections were double stained forLI and CTb immunoreactive retrograde cells. After three washes
n phosphate buffered saline containing 0.1% Triton X-100 (PBS-), the sections were incubated in a polyclonal rabbit anti-Fosntibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA;:5000 in 0.1 M PB containing 10% newborn calf serum, 1%ormal goat serum and 0.1% Triton) for 48–72 h at 4 °C. Theections were washed again in three changes of PBS-T and thenncubated in biotinylated anti-rabbit immunoglobulin (Dako Ltd.,ly, UK; 1:500 in PBS-T) for 1–2 h at room temperature and
hen washed as before and transferred to extravidin-peroxidaseSigma-Aldrich; 1:1000 in PBS-T) for 1–2 h. Sections wereashed again in three changes of PB. The peroxidase was thenisualized using 3,3-diaminobenzidine tetrahydrochloride (DAB;.015%; Sigma-Aldrich) with nickel intensification and glucosexidase to generate the hydrogen peroxide. The sections wereashed again in PB and transferred to antibody to CTb (Quadrat-ch Ltd., Surrey, UK; 1:20,000) for 24–48 h. After further washing
n PB, sections were transferred to anti-sheep immunoglobulin1:500 in PB, as above), and left for 1–2 h after which they wereashed and transferred to extravidin peroxidase as before. Theeroxidase was visualized using DAB with glucose oxidase to
enerate the hydrogen peroxide but without nickel intensification.
D
Irta[sC
fp1bmhtwcabinwPtt
S
DVbpsDtpmwe
Lsct
Fel
twFmwnc
ntiwnhadwtpioswpDFhwhfrtlwP
Lfo
ItnuF
Tp
NNNNNNN
a
D. M. Parry et al. / Neuroscience 152 (2008) 1076–10851078
ata collection
n 13 animals given slow heating ramps and 13 given fast heatingamps, Fos-labeled nuclei were counted in all sections throughouthe L3–L5 levels of the spinal cord and Fos-positive neuronsssigned to the spinal cord region at which they were found (LI, LIIsuperficial dorsal horn], LIII–VI [deep dorsal horn] and the lateralpinal nucleus (LSN)). Any neuron that was double labeled withTb was also recorded.
For the hypothalamus, a total of 17 animals were analyzed afterast heating ramps, 13 from the above group plus four extra. Fos-ositive neurons were counted in every third section (i.e. every20 �m) throughout the hypothalamus from 1.2–2.0 mm caudal toregma and the locations of individual Fos-positive neurons wereapped onto representative drawings of transverse sections. Theypothalamus was sub-divided into stylized regions with reference tohe atlas of Bleier et al. (1979) and Fos-immunoreactive neuronsere assigned to the appropriate region. Labeled neurons wereounted throughout the rostrocaudal extent of each of the regionsnd mean values (�S.E.M) were calculated. Counts were made onoth the right and left sides of the hypothalamus. Statistical compar-
sons were made between the absolute numbers of Fos-positiveeurons in each hypothalamic region and all Fos-positive neuronsere examined for double-labeling with CTb from the VL or DL/LAG. Double-labeled neurons were mapped onto the representative
ransverse sections and the number within each region of the hypo-halamus was counted as described above.
tatistical analysis
ata in bar charts are displayed as medians�interquartile ranges.alues in the text and in tables are means�S.E.M. Comparisonsetween multiple groups were made using Kruskal-Wallis non-arametric variation of one-way ANOVA. Data sets in which aignificant change was observed were further compared usingunn’s multiple comparison post hoc test. Comparisons between
wo groups were made using Mann-Whitney U test, while com-arisons between left and right sites of the hypothalamus wereade using Wilcoxon matched pairs test. All statistical analysisas performed using GraphPad Prism 4.0. P�0.05 was consid-red to represent a significant difference.
RESULTS
aminar localization of spinal neurons activated bylow versus fast rates of skin heating and theolumnar organization of their projectionso the PAG
ast and slow rates of skin heating of the hind paw dorsumvoked significantly more (P�0.05) fos in the ipsilateral
able 1. Laminar localization of neurons activated by slow versusercentages of neurons double labeled from the DL/L and the VL PA
o. of fos �ve neurons in unstimulated control rats (n�4)o. of fos �ve neurons in response to SLOW rates of skin heating (no. and (%) double-labeled neurons from DL/L-PAG (n�7)o. and (%) double-labeled neurons from VL-PAG (n�6)o. of fos �ve neurons in response to FAST rates of skin heating (n�
o. and (%) double-labeled neurons from DL/L-PAG (n�7)o. and (%) double-labeled neurons from VL-PAG (n�6)
Fos-positive neurons were counted in all sections (40 �m) from L3–Lnd III–VI; deep dorsal horn).
umbar segments 3–5 than in unstimulated control animals s
hroughout laminae I–VI and the LSN (Table 1). Consistentith previous studies that allowed 2 h for the induction ofos protein (Coggeshall, 2005), there were significantlyore neurons in the superficial (laminae I and II) comparedith the deep dorsal horn (laminae III–VI) in response tooxious heat stimulation (P�0.01; Mann-Whitney test,omparison not shown).
Fig. 1A and B shows the proportions of Fos-positiveeurons, following either fast or slow rates of skin heatinghat were double labeled with retrograde tracer injectednto either the VL- or DL/L-PAG. Double-labeled neuronsere found in lamina I and in the LSN. No double-labeledeurons were encountered in lamina II or the deep dorsalorn. In animals stimulated with slow rates of skin heating,significantly greater proportion of lamina I neurons were
ouble labeled with tracer injected into the VL-comparedith the DL/L-PAG (P�0.05). A similar trend was seen in
he LSN, although this did not reach significance. Exam-les of single- and double-labeled (Fos plus CTb) neurons
n the superficial dorsal horn and LSN following slow ratesf skin heating and injection of CTb into the VL-PAG arehown in Fig. 2. Following fast rates of skin heating thereas no significant difference in the proportions of Fos-ositive neurons retrogradely labeled from either the VL- orL/L-PAG. However, a significantly greater proportion ofos-positive neurons (P�0.05) following fast rates of skineating were retrogradely labeled from the DL/L-PAG,hen compared with those activated by slow rates of skineating (Fig. 1C). In contrast, no significant difference wasound in proportions of neurons activated by fast and slowates of skin heating that were retrogradely labeled fromhe VL-PAG. Examples of single (Fos or CTb) and doubleabeling (Fos and CTb) from an individual experiment inhich CTb was injected into either the DL/L- or the VL-AG are illustrated in Fig. 3.
ocalization of hypothalamic neurons activated byast rates of skin heating and the columnarrganization of their projections to the PAG
n all animals Fos-positive neurons were found throughouthe hypothalamus, with no significant difference betweenumbers ipsilateral and contralateral to the noxious stim-lus. There was no significant difference in the numbers ofos-positive neurons in all areas under investigation (i.e.
s of skin heating, controls with no skin heating, and numbers and
amina I Lamina II Laminae III–VI LSN
17.3�11.6 2.5�0.9 15.3�8.0 0.5�0.358.5�115.5 129.5�24.2 84.6�37.2 19.7�2.758.3�19.4 (7.4) 0 (0) 0 (0) 3.4�0.9 (25.2)91.0�11.5 (14.7) 0 (0) 0 (0) 7.3�2.2 (41.7)43.8�71.7 54.5�10.2 51.3�8.0 28.5�3.952.4�11.0 (11.5) 0 (0) 0 (0) 9.1�2.3 (42.8)68.3�10.4 (11.8) 0 (0) 0 (0) 15�1.6 (48.9)
pressed as means�S.E.M. Laminae (I and II; superficial dorsal horn
fast rateG
L
�13) 7
13) 5
5, and ex
uprachiasmatic and supraoptic nuclei, the caudal and

lL(twRrwLwiFclccae1stvclidP
slT
pabbti
Tnroods
tpcn(nCPtlpvb
Fsob L/L- orM
D. M. Parry et al. / Neuroscience 152 (2008) 1076–1085 1079
ateral areas of the anterior hypothalamus (CAAH andAAH respectively), the dorsal anterior hypothalamic areaAHAd) and the paraventricular nucleus), with the excep-ion of the lateral hypothalamic area (LHA), in which thereere significantly fewer Fos-positive neurons (P�0.05).egions of the hypothalamus in which double-labeled neu-
ons were found (i.e. Fos-positive neurons also labeledith CTb) are shown in Table 2, together with data from theHA. The highest proportions of Fos-positive neurons thatere double labeled with CTb from the PAG were located
n the LAAH. Here, although there were similar numbers ofos-positive neurons both ipsilateral (‘L’ in Table 2) andontralateral (‘R’ in Table 2) to the peripheral noxious stimu-us, significantly more double-labeled neurons were localizedontralateral to the noxious stimulus (P�0.01). In LAAH,ontralateral (‘R’ in Table 2) to the noxious stimulus, 20.1%nd 5.7% of neurons projected to the ipsilateral and contralat-ral VL-PAG respectively. Of fos positive neurons in LAAH,2.2% were contralateral (‘R’ in Table 2) to the noxioustimulus projected to the ipsilateral DL/L-PAG and 3.9% tohe contralateral DL/L-PAG. Significantly more neurons acti-ated by fast rates of skin heating projected to the VL-PAGompared with its DL/L sector (P�0.05). Locations of double-abeled neurons (Fos plus CTb) in the hypothalamus follow-ng fast rates of skin heating and injections of CTb at aepressor site in the VL-PAG and a pressor site in the DL/L-AG in individual animals are shown in Fig. 4.
In contrast, in all of the other hypothalamic sub-divi-ions, less than 3.1% of neurons were found to be doubleabeled despite high numbers of Fos-positive neurons.
ig. 1. Histograms to illustrate columnar organization of projections tokin heating. Percentages of fos positive neurons in Lam I, II (superfir fast rates of skin heating, with projections to VL PAG and/or DL/L PAy fast versus slow rates of skin heating and projecting to either Dann-Whitney test. Lam�lamina/e.
wo single examples of hypothalamic labeling, one with p
rojections to the VL PAG and the other to the DL/L PAGre shown in Fig. 4. The PAG injection sites are illustratedelow the relevant line drawings and superimposed is thelood pressure response evoked by injection of DLH athat site. The panel at the bottom illustrates the PAGnjection sites in all animals.
DISCUSSION
he data presented here provide further evidence thatociceptive inputs of different qualities target particularegions of the PAG. We hypothesize that this level ofrganization enables the coordination of specific patternsf autonomic response that are appropriate to meet theemands imposed by stressors of different behavioralignificance.
These studies are the first to describe, and compare,he columnar organization of inputs to the PAG from spinalrojection neurons that are activated by A- versus C-noci-eptive inputs. The main findings are that (i) superficial butot deep dorsal horn neurons project directly to the PAG,ii) in the superficial dorsal horn lamina I, but not lamina II,eurons are the source of input to the PAG and that-nociceptor-activated lamina I neurons target the VL-AG compared with its DL/L-sector, (iii) a greater propor-
ion of A-fiber compared with C-fiber-nociceptor-activatedamina I neurons project to the DL/L-PAG, (iv) a largeroportion (25–50%) of neurons in the LSN that are acti-ated by A- or C-nociceptor stimulation project to the PAG,ut there is no clear columnar organization of these
from spinal neurons activated by either slow (A) or fast (B) rates ofl horn), Lam III–VI (deep dorsal horn) and LSN, in response to slowmparisons of the total number of spinal dorsal horn neurons activated
VL-PAG. Values are medians and inter-quartile ranges. * P�0.05;
the PAGcial dorsaG. (C) Co
rojections.

hbrgD
M
In(1aicii
srFtdsp4bn
Lso
On
FP ). Fos-lab
D. M. Parry et al. / Neuroscience 152 (2008) 1076–10851080
We also report that neurons throughout the rostralypothalamus are activated by A-nociceptor stimulation,ut those that project to the PAG are localized in a discreteegion of the anterior hypothalamus, LAAH, from where areater proportion project to the VL-PAG as opposed to itsL-sector.
ethodological considerations
nduction of Fos has been used widely as a marker ofeuronal activation in the CNS, including higher centerse.g. Bandler et al., 2000; Bullitt, 1989; Clement et al.,996; Keay et al., 1994). The main attraction of the Fospproach is that it reveals whole populations of depolar-
zed neurons. However, an important limitation is that Fosan be induced by various stimuli other than that undernvestigation. As a result, care was taken to minimize FLI
A
B
B
ig. 2. Photomicrograph to show CTb- and Fos-labeled neurons in thAG. High magnification insets are from lamina I (B) and the LSN (C
nduced by non-specific stimuli, such as handling and p
tress. Therefore, in the present study, rats were handledegularly during the period between tracer injection andos induction in order to reduce FLI evoked by stress on
he day of the experiment. Experimental procedures un-ertaken at the onset of anesthesia may also evoke non-pecific Fos expression. In the present study, FLI inducedrior to anesthesia was minimized by allowing a delay ofh between anesthetic induction and perfusion, as it has
een reported that FLI in diencephalic neurons returns toear basal levels 3 h post stimulus (Bon et al., 1997).
ocalization of dorsal horn neurons activated bylow and fast rates of skin heating and the columnarrganization of their projections to the PAG
ur observation that the majority of neurons activated by aoxious heat stimulus are located in the superficial, com-
C
horn following slow ramp stimulation and tracer injection into the VLeled neurons (open arrows), double-labeled neurons (filled arrows).
C
e dorsal
ared with the deep, dorsal horn is consistent with previ-
olaa(nonbtlS
riiterHwa
dlrbCDnePsdtntd1at((Vttc
drCcnitrCnttPVn
No
Iann(
FsnPcr
D. M. Parry et al. / Neuroscience 152 (2008) 1076–1085 1081
us reports from this (Koutsikou et al., 2007) and otheraboratories that have used the expression of Fos proteins a marker of neuronal activity (Bullitt, 1989; Williams etl., 1990; Coggeshall, 2005). As reported previouslyKoutsikou et al., 2007), there is a tendency for moreeurons to be activated by slow compared with fast ratesf skin heating and for there to be more Fos-positiveeurons in lamina I compared with lamina II. As reportedy others, superficial dorsal horn neurons that project tohe contralateral PAG are located almost exclusively inamina I (Keay et al., 1997; Gauriau and Bernard, 2002;
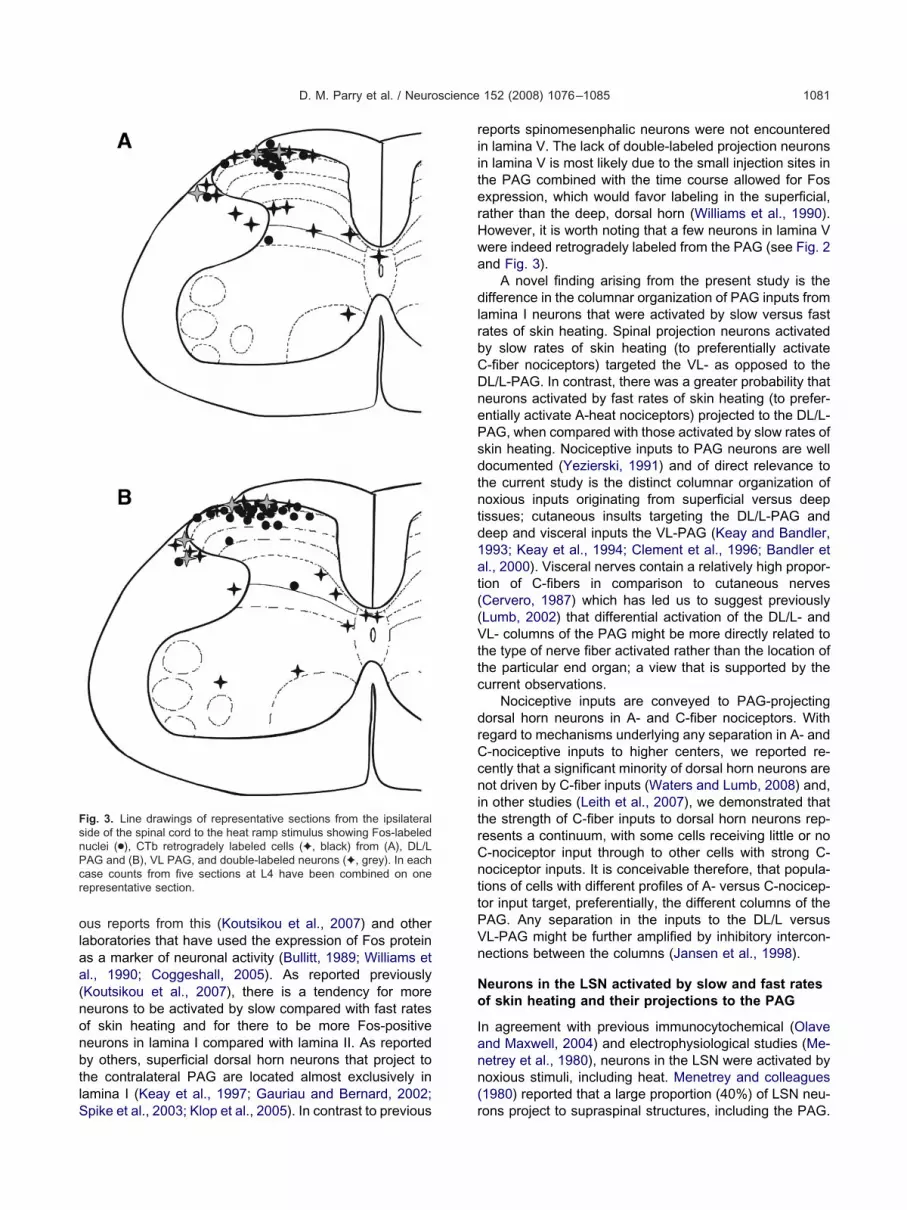
ig. 3. Line drawings of representative sections from the ipsilateralide of the spinal cord to the heat ramp stimulus showing Fos-labeleduclei (●), CTb retrogradely labeled cells (✦, black) from (A), DL/LAG and (B), VL PAG, and double-labeled neurons (✦, grey). In eachase counts from five sections at L4 have been combined on oneepresentative section.
pike et al., 2003; Klop et al., 2005). In contrast to previous r
eports spinomesenphalic neurons were not encounteredn lamina V. The lack of double-labeled projection neuronsn lamina V is most likely due to the small injection sites inhe PAG combined with the time course allowed for Fosxpression, which would favor labeling in the superficial,ather than the deep, dorsal horn (Williams et al., 1990).owever, it is worth noting that a few neurons in lamina Vere indeed retrogradely labeled from the PAG (see Fig. 2nd Fig. 3).
A novel finding arising from the present study is theifference in the columnar organization of PAG inputs from
amina I neurons that were activated by slow versus fastates of skin heating. Spinal projection neurons activatedy slow rates of skin heating (to preferentially activate-fiber nociceptors) targeted the VL- as opposed to theL/L-PAG. In contrast, there was a greater probability thateurons activated by fast rates of skin heating (to prefer-ntially activate A-heat nociceptors) projected to the DL/L-AG, when compared with those activated by slow rates ofkin heating. Nociceptive inputs to PAG neurons are wellocumented (Yezierski, 1991) and of direct relevance tohe current study is the distinct columnar organization ofoxious inputs originating from superficial versus deepissues; cutaneous insults targeting the DL/L-PAG andeep and visceral inputs the VL-PAG (Keay and Bandler,993; Keay et al., 1994; Clement et al., 1996; Bandler etl., 2000). Visceral nerves contain a relatively high propor-ion of C-fibers in comparison to cutaneous nervesCervero, 1987) which has led us to suggest previouslyLumb, 2002) that differential activation of the DL/L- andL- columns of the PAG might be more directly related to
he type of nerve fiber activated rather than the location ofhe particular end organ; a view that is supported by theurrent observations.
Nociceptive inputs are conveyed to PAG-projectingorsal horn neurons in A- and C-fiber nociceptors. Withegard to mechanisms underlying any separation in A- and-nociceptive inputs to higher centers, we reported re-ently that a significant minority of dorsal horn neurons areot driven by C-fiber inputs (Waters and Lumb, 2008) and,
n other studies (Leith et al., 2007), we demonstrated thathe strength of C-fiber inputs to dorsal horn neurons rep-esents a continuum, with some cells receiving little or no-nociceptor input through to other cells with strong C-ociceptor inputs. It is conceivable therefore, that popula-ions of cells with different profiles of A- versus C-nocicep-or input target, preferentially, the different columns of theAG. Any separation in the inputs to the DL/L versusL-PAG might be further amplified by inhibitory intercon-ections between the columns (Jansen et al., 1998).
eurons in the LSN activated by slow and fast ratesf skin heating and their projections to the PAG
n agreement with previous immunocytochemical (Olavend Maxwell, 2004) and electrophysiological studies (Me-etrey et al., 1980), neurons in the LSN were activated byoxious stimuli, including heat. Menetrey and colleagues1980) reported that a large proportion (40%) of LSN neu-
ons project to supraspinal structures, including the PAG.
A2Psnruftpslsic
Hro
Ofaprargwr(2(pluctm1i
nmogrVvphPIitiA
F
NcamcgtTgflctaisp1ondcr
T
H
NNN
N
sn
i
(ation.
D. M. Parry et al. / Neuroscience 152 (2008) 1076–10851082
gain this is consistent with the current studies in which5–50% of neurons in the LSN were found to project to theAG. However, unlike projections from the superficial dor-al horn, there was no significant difference in the colum-ar organization of C- versus A-fiber inputs from this spinalegion to the PAG. The role of LSN in nociception is largelynknown (Olave and Maxwell, 2004) and as such theunctional significance of this ascending pathway remainso be determined. However the observation, in this andrevious studies, that noxious stimulation of peripheraltructures provides a powerful input to LSN, and that aarge proportion of neurons in this region project to brain-tem regions with established roles in nociceptive process-ng, indicates that the role of this neglected region in no-iceptive processing merits further investigation.
ypothalamic neurons activated by slow and fastates of skin heating and the columnar organizationf their projections to the PAG
nly data relating to hypothalamic neurons activated byast rates of skin heating are included here, as informationrising from slow rates of skin heating has been reportedreviously (Lumb et al., 2002). Fos-positive neurons inesponse to fast rates of skin heating (to preferentiallyctivate A-heat nociceptors) were located throughout theostral hypothalamus, with similar numbers in different re-ions. These findings are similar to other studies in whiche used different modalities of noxious stimulation; slow
ates of skin heating (Lumb et al., 2002), noxious visceralSnowball et al., 2000) and noxious somatic (Parry et al.,002) stimuli. As such, our data support other studiesBullitt, 1989; Traub et al., 1996; Rodella et al., 1998) thatrovide evidence for strong nociceptive input to hypotha-
amic neurons; findings that are consistent with the mod-latory effects of noxious stimulation on diverse functionsoordinated by neurons in different regions of the hypo-halamus. The PAG is a major target of projections fromany hypothalamic regions (Saper et al., 1978; Beitz,982; Holstege, 1987; Rizvi et al., 1996). However, strik-
able 2. Numbers of Fos-positive neurons in different regions of the
ypothalamic region LAAH
L R
umber of Fos �ve neurons (controls; n�6) 12�4† 17�
umber of Fos �ve neurons (fast ramps; n�17) 52�4.8 49�
o. and (%) of double-labeled neurons fromDL/L-PAG (n�7)
2 (3.9) 7 (1
o. and (%) of double-labeled neurons fromVL-PAG (n�10)
3 (5.7) 9 (2
Also numbers and percentages of Fos-positive neurons double-labection between 1.2 and 2.0 mm caudal to Bregma and those doubeurons and double-labeled neurons in each region of the hypothalamSignificant difference between the number of fos-positive neurons
ndicated with † P�0.01 and ‡ P�0.001 (Mann-Whitney test).Significant difference in the expression of Fos between left (L) and
Wilcoxon matched pairs test).Right�ipsilateral to PAG and contralateral to peripheral heat stimul
ngly and as described previously, neurons activated by T
oxious stimuli that project to the PAG were localizedainly in the LAAH (Lumb et al., 2002), where up to 20%f Fos-positive neurons were double-labeled with retro-rade tracer injected into the PAG. Significantly more neu-ons activated by fast rates of skin heating projected to theL-PAG (�20%) compared with the DL/L-sector (�20%ersus �12%). In a previous study, remarkably similarroportions of neurons activated by slow rates of skineating were found to project to the VL- versus the DL/L-AG (�21% versus 11% respectively; Lumb et al., 2002).
n other words, although there is evidence for a separationn the columnar organization of heat nociceptive input fromhe LAAH to the PAG, it is the VL-PAG that is targetedrrespective of whether the input originates from C- or-heat nociceptors.
unctional significance
eurons in the different functional columns of the PAGoordinate emotional coping strategies that are appropri-te to meet the demands imposed by different environ-ental and emotional stressors. Neurons in the DL/L-
olumns coordinate active coping strategies that include aeneral sympathoexcitation resulting in hypertension,achycardia and redistribution of regional blood flow.hese alterations in autonomic outflow support emer-ency, defensive behaviors such as confrontation or fight/ight responses; an engagement with the environment. Inontrast, neurons in the VL-PAG coordinate passive emo-ional coping strategies. The response is characterized by
disengagement from the external environment, whichncludes decreased responsiveness to tactile or visualtimuli (quiescence, immobility, hyporeactivity) and sym-athoinhibition (Bandler and Depaulis, 1991; Morgan et al.,991; Depaulis et al., 1994). Noxious stimuli are one formf environmental stressor that powerfully activate PAGeurons and it has been suggested that noxious stimuli ofifferent behavioral significance (escapable versus ines-apable) trigger activity in the DL/L- and VL- columnsespectively to coordinate appropriate coping behaviors.
mus in control animals and following fast ramp stimulation
CAAH AHAd LHA
L R L R L R
6�2‡ 8�2‡ 19�5‡ 20�6† 11�2 7�232�4.6 38�5.0 50�6.1 48�6.7 14�2.2 12�1.70 (0) 1 (2.4)* 0 (0) 1 (2.2) 0 (0) 0 (0)
0 (0) 0 (0) 1 (2.2) 1 (2.1) 0 (0) 0 (0)
retrograde tracer. Fos-positive neurons were counted in every thirdwith CTb recorded. For each animal total numbers of Fos positivecombined. Fos-positive neurons are expressed as means�S.E.M.
ol and experimental animals in each region of the hypothalamus is
sides of the hypothalamus is indicated by * P�0.05 and ** P�0.01
hypothala
3†
4.92.2)*
0.1)**
eled withle labeled
us werein contr
right (R)
his view is supported by the observation that brief nox-

FdpDS
D. M. Parry et al. / Neuroscience 152 (2008) 1076–1085 1083
ig. 4. Representative sections of the rostral hypothalamus to indicate the locations of Fos-positive neurons in individual experiments that wereouble labeled from the VL or the DL/L PAG. The PAG injection sites are illustrated below the relevant line drawings and superimposed is the bloodressure response evoked by injection of DLH at that site. Bottom panels illustrate the locations of all injection sites in the VL-PAG (left) and theL/L-PAG. Numbers indicate distance in mm caudal to bregma, and the different regions of the hypothalamus are shown at the top. Abbreviations:
ON, supraoptic nucleus; PVN, paraventricular nucleus.
iaeK2o(Ttiifb(Cd
dascrurpkstWsCntcSidhcrhpof(thon
ATeS
B
B
B
B
B
B
B
B
C
C
C
C
C
C
D
G
H
J
K
K
K
K
D. M. Parry et al. / Neuroscience 152 (2008) 1076–10851084
ous cutaneous (escapable) stimuli activate the DL/L-PAGnd prolonged cutaneous, visceral and deep noxious (in-scapable) stimuli the VL-PAG (Keay and Bandler, 1993;eay et al., 1994; Clement et al., 1996; Bandler et al.,000). A- and C-fiber nociceptors convey different qualitiesf the pain signal, first (pricking, well localized) and secondburning, diffuse) pain respectively (Schady et al., 1983;orebjork and Ochoa, 1990; Magerl et al., 2001). Differen-
ial organization of A- and C-nociceptive input to the PAGs therefore consistent with the concept that escapable andnescapable noxious stimuli trigger activity in the differentunctional columns of the PAG; A-fiber inputs providingehaviorally useful information that drive active copingcoordinated by neurons in the DL/L column) and diffuse-fiber inputs triggering quiescence and immobility (coor-inated by neurons in the VL column).
Our previous findings that hypothalamic neuronsriven by noxious somatic stimuli target the DL/L-PAGnd, in comparison, those activated by noxious visceraltimuli the VL-PAG have led us to suggest that, underertain circumstances, neurons in the hypothalamus mayeinforce, or fine tune, the functioning of the different col-mns of the PAG in a behaviorally meaningful way. In thisespect it is notable that nociceptive hypothalamic-PAGrojection neurons are located mainly in the LAAH; an areanown to coordinate patterns of change in autonomic andensory function that are similar to those coordinated fromhe different columns of the PAG (Lumb and Lovick, 1993;
orkman and Lumb, 1997; Simpson et al., 2007). At firstight, the current finding that neurons activated by A- and-nociceptors (Lumb et al., 2002), target the VL-PAG doesot support this view. However it must be borne in mindhat, although many projections from LAAH to the differentolumns of the PAG are excitatory (Rizvi et al., 1996;emenenko and Lumb, 1999) we cannot exclude a role for
nhibitory projections that may selectively influence theifferent columns to preserve the functional organization ofypothalamic–PAG connections. Also, hypothalamic–PAGircuits that include the VL column have roles in thermo-egulation (Yoshida et al., 2005) and the data presentedere were generated using heat stimuli. Therefore theresent findings may result, at least in part, from activationf thermoregulatory pathways. This could account for dif-erences in the organization of circuits activated by pinchParry et al., 2002) and those activated by fast heat rampso preferentially activate A-heat nociceptors as reportedere. Future studies in which modalities of stimulationther than heat are used to selectively activate A-fiberociceptors will help resolve this issue.
cknowledgments—The work was supported by the Wellcomerust. The authors thank Annette Goodall and Simon Lishman forxpert technical help. Simon McMullan was a University of Bristolcholar.
REFERENCES
andler R, Depaulis A (1991) Midbrain periaqueductal gray control ofdefensive behavior in the cat and the rat. In: The midbrain periaq-
ueductal gray matter: functional, anatomical and neurochemicalorganization (Depaulis A, Bandler R, eds), pp 175–198. New York:Plenum Press.
andler R, Keay KA, Floyd N, Price J (2000) Central circuits mediatingpatterned autonomic activity during active vs. passive emotionalcoping. Brain Res Bull 53:95–104.
andler R, Shipley MT (1994) Columnar organization in the midbrainperiaqueductal gray: modules for emotional expression? TrendsNeurosci 17:379–389.
eitz AJ (1982) The organization of afferent projections to the midbrainperiaqueductal gray of the rat. Neuroscience 7:133–159.
ernard JF, Bandler R (1998) Parallel circuits for emotional copingbehaviour: new pieces in the puzzle. J Comp Neurol 401:429–436.
leier R, Cohn P, Siggelkow IR (1979) A cytoarchitectonic atlas of thehypothalamus and hypothalamic third ventricle of the rat. In: Hand-book of the hypothalamus (Morgane PJ, Panksepp J, eds), pp137–229. New York: Marcel Dekker.
on K, Lanteri-Minet M, de Pommery J, Michiels JF, Menetrey D(1997) Cyclophosphamide cystitis as a model of visceral pain inrats: minor effects at mesodiencephalic levels as revealed by theexpression of c-fos, with a note on Krox 24. Exp Brain Res113:249–264.
ullitt E (1989) Induction of c-fos-like protein within the lumbar spinalcord and thalamus of the rat following peripheral stimulation. BrainRes 493:391–397.
arrive P (1993) The periaqueductal gray and defensive behavior:functional representation and neuronal organization. Behav BrainRes 58:27–47.
ervero F (1987) Fine afferent fibres from viscera and visceral pain:anatomy and physiology of viscero-somatic convergence. In: Fineafferent nerve fibers and pain (Schmidt RF et al., eds), pp 322–331. Weinheim: VCH Publishers.
ervero F, Tattersall JE (1987) Somatic and visceral inputs to thethoracic spinal cord of the cat: marginal zone (lamina I) of thedorsal horn. J Physiol 388:383–395.
lement CI, Keay KA, Owler BK, Bandler R (1996) Common patternsof increased and decreased fos expression in midbrain and ponsevoked by noxious deep somatic and noxious visceral manipula-tions in the rat. J Comp Neurol 366:495–515.
lement J, Van Ranst M, Leirs H (2000) The results of tick-boneencephalitis (TBE)-vaccination. Am J Trop Med Hyg 62:325–326.
oggeshall RE (2005) Fos, nociception and the dorsal horn. ProgNeurobiol 77:299–352.
epaulis A, Keay KA, Bandler R (1994) Quiescence and hyporeactiv-ity evoked by activation of cell bodies in the ventrolateral midbrainperiaqueductal gray of the rat. Exp Brain Res 99:75–83.
auriau C, Bernard JF (2002) Pain pathways and parabrachial circuitsin the rat. Exp Physiol 87:251–258.
olstege G (1987) Some anatomical observations on the projectionsfrom the hypothalamus to brainstem and spinal cord: an HRP andautoradiographic tracing study in the cat. J Comp Neurol 260:98–126.
ansen ASP, Farkas E, Sams JM, Loewy AD (1998) Local connectionsbetween the columns of the periaqueductal gray matter: a case forintrinsic neuromodulation. Brain Res 784:329–336.
eay KA, Bandler R (1993) Deep and superficial noxious stimulationincreases Fos-like immunoreactivity in different regions of the mid-brain periaqueductal grey of the rat. Neurosci Lett 154:23–26.
eay KA, Bandler R (2002) Distinct central representations of ines-capable and escapable pain: observations and speculation. ExpPhysiol 87:275–279.
eay KA, Clement CI, Owler B, Depaulis A, Bandler R (1994) Con-vergence of deep somatic and visceral nociceptive informationonto a discrete ventrolateral midbrain periaqueductal gray region.Neuroscience 61:727–732.
eay KA, Feil K, Gordon BD, Herbert H, Bandler R (1997) Spinalafferents to functionally distinct periaqueductal gray columns in therat: an anterograde and retrograde tracing study. J Comp Neurol
385:207–229.
K
K
L
LL
L
L
L
M
M
M
M
O
P
R
R
S
S
S
S
S
S
T
T
W
W
W
Y
Y
Y
D. M. Parry et al. / Neuroscience 152 (2008) 1076–1085 1085
lop EM, Mouton LJ, Hulsebosch R, Boers J, Holstege G (2005) In catfour times as many lamina I neurons project to the parabrachialnuclei and twice as many to the periaqueductal gray as to thethalamus. Neuroscience 134:189–197.
outsikou S, Parry DM, MacMillan FM, Lumb BM (2007) Laminarorganization of spinal dorsal horn neurons activated by C- versusA-heat nociceptors and their descending control from the periaq-ueductal grey in the rat. Eur J Neurosci 26:943–952.
eith JL, Wilson AW, Donaldson LF, Lumb BM (2007) COX-1 derivedprostaglandins in the periaqueductal grey differentially control C-versus A-fibre-evoked spinal nociception. J Neurosci 27:11296–11305.
ewis T (1942) Pain. London: MacMillan.ovick TA (1993) Integrated activity of cardiovascular and pain regu-
latory systems: role in adaptive behavioural responses. Prog Neu-robiol 40:631–644.
umb BM (2002) Inescapable and escapable pain is represented indistinct hypothalamic-midbrain circuits: specific roles for Adelta-and C-nociceptors. Exp Physiol 87:281–286.
umb BM, Lovick TA (1993) The rostral hypothalamus: an area for theintegration of autonomic and sensory responsiveness. J Neuro-physiol 70:1570–1577.
umb BM, Parry DM, Semenenko FM, McMullan S, Simpson DA(2002) C-nociceptor activation of hypothalamic neurones and thecolumnar organisation of their projections to the periaqueductalgrey in the rat. Exp Physiol 87:123–128.
agerl W, Fuchs PN, Meyer RA, Treede RD (2001) Roles of capsa-icin-insensitive nociceptors in cutaneous pain and secondary hy-peralgesia. Brain 124:1754–1764.
cMullan S, Simpson DA, Lumb BM (2004) A reliable method for thepreferential activation of C- or A-fibre heat nociceptors. J NeurosciMethods 138:133–139.
enetrey D, Chaouch A, Besson JM (1980) Location and properties ofdorsal horn neurons at origin of spinoreticular tract in lumbarenlargement of the rat. J Neurophysiol 44:862–877.
organ MM, Gold MS, Liebeskind JC, Stein C (1991) Periaqueductalgray stimulation produces a spinally mediated, opioid antinocicep-tion for the inflamed hindpaw of the rat. Brain Res 545:17–23.
lave MJ, Maxwell DJ (2004) Axon terminals possessing alpha2C-adrenergic receptors densely innervate neurons in the rat lateralspinal nucleus which respond to noxious stimulation. Neuro-science 126:391–403.
arry DM, Semenenko FM, Conley RK, Lumb BM (2002) Noxioussomatic inputs to hypothalamic-midbrain projection neurones: acomparison of the columnar organisation of somatic and visceralinputs to the periaqueductal grey in the rat. Exp Physiol 87:117–122.
izvi TA, Murphy AZ, Ennis M, Behbehani MM, Shipley MT (1996)Medial preoptic area afferents to periaqueductal gray medullo-output neurons: a combined Fos and tract tracing study. J Neurosci
16:333–344.odella L, Rezzani R, Gioia M, Tredici G, Bianchi R (1998) Expressionof Fos immunoreactivity in the rat supraspinal regions followingnoxious visceral stimulation. Brain Res Bull 47:357–366.
aper CB, Swanson LW, Cowan WM (1978) The efferent connectionsof the anterior hypothalamic area of the rat, cat and monkey.J Comp Neurol 182:575–599.
chady WJ, Torebjork HE, Ochoa JL (1983) Peripheral projections ofnerve fibres in the human median nerve. Brain Res 277:249–261.
emenenko FM, Lumb BM (1999) Excitatory projections from theanterior hypothalamus to periaqueductal gray neurons that projectto the medulla: a functional anatomical study. Neuroscience94:163–174.
impson DAA, Headley PM, Lumb BM (2007) Selective inhibition fromthe anterior hypothalamus of C- vs A-fibre mediated spinal noci-ception. Pain, doi:10.1016/j.pain. 2007.07.008.
nowball RK, Semenenko FM, Lumb BM (2000) Visceral inputs toneurons in the anterior hypothalamus including those that projectto the periaqueductal gray: a functional anatomical and electro-physiological study. Neuroscience 99:351–361.
pike RC, Puskar Z, Andrew D, Todd AJ (2003) A quantitative andmorphological study of projection neurons in lamina I of the ratlumbar spinal cord. Eur J Neurosci 18:2433–2448.
orebjork HE, Ochoa JL (1990) New method to identify nociceptorunits innervating glabrous skin of the human hand. Exp Brain Res81:509–514.
raub RJ, Sengupta JN, Gebhart GF (1996) Differential c-fos expres-sion in the nucleus of the solitary tract and spinal cord followingnoxious gastric distention in the rat. Neuroscience 74:873–884.
aters AJ, Lumb BM (2008) Descending control of spinal nociceptionfrom the periaqueductal grey distinguishes between neurons withand without C-fibre inputs. Pain 134:32–40.
illiams S, Evan GI, Hunt SP (1990) Changing patterns of c-fosinduction in spinal neurons following thermal cutaneous stimulationin the rat. Neuroscience 36:73–81.
orkman BJ, Lumb BM (1997) Inhibitory effects evoked from theanterior hypothalamus are selective for the nociceptive responsesof dorsal horn neurons with high- and low-threshold inputs. J Neu-rophysiol 77:2831–2835.
eomans DC, Proudfit HK (1996) Nociceptive responses to high andlow rates of noxious cutaneous heating are mediated by differentnociceptors in the rat: electrophysiological evidence. Pain 68:141–150.
ezierski RP (1991) Somatosensory input to the periaqueductal gray:a spinal relay to descending control center. In: The midbrain peri-aqueductal gray matter: functional, anatomical and neurochemicalorganization (Depaulis A, Bandler R, eds), pp 365–386. New York:Plenum Press.
oshida K, Konishi M, Nagashima K, Saper CB, Kanosue K (2005)Fos activation in hypothalamic neurons during cold or warmexposure: projections to periaqueductal gray matter. Neuroscience
133:1039–1046.(Accepted 11 January 2008)(Available online 19 January 2008)