selective attention: a reevaluation of the implications of negative priming
TRANSCRIPT

Psychological Review Copyright 1998 by the American Psychological Association, Inc. 1998, Vol. 105, No. 2, 203-229 0033-295X/98/$3.00
Selective Attention: A Reevaluation of the Implications of Negative Priming
Bruce Milliken McMaster University
Steve Joordens University of Toronto at Scarborough
Philip M. Merikle University of Waterloo
Adriane E. Seiffert Harvard University
The notion that inhibitory processes play a critical role in selective attention has gained wide support. Much of this support derives from studies of negative priming. The authors note that the attribution of negative priming to an inhibitory mechanism of attention draws its support from a common assumption underlying priming procedures, together with the procedure that has been used to measure negative priming. The results from a sexies of experiments demonstrate that selection between 2 competing prime items is not required to observe negative priming. This result is demonstrated across several experiments in which participants named 1 of 2 items in a second display following presentation of a single-item prime. The implications of these results for existing theories of negative priming are discussed, and a theoretical framework for interpreting negative priming and several related phenomena is forwarded.
Information about the surrounding environment is processed selectively. The area of study that concerns itself with the ability to respond selectively to relevant sources of information, while ignoring irrelevant sources of information, is that of selective attention. Its importance to adaptive behavior, together with its complexity, has made selective attention one of the most studied areas in cognitive psychology.
One of the experimental methods used to study selective atten- tion is commonly known as the priming procedure. This proce- dure requires participants to respond to a probe stimulus that is either related or unrelated to a previous prime stimulus. Differ- ences in the latency or accuracy of responses to related and unrelated probes are often referred to as priming effects. A variety of methods have been used to demonstrate that priming effects can be modulated by attentional processing (e.g., Neely, 1977; Posner & Snyder, 1975; Tipper, 1985). These attentional
Bruce Milliken, Department of Psychology, McMaster University, Hamilton, Ontario, Canada; Steve Joordens, Department of Psychology, University of Toronto at Scarborough, Scarborough, Ontario, Canada; Philip M. Merikle, Department of Psychology, University of Waterloo, Waterloo, Ontario, Canada; Adriane E. Seiffert, Department of Psychol- ogy, Harvard University.
The research reported in this article was supported by the Natural Science and Engineering Research Council of Canada.
We thank Adrienne Rock for help with data collection and Lee Brooks, Jim Debner, Jim Enns, Larry Jacoby, Pierre Jolicoeur, and Steve Tipper for useful discussions of this work. We also thank Tram Neill for helpful reviews on a version of this article.
Correpsondence concerning this article should be addressed to Bruce Milliken, Department of Psychology, McMaster University, Hamilton, Ontario, Canada LSS 4K1. Electronic mail may be sent to millike@ mcmall.cis.mcmaster.ca.
modulations of priming effects are, in turn, used to make infer- ences regarding fundamental attentional mechanisms.
In this article we are concerned primarily with such inferences as those drawn on the basis of a phenomenon known as negative priming (see Neill, 1977; Tipper, 1985). Negative priming is conventionally defined by slowed responses to a probe stimulus that is identical or related to an ignored, distracting dimension of a preceding prime stimulus. This empirical effect has been cited as favoring the view that selective attention is achieved both by amplifying the signal associated with relevant informa- tion and by inhibiting signals associated with irrelevant informa- tion. The notion that focused attention is achieved by actively preventing distraction has a long history in the field of psychol- ogy (James, 1890/1950; see also Broadbent, 1958; Moran & Desimone, 1985; Walley & Weiden, 1973) - - a history that has fueled research on negative priming over the past decade.
The experiments reported in this article focus on the theoreti- cal framework by which negative priming is conventionally at- tributed to an inhibitory selection mechanism. In particular, we report negative priming effects under experimental conditions that challenge this attribution and that warrant a reevaluation of the implications of negative priming for selective attention. The reevaluation that we offer includes a new theoretical framework that accommodates data from studies of negative priming and that can be extended to account for data from several phenomena that appear related to negative priming.
At the heart of our alternative framework is skepticism con- cerning a widely adopted assumption used to interpret priming effects. In particular, the second of two stimuli, the probe, is often regarded as an innocuous measuring instrument of the psychological state of the observer prior to its onset. In contrast to this view, it is assumed that priming effects can be modulated qualitatively by processing that occurs during computation of a response to the probe. From this standpoint, our theoretical
203

204 M1LLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
approach is consistent with recent proposals that priming effects are a phenomenon of memory retrieval (see Kahneman, Treis- man, & Gibbs, 1992; Logan, 1988; Ratcliff & McKoon, 1988; Whittlesea & Jacoby, 1990) and, in particular, with the proposal of Neill, Valdes, Terry, and Gorfein (1992) that retrieval pro- cesses play an important role in determining negative priming.
At the same time, the recent focus on the role of retrieval processes (e.g., Neill et al., 1992) has obscured the role that attention plays in determining negative priming. Our alternative theoretical framework indicates how attentional processing dur- ing retrieval helps to explain the qualitatively different priming effects often observed for previously ignored and previously attended primes. The critical explanatory tool that is inherent in our altemative account, and that is not inherent in prior theo- retical accounts, is an orienting system that detects discrepancies between the perceptual representation of the current probe and the memorial representation of the preceding prime.
We begin with a brief summary of the selective attention and negative priming literature. This discussion is followed by an introduction to a troubling property of negative priming that motivated the experiments and the alternative theory offered here.
Selective Attention
The experimental study of selective attention is often cited as starting with the work of Cherry (1953). In an influential series of studies, Cherry examined the ability of participants to attend to one of two simultaneously presented auditory signals. Two verbal messages were played to participants, one to each ear, and participants were asked to shadow (i.e., repeat aloud) the material that was presented to just one ear. Cherry noted that when participants had finished the shadowing task, they had little memory for the semantic content of the nonshadowed mes- sage. However, participants were often able to recall details regarding the physical characteristics of that message.
Broadbent's (1958) filter theory of selective attention is well- known as a formal account of these and other related data. Broadbent proposed that the gross physical characteristics of external stimuli are analyzed in parallel and that the results of this initial preattentive analysis are subject to a selective filter. The role of the filter is to ensure that only relevant sources of information are allowed further elaborative limited-capacity processing. Access to meaning is purported to be a product of this elaborative processing. As a result, Broadbent's filter model came to be known as an "early" selection model. Selection is early in Broadbent's model because serial processing of stimuli begins at a stage that precedes access to meaning.
A second important property of Broadbent's (1958) model concerns the fate of irrelevant stimuli. Beyond the point at which relevant stimuli are selected for further processing, the products of preattentive processing on irrelevant stimuli are presumed to decay passively. Thus, according to Broadbent's model of selection, the critical distinction between processing of relevant and irrelevant information is that only relevant information is subject to the elaborative processing required to gain access to meaning.
After the publication of Broadbent's (1958) filter model, sev- eral investigators suggested that access to meaning can, in fact,
occur for irrelevant stimuli (e.g., Moray, 1959; Treisman, 1960, 1964). For example, Treisman (1960) reported that participants often continued to shadow a message that was abruptly switched from one ear to the other, even though they had been instructed to attend only to the ear to which the shadowed message was initially presented. That participants are able to switch attention to the message of the nonattended ear on the basis of its semantic content suggests that participants may have earlier access to the meaning of nonattended messages than is proposed in Broad- bent's model.
Findings such as these led to alternative models of selective attention, such as Treisman's (1964) attenuation model and Deutsch and Deutsch's (1963) response selection model. For the most part, these theories of selective attention differed from that of Broadbent (1958), and from one another, in the analysis that was assumed to take place for irrelevant stimuli. However, in the present context this difference can be construed as one of degree rather than of character. That is, all three models (Broadbent, 1958; Deutsch & Deutsch, 1963; Treisman, 1964) characterize unattended information by reference to the level of representation achieved before some form of filtering takes place. Beyond that critical stage, representations of unattended events are presumed not to be processed further. Thus, the criti- cal processing distinction between attended and unattended events is that, beyond some critical stage, active processing of attended events continues, whereas that for unattended events ceases.
Within this framework, attention results in continued ampli- fication of the signal of a relevant stimulus. This attentional amplification may result in a more highly excited internal repre- sentation of an attended stimulus. Morton (1969) suggested that this increased activation level triggers access to higher level semantic information. Thus, recognition of a word is proposed to take place when the internal representation, or logogen, for that concept reaches a threshold state of excitation. Application of this theoretical framework to the issue of selective attention leads to a simple redescription of the selection process. Specifi- cally, selective attention to a relevant stimulus, or target, among other irrelevant alternatives, or distractors, can be presumed to occur when the internal representation of the target is some criterion amount more excited than that of each of the distractors.
It should be evident that the models of selective attention described earlier achieve a criterion difference between target and distractor activation states by the continued active pro- cessing of target items. It should be equally evident that an alternative, and arguably more efficient, way to produce such a difference would be to actively enhance processing of the target and to actively suppress processing of the distractor. In theory, then, selection may be achieved by continued excitatory pro- cessing of a target representation, or it may involve both excit- atory processing of targets and inhibitory processing of distractors.
The notion that inhibitory mechanisms play a role in selective attention has gained considerable support from studies of nega- tive priming (Tipper, 1985). As the negative priming effect is the focus of the empirical work reported in this article, a brief and selective review of the relevant literature is provided. More comprehensive reviews of the negative priming literature have

NEGATIVE PRIMING 205
been provided by Fox (1995), May, Kane, and Hasher (1995), and Neill, Valdes, and Terry (1994).
Negative Pr iming
Negative priming is defined by slow responses to stimuli, or properties of stimuli, that have previously been ignored. This empirical finding was first demonstrated by Dalrymple-Alford and Budayr ( 1966; see also Greenwald, 1972). These investiga- tors found that the time required to name the ink color of color words (Stroop, 1935) presented in lists is particularly slow when the ink color of an item corresponds to the color word of the immediately previous item. Neill (1977) showed subsequently that this negative effect generalizes beyond a whole-list proce- dure to one in which response time is measured on an item-by- item basis.
Negative priming gained wider interest following a series of studies reported by Tipper and colleagues (Allport, Tipper, & Chmiel, 1985; Tipper, 1985; Tipper & Cranston, 1985). They demonstrated that negative priming generalizes beyond the Stroop task, suggesting it may reflect a fundamental characteris- tic of selective attention. In one set of studies, Tipper and Crans- ton showed that response time to identify one of two overlapping letters is slowed when the to-be-named letter is the same as a letter that was ignored in the immediately preceding display (Tipper & Cranston, 1985).
Subsequent studies showed a similar effect in a task requiring identification of one of two overlapping line drawings of natural objects (Allport et al., 1985; Tipper, 1985). Examples of these line drawings are displayed in Figure 1. The critical conditions in Tipper's (1985) Experiment 3 were defined by the relation between target and distractor items in a prime display and the target item in a following probe display. In the attended repeti- tion condition, the prime target was identical to the probe target; in the ignored repetition condition, the prime distractor was identical to the probe target; and in the control condition, the prime and probe items were unrelated to one another.
Tipper (1985) reported that participants responded faster in the attended repetition condition than in the control condition and slower in the ignored repetition condition than in the control condition. These results provided an important link to more conventional studies of priming and, in doing so, highlighted the potential implications of negative priming for selective atten- tion. In particular, Tipper ( 1985; see also Neill, 1977) suggested that selective attention to a target item may be achieved, in part, by the active inhibition of representations associated with distracting items.
The Distractor Inhibition Hypothesis
The logic for attributing negative priming to an inhibitory process is an extension of logic often used to interpret more conventional priming effects. In conventional studies of prim- ing, a probe event follows presentation of an identical or related prime. Typically, responses to the probe are faster or more accu- rate when it is identical (e.g., Jacoby, 1983; Jacoby & Dallas, 1981; Scarborough, Cortese, & Scarborough, 1977; Scarbor- ough, Gerard, & Cortese, 1979) or semantically related to the
prime (e.g., Meyer & Schvaneveldt, 1971; Meyer, Schvane- veldt, & Ruddy, 1975) than when it is unrelated to the prime.
As the terminology of the priming procedure suggests, facili- tated performance for repeated or related probes is commonly attributed to the persisting effect of activation of the internal representation of the prime and its semantic neighbors (Col- lins & Loftus, 1975; but see Becker, Moscovitch, Behrmann, & Joordens, 1997; Joordens & Becker, 1997; Joordens & Besner, 1992; Kahneman et al., 1992; Masson, 1991; McNamara, 1994; Ratcliff & McKoon, 1988; Whittlesea & Jacoby, 1990, for recent discussions of alternatives). By this view, responses are faster to identical or related probes than to unrelated probes because persistence of activation from the prime implies that less activa- tion of the probe's representation is required for it to reach its threshold of recognition (Morton, 1969).
An assumption that underlies this conception of priming phe- nomena is that response times to probes are directly related to the activation state of their internal representations just prior to their onset. By this logic, if slower responses are observed to repeated or related probes than to control probes, then one must conclude that the internal representation of the repeated or re- lated probe was inhibited, rather than excited, as a result of its presentation as a prime. This is precisely the logic underlying the attribution of negative priming to an inhibitory mechanism. Of course, what makes this logic so compelling is the procedure by which negative priming is measured. Specifically, the critical experimental condition used to measure negative priming is one in which the probe target corresponds to the ignored distracting dimension of the previous prime. Selective attention to one of two primes, then, may require the active inhibition of the internal representation of the irrelevant prime to a level below its base- line state. If that subbaseline activation state persists to the time of the probe display, then responses to probes that are identical or related to the ignored prime may be slower than those to control items.
The distinction between a selection process in which the acti- vation state of distractors is suppressed below baseline, and one in which the activation state of distractors decays passively to baseline, is depicted in Figure 2. If priming effects reflect the relative activation states of related and unrelated probes, then negative priming cannot be accounted for by the passive decay model depicted at the top of Figure 2 but can be accounted for by the active inhibitory model as depicted at the bottom of Figure 2.
The Burgeoning Negative Priming Literature
The sprinkle of articles in which negative priming was first documented has more recently given way to a torrent of research activity. Negative priming has now been observed in tasks re- quiring counting (Driver & Tipper, 1989), same-different letter matching (Neill, Lissner, & Beck, 1990), abstract shape match- ing (DeSchepper & Treisman, 1991), lexical decision (Fuen- tes & Tudela, 1992; Yee, 1991), spatial localization (Tipper, Brehaut, & Driver, 1990), and reaching (Tipper, Lortie, & Baylis, 1992), to name a few. To the extent that negative priming in each of these tasks is determined by the same underlying mechanism, the negative priming experimental paradigm is

206 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
PRIMES .oO;;°~
Au un=~II~HED REF,m: u niuOIM
: .,~'° •
IGNORED REPe o, f/ON
PROBE
Figure 1. Examples of the stimuli used by Tipper (1985). Target items are drawn by using solid lines, and distractor items are drawn by using dotted lines. In the actual experiments the stimuli were displayed as red and green. In the control condition, the target and distractor items in the probe display are different from those in the prime display; in the ignored repetition condition, the probe target is the same as the prime distractor. From "The Negative Priming Effect: Inhibitory Effects of Ignored Primes," by S. P. Tipper, 1985, Quarterly Journal of Experimental Psychology, 37A, p. 584. Copyright 1985 by The Experimental Psychology Society. Reprinted by permission of The Experimental Psychology Society.
clearly a very powerful one. That is, it may reflect a highly general and potentially important cognitive process.
At the same time, the growing interest in negative priming has led to a complex empirical literature. One of the conse- quences of this complexity, as is often the case when an empiri- cal phenomenon gains popularity, is that research has been di- rected increasingly at understanding negative priming rather than at understanding selective attention. Despite this empirical fo- cus, a comprehensive account of all negative priming data has proved elusive. For example, Neill and Valdes (1996) recently reviewed six separate accounts of negative priming but con- cluded that none satisfactorily explains all negative priming effects that have been reported.
Two general explanations for why negative priming has defied explanation may be contrasted. On one hand, negative priming may have more than one cause. Indeed, several recent articles have suggested a multideterminant approach in explaining nega- tive priming (Fox, 1995; Kane et al., 1997; May et al., 1995; Milliken, Tipper, & Weaver, 1994; Neill & Valdes, 1996). On the other hand, negative priming may have defied explanation because it is fundamentally misunderstood. The experiments reported in this article suggest that this second possibility should not be dismissed. These experiments were motivated by a prop- erty of negative priming that, in our view, has never been ex- plained satisfactorily.
The Empirical Puzzle
A particularly puzzling property of negative priming was first reported by Lowe (1979; see also Allport et al., 1985; Moore, 1994; Tipper & Cranston, 1985). Using the Stroop procedure (Dalrymple-Alford & Budayr, 1966; Neill, 1977), Lowe showed that whether negative priming occurs can depend on the selec- tion requirements of the probe, the second of two stimuli on each trial. In one experiment, Lowe presented primes that con- tained an incongruent Stroop stimulus (e.g., the word BLUE in red ink) and probes that contained either a second incongruent Stroop stimulus or a simple color patch, comprised of a row of colored disks. When the probe contained a Stroop stimulus, negative priming was observed. However, when the probe con- tained a simple color patch, positive priming was observed.
Tipper and Cranston (1985, Experiment 3; see also Allport et al., 1985, Experiment 9) reported a similar result in a task requiring letter identification. In the ignored repetition condition of their experiment, participants named a single probe letter that was the same as the letter that was ignored in the previous prime display. Responses in this condition were significantly faster than those in the appropriate control condition. Importantly, Tipper and Cranston concluded that this result makes untenable any simple model of inhibition that presumes that the activation state of a distracting item's representation is suppressed below its resting level. Clearly, a comprehensive account of negative
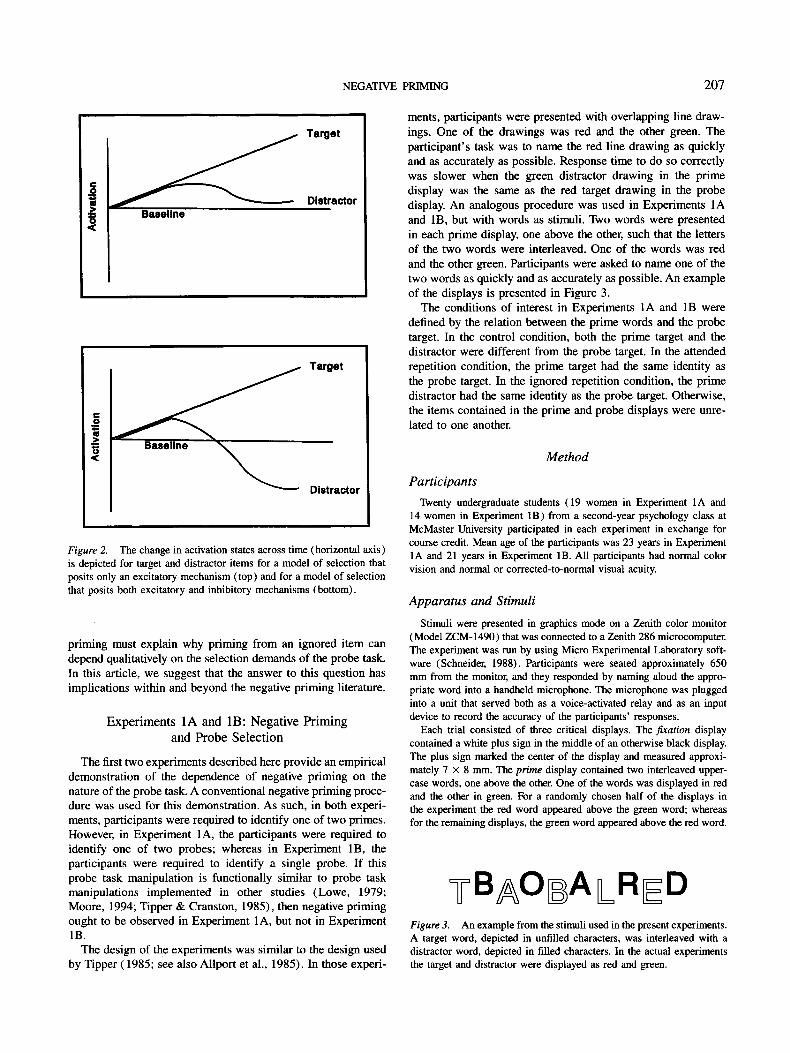
NEGATIVE PRIMING 207
<
Target
Dlstractor Baseline
~ Target
Diutraetor
Figure 2. The change in activation states across time (horizontal axis) is depicted for target and distractor items for a model of selection that posits only an excitatory mechanism (top) and for a model of selection that posits both excitatory and inhibitory mechanisms (bottom).
priming must explain why priming from an ignored item can depend qualitatively on the selection demands of the probe task. In this article, we suggest that the answer to this question has implications within and beyond the negative priming literature.
E x p e r i m e n t s 1A and 1 B: Nega t ive P r iming
and P robe Se l ec t i on
The first two experiments described here provide an empirical demonstration of the dependence of negative priming on the nature of the probe task. A conventional negative priming proce- dure was used for this demonstration. As such, in both experi- ments, participants were required to identify one of two primes. However, in Experiment 1A, the participants were required to identify one of two probes; whereas in Experiment 1B, the participants were required to identify a single probe. If this probe task manipulation is functionally similar to probe task manipulations implemented in other studies (Lowe, 1979; Moore, 1994; Tipper & Cranston, 1985), then negative priming ought to be observed in Experiment 1A, but not in Experiment lB.
The design of the experiments was similar to the design used by Tipper (1985; see also Allport et al., 1985). In those experi-
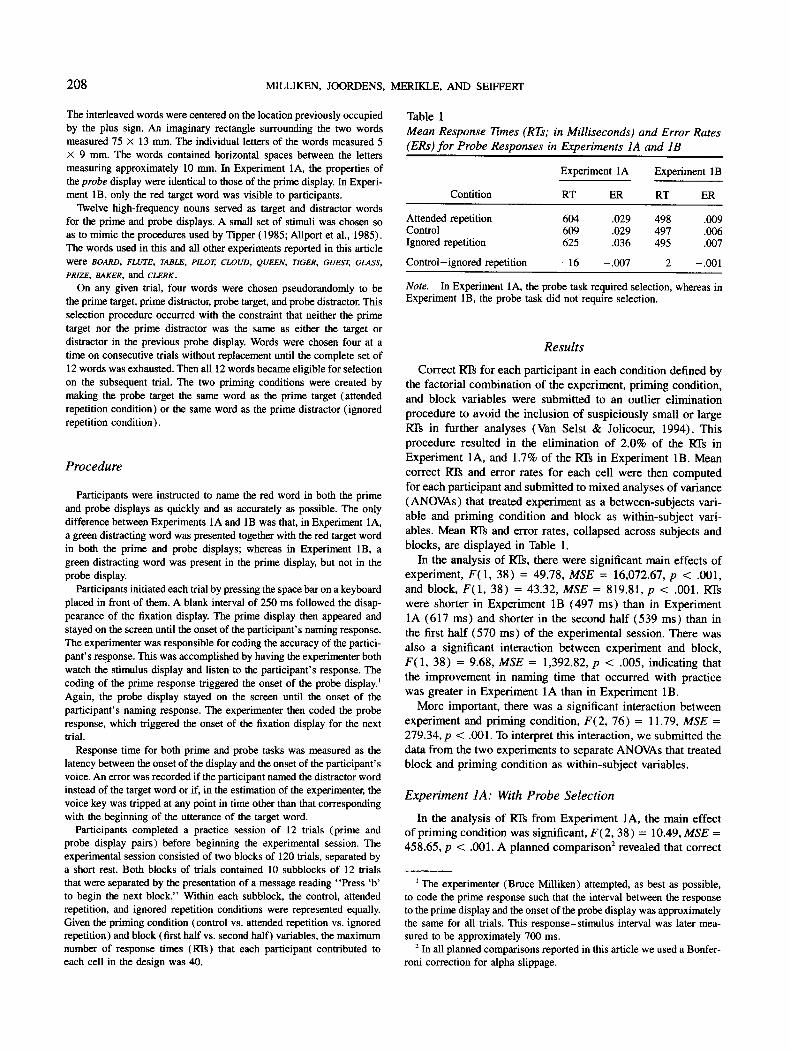
ments, participants were presented with overlapping line draw- ings. One of the drawings was red and the other green. The participant's task was to name the red line drawing as quickly and as accurately as possible. Response time to do so correctly was slower when the green distractor drawing in the prime display was the same as the red target drawing in the probe display. An analogous procedure was used in Experiments 1A and 1B, but with words as stimuli. Two words were presented in each prime display, one above the other, such that the letters of the two words were interleaved. One of the words was red and the other green. Participants were asked to name one of the two words as quickly and as accurately as possible. An example of the displays is presented in Figure 3.
The conditions of interest in Experiments 1A and 1B were defined by the relation between the prime words and the probe target. In the control condition, both the prime target and the distractor were different from the probe target. In the attended repetition condition, the prime target had the same identity as the probe target. In the ignored repetition condition, the prime distractor had the same identity as the probe target. Otherwise, the items contained in the prime and probe displays were unre- lated to one another.
Method
Participants
Twenty undergraduate students (19 women in Experiment 1A and 14 women in Experiment IB) from a second-year psychology class at McMaster University participated in each experiment in exchange for course credit. Mean age of the participants was 23 years in Experiment 1A and 21 years in Experiment lB. All participants had normal color vision and normal or corrected-to-normal visual acuity.
Apparatus and Stimuli
Stimuli were presented in graphics mode on a Zenith color monitor (Model ZCM-1490) that was connected to a Zenith 286 microcomputer. The experiment was run by using Micro Experimental Laboratory soft- ware (Schneider, 1988). Participants were seated approximately 650 mm from the monitor, and they responded by naming aloud the appro- priate word into a handheld microphone. The microphone was plugged into a unit that served both as a voice-activated relay and as an input device to record the accuracy of the participants' responses.
Each trial consisted of three critical displays. The fixation display contained a white plus sign in the middle of an otherwise black display. The plus sign marked the center of the display and measured approxi- mately 7 × 8 mm. The prime display contained two interleaved upper- case words, one above the other. One of the words was displayed in red and the other in green. For a randomly chosen half of the displays in the experiment the red word appeared above the green word; whereas for the remaining displays, the green word appeared above the red word.
TBAO[BA LRED Figure 3. An example from the stimuli used in the present experiments. A target word, depicted in unfilled characters, was interleaved with a distractor word, depicted in filled characters. In the actual experiments the target and distractor were displayed as red and green.
208 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
The interleaved words were centered on the location previously occupied by the plus sign. An imaginary rectangle surrounding the two words measured 75 × 13 mm. The individual letters of the words measured 5 × 9 mm. The words contained horizontal spaces between the letters measuring approximately 10 mm. In Experiment 1A, the properties of the probe display were identical to those of the prime display. In Experi- ment 1B, only the red target word was visible to participants.
Twelve high-frequency nouns served as target and distractor words for the prime and probe displays. A small set of stimuli was chosen so as to mimic the procedures used by Tipper (1985; Allport et al., 1985). The words used in this and all other experiments reported in this article were BOARD, FLUTE, TABLE, PILOT, CLOUD, QUEEN, TIGER, GUEST, GLASS, PRIZE, BAKER, and CLERK.
On any given trial, four words were chosen pseudorandornly to be the prime target, prime distractor, probe target, and probe distractor. This selection procedure occurred with the constraint that neither the prime target nor the prime distractor was the same as either the target or distractor in the previous probe display. Words were chosen four at a time on consecutive trials without replacement until the complete set of 12 words was exhausted. Then all 12 words became eligible for selection on the subsequent trial. The two priming conditions were created by making the probe target the same word as the prime target (attended repetition condition) or the same word as the prime distractor (ignored repetition condition).
Procedure
Participants were instructed to name the red word in both the prime and probe displays as quickly and as accurately as possible. The only difference between Experiments 1A and 1B was that, in Experiment 1A, a green distracting word was presented together with the red target word in both the prime and probe displays; whereas in Experiment 1B, a green distracting word was present in the prime display, but not in the probe display.
Participants initiated each trial by pressing the space bar on a keyboard placed in front of them. A blank interval of 250 ms followed the disap- pearance of the fixation display. The prime display then appeared and stayed on the screen until the onset of the participant's naming response. The experimenter was responsible for coding the accuracy of the partici- pant's response. This was accomplished by having the experimenter both watch the stimulus display and listen to the participant's response. The coding of the prime response triggered the onset of the probe display) Again, the probe display stayed on the screen until the onset of the participant's naming response. The experimenter then coded the probe response, which triggered the onset of the fixation display for the next trial.
Response time for both prime and probe tasks was measured as the latency between the onset of the display and the onset of the participant's voice. An error was recorded if the participant named the distractor word instead of the target word or if, in the estimation of the experimenter, the voice key was tripped at any point in time other than that corresponding with the beginning of the utterance of the target word.
Participants completed a practice session of 12 trials (prime and probe display pairs) before beginning the experimental session. The experimental session consisted of two blocks of 120 trials, separated by a short rest. Both blocks of trials contained 10 subblocks of 12 trials that were separated by the presentation of a message reading "Press 'b' to begin the next block." Within each subblock, the control, attended repetition, and ignored repetition conditions were represented equally. Given the priming condition (control vs. attended repetition vs. ignored repetition) and block (first half vs. second half) variables, the maximum number of response times (RTs) that each participant contributed to each cell in the design was 40.
Table 1
Mean Response Times (RTs; in Milliseconds) and Error Rates (ERs) for Probe Responses in Experiments 1A and 1B
Experiment 1A Experiment 1B
Contition RT ER RT ER
Attended repetition 604 .029 498 .009 Control 609 .029 497 .006 Ignored repetition 625 .036 495 .007
Control-ignored repetition - 16 -.007 2 -.001
Note. In Experiment 1A, the probe task required selection, whereas in Experiment 1B, the probe task did not require selection.
Results
Correct RTs for each participant in each condition defined by the factorial combinat ion of the experiment, priming condition, and block variables were submit ted to an outlier el imination procedure to avoid the inclusion of suspiciously small or large RTs in further analyses (Van Selst & Jolicoeur, 1994). This procedure resulted in the el imination of 2.0% of the RTs in Experiment 1A, and 1.7% of the RTs in Experiment lB. Mean correct RTs and error rates for each cell were then computed for each participant and submitted to mixed analyses of variance (ANOVAs) that treated experiment as a between-subjects vari- able and pr iming condition and block as within-subject vari- ables. Mean RTs and error rates, collapsed across subjects and blocks, are displayed in Table 1.
In the analysis of RTs, there were significant main effects of experiment, F (1 , 38) = 49.78, MSE = 16,072.67, p < .001, and block, F (1 , 38) = 43.32, MSE = 819.81, p < .001. RTs were shorter in Experiment 1B (497 ms) than in Experiment 1A (617 ms) and shorter in the second half (539 ms) than in the first half (570 ms) of the experimental session. There was also a significant interaction between experiment and block, F (1 , 38) = 9.68, MSE = 1,392.82, p < .005, indicating that the improvement in naming time that occurred with practice was greater in Experiment 1A than in Experiment lB.
More important, there was a significant interaction between experiment and pr iming condition, F (2 , 76) = 11.79, MSE = 279.34, p < .001. To interpret this interaction, we submitted the data f rom the two experiments to separate ANOVAs that treated block and pr iming condition as within-subject variables.
Experiment 1A: With Probe Selection
In the analysis of RTs from Experiment 1A, the main effect of pr iming condition was significant, F ( 2 , 38) = 10.49, MSE = 458.65, p < .001. A planned comparison 2 revealed that correct
i The experimenter (Bruce Milliken) attempted, as best as possible, to code the prime response such that the interval between the response to the prime display and the onset of the probe display was approximately the same for all trials. This response-stimulus interval was later mea- sured to be approximately 700 ms.
2 In all planned comparisons reported in this article we used a Bonfer- roni correction for alpha slippage.

NEGATIVE PRIMING 209
responses were significantly slower in the ignored repetition condition (625 ms) than in the control condition (609 ms), t (19) = 3.43, p < .01. However, the positive difference between RTs in the control condition and those in the attended repetition condition (604 ms) was not statistically reliable (p > .10).
Experiment 1B: Without Selection
In the analysis of RTs from Experiment 1B, the main effect of priming condition was not significant. However, given its theoretical importance, a planned comparison was performed for RTs in the control (497 ms) and ignored repetition (495 ms) conditions. This comparison revealed a nonsignificant dif- ference (p > .10). The difference in mean RT between the control and attended repetition conditions was also not signifi- cant ( F < 1).
In the overall analysis of error rates, there was only one significant effect. More errors were made in Experiment 1A (.032) than in Experiment 1B (.007), F(1 , 38) = 9.41, MSE = 37.90, p < .005.
Discussion
The critical difference between Experiments 1A and 1B is the contrast between ignored repetition effects in the two experi- ments. When the probe task required participants to name a red target and to ignore a green distractor (Experiment 1A), nega- tive priming was observed. In contrast, when the probe task required the naming of a lone red target word (Experiment 1B), negative priming was not observed. These different patterns of results are consistent with previous research that has demon- strated a dependence of priming effects on the selection require- ments of the probe task (Lowe, 1979; Moore, 1994; Tipper & Cranston, 1985). As such, the results of Experiments 1A and 1B demonstrate that the interleaved word procedure leads to ignored repetition effects that behave in a manner similar to those measured with overlapping letters and pictures (Tipper, 1985; Tipper & Cranston, 1985) and Stroop stimuli (Lowe, 1979; Neill, 1977).
A second result from Experiments 1A and 1B deserves com- ment. It may seem surprising that significant positive priming was not observed in the attended repetition condition in either experiment. This result is not consistent with the results reported by Tipper (1985), who used overlapping line drawings. In sev- eral experiments, Tipper (1985; see also Allport et al., 1985) observed that positive priming in the attended repetition condi- tion was larger than the corresponding negative priming effect found in the ignored repetition condition. Although the attended repetition effect in Experiment 1A could conceivably have reached significance had the experiment more power, negative priming that was due to ignored repetition is clearly the larger of the two effects. This anomaly is noteworthy and is addressed both in Experiment 4 and in the General Discussion section.
Account ing for the Dependence of Negative Pr iming on Probe Select ion
Much research on negative priming has been carried out with- out regard for the dependence of negative priming on probe
selection (but see Lowe, 1979; Milliken & Joordens, 1996; Moore, 1994; Neill, Terry, & Valdes, 1994; Neill & Westberry, 1987; Tipper & Cranston, 1985). At the same time, a compre- hensive theoretical account of negative priming must explain this dependence. Two such extant accounts play an important role in theoretical discussion in this article. These two accounts are described in turn and are then contrasted with a novel alter- native framework for interpreting negative priming.
The Response Blocking Account
Tipper and Cranston (1985) suggested that inhibitory pro- cesses may not suppress the activation state of the representation of a distractor but instead may block the translation of an active perceptual representation into a response code. By this view, selection between target and distractor at the time of the prime display establishes the response block. When a probe task also requires selection between a target and distractor, Tipper and Cranston suggested that participants maintain the "selection state" adopted in the prime task to cope with the selection demands of the probe task. The response blocking mechanism is presumed to be part of the selection state and is therefore maintained for probes that require selection. However, the selec- tion s ta te--and thus the blocking of response to the previous prime distractor--can be abandoned for probes that do not require selection. With the abandonment of the selection state, an otherwise active perceptual code is revealed, thus explaining why positive priming can occur for an ignored prime.
Tipper and Cranston's ( 1985 ) selection state may be regarded as a strategic attentional set, one element of which is the re- sponse blocking process that causes negative priming. Indeed, previous studies of the role of probe selection indicate that strategic factors do play a role. For example, one might reason- ably ask whether negative priming is related to response conflict that occurs when a probe contains a distractor that competes with the target for the control of action. Lowe (1979) first investigated this issue in a Stroop color naming task. When probes that could not elicit response conflict (e.g., a random letter string presented in colored ink) and probes that could elicit response conflict (e.g., a Stroop stimulus) were mixed randomly within an experimental session, so that they were difficult to discriminate from one another, negative priming oc- curred for both probe types. However, when the conflict and nonconflict probes were presented in separate blocks, negative priming was observed for conflict probes, and positive priming was observed for nonconflict probes. Strikingly similar data were reported recently by Moore (1994). Together, the data of Lowe and Moore suggest that the dependence of negative prim- ing on probe selection does not occur as a direct result of competition between target and distractor. Rather, the selection task appears to influence the strategic set of participants, and negative priming hinges on this strategic set.
The Episodic Retrieval Account
Neill et al. (1992; see also Neill & Valdes, 1992) offered an account of negative priming in which retrieval of episodic repre- sentations plays a critical role. Their model borrows from Logan's (1988) instance theory of automaticity. Logan suggested that per-

210 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
formance is determined by the faster of two processes: an auto- matic retrieval process and a more controlled algorithmic process. By this view, onset of a probe stimulus cues the retrieval of previous processing instances involving that stimulus. Neill et al. (1992) proposed that if response information is encoded as part of the prime episode, then responses to previously ignored probes may be slowed by the retrieval of that episode. For example, the prime episode may contain an ignore-this~stimulus tag that is linked to the prime distracto~ On ignored repetition trials, this response information may be retrieved and lead to conflict with the current goal of responding to the probe target. Altematively, Neill et al. suggested that failure to encode response information for the prime distractor may lead participants to rely on the slower algorithmic processing route rather than the faster automatic pro- cessing route. In either case, slower responses to probes that corre- spond to previously ignored primes are explained by reference to an automatic retrieval process that hinders rather than helps performance.
The dependence of negative priming on probe selection can be accommodated by the episodic retrieval account in either of two ways. Both make reference to an increased contribution of automatic retrieval when the probe task requires selection. One possibility is that the selection task slows the algorithmic pro- cess, which in turn would result in the automatic retrieval pro- cess winning the processing race on a larger proportion of trials. A second possibility is that the contextual similarity of a prime task that requires selection and a probe task that also requires selection results in a greater contribution of automatic retrieval to performance than occurs when the probe task does not require selection. In either case, if retrieval of conflicting response infor- mation causes negative priming, then an increased contribution of automatic retrieval to performance ought to increase negative priming.
A Temporal Discrimination Account
In contrast to the two theoretical accounts described earlier, we propose that negative priming does not provide a measure of the prior act of "selecting against" a prime distractor. Instead, we argue that negative priming is an emergent consequence of a dis- crimination process that is inherent to memory retrieval. 3 Impor- tantly, this discrimination process has an attentional basis. As such, our alternative account explains negative priming by reference to the attentional modulation of a retrieval process.
For the purpose of introduction to this discrimination process, consider the experience of suddenly realizing that an appreciable stretch of highway has been navigated successfully without the abihty to remember the details of the environment through which the highway passes. This phenomenon is relatively common, and yet it may alarm the driver. What is alarming about this experience is the thought that had an unpredictable event occurred during that stretch of driving, it would not have been responded to appro- priately because of inattentiveness. Although this concern is not entirely unwarranted, it may be exaggerated by the belief that if perceptual detail cannot be remembered, then it was not analyzed and encoded, and, further, that it would not have been encoded even if something unexpected were to have occurred. In fact, at the first sign of an unexpected event, an orienting system is likely to alert the driver that attentive perceptual analysis is required. In
practice, then, the driver is unlikely to miss the flashing red signal indicating a rare crossing of highways.
This anecdote provides one answer to how people cope with more information than can possibly be analyzed at any one point in time. In many situations, behavior does not rely on detailed perceptual analysis but instead proceeds relatively "automati- cally" by virtue of the retrieval of prior well-learned actions. When prior learning is an inappropriate basis for action, an orient- ing system alerts the observer to engage in more detailed percep- tual analysis. In other words, novel or unexpected stimuli may be responded to on the basis of new learning rather than on reinstated old learning. By this general view, the discrimination of a stimulus as known or unknown must play a critical role in determining how a response to that stimulus is computed.
With this general framework in mind, we suggest that negative priming results from an attention system "deciding" whether a response to a probe stimulus is "known" and can be retrieved directly from memory or whether it must be "learned" by more analytic means. Thus, in agreement with Neill et al. (1992; see also Logan, 1988), a distinction between two different pro- cessing solutions is central to the temporal discrimination frame- work forwarded here. Which of two processing solutions guides behavior is determined by the categorization of the probe target as "old" or "new." Response to a probe categorized as old is retrieved directly from memory, whereas response to a probe categorized as new relies on perceptual analysis. Most im- portant, ambiguity in this temporal discrimination process can impede performance.
To apply this logic, one must consider the three types of probes often tested in studies of negative priming. (a) In the control condition, the probe target is unrelated to the previous prime. The temporal distinctiveness of an unrelated probe target is presumed to facilitate its categorization as new. (b) In the attended repetition condition, the probe target is related to the attended prime target. The similarity of the probe target to an attended prime is presumed to facilitate its categorization as old. Finally, (c) in the ignored repetition condition, the probe target is related to the previously ignored prime distractor. The familiarity of the probe target is presumed to rule out its quick categorization as new but at the same time is an insufficient basis for its categorization as old. This ambiguity in the temporal discrimination process for ignored repetition trials is presumed to underlie negative priming.
The bases for ambiguity in a temporal discrimination process are noted explicitly in Figure 4. An orienting system is presumed to be responsible for detecting differences between a current probe and the memorial representation of a preceding prime, whereas an automatic retrieval process is responsible for de- tecting similarity between a current probe and a preceding prime. Stated in terms consistent with the notion of temporal discrimination, the orienting system is responsible for temporal
3 The temporal discrimination account forwarded here can be regarded as one specific method of explaining interference by reference to oppo- nent processes. Although opponent process have been discussed widely in psychology, our adoption of such a framework here can be traced to the joint influence of Johnston and Hawley (1994) and a personal communication with Steve Link (March 1996) regarding psychophysical studies of the discrimination of same from different.

NEGATIVE PRIMING 211
Orienting Automatic Retrieval
Attended Repetition Ignored Repetition Unrepeated (Control) +
+
Figure 4. The three conditions often tested in studies of negative prim- ing are listed together with an indication of whether that condition would benefit (depicted with a + sign) or not benefit (depicted with a - sign) from an orienting process and from an automatic retrieval process.
differentiation, whereas the retrieval system is responsible for temporal integration. The ambiguity that results in negative priming may then occur when these two opponent processes are weighted relatively equally.
It is worth noting that the temporal discrimination process intro- duced here can also be regarded as a process of comparison between the perceptual representation of the probe and the memo- rial representation of the preceding prime. This comparison pro- cess results in either a "same" or a "different" decision. The property of this comparison process that provides the basis for explaining negative priming is that the time required to make a decision can be nonmonotonically related to the match between representations. This same property has been noted in the psycho- physical literature on same-different discrimination (Cart- wright & Festingel; 1943; Link, 1992), in the literature on sentence verification (Kintsch, 1980), and can be demonstrated by refer- ence to many decision processes that confront us from day to day. Consider the decision that regularly confronts the consumer. The product that matches perfectly with the taste of the consumer may be bought quickly, whereas the product that mismatches starkly with the taste of the consumer may be discarded just as quickly. In contrast, a product that neither matches perfectly nor mismatches starkly with the taste of the consumer may be pondered over for some time before a decision to buy or discard is made.
Alternatively, consider the task taken on by a graduate student entering a doctoral program. Some would argue that the efficiency with which a student completes such a program depends, in part, on the "match" between the interests of the student and those of the laboratory in which the student works. Of course, the best possible scenario is that the interests of the student match perfectly with those of the laboratory chosen on entrance to the program. However, in some cases the match is imperfect and resuks in a switch to a different laboratory. Note that, in many circumstances, for the student who switches laboratories it is best to detect the initial mismatch in interests quickly. The quick detection of a mismatch ensures that progress is not delayed appreciably.
Again, the point of most relevance to the present discussion is that, under some conditions, the time required to complete the program will be nonmonotonically related to the quality of the initial match between student and laboratory. As depicted at the top of Figure 5, one possible scenario is that a good initial match results in the fastest completion of the program, a poor initial match results in slightly slower completion of the pro- gram, and an intermediate quality initial match, which in turn results in a slow switch to a new laboratory, results in slower
completion still. We draw a parallel between the intermediate quality match between student and laboratory, which can result in delayed program completion, and the intermediate quality match between the perceptual representation of a probe target and the memorial representation of an ignored prime. In particu- lar, we assume that negative priming occurs because this match may be insufficient to result in a response based on memory retrieval but is sufficient to interfere with an orienting process.
In the example provided here, note that the nonmonotonic relation between the quality of initial match and the time re- quired to finish the program is a product of the discrimination process itself. In other words, if the student were to enter the program and not discriminate between a good and a poor match, instead persevering through the program regardless of the qual- ity of match, one might expect the time to complete the program to be directly and monotonically related to the quality of that match--good matches resulting in short completion times, and poor matches resulting in long completion times (see bottom of Figure 5).
One of the critical issues addressed in this article concerns the dependence of negative priming on probe selection. When a probe task does not require selection, negative priming is often not observed. To explain this result, we assume that a probe selection task forces the participant to engage in a discrimination process that is analogous to that of the careful graduate student. In the case of the graduate student, discriminating between a good match and a poor match may be particularly critical if failure to discriminate is likely to result in a setback at some later point in time. On the other hand, if a laboratory has gained the reputation that its students are highly likely to succeed, then the student may well not be as vigilant to a potential mismatch. In other words, whether the time required to complete the pro- gram is monotonically or nonmonotonically related to the qual- ity of match between student and laboratory is determined by the discriminative strategy of the participant.
By analogy, we assume that requiring participants to respond selectively to one of two probes introduces a source of error to performance. In response to this potential error, participants adopt the strategy of using automatic retrieval processes dis- criminatively. For example, when a probe task contains both a target and a distractor, participants rely on automatic retrieval processes only when the match to a retrieved representation is of high quality. The motive for this discriminative strategy is the avoidance of naming responses that correspond to the probe distractor rather than to the probe target. The discriminative use of automatic retrieval then results in the nonmonotonic relation between performance and the quality of match between the probe and memory (see top of Figure 5). In contrast, when the probe task requires identification of a lone probe word, participants may be much less discriminative of the match be- tween the probe and memory, resulting in performance that is monotonically related to the match between the probe and mem- ory (see bottom of Figure 5).
Temporal Discrimination: Its Relation to Other Theories
The temporal discrimination process described here is subject to the participant's control and, as such, can be implemented

2 1 2 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
Quality of Match Time to completion
G o o d (Attended Repetition)
Intermediate (i~aor~ Rq~aon)
Poor ( u ~ t e d )
Mismatch deU~l j/(o~e~g r~ponsO
Mi~atch detected
Time to completion Quality of Match
Good I I
I I Intermediate
Poor ,
Figure 5. The hypothetical relation between the time required to complete a graduate program and the quality of match between the interests of the student and laboratory. The total time to complete the program is represented by the horizontal length of the three thin rectangles, with the time that passes before a mismatch detection occurs represented by the length of the black portion of the rectangles. At the top of the figure, it is assumed that the student actively discriminates whether the quality of match satisfies a criterion and will switch laboratories if the criterion is not met. Under some conditions, this discriminative strategy can result in a nonmonotonic relation between the quality of initial match and the time required to complete the program. The labels in parentheses indicate the parallel between the discriminative strategy of the graduate student and the attentional strategy that results in positive priming for attended repetition trials and negative priming for ignored repetition trials. At the bottom of the figure, the assumed monotonic relation between quality of match and time to completion is depicted. This monotonic relation is presumed to parallel the positive priming effects that occur without the discriminative attentional set.
as part of a strategic attentional set. Given that both an atten- tional set and automatic retrieval processes are implicated in this theoretical account, it is worth noting explicitly the relation between the temporal discrimination account and the response blocking and episodic retrieval accounts described earlier. The characterization of these three accounts presented in Figure 6 has two purposes. One purpose is to make clear that the temporal
discrimination account draws on ideas that have played a forma- tive role in prior theoretical work (Neill et al., 1992; Tipper & Cranston, 1985). Note again that both an attentional set, which plays an important role in Tipper and Crans ton ' s response blocking account, and automatic retrieval processes, which play a critical role in Neill et al. 's episodic retrieval account, are adopted within the temporal discrimination account of negative

NEGATIVE PRIMING 213
Resuonse Blockin2 Euisodic Retrieval 1. Response Block 1. "Ignore this stimulus" tag 2. Attentional Set [
(Selection State) ] [2. Automatic Retrieval]
. Temporal Discrimination ]
(temporal integration)
Figure 6. The temporal discrimination framework acknowledges the importance of an attentional set, as pointed out in Tipper and Cranston's (1985) response blocking account, and of automatic retrieval processes, as pointed out in Neill et al.'s (1992) episodic retrieval account. How- ever, response blocks and tags that putatively result from ignoring a prime do not play a role in the temporal discrimination framework.
priming. The second purpose is to make clear the distinction between previous accounts and the temporal discrimination ac- count forwarded here. To this end, note that two mechanisms often used to explain negative priming, a response block and an ignore-this-stimulus tag, do not play a role in the alternative framework forwarded here.
Negative Priming Without Prime Selection
The most noteworthy property of the temporal discrimination account forwarded here is that it explains negative priming with- out reference to selection processes that occur during the prime task. At the same time, note that it must be the case that atten- tional processing during the prime task can modulate priming effects; the qualitative contrast between ignored and attended repetition effects reported by Tipper (1985) and others is a provocative and important finding.
However, rather than attribute positive priming to an excit- atory attentional process, and negative priming to an inhibitory attentional process, we propose that both effects may result from changes in the contribution to performance of a single process. In particular, the contribution of automatic retrieval to performance may be greatest in the attended repetition condi- tion, least in the unrepeated condition, and intermediate in the ignored repetition condition. This assumption states little more than that attention to a prime later facilitates its retrieval. As a result, the attention afforded a prime target in an attended repeti- tion condition may facilitate the categorization of the probe target as old, resulting in a relatively fast reinstatement of a prior learned response. In the unrepeated condition, the opposite decision may also be reached relatively quickly. In other words, a probe may be categorized as new relatively quickly when it is unrelated to a preceding prime. In this case, the quick categorization of the probe as new owes to relatively little oppo- sition from automatic retrieval, a process that provides evidence in favor of the opposite decision. Finally, this discrimination
process may resolve itself particularly slowly when the contribu- tion of the retrieval process is intermediate to that in the other two conditions. Again, on ignored repetition trials, the similarity of the probe target to a previously ignored prime may trigger retrieval processes that interfere with the categorization of the probe as new but that are insufficient to support retrieval of the correct response to the probe target.
In summary, we suggest that the contribution of automatic retrieval processes to performance in a negative priming task may vary along a single dimension for attended repetition, ig- nored repetition, and unrepeated conditions. Further, when parti- cipants engage in a discriminative attentional strategy that mod- ulates the contribution of automatic retrieval processes to perfor- mance, the contribution of automatic retrieval to a processing solution may be nonmonotonically related to response time. This discrimination process explains how the repetition effect observed for an unattended prime may differ qualitatively from that for an attended prime. This logic is critical to the present argument because it provides a method of attributing attended repetition effects to attentional processing of a prime, and ig- nored repetition effects to the absence of attentional processing of a prime, rather than to inhibition, a response block, or an ignore-this-stinmlus tag.
If the above-indicated theory is correct, then it ought to be possible to demonstrate negative priming without requiring par- ticipants to select overtly between a prime target and a distractor. Indeed, Milliken and Joordens (1996) reported two experiments in which participants were required to name one of two words following presentation of two briefly presented prime words. In contrast to the conventional procedure used to measure negative priming, participants were not required to respond to the prime display and were instead asked to respond only to the probe target. Negative priming was observed when the probe target was the same as either of the two primes. Of course, these results do not preclude the possibility that participants select in favor of one of the two primes and against the other prime, despite not being required to do so (see also Park & Kanwisher, 1994; Ortells & Tudela, 1996). This alternative hypothesis was ruled out by the experiments reported in this article. In our experiments, participants were required to respond to one of two interleaved probe words that followed the presentation of a single prime. Clearly, if negative priming can be observed following presentation of a single prime, then the argument that negative priming is directly related to the overt selection against a distractor and consequent response blocks or ignore-this-stim- ulus tags must be incorrect.
Experiments 2A, 2B, and 2C: Negative Pr iming Fol lowing Br ie f Single Pr imes
In these experiments, participants were required to respond by naming a red probe target, and ignoring a green probe dis- tractor, in displays identical to those used in Experiment 1A. Importantly, the prime displays did not contain interleaved red and green words. Rather, each prime display consisted of a single, briefly presented white word that was pattern masked both before and after its onset. Participants were not informed of the presence of the prime words until after the experimental session, and they typically expressed surprise when so informed.

214 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
The logic underlying the experiments was as follows. If a prime item must actively be selected against in favor of a prime target for negative priming to occur, then negative priming should not occur in these experiments. In other words, the use of a single, briefly exposed and masked prime allowed us to test whether negative priming might occur when a prime is presented but not attended rather than presented and "selected against." Note that our logic suggests a similarity between items
presented for brief durations and items presented for longer durations that are not attended. Data that support this view are provided later in this article (see also Merikle & Joordens, 1997).
Method
Participants
Twenty undergraduate students (12 women) participated in Experi- ment 2A, 24 undergraduate students ( 14 women) participated in Experi- ment 2B, and 20 undergraduate students (12 women) participated in Experiment 2C. All participants were drawn from the introductory psy- chology participant pool at McMaster University, and all participants took part in exchange for course credit. The mean age of participants was 19, 19, and 20 years in Experiments 2A, 2B, and 2C, respectively. All participants had normal color vision and normal or corrected-to- normal visual acuity.
Apparatus and Stimuli
The apparatus for all three experiments was identical to that used in Experiments 1A and 1B, as was the set of 12 words from which the prime and probe words were drawn. The stimuli were identical in Experi- ments 2A and 2B and were just slightly different in Experiment 2C, as described below.
Experiments 2A and 2B. The first display on each trial contained the fixation marker. The last display on each trial contained the probe stimulus. These two displays were identical to those used in Experiment 1A. The prime displays in these experiments actually consisted of three separate displays: a premask, a prime word, and a postmask, in sequence. The pre- and postmasks consisted of the symbols X, O, and & overlaying one another in the five spatially separate locations in which the letters of the prime could appear. The single prime word was the same size and format as the words that made up the probe display.
Experiment 2C. The stimuli used in Experiment 2C were identical to those used in Experiments 2A and 2B until the point in time at which the probe display appeared on the screen. In Experiment 2C, the probe display contained a rectangle that surrounded the probe target and dis- tractor words. The rectangle was red for half of the trials and green for half of the trials, and it measured 83 × 12 mm.
In all three experiments, three words were chosen pseudorandomly from the stimulus set on each trial and were assigned the roles of prime, probe target, and probe distractor. This selection procedure occurred with the constraint that the prime word not be the same as either the previous probe target or distractor. Words were chosen three at a time on consecutive trials without replacement until the complete list of 12 words was exhausted, after which all 12 words became eligible for selection on the subsequent trial. Repeated trials were created by as- signing the prime to be the same word as the probe target.
Procedure
Participants were instructed to focus their attention on the fixation marker at the beginning of each trial and to press the space bar to initiate
Figure 7. The procedure used in Experiment 2A. Participants started each trial by pressing the space bar on the computer keyboard and responded by naming the red item (unfilled letters) in just the probe display. The distracting item in both displays was green.
a trial. Participants were told that the first thing they would see on each trial would be a series of letter markers and that these letter markers (the premask, brief prime, and postmask) would flicker. Participants were told that they were to focus their attention on these letter markers because they indicated the location of the middle of the screen, which was where the probe display would appear. On the appearance of the probe display, participants were to name aloud the target word as quickly and as accurately as possible.
The timing of the sequence of displays is depicted in Figure 7. The fixation display remained on the screen until the participant initiated a trial. A blank interval of 500 ms followed. The premask then appeared for 500 ms, followed immediately by the prime word for 33 ms. The prime word was followed by a 500-ms postmask. After the postmask, the probe display appeared and remained on the screen until the onset of the response. The experimenter then coded the response, which trig- gered the onset of the fixation marker for the next trial.
At the end of the experimental session, participants were asked if they noticed anything peculiar about the letter markers (the masked prime). After informing the experimenter of their impression of the masked prime, participants were informed that the prime in fact consisted of a briefly presented and masked word. Participants were then asked again if they were aware at anytime during the experimental session that this had occurred. Participants who reported awareness of the prime words were asked to estimate approximately how often they were aware of the identity of the prime words. Also, they were asked if they noticed any relation between the prime display and probe target word.
Two critical conditions were tested in all three experiments. In the unrepeated condition, the probe target and distractor were different from the briefly presented prime. In the repeated condition, the probe target was the same as the briefly presented prime. In Experiments 2A and 2C, there were an equal number of unrepeated and repeated trials, whereas in Experiment 2B there were twice as many unrepeated as repeated trials. Thus, Experiments 2A and 2B were identical in procedure with the exception of the proportion of unrepeated to repeated trials.
The procedure for Experiment 2C differed from that for Experiment 2A only in the nature of the probe task. Whereas in Experiments 2A and 2B the target was always the red probe word, and the distractor always the green probe word, in Experiment 2C the probe target was red on half of the trials and green on the other half of the trials. The target color for each trial in Experiment 2C was defined by the color of the rectangular selection cue that surrounded the probe target and dis- tractor. If the selection cue was red, then participants were to name the

NEGATIVE PRIMING 215
red probe word; if the selection cue was green, then participants were to name the green probe word.
Participants in all experiments completed a practice session of 12 trials before beginning the experimental session. The experimental ses- sion consisted of 2 blocks of 120 trials, separated by a short rest. Each block contained 10 subblocks of 12 trials each. Within each subblock, the proportion of repeated to unrepeated trials was the same as for the experiment as a whole. Given the priming condition (unrepeated vs. repeated) and block (first half vs. second half) variables, the maximum number of RTs contributed by each participant to each cell in the design was 60 for the unrepeated and repeated conditions of Experiments 2A and 2C, 40 for the repeated condition of Experiment 2B, and 80 for the unrepeated condition of Experiment 2B.
Results
Participants' Reports o f Prime Awareness
In each of Experiments 2A and 2B, all but 2 participants reported no awareness of the prime words. In Experiment 2A, 1 participant reported being aware of the identity of the prime word on approximately 30% of the experimental trials and noted that, occasionally, the prime word was the same as the probe target word. A second participant reported noticing one experi- mental trial in which the prime may have contained a word. In Experiment 2B, one participant reported having been aware that the prime contained a briefly presented word, was able to iden- tify this word on only two or three trials, and did suspect that the probe target word was sometimes the same as the prime word. A second participant reported being aware that the prime contained a briefly presented word but reported not paying atten- tion to its identity. In Experiment 2C, no participants reported that they were aware that words were presented in the prime display, but 2 participants reported that they saw letter features on a small number of trials.
Table 2 Mean Response Times (RTs; in Milliseconds) and Error Rates (ERs) for the Unrepeated and Repeated Conditions in Experiments 2A, 2B, and 2C and Experiment 3
Exp. 2A Exp. 2B Exp. 2C Exp. 3
Condition RT ER RT ER RT ER RT ER
Unrepeated 604 .028 574 .036 810 .048 477 .014 Repeated 612 .024 580 .039 819 .048 472 .017
Difference -8 .004 -6 -.003 -9 .000 5 -.003
Note. In Experiments 2A, 2B, and 2C, the probe task required selection whereas in Experiment 3, the probe task did not require selection. Exp. = experiment.
were longer in the repeated condition (664 ms) than in the unrepeated condition (657 ms). Analysis of the individual ex- periments revealed significant main effects of priming condition in all three experiments, F( 1, 19) = 10.83, MSE = 106.36, p < .004, F(1, 23) = 5.17, MSE = 184.83, p < .04, and F(1, 19) = 9.13, MSE = 175.08, p < .008, for Experiments 2A, 2B, and 2C, respectively (see Table 2).
In the analysis of error rates, there were significant main effects of experiment, F(2, 61 ) = 4.98, MSE = 0.002, p < .05, and block, F( 1, 61 ) = 4.36, MSE = 0.0007, p < .05. More errors were made in the first half than in the second half of the experimental session, and post hoc tests indicated that more errors were made in Experiment 2C than in Experiment 2A (p < .05). Across Experiments 2A, 2B, and 2C, the mean error rates for the unrepeated (.037) and repeated conditions (.037) were identical.
Naming Task Performance
Correct RTs for each participant within each of the three experiments were submitted to the same outlier analysis as used in Experiments 1A and lB. This procedure resulted in the elimi- nation of 2.4% of the RTs in Experiment 2A, 2.5% of the RTs in Experiment 2B, and 1.4% of the RTs in Experiment 2C. Mean correct RTs and error rates for each cell within each experiment were then computed and submitted to mixed ANOVAs that treated experiment as a between-subjects variable and priming condition and block as within-subject variables. Mean RTs and error rates for both priming conditions, collapsed across subjects within experiments, are displayed in Table 2.
In the analysis of RTs, there were significant main effects of experiment, F(2, 61) = 68.68, MSE = 20,509.12, p < .001, and block, F(1, 61 ) = 101.40, MSE = 1,334.46, p < .001, as well as a significant interaction between experiment and block, F(2, 61 ) = 7.84, MSE = 1,334.46, p < .01. RTs were longer in the first half than in the second half of the experimental session, and this practice effect was larger in Experiment 2C than in Experiments 2A and 2B. In addition, post hoc qhkey tests revealed that RTs were longer in Experiment 2C than in Experiments 2A and 2B ( p < .05).
More important, there was a significant main effect of priming condition, F( 1, 61 ) = 23.41, MSE = 157.38, p < .001. RTs
Discussion
The results from these brief prime experiments demonstrate unambiguously that negative priming can occur without partici- pants having to 'select against' a distractor in the prime display. Across two experiments that varied only in the proportion of repeated items, and in a third experiment in which a different procedure for designating target items was used, response was slower to repeated than to unrepeated probe targets. As such, these results demonstrate that negative priming can be observed when a prime is presented but not attended rather than presented and overtly selected against in favor of a target.
The theoretical account offered here explains these data by reference to attentional processing during the retrieval of a re- sponse to the probe. Because the probe selection task requires selection between two interleaved probe words, participants are assumed not to rely indiscriminately on automatic retrieval pro- cesses to respond to the probe. Presumably, such a strategy would result in a substantial number of trials in which the dis- tractor rather than the target would be named. Instead, partici- pants adopt an attentional set that discriminates between stimuli for which a response can be retrieved directly from memory and stimuli for which a response relies on analytic perceptual processing. By this account, responses in the repeated condition of the present experiments are slow because they benefit neither

216 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
from the direct retrieval of a response from memory nor from the quick engagement in perceptual analysis that accompanies an orienting response. In effect, the slow responses on repeated trials occur because repeated probes are less temporally discrim- inable than unrepeated probes from the preceding prime. 4
Expe r imen t 3: Br ie f Pr imes Wi thou t Probe Selec t ion
The results of the brief prime experiments described pre- viously demonstrate that negative priming can occur in an exper- iment that precludes attribution of the effect to the selection against a distracting prime. More important, we have suggested that these negative priming effects are determined by the same process as that observed in experiments that do require selection against a prime distractor. The present experiment was con- ducted to provide converging evidence for this claim.
If the negative priming effects observed with prime selection, on the one hand, and without prime selection, on the other hand, share a common cause, then manipulation of a variable that affects negative priming observed with prime selection should also affect the negative priming observed without prime selec- tion. To test this hypothesis, we conducted an experiment that was similar to the brief prime experiments reported earlier, but in which there was no distractor in the probe display. Our predic- tion was that negative priming would not occur under these conditions.
Me~od
Participants
Twenty (17 women) undergraduate students from a second-year psy- chology course at McMaster University participated in the experiment for course credit. Mean age of the participants was 21 years. All partici- pants had normal color vision and normal or corrected-to-normal visual acuity.
Procedure
This experiment was identical to Experiment 2A, with the exception that the distracting probe word was displayed in the same color as the background of the computer monitor. As such, participants were required to name only a single red probe word on each trial. As in Experiment 2A, there were equal numbers of unrepeated and repeated trials in the experimental session.
Results
Participants' Reports of Prime Awareness
One participant reported being aware of the identity of the prime word on approximately 90% of the trials. Further, this participant reported using the prime word to predict the occur- rence of that same word in the probe display. 5 A second partici- pant reported seeing a word during the prime display on a small number of trials but having ignored the prime display for the most part and therefore not noticing a relation between the prime and probe displays. The remaining participants reported not being aware of the prime words.
Naming Task Performance
Correct RTs for each participant in each cell defined by the factorial combination of the priming condition and block vari- ables were submitted to the same outlier analysis as used in earlier experiments. This procedure resulted in the elimination of 1.7% of the RTs. Mean correct RTs and error rates for each cell were then computed for each participant and submitted to repeated measure ANOVAs that treated priming condition and block as within-subject variables. Mean Rig and error rates for the two priming conditions, collapsed across subjects, are dis- played in Table 2.
In the analysis of RTs, the main effect of priming condition was significant, F(1 , 19) = 5.74, MSE = 66.07, p < .03. Correct responses were significantly faster in the repeated condi- tion (472 ms) than in the unrepeated condition (477 ms). The main effect of block was marginally significant, F( 1, 19) = 3.96, MSE = 749.64, p < .07, with faster responses in the second half (468 ms) than in the first half (480 ms) of the experimental session. Priming condition did not interact signifi- cantly with block ( F < 1 ). There were no significant effects in the analysis of error rates ( F < 1 ).
Discussion
This experiment was identical in procedure to Experiment 2A with the exception that participants named a lone red word in the probe display rather than naming a red word and ignoring a green word. When participants were asked to name one of two probe words in Experiment 2A, significant negative priming was observed. In the present experiment, significant positive priming was observed. The temporal discrimination account forwarded here explains the results from this experiment by reference to the attentional set adopted by participants during response to the probe task. In the present task, automatic re-
4 To those familiar with the negative priming literature, the results of Experiments 2A-2C may seem suspect in light of data reported pre- viously by Allport et al. ( 1985 ). Using stimuli that consisted of overlap- ping line drawings, Allport et al. found that if a prime display containing a target and distractor item was presented briefly and pattern masked, then priming effects from both the attended and ignored primes were positive rather than negative in direction. Yet, we have argued that nega- tive rather than positive priming occurs when a similar procedure is used with words as stimuli. There are a number of methodological differences that could account for the discrepancy between the present data and those reported by Allport et al. The most obvious difference has to do with the stimuli themselves. It is possible that differences in the relative speeds with which visual and semantic information is processed for words and pictures underlie the different results. Nevertheless, the point we are making is that the positive priming for pattern-masked primes reported by Allport et al. is consistent with the notion that prime selec- tion determines negative priming but does not necessitate such a conclu- sion. On the other hand, negative priming following a single briefly presented and pattern-masked word does require an interpretation in which selection between two simultaneously presented prime words plays no role.
5 All analyses reported for this experiment were also conducted after removing the data from this clearly atypical participant. However, the results from these analyses were identical whether or not this partici- pant's data were included.

NEGATIVE PRIMING 217
trieval processes are much less likely than in experiments that require probe selection to result in errors in response. As such, the relative performance in repeated and unrepeated conditions does not take on the properties of the temporal discrimination process described here and instead maps monotonically onto the quality of match between prime and probe.
Expe r imen t 4: Read ing Versus Ignor ing
The purpose of the brief prime experiments was to demon- strate that negative priming can occur when attentive processing of a single prime is limited by duration of presentation. In contrast, in the present experiment, attentive processing of a single prime was manipulated by instruction. The prime words were displayed for 200 ms, rather than for 33 ms, and partici- pants were asked either to attend or to ignore the prime.
This experiment was motivated by the results of two pilot studies. In one of these studies, participants responded by nam- ing one of two probes following a single prime. However, in contrast to the brief prime experiments, the prime was presented for 200 ms. The longer exposure duration of the prime allowed participants to attend to its identity. However, following partici- pation in the study, 9 of 10 participants reported having ignored the identity of the prime voluntarily. Eight of these 9 participants responded more slowly on repeated than on unrepeated trials. In contrast, the 1 participant who reported having read most of the primes responded more quickly on repeated trials. In the second pilot study, we confirmed that the nature of the probe task was critical to this negative priming effect. Ten participants completed an experiment that was identical to the previous one, with the exception that the green distracting word was removed from the probe display. Note that the procedure then consisted of a single prime followed by a single probe, with participants instructed to name the probe as quickly and as accurately as possible. Seven of 10 participants reported predominantly ignor- ing the identity of the prime, yet the group produced a reliable positive priming effect. This positive priming effect demon- strates that there is nothing inherent in the present procedure that makes it difficult to measure repetition priming, as conven- tionally defined.
The results from these pilot studies again highlight the fact that the direction of a repetition effect can depend critically on the nature of the probe task. Most participants in both studies reported having ignored rather than attended to the identity of the prime, yet the groups produced repetition effects that were opposite in sign. These data suggest that attentional processing of a single prime has consequences that are analogous to those observed in the attended repetition condition of studies in which the conventional prime selection procedure is used. More im- portant, ignoring a single prime may have consequences that are analogous to those observed in the ignored repetition condi- tion of studies in which a prime selection procedure is used. In other words, the target-distractor prime status of the repeated probe appears not to dictate the direction of priming effects. Rather, as noted earlier in the article, encoding with versus with- out attention may result in qualitatively different priming effects.
To follow up the results of these pilot studies, we conducted the following experiment in which participants responded to one of two probes that followed a single prime. In contrast to
the pilot studies, we attempted to manipulate the manner in which participants processed the prime rather than allowing them to decide on their own. Half of the participants in this experiment were asked to read the prime to themselves, and the other half were asked to ignore the prime. Negative priming was predicted for the group instructed to ignore the prime, and positive priming was predicted for the group instructed to read the prime.
Method
Participants
Twenty (14 women) undergraduate students from a second-year psy- chology course at McMaster University participated in the experiment for course credit. Half of the participants were assigned to the read group, and the other half to the ignore group. Mean age of the participants was 22 years. All participants had normal color vision and normal or corrected-to-normal visual acuity.
Procedure
The apparatus for the experiment was identical to that used in all previously described experiments. The stimuli used were identical to those in Experiments 2A and 2B, with the exception that the exposure duration of the prime word was lengthened from 33 to 200 ms. Details of the procedure for this experiment that differed from previous experi- ments are as follows.
Participants were assigned to one of two groups. Participants in the read group were told that the first thing to appear on each trial would be a pattern mask. Then a single white word would appear and partici- pants were not to let this word pass by without reading it to themselves. After the presentation of this word, a mask appeared, then disappeared, and was followed by the probe display. Participants were asked to name the red word in the probe display as quickly and as accurately as possible. The instruction to read the prime word was then repeated to participants before having them move on to the practice and experimental sessions.
Participants in the ignore group were told that the first thing to appear on each trial would be a pattern mask. Then a single white word would appear and disappear, followed by another pattern mask. Participants were told that they could ignore all of these displays because their task was to respond to the display that followed. They were told that in the following display there would be two interleaved words, one red and one green, and they were to name the red one as quickly and as accurately as possible.
After the experimental session, the experimenter addressed partici- pants in both groups as follows: "I realize that you were asked to (read-ignore) the prime word, but I'd like to ask you this anyway." Participants were then asked to estimate the percentage of trials in which they read the prime word and to estimate the percentage of trials in which the prime word was the same as the probe target. The correct answer to the second question was 50%. Participants in the read group were expected to provide more accurate estimates of this value than participants in the ignore group.
Results
Estimates of the percentage of prime words read by partici- pants in the read and ignore groups were submitted to a single- factor between-subjects ANOVA. The effect of group was sig- nificant, F(1 , 18) = 93.31, MSE = 244.51, p < .001. Partici- pants who were instructed to read the prime words estimated that they read more (90%) primes than did participants who
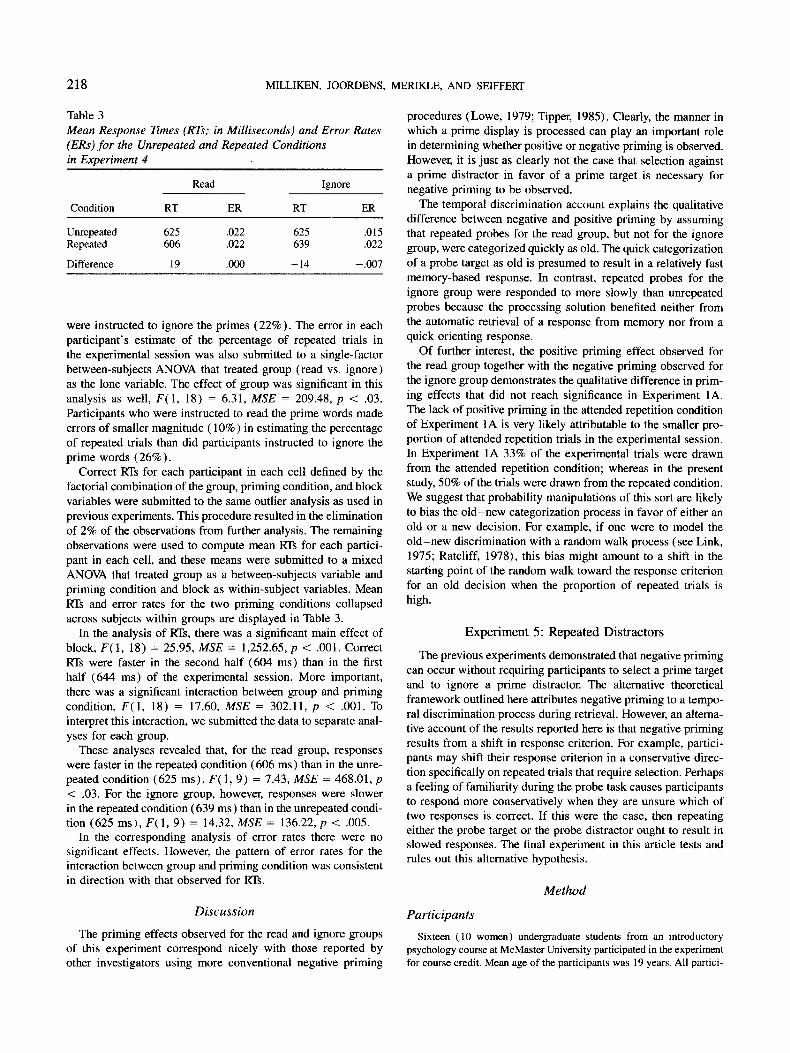
218 MILLIKEN, JOORDENS, MERIKLE, AND SE1FFERT
Table 3 Mean Response Times (RTs; in Milliseconds) and Error Rates (ERs) for the Unrepeated and Repeated Conditions in Experiment 4
Read Ignore
Condition RT ER RT ER
Unrepeated 625 .022 625 .015 Repeated 606 .022 639 .022
Difference 19 .000 - 14 -.007
were instructed to ignore the primes (22%). The error in each participant's estimate of the percentage of repeated trials in the experimental session was also submitted to a single-factor between-subjects ANOVA that treated group (read vs. ignore) as the lone variable. The effect of group was significant in this analysis as well, F(1, 18) = 6.31, MSE = 209.48, p < .03. Participants who were instructed to read the prime words made errors of smaller magnitude (10%) in estimating the percentage of repeated trials than did participants instructed to ignore the prime words (26%).
Correct RTs for each participant in each cell defined by the factorial combination of the group, priming condition, and block variables were submitted to the same outlier analysis as used in previous experiments. This procedure resulted in the elimination of 2% of the observations from further analysis. The remaining observations were used to compute mean RTs for each partici- pant in each cell, and these means were submitted to a mixed ANOVA that treated group as a between-subjects variable and priming condition and block as within-subject variables. Mean RTs and error rates for the two priming conditions collapsed across subjects within groups are displayed in Table 3.
In the analysis of RTs, there was a significant main effect of block, F(1, 18) = 25.95, MSE = 1,252.65, p < .001. Correct RTs were faster in the second half (604 ms) than in the first half (644 ms) of the experimental session. More important, there was a significant interaction between group and priming condition, F(1, 18) = 17.60, MSE = 302.11, p < .001. To interpret this interaction, we submitted the data to separate anal- yses for each group.
These analyses revealed that, for the read group, responses were faster in the repeated condition (606 ms) than in the unre- peated condition (625 ms), F( 1, 9) = 7.43, MSE = 468.01, p < .03. For the ignore group, however, responses were slower in the repeated condition (639 ms) than in the unrepeated condi- tion (625 ms), F(1, 9) = 14.32, MSE = 136.22, p < .005.
In the corresponding analysis of error rates there were no significant effects. However, the pattern of error rates for the interaction between group and priming condition was consistent in direction with that observed for RTs.
Discussion
The priming effects observed for the read and ignore groups of this experiment correspond nicely with those reported by other investigators using more conventional negative priming
procedures (Lowe, 1979; Tipper, 1985). Clearly, the manner in which a prime display is processed can play an important role in determining whether positive or negative priming is observed. However, it is just as clearly not the case that selection against a prime distractor in favor of a prime target is necessary for negative priming to be observed.
The temporal discrimination account explains the qualitative difference between negative and positive priming by assuming that repeated probes for the read group, but not for the ignore group, were categorized quickly as old. The quick categorization of a probe target as old is presumed to result in a relatively fast memory-based response. In contrast, repeated probes for the ignore group were responded to more slowly than unrepeated probes because the processing solution benefited neither from the automatic retrieval of a response from memory nor from a quick orienting response.
Of further interest, the positive priming effect observed for the read group together with the negative priming observed for the ignore group demonstrates the qualitative difference in prim- ing effects that did not reach significance in Experiment 1A. The lack of positive priming in the attended repetition condition of Experiment 1A is very likely attributable to the smaller pro- portion of attended repetition trials in the experimental session. In Experiment 1A 33% of the experimental trials were drawn from the attended repetition condition; whereas in the present study, 50% of the trials were drawn from the repeated condition. We suggest that probability manipulations of this sort are likely to bias the old-new categorization process in favor of either an old or a new decision. For example, if one were to model the old-new discrimination with a random walk process (see Link, 1975; Ratcliff, 1978), this bias might amount to a shift in the starting point of the random walk toward the response criterion for an old decision when the proportion of repeated trials is high.
Experiment 5: Repeated Distractors
The previous experiments demonstrated that negative priming can occur without requiring participants to select a prime target and to ignore a prime distractor. The alternative theoretical framework outlined here attributes negative priming to a tempo- ral discrimination process during retrieval. However, an alterna- tive account of the results reported here is that negative priming results from a shift in response criterion. For example, partici- pants may shift their response criterion in a conservative direc- tion specifically on repeated trials that require selection. Perhaps a feeling of familiarity during the probe task causes participants to respond more conservatively when they are unsure which of two responses is correct. If this were the case, then repeating either the probe target or the probe distractor ought to result in slowed responses. The final experiment in this article tests and rules out this alternative hypothesis.
Method
Participants
Sixteen (10 women) undergraduate students from an introductory psychology course at McMaster University participated in the experiment for course credit. Mean age of the participants was 19 years. All partici-

NEGATIVE PRIMING
Table 4 Mean Response Times (RTs; in Milliseconds) and Error Rates (ERs) for the Unrepeated, Target Repeated, and Distractor Repeated Conditions in Experiment 4
Stimulus onset asynchrony (ms)
1 ~ 2 ~ 4 ~
Condition RT ER RT ER RT ER
Unrepeated 572 .015 559 .021 542 .029 Target repeated 591 .034 571 .046 564 .040 Distractor repeated 553 .003 532 .013 536 .022 Repetition effects
Unrepeated-target repeated - 19 -.019 - 12 -.025 -22 -.011 Unrepeated-distractor repeated 19 .012 27 .008 6 .007
219
pants had normal color vision and normal or corrected-to-normal visual acuity.
Procedure
All aspects of the procedure of this experiment were identical to those in preceding experiments with the following exceptions. Participants responded to the red probe target and ignored the green probe distractor on all experimental trials. A single prime word preceded the probe display. The presentation duration of the prime word was 50 ms, and the prime was not masked. An interval of 50, 150, or 350 ms followed presentation of the prime, creating stimulus onset asynchronies (SOAs) of 100, 200, and 400 ms. The probe presentation then followed and remained on the screen until onset of the participants' naming response. There were three priming conditions tested in this experiment. In the anrepeated condition neither the probe target nor the probe distractor was related to the prime, in the target repeated condition the probe target was the same as the prime, and in the distractor repeated condition the probe distractor was the same as the prime. Trials were run in subblocks of 18, within which each combination of the priming condition and SOA variables was represented twice. The experimental session consisted of two blocks of 180 trials, preceded by a practice session of 18 trials.
Results
Correct RTs for each participant in each cell defined by the factorial combination of the priming condition, SOA, and block variables were submitted to the same outlier analysis as used in previous experiments. This procedure resulted in the elimination of 2% of the observations from further analysis. The remaining observations were used to compute mean RTs for each partici- pant in each cell, and these means were submitted to a repeated measures ANOVA that treated priming condition, SOA, and block as within-subject variables. Mean RTs and error rates, collapsed across subjects, are displayed in Table 4.
In the analysis of RTs, there were significant main effects of block and SOA, F(1 ,15) = 28.95, MSE = 3,035.04, p < .001 and F (2 , 30) = 20.15, MSE = 760.61, p < .001, respectively. RTs decreased with increasing SOA (see Table 4) and were faster in the second block than in the first block of the experi- mental session. More important, there was a significant main effect of priming condition, F (2 , 30) = 34.58, MSE = 847.36, p < .001. Subsequent planned comparisons revealed that re- sponses were slower in the target repeated condition (575 ms) and faster in the distractor repeated condition (541 ms) than in
the unrepeated condition (558 ms) , F ( 1, 15) = 18.90, MSE = 810.85, p < .001 and F (1 , 15) = 18.87, MSE = 741.47, p < .001, respectively. There was also a significant interaction be- tween priming condition and SOA, F(4 , 60) = 3.33, MSE = 262.33, p < .05. This interaction appears to be due to relatively small variations in the pattern of priming effects described ear- lier across the three SOAs (see Table 4).
In the analysis of error rates, the only significant effect was the main effect of priming condition, F (2 , 30) = 10.28, MSE = 17.76, p < .001. Planned comparisons revealed that more errors were made in the target repeated condition (.040) than in the unrepeated condition ( .022), F (1 , 15) = 9.63, MSE = 16.47, p < .01. Marginally fewer errors were made in the dis- tractor repeated condition (.013) than in the unrepeated condi- tion (p < .10). These error rate effects were consistent in direc- tion with those observed in the response time analysis.
Discussion
The present experiment was similar to others described in this article. Participants were asked to respond by naming one of two interleaved words in a probe display following presenta- tion of a single prime. However, in addition to the unrepeated and repeated conditions, a distractor repetition condition was added to this experiment. In this condition, the single prime was the same as the probe distractor. Importantly, responses to the probe target were slower in the target repeated condition than in the unrepeated condition but were faster in the distractor repeated condition than in the unrepeated condition. Note that if participants shifted their response criterion in a conservative direction any time a repeated stimulus appeared, then responses in the distractor repetition condition should also have been slower, rather than faster, than those in the unrepeated condition. This result, as well as many others like it (e.g., Neumann & DeSchepper, 1991; Tipper, Bourque, Anderson, & Brehaut, 1989), suggests that repetition of a stimulus can reduce its com- petitiveness for access to a limited-capacity response system. According to the temporal discrimination framework, this re- duced competitiveness occurs because repeated probes are less discriminable than unrepeated probes from preceding primes. In short, if participants are attentionally set to capitalize on the distinctiveness of an unrepeated probe, then repeated stimuli may be responded to more slowly and ignored more efficiently.

220 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
General Discuss ion
Over the past decade negative priming has gained wide accep- tance as a measure of inhibitory attentional processing. At the same time, although negative priming is presumed to measure inhibition that occurs during selection of a prime target, the effect often depends on properties of the following probe task. This result suggested to researchers some time ago (Lowe, 1979; Tipper & Cranston, 1985) that the simplest, and perhaps most intuitively appealing, of inhibitory accounts does not provide a satisfactory explanation of negative priming. The task of provid- ing an alternative account that better explains the cause of nega- tive priming has proved a difficult one (see Neill & Valdes, 1996). The experiments reported here suggest that one of the sources of this difficulty stems from a misunderstanding con- cerning the procedure required to measure negative priming. The principal finding reported in this article is that negative priming can be observed following presentation of a single prime. This result contradicts the widely held view that negative priming is caused by the act of selecting against a prime dis- tractor in favor of a prime target.
The dependence of the negative priming effects reported here on properties of the probe task is a finding of particular impor- tance. As mentioned previously in this article, these results par- allel numerous findings that have been observed in which the more conventional prime selection procedure for measuring negative priming was used (e.g., Lowe, 1979; Moore, 1994; Tipper, 1985; Tipper & Cranston, 1985). This parallel argues against the view that we have created a distinct negative repeti- tion effect that deserves its own idiosyncratic explanation. Fur- ther, the dependence of negative priming on probe selection suggests that the attribution of attended and ignored repetition effects to the persisting effects of complementary excitatory and inhibitory attentional processes is incorrect.
As an alternative to extant accounts, we forwarded a temporal discrimination framework for explaining negative priming. One of the notable properties of the temporal discrimination frame- work is that, although two processes are implicated, qualitatively different priming effects for attended and ignored repetition conditions can be explained by reference to the modulation of a single process. In particular, attention to a prime is presumed to increase the contribution of automatic retrieval to perfor- mance such that this contribution may be greatest on attended repetition trials, least on unrepeated trials, and intermediate on ignored repetition trials. Consequently, probes may be catego- rized quickly as old in the attended repetition condition and as new in the unrepeated condition. Negative priming occurs on ignored repetition trials because the similarity of the ignored prime to the subsequent probe target impedes its categorization as new and yet is insufficient to support its categorization as old. In effect, slowed responses result from ambiguity as to whether a probe should be responded to by reinstating prior learning or by engaging in new learning.
Is a New Theoretical Framework Necessary?
The temporal discrimination framework provides a novel ex- planation of negative priming. However, if an existing theoretical account can accommodate the present data in a straightforward
manner, then it may be argued that a novel interpretation is unnecessary. In this section we consider whether the present data can be accommodated by two of the predominant extant accounts of negative priming.
The Distractor Inhibition Hypothesis
As mentioned in the introduction of this article, the simplest of distractor inhibition accounts was rendered untenable long ago (Lowe, 1979; Tipper & Cranston, 1985). However, variants of the distractor inhibition hypothesis have evolved that are able to account for various complexities of the empirical literature on negative priming. Might the distractor inhibition hypothesis also accommodate negative priming following presentation of a single prime? Rather than selecting against a distractor in the prime display, it may be argued that participants in the present studies selected against the prime display as a whole. By this logic, because the lone prime word was irrelevant to the goal of naming a red probe word, it may have been actively ignored in much the same way as is a prime distractor in conventional negative priming tasks. This explanation implies that selective attention to one of two sources of information that are presented one after another may have similar consequences to selective attention to one of two sources of information that are presented simultaneously. The implications of this argument for two vari- ants of the distractor inhibition hypothesis are described below.
Response blocking. Recall that Tipper and Cranston's (1985) response blocking account attributes negative priming to a response block that is engaged to aid selection of a prime target. This response block is presumed to be maintained for probe tasks that require selection but that can be abandoned for probe tasks that do not require selection. Although this response blocking account explains negative priming reasonably well when the prime task requires selection, note the implication of this model for explaining the single prime results reported here. Ignoring a single prime would have to engage the response block. The response block would then remain activated for probe tasks that require selection but would be abandoned for probe tasks that do not require selection. Consequently, this account provides an unparsimonious explanation for positive priming when a single probe follows a single ignored prime; that is, a response block must be engaged and later abandoned to reveal positive priming. A more parsimonious account explains positive priming effects without reference to blocking processes and instead explains negative priming by reference to processes en- gaged in response to a probe task that requires selection.
The Houghton and Tipper (1994) model. An alternative in- hibition account of negative priming was offered by Houghton and Tipper (1994). This account derives from a connectionist model in which target and distractor representations are differen- tiated from one another by both excitatory and inhibitory pro- cesses. Importantly, the representation of the distractor is not immediately inhibited to a subbaseline state of activation. In- stead, inhibition acts as a counterforce against the bottom-up excitatory effect of the physically present distractor. Thus, while the stimulus is in view, the bottom-up excitation provided by its presence reaches a state of equilibrium with respect to the inhibitory counterforce. The inhibitory counterforce allows the target to become more actively represented than the distractor

NEGATIVE PRIMING 221
but does not preclude an active representation of the distractor while the selection of an action is taking place. This aspect of the Houghton and Tipper model provides a solution to a nagging drawback of simpler variants of the distractor inhibition hypoth- esis. In particular, objects must often remain actively represented in spite of their role as distractors. For example, imagine reach- ing around one object (the distractor) to grab another (the target; see Tipper et al., 1992). Somehow, incorrect reaching move- ments toward the distractor mast be prevented. At the same time, the distractor must be taken into account in the correct reaching behavior to the target. Without an active representation of the distractor, the computed reach toward the target could conceiv- ably send the hand through the space occupied by the distractor.
According to the Houghton and Tipper (1994) model, then, selection of a prime target for action occurs on the basis of a difference in activation for targets and distractors, without ini- tially producing a subbaseline state of activation for the dis- tractors. However, on offset of the prime stimalus, the inhibition directed toward the distractor loses its excitatory counterforce, and an inhibitory rebound occurs. During this inhibitory re- bound the activation level of the distractor falls below its resting level, thus explaining how negative priming occurs.
The Houghton and Tipper (1994) model does provide an explanation for the dependence of negative priming on probe selection. The model assumes that ignoring a prime distractor results in a subsequent competitive disadvantage for an identical or related probe target. However, this competitive disadvantage may only realize itself (as negative priming) when the probe task involves selection between two highly competitive re- sponses, such as occurs for simultaneously present targets and distractors. In other words, the criterion difference in activation required for selection to occur may be drawn out over time when the probe task requires selection but occurs immediately when the probe task does not require selection. When this crite- rion difference in activation is achieved immediately on onset of the probe, the competitive disadvantage afforded by distractor inhibition (negative priming) may no longer be measurable.
Generally speaking, then, the Houghton and Tipper (1994) model accounts for the dependence of negative priming on probe selection by reference to response competition between the probe target and distractor. However, as noted earlier in this article, there is considerable evidence that a strategic response to impending response competition, rather than response compe- tition itself, is responsible for the dependence of negative prim- ing on probe selection (Lowe, 1979; Moore, 1994). The flexi- bility afforded by strategic processes seems necessary to ac- count for negative priming following an ignored prime under some conditions and positive priming following ignored primes under other conditions. As currently instantiated, the Houghton and Tipper model does not provide a means of explaining posi- tive priming for a previously inhibited prime. 6
The Episodic Retrieval Hypothesis
According to Neill et al.'s (1992) episodic retrieval hypothe- sis, negative priming is caused by the retrieval of prior pro- cessing episodes in which response information is encoded. Thus, responses on ignored repetition trials may be slowed by an ignore-this-stimulus tag associated with the retrieved repre-
sentation of the prime distractor. As argued for the distractor inhibition hypothesis, it is conceivable that participants attach an ignore-this-stimulus tag to a single prime in much the same way they might for a prime distractor. If this response tag is retrieved during the probe task, then negative priming ought to occur. To evaluate this account in light of the present data, it is instructive to consider the two ways in which the episodic re- trieval model explains the dependence of negative priming on probe task selection.
One way in which the episodic retrieval model accounts for the dependence of negative priming on probe selection is by reference to the contextual similarity of a two-item probe dis- play to the conventional two-item prime display. This particular account does not fare well in light of the present data. We observed negative priming for two-item probe displays that fol- lowed one-item prime displays and positive priming for one- item probe displays that followed one-item prime displays. These results are not consistent with a contextual similarity account in which similarity is defined by the relative numbers of prime and probe items. 7
A second way in which the episodic retrieval model accounts for the dependence of negative priming on probe selection refers directly to the relative speeds of the automatic retrieval and controlled algorithmic processing routes. Recall that perfor- mance is presumed to depend on the faster of these two pro- cessing routes (Logan, 1988). As a result, any manipulation that slows the algorithmic process ought to result in a larger contribution to performance by automatic retrieval processes. If negative priming results from the retrieval of conflicting re- sponse information, and if the presence of a probe distractor slows algorithmic processing, then negative priming might well depend on the presence of a probe distractor.
One implication of this account is that the relative contribu- tion of controlled processes to performance must be greater for probe tasks that do not require selection than for probe tasks that do require selection. We know of no direct evidence to contradict this assumption. At the same time, this assumption is precisely the opposite to that adopted in the temporal discrimi- nation account. Our assumption is that negative priming occurs when automatic retrieval processes contribute to a processing solution, but when they are insufficient to provide a basis for response. However, the insufficiency of automatic retrieval as a basis for response stems from a controlled strategic response to a probe task that requires selection. Presumably, a probe task
6 Although positive priming for ignored-repeated items presents a unique challenge for the Houghton and Tipper (1994) model, incorpora- tion of the notion that only behavioral goal-related properties of dis- tractors are inhibited may provide a solution. In this manner, representa- tions associated with other properties of the distractor may support faster responses to ignored-repeated items under appropriate conditions (see Milliken et al., 1994). As noted, this version of the model has yet to be instantiated.
7A contextual similarity account in which similarity is defined by whether an item is ignored in both prime and probe displays would fare better (W. T. Neill, personal communication, November 1, 1996). For example, when participants ignore single primes, the presence of a dis- tracting probe (also an ignored item) might reinstate the processing context of the prime display. On repeated trials, the process of ignoring the prime would be retrieved, which might cause negative priming.

222 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
that does not require selection can be responded to efficiently without this strategy, by reliance on automatic retrieval processes.
Contrasting the episodic retrieval and temporal discrimina- tion hypotheses. Given the well-defined distinction between the episodic retrieval and temporal discrimination accounts noted previously, it is worth considering whether extant data lean toward one theory or the other. Again, the episodic retrieval theory predicts that increases in the contribution of automatic retrieval to performance ought to increase negative priming. It was pointed out earlier that contextual similarity, defined by the number of prime and probe items presented, cannot explain the dependence of negative priming on probe selection. However, recent data reported by Neill (1997) demonstrate that negative priming can depend on other contextual similarities between the prime and probe tasks. In Neill 's (1997) study, the temporal onset of target and distracting flanker stimuli (Eriksen & Erik- sen, 1974) was varied such that onsets for the target and dis- tractors were sometimes synchronous and other times asynchro- nous. Importantly, negative priming was observed when the on- sets of targets and distractors were either synchronous in both prime and probe tasks or asynchronous in both prime and probe tasks. These data do suggest that retrieval processes play an important role in determining negative priming and thus are quite consistent with the episodic retrieval account.
The temporal discrimination account can explain this finding by assuming that contextual dissimilarity between prime and probe makes both repeated and unrepeated probes easily dis- criminable from preceding primes. In other words, contextually dissimilar probes may be categorized quickly as new regardless of whether they are repeated or unrepeated. In contrast, repeated probes may be discriminated slowly from preceding primes only when they are contextually similar to those primes. Thus, both the episodic retrieval account and the temporal discrimination account can explain increases in negative priming for contextu- ally similar primes and probes.
The temporal discrimination account makes the further pre- diction that, under some circumstances, negative priming ought to be larger for contextually dissimilar primes and probes. This claim follows from the notion that contextual dissimilarity be- tween prime and probe favors categorization of the probe as new, whereas contextual similarity favors its categorization as old. Thus, a probe stimulus that matches one stimulus dimension of the prime, but mismatches on a second dimension of the prime, may be difficult to categorize as old or new.
Indeed, this result has been demonstrated in the spatial variant of the negative priming procedure. Similar to the results reported here, Park and Kanwisher (1994) demonstrated negative prim- ing in a spatial localization task without requiring participants to select overtly between a prime target and a distractor. How- ever, this result was observed only when the probe target symbol mismatched with the symbol that appeared at the same location in the prime display. Park and Kanwisher subsequently attributed negative priming in spatial localization tasks to mismatching features associated with a single location (see Kahneman et al., 1992) rather than to distractor inhibition.
In contrast, the temporal discrimination account assumes that feature mismatching effects play a supporting role in a larger story. Thus, rather than attributing negative priming to feature
mismatches (for evidence against this hypothesis, see Milliken et al., 1994; Tipper, Weaver, & Milliken, 1995), we suggest that negative priming results from the processing consequences of feature mismatches in the categorization of probes as old or new. From this standpoint, the contribution of mismatching to negative priming fits nicely within the temporal discrimination framework. However, because contextual dissimilarity should decrease the contribution of automatic retrieval processes to performance, the episodic retrieval account must, at the very least, invoke some other principle to account for such effects.
Absence of response information? To this point, our discus- sion of the episodic retrieval account (Neill et al., 1992) has focused on the view that negative priming results from the re- trieval of conflicting response information, such as an ignore- this-stimulus tag. However, Neill et al. did identify an alternative version of their account, by which negative priming might result from the absence of encoded response information for the prime distractor. Presumably, the absence of response information for the prime distractor could result in reliance on the slower con- trolled processing route in the ignored repetition condition.
Note that the absence of response information version pro- vides a very different explanation for negative priming than does the response tagging version of the episodic retrieval the- ory. Further, this version of the episodic retrieval account re- quires an explanation as to why responses in an unrepeated condition are faster than those in an ignored repetition condition. The answer to this question must be that responses in an unre- peated condition sometimes rely on the faster automatic retrieval route or that the controlled processing route wins the race with automatic retrieval processes more quickly in the unrepeated condition than in the ignored repetition condition. The latter of these solutions is consistent with the temporal discrimination account forwarded here. However, neither of the solutions pre- sented here follows directly from Logan's (1988) instance the- ory of automaticity. As such, the absence of response informa- tion version of the episodic retrieval account requires specifica- tion beyond that provided to date. This critique is not meant to discourage such specification. Given the data presented in the brief prime experiments of this article, absence of response information, rather than conflicting response information, seems the more viable version of Neill et al.'s (1992) episodic retrieval account.
Temporal Discrimination and Negative Priming
Earlier in this article we noted that both an attentional set (see Tipper & Cranston, 1985) and retrieval processes (see Neill et al., 1992) are necessary components of an explanation of negative priming ( see Figure 6). The temporal discrimination framework outlined here incorporates both of these components and, as such, is a favorable acknowledgment of prior theoretical work. At the same time, the temporal discrimination framework provides a method of explaining negative priming that focuses directly on the process of retrieval rather than on processes involved in selection against an irrelevant prime. As such, we suggest that negative priming may be explained without refer- ence to inhibition of the representation of a prime distractor or to inhibitory response tags that are encoded together with the representation of the prime distractor.

NEGATIVE PRIMING 223
To this point, our discussion of the temporal discrimination framework has primarily concerned the dependence of negative priming on attention during the prime and probe tasks. This section is devoted to a more comprehensive discussion of the temporal discrimination framework and empirical properties of negative priming.
Attended Versus Ignored Repetition Effects
The empirical result most responsible for the wide interest in negative priming is not the negative priming effect itself but rather the contrast between negative priming on ignored repetition trials and positive priming on attended repetition trials (see Lowe, 1979; Neill, 1997; Tipper, 1985). This contrast maps directly onto the theory that selection involves both excitation of target representations and inhibition of distractor representa- tions. If this contrast were always observed, then it would pro- vide compelling evidence in favor of the distractor inhibition hypothesis.
Of course, one of the principal themes of the present article is that negative priming is not always observed for previously ignored primes. Just as importantly, and perhaps more surpris- ingly, positive priming is not always observed for previously attended primes. Although positive priming appears robust for previously attended line drawings (Allport et al., 1985; Sulli- van & Faust, 1993; Tipper, 1985), it was less robust in the experiments reported in this article. More important, negative attended repetition effects have been reported in studies of both word identification (Dark & Schmidt, 1996; Sullivan, Faust, & Balota, 1995) and spatial localization (e.g., Shapiro & Loughlin, 1993).
These data make a clear and important point: The sign of priming effects is not directly related to whether a prime was attended or ignored. A safer assumption is that attention during encoding has an indirect effect on the sign of priming effects. In particular, attention during encoding may facilitate retrieval of the prime when responding to an identical or related probe. In many, but not all, conditions the advantage during retrieval afforded by attention during encoding results in an attended repetition effect that is positive in sign. Similarly, the relatively poor retrieval that owes to ignoring, rather than to attending, a prime often results in an ignored repetition effect that is negative in sign. Importantly, it is assumed that the ignored repetition effect is not related directly to the process of ignoring but in- stead occurs because unrepeated probes are more easily discrim- inable than ignored repetition probes from the primes that pre- cede them.
Persistence of Negative Priming
One of the more profound reports of negative priming to date is that of DeSchepper and Treisman ( 1991, 1996; Treisman & DeSchepper, 1996). Their participants were required to discrim- inate whether one of two overlapping nonsense shapes was the same as or different from a standard shape that was presented beside the comparison pair. They found that participants made same-different discriminations more slowly when the target of comparison on one trial was the same as one that had been ignored during a previous trial. Furthermore, they reported that
negative priming effects of this sort can survive a lag between the prime and probe display of as long as 30 days. Treisman and DeSchepper attributed the negative priming effects to the retrieval of action tags that are stored together with the represen- tation of the shapes. This account is similar to the Neill et al. (1992) view that negative priming owes to the retrieval of conflicting response information. If differential availability of conflicting response information for unrepeated and ignored repetition conditions determines the negative priming effect, then one must assume that this difference in availability is re- tained over lags of up to a month.
However, examination of the Treisman and DeSchepper data (1996, Figure 2.5) suggests that negative priming may outlast participants' memory for response information from the prime episode. This conclusion is drawn on the basis of a comparison of attended and ignored repetition effects for the group of parti- cipants that produced a negative priming effect following a lag of 1 month between prime and probe presentations. Importantly, although not statistically significant, the attended repetition ef- fect was nearly as large and negative in direction as the ignored repetition effect following the I-month lag. More generally, as the lag between prime and probe presentations increased from 1 day to 1 week to 1 month, the ignored repetition and attended repetition effects differed by increasingly less, until at the 1- month lag the difference appears negligible. If it is assumed that the difference between these two effects indicates the degree to which response information was retained, then one mast con- clude that little response information is available after a lag of 1 month between the prime and probe tasks. Yet, these two priming effects appear to converge on a negative value rather than on a value of zero, as the lag between prime and probe presentations increases.
Within the temporal discrimination framework forwarded here, all that mast be retained across a lag of 1 month for negative priming to be observed is memory for the shape pre- sented 1 month earlier. Negative priming would then be attrib- uted to automatic retrieval of a shape encoded 1 month earlier, which interferes with the perceptual analysis performed by the participant 1 month later, to perform the same-different dis- crimination task.
Individual Differences
Another reason that negative priming has gained wide interest is studies that have demonstrated diminished negative priming in clinical and developmental populations. Negative priming has been shown to be less robust in children (Tipper et al., 1989), older adults (Hasher, Stoltzfus, Zacks, & Rypma, 1991; McDowd & Oseas-Kreger, 1991; Tipper, 1991; see also Sulli- van & Faust, 1993), schizophrenic patients (Beech, Powell, McWilliams, & Claridge, 1989), participants who report high cognitive failure (Tipper & Baylis, 1987), depressed patients (Benoit et al., 1992), and Alzheimer's patients (Sullivan et al., 1995). For the most part, it has been argued that the reduced negative priming in these populations reflects a deficit in inhibi- tory attentional processing. Of course, this conclusion is based on the distractor inhibition hypothesis.
One implication of this approach is that deficits in a wide array of clinical populations are attributed to a single cause.

224 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
This approach encourages comparison of the cognitive deficits of, say, schizophrenic patients and older adults. Through this comparison an appreciation of similarities between the deficits of populations that differ markedly in other behavioral respects may be gained. For example, if negative priming is attributed to the persisting effect of an inhibitory selection mechanism, then all of the populations cited previously can be regarded as suffering from deficits in this selection mechanism or in an inhibitory resource on which this selection mechanism depends.
An appreciation of the commonality in cognitive deficits that characterize various populations is important. However, the ob- servation of similarities among populations makes even more intriguing the differences amOng those populations. In other words, if negative priming falls to occur in markedly different populations because of a common deficit, then an explanation is required for why such a wide variety of pathologies accompa- nies that deficit.
Although this issue is not resolved here, we point out that this issue can be addressed fruitfully in further research if a theoretical framework is adopted that allows a common behav- ioral finding to emerge for a variety of reasons. We have sug- gested that negative priming is an emergent result of a temporal discrimination process. One of the alms of casting our frame- work in these terms is so that future researchers might examine various parameters of this discrimination process, such as biases favoring either integration or differentiation, as well as the abil- ity to control these biases. Within this framework, the variety of clinical and developmental populations that fail to demonstrate negative priming might at some point be explained by reference to different factors that affect the temporal discrimination process.
For example, a first distinction that might be made is between populations for whom temporal discrimination is not possible, because of a biological deficit, and populations for whom tem- poral discrimination may be ineffective, perhaps because of immaturity or aging. This distinction is worth noting in refer- ence to data reported recently on negative priming in older adults (Kane et al., 1997). Kane et al. reported several experiments in which negative priming was observed for older adults only in conditions designed to encourage episodic retrieval. Thus, deg- radation of the probe target, reduction of the probe exposure duration, and inclusion of attended repetition trials in the experi- mental session all led to negative priming in older adults. Under a host of other conditions, negative priming has been shown not to occur in an older population (but see Kramer, Humphrey, Larish, Logan, & Strayer, 1994; Sullivan & Faust, 1993; Sullivan et al., 1995).
To explain these data, Kane et al. (1997) suggestedthat nega- tive priming can be caused either by distractor inhibition or by the retrieval of conflicting response information (Neill et al., 1992). Further, when negative priming is observed in older adults, Kane et al. concluded that it must be due to episodic retrieval processes rather than to distractor inhibition. Although this conclusion explains why negative priming might be ob- served in older adults without compromising the view that they suffer from deficient inhibitory processing, it invites debate over which negative priming effects are due to one or the other pro- cess. An alternative framework for explaining negative priming
ought to provide a parsimonious account of these data that preempt such debates.
To this end, these data can be accommodated within the pres- ent framework by assuming that, under some conditions, older adults may not engage in the same temporal discrimination strat- egy as younger adults. For example, if older participants are less efficient than younger participants at discriminating old from new, which seems a modest assumption, then the experi- mental conditions required to induce a temporal discrimination strategy might be expected to differ for the younger and the older adults. For example, inclusion of attended repetition trials in the experimental session may encourage the older participants to discriminate between trials in which the target repeats and all other trials. If automatic retrieval processes are relied on to respond only to the attended repetition trials, then the better discriminability of unrepeated than ignored repeated probes from the prime episode should result in negative priming. In general, the different results for younger and older adults re- ported by Kane et al. (1997) may imply that the older adults, but not the younger adults, must be coaxed by the experimental method to discriminate between automatic retrieval and percep- tual analysis as bases for responding.
In the case of clinical populations, such as schizophrenic patients, reduced negative priming may reflect a deficit that cannot be overcome by attentional, strategies. Cohen and Servan- Schreiber (1992) have argued that schizophrenic patients suffer from an inability to use an internal representation of context caused by a reduction of dopaminergic activity in prefrontal cortex. One of the implications of this deficit is that the represen- tation of current context cannot be used to override the effects of prior experience. As such, perseverative error marks the be- havior of the schizophrenic individual. The temporal discrimina- tion framework is consistent with this view. Cast in the terminol- ogy used here, it is assumed that schizophrenic patients are unable to implement a strategy that differentiates behavior that is based on the current context and that which is based on prior experience. In effect, whereas normal participants may bind the processing of the probe target to the current context, thus ruling out perseveration as a source of error in performance, this strat- egy may not be available to the schizophrenic patient.
Perhaps most important, this theoretical approach stresses that negative priming not be interpreted as reflecting the contri- bution to performance of any one process. Instead, we offer the interpretation that negative priming is an emergent result of a discrimination process that, when operating efficiently, allows behavior to be directed as efficiently as possible toward both familiar and novel stimuli. Future work on individual differences would benefit from examination of the dynamic properties of this discrimination process to better understand the relation be- tween findings that have been observed across a variety of clini- cal and developmental populations.
Speed-Accuracy Criteria
Neill and Westberry (1987) demonstrated that whether nega- tive or positive priming occurs in the Stroop task can depend on the speed/accuracy criterion adopted by participants. In the Neill and Westberry (1987) study, participants in one group were given "strict accuracy instructions," whereas participants

NEGATIVE PRIMING 225
in the other group were given "lax accuracy instructions," thereby encouraging error rates of less than 5% and approxi- mately 10% for the two groups, respectively. The group given strict accuracy instructions produced a significant negative priming effect, whereas the group given lax accuracy instruc- tions produced a nonsignificant trend in the opposite direction. The temporal discrimination framework accounts for this result by assuming that participants engage in an attentive discrimina- tion process precisely because an accurate response is required. If a higher error rate were tolerated, then faster but more error- prone automatic processes could be relied on as a basis for response. As noted earlier in this article, indiscriminate reliance on automatic processes can be expected to produce positive rather than negative priming.
Set Size Effects
Malley and Strayer (1995) reported recently that negative priming is most robust when small stimulus sets are used. It is assumed that the use of small stimulus sets increases the contribution to performance of the temporal discrimination pro- cess described here. In effect, when few stimuli are used in an experiment, the participant's task is less one of generating a hypothesis about what the target stimulus may be and is more one of discriminating among a small number of highly available or primed responses. Efficient performance in such a task re- quires not just an accessible response but a response that is accessible for the right reason; namely, because it corresponds to the current target. As a result, a significant proportion of the time required to respond in tasks in which few stimuli, or few responses, are used (e.g., same-different or word-nonword tasks; see DeSchepper & Treisman, 1996; Fuentes & "lhdela, 1992; Yee, 1991) is devoted to discriminating between current perceptual sources of influence and past memorial sources of influence on performance. In other words, the temporal discrimi- nation process forwarded here is presumed to discriminate be- tween perceptual and memorial sources of influence on perfor- mance (see Jacoby, Kelley, & Dywan, 1989; Johnson, Hash- troudi, & Lindsay, 1993).
In general, the attribution of negative priming to a discrimina- tive process is consistent with the Selective Construction and Preservation of Experiences (SCAPE) formulation of memory forwarded recently by Whittlesea (1997). Whittlesea distin- guishes between the production and evaluation activities that make up a constructive retrieval process. Within Whittlesea's SCAPE framework, the temporal discrimination process de- scribed here may be regarded as an evaluation tool that acts on a produced mental event. Negative priming is then subject to stimulus set size effects because the evaluative function of mem- ory takes on an increased importance with small stimulus sets.
Probe Selection and Temporal Discrimination
One of the critical properties of the putative temporal discrim- ination process is that it may be engaged strategically in re- sponse to task demands. Requiring the participant to respond to one of two interleaved words is one such demand. However, it follows that other task demands might also result in the same strategy. From this perspective, it should not be surprising that
negative priming requires probe selection in some procedures but not in others (see Fuentes & Tadela, 1992; Neill et al., 1994; Neill & Westberry, 1987; Yee, 1991). The precise conditions that lend themselves to implementation of this discrimination process are an interesting and open empirical issue.
Temporal Discrimination and Other-Related Phenomena
Although the discussion to this point has focused on negative priming, our primary motivation was to develop a theoretical framework that could extend beyond the procedurally defined boundaries of any one experimental phenomenon. As a result, the appeal of the temporal discrimination framework ought to depend on its potential to explain other-related experimental phenomena as well. This section describes two such phenomena that appear compatible with the temporal discrimination framework.
Inhibition of Return
Posner and Cohen (1984) demonstrated that the time to detect a visual target at a previously cued location can be slower than that to detect a target at a previously uncued location. This effect is commonly referred to as inhibition of return. In a typical study, an attentional cue consists of the abrupt bright- ening of one of two peripherally marked locations on a computer screen, located to the left and right of fixation. After a variable time interval following this cue, a target is displayed either in the cued location or in the uncued location. The participant is usually required to detect the onset of the target (Posner & Cohen, 1984; Rafal, Calabresi, Brennan, & Sciolto, 1989; Tip- per, Driver, & Weaver, 1991) or to indicate its location (Maylor, 1985). When the temporal interval between the onset of the cue and the onset of the target is short, response times are faster for targets appearing at the cued location than for targets appearing at the uncued location. However, for temporal intervals between cue and target of longer than 300 ms, response times are longer for targets at the cued location.
The benefit of cuing observed at short temporal intervals is commonly attributed to the automatic capture of attention by the attentionai cue. Thus, if attention moves to the location of the cue prior to the onset of the target, then detection of the target may benefit from occurring at an attended location. The cost of attentional cuing observed at longer temporal intervals is often attributed to inhibition that lingers at the cued location after attention has been disengaged and reallocated to fixation. It has been argued that the inhibitory tagging of previously attended locations fosters an adaptive environmental sampling strategy. Thus, after having sampled one location while search- ing for food, the adaptive organism should sample new locations in subsequent searches rather than resampling old locations.
Although acknowledging the importance of novelty, theoreti- cal accounts of inhibition of return often explain attentional cuing effects by reference to representation states that precede the onset of the target. Thus, the benefit of attentional cuing at short temporal intervals between cue and target occurs because attention was already at the location when the target appeared. Similarly, inhibition of return is attributed to an inhibitory tag,

226 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
or to an inhibition of the orienting system, that occurs following the disengaging of attention but prior to the onset of the target. In contrast, we suggest that these attentional effects may not be determined prior to the onset of the target and instead are more directly explained by reference to processes that occur following presentation of the target, but prior to the point in time at which the target onset is registered consciously. In other words, al- though an abrupt visual target may give the impression that it can be detected spontaneously on onset, even this simplest of perceptual events can be thought of as the emergent consequence of a constructive retrieval process.
The qualitatively different cuing effects observed at short and long temporal intervals between cue and target can be inter- preted in a similar manner to the qualitatively different repetition effects observed for ignored and attended repetition conditions in the negative priming paradigm. Thus, the benefit for cued targets at short cue-target intervals may be attributed to the fast categorization of the target as having appeared at the same location as the cue. In effect, the cue and target are grouped together by their spatiotemporal proximity. In contrast, the cost for cued targets at long cue-target intervals may be attributed to the fast categorization of uncued targets as new. In other words, fast responses to uncued targets occur because such targets, or the actions that such targets activate (e.g., oculomotor activation; see Rafal et al., 1989), are easily discriminable from- the preceding cue or from the actions that such cues activate.
If the same processes underlie negative priming and inhibition of return, then the spatial localization variant of the negative priming paradigm ought to provide a bridge between these two phenomena. Indeed, a recent series of studies that successively stripped the spatial negative priming procedure of its distractors revealed that a single prime that provides no predictive informa- tion about the location of a subsequent single probe is sufficient to observe a robust negative priming effect (Milliken, 1995; Milliken, Tipper, Houghton, & Lupianez, 1997; see also Park & Kanwisher, 1994). Although the inhibition of return literature has its share of peculiarities, these data suggest that distinctions drawn between negative priming and inhibition of return may well be related to procedure rather than to process.
Inhibitory Semantic Priming in Lexical Decision
An inhibitory priming effect that is not commonly ascribed to the same mechanism as negative priming has been reported in the lexical-decision task (Carr & Dagenbach, 1990; Dagenbach, Carr, & Barnhardt, 1990; Dagenbach, Carr, & Wilhelmsen, 1989). Dagenbach et al. (1989) showed that both positive and negative semantic priming effects can result from near-threshold presentation of primes in a lexical-decision task. Which of the two results was observed in their studies depended on the nature of a threshold setting procedure that preceded the lexical-deci- sion task.
In prior studies, Marcel (1983) used a presence-absence threshold setting procedure to establish a prime duration at which participants would remain unaware of the prime. In those studies, positive semantic priming for masked primes was ob- served. Dagenbach et al. (1989) also observed positive priming by using this threshold setting procedure. However, when they used a threshold setting task that required participants to judge
which of two probes was semantically related to a masked prime, a negative semantic priming effect was observed.
The different direction of priming effects observed in the Marcel (1983) and Dagenbach et al. (1989) studies must be attributed to different strategic sets established in the threshold setting procedures of the two studies. In particular, training participants to match a target back to a prime teaches partici- pants to discriminate between perceptual information from the current stimulus and memorial information that owes to the prime stimulus. When a prime is encoded without attentive anal- ysis, this discrimination strategy results in slower rather than faster responses to related probes. In other words, rather than use the familiarity of a related target as evidence favoring a word response (see Ratcliff & McKoon, 1988), participants appear to have been taught by the threshold setting procedure to differentiate between activation that owes to a target stimulus and activation that owes to a related prime stimulus. Viewed in this light, the contrast between negative and positive priming in these lexical-decision studies is analogous to the dependence of negative priming on probe task selection examined here. In the Dagenbach et al. (1989) study, and in the experiments reported here in which a selection probe task was used, opposition inher- ent to temporal discrimination resulted in slower responses to probes that were related to previous primes. In the Marcel study, and in the experiments reported here in which a lone probe item was used, positive priming effects may be attributed to the predominance of automatic retrieval processes.
Temporal Discrimination and Selective Attention
Negative priming captured, and has maintained, the interest of a wide number of researchers for several reasons. Foremost among these reasons is that, at an intuitive level, it seems an empirical window to a theoretical question that has troubled psychologists from the time of William James ( 1890/1950): Is the focus of attention a product of active processes that filter, or push into the phenomenal background, information that is irrelevant to current thought and behavior? Certainly, the slowed responses to previously ignored stimuli that define negative priming are consistent with the view that representations of these ignored stimuli are subject to active inhibitory processing. At the same time, early studies of negative priming provided unambiguous evidence against intuitively sensible inhibitory- based explanations of negative priming (Lowe, 1979; Tipper & Cranston, 1985).
Recent work has focused on the possibility that retrieval pro- cesses play a critical role in determining negative priming (Neill et al., 1992), and there are now considerable data to support this view (Neill, 1997; Neill & Valdes, 1992; Neill et al., 1992; see also Milliken et al., 1994; Park & Kanwisher, 1994). Yet, to those interested in attention, losing the negative priming phe- nomenon to those interested in memory is bitter medicine. One of the purposes that we hope this article serves is to point out that negative priming does not cease to be an attentional phenomenon because it has been shown to be a phenomenon of memory retrieval as well. Memory and attention are concepts that would profit from being studied together rather than in isolation.
The principal observations reported in this article, comple-

NEGATIVE PRIMING 227
mented by an appreciation of related work and discussion by Johnston and Hawley (1994), prompted us to develop a tempo- ral discrimination framework for explaining negative priming. The temporal discrimination framework blends attentional and memorial processes and recasts the significance of negative priming and other empirical phenomena to the broader concep- tual issue of selective attention.
The fundamental question underlying research in selective attention concerns how people avoid being overloaded by the seemingly limitless information available at any given time. One initial answer to this question is that very little of this input is processed in a resource-demanding analytic manner. Instead, a great deal of behavior is determined "automat ical ly" by the relatively effortless reinstatement of prior learning. However, for an organism to survive it must be able to respond efficiently to situations in which a prior learned response is inaccessible. Following many others, we have suggested that an orienting system ensures that new learning occurs efficiently (Grossberg, 1994; Sokolov, 1963). Cast within this framework, negative priming provides a fascinating example of opposition between two fundamental objectives of an adaptive system: efficient rein- statement of old learning and efficient engagement in new learning.
References
Allport, D. A., Tipper, S. P., & Chmiel, N. (1985). Perceptual integration and post-categorical filtering. In M. I. Posner & O. S. M. Marin (Eds.), Attention and performance X1 (pp. 107-132). Hillsdale, N J: Erlbaum.
Becker, S., Moscovitch, M., Behrmann, M., & Joordens, S. (1997). Long-term semantic priming: A computational account and empirical evidence. Journal of Experimental Psychology: Learning, Memory, and Cognition, 23, 1059-1082.
Beech, A. R., Powell, T. J., McWilliams, J., & Claridge, G. S. (1989). Evidence of reduced "cognitive inhibition" in schizophrenia. British Journal of Clinical Psychology, 28, 110-116.
Benoit, G., Fortran, L., Lemelin, S., LaPlante, L., Thomas, J., & Everett, J. (1992). L' attention selective dans la depression majeure: Ralentisse- ment clinique et inhibition cognitive [Selective attention in major depression: Clinical slowing and cognitive inhibition]. Canadian Journal of Psychology, 46, 41-52.
Broadbent, D. E. (1958). Perception and communication. London: Per- gamon Press.
Carr, T. H., & Dagenbach, D. (1990). Semantic priming and repetition priming from masked words: Evidence from a center-surround atten- tional mechanism in perceptual recognition. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 341-350.
Cartwright, D., & Festinger, L. (1943). A quantitative theory of decision. Psychological Review, 50, 596-621.
Cherry, E. C. (1953). Some experiments upon the recognition of speech, with one and with two ears. Journal of the Acoustical Society of America, 25, 975-979.
Cohen, J. D., & Servan-Schreiber, D. (1992). Context, cortex, and dopa- mine: A connectionist approach to behavior and biology in schizo- phrenia. Psychological Review, 99, 45-77.
Collins, A. M., & Loftus, E. L. (1975). A spreading-activation theory of semantic processing. Psychological Review, 82, 407-428.
Dagenbach, D., Carr, T. H., & Barnhardt, T. M. (1990). Inhibitory se- mantic priming of lexical decisions due to failure to retrieve weakly activated codes. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 328-340.
Dagenbach, D., Carr, T. H., & Wilhelmsen, A. (1989). Task-induced
strategies and near-threshold priming: Conscious effects on uncon- scious perception. Journal of Memory and Language, 28, 412-443.
Dalrymple-Alford, E. C., & Budayr, B. (1966). Examination of some aspects of the Stroop Color-Word Test. Perceptual & Motor Skills, 23, 1211-1214.
Dark, V. J., & Schmidt, P. A. (1996, November). Negative priming and stimulus activation. Paper presented at the 37th annual meeting of the Psychonomic Society, Chicago.
DeSchepper, B., & Treisman, A. ( 1991, November). Novel visual shapes in negative priming. Paper presented at the 32rid annual meeting of the Psychonomic Society, San Francisco.
DeSchepper, B., & Treisman, A. (1996). Visual memory for novel shapes: Implicit coding without attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 22, 27-47.
Deutsch, J. A., & Deutsch, D. (1963). Attention: Some theoretical con- siderations. Psychological Review, 70, 80-90.
Driver, J., & Tipper, S. P. (1989). On the nonselectivity of "selective" seeing: Contrasts between interference and priming in selective atten- tion. Journal of Experimental Psychology: Human Perception and Performance, 15, 304-314.
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16, 143-149.
Fox, E. (1995). Negative priming from ignored distractors in visual selection: A review. Psychonomic Bulletin and Review, 2, 145-173.
Fuentes, L. J., & Tudela, P. (1992). Semantic processing of foveally and parafoveally presented words in a lexical decision task. Quarterly Journal of Experimental Psychology, 45A, 299-322.
Greenwald, A.G. (1972). Evidence of both perceptual filtering and response suppression for rejected messages in selective attention. Journal of Experimental Psychology, 94, 58-67.
Grossberg, S. (1994). The attentive brain. American Scientist, 83, 438- 449.
Hasher, L., Stoltzfus, E. R., Zacks, R. T., & Rypma, B. ( 1991 ). Age and inhibition. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 163-169.
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 53-112). Orlando, FL: Academic Press.
Jacoby, L. L. (1983). Perceptual enhancement: Persistent effects of an experience. Journal of Experimental Psychology: Learning, Memory, and Cognition, 9, 21-38.
Jacoby, L. L., & Dallas, M. (1981). On the relationship between auto- biographical memory and perceptual learning. Journal of Experimen- tal Psychology: General, 3, 306-340.
Jacoby, L. L., Kelley, C. M., & Dywan, J. (1989). Memory attributions. In H. L. Roediger III & E I. M. Craik (Eds.), Varieties of memory and consciousness: Essays in honor of Endel Tulving (pp. 391-422 ). Hillsdale, NJ: Erlbaum.
James, W. (1950). Principles of psychology. New York: Holt. (Original work published 1890)
Johnson, M. K., Hashtroudi, S., & Lindsay, D.S. (1993). Source moni- toring. Psychological Bulletin, 114, 3-28.
Johnston, W. A., & Hawley, K. J. (1994). Perceptual inhibition of ex- pected inputs: The key that opens closed minds. Psychonomic Bulletin and Review, 1, 56-72.
Joordens, S., & Becker, S. (1997). The long and short of semantic priming effects in lexical decision. Journal of Experimental Psychol- ogy: Learning, Memory, and Cognition, 23, 1083-1105.
Joordens, S., & Besner, D. (1992). Priming effects that span an interven- ing unrelated word: Implications for models of memory representation and retrieval. Journal of Experimental Psychology: Learning, Mem- ory, and Cognition, 18, 483-491.

228 MILLIKEN, JOORDENS, MERIKLE, AND SEIFFERT
Kahneman, D., Treisman, A., & Gibbs B.J. (1992). The reviewing of object files: Object-specific integration of information. Cognitive Psychology, 24, 175-219.
Kane, M. J., May, C. P., Hasher, L., Rahhal, T., & Stoltzfus, E. R. (1997). Dual mechanisms of negative priming. Journal of Experimental Psy- chology: Human Perception and Performance, 23,632-650.
Kintsch, W. (1980). Semantic memory: A tutorial. In R. S. Nickerson ( Ed.), Attention and performance VIII (pp. 595-620). Hillsdale, NJ: Erlbaum.
Kramer, A. E, Humphrey, D. G., Larish, J. E, Logan, G. D., & Strayer, D. L. (1994). Aging and inhibition: Beyond a unitary view of inhibi- tory processing in attention. Psychology and Aging, 9, 491-512.
Link, S. W. (1975). The relative judgment theory of two choice response time. Journal of Mathematical Psychology, 12, 114-135.
Link, S. W. (1992). The wave theory of difference and similarity. Hills- dale, NJ: Erlbaum.
Logan, G.D. (1988). Toward an instance theory of automatization. Psychological Review, 95, 492-527.
Lowe, D. G. (1979). Strategies, context and the mechanisms of response inhibition. Memory & Cognition, 7, 382-389.
Malley, G. B., & Strayer, D. L. (1995). Effect of stimulus repetition on positive and negative identity priming. Perception & Psychophysics, 57, 657-667.
Marcel, A. J. (1983). Conscious and preconscious perception: Experi- ments on visual masking and word recognition. Cognitive Psychology, 15, 197-237.
Masson, M. E. J. ( 1991 ). A distributed memory model of context effects in word recognition. In D. Besner & G. W. Humphreys (Eds.), Basic processes in reading: Visual word recognition (pp. 233-263). Hills- dale, NJ: Erlbaum.
May, C. P., Kane, M. J., & Hasher, L. (1995). Determinants of negative priming. Psychological Bulletin, 118, 35-54.
Maylor, E. A. (1985). Facilitatory and inhibitory components of orient- ing in visual space. In M. I. Posner & O. S. M. Marin (Eds.),Attention and performance XI (pp. 189-204). Hillsdale, NJ: Erlbaum.
McDowd, J. M. & Oseas-Kreger, D. M. (1991). Aging, inhibitory pro- cesses, and negative priming. Journal of Gerontology, 46, 340-345.
McNamara, T.P. (1994). Theories of priming: II. Types of primes. Journal of Experimental Psychology: Learning, Memory, and Cogni- tion, 20, 507-520.
Merikle, P.M., & Joordens, S. (1997). Parallels between perception without attention and perception without awareness. Consciousness and Cognition, 6, 219-236.
Meyer, D. E., & Schvaneveldt, R. W. ( 1971 ). Facilitation in recognizing pairs of words: Evidence of a dependence between retrieval opera- tions. Journal of Experimental Psychology, 90, 227-234.
Meyer, D. M., Schvaneveldt, R.W., & Ruddy, M.G. (1975). Loci of contextual effects on visual word recognition. In P. M. A. Rabbitt & S. Domic (Eds.), Attention and performance V (pp. 98-118). San Diego, CA: Academic Press.
Milliken, B. ( 1995, June). Allocating attention and not allocating atten- tion: Ignoring. Paper presented at the meeting of the Canadian Society for Brain, Behavior, and Cognitive Science, Halifax, Nova Scotia, Canada.
Milliken, B., & Joordens, S. (1996). Negative priming without overt prime selection. Canadian Journal of Experimental Psychology, 50, 333-346.
Milliken, B., Tipper, S. P., Houghton, G., & Lupianez, J. (1997). At- tending, ignoring, and repetition: On the relation between negative priming and inhibition of return. Manuscript submitted for publication.
Milliken, B., Tipper, S.P., & Weaver, B. (1994). Negative priming in a spatial localization task: Feature mismatching and distractor inhibi-
tion. Journal of Experimental Psychology: Human Perception and Performance, 20, 624-646.
Moore, C. M. (1994). Negative priming depends on probe-trial conflict: Where has all the inhibition gone? Perception & Psychophysics, 56, 133-147.
Moran, J., & Desimone, R. (1985, August). Selective attention gates visual processing in the extrastriate cortex. Science, 229, 782-784.
Moray, N. (1959). Attention in dichotic listening: Affective cues and the influence of instructions. Quarterly Journal of Experimental Psy- chology, 11, 56-60.
Morton, J. (1969). Interaction of information in word recognition. Psy- chological Review, 76, 165-178.
Neely, J. H. (1977). Semantic priming and retrieval from lexical mem- ory: Roles of inhibitionless spreading activation and limited-capacity attention. Journal of Experimental Psychology: General, 106, 226- 254.
Neill, W.T. (1977). Inhibition and facilitation processes in selective attention. Journal of Experimental Psychology: Human Perception and Performance, 3, 444-450.
Neill, W. T (1997). Episodic retrieval in negative priming and repetition priming. Journal of Experimental Psychology: Learning, Memory, and Cognition, 23, 1291-1305.
Neill, W.T., Lissner, L.S., & Beck, J.L. (1990). Negative priming in same-different matching: Further evidence for a central locus of inhibition. Perception & Psychophysics, 48, 398-400.
Neill, W. T., Terry, K.M., & Valdes, L.A. (1994). Negative priming without probe selection. Psychonomic Bulletin and Review, 1, 119- 121.
Neill, W. T., & Valdes, L. A. (1992). The persistence of negative prim- ing: Steady-state or decay? Journal of Experimental Psychology: Learning, Memory, and Cognition, 18, 565-576.
Neill, W. T., & Valdes, L. A. (1996). Facilitatory and inhibitory aspects of attention. In A. E Kramer, M. Coles, & G. D. Logan (Eds.), Con- verging operations in the study of visual selective attention (pp. 77- 106). Washington, DC: American Psychological Association.
Neill, W. T., Valdes, L. A., & Terry, K. M. (1994). Selective attention and the inhibitory control of cognition. In E N. Dempster & C.J. Brainerd (Eds.), New perspectives on interference and inhibition in cognition (pp. 207-261 ). New York: Academic Press.
Neill, W. T., Valdes, L. A., Terry, K. M., & Gorfein, D. S. (1992). The persistence of negative priming: II. Evidence for episodic trace re- trieval. Journal of Experimental Psychology: Learning, Memory, and Cognition, 18,993-1000.
Neill, W.T., & Westberry, R.L. (1987). Selective attention and the suppression of cognitive noise. Journal of Experimental Psychology: Learning, Memory and Cognition, 13, 327-334.
Neumann, E., & DeSchepper, B. G. ( 1991 ). Costs and benefits of target activation and distractor inhibition in selective attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 1136-1145.
Ortells, J. J., & Tudela, E (1996). Positive and negative semantic prim- ing of attended and unattended parafoveal words in a lexical decision task. Acta Psychologica, 94, 209-226.
Park, J., & Kanwisher, N. (1994). Negative priming for spatial location: Identity mismatching, not distractor inhibition. Journal of Experimen- tal Psychology: Human Perception & Performance, 20, 613-623.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Houma & D. G. Bouwhuis (Eds.), Attention and performance X: Control of language processes. Hillsdale, N J: Eflbaum.
Posner, M. I., & Snyder, C. R. R. (1975). Attention and cognitive con- trol. In R. L. Solso (Ed.), The Loyola Symposium: Information pro- cessing and cognition (pp. 531-556). Hillsdale, NJ: Erlbaum.
Rafal, R. D., Calabresi, E, Brennan, C., & Sciolto, T. (1989). Saccade preparation inhibits reorienting to recently attended locations. Journal

NEGATIVE PRIMING 229
of Experimental Psychology: Human Perception and Performance, 15, 673-685.
Ratcliff, R. (1978). A theory of memory retrieval. Psychological Re- view, 85, 59-108.
Ratcliff, R., & McKoon, G. (1988). A retrieval theory of priming in memory. Psychological Review, 101, 177-184.
Scarborough, D.L., Cortese, D., & Scarborough, H.S. (1977). Fre- quency and repetition effects in lexical memory. Journal of Experi- mental Psychology: Human Perception and Performance, 3, 1 - 17.
Scarborough, D. L., Gerard, L., & Cortese, C. (1979). Accessing texical memory: The transfer of word repetition effects across task and mo- dality. Memory & Cognition, Z 3-12.
Schneider, W. (1988). Micro-experimental laboratory: An integrated system for IBM PC compatibles. Behavior Research Methods, Instru- ments, and Computers, 20, 206-217.
Shapiro K. L., & Loughlin, C. (1993). The locus of inhibition in the priming of static object: Object token versus location: Journal of Experimental Psychology: Human Perception and Performance, 19, 352-363.
Sokolov, E. N. (1963). Perception and the conditioned reflex. New York: Pergamon.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18, 643-662.
Sullivan, M. P., & Faust, M. E. (1993). Evidence for identity inhibition during selective attention in old adults. Psychology and Aging, 8, 589-598.
Sullivan, M.P., Faust, M.E., & Balota, D. (t995). Identity negative priming in old adults and individuals with dementia of the Alzheimer's type. Neuropsychology, 9, 537-555.
Tipper, S. P. (1985). The negative priming effect: Inhibitory effects of ignored primes. Quarterly Journal of Experimental Psychology, 37A, 571-590.
Tipper, S. P. (1991). Less attentional selectivity as a result of declining inhibition in older adults. Bulletin of the Psychonomic Society, 29, 45 -47.
Tipper, S. P., & Baylis, G. C. (1987). Individual differences in selective attention: The relation of priming and interference to cognitive failure. Personality and Individual Differences, 8, 667-675.
Tipper, S. P., Bourque, T., Anderson, S., & Brehaut, J. C. (1989). Mecha- nisms of attention: A developmental study. Journal of Experimental Child Psychology, 48, 353-378.
Tipper, S. P., Brehaut, J. C., & Driver, J. (1990). Selection of moving and static objects for the control of spatially directed action. Journal
of Experimental Psychology: Human Perception and Performance, 16, 492-504.
Tipper, S. P., & Cranston, M. (1985). Selective attention and priming: Inhibitory and facilitatory effects of ignored primes. Quarterly Jour- nal of Experimental Psychology, 37A, 591-61 I.
Tipper, S. P., Driver, J., & Weaver, B. ( 1991 ). Object-centered inhibition of return of visual attention. Quarterly Journal of Experimental Psy- chology, 37A, 591-611.
Tipper, S.P., Lortie, C., & Baylis, G. C. (1992). Selective reaching: Evidence for action-centered attention. Journal of Experimental Psy- chology: Human Perception and Performance, 18, 891-905.
Tipper, S., Weaver, B., & MiUiken, B. (1995). Spatial negative priming without mismatching: A reply to Park and Kanwisher. Journal of Experimental Psychology: Human Perception and Performance, 21, 1220-1229.
Treisman, A. M. (1960). Contextual cues in selective listening. Quar- terly Journal of Experimental Psychology, 12, 242-248.
Treisman, A. M. (1964). Monitoring and storage of irrelevant messages and selective attention. Journal of Verbal Learning and Verbal Behav- ior, 3, 449-459.
Treisman, A., & DeSchepper, B. (1996). Object tokens, attention, and visual memory. In T. Inni & J. McClelland (Eds.), Attention and performance XVI (pp. 15-46). Cambridge, MA: MIT Press.
Van Selst, M.A., & Jolicoeur, P. (1994). A solution to the effect of sample size on outlier elimination. Quarterly Journal of Experimental Psychology, 47A, 631-650.
Walley, R. E., & Weiden, T. D. (1973). Lateral inhibition and cognitive masking: A neuropsychological theory of attention. Psychological Review, 80, 284-302.
Whittlesea, W. A. (1997). Production, evaluation, and preservation of experiences: Constructive processing in remembering and perfor- mance tasks. In D. Medin (Ed.), Psychology of learning and motiva- tion (Vol. 37, pp. 211-264). San Diego, CA: Academic Press.
Whittlesea, B. W. A., & Jacoby, L. L. (1990). Interaction of prime repe- tition with visual degradation: Is priming a retrieval phenomenon? Journal of Memory and Language, 29, 546-565.
Yee, P. L. ( 1991 ). Semantic inhibition of ignored words during a figure classification task. The Quarterly Journal of Experimental Psychol- ogy, 43A, 127-153.
Received March 17, 1995 Revision received July 9, 1997
Accepted July 9, 1997 I